Submitted:
23 December 2023
Posted:
25 December 2023
You are already at the latest version
Abstract
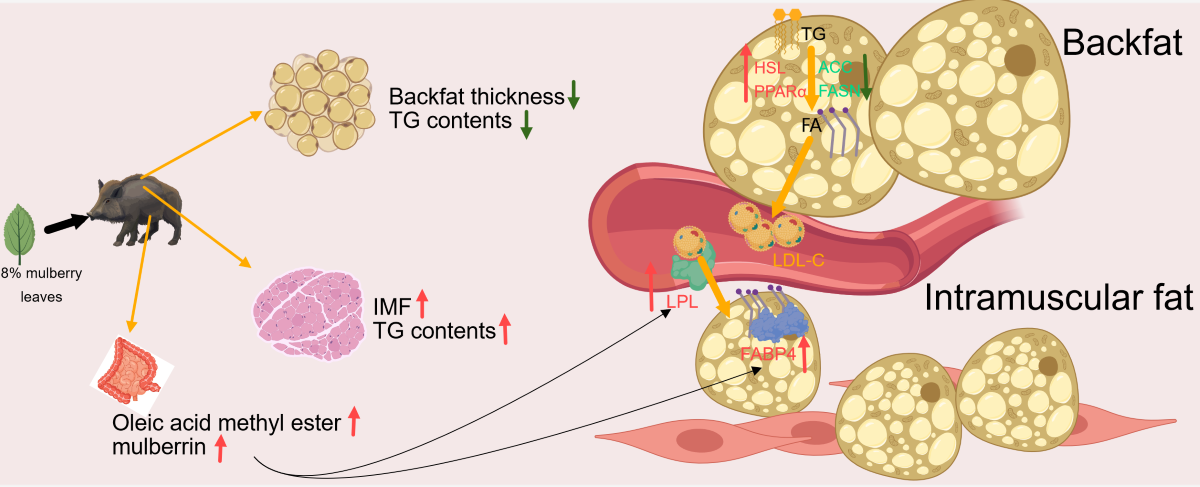
Results showed that mulberry leaves could significantly reduce backfat thickness (P<0.05) and increase intramuscular fat (IMF) content (P<0.05). Lipidomics analysis showed that mulberry leaves improved lipid profile composition and increased the proportion of triglycerides (TG). Interestingly, IMF content was positively correlated with acyl C18:2 and negatively correlated with C18:1 of differential TGs. In addition, cecal microbiological analysis showed that mulberry leaves could increase the abundance of bacteria such as UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, Limosilactobacillus, and so on. At the same time improve the L-tyrosine-baton rouge ester, Oleic acid methyl ester, 21-deoxycortisol, N-acetyldihydrosphingosine, Mulberrin relative abundance. In this study, we found that mulberry leaves could extremely significantly increasing the mRNA relative expression of lipoprotein lipase (LPL), fatty acid-binding protein 4 (FABP4) and peroxisome proliferators-activated receptor γ (PPARγ) in muscle (P<0.01), significantly increasing mRNA relative expression of diacylglycerol acyltransferase 1 (DGAT1) (P<0.05) and significantly decreasing the mRNA relative expression of acetyl CoA carboxylase (ACC) (P<0.05), and aignificantly upregulated hormone-sensitive triglyceride lipase (HSL), peroxisome proliferators-activated receptor α (PPARα) mRNA relative expression (P<0.05) in backfat. In addition mulberry leaves could increased serum leptin and adiponectin (P<0.01). Ultimately lead to up-regulation of IMF and down-regulation of backfat.
Keywords:
lipidomic 1
; intramuscular fat 2
; mulberry leaves 3
; triglyceride 4
; lipid profile 5
1. Introduction
Pork is the most produced and most widely consumed type of meat product in the world and the main source of protein intake for humans because of its rich and balanced nutrition [1,2]. The quality and safety of pork are closely related to people’s lives. With the rise of large-scale swine farms worldwide, although the production of pork has been guaranteed, the taste and quality of pork have been continuously reduced [3,4]. Intramuscular fat (IMF) is an important indicator of pork quality, one of the important traits affecting pork quality, closely related to flavor, juiciness, tenderness, etc., determining the quality and nutritional value of pork [5]; however, it has not attracted sufficient attention on some pig farms. With the increasing demand for higher-quality meat, the IMF content of meat has gradually increased [6]. However, a higher IMF content is often accompanied by a higher backfat thickness, and excessive backfat seriously affects the lean meat percentage. Therefore, balancing the IMF and backfat is an extremely important issue [7].
Improving meat quality by improving feed formulations is currently one of the mainstream strategies [8]. In many countries, the shortage of raw feed materials and the continuous improvement of people’s requirements for meat quality are the main pressures that inhibit the development of the breeding industry [9]. Therefore, finding a raw feed material that can not only improve meat quality but also be used as a regular feed substitute is the main research focus of the livestock industry. Mulberry leaf plants of the genus Morusare widely distributed in northwest China and the south, Japan, Europe, Southeast Asia, and other regions [10]. They have a high protein content (about 25–30%) and are rich in a variety of vitamins, amino acids, and mulberry leaf polysaccharide, and are widely used in Asia as a new protein feed [11]. Related studies have shown that 8-10% mulberry leaf supplementation can effectively improve pork quality [12,13]. Related studies have shown that mulberry leaves have positive effects on lowering blood lipids, blood pressure, immunity, and inflammation; therefore, they are often used as a traditional Chinese medicine or functional food [14,15]. The Yuxi black pig, a central Chinese pig breed, has roughage resistance, disease resistance, and good meat quality features. Preliminary evidence shows that mulberry leaves can significantly improve the quality of Yuxi black pig meat and reduce backfat ketone bodies, but the specific mechanism is unclear [13,16].
Lipidomics, as an emerging omics technology, is important for exploring the composition of lipid profiles. Lipidomics plays an important role in exploring the regulatory mechanisms of lipid metabolism by screening, classifying, and refining differentially expressed lipid molecules. The gut microbiota refers to the assemblage of microorganisms in the gut. Alterations in the composition of these gut microorganisms can impact the physiology, immune, and nutrient absorption of the host. Conversely, changes in the health status of the host can lead to corresponding changes in gut microbiota composition [17]. Exploring the relationship between gut microbiota, gut metabolites, and lipid metabolism has been a hot topic [18,19]. Metabolomics techniques based on high performance gas chromatography-mass spectrometry (HPGLC-MS) are becoming a common tool for the analysis of volatile and non-volatile compounds, and are widely used in the identification of food products and the analysis of metabolites for the characterization of food products [20].
In this study, the effect of mulberry leaves on IMF was explored through comprehensive multi-omics analysis, and the relationship between differential lipid molecules, gut microbiota, and gut metabolites was explored to screen out gut microbiota and metabolites that may have an impact on IMF. Finally, possible regulatory mechanisms were speculated by exploring the relative expression differences of important regulatory genes in lipid metabolism.
2. Article Types
Original Research
3. Materials and Methods
3.1. Ethics Approval
The animal study was approved by the Institutional Animal Ethics Committee of Henan University of Science and Technology (approval 2022-02-023).
3.2. Preparation of Animals and Samples
All procedures involved in animal experiments were performed in accordance with the Animal Protection and Welfare Guidelines of the Ministry of Agriculture of China. Yuxi black pigs used in the experiments were purchased from Luoyang Luanchuan Farming Co., Ltd., Luoyang China. Sixty Yuxi black pigs weighing approximately 40 kg were randomly assigned to four identical rooms, with 15 pigs in each room. One group was fed a diet without mulberry leaves, the other three were fed 8% fermented mulberry leaves based on the standard diet. The experimental period was 120 days, with 7 days of pre-experimentation. The nutrient composition of the feed is shown in Table S1. Mulberry leaves were purchased from the Henan Sangbio Agricultural Company. After 120 days of feeding, the animals were weighed and fasted for 1 day before slaughter, and approximately 200 g of longissimus thoracis and backfat tissue at the fifth to sixth rib and cecal contents were collected.
3.3. Determination of Meat Quality
Body weight was determined prior to slaughter, followed by backfat thickness, marbling score, pH, meat color, IMF content, and triglycerides (TG) content of muscle and backfat. The average backfat thickness of the first rib, last lumbar spine, and last rib was used as the backfat thickness. The marbling score was determined based on a cross-sectional section of the muscle at the first lumbar spine, ranging from 0 to 5, with higher scores indicating more abundant IMF content. The color of the longest dorsal muscle was determined using OPTO-STAR (Germany, Denmark), and the meat color parameters included L* (lightness), a* (intensity), and b* (yellowness).
The pH-value of the longest dorsal muscle was determined 40-50 min after slaughter and 24 h after slaughter (HI99163, HANNA, Italy). The IMF content was measured using a Soxhlete extractor (SX-360, OPSIS, Sweden). Triglyceride content was determined by the triglyceride kit (EY-01H2206, Shanghai, China)
3.4. Lipidomics Analysis
Thirty milligrams of the sample (referring to the fat-rich part of the LT muscle) was weighed and homogenized by adding an internal standard (Lyso PC-17:0) and 280 μL of methanol-water (V: V = 1:1) for 2 min (60 Hz), followed by vortexing with chloroform for 30 s and centrifugation for 10 min (12,000 × g, 4°C). The lower chloroform layer was removed, 280 μL of chloroform-methanol (V: V = 2:1) was added, swirled for 30 s, ultrasonic extraction for 20 min, allowed to rest for 20 min, then centrifuged for 10 min (12,000 rpm, 4°C) to extract lipids, wave-dried, and redissolved with 280 μL isopropanol-methanol. After centrifugation for 15 min (12,000 × g, 4°C), 150 μL of the supernatant was loaded into a lined LC-MS injection vial for LC-MS analysis.
The column temperature was set at 55°C, while mobile phase A consisted of acetonitrile: water at a ratio of 6:4 (v/v), and mobile phase B consisted of isopropanol: acetonitrile at a ratio of 9:1 (v/v). The flow rate was maintained at 0.26 mL/min, with an injection volume of 2 µL in positive ion mode and 2 μL in negative ion mode set to be 4 μL. The samples were separated using Dionex U3000 ultra-high-performance liquid chromatography (UHPLC) (Thermo Scientific™) and analyzed by mass spectrometry using Q Exactive Plus (Thermo Scientific™). The detection employed heated electrospray ionization operating in both positive and negative ion modes.
3.5. Total RNA Extraction and qPCR
TRIzol reagent (Servicebio, China) was used to extract total RNA from subcutaneous fat and IMF, and a spectrophotometer (NanoDrop 2000, Thermo Fisher) was used to determine the absorbance of RNA at 260 nm and 280 nm to ensure the purity and concentration of RNA. The RNA was then reverse-transcribed using the SweScript RT I First Strand cDNA Synthesis Kit (Servicebio, China) and random primers. QPCR was performed using a Bio-Rad CFX Connect PCR fluorescence quantitative PCR instrument with SYBR Green qPCR Master Mix and gene-specific primers (Table S2), and relative expression was calculated using the 2−ΔΔCt method [21].
3.6. H&E staining, Oil Red Staining, and BODIPY Staining
H&E staining: Backfat tissue samples were placed in fixative for 1 day, embedded in paraffin, and made into 10 μm frozen sections. The sections were sequentially immersed in xylene I and II for 15 min, then sequentially immersed in 95%, 85%, and 70% ethanol for 5 min, and then in distilled water for 5 min. The eosin staining solution was added to the tissue sample sections to allow the tissue to be fully stained for 3 min. The slices were sequentially placed in 95% alcohol I, 95% alcohol II, absolute alcohol I, absolute alcohol II, xylene I, and xylene II for 5 min, then dried and sealed with neutral glue, and finally observed under a microscope.
Oil red staining: The muscle tissue sections were fixed in 4% paraformaldehyde fixator at 37 ℃ for 20–30 min, washed with 70% ethanol solution for 5 s, stained with oil red staining solution for 5–10 min, washed with 70% ethanol solution to remove the excess staining solution, and washed with precooled phosphate-buffered saline (PBS) three times; subsequently, the sections were immersed in hematoxylin solution for 2 min, rinsed with PBS for another 30 s, and finally observed under a microscope.
BODIPY staining: Muscle tissue sections were fixed with 4% formaldehyde or 4% acetaldehyde, then incubated with BODIPY for 30 min, and cells were washed three times with PBS or other buffers before being photographed using a fluorescence microscope.
3.7. DNA Extraction and 16SrRNA Gene Amplification and Sequencing
DNA extraction from the cecal contents was performed using the Intestinal Fecal DNA Extraction Kit Omega D4015-02 (Omega Bio-Tek, Beijing, China). Subsequent PCR amplification was performed using TransStart FastPfu DNA Polymerase (TransGen AP221-02, Beijing, China) with specific primers and barcodes. The PCR products were quantified using the QuantiFluor™ -ST Blue fluorescent Quantitative System (Promega), followed by Illumina PE250 library construction and Illumina PE250 sequencing. The PE reads obtained by Illumina PE250 sequencing were first assembled according to the overlap relationship, and sequence quality was controlled and filtered simultaneously. After distinguishing the samples, OTU cluster and taxonomic analyses were performed. The sequencing depth was determined based on taxonomic information, and statistical analyses of the community structure were performed at each taxonomic level.
3.8. Untargeted Metabolomics Assays
3.9. Statistical Analysis
The experimental data are expressed as the mean ± standard deviation (SD) of three independent experiments. Statistical analyses were performed using SPSS 22.0. GraphPad software (9.0), and Origin 2022 were used for correlation analysis and data visualization, and P values less than 0.05 were considered significant.
4. Results
4.1. Effect of Mulberry Leaves on Growth Performance and Meat Quality
The results of growth performance showed that a diet supplemented with 8% mulberry leaves did not affect the growth performance of Yuxi black pigs (Table S3). Meat quality measurements showed that the marbling score and lightness (L*) of muscle tissue in the mulberry leaf group were increase significantly, whereas pH, shear force, cleavage (a*), and yellowness (b*) were not significantly changed (Table 1). In addition, we found that mulberry leaves significantly increased IMF and TG in muscle and significantly reduced backfat thickness and TG in backfat (P < 0.05) (Figure 1A–D). Oil red O staining and BODIPY staining of muscle tissue also showed that the fat content of muscle in the ML group was significantly higher than that in the CON group (P < 0.01) (Figure 1E, F). The results of H&E staining of adipose of backfat showed that the size of adipocytes in the backfat of the ML group was significantly lower than that in the CON group (P < 0.01) (Figure 1E, G). The above results show that feeding a diet supplemented with 8% mulberry leaves could improve IMF and reduce backfat thickness in Yuxi black pigs.
4.2. Distribution of Lipid Profiles of Muscle Tissue and Screening of Differential Lipid Molecules
The significant difference in IMF between the CON and ML groups made us eager to understand specific lipid distribution. Principal component analysis (PCA) showed significant differences in lipid profiles between the CON and ML groups (Figure 2A). The lipid compositions of the CON and ML groups are shown in Figure 2B–D. The results of lipid profiling showed that the proportions of TG, Cer, and FA in the ML group were significantly higher than those in the CON group, whereas the proportions of PC, PS, and SM were significantly lower than those in the CON group (Figure 2E–I). In addition, the abundances of TG, Cer, and FA in the ML group were significantly higher than those in the CON group, while the abundances of PC, PS, and LSM were significantly lower than those in the CON group (Figure 2J–N).
A total of 777 lipid molecules were identified, and 110 differential lipid molecules (P < 0.05, FC > 2) were screened (Figure 3A). The heat map results of the top 20 differentially expressed lipid molecules in abundance showed that compared with the CON group, TG(18:1/18:2/18:2), TG(18:0/16:0/18:1), TG(16:0/18:2/18:2) showed an upward trend in the ML group, while TG(16:0/14:0/18:1), TG(18:0/18:1/18:1), TG(18:1/18:1/18:1) showed a downward trend in the ML group (Figure 3B). Correlation analysis of different lipid species showed that IMF was significantly positively correlated with TG but negatively correlated with PC and LSM (Figure 3C). The differential lipid molecules were classified. A heat map was drawn, and the results showed that the lipid molecules with significantly upregulated expressions were mainly GL, PS, PI, LPC, LPE, and SP. By contrast, the lipid molecules with significantly downregulated expressions were mainly PC and PE (Figure 3D–J).
The above results show that TG is the most abundant lipid molecule with significant differences between groups; therefore, it is necessary to analyze TG more deeply. The number of carbon atoms in the acyl chain of TG is generally between 4 and 26, with = (18:1), (18:2), (16:0), and (16:1) being the most important acyl chains, accounting for more than 70%. Through correlation analysis of the major acyl chains of TG, we found that (18:1) was negatively correlated with (18:2) and (16:0) and positively correlated with (16:1), (18:2) was positively correlated with (16:0), and (16:0) was negatively correlated with (16:0) (Figure 3K). Subsequently, through correlation analysis of the acyl chain of TG and the IMF content, we found that the IMF content was positively correlated with (18:2) and (16:0) and negatively correlated with (18:1) and (16:1) (Figure 3L).
The above results showed that feeding a diet supplemented with 8% mulberry leaves could improve the lipid profile of muscle mainly by increasing TG levels, among which TG (16:0/16:0/18:1), TG (18:0/16:0/18:1), and TG (18:0/16:0/18:1) were the main lipid molecules with upregulated expressions. At the same time, we found that IMF content was positively correlated with C18:2 in TG and negatively correlated with C18:1 in TG.
4.3. Effects of Mulberry Leaves on Gut Microbiota
To explore the effects of mulberry leaves on the cecal bacterial composition of Yuxi black pigs, cecal bacterial diversity was analyzed. We detected and screened 720,798 sequences. OTU analysis of non-repetitive sequences with 97% similarity obtained 4496 OTUs, and the rarefaction curve tended to be flat (Figure S1). The results of the diversity analysis of colony composition showed that the Chao1 index, Shannon index, and Simpson index were not significantly different between the CON and ML groups (Figure S2). At the phylum level, Bacteroidetes levels were significantly increased (P < 0.05). The levels of Proteobacteria and Campylobacterota were significantly decreased (P < 0.01). By contrast, the levels of Firmicutes and Actinobacteria were not significantly changed (Figure 4B). At the genus level, the top 20 bacterial genera in the CON and ML groups were plotted in stacked bar graphs, as shown in Figure 4C. The top five were Lachnospiraceae_unclassified, UCG-005, [Eubacterium]_coprostanoligenes_group_norank, Lactobacillus, and Phascolarctobacterium. Subsequently, a heat map was drawn for the top 30 differential bacteria between the CON and ML groups, and the results showed that the levels of UCG-005, Muribaculaceae_norank, Prevotella showed an increasing trend, and the levels of Campylobacter, Prevotella_7, and Agathobacter showed a decreasing trend (Figure 4D). Correlation analysis between differential bacteria and lipid species showed that UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, Prevotella, Ruminococcaceae_uncultured, Limosilactobacillus, and Monoglobus were significantly positively correlated with IMF and TG (Figure 4E). The correlation analysis between different bacteria and acyl groups of TG showed that UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, and Ruminococcaceae_uncultured were positively correlated with precursor of linoleic acid (C18:2) and negatively correlated with oleic acid precursors (C18:1) (Figure 4F). The above results showed that mulberry leaves may regulate IMF by improving the relative abundance of UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, and Ruminococcaceae_uncultured.
4.4. Untargeted Metabolomics Analysis of Cecal Contents
To explore the effects of mulberry leaves on cecal contents, an untargeted metabolomic analysis of cecal contents was performed. Pie chart based on the number of various compounds in the contents of the cecum; the top 3 compounds were organic acids and their derivatives, lipids and lipid-like molecules, organic heterocyclic compounds (Figure 5A). We screened 338 differential compounds (P < 0.05, FC > 2) and generated a volcano plot (Figure 5B). Heat maps of the top 30 differentially expressed compounds were plotted. The results showed that the expressions of maltose, acetylcarnitine, and creatine were significantly downregulated, and the expressions of 1-stearoyl-2-linoleoyl-sn-glycerol and traumatic acid were significantly upregulated (Figure 5C). Subsequently, the differential compounds were classified, heat maps were drawn sequentially, and the results are shown in Figure 5D–L. The correlation results of different compounds with IMF and lipid species showed that L−tyrosine−ethyl ester, oleic acid methyl ester, 21−deoxycortisol, N−acetyldihydrosphingosine, and mulberrin were positively correlated with TG, creatinine and pheniramine were negatively correlated with TG (Figure 5M). The correlation results of different compounds with acyl of TG showed that L-tyrosine–ethyl ester, oleic acid methyl ester, 21−deoxycortisol, N−N-acetyldihydrosphingosine, and mulberrin were significantly positively correlated with precursor of linoleic acid (C18:2) and negatively correlated with oleic acid precursors (C18:1) (Figure 5N). The above results show that the mulberry leaves may, through regulating the cecum L-tyrosine-baton rouge ester, Oleic acid methyl ester, 21-deoxycortisol, N-acetyldihydrosphingosine, and mulberrin, then affect the IMF content.
4.5 Expression of Genes Involved in Lipid Metabolism and Changes in Regulatory Factors in Serum
To further explore the mechanism by which mulberry leaf improves meat quality, we measured the genes related to lipid metabolism and some serum indicators, including genes related to fatty acid synthesis, acetyl CoA carboxylase (ACC), fatty acid synthase (FASN)), genes related to fatty acid transport, lipoprotein lipase (LPL), fatty acid-binding protein 4 (FABP4) genes related to triglyceride synthesis, diacylglycerol acyltransferase 1 (DGAT1), genes related to triglyceride breakdown, hormone-sensitive triglyceride lipase (HSL), and key genes for prelipid cell differentiation, peroxisome proliferator-activated receptor γ (PPARγ). The results showed that the expression of ACC was significantly downregulated in backfat (P < 0.05), and the expressions of peroxisome proliferator-activated receptor α (PPARα) and HSL were significantly upregulated (P < 0.05) (Figure 6A–D). The expressions of FABP4, LPL, DGAT1, and PPARγ were significantly upregulated in muscle tissue (P < 0.01), whereas those of ACC and FASN were not significantly different (Figure 6E–J). The results of serum indices showed that the expressions of low-density lipoprotein cholesterol (LDL-C) and insulin were significantly downregulated (P < 0.01), the expression of high-density lipoprotein cholesterol (HDL-C) was significantly upregulated (P < 0.05), and the expressions of leptin, adiponectin, and free fatty acid were significantly upregulated (P < 0.01) (Figure 6K–P). These results indicate that mulberry leaves may inhibit backfat deposition by inhibiting the relative expression of ACC in backfat and promoting PPARα and HSL expression. Meanwhile, mulberry leaves may also increase IMF by promoting FABP4, LPL, and PPARγ expression in the muscles. The increase in serum leptin and adiponectin levels may also be one of the factors regulating lipid deposition in muscles and backfat.
5. Discussion
The above study found that mulberry leaves can significantly improve the meat quality of pork. Mulberry leaves are often used in traditional Chinese medicine to reduce blood glucose and lipid levels [10]. More attention has been paid to their application in non-alcoholic hepatitis and dyslipidemia, so their potential to improve animal meat quality has been ignored. Backfat thickness is an important indicator of performance and meat quality. Excessive backfat often has a negative impact on the reproductive performance, immunity, and meat quality of pigs [24,25]. Reducing backfat is key to improving the lean meat percentage, relevant studies have found that mulberry leaves can inhibit the deposition of backfat by promoting the secretion of leptin and the expression of the lipolysis gene HSL in the backfat tissue of pigs and inhibiting the expression of the fatty acid synthesis gene FASN, which is consistent with our results [16]. Leptin, a cytokine secreted by adipocytes, can promote lipolysis and inhibit fat deposition, which may be related to the regulation of backfat by mulberry leaves [26]. In addition, mulberry leaves may regulate fat deposition through the de novo fatty acid synthesis (DNL) pathway. ACC and FASN are the key genes upstream of the DNL pathway, we found that mulberry leaves had a significant inhibitory effect on ACC in backfat tissue, which may be one of the reasons for the inhibition of backfat deposition by mulberry leaves [27,28]. In addition, related studies have found that mulberry leaf polysaccharides (MLP) may increase the activity of brown adipocytes, accelerate fat consumption, and inhibit fat deposition, which may be one of the mechanisms by which MLP inhibits backfat deposition [29]. In addition, MLP can inhibit the expression of pancreatic lipase and reduce the intestinal absorption ability of dietary fat, thus playing an anti-obesity role [30].
IMF is an important factor in determining meat tenderness. Factors that affect fat deposition include genetics, nutrition, age, and environment [31,32]. There are some differences in fat deposition at different sites, but the specific mechanism remains unclear. Backfat and IMF content are important indicators to measure meat quality. A higher IMF content, marbling score, and lower backfat content are often preferred by consumers. However, backfat and IMF often show a positive correlation, so it is very important to balance IMF and backfat. Improving meat quality through dietary formulation is currently the mainstream strategy. Previous studies have found that mulberry leaves may promote IMF deposition and improve amino acid composition, which is consistent with our experimental results; however, its specific mechanism of action remains unclear [13]. Previous studies have found that some flavonoids in the mulberry leaf extract can significantly stimulate the differentiation of 3T3-L1 cells and promoted adiponectin secretion, which may be related to the promotion of IMF deposition by mulberry leaves [33]. Our results also showed that mulberry leaf extract significantly increased the serum levels of adiponectin and adiponectin, which are cytokines secreted during adipocyte differentiation, promoted the expression of PPARγ, and inhibited insulin resistance [34]. This difference in IMF deposition may be related to the regulation of adiponectin levels in mulberry leaves. We found that mulberry leaves significantly increased the expression of LPL, FABP4, and DGAT1, which are key genes for cellular fatty acid uptake and the rate-limiting gene for triglyceride synthesis [35,36]. These results suggest that mulberry leaves may promote fatty acid uptake and TG synthesis by upregulating the expression of LPL, FABP4, and DGAT1, thereby increasing the IMF. Fatty acid oxidation is an issue related to food quality, and there are many ways to inhibit fatty acid oxidation. Rosemary extract has been reported to reduce lipid oxidation in meat to prolong storage time and improve the fatty acid composition of meat, and supplementation with α-tocopherol has been reported to alter the body's polyunsaturated fatty acid profile and reduce oxidative stress indices [37,38]. Lipid nutrition is a kind of oil-in-water complex in meat, and there are significant differences in the content and composition of lipid nutrients between different animals, with the general content ranging from 5 to 35%, the lipid nutrition of pork can be as high as 20% to 35%, beef 8% to 25%, calf meat 4% to 12%, and sheep meat 8% to 25% [39-41]. Mammalian lipid nutrition is higher in saturated fatty acids than avian. pig fat contains about 45% saturated fatty acids, while chicken fat contains only 56%. water retentionr, which is the ability of meat to retain all or part of its moisture, was reported to be significantly lower in dripping loss for loins of pigs with higher marbling scores than for those with lower marbling scores, but there was no difference in cooking loss. In contrast, loin meat of Large White and Long White pigs with higher fat content showed higher cooking loss than low-fat pork, but no significant difference in dripping loss [42,43].
The development of lipidomics has allowed us to better understand subtle differences in lipid composition and lipid deposition, and the lipidomics and metabolomics has played an important role in exploring microscopic changes in lipids and metabolites, and finding possible biomarkers and tracing upstream pathways through deep correlation of differentiators with traits can help to elucidate the underlying mechanisms [44-46]. In this study, we used lipidomics technology to analyze the differences in lipid molecules in muscle and identified the main differential lipid molecules as TG, PE, PC, and TG. In this study, we found that dietary addition of mulberry leaves enhanced muscle C18:2 content and C18:2 was significantly positively correlated with IMF, and linoleic acid, as a precursor of C18:2 acyl group in lipid molecules, may be potentially associated with IMF, and conjugated linoleic acid, as a heterodimer of linoleic acid, has been reported to increase the deposition of IMF in animals through activation of the PPARα/FAPB4 signaling pathway [47], and palmitic acid as C18:0 precursor was also positively correlated with IMF. It has been reported that palmitic acid can induce endoplasmic reticulum stress through activation of the NF-κB signaling pathway, which in turn leads to the deposition of lipid tissues and promotes the release of exosome miR-4431. The above study suggests that mulberry leaves may lead to IMF deposition by increasing linoleic acid and palmitic acid [48,49].
Previous studies have found that after adding 10% mulberry leaves to the pig diet, the C18:2 ratio content of free fatty acids in muscle was significantly increased, and the C18:1 ratio content was significantly decreased, which is similar to the results of our study on the acyl group of muscle TG [50]. There are two possible reasons for this finding. The higher cellular uptake of linoleic acid compared with oleic acid results in a higher C18:2 ratio and a lower C18:1 ratio in the TG. Another reason is that with the increase in IMF, fatty acids synthesized by the DNL pathway or taken up from the blood circulation are more likely to be converted into linoleic acid rather than oleic acid in cells. Relevant studies have found that fatty acids are more inclined to be converted into long-chain polyunsaturated fatty acids in mature 3T3-L1 cells, but the specific mechanism remains unclear [51]. Previous studies on meat quality analysis have often focused on exploring the distribution and differences of free fatty acids in muscle. However, the content of free fatty acids in muscle is much lower than that of TG, and the composition of free fatty acids is unstable and easily regulated by a variety of factors; therefore, the analysis of the TG acyl chain is more intuitive.
The cecum microbiota is a central factor in the regulation of lipid metabolism, and immune ability (Adak & Khan, 2019; Schoeler & Caesar, 2019). However, the effect of mulberry leaves on the gut microbiota and the regulatory mechanism between gut microbiota and lipids are still unclear. Firmicutes and Bacteroidetes are the main microorganisms in the mammalian gut, and studies have found that the ratio of Firmicutes to Bacteroidetes is positively correlated with obesity [52,53]. Mulberry leaves significantly increased the abundance of Bacteroidetes in the cecum, leading to a decrease in the ratio of Firmicutes to Bacteroidetes, which may be related to the anti-obesity effect of mulberry leaves. We found that UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, Prevotella, Limosilactobacillus, Ruminococcaceae_uncultured, and Monoglobus were significantly positively correlated with IMF and TG, among which UCG−005, Muribaculaceae_norank, Prevotellaceae_NK3B31_group, and Ruminococcaceae_uncultured were positively correlated with precursor of linoleic acid (C18:2), negatively correlated with oleic acid precursors (C18:1). It has been reported that UCG−005 is correlated with circulating acetate [54], and Prevotellaceae_NK3B31_group is often regarded as the main producer of butyrate [55]. Scfas are often closely related to meat quality regulation and lipid metabolism, suggesting that mulberry leaves may regulate lipid deposition in pig muscle by changing the composition of bacterial flora [56]. We found that mulberrin, oleic acid methyl ester in the cecal contents, were positively correlated with IMF and significantly negatively correlated with oleate. Gut metabolites are closely related to multiple physiological processes and metabolic pathways in the host [57]. In the study, a variety of gut-differentiated compounds were found to be associated with IMF, among which mulberry leaf flavonoids should be emphasized as mulberry leaf-derived compounds, and flavonoids in mulberry leaves are closely related to lipid metabolism, flavonoids in mulberry leaves have been reported to promote adipocyte differentiation by increasing the expression of PPARγ and FABP4 genes, which act as candidate genes for the regulation of IMF, and the results suggest that flavonoids in fermented mulberry leaves may act as potential modulators to regulate IMF deposition by promoting the expression of FABP4 [33]. Oleic acid methyl ester may act as a precursor of C18:2 after intestinal uptake into the blood circulation; however, the specific mechanism is not clear.
6. Conclusions
In conclusion, feeding an 8% mulberry leaf diet may inhibit backfat deposition in Yuxi black pigs by inhibiting the mRNA relative expression of ACC of the DNL pathway, promoting leptin synthesis, affecting the composition ratio of C18:1 and C18:2 in lipid metabolism by promoting LPL, FABP4, and DGAT1 mRNA relative expression in muscle tissue, and finally affecting IMF deposition. The deeper mechanisms of follow-up are not yet explored . In addition, mulberry leaves may also affect pork fat deposition by regulating intestinal microbiota and intestinal metabolites. These results show that the improvement of meat quality by mulberry leaves is obvious and also lays the foundation for further exploration of the specific mechanism of mulberry leaf regulation of meat quality and the specific components of mulberry leaves that play a regulatory role.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Conceptualization, Junjie Hou and Xiang Ji; methodology, Junjie Hou; software, Junjie Hou; validation, Zhen Song, Binjie Wang and Kangle Sun; formal analysis, Haibo Wei; investigation, Fengyun Wen; resources, Fengyun Wen; data curation, Junjie Hou; writing—original draft preparation, Junjie Hou;.
Funding
Please add: This work was supported by the grants from Program of the National Natural Science Foundation of China (U-1804118), a Program of the Henan Province Higher Education Key Teacher Training (2018-GGJS052), Science and technology project of the Education Department of Henan Province (222102110088).
Institutional Review Board Statement
The animal study was approved by the Institutional Animal Ethics Committee of Henan University of Science and Technology (approval 2022-02-023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data were available on request.
Acknowledgments
We sincerely thank Henan sangbio Agricultural Technology Co for providing fermented mulberry leaves, and Yuxi black pigs provided by Luoyang Yiluan Agricultural and Livestock Development C.
Conflicts of Interest
The authors declare no conflict of interest.
References
- González-García, S.; Esteve-Llorens, X.; Moreira, M.T.; Feijoo, G. Carbon footprint and nutritional quality of different human dietary choices. Sci Total Environ 2018, 644, 77–94. [Google Scholar] [CrossRef]
- Reig, M.; Aristoy, M.C.; Toldrá, F. Variability in the contents of pork meat nutrients and how it may affect food composition databases. Food Chem 2013, 140, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.M.; Smith, L.M. Review: Smart agri-systems for the pig industry. Animal 2022, 16 Suppl 2, 100518. [Google Scholar] [CrossRef]
- Kong, C.; Yang, L.; Gong, H.; Wang, L.; Li, H.; Li, Y.; Wei, B.; Nima, C.; Deji, Y.; Zhao, S.; et al. Dietary and Food Consumption Patterns and Their Associated Factors in the Tibetan Plateau Population: Results from 73 Counties with Agriculture and Animal Husbandry in Tibet, China. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Malgwi, I.H.; Halas, V.; Grünvald, P.; Schiavon, S.; Jócsák, I. Genes Related to Fat Metabolism in Pigs and Intramuscular Fat Content of Pork: A Focus on Nutrigenetics and Nutrigenomics. Animals (Basel) 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Fortin, A.; Robertson, W.M.; Tong, A.K. The eating quality of Canadian pork and its relationship with intramuscular fat. Meat Sci 2005, 69, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Y.; Wu, Z.; Xiong, X.; Zhang, J.; Ma, J.; Xiao, S.; Huang, L.; Yang, B. Subcutaneous and intramuscular fat transcriptomes show large differences in network organization and associations with adipose traits in pigs. Sci China Life Sci 2021, 64, 1732–1746. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, J.; Francisco, A.; Portugal, A.P.; Paulos, K.; Dentinho, M.T.; Almeida, J.M.; Regedor, L.; Fialho, L.; Cachucho, L.; Jerónimo, E.; et al. Effects of partial substitution of grain by agroindustrial byproducts and sunflower seed supplementation in beef haylage-based finisher diets on growth, in vitro methane production and carcass and meat quality. Meat Sci 2022, 188, 108782. [Google Scholar] [CrossRef] [PubMed]
- Fixen, P.E.; Johnston, A.M. World fertilizer nutrient reserves: a view to the future. J Sci Food Agric 2012, 92, 1001–1005. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Q.; Zhu, S.; Liu, B.; Liu, F.; Xu, Y. Mulberry leaf (Morus alba L.): A review of its potential influences in mechanisms of action on metabolic diseases. Pharmacol Res 2022, 175, 106029. [Google Scholar] [CrossRef]
- Ma, G.; Chai, X.; Hou, G.; Zhao, F.; Meng, Q. Phytochemistry, bioactivities and future prospects of mulberry leaves: A review. Food Chem 2022, 372, 131335. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Peng, Y.; He, J.; Xiao, D.; Chen, C.; Li, F.; Huang, R.; Yin, Y. Dietary mulberry leaf powder affects growth performance, carcass traits and meat quality in finishing pigs. J Anim Physiol Anim Nutr (Berl) 2019, 103, 1934–1945. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Xiao, Y.; Peng, Y.; He, J.; Chen, C.; Xiao, D.; Yin, Y.; Li, F. Mulberry leaf powder regulates antioxidative capacity and lipid metabolism in finishing pigs. Anim Nutr 2021, 7, 421–429. [Google Scholar] [CrossRef]
- Parida, I.S.; Takasu, S.; Nakagawa, K. A comprehensive review on the production, pharmacokinetics and health benefits of mulberry leaf iminosugars: Main focus on 1-deoxynojirimycin, d-fagomine, and 2-O-ɑ-d-galactopyranosyl-DNJ. Crit Rev Food Sci Nutr 2023, 63, 3468–3496. [Google Scholar] [CrossRef]
- Cui, W.; Luo, K.; Xiao, Q.; Sun, Z.; Wang, Y.; Cui, C.; Chen, F.; Xu, B.; Shen, W.; Wan, F.; et al. Effect of mulberry leaf or mulberry leaf extract on glycemic traits: a systematic review and meta-analysis. Food Funct 2023, 14, 1277–1289. [Google Scholar] [CrossRef]
- Fan, L.; Peng, Y.; Wu, D.; Hu, J.; Shi, X.; Yang, G.; Li, X. Dietary supplementation of Morus nigra L. leaves decrease fat mass partially through elevating leptin-stimulated lipolysis in pig model. J Ethnopharmacol 2020, 249, 112416. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev 2017, 81. [Google Scholar] [CrossRef] [PubMed]
- Edwards, B.R. Lipid Biogeochemistry and Modern Lipidomic Techniques. Ann Rev Mar Sci 2023, 15, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, W.; Li, Z.; Xia, Y.; Ouyang, Z. Enabling High Structural Specificity to Lipidomics by Coupling Photochemical Derivatization with Tandem Mass Spectrometry. Acc Chem Res 2021, 54, 3873–3882. [Google Scholar] [CrossRef]
- Hillesheim, E.; Brennan, L. Distinct patterns of personalised dietary advice delivered by a metabotype framework similarly improve dietary quality and metabolic health parameters: secondary analysis of a randomised controlled trial. Front Nutr 2023, 10, 1282741. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat Bioinforma Biomath 2013, 3, 71–85. [Google Scholar] [PubMed]
- Liu, W.; Chen, L.; Miao, K.; You, Y.; Li, J.; Lu, J.; Zhang, Y. Identification and validation of diagnostic biomarkers for intrahepatic cholestasis of pregnancy based on untargeted and targeted metabolomics analyses of urine metabolite profiles. BMC Pregnancy Childbirth 2023, 23, 828. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, L.; Mi, L. Effects of Leymus chinensis hay and alfalfa hay on growth performance, rumen microbiota, and untargeted metabolomics of meat in lambs. Front Vet Sci 2023, 10, 1256903. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, H.; Billon, Y.; Brossard, L.; Faure, J.; Gatellier, P.; Gondret, F.; Labussière, E.; Lebret, B.; Lefaucheur, L.; Le Floch, N.; et al. Review: divergent selection for residual feed intake in the growing pig. Animal 2017, 11, 1427–1439. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiong, X.; Wu, X.; Liu, G.; Zhou, K.; Yin, Y. Effects of stocking density on growth performance, blood parameters and immunity of growing pigs. Anim Nutr 2020, 6, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liao, S.; Pang, D.; Li, E.; Liu, T.; Liu, F.; Zou, Y. The transported active mulberry leaf phenolics inhibited adipogenesis through PPAR-γ and Leptin signaling pathway. J Food Biochem 2022, 46, e14270. [Google Scholar] [CrossRef]
- He, L.; Xing, Y.; Ren, X.; Zheng, M.; Yu, S.; Wang, Y.; Xiu, Z.; Dong, Y. Mulberry Leaf Extract Improves Metabolic Syndrome by Alleviating Lipid Accumulation In Vitro and In Vivo. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Huang, T.; Jing, R.; Wen, J.; Cao, M. Mulberry leaf extract reduces abdominal fat deposition via adenosine-activated protein kinase/sterol regulatory element binding protein-1c/acetyl-CoA carboxylase signaling pathway in female Arbor Acre broilers. Poult Sci 2023, 102, 102638. [Google Scholar] [CrossRef]
- Li, R.; Xue, Z.; Li, S.; Zhou, J.; Liu, J.; Zhang, M.; Panichayupakaranant, P.; Chen, H. Mulberry leaf polysaccharides ameliorate obesity through activation of brown adipose tissue and modulation of the gut microbiota in high-fat diet fed mice. Food Funct 2022, 13, 561–573. [Google Scholar] [CrossRef]
- Li, R.; Xue, Z.; Jia, Y.; Wang, Y.; Li, S.; Zhou, J.; Liu, J.; Zhang, M.; He, C.; Chen, H. Polysaccharides from mulberry (Morus alba L.) leaf prevents obesity by inhibiting pancreatic lipase in high-fat diet induced mice. Int J Biol Macromol 2021, 192, 452–460. [Google Scholar] [CrossRef]
- Cesar, A.S.; Regitano, L.C.; Koltes, J.E.; Fritz-Waters, E.R.; Lanna, D.P.; Gasparin, G.; Mourão, G.B.; Oliveira, P.S.; Reecy, J.M.; Coutinho, L.L. Putative regulatory factors associated with intramuscular fat content. PLoS One 2015, 10, e0128350. [Google Scholar] [CrossRef] [PubMed]
- Moisá, S.J.; Shike, D.W.; Faulkner, D.B.; Meteer, W.T.; Keisler, D.; Loor, J.J. Central Role of the PPARγ Gene Network in Coordinating Beef Cattle Intramuscular Adipogenesis in Response to Weaning Age and Nutrition. Gene Regul Syst Bio 2014, 8, 17–32. [Google Scholar] [CrossRef]
- Naowaboot, J.; Chung, C.H.; Pannangpetch, P.; Choi, R.; Kim, B.H.; Lee, M.Y.; Kukongviriyapan, U. Mulberry leaf extract increases adiponectin in murine 3T3-L1 adipocytes. Nutr Res 2012, 32, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yuan, W.; Peng, X.; Wang, M.; Xiao, J.; Wu, C.; Luo, L. PPAR γ/Nnat/NF-κB Axis Involved in Promoting Effects of Adiponectin on Preadipocyte Differentiation. Mediators Inflamm 2019, 2019, 5618023. [Google Scholar] [CrossRef] [PubMed]
- Boss, M.; Kemmerer, M.; Brüne, B.; Namgaladze, D. FABP4 inhibition suppresses PPARγ activity and VLDL-induced foam cell formation in IL-4-polarized human macrophages. Atherosclerosis 2015, 240, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.Z.; Fang, J.C.; Zhang, J.S.; Zhang, L.M.; Xu, C.; Xu, H.Y.; Shao, J.; Xia, G.J. Correlations between single nucleotide polymorphisms in FABP4 and meat quality and lipid metabolism gene expression in Yanbian yellow cattle. PLoS One 2020, 15, e0234328. [Google Scholar] [CrossRef]
- Opgenorth, J.; Sordillo, L.M.; VandeHaar, M.J. Colostrum supplementation with n-3 fatty acids and α-tocopherol alters plasma polyunsaturated fatty acid profile and decreases an indicator of oxidative stress in newborn calves. J Dairy Sci 2020, 103, 3545–3553. [Google Scholar] [CrossRef]
- Yagoubi, Y.; Joy, M.; Ripoll, G.; Mahouachi, M.; Bertolín, J.R.; Atti, N. Rosemary distillation residues reduce lipid oxidation, increase alpha-tocopherol content and improve fatty acid profile of lamb meat. Meat Sci 2018, 136, 23–29. [Google Scholar] [CrossRef]
- Fu, Y.; Cao, S.; Yang, L.; Li, Z. Flavor formation based on lipid in meat and meat products: A review. J Food Biochem 2022, 46, e14439. [Google Scholar] [CrossRef]
- Mapiye, C.; Aldai, N.; Turner, T.D.; Aalhus, J.L.; Rolland, D.C.; Kramer, J.K.; Dugan, M.E. The labile lipid fraction of meat: from perceived disease and waste to health and opportunity. Meat Sci 2012, 92, 210–220. [Google Scholar] [CrossRef]
- Mariutti, L.R.; Bragagnolo, N. Influence of salt on lipid oxidation in meat and seafood products: A review. Food Res Int 2017, 94, 90–100. [Google Scholar] [CrossRef]
- Bloukas, I.; Honikel, K.O. The influence of mincing and temperature of storage on the oxidation of pork back fat and its effect on water- and fat-binding in finely comminuted batters. Meat Sci 1992, 32, 215–227. [Google Scholar] [CrossRef]
- Zheng, B.; Li, X.; Hao, J.; Xu, D. Meat systems produced with Monascus pigment water-in-oil-in-water multiple emulsion as pork fat replacers. Food Chem 2023, 402, 134080. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Gross, R.W. The foundations and development of lipidomics. J Lipid Res 2022, 63, 100164. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.K.; Catanese, S.; Emond, P.; Corcia, P.; Blasco, H.; Pisella, P.J. Metabolomics and lipidomics approaches in human tears: A systematic review. Surv Ophthalmol 2022, 67, 1229–1243. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, B.; Lam, S.M.; Shui, G. Integration of lipidomics and metabolomics for in-depth understanding of cellular mechanism and disease progression. J Genet Genomics 2020, 47, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; You, R.; Lv, Y.; Liu, H.; Yang, G. Conjugated linoleic acid regulates adipocyte fatty acid binding protein expression via peroxisome proliferator-activated receptor α signaling pathway and increases intramuscular fat content. Front Nutr 2022, 9, 1029864. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Mai, K.; Ai, Q. Palmitic acid activates NLRP3 inflammasome through NF-κB and AMPK-mitophagy-ROS pathways to induce IL-1β production in large yellow croaker (Larimichthys crocea). Biochim Biophys Acta Mol Cell Biol Lipids 2023, 159428. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.R.; Teng, F.Y.; Fan, W.; Xu, B.T.; Li, X.Y.; Tan, X.Z.; Guo, M.; Gao, C.L.; Zhang, C.X.; Jiang, Z.Z.; et al. BDH1-mediated βOHB metabolism ameliorates diabetic kidney disease by activation of NRF2-mediated antioxidative pathway. Aging (Albany NY) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Liu, X.; Xu, J.; Li, F.; Wang, J.; Zhang, X.; Yang, X.; Wang, L.; Ma, S.; Li, D.; et al. Effects of Silage Diet on Meat Quality through Shaping Gut Microbiota in Finishing Pigs. Microbiol Spectr 2023, 11, e0241622. [Google Scholar] [CrossRef]
- Wunderling, K.; Zurkovic, J.; Zink, F.; Kuerschner, L.; Thiele, C. Triglyceride cycling enables modification of stored fatty acids. Nat Metab 2023, 5, 699–709. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Wang, R.; Ji, G.; Elmassry, M.M.; Zabet-Moghaddam, M.; Vellers, H.; Hamood, A.N.; Gong, X.; Mirzaei, P.; Sang, S.; et al. Dietary supplementation of gingerols- and shogaols-enriched ginger root extract attenuate pain-associated behaviors while modulating gut microbiota and metabolites in rats with spinal nerve ligation. J Nutr Biochem 2022, 100, 108904. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cao, M.; Wei, Z.; Mei, J.; Zhang, Y.; Li, M.; Li, M.; Zhang, Y.; Wang, Z. The protective effect of Buzhong Yiqi decoction on ischemic stroke mice and the mechanism of gut microbiota. Front Neurosci 2022, 16, 956620. [Google Scholar] [CrossRef]
- Zhang, H.; Qin, S.; Zhang, X.; Du, P.; Zhu, Y.; Huang, Y.; Michiels, J.; Zeng, Q.; Chen, W. Dietary resistant starch alleviates Escherichia coli-induced bone loss in meat ducks by promoting short-chain fatty acid production and inhibiting Malt1/NF-κB inflammasome activation. J Anim Sci Biotechnol 2022, 13, 92. [Google Scholar] [CrossRef]
- Krishnamurthy, H.K.; Pereira, M.; Bosco, J.; George, J.; Jayaraman, V.; Krishna, K.; Wang, T.; Bei, K.; Rajasekaran, J.J. Gut commensals and their metabolites in health and disease. Front Microbiol 2023, 14, 1244293. [Google Scholar] [CrossRef]
Figure 1.
Effects of mulberry leaves on backfat and IMF. (A) Determination of IMF content in the CON and ML groups. (B) TG measurement in the muscles of the CON and ML groups. (C) Determination of backfat thickness in the CON and ML groups. (D) Determination of TG in the backfat of the CON and ML groups. (E) Oil red O staining, BODIPY staining of muscle tissue, and H&E staining of the backfat. (F) Determination of lipid droplet area in the muscle. (G) Determination of the diameter of adipocytes in the backfat. Results are presented as mean ± SEM, n = 8. *P < 0.05, **P < 0.01.
Figure 1.
Effects of mulberry leaves on backfat and IMF. (A) Determination of IMF content in the CON and ML groups. (B) TG measurement in the muscles of the CON and ML groups. (C) Determination of backfat thickness in the CON and ML groups. (D) Determination of TG in the backfat of the CON and ML groups. (E) Oil red O staining, BODIPY staining of muscle tissue, and H&E staining of the backfat. (F) Determination of lipid droplet area in the muscle. (G) Determination of the diameter of adipocytes in the backfat. Results are presented as mean ± SEM, n = 8. *P < 0.05, **P < 0.01.
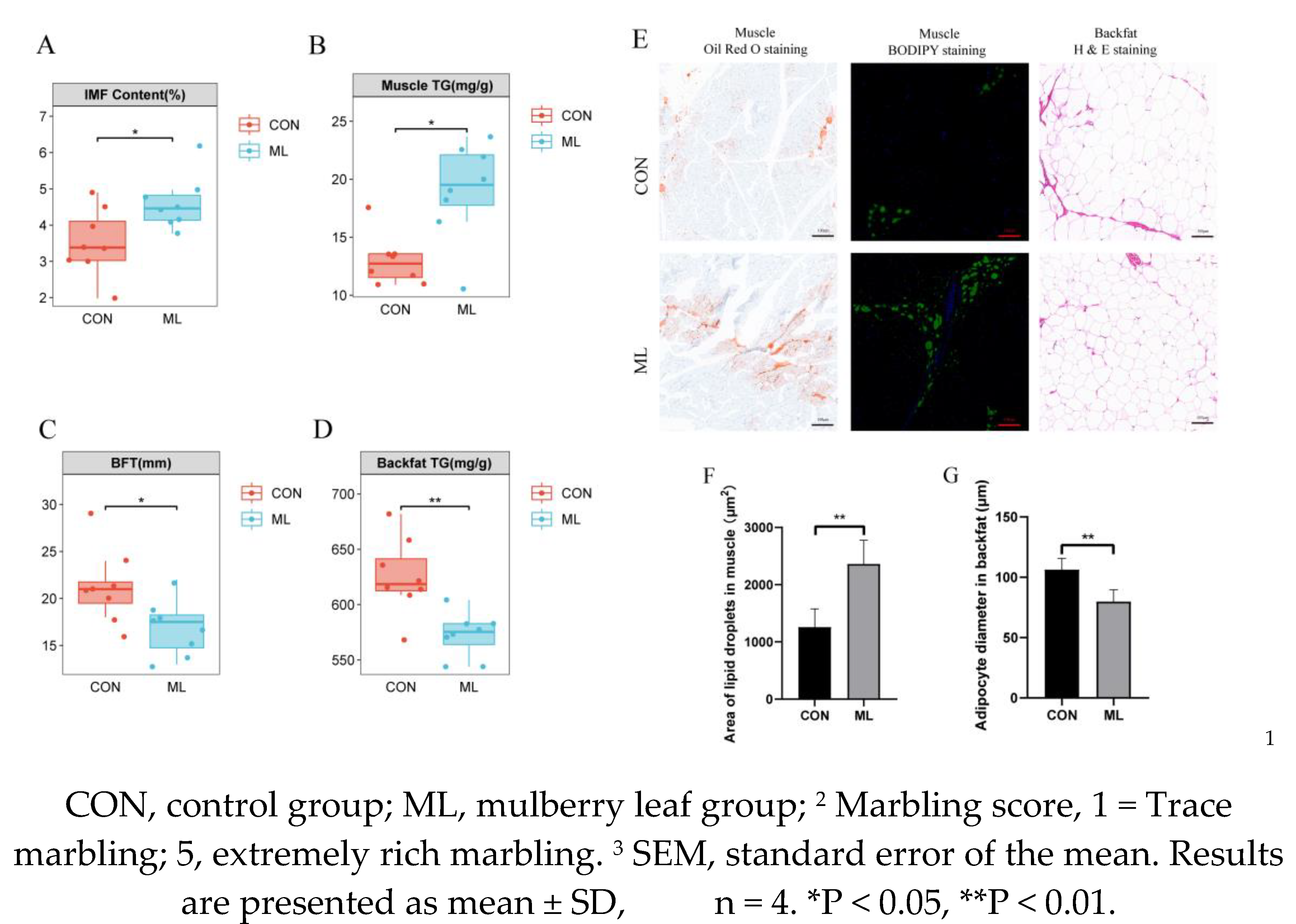
Figure 2.
Effect of mulberry leaves on muscle lipid profile composition. (A) PCA analysis. (B, C) Proportions of different lipid species in the CON and ML groups. (D) Abundance of different lipid species in the CON and ML groups. (E–I) Proportions of glycerolipids (E), glycerophospholipids (F), sphingolipids (G), fatty acids (H), and saccharolipids (I) in CON and ML groups. (J–N) Abundance of glycerolipids (J), glycerophospholipids (K), sphingolipids (L), fatty acids (M), and saccharolipids (N) in the CON and ML groups.
Figure 2.
Effect of mulberry leaves on muscle lipid profile composition. (A) PCA analysis. (B, C) Proportions of different lipid species in the CON and ML groups. (D) Abundance of different lipid species in the CON and ML groups. (E–I) Proportions of glycerolipids (E), glycerophospholipids (F), sphingolipids (G), fatty acids (H), and saccharolipids (I) in CON and ML groups. (J–N) Abundance of glycerolipids (J), glycerophospholipids (K), sphingolipids (L), fatty acids (M), and saccharolipids (N) in the CON and ML groups.
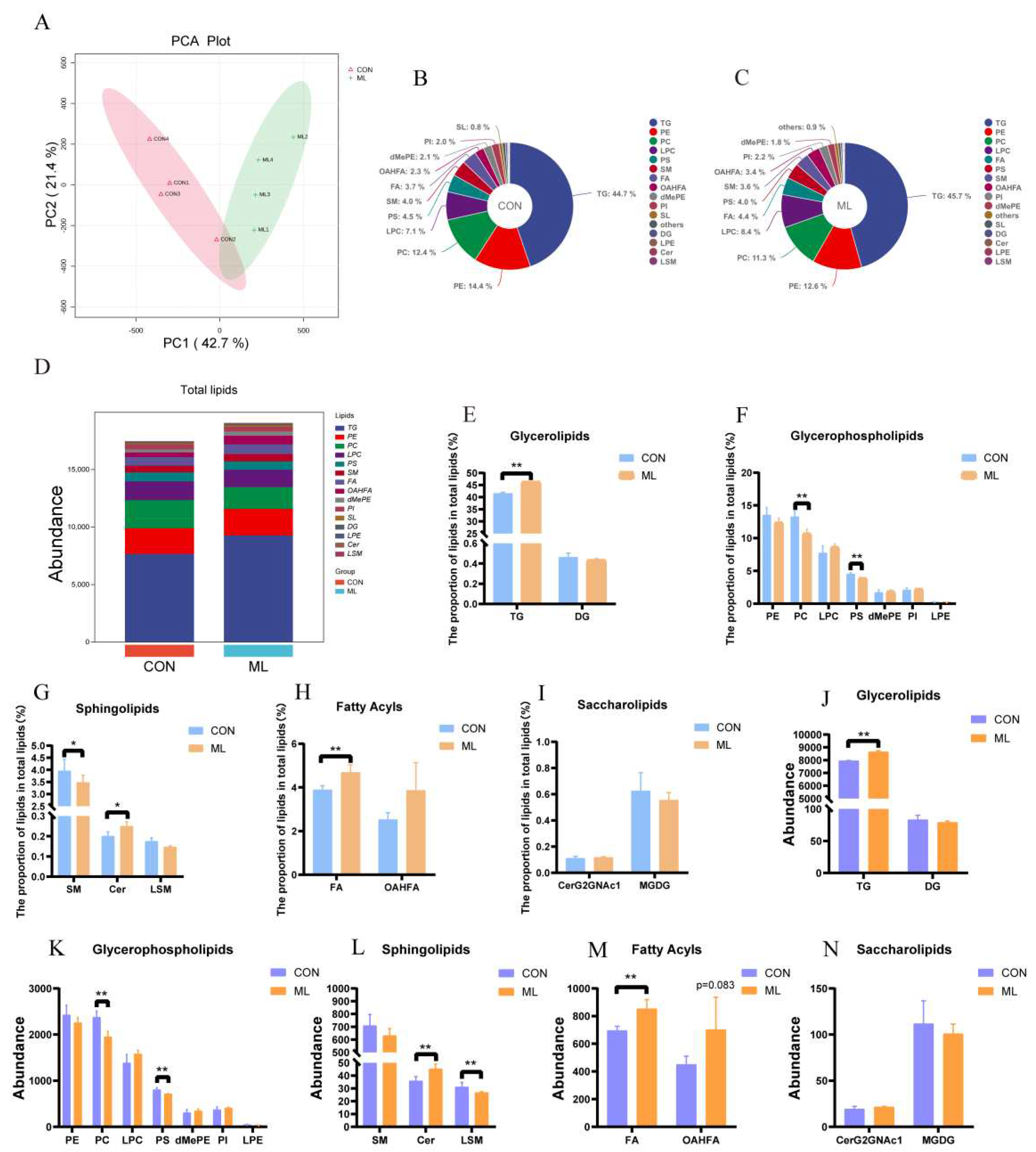
Figure 3.
Effects of mulberry leaves on muscle lipid molecules (A) Volcano plots of lipid molecules. (B) Heat map of top 20 differentially expressed lipid molecules. (C) Heat map of correlations for different lipid species. (D–J) Heat maps of GL (D), PS and PI (E), LPC and LPE (F), PC (G), PE (H), SP (I), and other lipids (J) in the CON and ML groups. (K) Heat map of the correlation between the acyl groups of TG. (L) Heat map of the correlation between the acyl groups of TG, IMF, and lipid species.
Figure 3.
Effects of mulberry leaves on muscle lipid molecules (A) Volcano plots of lipid molecules. (B) Heat map of top 20 differentially expressed lipid molecules. (C) Heat map of correlations for different lipid species. (D–J) Heat maps of GL (D), PS and PI (E), LPC and LPE (F), PC (G), PE (H), SP (I), and other lipids (J) in the CON and ML groups. (K) Heat map of the correlation between the acyl groups of TG. (L) Heat map of the correlation between the acyl groups of TG, IMF, and lipid species.
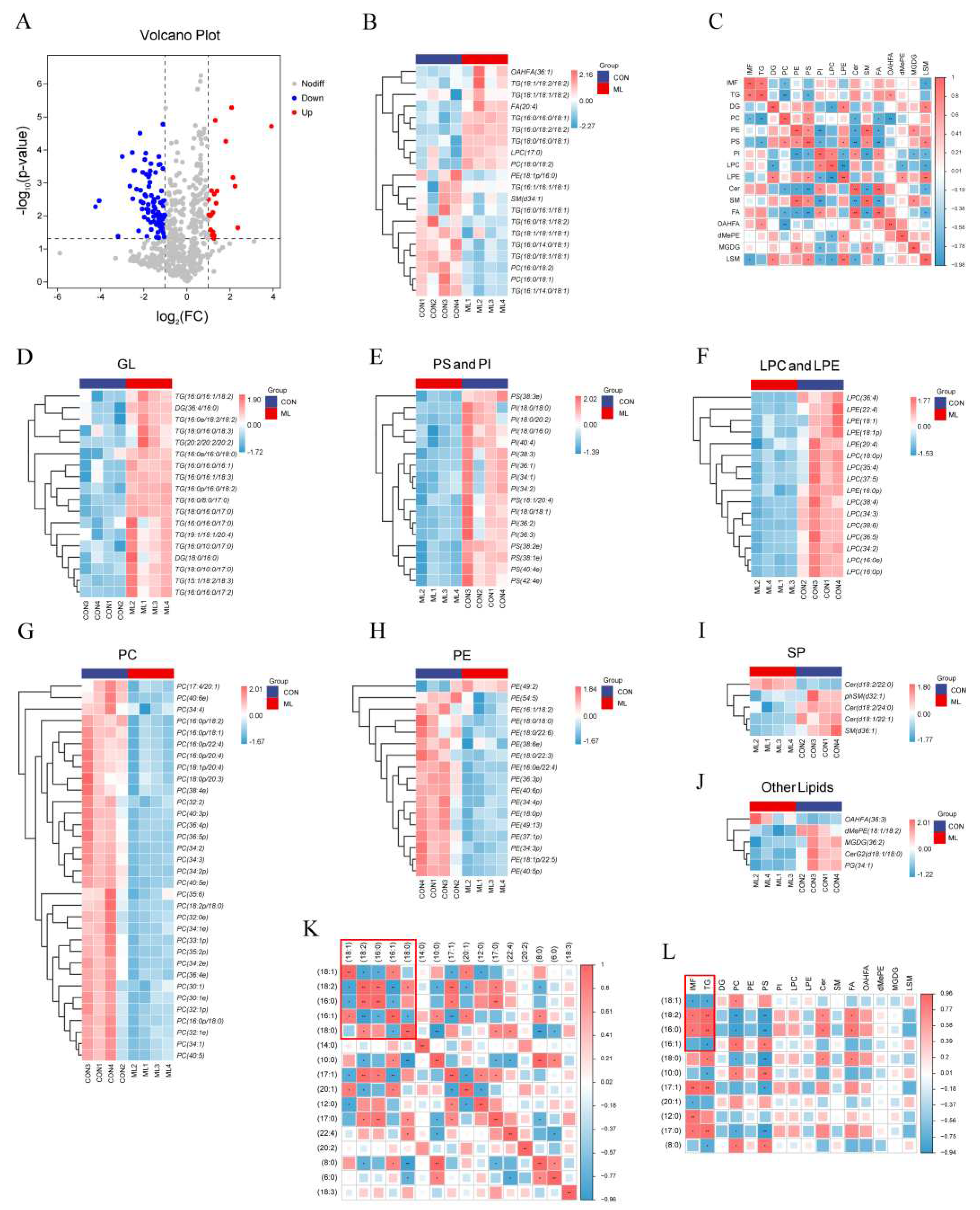
Figure 4.
Effects of mulberry leaves on cecum microbes. (A) The proportion of bacteria at the phylum level. (B) Bacterial abundance at the phylum level. (C) The proportion of bacteria at the genus level. (D) Heat map of the top 30 most abundant bacteria. (E) Heat map showing the correlation between the differential bacteria and lipid species. (F) Heat map of the correlation between the differential bacteria and acyl groups of TG.
Figure 4.
Effects of mulberry leaves on cecum microbes. (A) The proportion of bacteria at the phylum level. (B) Bacterial abundance at the phylum level. (C) The proportion of bacteria at the genus level. (D) Heat map of the top 30 most abundant bacteria. (E) Heat map showing the correlation between the differential bacteria and lipid species. (F) Heat map of the correlation between the differential bacteria and acyl groups of TG.
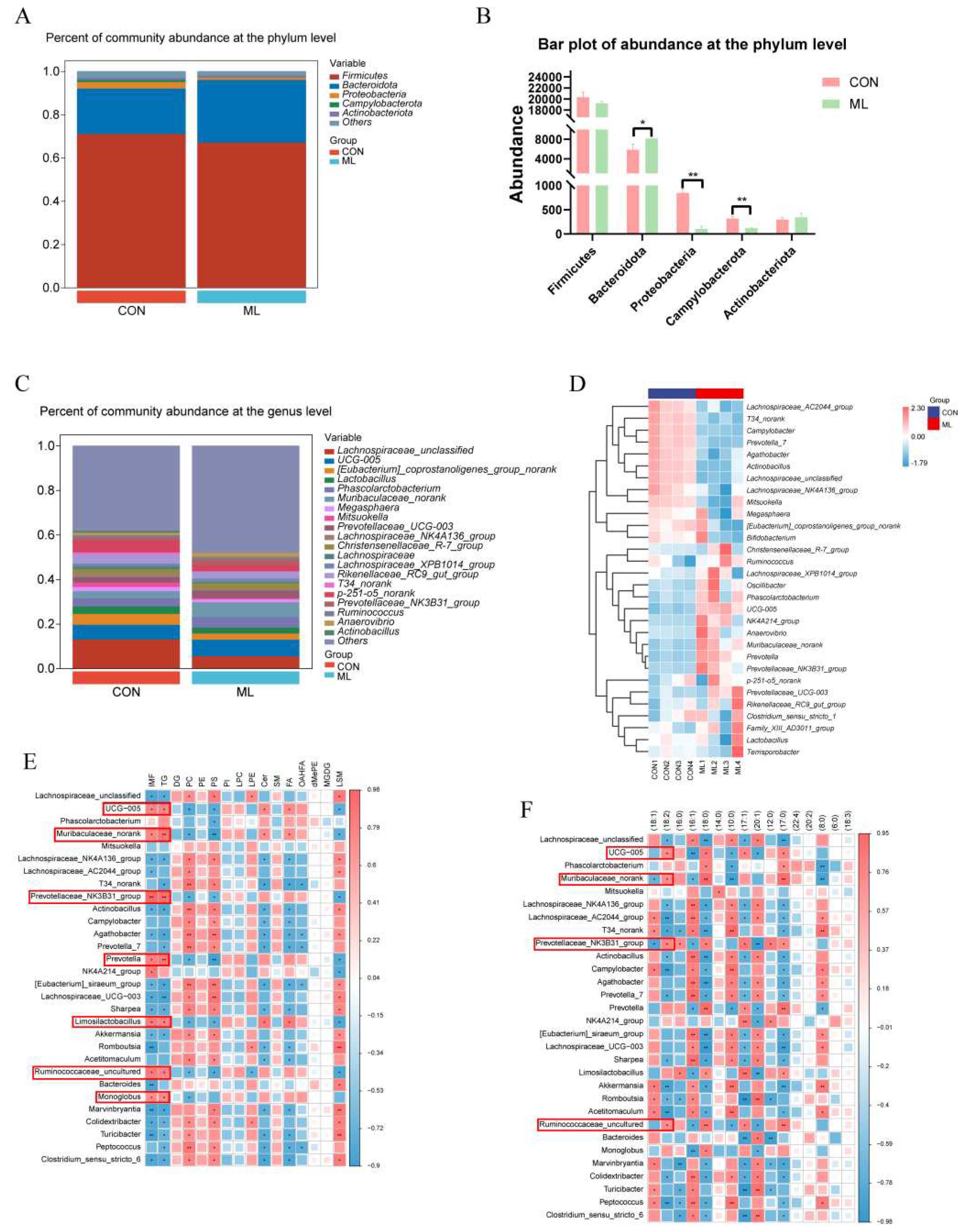
Figure 5.
Effects of mulberry leaves on cecal metabolites. (A) Proportional composition of cecal metabolites. (B) Volcano plot of cecal metabolites. (C) Heat map of top 30 differentially expressed metabolites. (D–L) Heat map of various differential cecal metabolites. (M) Heat map of the correlation between differential cecal metabolites and lipid species. (N) Heat map of the correlation between differential cecal metabolites and acyl groups of TG.
Figure 5.
Effects of mulberry leaves on cecal metabolites. (A) Proportional composition of cecal metabolites. (B) Volcano plot of cecal metabolites. (C) Heat map of top 30 differentially expressed metabolites. (D–L) Heat map of various differential cecal metabolites. (M) Heat map of the correlation between differential cecal metabolites and lipid species. (N) Heat map of the correlation between differential cecal metabolites and acyl groups of TG.

Figure 6.
Effect of mulberry leaves on the expression of key genes involved in lipid metabolism and serum indicators. (A–D) Effect of mulberry leaves on the relative expression of ACC, FASN, PPARα, and HSL in the backfat. (E–J) Effects of mulberry leaves on the relative expression of ACC, FASN, DGAT1, PPARγ, LPL, and FABP4 in the muscle. (K–P) Effects of mulberry leaves on serum FFA, LDL-C, HDL-C, insulin, adiponectin, and leptin.
Figure 6.
Effect of mulberry leaves on the expression of key genes involved in lipid metabolism and serum indicators. (A–D) Effect of mulberry leaves on the relative expression of ACC, FASN, PPARα, and HSL in the backfat. (E–J) Effects of mulberry leaves on the relative expression of ACC, FASN, DGAT1, PPARγ, LPL, and FABP4 in the muscle. (K–P) Effects of mulberry leaves on serum FFA, LDL-C, HDL-C, insulin, adiponectin, and leptin.
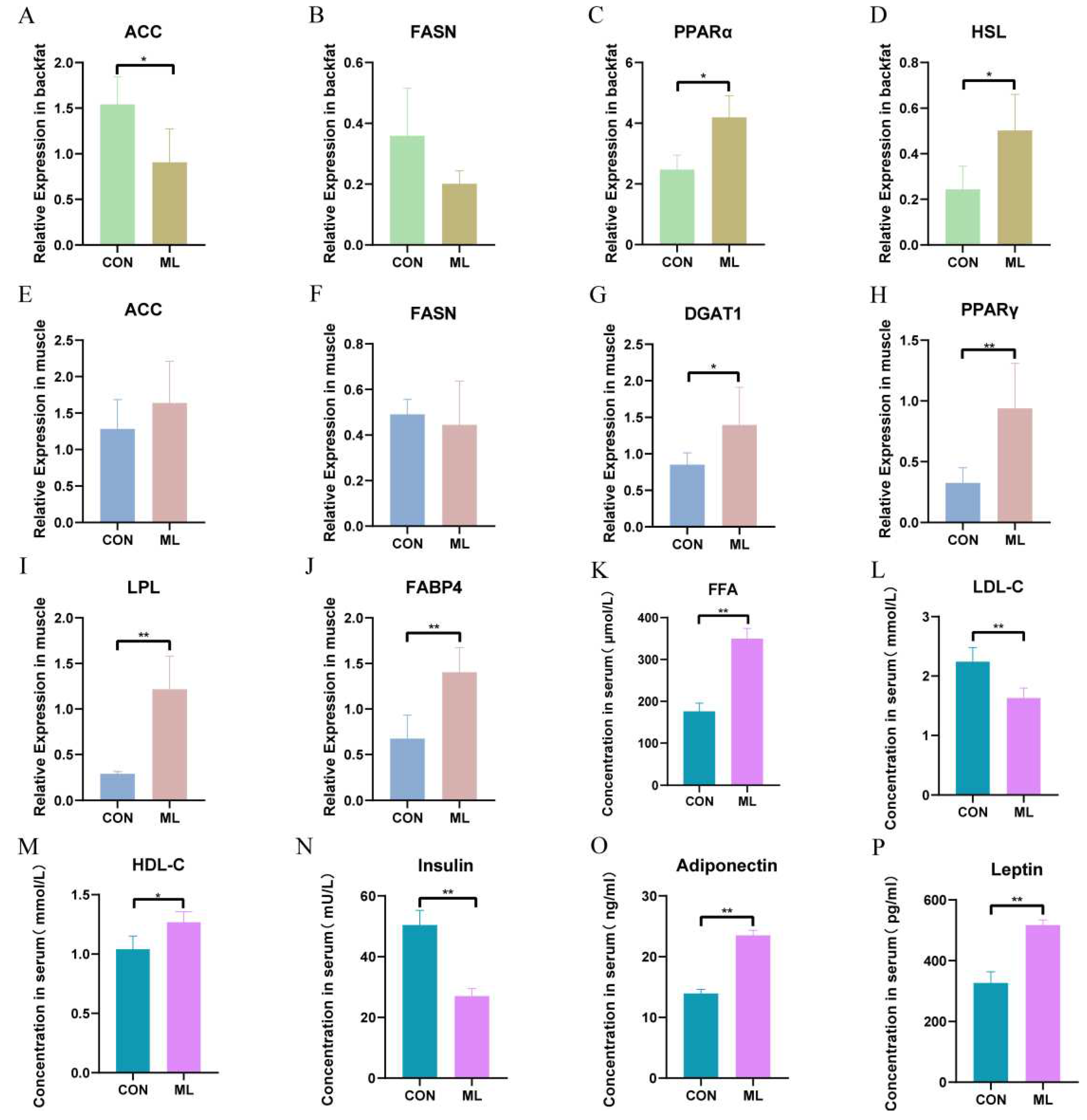
Figure 7.
A working model of the effect of mulberry leaves on the nutritional quality of fat in Yuxiblack pigs.
Figure 7.
A working model of the effect of mulberry leaves on the nutritional quality of fat in Yuxiblack pigs.
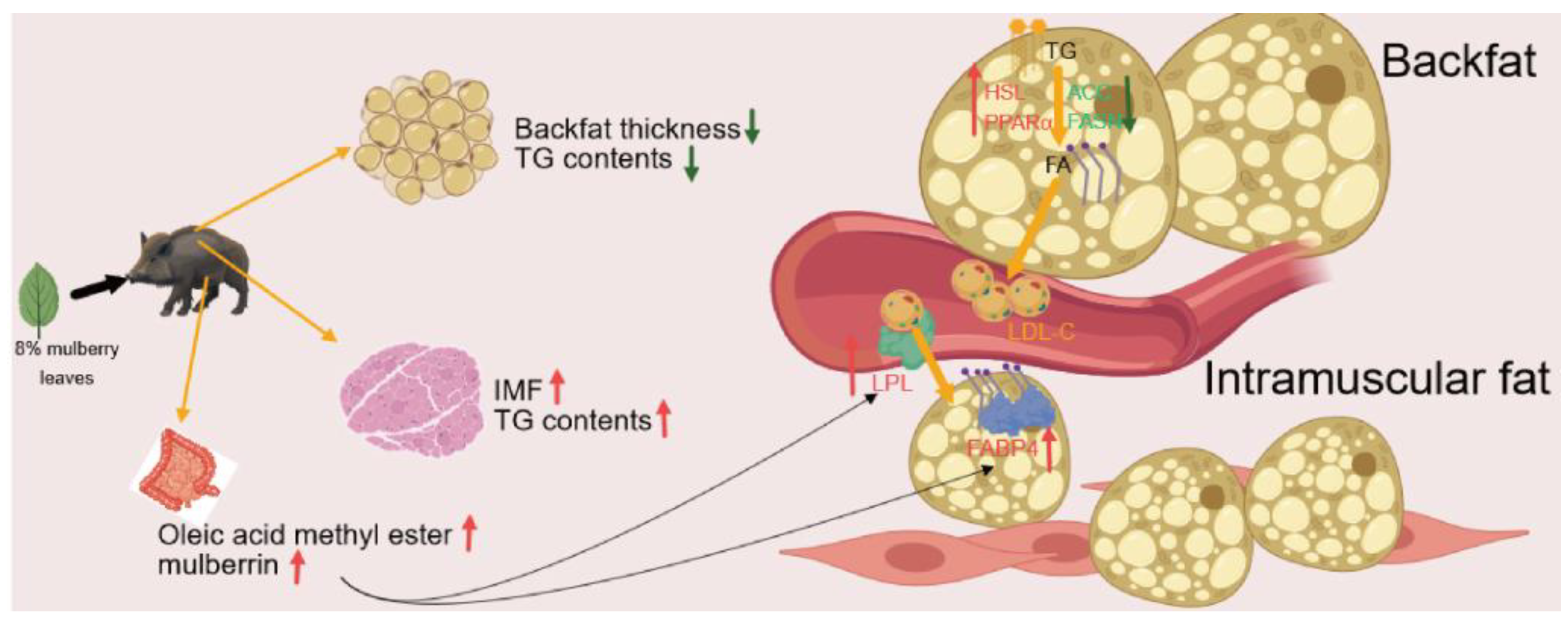

Table 1.
Effects of mulberry leaves on the meat quality of Yuxi pigs1.
| Meat quality traits | CON | ML | SEM3 | P-Value |
| Lightness(L*) | 50.182 ± 1.595 | 52.652 ± 0.929* | 0.923 | 0.036 |
| Redness(a*) | 13.770 ± 1.435 | 13.210 ± 1.686 | 1.107 | 0.633 |
| Yellowness(b*) | 6.175 ± 1.786 | 5.255 ± 1.638 | 1.212 | 0.476 |
| Marbling score2 | 3.275 ± 0.320 | 4.075 ± 0.435* | 0.270 | 0.025 |
| pH | 6.260 ± 0.232 | 6.118 ± 0.284 | 0.183 | 0.467 |
| Shear force | 19.73 ± 2.275 | 21.28 ± 1.359 | 1.325 | 0.287 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



