
Submitted:
26 December 2023
Posted:
27 December 2023
You are already at the latest version
Abstract
In recent years, extensive research has delved into the pathophysiology of local reactions triggered by Bothrops snake venoms. Even though antivenom works well at reducing death and systemic effects, it is still not very effective in treating local reactions because the venom works so quickly, and antivenom cannot fix injuries that have already been triggered. This might be attributed to certain molecules amplifying the venom-induced innate response. While evidence suggests endogenous mediators at the venom site play a role in this envenomation, in Brazil, concurrent use of anti-inflammatories or other drugs associated with the antivenom remains uncommon. The aim of this study was to determine how various inflammatory mediators affected the leukocyte-endothelium interaction (LEI) following a B. jararaca bite. According to our findings, nitric oxide and cyclooxygenase pathway eicosanoids significantly contribute to alteration of LEI induced by B. jararaca venom. Conversely, lipoxygenase-mediated eicosanoids, histamine, or serotonin had minimal participation. Markedly, treatment with dexamethasone alongside antivenom mitigated alterations of LEI induced by the B. jararaca snake venom. The limited efficacy of the antivenom in managing Bothrops venom-induced local reactions emphasizes the critical need for supplementary treatments to enhance therapeutic outcomes.
Keywords:
Bothrops jararaca
; Snakebite
; Local Lesions
; Inflammation
; Leukocyte-endothelium interactions
; Antivenom
; Anti-inflammatory drugs
; Microcirculation.
1. Introduction
Snakebites are a serious global health issue prevalent among poor populations, causing significant social problems [1,2,3]. The World Health Organization (WHO) estimates approximately 2.7 million snakebite incidents worldwide annually, resulting in about 81,000 to 138,000 fatalities and 400,000 survivors with enduring disabilities [4,5].
With high mortality and morbidity rates, snakebites primarily affect underprivileged populations in tropical and subtropical areas, according to the WHO, which recognizes them as a neglected tropical disease. Since 2017, snakebite envenoming has been listed among the 20 neglected tropical diseases, prompting a strategic plan to reduce fatalities by 50% by 2030. Despite being preventable and treatable, research investment in this area remains remarkably low [6]. The Brazilian Academy of Sciences (BAS) has identified accidents involving venomous animals as a neglected disease in Brazil. One directive of BAS prioritizes supporting research on drug combinations for treating toxin-induced accidents [7].
In Brazil, an average of 29,000 snakebite cases occurs annually, resulting in approximately 120 fatalities per year and around 600 cases with curable sequelae, excluding unreported cases. Among venomous snakes, the Viperidae family, particularly the subfamilies Crotalinae (including snakes of the genera Crotalus, Bothrops, and Lachesis), are noteworthy [8,9].
Bothrops snakebites causes systemic reactions, such as severe blood clotting disorders [10,11], which are like symptoms of disseminated intravascular coagulation [12]. Complications such as hypotension and hypovolemic shock can lead to fatalities. Severe local reactions comprising edema, pain, hemorrhage, and necrosis are common, often resulting in substantial tissue loss and potential amputations [13,14]. These reactions trigger inflammatory responses by starting up signaling pathways that cause the transcription of inflammatory genes, such as cytokines and eicosanoids [15]. This causes endothelial activation with the expression of lectins and adhesion molecules, which causes the interaction of leukocytes with the endothelium, resulting in firm adhesion and cell migration [16].
The pathogenesis of local reactions caused by Bothrops venoms is still not fully understood because they are made up of a lot of different toxin classes that can work alone or together through different pathways in the body [17,18].
The only effective treatment for snakebite envenomation is specific antivenom [6,19,20]. Despite its efficacy in reducing lethality and reversing systemic effects, antivenom inadequately addresses local reactions, causing severe sequelae [10,11,13]. This deficiency arises from the rapid onset of these local actions and due the fact that the antivenomcannot reverse established or triggered injuries or neutralize endogenous mediators involved in the process [10,21].
Essential anti-inflammatory drugs include glucocorticoids (steroids), which inhibit early and late inflammatory manifestations, and non-steroidal drugs, among the most widely used globally [22]. The literature shows that treating animals with anti-inflammatories, particularly phospholipase A2 and cyclooxygenase (COX) inhibitors, significantly reduces edema induced by Bothrops venoms [23,24,25].
Despite evidence implicating endogenous mediators released at the venom injection site, using anti-inflammatories or other drugs alongside serotherapy remains uncommon in Brazil [14]. The limited effectiveness of serotherapy in treating Bothrops venom-induced local reactions prompts the search for complementary treatments to improve this condition. In recent decades, many studies have sought to understand local reactions caused by Bothrops venoms and explore complementary therapies alongside antivenom. Making antivenoms better and more effective [26], using protease inhibitors [27], identifying animals resistant to snake venom [28], and utilizing therapeutic combinations that work with serotherapy [25,29] are a few of these.
Therefore, the purpose of this study was to ascertain which inflammatory mediators are involved in the alterations of the leukocyte-endothelium interactions following a Bothrops jararaca experimental envenomation. The goal was to find possible treatments that could be used along with antivenom to help reverse the severe effects of the venom's quick action.
The results have shown that alterations in the leukocyte-endothelium interactions induced by the Bothrops jararaca venom are mediated mainly by cyclooxygenase-derived eicosanoids and that the association of dexamethasone with the antivenoms could reduce this effect.
2. Materials and Methods
2.1. Animals
Male Swiss mice weighing 20–25 g were used. Animals were maintained for two days at the laboratory before experiments. The Ethical Committee for Use of Animals of the Butantan Institute (protocol no. 199/05) approved all experimental procedures in accordance with the ethical guidelines put forth by the International Society of Toxinology and the Brazilian College of Experimental Animals.
2.2. Bothrops jararaca venom (BjV)
The Laboratory of Herpetology at the Butantan Institute provided a pool of lyophilized venom from various adult specimens of B. jararaca snakes. The venom was stored at -20 °C, and solutions (w/v) were prepared immediately before use in sterile saline.
2.3. Antivenom
The Butantan Institute produced the Bothrops antivenom (BAV) that was kept at 8 °C until use. A combination of snake venoms from Bothrops jararaca, B. jararacussu, B. moojeni, B. neuwiedi, and B. alternatus is used in horse immunization to produce BAV. The antivenom is subjected to multiple treatments during the production process to get the F(ab’)2 fraction of IgG. BAV is available in flasks of 10 mL, and according to the supplier, each mL neutralizes 5 mg of the Bothrops jararaca reference venom.
2.4. Treatment of animals
Groups of animals (n= 5 per group) received the following treatments 30 minutes or 1 hour before or after the s.c. injection of BjV into the scrotal bag:
1) Dexamethasone (a corticosteroid Decadron, Promade, Brazil; 1 mg/kg dissolved in sterile saline, i.p.) 1h before or after the s.c. injection of BjV [25];
2) Indomethacin (a cyclooxygenase inhibitor, Sigma-3 mg/kg dissolved in 1M TRIS buffer, pH 8, at 37 °C and sterile saline solution (1:10), i.p.), 30 minutes before the BjV was injected subcutaneously [25].
3) Celecoxib (a cyclooxygenase-2 inhibitor, Celebra®, Pfizer, Brazil) 30 mg/kg. For each tablet containing 200mg, 2.5 mL of a 1% carboxymethyl (CMC) solution is added, v.o., 30 minutes before the s.c. injection of BjV [30].
4) L-NG-nitro arginine methil ester-L-NAME (a nonspecific inhibitor of the enzyme nitric oxide synthase (NOS), Sigma-100 mg/kg, dissolved in sterile saline, s.c.) 30 minutes before the s.c. injection of BjV [31];
5) Aminoguanidine hydrochloride (a selective enzyme inhibitor inducible nitric oxide (iNOS), Sigma- 50 mg/kg dissolved in sterile saline, i.p.) 30 minutes before the s.c. injection of BjV [31];
6) Promethazine hydrochloride (an H1 receptor inhibitor, Sigma, 10 mg/kg dissolved in sterile saline, i.p.) 30 minutes before the s.c. injection of BjV [32];
7) Methysergide maleate salt (a serotonin 5HT receptor antagonist, Sigma-0.8 mg/kg dissolved in sterile saline, i.p.) 30 minutes before the s.c. injection of BjV [32];
8) Nordihydroguaiaretic acid-NDGA (a 5-inhibitor lipoxygenase, Sigma, 30 mg/kg; for every 30 mg/kg of the compound, 2 mL of solution containing ethanol-saline (1:9) was added, the pH adjusted to 7.5 with 0.1N NaOH, i.p.) 30 minutes before the s.c. injection of BjV [32];
9) Bothrops antivenom- BAV (0.2 mL, i. v.). The antivenom was administered alone or in association with dexamethasone 1 h after the s.c. injection of BjV at the dose and routes specified. The control group received saline by the same routes instead. [25].
2.5. Intravital microscopy of murine cremaster venules
Intravital microscopy evaluated leukocyte-endothelial interactions in mouse cremaster venules. Five mice per group were injected under the skin of their scrotum with either 100 µL of sterile saline (as a control) or 1.0 mg/100 ml of BjV (raw or treated). The doses of BjV used were sub-hemorrhagic. After 2 or 24 h, animals were anesthetized by an injection (i.p.) of ketamine (100 mg/kg) and xylazine (10 mg/kg), and the cremaster muscle was exteriorized for microscopic examination in situ as described by Baez [33]. The microcirculation of the cremaster muscle was visualized in a transparent board window heated at 37 ºC, on which the anesthetized mouse was maintained. Leukocyte responses on cremaster muscle microcirculation were evaluated by light microscopy (Axioplan II, Carl Zeiss, Germany, equipped with Achroplan objectives of 10.0 longitudinal distance, 0.25 numeric apertures, and 1.60 optovar). Images were captured and digitalized on the computer for further analysis using the image analyzer software (KS 300, Kontron, Carl Zeiss, Germany). One post-capillary venule (30–40 mm in diameter) was selected at random. After the stabilization period (10 min), adhering leukocytes were counted for 5 minutes in a 100-µm vascular section. Adherent leukocytes were considered cells that remained immobile for at least 30 s within a given 100-µm vessel segment. It was possible to count leukocytes that had moved into tissue outside of vessels that was within 50 µm of all sides of the 100-µm segments of vessels that were being studied. When comparing adhered leukocytes, the results from groups that were studied two hours after the toxin injection were used. When comparing migrated leukocytes, the results were used 24 hours after the venom injection.
2.6. Statistical analysis
Results were expressed as mean ± standard error of the mean and analyzed by one-way ANOVA, followed by Tukey’s test. Differences in results were considered significant at p<0.05.
3. Results
BjV induces changes in the leukocyte-endothelium interaction in the mouse's microcirculation cremaster muscle, with a significant increase in adhered and migrated cells after 2 hours (Figure 1A) and 24 hours (Figure 1B), respectively, after its exposure.
When we pre-treated with dexamethasone (Dx) 1 hours before venom injection with the purpose of verifying the participation of arachidonic acid degradation metabolites in leukocyte-endothelium interaction events induced by BjV injection, we observed that the treatment could reduce the number of adhered and migrated cells in the groups analyzed at 2 and 24 hours, when compared with the control or with the group inflamed only with BjV (Figure 1A and Figure 1B).
An investigation into the role of cyclooxygenase enzymes (COX) in microcirculation cellular events was also performed using indomethacin (Indo) or celecoxib, which blocks products coming from the cyclooxygenase-2 (COX-2) pathway, 30 minutes before BjV was injected under the skin of mice's scrotums. Both drugs were efficient in inhibiting the number of adhered and migrated cells at both times evaluated (Figure 2A and 2B and Figure 3A and B).
Although celecoxib inhibited adhesion and migration events after 2 hours and 24 hours, respectively, it had no role in inhibiting migrated cells after 2 hours of exposure to the venom (Figure 3A and Figure 3B). The initial times of the leukocyte-endothelium interaction are predominantly cell adhesion, and in the later times, there is migration to the extravascular space since these interactions occur because of the expression of adhesion molecules that have sequential and orchestrated kinetics [33,34].
Other inhibitors that have been found effective in preventing microcirculation changes are those targeting the nitric oxide pathway. To determine their role in these changes, we gave mice L-NAME, which blocks nitric oxide synthase (NOS), or aminoguanidine, which blocks inducible nitric oxide synthase (iNOS). This was done for 30 minutes before injecting BjV into the subcutaneous tissue of the scrotum. Both inhibitors significantly reduced the number of adhered and migrated cells at 2 and 24 hours, as depicted in Figure 4 and Figure 5.
Using vasoactive amine inhibitors before the BjV treatment did not change the way the leukocytes and endothelium interacted. Cell adhesion and migration were not different in the groups that were first given promethazine (Pro), which blocks the histamine H1 receptor, or methysergide (Met), which blocks the serotonin (5-HT2) receptor. This was true at both time points studied (Figure 6A and B, Figure 7A and B).
When the lipoxygenase pathway inhibitor NDGA was used first, there were no changes in the number of cells that stuck to or moved through the post-capillary venules (Figure 8A and B).
All the drugs used per se did not cause significant changes in the leukocyte-endothelium interaction patterns (data not shown).
It was tested to see if dexamethasone could reverse the inflammatory effect that BjV causes by treating the mice with this drug an hour after BjV was injected under the skin in their scrotum. There were a lot fewer adherent cells after treatment with Dx compared to the group that was injected with venom, but there were no statistical differences between the Dx group and the control group (Figure 9A). This decrease was also observed in the cell migration event after 24 hours when compared to the poisoned group (Figure 9B).
One hour after the BjV injection, treatment with antivenom (BAV) was done to see how well the antivenom stopped the changes that the venom caused in the interaction between leukocytes and endothelium. However, we found the antivenom could not stop the microcirculatory alterations. There was an increase in adhered and migrated cells at both assessed times—2 and 24 hours—as shown in Figure 10A and Figure 10B. This increase in changes in leukocyte-endothelium interactions did not differ from the envenomed group and with no treatment.
Figure 10A and Figure 10B show that the group that was treated with BAV+Dx had fewer cells that stuck to and migrated. This means that the anti-inflammatory drug association lowers the number of white blood cells that are recruited even an hour after envenomation and the interactions between these cells and the endothelium.
4. Discussion
Using specific antitoxins is the only proven and recommended treatment by the World Health Organization for cases of snakebites [6]. Since introducing antivenom treatment, there has been a significant decrease in the number of deaths caused by non-medical accidents [14].
Despite the effectiveness of serotherapy in neutralizing systemic symptoms, local reactions induced by Bothrops venom do not always respond to such treatment. This is because of the rapid manifestation of these local reactions and the fact that antivenom agents cannot reverse the already established or triggered lesions nor neutralize the endogenous mediators involved in the process [10].
Bothrops venoms can induce inflammatory responses, activating signaling pathways that culminate in the transcription of inflammatory genes such as cytokines and eicosanoids, such as tumor necrosis factor-alpha (TNF-α), interleukin-1 (IL-1) and interleukin-6 (IL-6), with consequent release of vasoactive substances and consequent increase in vascular permeability, activation of endothelial cells, expression of adhesion molecules, which will trigger capture, rolling, firm adherence and cell migration [15,34,35,36,37,38].
Previous studies of our group have shown the pro-inflammatory action of the Bothrops jararaca snake venom [25,26] and that this action is because of the constituent metalloproteinases of the venom [27]. We have recently shown that Jar, Jar-C, and BnP1, different Snake Venom Metalloproteinases, can induce adhesion and cell migration in post-capillary venules in the cremaster muscle of mice, observed by intravital microscopy [39] and that the alterations in leukocyte-endothelium interaction occur because of the participation of the adhesive molecule ICAM-1 in early times and of PECAM-1, in later times [34].
In the present study, we note that both the pre- and post-treatment with dexamethasone could decrease the number of adhered and migrated cells in the two times analyzed when compared with the control (non-envenomed) and with the BjV-inflamed group (Figure 1 and Figure 9), demonstrating the participation of arachidonic acid degradation metabolites and the ability of dexamethasone to reverse changes in leukocyte-endothelium interactions induced by the BjV.
The eicosanoids originating from the cyclooxygenase pathway, but not those originating in the lipoxygenase route, mediate the endothelial leukocyte interaction induced by BjV. Studies have shown the participation of products originating from eicosanoids in cell migration induced by the venom of B. jararaca [40]. Also, concerning the role of glucocorticoids in leukocyte recruitment, it has been described that glucocorticoids reduce cell recruiting to the site of inflammation by inhibition of adhesion molecules, as they induce a rapid change in the surface of the CAMs, by a genomic mechanism, or by interfering with the expression of the ICAM-I adherence molecule in the IL-1-induced leucocyte adherence in the mouse mesentery [41,42,43]. It is known that the migration of inflammatory cells induced by Bothrops venoms is mediated by eicosanoids and that this recruitment depends on the expression of adhesion molecules such as ICAM-1, LECAM-1, LFA-1, PECAM-1, but not MAC-1 [40,44,45].
The expression of these adhesion molecules results in increased leukocyte-endothelium interaction, the first step before the process of inflammatory cell diapedesis [46], and this may be reflected in the modifications of cellular events observed after BjV injection.
The results got with pre-treatment with indomethacin (cyclooxygenase pathway inhibitor) and celecoxib (COX-2-specific inhibitors) reinforce the importance of the participation of the cyclooxygenase route, COX-2 in mediating the leukocyte-endothelium interaction induced by BjV, since these drugs are quite effective in inhibiting the cellular events inducted by the venom, a significant decrease in adhered cells in the 2nd hour and migrated in the 24th hour was observed compared to the non-treated groups, injected with the venom.
These data corroborate the literature, where it has already been showed that the pre-treatment of animals with anti-inflammatory drugs, inhibitors of phospholipase A2 and the COX pathway, results in a significant reduction in paw edema induced by Bothrops venoms [23,24,25,32]. This participation was also observed in mice for cyclooxygenase pathway products and the isoform COX-2 [47].
A body of research shows that nitric oxide (NO) plays a part in controlling the interaction between white blood cells and endothelium. It is possible to see more of these cellular events by blocking certain forms of the enzymes' endothelial (eNOS) and inducible nitric oxide synthase (iNOS) [31,49]. A study found that NOS inhibitors that do not target specific genes, like L-NAME, do not stop the production of ICAM-1, a molecule that helps white blood cells stick firmly to endothelium when lipopolysaccharide (LPS) or carrageenan is present [49]. However, we found that cellular events in the cremaster microcirculation muscle of mice were significantly slowed down after they were pre-treated with L-NAME, a NOS inhibitor, and aminoguanidine, a selective iNOS isoform inhibitor. This shows that these substances play a role in the interaction between white blood cells and the endothelium.
When applied topically to the spermatic fascia of rats, the BjV induces changes in leukocyte-endothelium interaction [50]. However, when the rats had been treated with L-NAME first, the number of rolling and adherent cells decreased [51]. Bothrops atrox venom is also described as inducing serum NO levels [52]. It is known that cytokines like INF-γ and TNF-α induce the expression of iNOS and that Bothrops jararaca venom can induce the release of both these cytokines and NO [53].
Different experimental conditions, administration routes, or inflammatory stimuli can determine opposing effects on NO expression, suggesting that this mediator often acts with an antagonistic action in relation to the inflammatory response [51,54].
As previously mentioned, the inflammatory reaction observed in BjV is because of the rapid action of the toxins in the venom and due to the rapid release of endogenous mediators. Therefore, we used other drugs to understand the mediators involved in the changes in microcirculation after Bothrops envenomation.
After injecting the animals with venom, the alterations in leukocyte-endothelium interaction did not change when promethazine, a histamine inhibitor, was given first. This suggests that this amine does not play a role in the interaction between leukocytes and endothelium caused by Bothrops jararaca venom. Studies show that this mediator does not also play a role in mediating the paw edema that this venom causes in mice [28,33]. In rats, the participation of histamine in mediating this edema has been verified [23].
In animals that had first received methysergide, there were no changes in the parameters of the leukocyte-endothelium interaction due to BjV. This also suggests that serotonin is not involved in this process.
Leukotrienes interact with neutrophil receptors on the leukocyte surface, which leads to chemotaxis and increased adhesion to endothelial cells, mainly through increased expression of β2 integrin [55]. But giving animals NDGA, which blocks the lipoxygenase pathway, did not stop the release of leukotrienes. This suggests that the products of the lipoxygenase pathway are not involved in the BjV-caused mediation process. On the other hand, products derived from the lipoxygenase pathway participate in the venom-induced migration of B. erythromelas and B. alternatus [44].
Pre- and post-treatment with BAV inhibited the formation of BjV-induced edema in mice [25]. However, this inhibition was not observed in the group that received BAV 45 minutes after the BjV injection. This result shows that BAV neutralizes the formation of edema; however, this reduction is more efficient when it is administered soon after envenomation [25]. Picolo et al. [56] noted the same result and showed that BAV caused a decrease in edema just when administered before injecting Bothrops jararaca venom into the rats' footpads.
Some studies suggest that the reason immunoglobulin therapy does not work to stop local lesions caused by Bothrops venoms is because it is hard for the immunoglobulin to get to the site of the lesion [57]. This would justify using only the Fab portion of total IgG in the treatment of snake envenomation [58]. There is, however, no significant difference in the reduction of local lesions like edema, hemorrhage, and myonecrosis between animals treated with all three types of serum. This is true whether the serums contained total IgG, only the F (ab')2 portion, or even the Fab portion [59,60].
In our data, we showed that the use of dexamethasone was beneficial in inhibiting the cellular events of the leukocyte-endothelium interaction induced by BjV, not only compared to the control group but also when compared to the groups that received only treatment with BAV. The same decreases were observed in animals treated with the combination of dexamethasone and BAV, like what was observed in animals treated with dexamethasone alone at both times studied (Figure 1 and Figure 9). When injected directly into a vein without being diluted, BAV can increase the interaction between white blood cells and the endothelium. Antivenin contains the preservative phenol, primarily responsible for this effect [26].
This association between dexamethasone and BAV accelerated edema regression and reduced muscle damage from B. jararaca and B. jararacussu venoms [29], as well as swelling caused by B. jararaca venom [25]. When B. atrox venom is the cause of tissue damage, the combination of this corticosteroid with antivenom is also beneficial, reducing inflammation and accelerating muscle regeneration [21].
A recent study on people who had been envenomed showed that giving anti-inflammatory drugs along with antivenom reduced the local effects of snakebites, including inflammatory symptoms [61]. However, treatment with dexamethasone, alone or combined with antivenom, could not prevent, or reduce hemorrhagic damage induced by this venom [21,32]. This outcome is in line with the hypothesis that eicosanoids are not involved in the pharmacological mechanisms causing local hemorrhage brought on by Bothrops jararaca venom.
It is known that BjV does not induce the secretion of endogenous corticosteroids or stimulation of the hypothalamic-pituitary-adrenal axis [62]. In this sense, the use of dexamethasone would be important to avoid inflammatory reactions; however, this corticosteroid does not alter the local hemorrhagic and coagulant activities of BjV and does not indicate its use as a replacement for antivenom. When dexamethasone is associated with BAV, it may help treat Bothrops jararaca envenomation better by lowering the inflammatory response (swelling and leukocyte influx) faster among people whom this snake has bitten.
The inefficiency attributed to the antivenin described above encourages studies of complementary therapies. In this sense, the association of anti-inflammatory drugs with serum in treating local inflammatory reactions present in Bothrops jararaca envenomation would be a rational alternative, which should be clinically tested.
Author Contributions
Conceptualization: B.C.Z., L.R.C.G.; Investigation: B.C.Z., Resources: B.C.Z., L.R.C.G; Supervision and Funding acquisition: L.R.C.G.; Writing original draft: B.C.Z., L.R.C.G.; Writing revision/editing: B.C.Z., L.R.C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação Butantan, BCZ were fellows from of Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- CAPES (process 308083/2013-0) and LRCG were research fellows of Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq (process 309392/2015-2).
Institutional Review Board Statement
All experimental procedures were conducted according to the ethical parameters proposed by the International Society of Toxinology and the Brazilian College of Experimental Animals and were approved by the Ethical Committee for the Use of Animals of Butantan Institute (nº 199/05), approval date (09/03/2005).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Conflicts of Interest
The authors have no conflict of interest regarding this work to declare.
References
- Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, Warrell DA. Snakebite envenoming. Nat Rev Dis Prim. 2017; 3: 17063. [CrossRef]
- Harrison RA, Hargreaves A, Wagstaff SC, Faragher B, Lalloo DG. Snake envenoming: A disease of poverty. PLoS Negl Trop Dis. 2009;3: e569. [CrossRef]
- Cruz LS, Vargas R, Lopes AA. Snakebite envenomation and death in the developing world. Ethn Dis. 2009 Spring;19: 1–5. (1 Suppl 1): S1-42-6. PMID: 19484874.
- Kasturiratne A, Wickremasinghe AR, De Silva N, Gunawardena NK, Pathmeswaran A, Premaratna R, et al. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Medicine. 2008. 11: e218. [CrossRef]
- World Health Organization (WHO). SEVENTY-FIRST WORLD HEALTH ASSEMBLY Resolution WHA71.5 on Addressing the burden of snakebite envenoming. 2018; 1: 24–26. Available in: http://apps.who.int/gb/ebwha/pdf_files/EB142/B142_R4-en.pdf.
- Wold Health Organization (WHO). Tenth Meeting of the Strategic and Technical Advisory Group on Neglected Tropical Diseases (STAG-NTD). 2017; 19. Available: http://www.who.int/about/licensing/copyright_form/en/index.html.
- Souza W de. Doenças negligenciadas. 1st ed. Souza W de, editor. Academia Brasileira de Ciências. Rio de Janeiro: Academia Brasileira de Ciências; 2010. [CrossRef]
- Chippaux JP. Epidemiology of envenomations by terrestrial venomous animals in Brazil based on case reporting: from obvious facts to contingencies. J Venom Anim Toxins Incl Trop Dis. 2015; 21: 13. [CrossRef]
- Araújo FAA, Santalúcia M, Cabral RF. Epidemiologia dos acidentes por animais peçonhentos. In: Cardoso JLC, França FOS, Fan HW, Málaque CMS, Haddad Jr V, editors. Animais peçonhentos no Brasil: Biologia, clínica e terapêutica dos acidentes. 1st ed. São Paulo: Sarvier; 2003. pp. 06-12.
- Rosenfeld G. Symptomatology, pathology, and treatment of snake bites in South America. In: Bücherl W, Buckley EE, editors. Venomous Animals and their Venoms. New York: Academic Press; 1971. pp. 345–384.
- Cardoso JLC, Fan HW, França FOS, Jorge MT, Leite RP, Nishioka SA, et al. Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bothrops jararaca) in São Paulo, Brazil. Q J Med. 1993; 86: 315–325.
- Marder VJ, Feinstein DI, Francis CW, Colman R. Consumptive thrombohemorrhagic disorders. In: Colman RW, Hirsh J, Marder VJ, Salzman EW, editors. Hemostasis and Thrombosis: Basic Principles and Clinical Practice. 3rd ed. Philadelphia: J.B. Lippincott Company; 1994. pp. 1023–1063.
- França FOS, Málaque CMS. Acidente botrópico. In: Cardoso JL, França FOS, Wen FH, Málaque CMS, Haddad Jr V, editors. Animais peçonhentos no Brasil: biologia, clínica e terapêutica dos acidentes. 1st ed. São Paulo: Sarvier; 2003. pp. 72–86.
- Ministério da Saúde do Brasil. Manual de diagnóstico e tratamento de acidentes por animais peçonhentos. Brasilia: FUNASA; 2001. p. 112.
- Moura-da Silva AM, Laing GD, Paine MJ, Dennison JM, Politi V, Cramptom JM, Theakston RD. Processingof pro-tumor necrosis factor-alpha by venom metalloproteinase: a hypothesis explaining local tissue damage follwing snakebite. Eur J Immunol. 1996; 26: 2000-5. [CrossRef]
- Ley, K, Laudanna, C, Cybulsky, MI, Nourshargh, S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nature reviews Immunology. 2007; 7: 678-89. [CrossRef]
- Gutiérrez JM, Rucavado A, Escalante T, Herrera C, Fernández J, Lomonte B, et al. Unresolved issues in the understanding of the pathogenesis of local tissue damage induced by snake venoms. Toxicon. 2018; 148: 123–131. [CrossRef]
- Gutiérrez JM. Comprendiendo los venenos de serpientes: 50 años de investigaciones en América Latina. Rev Biol Trop. http://creativecommons.org/licenses/by/3.0; 2002; 50:377–394.Available: http://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S0034-77442002000200002&lng=pt&nrm=iso&tlng=es.
- Williams D, Gutiérrez JM, Harrison R, Warrell DA, White J, Winkel KD, et al. The Global Snake Bite Initiative: an antidote for snake bite. Lancet. Elsevier Ltd.; 2010; 375(9708):89-91. [CrossRef]
- Warrell DA. Snake bite. Lancet [Internet]. 2010; 375(9708):77–88. Available in: https://linkinghub.elsevier.com/retrieve/pii/S0140673609617542.
- Santos Barreto GNL, de Oliveira SS, dos Anjos IV, Chalkidis H de M, Mourão RHV, da Silva AMM, et al. Experimental Bothrops atrox envenomation: Efficacy of antivenom therapy and the combination of Bothrops antivenom with dexamethasone. PLoS Negl Trop Dis.; 2017;11: e0005458. [CrossRef]
- Rang HP, Dale MM, Ritter JM, Moore PK. Farmacologia. 5 ed. Rio de Janeiro: Elsevier; 2004.
- Trebien HA, Calixto JB. Pharmacological evaluation of rat paw oedema induced by Bothrops jararaca venom. Agents Actions. 1989; 26(3-4): 292-300. [CrossRef]
- Barbosa AM, Do Amaral RO, Teixeira CF, Hyslop S, Cogo JC. Pharmacological characterization of mouse hind paw oedema induced by Bothrops insularis (jararaca ilhoa) snake venom. Toxicon. 2003; 42(5):515-23. [CrossRef]
- Araújo SD, De Souza A, Nunes FPB, Gonçalves LRC. Effect of dexamethasone associated with serum therapy on treatment of Bothrops jararaca venom-induced paw edema in mice. Inflamm Res. 2007; 56. [CrossRef]
- Zychar, B.C.; Castro, N.C.;Marcelino, J.R.;Gonçalves, L.R. Phenol used as a preservative in Bothrops antivenom induces impairment in leukocyte-endothelial interactions. Toxicon. 2008. 51:1151-57. [CrossRef]
- Zychar, BC; Dale, CS; Demarchi, DS; Gonçalves, LRC. Contribution of metalloproteases, serine proteases and phospholipases A2 to the inflammatory reaction induced by Bothrops jararaca crude venom in mice. Toxicon, 2010; 55:227-34. [CrossRef]
- Perales J, Amorim CZ, Rocha SLG, Domont GB, Moussatché H. Neutralization of the oedematogenic activity of Bothrops jararaca venom on the mouse paw by an antibothropic fraction isolated from opossum (Didelphis marsupialis) serum. Agents Actions. 1992;37: 250-9. [CrossRef]
- Patrão-Neto FC, Tomaz MA, Strauch MA, Monteiro-Machado M, Da Silva Rocha-Junior JR,Borges PA, et al. Dexamethasone antagonizes the in vivo myotoxic and inflammatory effects of Bothrops venoms. Toxicon. 2013; 69: 55–64. [CrossRef]
- Wallace JL. Distribution and expression of cyclooxygenases (COX) isoenzymes, their physiological roles, and the categorization of nonsteroidal anti-inflammatory drugs. Am J Med. 1999; 107:11-6. [CrossRef]
- Dal Secco D, Paron JA, Oliveira SHP, Ferreira SH, Silva JS, Cunha FQ. Neutrophil migration in inflammation: nitric oxide inhibits rolling, adhesion and induces apoptosis. Nitric Oxide. 2003; 9:153-64. [CrossRef]
- Gonçalves LR, Mariano M. Local haemorrhage induced by Bothrops jararaca venom: relationship to neurogenic inflammation. Mediat Inflamm. 2000; 9:101-7. [CrossRef]
- Baez S. An open cremaster muscle preparation for the study of blood vessels by in vivo microscopy. Microvasc Res. 1973; 5:384-94. [CrossRef]
- Zychar BC, Clissa PB, Carvalho E, Alves AS, Baldo C, Faquim-Mauro EL, Gonçalves LRC. Modulation of Adhesion Molecules Expression by Different Metalloproteases Isolated from Bothrops Snakes. Toxins, 2021; 13: 803. [CrossRef]
- Clissa PB, Laing GD, Theakston RD, Mota I, Taylor MJ, Moura-da-Silva AM. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on pro-inflammatory cytokines released by murine peritoneal adherent cells. Toxicon. 2001; 39(10):1567-73. [CrossRef]
- Zamuner SR, Zuliani JP, Fernandes CM, Gutiérrez JM, de Fátima Pereira Teixeira C. Inflammation induced by Bothrops asper venom: release of proinflammatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon. 2005 Dec 1; 46(7):806-13. [CrossRef]
- Clissa, PB, Lopes-Ferreira, M, Della-Casa, MS, Farsky, SHP, Moura-da Silva, AM. Importance of jararhagin disintegrin-like and cysteine-rich domains in the early events of local inflammatory response. Toxicon. 2006; 47: 591. [CrossRef]
- Fernandes, CM, Zamuner, SR, Zuliani, JP, Rucavado, A, Gutiérrez, JM, Teixeira, CFP. Inflammatory effects of BaP1 a metalloproteinase isolated from Bothrops asper snake venom: Leukocyte recruitment and release of cytokines. Toxicon. 2006; 47: 549. [CrossRef]
- Zychar BC, Clissa PB, Carvalho E, Baldo C, Gonçalves LRC. Leukocyte recruitment induced by snake venom metalloproteinases: Role of the catalytic domain. Biochem Biophys Res Commun. 2020 Jan 8; 521(2):402-407. Epub 2019 Oct 25. [CrossRef]
- Búrigo AC, Calixto JB, Medeiros YS. Pharmacological profile of rat pleurisy induced by Bothrops jararaca venom. J Pharm Pharmacol. 1996; 48(1):106-11. [CrossRef]
- Ptzalis C, Pipitone N, Perretti M. Regulation of Leukocyte-Endothelial Interactions by Glucocorticoids. Annals of the N.Y. Acad Sci. 2002; 966:108-18. [CrossRef]
- Czock D, Keller F, Rasche FM, Haussler U. Pharmacokinetics and pharmacodynamics of systemically administered glucocorticoids. Clin Pharmacokinet. 2005; 44(1):61-98. [CrossRef]
- Tailor A, Tomlinson A, Salas A, Panés J, Granger DN, Flower RJ, Perretti M. Dexamethasone inhibition of leukocyte adhesion to rat mesenteric postcapillary venules: role of intercellular adhesion molecule 1 and KC. Gut. 1999; 45:705-12. [CrossRef]
- Flores CA, Zappellini A, Prado-Franceschi J. Lipoxygenase-derived mediators may be involved in in vivo neutrophil migration induced by Bothrops erythromelas and Bothrops alternatus venoms. Toxicon. 1993;31(12):1551-9. [CrossRef]
- Zamuner SR, Teixeira CF. Cell adhesion molecules involved in the leukocyte recruitment induced by venom of the snake Bothrops jararaca. Med Inflamm. 2002;11(6):351-7. [CrossRef]
- Rampart M. Neutrophil-Endothelial Cell Interations. In: Brain SD. The Handbook of Immunopharmacology. Immunopharmacology of Microcirculation. San Diego: Academic Press; 1994.
- Olivo RA, Teixeira CFP, Wallace JL, Gutiérrez JM, Zamuner SR. Role of cyclooxigenases in oedema-forming activity of bothropic venoms. Toxicon. 2007;49: 670-7. [CrossRef]
- Kubes P, Suzuki M, Granger DN. Nitric Oxide: An endogenous modulator of leukocyte adhesion. Proc Natl Acad Sci USA. 1991; 88:4651-55. [CrossRef]
- Dal Secco D, Moreira AP, Freitas A, Silva JS, Rossi MA, Ferreira SH, Cunha FQ. Nitric oxide neutrophil migration by a mechanism dependent on ICAM-1: Role of soluble guanylate cyclase. Nitric Oxide. 2006; 15:77-86. [CrossRef]
- Farsky SH, Goncalves LRC, Cury Y. Characterization of local tissue damage evoked by Bothrops jararaca venom in the rat connective tissue microcirculation: an intravital microscopic study. Toxicon. 1999; 37(7):1079-83. [CrossRef]
- Farsky SH, Borelli P, Fock RA, Proto SZ, Ferreira Jr JMC, Mello SBV. Chronic blockade of nitric oxide biosynthasis in rats: effect on leukocyte endothelial interaction and on leukocyte recrutaiment. Inflamm Res. 2004; 53:442-52. [CrossRef]
- Barros SF, Friedlanskaia I, Petricevich VL, Kipini TL. Local inflammation, lethality and citokine release in mice injected with Bothrops atrox venom. Med Inflamm. 1998; 7:339-46. [CrossRef]
- Petricevich VL, Teixeira CF, Tambourgi DV, Gutiérrez JM. Increments in serum cytokine and nitric oxide levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxicon. 2000; 38:1253-66. [CrossRef]
- Geffner JR, Analía ST, D’Elia I, Diament M, Klein D, Giordano M. Involvement of nitric oxide in the regulation of peripheral blood leukocyte counts. J Leuk Biol. 1995; 58:391-4. [CrossRef]
- Palmblad J, Lindstron P, Lemer R. Leukotriene B4 induced hyperadhesiveness of endothelial cells for neutrophils. Biochem Biophys Res Commun. 1990;166(2) 848-51. [CrossRef]
- Picolo G, Chacur M, Gutiérrez JM, Teixeira CF, Cury Y. Evaluation of antivenoms in the neutralization of hyperalgesia and edema induced by Bothrops jararaca and Bothrops asper snake venoms. Braz J Med Biol Res. 2002; 35(10):1221-8. [CrossRef]
- Battellino C, Piazza R, Da Silva AMM, Cury Y, Farsky SHP. Assessment of efficacy of bothropic antivenom therapy on microcirculatory effects induced by Bothrops jararaca snake venom. Toxicon. 2003; 41(5):583-93. [CrossRef]
- Landon J, Smith DC. Development of novel antivenoms based on specific ovine Fab. In: Bon C, Goyffon M, editors. Evenomings and their treatments. Lyon: Editions Foudation Maecel Mérieux; 1995. p. 173.
- Gutiérrez JM, Leon G, Rojas G, Lomonte B, Rucavado A, Chaves F. Neutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon. 1998; 36(11):1529-38. [CrossRef]
- Chaves F, Loría GD, Salazar A, Gutiérrez JM. Intramuscular administration of antivenoms in experimental envenomation by Bothrops asper: comparison between Fab and IgG. Toxicon. 2003; 41:237-44. [CrossRef]
- Dorooshi G, Javid ZN, Meamar R, Farjzadegan Z, Nasri M, and Eizadi-Mood N. Evaluation of the effects of Anti-Inflammatory drugs on local and systemic manifestations of snakebite: A cross-sectional study. J Venom Res. 2021; 25; 11:21-25. eCollection 2021. PMID: 34123361; PMCID: PMC8169030.
- Cury Y, Teixeia CF, Farsky SH. Lack of effect of endogenous corticosteroids on the acute inflammatory reaction (edema) induced by Bothrops jararaca venom (BjV) in rats. Toxicon. 1997; 35(5):773-6. [CrossRef]
Figure 1.
Effect of pre-treatment with dexamethasone (Dx) on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg) or saline (S) 1 hour before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
Figure 1.
Effect of pre-treatment with dexamethasone (Dx) on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg) or saline (S) 1 hour before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
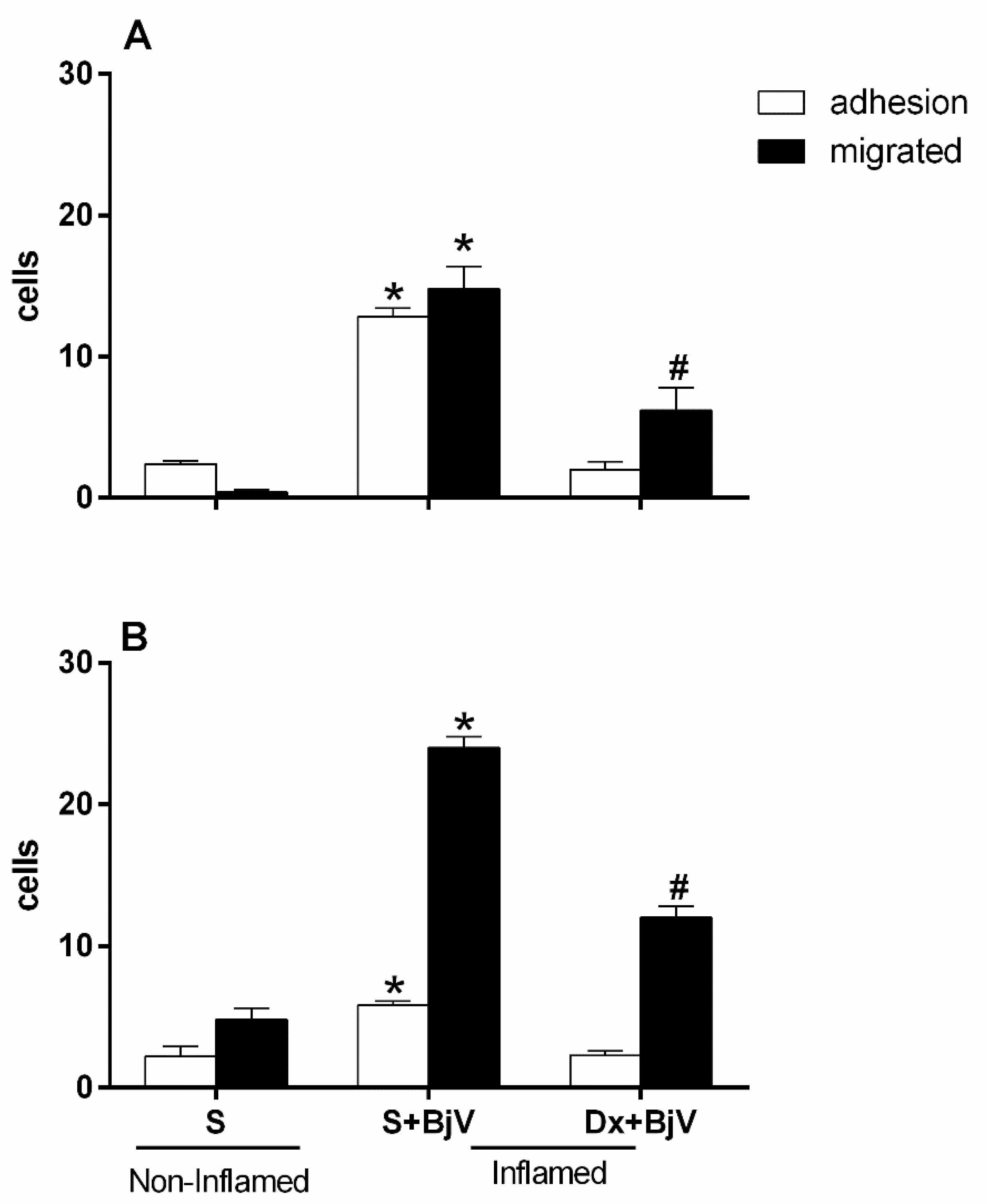
Figure 2.
Effect of pretreatment with indomethacin (Indo) on the interaction between white blood cells and endothelium in postcapillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with indomethacin (Indo, 3 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
Figure 2.
Effect of pretreatment with indomethacin (Indo) on the interaction between white blood cells and endothelium in postcapillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with indomethacin (Indo, 3 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
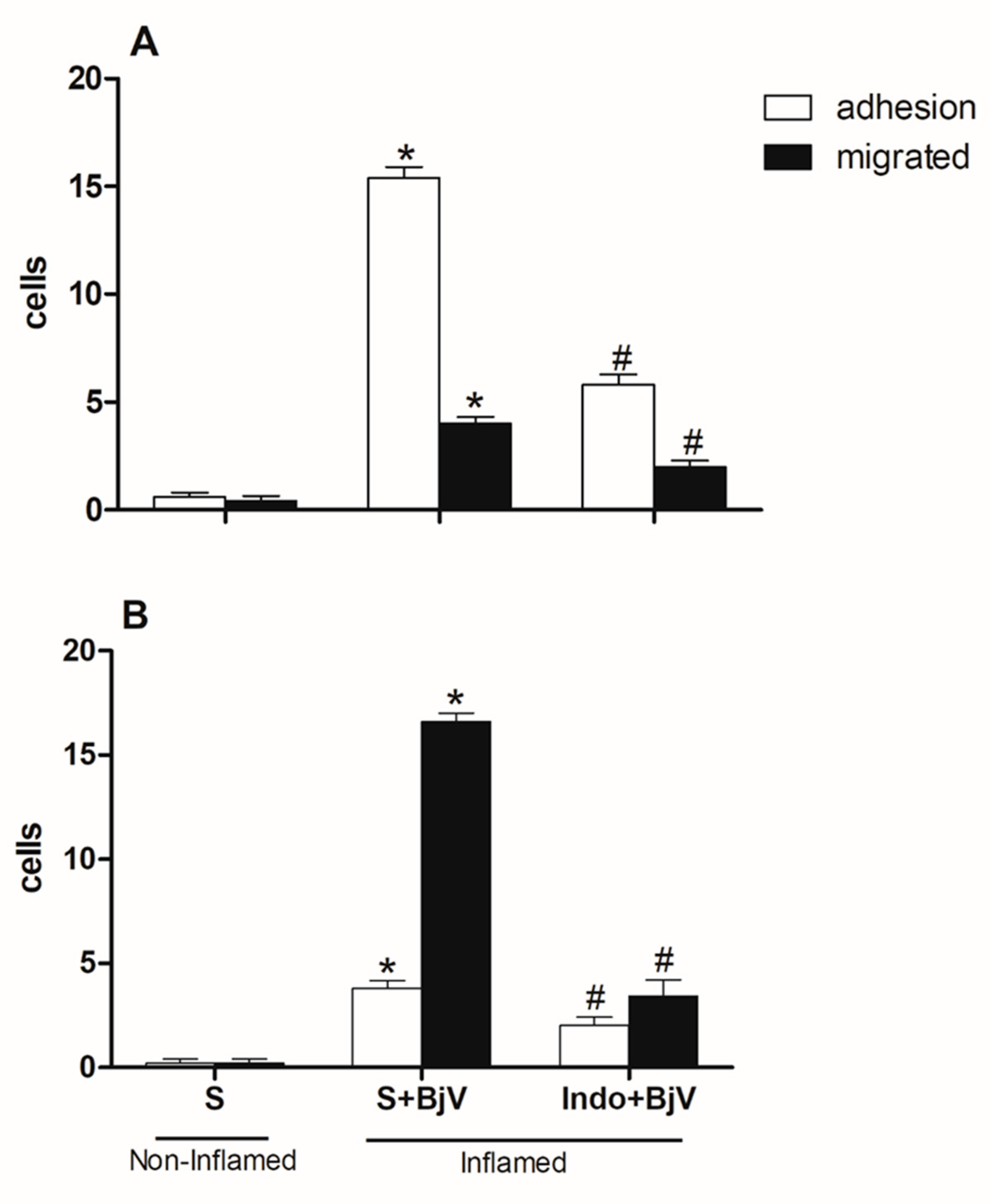
Figure 3.
The impact of pre-treatment with celecoxib on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with celecoxib (30 mg/kg) or carboxymethyl cellulose (CMC) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
Figure 3.
The impact of pre-treatment with celecoxib on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with celecoxib (30 mg/kg) or carboxymethyl cellulose (CMC) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
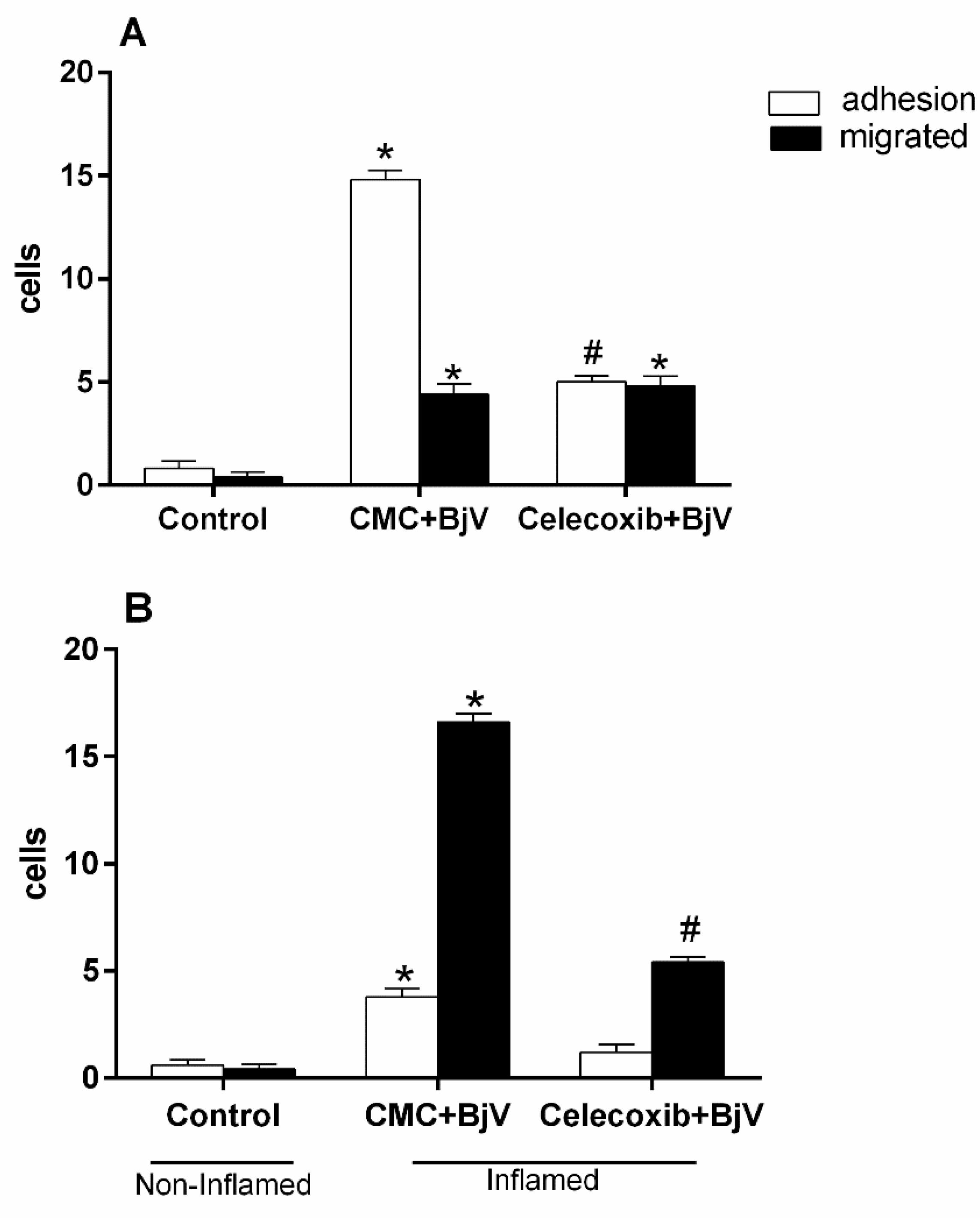
Figure 4.
The impact of pretreatment with L-NAME on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with L-NAME (100 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group and the group injected with BjV.
Figure 4.
The impact of pretreatment with L-NAME on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with L-NAME (100 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group and the group injected with BjV.
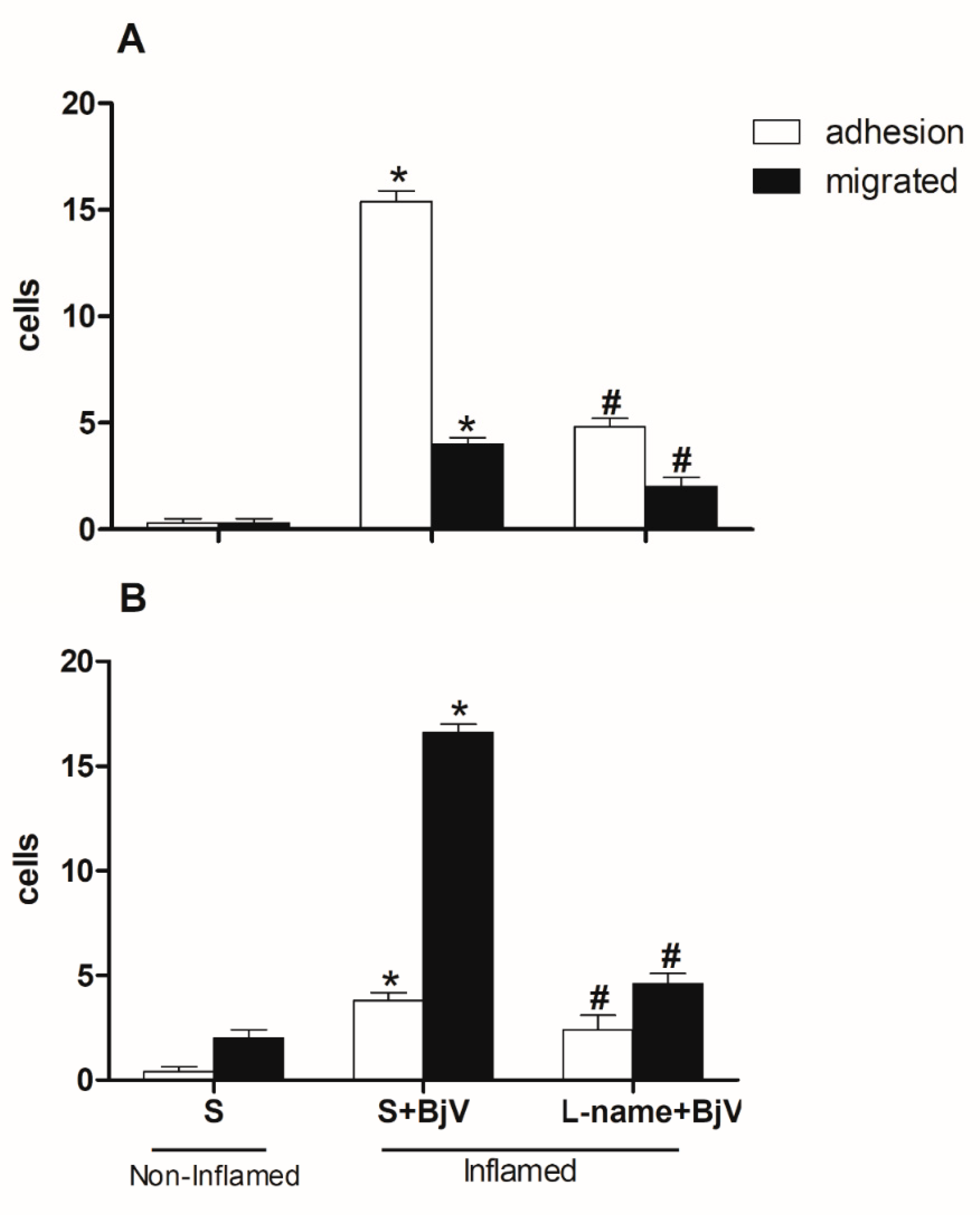
Figure 5.
Effect of pre-treatment with aminoguanidine (Amino) on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with aminoguanidine (Amino, 50 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
Figure 5.
Effect of pre-treatment with aminoguanidine (Amino) on the interaction between white blood cells and endothelium in post-capillary venules in the microcirculation of mouse cremaster muscle triggered by BjV. The animals received treatment with aminoguanidine (Amino, 50 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group, and the group injected with BjV.
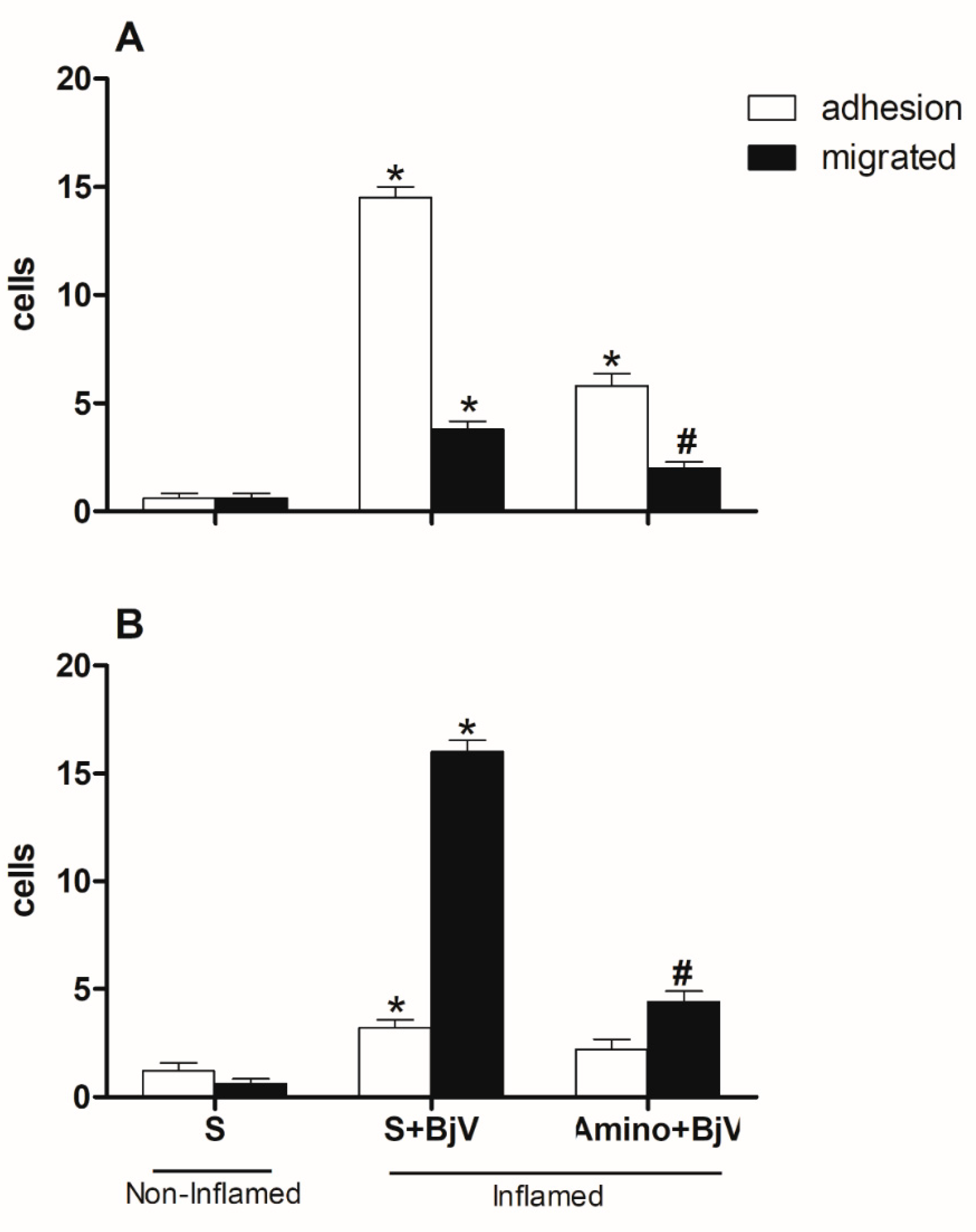
Figure 6.
Effect of pre-treatment with promethazine (Pro) on the leukocyte-endothelium interaction in post-capillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with promethazine (Pro, 10 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group.
Figure 6.
Effect of pre-treatment with promethazine (Pro) on the leukocyte-endothelium interaction in post-capillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with promethazine (Pro, 10 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group.
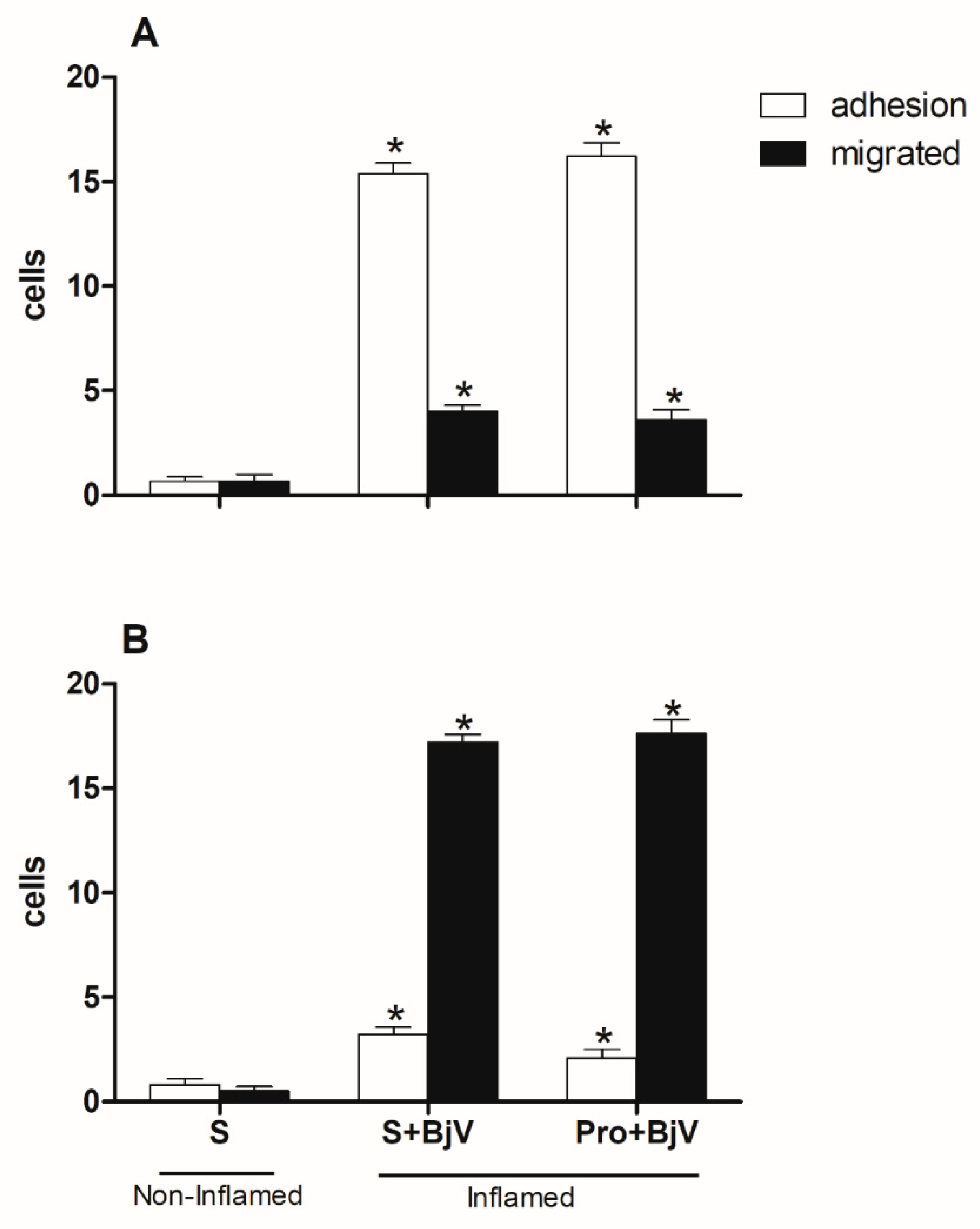
Figure 7.
Effect of pre-treatment with methysergide (Met) on the leukocyte-endothelium interaction in post-capillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with methysergide (Met, 0.8 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group.
Figure 7.
Effect of pre-treatment with methysergide (Met) on the leukocyte-endothelium interaction in post-capillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with methysergide (Met, 0.8 mg/kg) or saline (S) 30 minutes before the injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of BjV injection, the cremaster muscle was exposed, and intravital microscopy was used to look at how cells adhered and moved. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group.
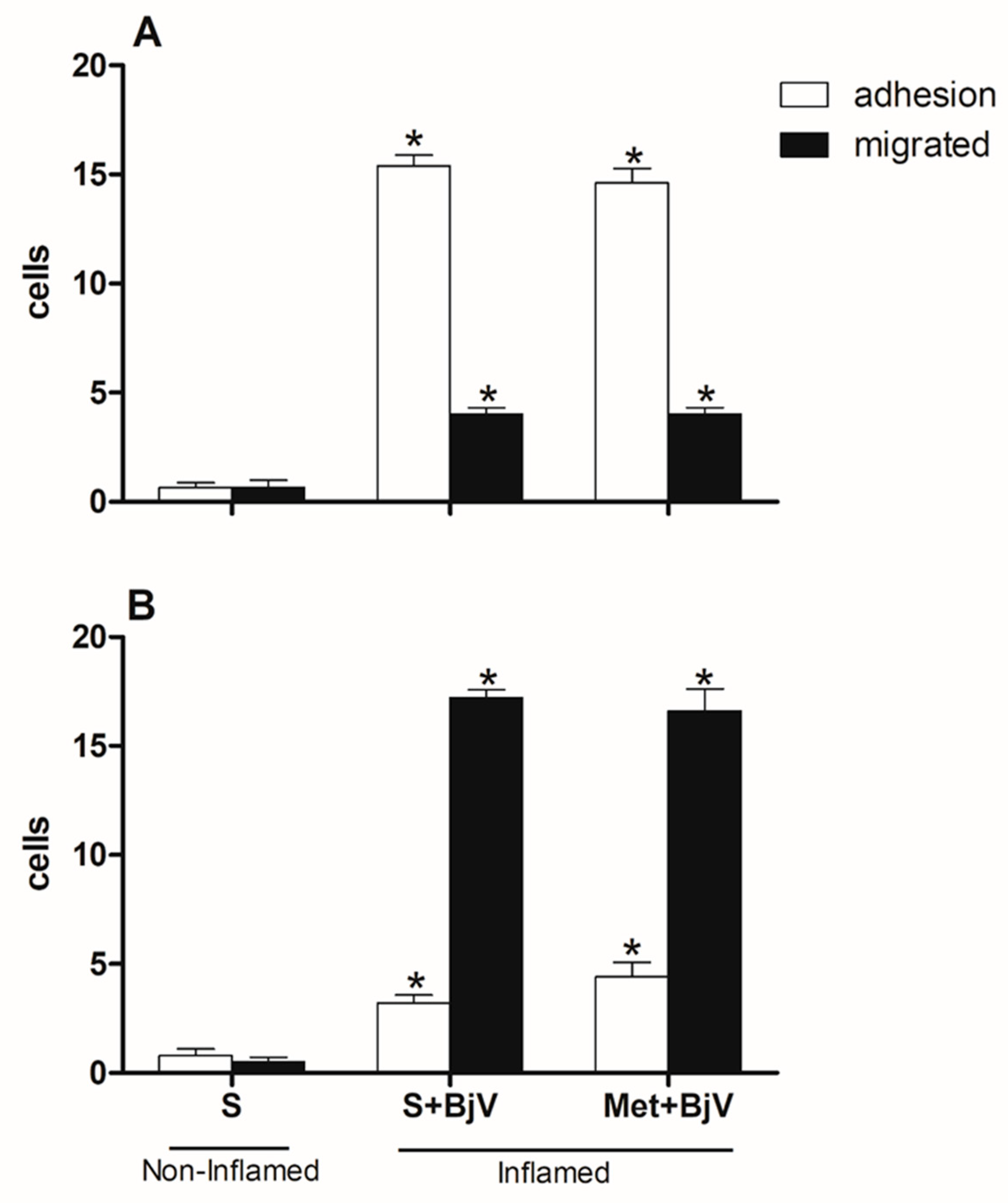
Figure 8.
Effect of pretreatment with nordihydroguaiaretic acid (NDGA) on the leukocyte-endothelium interaction in postcapillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with NDGA (30 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. The cremaster muscle was exposed 2 hours (A) or 24 hours (B) after being injected with BjV, and intravital microscopy was used to look at cellular events like adhesion and migration. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group and the group injected with BjV.
Figure 8.
Effect of pretreatment with nordihydroguaiaretic acid (NDGA) on the leukocyte-endothelium interaction in postcapillary venules in the microcirculation of mouse cremaster muscle induced by BjV. The animals received treatment with NDGA (30 mg/kg) or saline (S) 30 minutes before the injection of BjV (1 μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. The cremaster muscle was exposed 2 hours (A) or 24 hours (B) after being injected with BjV, and intravital microscopy was used to look at cellular events like adhesion and migration. The results are presented as the means ± S.E.M. (n = 5). *p < 0.05 compared to the control group. #p<0.05 compared to the control group and the group injected with BjV.
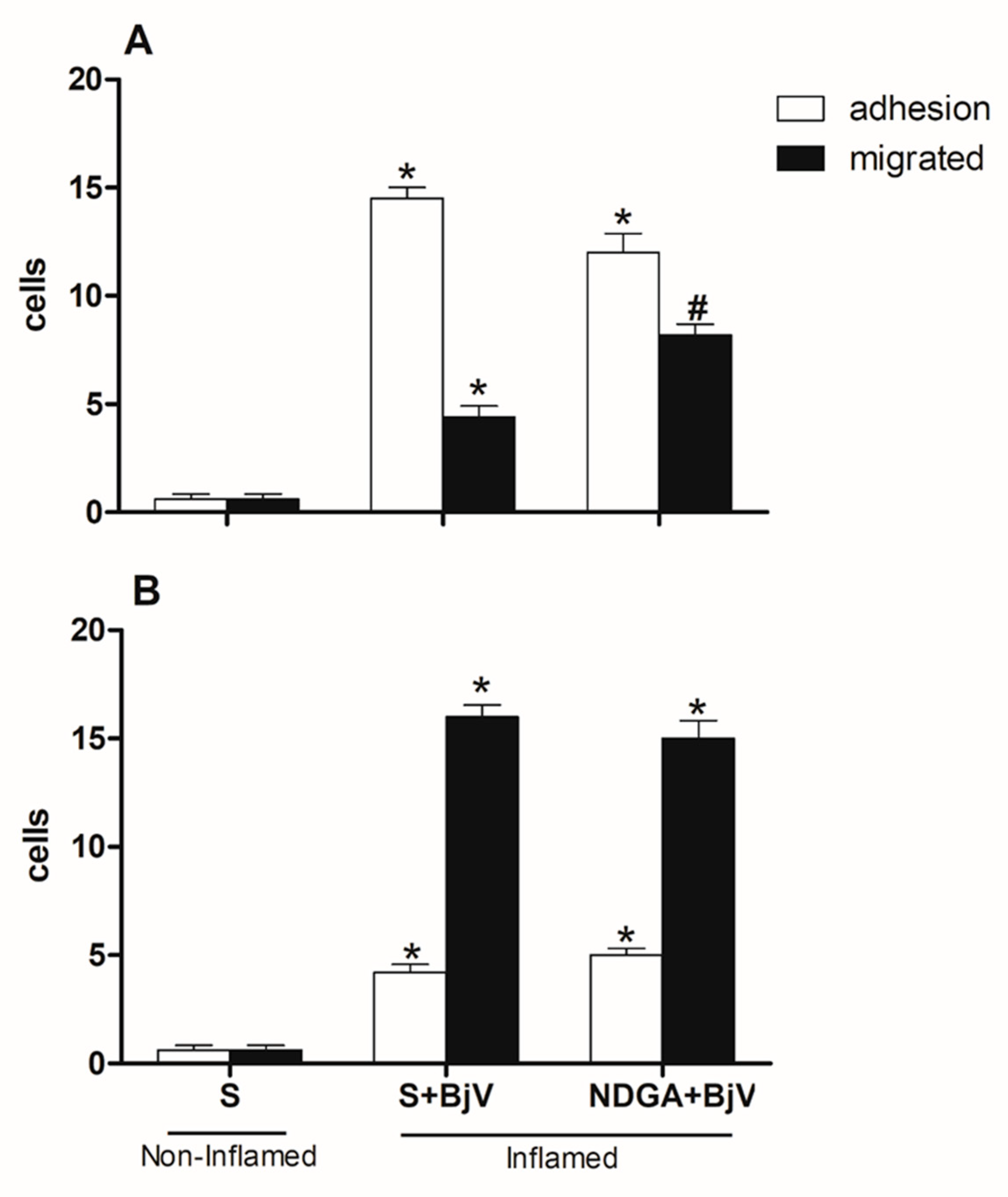
Figure 9.
Effect of post-treatment with dexamethasone (Dx) on leukocyte-endothelium interaction in post-capillary venules in the muscular microcirculation of the mouse cremaster induced by BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg) or saline (S) 1 hour after injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of envenomation, the cremaster muscle was exposed, and intravital microscopy was used to look at cellular events like adhesion and migration. Results are presented as means ± SEM. (n = 5). *p< 0.05 compared to the control group; #p<0.05 compared to other groups.
Figure 9.
Effect of post-treatment with dexamethasone (Dx) on leukocyte-endothelium interaction in post-capillary venules in the muscular microcirculation of the mouse cremaster induced by BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg) or saline (S) 1 hour after injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of envenomation, the cremaster muscle was exposed, and intravital microscopy was used to look at cellular events like adhesion and migration. Results are presented as means ± SEM. (n = 5). *p< 0.05 compared to the control group; #p<0.05 compared to other groups.
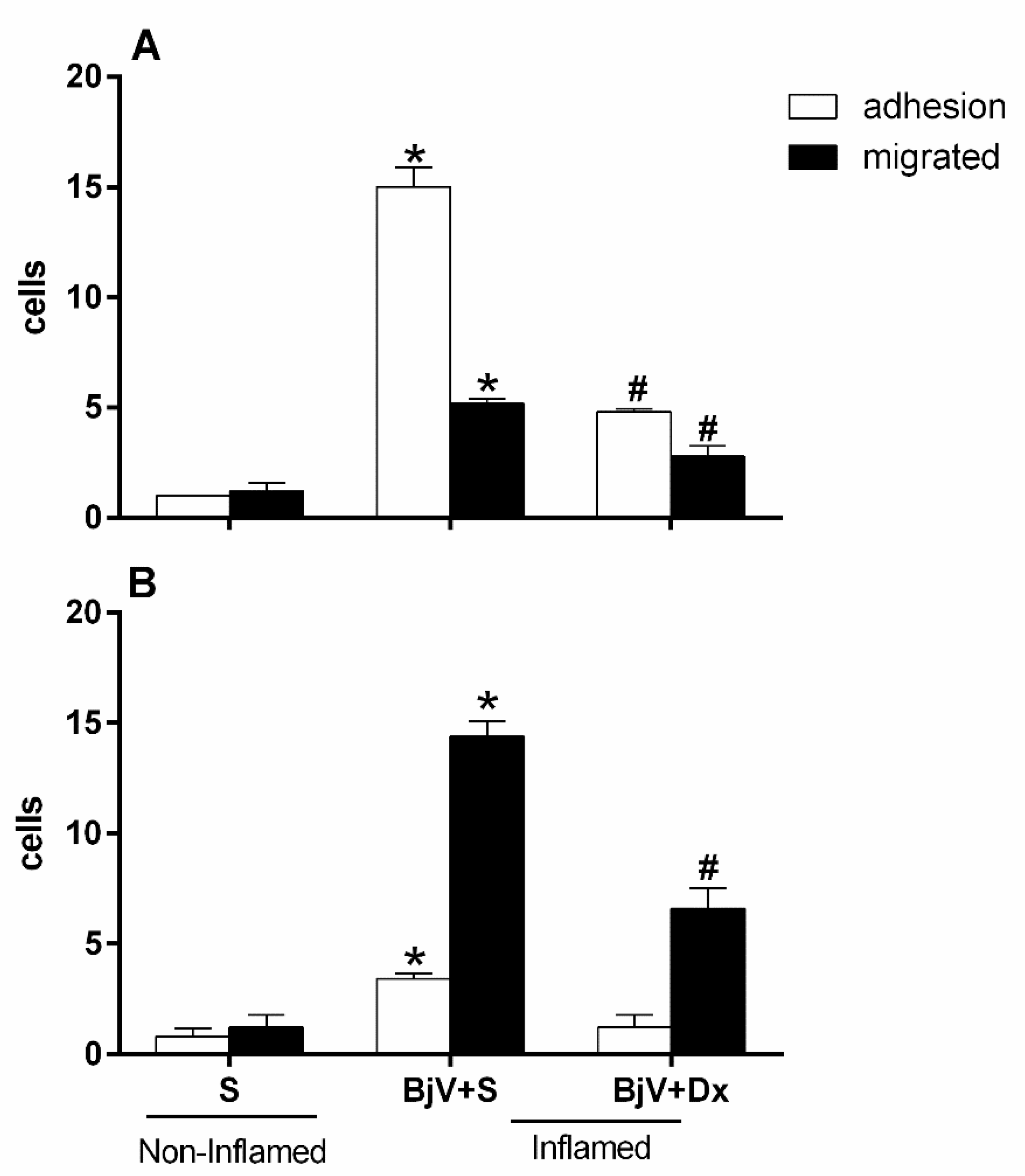
Figure 10.
Effect of combining treatment with dexamethasone and Bothrops antivenom (BAV) on the interaction between endothelium and white blood cells in the cremaster muscle post-capillary venules of mice injected with BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg i.p.), BAV (200 μL, i.v.) or saline (i.v. or i.p.), 1h after injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of envenomation, the cremaster muscle was exposed, and intravital microscopy was used to look at cellular events like adhesion and migration. The results are presented as the means ± S.E.M. (n = 5). *p< 0.05 compared to the control group; #p<0.05 compared to the other groups.
Figure 10.
Effect of combining treatment with dexamethasone and Bothrops antivenom (BAV) on the interaction between endothelium and white blood cells in the cremaster muscle post-capillary venules of mice injected with BjV. The animals received treatment with dexamethasone (Dx, 1.0 mg/kg i.p.), BAV (200 μL, i.v.) or saline (i.v. or i.p.), 1h after injection of BjV (1μg/100 μL) or saline (S) into the subcutaneous tissue of the scrotum. After 2 hours (A) or 24 hours (B) of envenomation, the cremaster muscle was exposed, and intravital microscopy was used to look at cellular events like adhesion and migration. The results are presented as the means ± S.E.M. (n = 5). *p< 0.05 compared to the control group; #p<0.05 compared to the other groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



