Submitted:
26 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Plastid-encoded RNA polymerase (PEP) forms a multisubunit complex in operating chloroplasts, where PEP subunits and a sigma factor are tightly associated with 12 additional nuclear-encoded proteins. Mutants with disrupted genes encoding PEP-associated proteins (PAPs) provide unique tools for deciphering mutual relationships among phytohormones. A block of chloroplast biogenesis in Arabidopsis pap mutants specifying highly altered metabolism in white tissues induced dramatic fluctuations in the content of major phytohormones and their metabolic genes, whereas hormone signaling circuits mostly remained functional. Reprogramming of the expression of biosynthetic and metabolic genes contributed to a greatly increased content of salicylic acid (SA) and a concomitant decrease in 1-aminocyclopropane-1-carboxylic acid (ACC) and oxophytodienoic acid (OPDA), precursors of ethylene and jasmonic acid, respectively, in parallel to reduced levels of abscisic acid (ABA). The lack of differences in the free levels of indole-3-acetic acid (IAA) between the pap mutants and wild-type plants was accompanied by fluctuations in the contents of IAA precursors and conjugated forms as well as multilayered changes in the expression of IAA metabolic genes. Along with cytokinin (CK) overproduction, all of these compensatory changes aim to balance plant growth and defense systems to ensure viability under highly modulated conditions.
Keywords:
Arabidopsis thaliana
; phytohormones
; gene expression
; PEP-associated proteins
; mutants
1. Introduction
The regulation of organellar gene expression in plants is principally performed by proteins encoded by nuclear genes [1]. Of these, components of the plastid transcription machinery play a fundamental role in fine-tuned regulation of the activity of the plastid genome to adjust it to developmental and environmental changes. This phenomenon is especially relevant for the transcriptional response to phytohormones, which harmonize the expression of nuclear and plastid genomes under various challenges.
In Arabidopsis, the specific changes in plastid genome expression are governed by three plastid RNA polymerases. Two monomeric nuclear-encoded phage-type RNA polymerases (NEPs), RPOTp and RPOTmp, are mainly responsible for the transcription of housekeeping genes. The third one, plastid-encoded eubacterial type RNA polymerase (PEP), transcribes photosynthesis genes. PEP activity is regulated by 6 nuclear-encoded sigma transcription factors and requires a set of PEP-associated proteins (PAPs) essential for the proper assembly and/or stability of the PEP complex [2]. Research over the last decade has revealed that PAPs are highly diverse in terms of their activities and structure and contain functional groups involved in DNA/RNA metabolism, redox regulation and ROS protection [3]. However, independent of their function, PAPs exhibit a high degree of coexpression and likely generate a regulon. Inactivation of any PAP gene results in an albino or ivory phenotype accompanied by elevated NEP activity and increased expression of NEP-dependent chloroplast genes [4]. In the light, pap mutants develop a normal photomorphogenetic phenotype and plastids without thylakoid formation [5]. A block of chloroplast biogenesis generates severe stress induced by dysregulation of the plant defense system and has a broad impact on hormone signaling circuits [6]. Therefore, PAPs may be an exclusive tool for dissecting interactions between chloroplast biogenesis and phytohormone pathways. Their interplay, in turn, can modulate the expression of nuclear genes encoding proteins locations.
The role of phytohormones in this mutual exchange of information between chloroplasts and the nucleus is far from understood. A detailed examination revealed the differential effects of exogenously applied phytohormones on the transcript levels of genes related to the plastid transcription machinery [7]. Unlike CKs and IAA, which have stimulatory effects, ABA, methyl jasmonate and SA repress the transcript accumulation of PAP genes in wild-type, whereas no reproducible alterations are observed with gibberellic acid (GA3), brassinolide (BL) or the ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC). It should be noted, however, that the transcriptional responses to exogenous hormone applications depend on age and developmental stage and may be near saturation in wild-type (WT) plants.
Gene expression profiling of the pap7 mutant recently presented by Grübler et al. [6] revealed several hormone-related genes regulated by biogenic retrograde signals from plastids. Among them, two highly downregulated genes, LOX2 (lAt3G45140) and AOS (At5G42650), encode lipoxygenase and allene oxide synthase, respectively, key enzymes involved in the production of oxophytodienoic acid (OPDA), a precursor of peroxisomal jasmonic acid. On the other hand, the expression of ACO3 (AT1g12010) and CORI3 (AT4G23600), involved in ethylene biosynthesis, was upregulated and downregulated, respectively, indicating that disruption of PAP genes may have both negative and positive effects on hormone-related genes.
Recently, we showed that impaired expression of the PAP1 and PAP6 genes in Arabidopsis contributed to the transition of CK-dependent accumulation of chloroplast gene transcripts and transcripts of the genes for plastid transcription machinery from positive to negative, although the CK signaling in the mutants remained practically unperturbed [8]. The opposite response was due to hormone overproduction as a consequence of the constitutive overexpression of the genes involved in CK synthesis and degradation. Elevated CK content could be the result of a compensatory mechanism allowing plants to promote morphogenesis in the absence of regular photosynthesis.
Extensive crosstalk and signal integration among growth-regulating hormones imply that CK overproduction may affect the transcript levels of genes involved in the metabolism of other hormones. A complex network of such interactions suggests that long-term effects of the altered content of any hormone can induce a ‘‘domino effect,’’ resetting many systems within the plant [9]. Compensatory processes aimed at maintaining hormone homeostasis in pap mutants are supposed to launch rearrangements of transcriptional programs to ensure plant viability under highly modulated conditions. At least in part, these differences can be interpreted as a consequence of retrograde signaling from defective plastids.
In this work, we applied highly sensitive analysis of phytohormone content and qRT–PCR to examine the effect of pap mutations on plant hormonal status and possible implications for the expression of chloroplast and chloroplast-related nuclear genes.
2. Results
2.1. Disruption of pap genes contributes to altered hormone levels
We employed two mutants with disrupted PAP genes, pap1/ptac3 (NASC 639260) and pap6/fln1 (NASC 305140), which exhibit features typical of so-called “PAP syndrome” [5]. Both mutants had albino phenotypes (Figure 1A, 1B) with reduced PEP-dependent plastid transcription and were able to grow only in the presence of a carbon source. The PAP1 protein belongs to a group implicated in DNA/RNA metabolism. It contains a SAP DNA-binding domain and a PPR motif and was shown to bind in a light-dependent manner to the transcribed regions of all PEP-dependent photosynthesis and rRNA genes [10]. The fructokinase-like protein PAP6/FNL2 is a member of a functional group engaged in redox regulation. PAP6/FLN2 is a target of PAP10/TrxZ, a redox-active thioredoxin, with which it interacts via cysteine bonds to form disulfide bonds [11,12]. However, complementation analysis suggested that PAP6 and PAP10 are structural rather than functional components of the PEP complex [3].
Previous studies have indicated that disruption of PAP genes results in elevated levels of cytokinins and their precursors [8]. In contrast, the levels of the CK antagonist abscisic acid (ABA) were significantly reduced as shown by our experiments. Both mutants accumulated twofold less ABA than did the wild type and had greatly decreased levels of ABA metabolites, namely, phaseic acid (PA), 9-hydroxy-ABA (9OH-ABA) and dihydrophaseic acid (DPA), in pap6 (Table 1). Similar trends, even more highly enhanced, were observed for the levels of ACC, a precursor of ethylene in pap1. The mutants contained no more than 3-10% of the wild-type values.
On the other hand, the contents of benzoic acid (BzA) and notably of salicylic acid (SA) significantly increased. The pap1 mutant accumulated 3 times more SA, and the pap6 mutant accumulated 4 times more SA than did the wild type, which indicates the possible activation of SA biosynthesis. There were no significant differences in the levels of jasmonic acid (JA) or its bioactive metabolite JA-isoleucine (JA-Ile) between the wild type and the mutants. In addition, the content of the JA precursor cis-oxophytodienoic acid (cisOPDA) decreased, especially in pap6.
Analysis of the pap mutants revealed no differences in free IAA levels when compared with those of the wild type; however, the amounts of the IAA precursor indole-3-acetonitrile (IAN) and IAA metabolites, including IAA-glucose ester (IAA-GE), oxo-IAA, and oxo-IAA-glucose ester (OxIAA-GE), tended to increase, albeit not always significantly. The levels of naturally occurring phenylacetic acid (PAA) also increased in the mutants. However, the biological significance of PAA is not completely understood. In addition to its auxin activity, PAA has been hypothesized to regulate the effects of free IAA by inhibiting polar auxin transport [13]. Notably, the content of IAA-aspartate (IAA-Asp) was significantly lower in both pap1 and pap6 than in the WT. A decrease in IAA-Asp content in pap mutants suggested a bias toward enhanced accumulation of auxin storage forms, which are postulated to regulate auxin homeostasis in growth and development [14].
The expression of genes involved in hormone metabolism is altered in pap mutants
Since the levels of various hormones are altered in pap mutants compared to those in WT plants, it is conceivable that the expression of at least some of the genes involved in their synthesis and metabolism was altered in these mutants. However, the search for the particular genes responsible for the altered hormonal status of pap mutants is impeded by the complexity of the metabolism of some phytohormones coupled with the specific regulation of different members of hormone gene families. To determine whether the transcript abundance of hormone-related genes and their response to treatment with exogenous phytohormones are associated with the levels of the corresponding hormones, we examined the expression of the key biosynthesis genes. Our studies were based on generally accepted concepts of hormone biosynthetic and signaling pathways and, in part, on publicly available transcriptomic data for pap7, taking into account the coregulation paradigm of PAP genes [6].
2.1.1. JA and SA
Consistent with the previously reported data for pap7, a block of chloroplast biogenesis had a strong impact on two genes encoding key enzymes for OPDA production, -LOX2 and AOS. We performed a qRT‒PCR assay to determine whether these two genes exhibited the same expression patterns in the pap1 and pap7 mutants. According to our analysis, the expression of both genes was downregulated 3-5-fold compared to that in the WT (Figure 2A). In addition, steady-state levels of the JA marker gene Thi2.1 (At1g72260) and the JA and ET marker gene PDF1.2 (At5G44420), which encode the antimicrobial proteins thionin and defensin, respectively, strongly decreased in both the mutants and in pap7, as shown by the transcriptomic data.
OPDA production is tightly regulated by SA, which was shown to decrease the expression of the genes encoding the OPDA biosynthetic enzymes LOX2, AOC2, and AOS in Arabidopsis [15]. OPDA levels can in turn affect the transcriptional activity of SA-related genes. In pap mutants, the decrease in the expression of OPDA synthesis genes was associated with the absence of thylakoid lipids, precursors of allene oxide [6]; therefore, jasmonate appears to act upstream of salicylate (Figure 2B). As can be judged from the results of our experiments, elevated levels of SA in pap mutants corresponded to increased expression of the SA biosynthesis-related gene ICS1 (isorchorismate synthase 1), which encodes a crucial enzyme in the SA synthesis pathway that accounts for 90% of SA production [16]. Elevated transcript levels of ICS1 were accompanied by increased expression of the SA signaling gene NPR1 (Non-expressor of pathogenesis-related genes1), a master regulator of downstream SA signaling [17], and the SA marker gene PR1 (PATHOGENESIS-RELATED 1) [18]. These data support the antagonistic relationships between SA and OPDA, which are involved in plant defense responses.
2.1.2. IAA
Chronic SA overproduction can be associated with a concomitant reduction in auxin biosynthesis, transport, and signaling [19]. According to the transcriptomic data, several auxin-responsive genes, AT1G29430, AT4G12980, AT1G34310 (AXLIN RESPONSE FACTOR 12 and SAUR21), were repressed in pap7, with an expression change exceeding the threshold of 1 (log2). Downregulation of SAUR21, which encodes SMALL AUXIN UP RNA, was also confirmed in our experiments by qRT‒PCR for pap1 and 6 (Figure 2C). In addition, it has been reported that SA repression of auxin-related genes has no significant effect on free auxin levels [20]. This finding is consistent with the almost identical content of bioactive IAA in the WT and pap mutant strains detected in our studies (Table 1). In accord with this, the expression of the TAA and YUCCA genes, which encode major steps of IAA biosynthesis from tryptophan via the IPA (indole-3-pyruvic acid) pathway [21], did not significantly differ between pap7 and the WT.
It should be noted, however, that only a small fraction of auxin exists as free, active signaling molecules [13]. Therefore, altered levels of auxin precursors and conjugated forms may contribute to the maintenance of auxin homeostasis in pap mutants under conditions of arrested chloroplast biogenesis. In this regard, the decreased expression of SUR1 (SUPER ROOT1) is of interest. SUR1 encodes the enzyme involved in the conversion of the IAA precursor IAOx to indole glucosinolate (IGS) in the side branch of the IAOx pathway [21]. Inactivation of SUR1 disrupts IGS biosynthesis and promotes accumulation of the upstream intermediate IAOx, which was elevated in pap1 and 6 (Table 1).
In addition, the expression of Gretchen Hagen 3 (GH3.9), a member of the family of genes encoding amido synthases, was strongly downregulated in the pap mutants, which was in line with the significantly reduced levels of IAA-Asp (Figure 2C; Table 1). Amido synthases conjugate amino acids to carboxyl groups of small molecules, including auxin [22]. However, the dependence of this repression solely on auxin is ambiguous since the reduced expression of GH3.9 may also be related to SA or JA metabolism. Therefore, the influence of IAA production and its effects are multifaceted and aim to balance plant growth and defense systems through cross interactions with other plant hormones.
2.1.3. Ethylene and ABA
Next, we analyzed the expression of ethylene biosynthesis genes. Ethylene is produced from its general precursor S-adenosyl-L-methionine (SAM) by a two-step biosynthesis route. In the first step, SAM is converted to ACC and 5’-methylthioadenosine (MTA) by ACC synthase (ACS). In the second step, ACC is converted to ethylene, CO2 and cyanide by ACC oxidase [23]. The ACSs constitute a multigene family that is differentially expressed in plants. The transcript accumulation of two members of the ACS family, ACS4 and ACS8, was highly repressed in the pap1 and pap6 mutants, consistent with the drastically reduced ACC content (Figure 2D; Table 1). Both genes also displayed low expression levels in pap 7 (-1.0 log2-fold for ACS4 and -2.1 for ACS8) [6]. Along with these findings, the accumulation of ACO3 transcripts in pap7, as well as in pap1 and pap6, was twice as high as that in the wild type. This result may reflect the specific induction of ACO3 in response to a decrease in the level of ACC.
Among the multiple members of the ethylene response factor (ERF) gene family, ERF1 and ERF14 had more than a 50% decrease in the magnitude of change in expression in the pap mutants compared to the WT (Figure 2D). These two trans-factors are of particular interest because they were shown to function in ethylene/jasmonic acid plant defense responses [24]. In addition, ERF1 upregulated specific suites of genes in response to different abiotic stresses [25]. Since both ERF1 and ERF14 are key elements in the integration of ethylene and JA signals, their suppression in pap mutants, as well as impairment of ACC synthesis, may also be a consequence of a decrease in OPDA production.
The ethylene biosynthesis genes ACS4 and ACS8 were shown to be negatively regulated in Arabidopsis by ABA through ABI4-mediated transcriptional repression [26]. The expression of ABI4, as well as that of the ABA marker gene RD29, decreased in the pap mutants (Figure 2E). In parallel, among the genes of the NCED family that control the entire process of ABA biosynthesis by regulating the rate-limiting step [27], NCED4 was significantly downregulated in pap7 (-1.66 log2) [6]. According to our qRT‒PCR assays, in addition to NCED3 and ABA2, NCED4 was also strongly repressed. A decrease in NCED expression as well as upregulation of the ABA catabolism gene CYP707A1 could account for the reduced ABA levels, thus demonstrating a positive feedback interaction between ethylene, OPDA and ABA in the mutants.
In summary, arrest of plastid development induces reprogramming of the expression of hormone biosynthetic and metabolic genes in pap mutants, which in turn leads to concomitant defects in the content and likely interplay between various hormones.
2.2. Pap mutants exhibited partially altered responses to hormone treatments
Next, we analyzed whether the modified hormone status of pap mutants affects hormone signaling and response to hormone treatment. As previously indicated, the expression levels of the ABA marker RD29, the ethylene and JA marker PDF1,2 and the IAA marker IAA19 were lower in the pap mutants than in the WT plants. The relative induction of these genes exceeded that of the WT plants, indicating increased responsiveness of the mutants to exogeneous hormones (Figure 3A). In contrast, the expression of the SA marker gene PR1 was downregulated following the application of exogenous hormones in accordance with the activation of a positive feedback loop, diminishing the uncontrolled activation of hormone signaling. Overall, these results indicate that hormone signaling is still functional in pap mutants.
Several bioassays were used to assess the sensitivity of the strains to ACC, ABA, IAA, metJA and SA. For shoot elongation tests, the seedling were grown for 6 days in the dark in media supplemented with a range of hormone concentrations. Homozygous pap1 and pap6 hypocotyls were identified based on the white phenotype of the cotyledons after they had been cultivated for an additional 24 h under light. The hypocotyl lengths of the mutants did not significantly change relative to those of the WT plants after treatment with IAA, SA, ACC or metJA (Table S1). Therefore, at the early stages of development, the plants responded to hormonal treatment as did the wild-type plants. This finding is consistent with the observation that these mutations do not affect the skotomorphogenic program and that pap mutants develop normally within the first few days even without sugar [5].
Compared with those of Col-0, the roots of the pap1 and pap6 plants were significantly shorter (41.9±13.3% and 43.8±6.6%, respectively) (Figure 1B). In the presence of SA, ABA and IAA, the relative decreases in lengths were similar to those in the wild-type plants. However, when grown on plates containing various concentrations of ACC or metJA, both mutants exhibited considerably shortened roots compared with corresponding decreases in Col-0 (Figure 3B, 3C; Table S2). From these results, we conclude that the mutants were more sensitive to treatment with these hormones.
Since the mutants were deprived of chlorophyll, the ability of the tested hormones to retard or accelerate senescence in detached leaves was evaluated by changes in protein content. Under SA and IAA treatments, protein degradation did not significantly differ from that in water-treated samples for either the wild type or the mutants (Figure 3C, Table S3). ABA, ACC, and metJA contributed to an accelerated decrease in protein content, and the decrease was significantly greater in mutants following the application of metJA and ACC. Therefore, pap mutations affect the ability of ACC and metJA to accelerate leaf senescence.
It thus appears that hormones may play specific roles in various stages of development in pap mutants, participating in some aspects of developmental processes. At each stage, the physiological relevance of the altered sensitivity to hormones should be considered separately.
Our previous studies revealed the important regulatory role of plant phytohormones in regulating the expression of genes related to the chloroplast transcription machinery [7,28]. However, pap mutations did not substantially modify the response to hormone application. The transcript accumulation of NEP, SIG1-4,6 and operational PAP genes was upregulated by IAA but did not respond to or was slightly activated by ACC treatment (Figure 4, Table S4). The genes were repressed by metJA and in part by ABA. The exceptions were CKA4, which encodes the Ser/Thr protein kinase cPCK2, and stress-responsive SIG5, which were upregulated by ABA in both the WT and mutant plants, consistent with the upregulation of these genes by ABA shown by Wang et al. [29] and Yamburenko et al. [30]. In parallel, the latter two genes were downregulated by IAA, exhibiting a special type of hormonal regulation. SA treatment did not affect the expression of the genes involved in the chloroplast transcription machinery or contributed to their slight suppression.
Considering the highly reduced expression of genes with PEP promoters (class I) and their residual transcript levels, our studies were mainly focused on modulations of the expression of selective NEP-dependent (class III) and NEP- and PEP-dependent (class II) genes. The expression of the studied chloroplast-encoded genes was gene specific and in part divergent from that in the WT plants. In particular, under our experimental design, rpoB (the beta subunit of PEP) was significantly upregulated by IAA, while clpP (an ATP-dependent protease) and accD (an acetyl-CoA carboxylase) were repressed by metJA and ABA only in the mutants; these genes did not respond reliably to hormone treatment in the WT plants. These data confirmed the increased sensitivity of the mutants to IAA, metJA, and ABA. In both the WT and mutant strains, no reproducible changes were observed when the samples were exposed to SA or ACC treatment.
3. Discussion
Mutants with functionally inactivated PAPs provide unique tools for the dissection of hormone response networks in plants. The dramatic fluctuations in the contents of major phytohormones detected in our studies suggest that changes in hormone interplay occur mainly at the level of hormone metabolism, while signaling circuits mostly remain operational. The most marked differences between the Pap mutants and the wild type relate to the greatly increased SA content and the concomitant decreases in ACC and OPDA, which are precursors of ethylene and jasmonic acid, respectively, as well as a significant reduction in ABA levels. In addition, we previously detected CK overproduction as a result of constitutive overexpression of genes involved in the synthesis and degradation of CK [8].
The magnitude and direction of the changes in expression of hormone metabolism genes suggest that they are triggered by altered sink/source relationships induced by nonautotrophic metabolism in albino plants. A recent transcriptomic study of pap7 revealed that a block in chloroplast biogenesis had a significant impact on metabolic genes related to starvation processes and the mobilization of storage energy [6]. Thus, the transcriptomic data revealed the induction of several genes encoding proteins involved in sucrose degradation and transport and variably regulated fatty acid metabolism. In particular, a lack of linoleic acid in arrested plastids contributed to the downregulation of the allene oxide pathway and the production of OPDA, the precursor of jasmonic acid, which is highly important for the pathogen defense system. These results are also consistent with microarray data from the Arabidopsis immutans (im) mutant, which has green and white sectoring due to the action of the nuclear recessive gene IMMUTANS (IM). The expression of genes encoding key enzymes of the allene oxide pathway (AOS, AOC1, AOC4 and LOX2) was strongly repressed in the white sectors of the mutant (4-10-fold) compared to that in the WT leaves [31].
Severe stress generated by arrested plastids activates different protective systems in response to the highly altered metabolism of white tissues. SA, the production of which has been strongly increased in pap mutants, is known to play a central role in local and systemic acquired resistance against biotrophic pathogens, while the JA-mediated response coupled with the ethylene signaling pathway contributes to defense against necrotrophic pathogens [17]. The SA- and ethylene/JA-mediated defense pathways are mutually antagonistic, which likely accounts for the elevated SA levels in the mutants with reduced OPDA and ACC contents. Hence, we speculate that SA accumulation can be treated as a response to enhanced susceptibility to biotrophic pathogens such as Pseudomonas syringae. Similarly, increased vulnerability has been detected for the white sectors of the im mutant with reduced lignin and cellulose microfibrils, and alterations in galactomannans and the decoration of xyloglucan [32].
However, the involvement of SA in the regulation of genes associated with pathogenesis may not be the only factor determining its increased content. Recent evidence suggests that SA can act antagonistically to the inhibition of plastid biogenesis by promoting the accumulation of photosynthesis-associated proteins [33]. Exogenous SA partially restored the level of chlorophyll in norflurazon (NF)-treated Arabidopsis plants and in the plastid protein import2 (ppi2) mutant, which had reduced SA and JA contents and an albino phenotype, and in addition, it increased the levels of some photosynthesis-associated proteins. In accordance with the positive effect of SA on chloroplast biogenesis, light regulation of PhANGs (with the exception of LHCB genes) and other nuclear gene groups was fully functional in the pap7 mutant, indicating that a block in chloroplast biogenesis does not repress their expression [5]. Conversely, SA is known to promote senescence and decrease the level of chlorophyll [34]. These findings suggested that SA may play opposite roles in the regulation of chlorophyll content, which are hypothesized to be determined by specific signaling pathways activated by SA [33].
The role of SA in balancing stress responses and growth can be at least partially realized via interactions with auxin and CK. SA attenuates plant growth by regulating the biosynthesis, transport, and signaling of auxin [35], which is consistent with the downregulation of several IAA-related genes observed in our studies (SAUR21, GH3.9 and several auxin-responsive genes). On the other hand, elevated SA levels in pap mutants were superimposed on increased levels of certain auxin precursors and conjugated forms, as well as concomitant enhanced accumulation of CK metabolites and precursors [8]. The ability of CK to increase ICS1 expression was shown by Choi et al. [36]. In response to non-cytokinin-secreting pathogens, ICS1 was hyperactivated in the presence of CKs. Hence, SA-mediated responses are multilayered and involve both the reinforcement of defense mechanisms and the activation of plant growth through crosstalk with other plant hormones, such as auxin and CK.
An elevated content of CK precursors and metabolites in pap mutants may augment sink activity and promote morphogenesis in the absence of normal photosynthesis. The ability of CK to create new source‒sink relationships and increase the nutrient sink activity of plant tissues to support their growth was first described by Mothes [37,38]. We therefore hypothesize that alterations in the levels of growth-promoting hormones in pap mutants may be the result of sink demand in non-photosynthesizing tissues to optimize plant growth.
Along with the upregulation of genes encoding phytohormones with growth-promoting activity, genes encoding hormones with inhibitory effects were largely suppressed. This group, which includes ethylene-, ABA-, and JA-related genes, triggers phenotypic changes in response to biotic and abiotic stresses under a variety of specific environmental conditions. The downregulation of these genes and the parallel decrease in hormone content imply that uncontrolled activities of the aforementioned hormones may contribute to severe physiological perturbations in pap mutants and failure to vegetate even on Suc-supplemented media. This finding is consistent with the overall trend suggesting that crosstalk between growth-promoting and growth-repressing hormones often opposes each other.
The specific feedback loops regulating the activity of various hormones in pap mutants can be attributed to the impact of retrograde signals from arrested albino plastids. Plastidial control of nuclear-encoded hormone-associated genes is partly explained by the fact that plastids are the sites of synthesis of a number of hormones, including CK, ABA, and SA. Most of the genes identified in the “Hormone” subset of the pap7 mutant were shown to encode proteins with non-plastidial locations, which was ascribed to the broad impact of retrograde biogenic signals on the hormone signaling network [6].
Changes in the hormonal status of pap mutants can remodel the responses of plastid-encoded genes to hormone treatment from positive to negative and vice versa. Thus, the disruption of PAP genes contributed to the abolishment of the positive CK effect on the accumulation of chloroplast gene transcripts and transcripts of the genes for plastid transcription machinery [8]. However, SA treatment did not affect the accumulation of plastid gene transcripts, probably due to the optimal concentration of SA, whereas at least some chloroplast genes in pap mutants were found to be more sensitive to metJA and ABA, consistent with enhanced signaling responses to these hormones.
4. Materials and methods
4.1. Plant material, growth conditions and hormone treatment
Arabidopsis thaliana ecotype Columbia 0 was used in all the experiments. The pap1 (NASK ID N 639260) and pap6 (NASK ID N305140-45) mutants were obtained from the European Arabidopsis Stalk Center. Seeds were sown on MS media supplemented with 2% sucrose and grown in a controlled-environment chamber under a 16-h light and 8-h dark photoperiod at an illumination of 100 μmol·m-2·s-1 and a temperature of 23°C for 4 weeks. For hormone treatment, plants were sprayed with solution of hormones or an equal aliquot of ethanol and collected after 3 h of exposure. The compounds assayed were abscisic acid (ABA, 5x10-5 M), indole-3-acetic acid (IAA; auxin, 10-6 M), 1-aminocyclopropane-1-carboxylic acid (ACC; ethylene precursor, 10-5 M), salicylic acid (SA, 10-5 M) and methyl jasmonate (MetJA 5x10-5 M). The concentrations of the active reagents and treatment times were selected based on preliminary experiments. Leaves from at least six different plants were harvested for each probe to average the results, after which the probes were stored at -80°C.
4.2. Hormone Extraction, Purification, and Determination
Frozen samples (100 mg FW) were homogenized and extracted with cold (–20°C) methanol/water/formic acid (15/4/1, v/v/v) as described in Dobrev and Kaminek [39] and Dobrev and Vankova [40]. Phytohormones were separated with a reverse-phase–cation exchange SPE column (Oasis-MCX, Waters) into the acid fraction by elution with methanol and into the basic fraction by elution with 0.35 M NH4OH in 60% methanol, which were used for the determination of auxins, ABA, SA, jasmonates, and ACC. Data processing was performed with the Analyst 1.5 software package (Applied Biosystems, Waltham, MA, USA).
4.3. Quantitative RT‒PCR
Total RNA was isolated from 30-day-old rosette leaves using the TRIzol (Thermo Fisher Scientific, Waltham, MA, USA) method. RT‒PCR was performed using a LightCycler 96 (Roche, Roche, Basel, Switzerland). with hot start SYBR Green I technology and gene-specific primers according to the protocol described previously [41]. The primers used are listed in the Supporting Information section (Table S5). The following standard thermal profile was used for all PCRs: 95°C for 5 min; 40 cycles of 95°C for 15 s, 58°C for 15 s and 72°C for 25 s. UBQ10 (at5g53300) and protein phosphatase 2A (PP2A) regulatory subunit A2 (at3g25800) were used as internal controls. The primers used are presented in Table S5.
4.4. Growth sensitivity assays
For the hypocotyl elongation assay, seeds were grown in darkness for 4 days on MS media supplemented with various concentrations of phytohormones. Homozygous pap1 and pap6 plants were identified based on the white phenotype of the cotyledons after they had been cultivated for additional 24 h under light.
For the root elongation assay, 7-day-old seedlings were subsequently transferred to MS plates supplemented with various concentrations of hormones and grown vertically for another 4 days under 16 h light/8 h dark conditions. Measurements were performed in triplicate with 20 seedlings for each experiment.
A protein retention assay was performed with rosette leaves excised from the 3rd and 4th layers of 5-week-old plants. The leaves were placed on filters moistened with hormone solutions or water with a solvent and kept for 3 days in darkness. Total soluble protein concentration was determined by the bicinchoninic acid (BCA) method using a Pierce BCA assay kit (Thermo Fisher Scientific, Waltham, MA, USA) and is expressed as µg/mm2. The protein content at the start of the experiment was taken as a reference and set at 100%.
4.5. Statistical Data Processing
The experiments were performed in triplicate. The significance of differences was estimated with one-way analysis of variance (ANOVA) followed by Tukey's method using an online calculator (astatsa.com/OneWay ANOVA_with_Tukey HSD/) and Student’s t test. All the data are presented as the means ± standard errors (SEs).
5. Conclusions
Mutants of PEP-associated proteins provide an excellent opportunity to expand our understanding of possible interactions among phytohormones and clarify the mechanisms of retrograde signaling from defective plastids. The block of chloroplast biogenesis and the ability to vegetate only in the presence of an external carbon source cause reprogramming of the expression of hormone-related genes and specific alterations in hormonal status, which play important roles in plant growth strategies. Elevated levels of SA, which balance stress responses and plant growth, are combined in pap mutants with increased levels of certain auxin precursors and conjugated forms, as well as a concomitant increase in the accumulation of CK metabolites and precursors that collectively promote morphogenesis in the absence of normal photosynthesis. Conversely, the levels of JA, ethylene and ABA, which negatively regulate morphogenesis, were significantly lower in the transgenic plants than in the WT plants. Hence, the interplay between growth-promoting and growth-suppressing hormones remains opposite in pap mutants, despite specific multilevel changes in hormone metabolic pathways. Pap mutations do not substantially modify the responses of chloroplast genes to hormone application, although in some cases, the effects are gene specific and are selectively divergent from those in green plants.
Taken together, our results indicate that the transition from autotrophic to heterotrophic metabolism in pap mutants induces a concerted transcriptomic response promoting versatile shifts in hormone metabolic pathways. Even though the exact set of specific transcriptional targets of these hormone-related changes still needs to be determined, the fact that altered hormone status can ensure the viability of nonphotosynthetic plants under modulated conditions is intriguing and suggests bifurcated pathways of hormone-related growth regulation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, N.V.K. and R.V.; Funding acquisition, V.V.K. and N.V.K.; Investigation, I.A.B., A.A.A., R.V., J.L. and N.V.K.; Project administration, V.V.K.; Writing—original draft, N.V.K.; Writing—review and editing, N.V.K., V.V.K., and R.V. All the authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the state assignment of the Ministry of Science and Higher Education of the Russian Federation (theme No. 122042700044-6).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Robles, P.; Quesada, V. Research Progress in the Molecular Functions of Plant mTERF Proteins. Cells. 2021, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Börner, T.; Aleynikova., A.Y.; Zubo, Y.O.; Kusnetsov, V.V. Chloroplast RNA polymerases: role in chloroplast biogenesis. Biochim. Biophys. Acta 2015, 1847, 761–769. [Google Scholar] [CrossRef]
- Liebers, M.; Pfannschmidt, T. Plastid RNA polymerases and nuclear-encoded proteins associated with them in Arabidopsis thaliana. Endocytobiosis and Cell Research 2017, 28, 20–32. [Google Scholar]
- Pfannschmidt, T.; Blanvillain, R.; Merendino, L.; Courtois, F.; Chevalier, F.; Liebers, M.; Lerbs-Mache, S. Plastid RNA polymerases: orchestration of enzymes with different evolutionary origins controls chloroplast biogenesis during the plant life cycle. J. Exp. Bot. 2015, 66, 6957–6973. [Google Scholar] [CrossRef] [PubMed]
- Grübler, B.; Merendino, L.; Twardziok, S. O.; Mininno, M.; Allorent, G.; Chevalier, F.; Ravanel, S. Light and plastid signals regulate different sets of genes in the albino mutant pap7-1. Plant Physiol. 2017, 175, 1203–1219. [Google Scholar] [CrossRef]
- Grübler, B.; Cozzi, C.; Pfannschmidt, T. A Core Module of Nuclear Genes Regulated by Biogenic Retrograde Signals from Plastids. Plants 2021, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Danilova, M.N.; Andreeva, A.A.; Doroshenko, A.S.; Kudryakova, N.V.; Kuznetsov, Vl. V.; Kusnetsov, V.V. Phytohormones Regulate the Expression of Nuclear Genes Encoding the Components of the Plastid Transcription Apparatus. Doklady Biochemistry and Biophysics 2018, 478, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, A.A.; Vankova, R.; Bychkov, I.A.; Kudryakova, N.V.; Danilova, M.N.; Lacek, J.; Pojidaeva, E.S.; Kusnetsov, V.V. Cytokinin-Regulated Expression of Arabidopsis thaliana PAP Genes and Its Implication for the Expression of Chloroplast-Encoded Genes. Biomolecules 2020, 10, 1658. [Google Scholar] [CrossRef] [PubMed]
- Nemhauser, J.L.; Hong, F.; Chory, J. Different plant hormones regulate similar processes through largely nonoverlapping transcriptional responses. Cell 2006, 126, 467–475. [Google Scholar] [CrossRef]
- Yagi, Y.; Ishizaki, Y.; Nakahira, Y.; Tozawa, Y.; Shiina, T. Eukaryotic-type plastid nucleoid protein pTAC3 is essential for transcription by the bacterial-type plastid RNA polymerase. Proc. Natl. Acad. Sci. USA 2012, 109, 7541–7546. [Google Scholar] [CrossRef]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Üstün, Ş.; Melzer, M.; Börnke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef] [PubMed]
- Gilkerson, J.; Perez-Ruiz, J. M.; Chory, J.; Callis, J. The plastid-localized pfkB-type carbohydrate kinases FRUCTOKINASE-LIKE 1 and 2 are essential for growth and development of Arabidopsis thaliana. BMC Plant Biology 2012, 12, 1–17. [Google Scholar] [CrossRef]
- Korasick, D.A.; Enders, T.A.; Strader, L.C. Auxin biosynthesis and storage forms. J. Exp. Bot. 2013, 64, 2541–2555. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.D.; Bandurski, R.S. Chemistry and Physiology of the bound auxin. Ann. Rev. Plant Physiol. 1982, 33, 403–430. [Google Scholar] [CrossRef]
- Leon-Reyes, A.; Van der Does, D.; De Lange, E.S.; Delker, C.; Wasternack, C.; Van Wees, S.C.; Ritsema, T.; Pieterse, C.M. Salicylate-mediated suppression of jasmonate-responsive gene expression in Arabidopsis is targeted downstream of the jasmonate biosynthesis pathway. Planta 2010, 232, 1423–1432. [Google Scholar] [CrossRef]
- Gawroński, P.; Górecka, M.; Bederska, M.; Rusaczonek, A.; Slesak, I.; Kruk, J.; Karpiński, S. Isochorismate synthase 1 is required for thylakoid organization, optimal plastoquinone redox status, and state transitions in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 3669–3679. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L-J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: DoWe Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y; Zhang, D.; Yan Chu, J.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Despre´ s, C. The Arabidopsis NPR1 Protein Is a Receptor for the Plant Defense Hormone Salicylic Acid. Cell Reports 2012, 1, 639–647. [Google Scholar] [CrossRef]
- Zhong, Q.; Hu, H.; Fan, B.; Zhu, C.; Chen, Z. Biosynthesis and Roles of Salicylic Acid in Balancing Stress Response and Growth in Plants. Int. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X. Salicylic acid inhibits pathogen growth in plants through repression of the auxin signaling pathway. Curr. Biol. 2007, 17, 1784–1790. [Google Scholar] [CrossRef]
- Kong, W.; Li, Y.; Zhang, M.; Jin, F.; Li, J. A Novel Arabidopsis MicroRNA Promotes IAA Biosynthesis via the Indole-3-acetaldoxime Pathway by Suppressing SUPERROOT1. Plant Cell Physiol. 2015, 56, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Hu, Y.; Aoi, Y.; Hira, H.; Ge, C.; Dai, X.; Kasahara, H.; Zhao, Y. Local conjugation of auxin by the GH3 amido synthetases is required for normal development of roots and flowers in Arabidopsis. Biochem. Biophys. Res. Commun. 2022, 589, 16e22. [Google Scholar] [CrossRef] [PubMed]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: a complex multilevel control circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef]
- Oñate-Sánchez, L.; Anderson, J.P.; Young, J.; Singh., K.B. AtERF14, a Member of the ERF Family of Transcription Factors, Plays a Nonredundant Role in Plant Defense. Plant Physiol. 2007, 143, 400–409. [Google Scholar] [CrossRef]
- Cheng, M-C.; Liao, P-M.; Kuo, W‒W.; Lin, T-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 Regulates Abiotic Stress-Responsive Gene Expression by Binding to Different cis-Acting Elements in Response to Different Stress Signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yu, Y.; Li, S.; Wang, J.; Tang, S.; Huang, R. Abscisic Acid Antagonizes Ethylene Production through the ABI4-Mediated Transcriptional Repression of ACS4 and ACS8 in Arabidopsis. Mol. Plant. 2016, 9, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Roychoudhury, A. Abscisic acid in plants under abiotic stress: crosstalk with major phytohormones. Plant Cell Rep. 2023, 42, 961–974. [Google Scholar] [CrossRef]
- Danilova, M.N.; Kudryakova, N.V.; Doroshenko, A.S.; Zabrodin, D.A.; Rakhmankulova, Z.F.; Oelmüller, R.; Kusnetsov, V.V. Opposite roles of the Arabidopsis cytokinin receptors AHK2 and AHK3 in the expression of plastid genes and genes for the plastid transcriptional machinery during senescence. Plant Mol. Biol. 2017, 93, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chang, H.P.; Hu, S.; Lu, X.T.; Yuan, C.Y.; Zhang, C.; Wang, P.; Xiao, W.J.; Xiao, L.T.; Xue, G.P.; Guo, X. Plastid casein kinase 2 knockout reduces abscisic acid (ABA) sensitivity, thermotolerance, and expression of ABA and heat stress responsive nuclear genes. J. Exp. Bot. 2014, 65, 4159–4175. [Google Scholar] [CrossRef]
- Yamburenko, M.V.; Zubo, Y.O.; Börner, T. Abscisic acid affects transcription of chloroplast genes via protein phosphatase 2C-dependent activation of nuclear genes: repression by guanosine-3′-5′-bisdiphosphate and activation by sigma factor 5. Plant J. 2015, 82, 1030–1041. [Google Scholar] [CrossRef]
- Aluru, M.R.; Zola, J.; Foudree, A.; Rodermel, S.R. Chloroplast Photooxidation-Induced Transcriptome Reprogramming in Arabidopsis immutans White Leaf Sectors. Plant Physiol. 2009, 150, 904–923. [Google Scholar] [CrossRef]
- Pogorelko, G.V.; Kambakam, S.; Nolan, T.; Foudree, A.; Zabotina, O.A.; Rodermel, S.R. Impaired Chloroplast Biogenesis in Immutans, an Arabidopsis Variegation Mutant, Modifies Developmental Programming, Cell Wall Composition and Resistance to Pseudomonas syringae. PLoS ONE 2016, 1, e0150983. [Google Scholar] [CrossRef] [PubMed]
- Hirosawa, Y; Tada, A.; Matsuura, T.; Mori, I.C.; Ogura, Y.; Hayashi, T.; Uehara, S.; Ito-Inaba, Y.; Inaba, T. Salicylic Acid Acts Antagonistically to Plastid Retrograde Signaling by Promoting the Accumulation of Photosynthesis-associated Proteins in Arabidopsis. Plant Cell Physiol. 2021, 62, 1728–1744. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Allakhverdiev, S.I.; Hurry, V.; Hüner, P.A. Stress-related hormones and glycinebetaine interplayin protection of photosynthesis under abiotic stress conditions. Photosyn. Res. 2015, 126, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Hu, H.; Fan, B.; Zhu, C.; Chen, Z. Biosynthesis and Roles of Salicylic Acid in Balancing Stress Response and Growth in Plants. Int. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Choi, J.; Huh, S.U.; Mikiko Kojima, M.; Sakakibara, H.; Paek, K-H.; Hwang, I. The Cytokinin-Activated Transcription Factor ARR2 Promotes Plant Immunity via TGA3/NPR1-Dependent Salicylic Acid Signaling in Arabidopsis Developmental. Cell 2010, 19, 284–295. [Google Scholar] [CrossRef]
- Mothes, K.; Engelbrecht, L.; Schűtte, H.R. Ǘber den Akkumulationvon α- minoisobuttersaure im Blatgewebe unter dem Einfluss von Kinetin. Physiol Plantarum 1961, 14, 72–76. [Google Scholar] [CrossRef]
- Mothes, K.; Engelbrecht, L. On the activity of a kinetin-like root factor. Life Sci. 1963, 11, 852–857. [Google Scholar] [CrossRef]
- Dobrev, P. I.; Kamınek, M. Fast and efficient separation of cytokinins from auxin and abscisic acid and their purification using mixed-mode solid-phase extraction. J. Chromatogr. A 2002, 950, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Dobrev, P. I.; Vankova, R. Quantification of abscisic acid, cytokinin, and auxin content in salt-stressed plant tissues. Methods Mol. Biol. 2012, 913, 251–261. [Google Scholar] [CrossRef]
- Bychkov, I.A.; Andreeva, A.A.; Kudryakova, N.V.; Pojidaeva, E.S.; Kusnetsov, V.V. The role of PAP4/FSD3 and PAP9/FSD2 in heat stress responses of chloroplast genes. Plant Sci. 2022, 322, 111359. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Phenotypes of pap mutants and wild-type plants grown on Murashige and Skoog (MS) media in Petri dishes for 4 weeks (A) under an illumination of 100 μmol·m-2·s-1 and a temperature of 23°C with a 16 h photoperiod. 1- WT: Col-0, 2- pap1, 3- pap6. (B) Root elongation assay. Seven-day-old seedlings were transferred to vertical MS plates and grown vertically for another 4 days under 16 h light/8 h dark conditions: 1, Col-0; 2 pap1; and 3, pap6.
Figure 1.
(A) Phenotypes of pap mutants and wild-type plants grown on Murashige and Skoog (MS) media in Petri dishes for 4 weeks (A) under an illumination of 100 μmol·m-2·s-1 and a temperature of 23°C with a 16 h photoperiod. 1- WT: Col-0, 2- pap1, 3- pap6. (B) Root elongation assay. Seven-day-old seedlings were transferred to vertical MS plates and grown vertically for another 4 days under 16 h light/8 h dark conditions: 1, Col-0; 2 pap1; and 3, pap6.

Figure 2.
Relative expression values of A. thaliana hormone biosynthesis genes in untreated 4-week-old wild-type plants and pap mutants. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1. Total RNA was isolated from rosette leaves and analyzed via qRT‒PCR using UBQ10 and PP2A as internal standards. The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test).
Figure 2.
Relative expression values of A. thaliana hormone biosynthesis genes in untreated 4-week-old wild-type plants and pap mutants. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1. Total RNA was isolated from rosette leaves and analyzed via qRT‒PCR using UBQ10 and PP2A as internal standards. The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test).
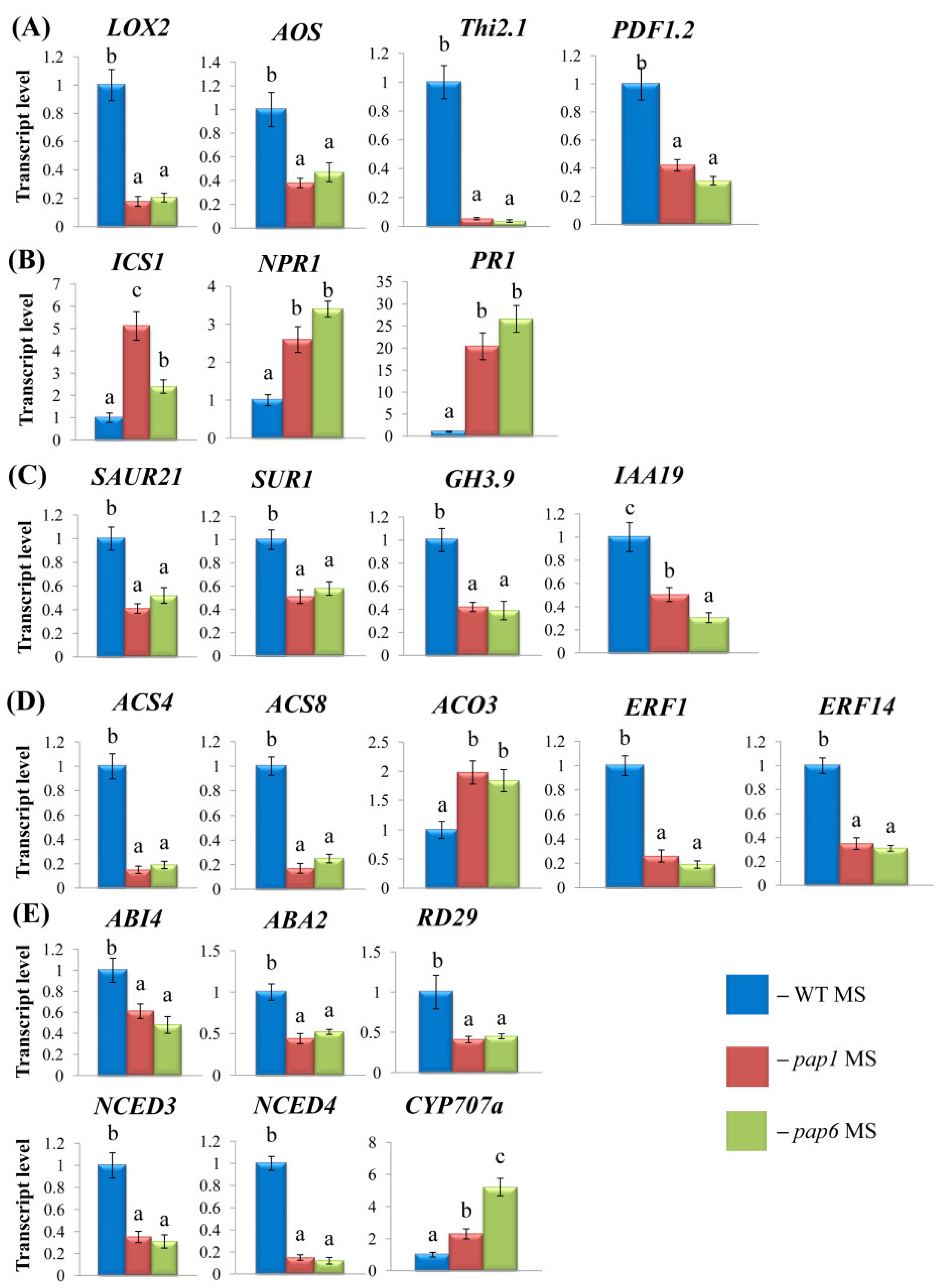
Figure 3.
(A) Induction of marker genes reflecting the activation of different hormonal pathways in 4-week-old wild-type plants and pap mutants. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1 and treated with solutions of hormones or an equal aliquot of ethanol for 3 h. Total RNA was isolated from rosette leaves and analyzed via relative quantitative RT‒PCR using UBQ10 and PP2A as internal standards. The data are presented as the means (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test). (B, C). Primary root growth inhibition in response to exogenous hormones. Seedlings were grown on MS media for 7 days under 16 h light/8 h dark conditions, subsequently transferred to vertical media plates supplemented with a range of concentrations of metJA and ACC and grown for another 4 days. Measurements were performed in triplicate with 20 seedlings for each experiment. Error bars represent SEs. (D) Sensitivity to hormone treatment during dark-induced senescence. Total protein content was measured in detached leaves (the 3rd and 4th layers) of A. thaliana WT plants and pap mutants incubated in the dark at 23°C on water or metJA (5x10-5 M) and ACC (10-5 M) for 3 days.
Figure 3.
(A) Induction of marker genes reflecting the activation of different hormonal pathways in 4-week-old wild-type plants and pap mutants. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1 and treated with solutions of hormones or an equal aliquot of ethanol for 3 h. Total RNA was isolated from rosette leaves and analyzed via relative quantitative RT‒PCR using UBQ10 and PP2A as internal standards. The data are presented as the means (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test). (B, C). Primary root growth inhibition in response to exogenous hormones. Seedlings were grown on MS media for 7 days under 16 h light/8 h dark conditions, subsequently transferred to vertical media plates supplemented with a range of concentrations of metJA and ACC and grown for another 4 days. Measurements were performed in triplicate with 20 seedlings for each experiment. Error bars represent SEs. (D) Sensitivity to hormone treatment during dark-induced senescence. Total protein content was measured in detached leaves (the 3rd and 4th layers) of A. thaliana WT plants and pap mutants incubated in the dark at 23°C on water or metJA (5x10-5 M) and ACC (10-5 M) for 3 days.
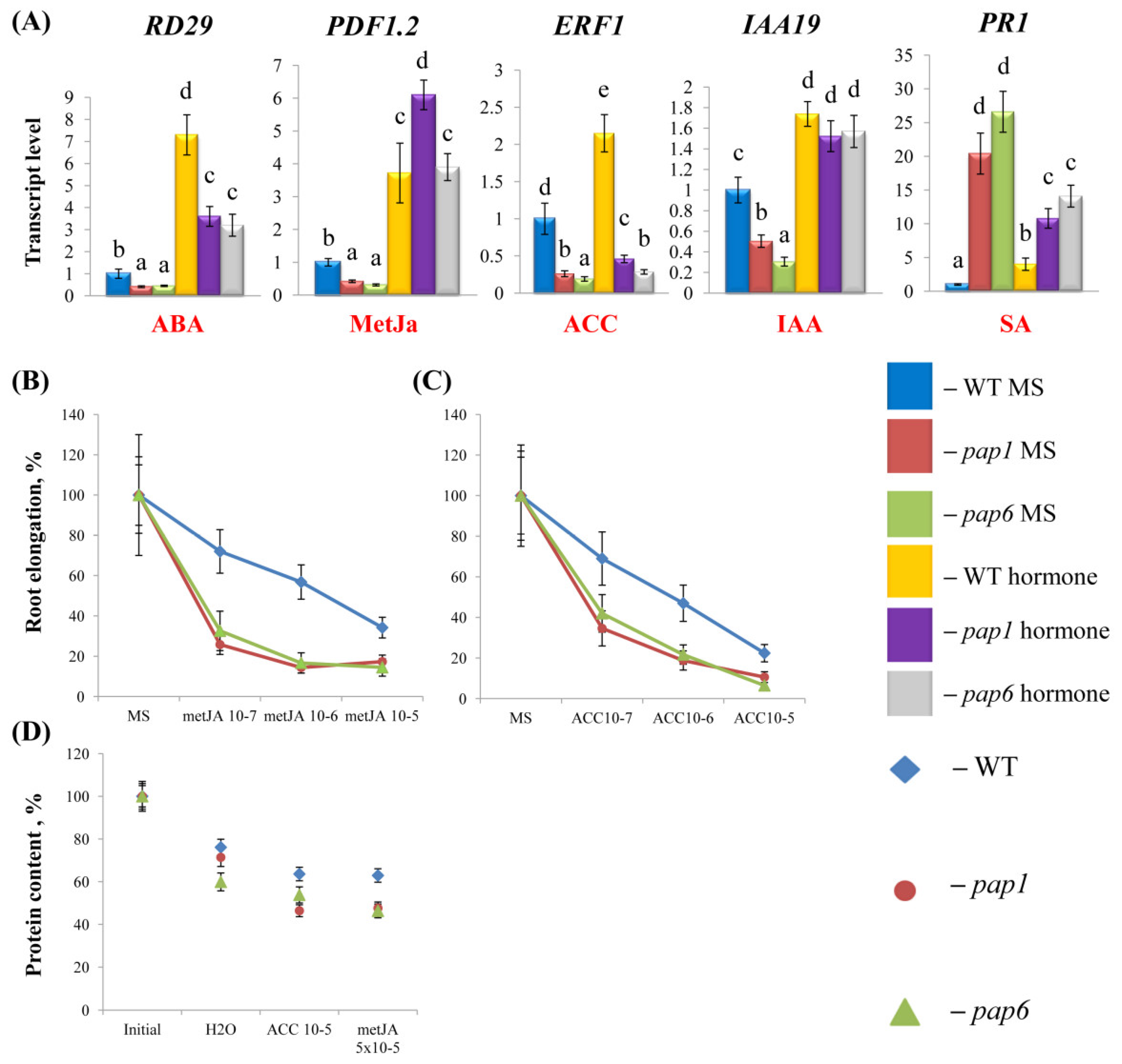
Figure 4.
Effect of hormone treatment on the relative expression values of genes related to the chloroplast transcription machinery and chloroplast encoded genes. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1 and treated with solutions of hormones or an equal aliquot of ethanol for 3 h. Total RNA was analyzed via quantitative RT‒PCR using UBQ10 and PP2A as internal standards. The data are presented as the means (n ≥ 3). The numbers in the “MS” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments. All the numerical data are presented in Table S4.
Figure 4.
Effect of hormone treatment on the relative expression values of genes related to the chloroplast transcription machinery and chloroplast encoded genes. WT and mutant plants were grown on MS media in Petri dishes for four weeks under a 16 h light/8 h dark photoperiod at 23°C with 100 μmol m-2 s-1 and treated with solutions of hormones or an equal aliquot of ethanol for 3 h. Total RNA was analyzed via quantitative RT‒PCR using UBQ10 and PP2A as internal standards. The data are presented as the means (n ≥ 3). The numbers in the “MS” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments. All the numerical data are presented in Table S4.
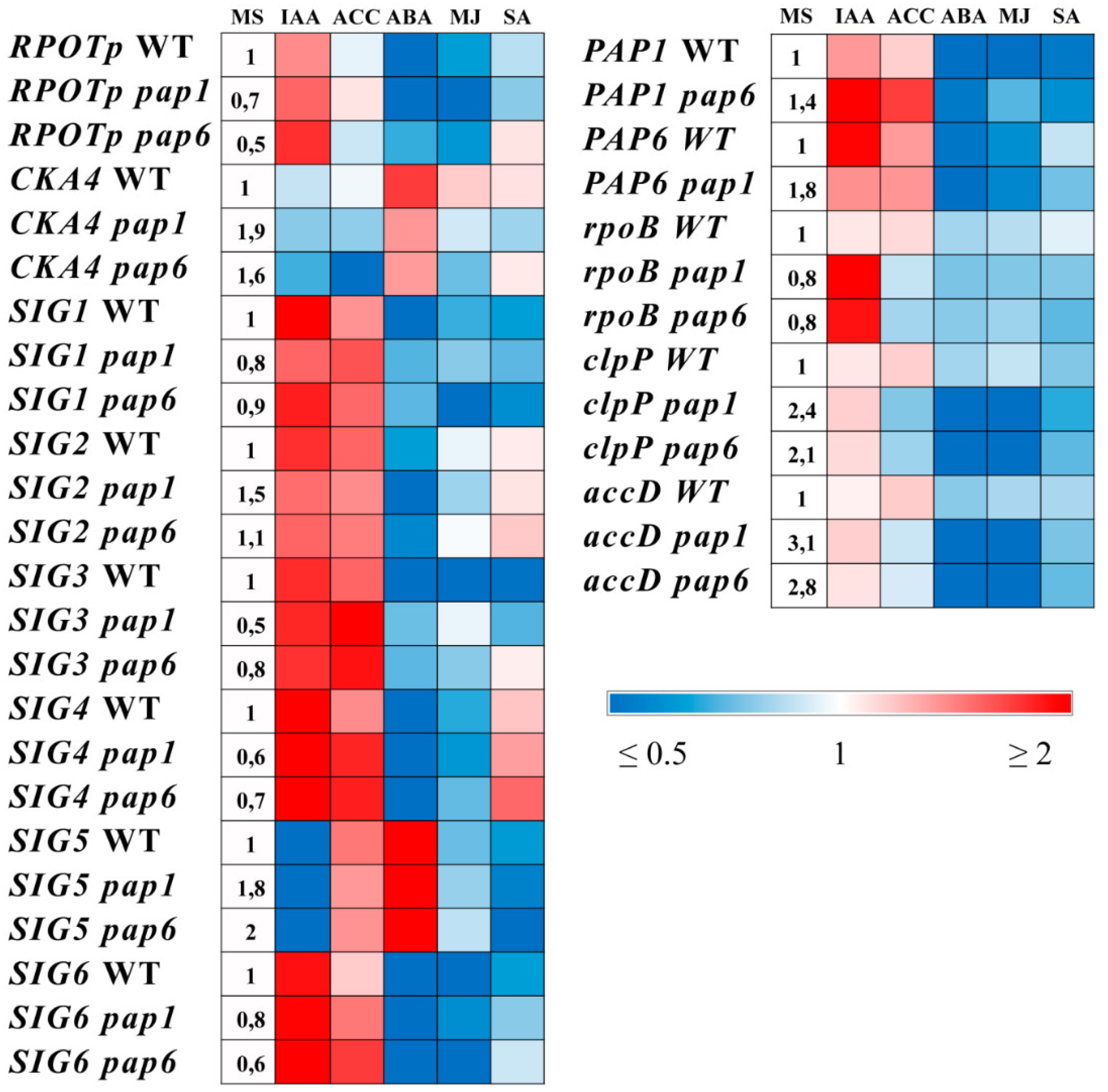

Table 1.
The impact of pap mutations on the phytohormone content (pmol/g fresh weight) of A. thaliana rosette leaves.
Table 1.
The impact of pap mutations on the phytohormone content (pmol/g fresh weight) of A. thaliana rosette leaves.
| Phytohormones | WT | pap1 | pap6 |
|---|---|---|---|
| ABA | 358.91±72.24 | 178.43±9.84* | 168.73±20.68* |
| PA | 84.39±12.01 | 14.64±2.62** | 6.73±2.16** |
| 9OH-ABA | 15.01±2.8546 | 2.70±0.46** | 1.14±1.14** |
| DPA | 31.22±7.04 | 29.49±7.02 | 18.76±3.91 |
| BzA | 655.04±61.87 | 1092.02±146.66* | 1160.01±90.71* |
| SA | 882.35±182.37 | 2588.49. ±498.01 | 3264.72±897.05* |
| JA | 151.58±29.71 | 145.89±30.01 | 140.03±13.77 |
| JA-Ileu | 16.31±3.27 | 9.3225±3.03 | 14.17±3.50 |
| cisOPDA | 137.52±7.42 | 94.85±24.90 | 58.77±13.50* |
| ACC | 338.17±86.45 | 11.00±2.63** | 34.28±7.05** |
| IAA | 67.44±2.89 | 64.63±6.54 | 66.74±4.22 |
| OxIAA-GE | 922.21±129.18 | 1461.79±151.29* | 1232.99±119.36 |
| PAA | 98.06±8.44 | 133.42±21.69 | 162.57±18.38 |
| IAA-Asp | 56.51±17.94 | 5.80±1.12* | 12.76±2.35* |
| IAA-GE | 20.11±1.91 | 33.34±5.95 | 32.02±5.82 |
| OxIAA | 248.51±38.26 | 312.15±47.86 | 326.04±26.21 |
| IAN | 4284.60 ±1411.27 | 5577.52±833.42 | 6208.17±2741.61 |
Abbreviations: ABA – abscisic acid, PA – phaseic acid, DPA – dihydrophaseic acid, 9OH-ABA – 9-hydroxy-ABA, BzA – benzoic acid, SA – salicylic acid, JA – jasmonic acid, JA-Ile – JA-isoleucine, cisOPDA – cis-oxophytodienoic acid, ACC – 1-aminocyclopropane-1-carboxylic acid, IAA – indole-3-acetic acid, IAA-GE – IAA-glucose ester, PAA – Phenylacetic acid, IAA-Asp – IAA-aspartate, OxIAA – oxo-IAA, OxIAA-GE – oxo-IAA-glucose ester, IAN – Indole-3-acetonitrile. Asterisks (*) indicate significant differences compared to WT (Student’s t test; * - P<0.05; ** - P<0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



