Submitted:
18 December 2023
Posted:
19 December 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Viruses rely on host cells to replicate their genomes and assemble new viral particles. Thus, they have evolved intricate mechanisms to exploit host factors. Host cells, in turn, have developed strategies to inhibit viruses, resulting in a nuanced interplay of co-evolution between virus and host. This dynamic often involves competition for resources crucial for both host cell survival and virus replication. Iron and iron-containing cofactors, including iron-sulfur clusters, are known to be a heavily battled resource during bacterial infections where control over iron can tug the war in favor of the pathogen or the host. It is logical to assume that viruses also engage in this competition. Surprisingly, our knowledge about how viruses utilize iron (Fe) and iron-sulfur (FeS) clusters remains limited. The handful of reviews on this topic primarily emphasize the significance of iron in supporting the host immune response against viral infections. The aim of this review, however, is to organize our current understanding of how viral proteins utilize FeS clusters, to give perspectives on what questions to ask next and to propose important avenues for future investigations.
Keywords:
iron-sulfur clusters
; HSC20
; HSPA9
; viral proteins
; SARS-CoV-2
; cytoplasmic iron-sulfur assembly machinery (CIA)
; virus-host interaction
; DNA replication and repair
1. Iron-Sulfur Clusters as Essential Cofactors in Cells from Bacteria to Human
Virtually every cell on Earth ingeniously harnesses FeS clusters as crucial cofactors in a multitude of biological processes.1, 2 FeS enzymes are integral to mitochondrial respiration, the TCA cycle, nitrogen fixation, DNA replication and repair, protein translation, heme and cofactor biosynthesis, and sensing and regulation of iron levels, among others.3-10 FeS clusters play both a structural and functional role by acting in electron transfer processes, oxidoreductive reactions, and protein conformational changes – an ability not inherently shared by other metal ions.2, 11, 12 It is likely that iron-sulfur clusters are utilized to a greater extent than our current observation suggests. These cofactors are prone to oxidative destabilization in oxygen-rich environments, such as those encountered during purification processes.1, 11, 13 While atmospheric oxygen levels hover around 21 percent, the interior of a mammalian cell typically ranges between 0.5 and 5 percent.14, 15 As a result, handling of iron-sulfur proteins following lysis of the cell requires special tools and protocols. FeS proteins are not only ubiquitous, but the roles performed by FeS clusters have remained remarkably conserved throughout evolution.16 Depending on the species, the bacterial NADH dehydrogenase complex contains seven to ten FeS clusters conserved in the eukaryotic mitochondrial respiratory complex I which typically ligates eight clusters.7, 17 Electron transfer between FeS clusters in complex I is coupled to proton pumping across the mitochondrial inner membrane, accounting for about 40% of the total protons pumped throughout the respiratory chain.7, 17 DNA replication proteins, including DNA polymerase-α, DNA polymerase-δ, DNA polymerase-ε, DNA primase, and DNA repair nucleases – NTHL1 and MUTYH – rely on iron-sulfur clusters for their imperative function in supporting genome maintenance and cell proliferation.3-6, 18
Cells have evolved intricate biosynthetic pathways to assemble FeS clusters and incorporate them into a vast array of proteins. De novo biogenesis of FeS clusters is a complex multi-step process highly conserved from bacteria to humans.16, 19, 20 In mammalian cells, the basic building block of all FeS clusters, the rhombic [2Fe-2S] cluster, is initially assembled on a main scaffold protein, ISCU.16, 19, 21-25 ISCU binds to a dimeric configuration of the cysteine desulfurase, NFS1, which enzymatically converts cysteine to alanine, while mobilizing inorganic sulfur for FeS cluster assembly.16, 19, 21, 22, 26 Specifically, in the catalytic reaction carried out by NFS1, sulfur from the thiol group of the substrate cysteine is mobilized to generate a persulfide intermediate on cysteine 138 of ISCU.23 NFS1 is stabilized by its essential binding partner ISD11.27-29 The identity of the iron donor for FeS cluster assembly in mitochondria remains controversial in the field. In the cytosol, PCBP1 and its binding partner, BOLA2, have been found to provide iron for [2Fe-2S] cluster assembly.30 PCBP1 can coordinate up to three ferrous iron ions and has been shown to function as a chaperone that delivers iron to the storage protein ferritin as well as to other cytosolic iron proteins.31-34 Additionally, two electrons are needed for [2Fe-2S] cluster assembly. In yeast and mitochondria, these reducing equivalents are provided by the ferredoxin/ferredoxin reductase complex.35, 36 The [2Fe-2S] cluster can then be utilized to generate much more complex FeS cofactors with different stoichiometries of iron and sulfur, including the most common the cubane [4Fe-4S] cluster (Figure 1A).1, 37 FeS clusters of more complex structures can be assembled on secondary carriers which in turn deliver the cofactors to subsets of recipient FeS proteins.16 Cubane [4Fe-4S] clusters can be generated by the reductive coupling of two [2Fe-2S] clusters37 and are commonly found in DNA and RNA enzymes where changes in the oxidation state of [4Fe-4S] clusters have been proposed to modulate affinity of binding to the DNA or RNA polyanion.3-6, 38 Cubane clusters function in complex electron relay systems where they allow facile electron transfer that is not coupled to changes in the cluster’s conformation due to the delocalization of electrons between iron atoms on the two planes of the cubic cluster.1, 39
Transfer of newly assembled FeS clusters to recipient apo-proteins is assisted and enhanced by an evolutionarily conserved chaperone/co-chaperone system.40-43 In human cells, the ATP-dependent Hsp70 chaperone, HSPA9, and its cognate J-domain protein, HSC20 (or HSCB), facilitate transfer of FeS clusters to recipient proteins (Figure 1B).22, 42, 44 The selection process for specifically allocating FeS clusters to only FeS apoproteins among the entire proteome has represented a long-standing question in the field. One mechanism has emerged in recent years that focuses on the specificity driven by the co-chaperone, HSC20, given that the multifunctional chaperone HSPA9 has promiscuous substrate-binding activity.43, 45, 46 HSC20 was found to recognize Leucine-Tyrosine-Arginine (LYR)-like motifs present in recipient FeS apo-proteins for direct binding and delivery of a FeS cluster.22, 42, 44 The C-terminal domain of HSC20 interacts directly with the FeS-bound ISCU, which is released from the initial pre-complex with NFS1, and a Histidine-Proline-Aspartate (HPD) tripeptide in the N-terminal J-domain of HSC20 activates the ATP hydrolysis by HSPA9, thereby accelerating cluster transfer.22, 42, 44, 47-49 Once transferred to the appropriate recipients, the current understanding is that the protein folds around the FeS cluster.50 A subset of nucleocytoplasmic FeS proteins, such as those involved in DNA replication and repair, purine and pyrimidine metabolism and tRNA modifications, require the specialized Cytoplasmic Iron-sulfur Assembly (CIA) complex to acquire their cofactors (Figure 1B).10, 51-53 The CIA complex is composed of MMS19, CIAO1, and FAM96B.51-53 MMS19 is essential for genome stability and DNA metabolism51, 52 and possesses four HEAT repeat motifs that have been proposed to mimic the DNA double helix, which could provide an appropriate docking site for DNA or RNA binding proteins.54 Binding of HSC20 to the LYR motif of CIAO1 likely bridges the CIA machinery to the de novo FeS complex.53 A model has been proposed, whereby a dimer of HSC20 engages simultaneously the LYR motif of CIAO1 that is bound to MMS19 and FAM96B, and the LYR sequence of an unfolded FeS-recipient apoprotein, thereby mediating the interaction of the FeS client with the CIA complex and facilitating cluster acquisition.16, 53 Overall, the relevance of the FeS biogenesis pathway for human health and disease has become particularly apparent during the past twenty years with the identification of several ultra-rare disorders caused by loss of function in the genes that encode the highly conserved FeS biogenesis proteins.16, 19, 55 Interestingly, thus far the in vivo physiological consequences of loss of function in any of the components of the CIA machinery have remained unknown. Exome sequencing has effectively contributed to uncover novel ultra-rare human conditions through the identification of genetic variants that have enabled a better understanding of disease mechanisms and the identification of potential targets for therapeutic interventions. It is likely that in the years to come many more human conditions will emerge due to loss of function in components of the FeS biogenesis pathway, which have yet to be associated with any known human diseases.
2. Viral Genome Evolution: Harnessing the Host Machinery and FeS Clusters
Unlike bacterial pathogens, many viruses evolved as genetic minimalists to evade host targeting strategies against their viral genomes.56, 57 Capsid virions and nucleocapsids also limit genome lengths and require the genome to squeeze into small protein capids.56, 58 To overcome these feats, many viruses have evolved complicated genomes with intricate replication and transcription mechanisms. Even the larger DNA viruses, which can be larger than the genomes of the smallest bacterium, do not encode all the necessary components for the production of progeny.59, 60 Instead of incorporating all of the necessary components into their minimalist genomes, viruses smartly use the host cell machinery. This extends to their utilization of the host cell FeS cluster assembly machinery. Why invest time, energy, and genome space into something already encoded by every potential host? This review showcases five diverse virus families that have been reported to encode FeS proteins.
3. FeS Clusters in Virally Encoded Proteins of Five Distinct Virus Families
3.1. The Rotavirus Nonstructural Protein 5, NSP5
The initial discovery of a virally encoded FeS protein, the rotavirus NSP5, was likely serendipitous, recognized by the appearance of an unexpected brown color during the purification process, indicating its potential coordination of a FeS cluster.61 The rotavirus nonstructural protein 5, NSP5, has been reported to ligate a [2Fe-2S] cluster based on distinctive features observed on Mossbauer spectroscopy.61 Rotaviruses mainly infect enterocytes of the small intestine villi and can cause severe diarrhea, which can be deadly in infants and immunocompromised patients.62 No specific treatments are available for rotavirus infections.62
Rotaviruses contain eleven double-stranded RNA (dsRNA) segments inside three-layered proteinaceous virions or virus particles.63 The total genome size is about 18 kilobases (kb), and each dsRNA segment is between 600 base pairs (bp) and 3,300 bp.63 Rotavirus NSP5 and NSP2 are responsible for forming liquid-like protein-RNA condensates that provide an appropriate environment for RNA replication, known as viroplasms (Figure 2).63-65 Never exposed to the cytoplasm, the dsRNA genome is transcribed and capped by VP1, the RNA-dependent RNA polymerase (RdRp), inside virions that have lost their outermost protein layer in the lysosomal lumen.63 Transcription of the dsRNA segments by the RdRp generates positive (+) sense single-stranded RNAs (ssRNA) which are extruded from the virions to be either translated by the host cell ribosomes or to serve as templates for replication.63 In the latter case, NSP2 binds the (+)ssRNAs in a sequence-specific manner and acts as an RNA chaperone by facilitating the folding of RNA into inter-segmented intermediates (Figure 2).63 Eleven single-stranded inter-segmented RNAs, NSP2 and NSP5, VP2 and VP1 form the inner core of a virion inside the viroplasms. Replication of (+)ssRNA templates occurs within these virion cores generating the dsRNA segments of the rotavirus genome.63 NSP5 has been proposed to act as a scaffold linking replication and transcription of the (+)ssRNA by binding to VP1 and the core structure formed by VP2.61, 63, 64 The virion core is encased in the triple-protein outer layer.63 There remain several unknowns in the current model of rotavirus replication, leaving much yet to be uncovered.
A recombinant rotavirus lacking NSP5 produced by reverse genetics was unable to form viroplasms and replicate viral RNA, while infection of a cell-line expressing wild-type NSP5 was able to rescue replication of the viral RNA.66 NSP5 has an N-terminal intrinsically disordered region, two oligomerization domains, and an amphipathic alpha-helical C-terminus.64 Additionally, NSP5 has been found to harbor ATPase activity.64 Interestingly, phosphorylation of multiple serine residues has been reported to modulate NSP5 function.64, 66 Two cysteine residues in NSP5 were found to coordinate a [2Fe-2S] cluster at the interface between NSP5 dimers.61 These residues are conserved among group A, C, and D rotaviruses, raising the possibility that other rotavirus NSP5 proteins may also harbor a rhombic cluster.61 Martin et al. showed that mutagenesis of the two cysteine residues responsible for FeS ligation did not hinder viroplasm formation.61 However, the NSP5 protein devoid of the iron-sulfur cluster had different binding patterns to ssRNA.61 The wild-type NSP5 exhibited two ssRNA binding sites, as detected by microscale thermophoresis, whereas both the NSP5 with mutated cysteines (into alanines) and an “oxidized-NSP5” had only one ssRNA binding site.61 The FeS cluster in NSP5 appears to be involved in the binding of ssRNA and might help in orchestrating the complex replication of the eleven genomic segments. 61 Despite this discovery being reported in 2013, there has been limited subsequent exploration into the role of the FeS cluster in rotavirus NSP5 and the potential advantages it confers. Further investigation into the utilization of FeS clusters in rotavirus replication could provide important insights into the mechanisms underlying rotavirus replication.
3.2. The Merkel Cell Polyomavirus small T antigen (sT)
The discovery of the Merkel cell Polyomavirus small T antigen as a FeS protein was a fortuitous finding as well, highlighted by the brown color of the purified protein indicative of the presence of iron.67 Merkel cell polyomavirus (MCPyV) is a small, circular double-stranded DNA (dsDNA) virus belonging to the Polyomaviridae family (Figure 3A).68, 69 The MCPyV viral genome can clonally integrate into the host genome which is a significant event in the development of Merkel cell carcinoma (MCC), a rare but aggressive skin cancer.70, 71 Upon infection, MCPyV relies heavily on the host cell DNA replication machinery within the cell nucleus.69 The viral DNA is released directly into the host cell’s nucleus, following entry.69 Subsequently, the cellular apparatus initiates the transcription of early genes from the MCPyV genome.69 Among the early genes are the regulatory proteins large T antigen (LT) and small T antigen (sT) that play a crucial role in genome replication and MCPyV-related carcinogenesis.68
Both LT and sT have an HSP40 chaperone J-like domain at their N terminus (Figure 3B).68, 72 LT has a large helicase domain and a DNA origin binding domain (OBD).68 LT initiates genome replication by binding to seven sequence-specific sites along 71 bp of the circular genome, allowing the cellular replication enzymes to reach the DNA template and initiate replication (Figure 3C,D).69 sT was initially thought to enhance replication solely by increasing LT stability through the inhibition of the the E3 ubiquitin ligase FBXW7.67, 73 sT, a 22 kDa protein with a T-antigen-like domain on its C-terminal half (Figure 3E), was found to ligate two FeS clusters, a [2Fe-2S] and [4Fe-4S], within two metal binding pockets composed of cysteine residues, as observed by Mossbauer spectroscopy.67 Tsang et al. showed that mutating the FeS ligating residues of sT rendered the protein ineffective in stimulating LT-mediated viral DNA replication, highlighting the essential roles of these residues for sT function.67 Moreover, these studies proposed that sT can stabilize LT without altering its protein levels. sT was shown to localize to active replication sites at the viral origin where LT is known to act, indicating a more direct role of sT in viral genome replication.67 A recent paper showed by knockout studies that while sT was not essential for initiating viral DNA replication, it played a crucial role in maintaining genome integrity and activating the transcription of early and late genes, specifically LT and VP1 – the primary capsid protein.72 Investigating the role of the FeS clusters for sT function could significantly advance our understanding of the function of sT in carcinogenesis.
3.3. The SARS-CoV-2 nsp12 and nsp13
Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), the causative agent of the COVID-19 pandemic, has been found to encode two FeS proteins that are at the core of the replication and transcription complex (RTC) (Figure 4A,B).74, 75 This discovery came into focus amid the pandemic’s heightened interest. The presence of LYR-like motifs and metal-binding sites in the viral proteins enabled the prospective prediction of FeS cluster binding, subsequently experimentally confirmed, a finding that suggests the existence of FeS proteins hiding in plain sight.
SARS-CoV-2 is a coronavirus of the order Nidovirales.76-78 Nidovirus genomes are some of the longest RNA genomes, typically consisting of ~30,000 base single-stranded RNA genomes (Figure 4C).76-78 Most RNA viruses trail behind these impressive genome lengths due to factors such as low-fidelity polymerases and the inherent instability of RNA.78-80 Coronaviruses stand out by encoding an exoribonuclease, an uncommon feature among RNA viruses, to counteract mutations as they arise during replication.76, 78, 79, 81 Upon entering a host cell and releasing its genome into the cytosol, coronaviruses utilize host ribosomes to translate two polypeptides directly from the positive sense-single-stranded RNA genome from two juxtaposed open reading frames defined through a ribosome frameshifting site.78, 82 The single polypeptides are cleaved by two viral proteases, namely nsp3 and nsp5, into sixteen non-structural proteins (nsps).78, 80, 83 A significant number of nsps assemble into the multi-subunit RTC, responsible for replicating the RNA genome and transcribing structural and accessory genes.78 The viral genome includes transcriptional regulatory sequences (TRS) located between the structural and accessory genes that are complementary to a 5’ leader sequence, enabling a template switching mechanism that skips large portions of the genome during transcription (Figure 4D).76, 78, 80 This complex mechanism ensures that each transcription product or subgenomic RNA (sgRNA) acquires a polyA tail and a 5’cap allowing for evasion of host defenses.76, 78, 80, 84, 85 Moreover, this mechanism potentially accommodates the diverse copy number needs for structural and accessory proteins.78, 85 The coronavirus replication and transcription complex is responsible for incredible feats that only viruses of the same order can achieve.
The heart of the replication and transcription complex of SARS-CoV-2 is nsp12, the catalytic subunit of the RNA-dependent RNA polymerase, which adopts a right-hand conformation with a unique N-terminal nidovirus-specific nucleotidyl transferase (NiRAN) domain.84 Nsp12 was found to ligate two [4Fe-4S]2+ clusters74 in the same sites that had been modeled as zinc centers in the available cryo-EM structures of the aerobically purified complex.86 One FeS cluster is at the interface between the NiRAN and the catalytic domain and the second in the fingers subdomain within the polymerase domain (Figure 4B).74 Using an in vitro primer extension assay with the minimal requirements of nsp12 and the two accessory factors, nsp7 and nsp8, Maio and colleagues showed that the iron-sulfur cluster in the catalytic domain is essential for replication.74 Substitution of the FeS ligating residues in the polymerase domain abolished the activity of the RdRp.74 The aerobically purified form of nsp12 containing zinc had significantly reduced polymerization efficiency and reduced affinity of binding to the RNA template.74 The template tested in this experiment was 1,000 times smaller than the SARS-CoV-2 RNA genome and had minimal secondary structure.74 Under these conditions, Zn was shown to be partially able to support function by potentially serving a structural role that preserved the integrity and tridimensional structure of the catalytic domain.74
Mutagenesis of the FeS ligating residues in the interface domain of nsp12 diminished its interaction with the SARS-CoV-2 helicase nsp13 without profoundly impairing the RNA polymerase activity.74 Nsp13 is a member of the 1B superfamily of helicases (SF1B) with an ATP-dependent unwinding activity that proceeds in the 5’ to 3’ direction.87 Notably, the directionality of nsp13 opposes that of nsp12 which extends the nascent RNA template in the 3’ to 5’ direction. Available cryo-EM structures of the SARS-CoV-2 RTC show two nsp13 protomers per copy of the RdRp (Figure 4A).88, 89 While the significance of this arrangement is still under debate, it has been proposed that one of the two nsp13 protomers may allow backtracking of the RdRp and template switching, while the other would enable the unwinding of downstream RNA secondary structures.89 Nsp13 is organized into five distinctive domains: an N-terminal Zinc Binding Domain (ZBD), a beta barrel 1B domain, a helical stalk and two RecA-like domains 1A and 2A, required for nucleotide binding and hydrolysis (Figure 4B).87, 90 Intriguingly, nsp13 was also found to ligate a [4Fe-4S]2+ cluster and two Zn2+ ions in three metal binding sites within its ZBD.75 Aerobically purified nsp13 ligates three zinc ions in its ZBD and has been shown to exhibit a significantly higher activity on DNA in in vitro enzymatic assays, despite being physiologically active on RNA.91-93. To investigate the effect of the Fe-S cluster on nsp13 binding to RNA versus DNA, Maio and colleagues performed electrophoretic mobility shift assays (EMSAs) using either anoxically purified nsp13 ligating a FeS in its ZBD (Zn2-[4Fe-4S]) or nsp13 purified aerobically and fully reconstituted with zinc (Zn3). 75 Nsp13-Zn2-[4Fe-4S] bound RNA substrates with an approximately 100-fold greater affinity than nsp13-Zn3.75Additionally, nsp13-Zn3 exhibited approximately 10-fold higher affinity for DNA than for RNA, consistent with results previously reported for the aerobically purified enzyme 93. These findings demonstrate that the [4Fe-4S] cluster greatly enhances the binding selectivity of nsp13 for its physiological substrate, RNA. Consistent with the enhanced affinity of nsp13-Zn2-[4Fe-4S] for RNA compared to nsp13-Zn3, functional assays revealed that the anoxically purified nsp13 ligating the cluster had greater unwinding activity than the aerobically purified enzyme that contained three zinc ions per protomer. 75 Loss of the Fe-S cluster impaired the unwinding activity of the helicase, likely as a result of diminished binding of the variant to the substrate.
The presence of a complex metal composition with both zinc and a FeS cluster, as in nsp13, has been thus far reported only once before, for the cleavage and polyadenylation specificity factor 30 94. Interestingly, nsp13 was found to acquire its [4Fe-4S] cluster through the recruitment of its LYK motif by the Fe-S biogenesis machinery,75 whereas the mechanism of zinc delivery to the protein is likely to depend on ZNG1, which has recently been reported to recognize a unique Cys6His2 zinc finger domain in zinc-dependent metalloproteins and to function as a GTPase metallochaperone that allocates Zn to specific recipient proteins 95. The ZBD of nsp13 contains a Cys6His2 domain, organized into a RING-like module that was found to ligate two zinc ions.75 The complete conservation of the LYK motif and of the Cys6His2 zinc finger domain in nsp13 from the seven human coronaviruses suggests that other coronaviruses may rely on similar pathways to incorporate the two zinc ions and a [4Fe-4S] cluster in their ZBDs.
It is noteworthy that the Fe-S clusters present in the SARS-CoV-2 RdRp and nsp13 share a Cys3His coordination that is typical of large protein complexes in which the clusters regulate inter- and intra-molecular electron transfer rates, such as in the NDUFS1 subunit of the NADH-ubiquinone dehydrogenase complex of the respiratory chain and in the NiFe and FeFe hydrogenases 96, 97. The distal Cys3His-ligated [4Fe-4S] cluster in the NiFe hydrogenases supports electron transfer ~ 1,000 times faster than the altered cluster with Cys4 coordination 96. The [4Fe-4S] clusters in the RdRp and nsp13 may play a role in coordinating the activities of the two enzymes during viral replication via redox switch mechanisms coupled either to changes in the relative affinities for the template, to conformational rearrangements similar to those proposed for DNA processing enzymes 4, or to both.
3.4. The Hepatitis B HBx Protein
Hepatitis B virus (HBV) poses a global health burden and contributes to liver-related disease and mortality.98 The World Health Organization estimates that one third of HBV infected patients develop no symptoms, another third non-icteric hepatitis, and the remaining third jaundice-related hepatitis with nausea, fatigue, and hepatomegaly.98, 99 Fulminant hepatitis develops in less than one percent of HBV-infected individuals which can lead to severe liver damage and death.98 In addition to acute hepatitis, there are also chronic cases which are predominant if HBV infection occurred at a young age.98 In chronic HBV infection, pathogenesis typically starts with liver inflammation, followed by fibrosis, cirrhosis and ultimately formation of hepatocellular carcinoma.98-100
HBV is an enveloped virus with a partially circular dsDNA genome of 3.2 kb that primarily infects primates with a specific tropism for hepatocytes.99, 101 Upon infection, the viral DNA enters the cell nucleus102 where it is converted into an episomal circular DNA molecule by cellular DNA repair mechanisms.99, 103, 104 The closed circular DNA serves as a template for transcription of the pre-genomic RNA, which is the precursor of the viral genomic dsDNA, and of four subgenomic viral RNAs which are translocated to the cytosol where they are translated.103 Capsid core structures consist of the pre-genomic RNA, the viral polymerase (P), and the HBV core capsid protein, (HBcAg).103 In these formations, the P enzyme carries out reverse transcription of pre-genomic RNA, converting it into a partially circular dsDNA genome.103 Normally, this reverse transcription process takes place inside capsids that envelop at the endoplasmic reticulum.99, 103, 105 Nevertheless, reverse transcription can also happen within capsid structures located in intracellular regions that are shuttled to the nucleus.99, 103
HBx, also known as the HBV X protein, plays several crucial roles in inducing changes within the cell that accumulate, ultimately leading to hepatocellular pathogenesis and the development of hepatocellular carcinoma.99 While the complete structure of HBx remains elusive, an in-silico 3D-modeling study (Figure 5A) revealed significant structural similarity between HBx and a DNA glycosylase.106 The 16.5 kDa X protein consists of an N-terminal transactivator domain involved in signal transduction and a C-terminal domain required for nuclear transcriptional activation of both virally encoded and host genes (Figure 5B).99 Within the C-terminal domain there are several mitochondrial localization sequences (Figure 5B).99 Moreover, different isoforms of HBx arise from two in-frame translation initiation codons, resulting in distinct N-termini.99 These isoforms may have differential localization and function within the cell. Nuclear HBx primarily activates transcription of viral encoded proteins, whereas cytoplasmic HBx targets signaling cascades and mitochondria.99 In mitochondria, studies have linked HBx to generation of reactive oxygen species (ROS) and a decrease in mitochondrial membrane potential.99 The generation of ROS is suggested to inhibit PTEN, a tumor suppressor, and is implicated in overall hepatocarcinogenesis.99
HBx has been associated with alterations in cellular iron homeostasis elicited by viral infection.107 When expressed in Huh7 cells, HBx led to a reduction in both protein and mRNA levels of transferrin receptor 1 (TFR1) while increasing the protein and mRNA levels of ferritin. 107 Both TFR1 and ferritin mRNA levels are under the control of the iron regulatory proteins (IRP1 and IRP2) that bind specific RNA secondary structures called iron-responsive elements (IRE).108-110 Notably, upon HBx expression in Huh7 cells, a significant decrease in the protein levels of IRP1 was observed, leading to the proposal that HBx may influence cellular iron homeostasis by affecting IRP1 levels.107
The biochemical and structural characterization of HBx has posed challenges, yet recent advancements have shed light on its properties. Notably, it has been discovered that HBx binds to a FeS cluster. 111 When solubility-tagged full-length HBx was purified, it was found to bind either a [2Fe-2S] or a [4Fe-4S] cluster, as confirmed by Mossbauer spectroscopy following growth in 57Fe-enriched media.111 The HBx-[2Fe-2S] was found to be stable upon exposure to atmospheric oxygen and could be converted to HBx-[4Fe-4S] upon addition of a strong reducing agent.111 This interconversion was found to be partially reversible, allowing HBx-[4Fe-4S] to revert to HBx-[2Fe-2S] when exposed to oxygen (Figure 5C), albeit with some cluster degradation resulting in the release of mononuclear Fe3+.111 The FeS cluster in HBx appears to persist even after multiple rounds of oxidation and reduction, despite the accumulation of iron released upon cluster degradation (Figure 5C).111 The release of mononuclear iron is likely acting as a source of ROS potentially outside mitochondria or even in the nucleus.111 The build-up of ROS resulting from iron release during HBx-cluster redox events may contribute to the severity of liver disease and the development of carcinogenesis.111
3.5. The Mimivirus GciS Protein
Mimiviruses are among the largest viruses found to date at roughly 750 nm in diameter with a linear dsDNA genome of about 12,000 kb.60 Their denotation originates from “Mimicking Microbes”, which was coined due to their ability to mimic bacteria in both size and appearance.60 Mimiviruses are Nucleocytoplasmic Large DNA viruses (NCLDV) which also includes Poxviridae and Asfarviridae, known to infect mammals.60, 112 Mimiviruses, however, infect Acanthamoeba, a unicellular amoeba.60 Their genomes consist of an impressive 1262 open reading frames that encode unusual proteins, including tRNA synthetases, translation peptide release factors, 6 tRNAs, DNA repair glycosylases, a UV-damage endonuclease, and type I and type II topoisomerases.60 It is likely not a coincidence that many of these pathways rely on FeS clusters within cells.
GciS is a mimivirus glycine and cysteine rich protein of about 6kDa that was recently found to ligate FeS clusters of both the [2Fe-2S]2+ and the linear [3Fe-4S]1+ geometry as assessed by Mossbauer spectroscopy on the Fe57-enriched purified protein (Figure 6A,B).113 The linear [3Fe-4S]1+ cluster represents a rare arrangement in biological systems.114 It may be an aerobically stabilized geometry in which loss of one Fe3+ atom would yield a [2Fe-2S] cluster or it may be a step in the process of reductive coupling toward a [4Fe-4S].114
While GciS lacks a predicted structure, it has demonstrated the ability to oligomerize.113 GciS ligates 0.23 [2Fe-2S]2+ clusters and 0.06 [3Fe-4S]1+ clusters per monomer as estimated from Inductively Coupled Plasma Mass Spectrometry (ICP-MS) results showing 0.65 Fe atoms per monomer. This suggests that cysteines from multiple monomers are required to bind even a single [2Fe-2S] cluster.113 In the case of glutaredoxins which have been shown to have similar FeS features, it's common for the FeS cluster to be bound by two cysteines from separate protein monomers along with two glutathione ligands.114 Villalta and colleagues give a glance into a potentially widespread FeS proteome in Mimiviruses.113 Further, they show that GciS-like proteins from clades A, B, C, and D of the Megavirinae family have UV-vis and EPR features suggestive of the presence of FeS clusters.113
4. Conclusion and Perspectives
4.1. Drawing Similarities
Table 1 outlines the five viruses thus far identified that encode FeS proteins, all of which are discussed in this review. A prevailing theme emerges, indicating that many of these viral FeS proteins are involved in genome replication. Viruses have very diverse and complex genomic organizations, collectively encoding an assorted array of polymerases to replicate their genomes.115 Viruses can be categorized according to their genome organization, as in the Baltimore Classification:
Group I: Double-stranded DNA viruses, (MCPyV, Mimivirus)
Group II: Single-stranded DNA viruses,
Group III: Double-stranded RNA viruses, (Rotavirus)
Group IV: Positive-sense single-stranded RNA viruses, (SARS-CoV-2)
Group V: Negative-sense single-stranded RNA viruses,
Group VI: Single-stranded RNA viruses with a DNA intermediate,
Group VII: Double-stranded DNA viruses with an RNA intermediate, (HBV)
Viruses must either encode genome replication proteins or manipulate cellular proteins not originally within their codebook to replicate their genomes. Incidentally, viruses encode some of the rarest polymerases and enzymes involved in nucleic acid processing. Further, within the same virus family, replication enzymes are the most well-conserved and viral polymerases also serve as a means to categorize viruses in terms of evolutionary characteristics.116, 117 The presence of FeS clusters within viral polymerases or other highly conserved proteins offers valuable insights to understand viral evolution and the changes they undergo over time.116, 117 Questions arise about the depth of conservation of viral FeS proteins and when they first emerged. Notably, all the virally encoded FeS proteins discussed in this review originate from viruses infecting eukaryotes. GciS stands out as the sole virally encoded FeS protein found in a virus infecting a non-mammalian organism. While little has been reported on FeS cluster utilization by viruses infecting other branches of the tree of life, it is likely that they exist.
Merkel Cell Polyomavirus and Hepatitis B viruses belong to distinct evolutionary families as evidenced by differences in their genome sequence homology: Polyomavirus falls under Polyomaviridae, while Hepatitis B virus is classified under Hepadnaviridae. However, they are related by the fact that they are non-retroviruses that are known to integrate into host genomes. HBV encodes a reverse transcriptase and integrates similarly to retroviruses.118 Merkel cell polyomaviruses LT may be responsible for clonal integration into host genomes during cellular mitosis.71 Additionally, virally encoded FeS proteins found in both viruses might contribute significantly to carcinogenesis. Both HBx and GciS demonstrate characteristics of iron-sulfur cluster scaffolds with the capability to enable cluster interconversion. Overall, these observations shed light on how viral proteins enable viruses to manipulate FeS cluster utilization by co-opting the cellular machinery responsible for their assembly.
4.2. So Why Do Viruses Utilize FeS Clusters?
FeS clusters offer benefits to viruses similar to those they provide to host cells, which possess intricate pathways for their assembly, for proper delivery to recipient proteins, and for repair of partially oxidized clusters. Given that virtually all cells depend on iron-sulfur clusters, it stands to reason that viruses infecting host cells would, either indirectly or as discussed in this review, directly rely on these clusters. However, it's worth noting that it is unclear whether all viruses encode FeS proteins. The question then arises: why have certain viruses evolved to directly utilize these cofactors while others might have not? Could the utilization of iron-sulfur clusters by viral proteins offer insights into the timing of their evolution and consequently, the evolution of the viruses that encode them?
Iron-sulfur clusters have been postulated to represent relics of early life on Earth, preceding a significant increase in atmospheric oxygen brought about by the emergence of photosynthetic organisms that took place around 2.1 to 2.4 billion years ago.119 Following the “Great Oxygen Event,” there was likely a widespread extinction event followed by evolutionary adaptations aimed towards utilizing oxidized Fe3+ while devising mechanisms to prevent the reactivity of free iron in cells. This reactivity could lead to the generation of harmful reactive oxygen species via Fenton chemistry.2, 13 This occurrence creates a paradox regarding the perceived benefits of FeS clusters in DNA replication proteins, as their presence could potentially heighten the risk of DNA damage due to reactive oxygen species. Despite this, numerous DNA and RNA binding proteins in bacteria, archaea, eukaryotes, and now viruses rely on FeS clusters, indicating an indispensable function of this cofactors for genome replication and maintenance.3-6 Viruses are important contributors to ecological gene pools, not only by integrating directly into host genomes and sometimes acquiring host genes, but also by influencing the natural selection of host genes that offer an immune advantage against viruses. Consequently, viruses adapt to counteract these evolving immune strategies. It is plausible that viral proteins utilizing FeS clusters are relics of ancient polymerases, ssRNA binding proteins, and helicases that employed FeS clusters in an era predating the "great oxygen event." These proteins have persisted throughout evolution due to the advantages they confer through natural selection.
4.3. A Note on Importance
Studying viral FeS proteins can offer valuable insights into mechanisms that are responsible for the fitness and adaptability of viruses. Additionally, exploring viral FeS proteins may broaden our knowledge about FeS clusters, their coordination geometries, redox chemistries, and more. For instance, biophysical studies on the mimivirus GciS protein have unveiled the presence in biological systems of the rare linear [3Fe-4S] geometries seldomly observed in vivo.
The identification of two FeS clusters in the SARS-CoV-2 RdRp highlighted the importance of FeS proteins in viral replication. This discovery is especially noteworthy because the RdRp, being a highly conserved protein in rapidly mutating coronaviruses, represents an ideal target for antiviral therapeutics, much needed during the global pandemic.120 The use of a FeS cluster-targeting technology was pioneered as an antiviral therapeutic strategy against SARS-CoV-2 infections,74, 120 marking a groundbreaking development likely to be followed by further advancements.
Presently, we stand at the cusp of an exponential discovery phase of novel FeS proteins. Many of the viral FeS proteins discussed in this review were stumbled upon accidentally. Now, armed with knowledge, we have a clearer understanding of where and how to search for candidate FeS proteins aided by the identification of evolutionarily conserved LYR-like motifs and potential metal binding sites in the structures of the proteins, when available, that can provide FeS cluster ligation sites.
Acknowledgments
The authors wish to thank the Intramural program of the Eunice Kennedy Shriver National Institute of Child Health and Human Development for support.
References
- Beinert, H.; Holm, R. H.; Munck, E. Iron-Sulfur Clusters: Nature’s Modular, Multipurpose Structures. Science 1997, 277(5326), 653–659. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.; Sabio, L.; Galvez, N.; Capdevila, M.; Dominguez-Vera, J. M. Iron chemistry at the service of life. IUBMB Life 2017, 69(6), 382–388. [Google Scholar] [CrossRef] [PubMed]
- Bartels, P. L.; Stodola, J. L.; Burgers, P. M. J.; Barton, J. K. A Redox Role for the [4Fe4S] Cluster of Yeast DNA Polymerase delta. J Am Chem Soc 2017, 139(50), 18339–18348. [Google Scholar] [CrossRef] [PubMed]
- Barton, J. K.; Silva, R. M. B.; O'Brien, E. Redox Chemistry in the Genome: Emergence of the [4Fe4S] Cofactor in Repair and Replication. Annu Rev Biochem 2019, 88, 163–190. [Google Scholar] [CrossRef] [PubMed]
- O'Brien, E.; Holt, M. E.; Thompson, M. K.; Salay, L. E.; Ehlinger, A. C.; Chazin, W. J.; Barton, J. K. The [4Fe4S] cluster of human DNA primase functions as a redox switch using DNA charge transport. Science 2017, 355 (6327). [CrossRef]
- Pinto, M. N.; Ter Beek, J.; Ekanger, L. A.; Johansson, E.; Barton, J. K. The [4Fe4S] Cluster of Yeast DNA Polymerase epsilon Is Redox Active and Can Undergo DNA-Mediated Signaling. J Am Chem Soc 2021, 143(39), 16147–16153. [Google Scholar] [CrossRef] [PubMed]
- Letts, J. A.; Sazanov, L. A. Clarifying the supercomplex: the higher-order organization of the mitochondrial electron transport chain. Nat Struct Mol Biol 2017, 24(10), 800–808. [Google Scholar] [CrossRef] [PubMed]
- Sickerman, N. S.; Ribbe, M. W.; Hu, Y. Nitrogenase Cofactor Assembly: An Elemental Inventory. Acc Chem Res 2017, 50(11), 2834–2841. [Google Scholar] [CrossRef] [PubMed]
- Sosorev, A. Y. Walking around Ribosomal Small Subunit: A Possible "Tourist Map" for Electron Holes. Molecules 2021, 26 (18).
- Lill, R.; Freibert, S. A. Mechanisms of Mitochondrial Iron-Sulfur Protein Biogenesis. Annu Rev Biochem 2020, 89, 471–499. [Google Scholar] [CrossRef]
- Holm, R. H.; Lo, W. Structural Conversions of Synthetic and Protein-Bound Iron-Sulfur Clusters. Chem Rev 2016, 116(22), 13685–13713. [Google Scholar] [CrossRef]
- Bromberg, Y.; Aptekmann, A. A.; Mahlich, Y.; Cook, L.; Senn, S.; Miller, M.; Nanda, V.; Ferreiro, D.; Falkowski, P. Quantifying structural relationships of metal-binding sites suggests origins of biological electron transfer. Science Advances 2022, 8(3984), 1–13. [Google Scholar] [CrossRef]
- Imlay, J. A. Iron-sulphur clusters and the problem with oxygen. Mol Microbiol 2006, 59(4), 1073–82. [Google Scholar] [CrossRef]
- McKeown, S. R. Defining normoxia, physoxia and hypoxia in tumours-implications for treatment response. Br J Radiol 2014, 87(1035), 20130676. [Google Scholar] [CrossRef]
- Thomas, K. A. Angiogenesis. In Encyclopedia of Cell Biology, 2016; pp 102-116.
- Maio, N.; Rouault, T. A. Outlining the Complex Pathway of Mammalian Fe-S Cluster Biogenesis. Trends Biochem Sci 2020, 45(5), 411–426. [Google Scholar] [CrossRef] [PubMed]
- Genova, M. L.; Lenaz, G. Functional role of mitochondrial respiratory supercomplexes. Biochim Biophys Acta 2014, 1837(4), 427–43. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B. L.; Zahn, K. E.; Hanley, J. P.; Wallace, S. S.; Dragon, J. A.; Doublie, S. Caught in motion: human NTHL1 undergoes interdomain rearrangement necessary for catalysis. Nucleic Acids Res 2021. [Google Scholar] [CrossRef] [PubMed]
- Maio, N.; Rouault, T. A. Mammalian iron sulfur cluster biogenesis: From assembly to delivery to recipient proteins with a focus on novel targets of the chaperone and co-chaperone proteins. IUBMB Life 2022, 74(7), 684–704. [Google Scholar] [CrossRef] [PubMed]
- Ciesielski, S. J.; Schilke, B. A.; Osipiuk, J.; Bigelow, L.; Mulligan, R.; Majewska, J.; Joachimiak, A.; Marszalek, J.; Craig, E. A.; Dutkiewicz, R. Interaction of J-protein co-chaperone Jac1 with Fe-S scaffold Isu is indispensable in vivo and conserved in evolution. J Mol Biol 2012, 417 (1-2), 1-12. [CrossRef]
- Chakrabbarti, M.; Lindahl, P. A. The utility of Mossbauer spectroscopy in eukaryotic cell biology and animal physiology. In Iron-sulfur clusters in chemistry and Biology, Rouault, T. A., Ed. Walter de Gruyter: Berlin/Boston, 2014; pp 49-71. [CrossRef]
- Maio, N.; Rouault, T. A. Iron-sulfur cluster biogenesis in mammalian cells: New insights into the molecular mechanisms of cluster delivery. Biochim Biophys Acta 2015, 1853(6), 1493–512. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tong, W.-H.; Hughes, R. M.; Rouault, T. A. Roles of the Mammalian Cytosolic Cysteine Desulfurase, ISCS, and Scaffold Protein, ISCU, in Iron-Sulfur Cluster Assembly. Journal of Biological Chemistry 2006, 281(18), 12344–12351. [Google Scholar] [CrossRef] [PubMed]
- Tong, W. H.; Rouault, T. A. Distinct iron-sulfur cluster assembly complexes exist in the cytosol and mitochondria of human cells. EMBO 2000, 19(21), 5692–5700. [Google Scholar] [CrossRef]
- Tong, W. H.; Rouault, T. A. Functions of mitochondrial ISCU and cytosolic ISCU in mammalian iron-sulfur cluster biogenesis and iron homeostasis. Cell Metab 2006, 3(3), 199–210. [Google Scholar] [CrossRef]
- Shi, R.; Proteau, A.; Villarroya, M.; Moukadiri, I.; Zhang, L.; Trempe, J. F.; Matte, A.; Armengod, M. E.; Cygler, M. Structural basis for Fe-S cluster assembly and tRNA thiolation mediated by IscS protein-protein interactions. PLoS Biol 2010, 8(4), e1000354. [Google Scholar] [CrossRef]
- Adam, A. C.; Bornhovd, C.; Prokisch, H.; Neupert, W.; Hell, K. The Nfs1 interacting protein Isd11 has an essential role in Fe/S cluster biogenesis in mitochondria. EMBO J 2006, 25(1), 174–83. [Google Scholar] [CrossRef]
- Shi, Y.; Ghosh, M. C.; Tong, W. H.; Rouault, T. A. Human ISD11 is essential for both iron-sulfur cluster assembly and maintenance of normal cellular iron homeostasis. Hum Mol Genet 2009, 18(16), 3014–25. [Google Scholar] [CrossRef]
- Terali, K.; Beavil, R. L.; Pickersgill, R. W.; van der Giezen, M. The effect of the adaptor protein Isd11 on the quaternary structure of the eukaryotic cysteine desulphurase Nfs1. Biochem Biophys Res Commun 2013, 440(2), 235–40. [Google Scholar] [CrossRef]
- Patel, S. J.; Frey, A. G.; Palenchar, D. J.; Achar, S.; Bullough, K. Z.; Vashisht, A.; Wohlschlegel, J. A.; Philpott, C. C. A PCBP1-BolA2 chaperone complex delivers iron for cytosolic [2Fe-2S] cluster assembly. Nat Chem Biol 2019, 15(9), 872–881. [Google Scholar] [CrossRef]
- Shi, H.; Bencze, K. Z.; Stemmler, T. L.; Philpott, C. C. A cytosolic iron chaperone that delivers iron to ferritin. Science 2008, 320(5880), 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Nandal, A.; Ruiz, J. C.; Subramanian, P.; Ghimire-Rijal, S.; Sinnamon, R. A.; Stemmler, T. L.; Bruick, R. K.; Philpott, C. C. Activation of the HIF prolyl hydroxylase by the iron chaperones PCBP1 and PCBP2. Cell Metab 2011, 14(5), 647–57. [Google Scholar] [CrossRef] [PubMed]
- Leidgens, S.; Bullough, K. Z.; Shi, H.; Li, F.; Shakoury-Elizeh, M.; Yabe, T.; Subramanian, P.; Hsu, E.; Natarajan, N.; Nandal, A.; Stemmler, T. L.; Philpott, C. C. Each member of the poly-r(C)-binding protein 1 (PCBP) family exhibits iron chaperone activity toward ferritin. J Biol Chem 2013, 288(24), 17791–802. [Google Scholar] [CrossRef] [PubMed]
- Frey, A. G.; Nandal, A.; Park, J. H.; Smith, P. M.; Yabe, T.; Ryu, M. S.; Ghosh, M. C.; Lee, J.; Rouault, T. A.; Park, M. H.; Philpott, C. C. Iron chaperones PCBP1 and PCBP2 mediate the metallation of the dinuclear iron enzyme deoxyhypusine hydroxylase. Proc Natl Acad Sci U S A 2014, 111(22), 8031–6. [Google Scholar] [CrossRef] [PubMed]
- Boniecki, M. T.; Freibert, S. A.; Muhlenhoff, U.; Lill, R.; Cygler, M. Structure and functional dynamics of the mitochondrial Fe/S cluster synthesis complex. Nat Commun 2017, 8(1), 1287. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Tonelli, M.; Frederick, R. O.; Markley, J. L. Human Mitochondrial Ferredoxin 1 (FDX1) and Ferredoxin 2 (FDX2) Both Bind Cysteine Desulfurase and Donate Electrons for Iron-Sulfur Cluster Biosynthesis. Biochemistry 2017, 56(3), 487–499. [Google Scholar] [CrossRef]
- Chandramouli, K.; Unciuleac, M. C.; Naik, S.; Dean, D. R.; Huynh, B. H.; Johnson, M. K. Formation and properties of [4Fe-4S] clusters on the IscU scaffold protein. Biochemistry 2007, 46(23), 6804–11. [Google Scholar] [CrossRef]
- Tse, E. C. M.; Zwang, T. J.; Barton, J. K. The Oxidation State of [4Fe4S] Clusters Modulates the DNA-Binding Affinity of DNA Repair Proteins. J Am Chem Soc 2017, 139(36), 12784–12792. [Google Scholar] [CrossRef] [PubMed]
- Pandelia, M. E.; Lanz, N. D.; Booker, S. J.; Krebs, C. Mossbauer spectroscopy of Fe/S proteins. Biochim Biophys Acta 2015, 1853(6), 1395–405. [Google Scholar] [CrossRef] [PubMed]
- Voisine, C.; Cheng, Y. C.; Ohlson, M.; Schilke, B.; Hoff, K.; Beinert, H.; Marszalek, J.; Craig, E. A. Jac1, a mitochondrial J-type chaperone, is involved in the biogenesis of Fe/S clusters in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 2001, 98(4), 1483–8. [Google Scholar] [CrossRef] [PubMed]
- Cupp-Vickery, J. R.; Peterson, J. C.; Ta, D. T.; Vickery, L. E. Crystal structure of the molecular chaperone HscA substrate binding domain complexed with the IscU recognition peptide ELPPVKIHC. J Mol Biol 2004, 342(4), 1265–78. [Google Scholar] [CrossRef] [PubMed]
- Maio, N.; Singh, A.; Uhrigshardt, H.; Saxena, N.; Tong, W. H.; Rouault, T. A. Cochaperone binding to LYR motifs confers specificity of iron sulfur cluster delivery. Cell Metab 2014, 19(3), 445–57. [Google Scholar] [CrossRef] [PubMed]
- Marszalek, J.; Craig, E. A. Interaction of client-the scaffold on which FeS clusters are build-with J-domain protein Hsc20 and its evolving Hsp70 partners. Front Mol Biosci 2022, 9, 1034453. [Google Scholar] [CrossRef] [PubMed]
- Maio, N.; Kim, K. S.; Singh, A.; Rouault, T. A. A Single Adaptable Cochaperone-Scaffold Complex Delivers Nascent Iron-Sulfur Clusters to Mammalian Respiratory Chain Complexes I-III. Cell Metab 2017, 25 (4), 945-953 e6. [CrossRef]
- Kampinga, H. H.; Craig, E. A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat Rev Mol Cell Biol 2010, 11(8), 579–92. [Google Scholar] [CrossRef] [PubMed]
- Sahi, C.; Craig, E. A. Network of general and specialty J protein chaperones of the yeast cytosol. Proc Natl Acad Sci U S A 2007, 104(17), 7163–8. [Google Scholar] [CrossRef]
- Bonomi, F.; Iametti, S.; Morleo, A.; Ta, D.; Vickery, L. E. Facilitated transfer of IscU-[2Fe2S] clusters by chaperone-mediated ligand exchange. Biochemistry 2011, 50(44), 9641–50. [Google Scholar] [CrossRef]
- Dutkiewicz, R.; Schilke, B.; Cheng, S.; Knieszner, H.; Craig, E. A.; Marszalek, J. Sequence-specific interaction between mitochondrial Fe-S scaffold protein Isu and Hsp70 Ssq1 is essential for their in vivo function. J Biol Chem 2004, 279(28), 29167–74. [Google Scholar] [CrossRef]
- Hoff, K. G.; Cupp-Vickery, J. R.; Vickery, L. E. Contributions of the LPPVK motif of the iron-sulfur template protein IscU to interactions with the Hsc66-Hsc20 chaperone system. J Biol Chem 2003, 278(39), 37582–9. [Google Scholar] [CrossRef]
- Rouault, T. A. Mammalian iron-sulphur proteins: novel insights into biogenesis and function. Nat Rev Mol Cell Biol 2015, 16(1), 45–55. [Google Scholar] [CrossRef]
- Gari, K.; Ortiz, A. M. L.; Borel, V.; Flynn, H.; Skehel, J. M.; Boulton, S. J. MMS19 links cytoplasmic iron-sulfur cluster assembly to DNA metabolism. Science 2012, 337. [Google Scholar] [CrossRef] [PubMed]
- Stehling, O.; Vashisht, A. A.; Mascarenhas, J.; Jonsson, Z. O.; Sharma, T.; Netz, D. J.; Pierik, A. J.; Wohlschlegel, J. A.; Lill, R. MMS19 assembles iron-sulfur proteins required for DNA metabolism and genomic integrity. Science 2012, 337(6091), 195–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, K. S.; Maio, N.; Singh, A.; Rouault, T. A. Cytosolic HSC20 integrates de novo iron-sulfur cluster biogenesis with the CIAO1-mediated transfer to recipients. Hum Mol Genet 2018, 27(5), 837–852. [Google Scholar] [CrossRef] [PubMed]
- Kassube, S. A.; Thoma, N. H. Structural insights into Fe-S protein biogenesis by the CIA targeting complex. Nat Struct Mol Biol 2020, 27(8), 735–742. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Rouault, T. A. Human iron-sulfur cluster assembly, cellular iron homeostasis, and disease. Biochemistry 2010, 49(24), 4945–56. [Google Scholar] [CrossRef] [PubMed]
- Crick, F. H. C.; Watson, J. D. Structure of Small Viruses. Nature 1956, 177, 473–475. [Google Scholar] [CrossRef]
- Wolf, Y. I.; Kazlauskas, D.; Iranzo, J.; Lucia-Sanz, A.; Kuhn, J. H.; Krupovic, M.; Dolja, V. V.; Koonin, E. V. Origins and Evolution of the Global RNA Virome. mBio 2018, 9 (6). [CrossRef]
- Twarock, R.; Luque, A. Structural puzzles in virology solved with an overarching icosahedral design principle. Nat Commun 2019, 10(1), 4414. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; De Lamballerie, X.; Yutin, N.; Asgari, S.; Bigot, Y.; Bideshi, D. K.; Cheng, X. W.; Federici, B. A.; Van Etten, J. L.; Koonin, E. V.; La Scola, B.; Raoult, D. "Megavirales", a proposed new order for eukaryotic nucleocytoplasmic large DNA viruses. Arch Virol 2013, 158(12), 2517–21. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Audic, S.; Robert, C.; Abergel, C.; Renesto, P.; Ogata, H.; Scola, B.; Suzan, M.; Claverie, J. The 1.2-Mb Genome Sequence of Mimivirus. Science 2004, 306, 1344 - 1350. [CrossRef]
- Martin, D.; Charpilienne, A.; Parent, A.; Boussac, A.; D'Autreaux, B.; Poupon, J.; Poncet, D. The rotavirus nonstructural protein NSP5 coordinates a [2Fe-2S] iron-sulfur cluster that modulates interaction to RNA. FASEB J 2013, 27(3), 1074–83. [Google Scholar] [CrossRef] [PubMed]
- Ozsari, T.; Bora, G.; Kaya, B.; Yakut, K. The Prevalence of Rotavirus and Adenovirus in the Childhood Gastroenteritis. Jundishapur J Microbiol 2016, 9(6), e34867. [Google Scholar] [CrossRef] [PubMed]
- Papa, G.; Borodavka, A.; Desselberger, U. Viroplasms: Assembly and Functions of Rotavirus Replication Factories. Viruses 2021, 13 (7). [CrossRef]
- Martin, D.; Ouldali, M.; Menetrey, J.; Poncet, D. Structural organisation of the rotavirus nonstructural protein NSP5. J Mol Biol 2011, 413(1), 209–21. [Google Scholar] [CrossRef]
- Fabbretti, E.; Afrikanova, I.; Vascotto, F.; Burrone, O. R. Two non-structural rotavirus proteins, NSP2 and NSP5, form viroplasm-like structures in vivo. Journal of General Virology 1999, 80, 333–339. [Google Scholar] [CrossRef]
- Papa, G.; Venditti, L.; Arnoldi, F.; Schraner, E. M.; Potgieter, C.; Borodavka, A.; Eichwald, C.; Burrone, O. R. Recombinant Rotaviruses Rescued by Reverse Genetics Reveal the Role of NSP5 Hyperphosphorylation in the Assembly of Viral Factories. J Virol 2019, 94 (1). [CrossRef]
- Tsang, S. H.; Wang, R.; Nakamaru-Ogiso, E.; Knight, S. A.; Buck, C. B.; You, J. The Oncogenic Small Tumor Antigen of Merkel Cell Polyomavirus Is an Iron-Sulfur Cluster Protein That Enhances Viral DNA Replication. J Virol 2016, 90(3), 1544–56. [Google Scholar] [CrossRef]
- Wendzicki, J. A.; Moore, P. S.; Chang, Y. Large T and small T antigens of Merkel cell polyomavirus. Curr Opin Virol 2015, 11, 38–43. [Google Scholar] [CrossRef]
- Kwun, H. J.; Guastafierro, A.; Shuda, M.; Meinke, G.; Bohm, A.; Moore, P. S.; Chang, Y. The minimum replication origin of merkel cell polyomavirus has a unique large T-antigen loading architecture and requires small T-antigen expression for optimal replication. J Virol 2009, 83(23), 12118–28. [Google Scholar] [CrossRef]
- Becker, J. C.; Stang, A.; DeCaprio, J. A.; Cerroni, L.; Lebbe, C.; Veness, M.; Nghiem, P. Merkel cell carcinoma. Nat Rev Dis Primers 2017, 3, 17077. [Google Scholar] [CrossRef]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P. S. Clonal Integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319. [Google Scholar] [CrossRef]
- Rapchak, K.; Yagobian, S. D.; Moore, J.; Khattri, M.; Shuda, M. Merkel cell polyomavirus small T antigen is a viral transcription activator that is essential for viral genome maintenance. PLoS Pathog 2022, 18(12), e1011039. [Google Scholar] [CrossRef]
- Kwun, H. J.; Shuda, M.; Feng, H.; Camacho, C. J.; Moore, P. S.; Chang, Y. Merkel cell polyomavirus small T antigen controls viral replication and oncoprotein expression by targeting the cellular ubiquitin ligase SCFFbw7. Cell Host Microbe 2013, 14(2), 125–35. [Google Scholar] [CrossRef]
- Maio, N.; Lafont, B. A. P.; Sil, D.; Li, Y.; Bollinger, J. M., Jr.; Krebs, C.; Pierson, T. C.; Linehan, W. M.; Rouault, T. A. Fe-S cofactors in the SARS-CoV-2 RNA-dependent RNA polymerase are potential antiviral targets. Science 2021, 373. [Google Scholar] [CrossRef]
- Maio, N.; Raza, M. K.; Li, Y.; Zhang, D.; Bollinger, J. M., Jr.; Krebs, C.; Rouault, T. A. An iron-sulfur cluster in zinc-binding domain of the SARS-CoV-2 helicase modulates its RNA binding and unwinding activities. PNAS 2023. [Google Scholar] [CrossRef] [PubMed]
- Malone, B.; Campbell, E. A.; Darst, S. A. CoV-er all the bases: Structural perspectives of SARS-CoV-2 RNA synthesis. Enzymes 2021, 49, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Woo, P. C.; Lau, S. K.; Huang, Y.; Yuen, K. Y. Coronavirus diversity, phylogeny and interspecies jumping. Exp Biol Med (Maywood) 2009, 234(10), 1117–27. [Google Scholar] [CrossRef]
- Robson, F.; Khan, K. S.; Le, T. K.; Paris, C.; Demirbag, S.; Barfuss, P.; Rocchi, P.; Ng, W. L. Coronavirus RNA Proofreading: Molecular Basis and Therapeutic Targeting. Mol Cell 2020, 79(5), 710–727. [Google Scholar] [CrossRef]
- Gribble, J.; Stevens, L. J.; Agostini, M. L.; Anderson-Daniels, J.; Chappell, J. D.; Lu, X.; Pruijssers, A. J.; Routh, A. L.; Denison, M. R. The coronavirus proofreading exoribonuclease mediates extensive viral recombination. PLoS Pathog 2021, 17(1), e1009226. [Google Scholar] [CrossRef] [PubMed]
- Brant, A. C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z. M. SARS-CoV-2: from its discovery to genome structure, transcription, and replication. Cell Biosci 2021, 11(1), 136. [Google Scholar] [CrossRef]
- Yan, L.; Yang, Y.; Li, M.; Zhang, Y.; Zheng, L.; Ge, J.; Huang, Y. C.; Liu, Z.; Wang, T.; Gao, S.; Zhang, R.; Huang, Y. Y.; Guddat, L. W.; Gao, Y.; Rao, Z.; Lou, Z. Coupling of N7-methyltransferase and 3'-5' exoribonuclease with SARS-CoV-2 polymerase reveals mechanisms for capping and proofreading. Cell 2021, 184 (13), 3474-3485 e11.
- Dos Ramos, F.; Carrasco, M.; Doyle, T.; Brierley, I. Programmed -1 ribosomal frameshifting in the SARS coronavirus. Biochem Soc Trans 2004, 32, 1081–1083. [Google Scholar] [CrossRef]
- Thiel, V.; Ivanov, K. A.; Putics, A.; Hertzig, T.; Schelle, B.; Bayer, S.; Weissbrich, B.; Snijder, E. J.; Rabenau, H.; Doerr, H. W.; Gorbalenya, A. E.; Ziebuhr, J. Mechanisms and enzymes involved in SARS coronavirus genome expression. J Gen Virol 2003, 84 (Pt 9), 2305–2315. [Google Scholar] [CrossRef]
- Hillen, H. S.; Kokic, G.; Farnung, L.; Dienemann, C.; Tegunov, D.; Cramer, P. Structure of replicating SARS-CoV-2 polymerase. Nature 2020, 584(7819), 154–156. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Lokugamage, K. G.; Makino, S. Viral and Cellular mRNA Translation in Coronavirus-Infected Cells. Adv Virus Res 2016, 96, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Kirchdoerfer, R. N.; Ward, A. B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat Commun 2019, 10(1), 2342. [Google Scholar] [CrossRef] [PubMed]
- Saikrishnan, K.; Powell, B.; Cook, N. J.; Webb, M. R.; Wigley, D. B. Mechanistic basis of 5'-3' translocation in SF1B helicases. Cell 2009, 137(5), 849–59. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Ge, J.; Zheng, L.; Zhang, Y.; Gao, Y.; Wang, T.; Huang, Y.; Yang, Y.; Gao, S.; Li, M.; Liu, Z.; Wang, H.; Li, Y.; Chen, Y.; Guddat, L. W.; Wang, Q.; Rao, Z.; Lou, Z. Cryo-EM Structure of an Extended SARS-CoV-2 Replication and Transcription Complex Reveals an Intermediate State in Cap Synthesis. Cell 2021, 184 (1), 184-193 e10.
- Chen, J.; Wang, Q.; Malone, B.; Llewellyn, E.; Pechersky, Y.; Maruthi, K.; Eng, E. T.; Perry, J. K.; Campbell, E. A.; Shaw, D. E.; Darst, S. A. Ensemble cryo-EM reveals conformational states of the nsp13 helicase in the SARS-CoV-2 helicase replication-transcription complex. Nat Struct Mol Biol 2022, 29(3), 250–260. [Google Scholar] [CrossRef] [PubMed]
- Singleton, M. R.; Dillingham, M. S.; Wigley, D. B. Structure and mechanism of helicases and nucleic acid translocases. Annu Rev Biochem 2007, 76, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Mickolajczyk, K. J.; Shelton, P. M. M.; Grasso, M.; Cao, X.; Warrington, S. E.; Aher, A.; Liu, S.; Kapoor, T. M. Force-dependent stimulation of RNA unwinding by SARS-CoV-2 nsp13 helicase. Biophys J 2021, 120(6), 1020–1030. [Google Scholar] [CrossRef]
- Jang, K. J.; Jeong, S.; Kang, D. Y.; Sp, N.; Yang, Y. M.; Kim, D. E. A high ATP concentration enhances the cooperative translocation of the SARS coronavirus helicase nsP13 in the unwinding of duplex RNA. Sci Rep 2020, 10(1), 4481. [Google Scholar] [CrossRef]
- Sommers, J. A.; Loftus, L. N.; Jones, M. P., 3rd; Lee, R. A.; Haren, C. E.; Dumm, A. J.; Brosh, R. M., Jr. Biochemical analysis of SARS-CoV-2 Nsp13 helicase implicated in COVID-19 and factors that regulate its catalytic functions. J Biol Chem 2023, 299(3), 102980. [Google Scholar] [CrossRef] [PubMed]
- Shimberg, G. D.; Michalek, J. L.; Oluyadi, A. A.; Rodrigues, A. V.; Zucconi, B. E.; Neu, H. M.; Ghosh, S.; Sureschandra, K.; Wilson, G. M.; Stemmler, T. L.; Michel, S. L. Cleavage and polyadenylation specificity factor 30: An RNA-binding zinc-finger protein with an unexpected 2Fe-2S cluster. Proc Natl Acad Sci U S A 2016, 113(17), 4700–5. [Google Scholar] [CrossRef]
- Weiss, A.; Murdoch, C. C.; Edmonds, K. A.; Jordan, M. R.; Monteith, A. J.; Perera, Y. R.; Rodriguez Nassif, A. M.; Petoletti, A. M.; Beavers, W. N.; Munneke, M. J.; Drury, S. L.; Krystofiak, E. S.; Thalluri, K.; Wu, H.; Kruse, A. R. S.; DiMarchi, R. D.; Caprioli, R. M.; Spraggins, J. M.; Chazin, W. J.; Giedroc, D. P.; Skaar, E. P. Zn-regulated GTPase metalloprotein activator 1 modulates vertebrate zinc homeostasis. Cell 2022, 185 (12), 2148-2163 e27. [CrossRef]
- Petrenko, A.; Stein, M. Distal [FeS]-Cluster Coordination in [NiFe]-Hydrogenase Facilitates Intermolecular Electron Transfer. Int J Mol Sci 2017, 18 (1). [CrossRef]
- Bak, D. W.; Elliott, S. J. Alternative FeS cluster ligands: tuning redox potentials and chemistry. Curr Opin Chem Biol 2014, 19, 50–8. [Google Scholar] [CrossRef]
- Burns, G. S.; Thompson, A. J. Viral hepatitis B: clinical and epidemiological characteristics. Cold Spring Harb Perspect Med 2014, 4(12), a024935. [Google Scholar] [CrossRef] [PubMed]
- Schollmeier, A.; Glitscher, M.; Hildt, E. Relevance of HBx for Hepatitis B Virus-Associated Pathogenesis. Int J Mol Sci 2023, 24 (5). [CrossRef]
- Brechot, C. Hepatitis B virus (HBV) and hepatocellular carcinoma. Journal of Hepatology 1987, 4, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Wu, H. Y.; Guy, J. S.; Yoo, D.; Vlasak, R.; Urbach, E.; Brian, D. A. Common RNA replication signals exist among group 2 coronaviruses: evidence for in vivo recombination between animal and human coronavirus molecules. Virology 2003, 315(1), 174–83. [Google Scholar] [CrossRef]
- Eckhardt, S. G.; Milich, D. R.; McLachlan, A. Hepatitis B virus core antigen has two nuclear localization sequences in the Arginine-rich carboxyl terminus. J Virol 1991, 65, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Tsukuda, S.; Watashi, K. Hepatitis B virus biology and life cycle. Antiviral Res 2020, 182, 104925. [Google Scholar] [CrossRef]
- Wei, L.; Ploss, A. Hepatitis B virus cccDNA is formed through distinct repair processes of each strand. Nat Commun 2021, 12(1), 1591. [Google Scholar] [CrossRef]
- Dane, D. S.; H. C. C., Virus-like particles in serum of patients with Australia-antigen associated hepatitis. The Lancet 1970, 295(7649), 649–698. [Google Scholar] [CrossRef]
- van Hemert, F. J.; van de Klundert, M. A.; Lukashov, V. V.; Kootstra, N. A.; Berkhout, B.; Zaaijer, H. L. Protein X of hepatitis B virus: origin and structure similarity with the central domain of DNA glycosylase. PLoS One 2011, 6(8), e23392. [Google Scholar] [CrossRef] [PubMed]
- Gu, J. M.; Lim, S. O.; Oh, S. J.; Yoon, S. M.; Seong, J. K.; Jung, G. HBx modulates iron regulatory protein 1-mediated iron metabolism via reactive oxygen species. Virus Res 2008, 133(2), 167–77. [Google Scholar] [CrossRef] [PubMed]
- Bettany, A. J.; Eisenstein, R. S.; Munro, H. N. Mutagenesis of the iron-regulatory element further defines a role for RNA secondary structure in the regulation of ferritin and transferrin receptor expression. Journal of Biological Chemistry 1992, 267(23), 16531–16537. [Google Scholar] [CrossRef]
- Aziz, N.; Munro, H. N. Iron regulates ferritin mRNA translation through a segment of its 5’ untranslated region. PNAS 1987, 84, 8478–8482. [Google Scholar] [CrossRef]
- Rouault, T. A. The role of iron regulatory proteins in mammalian iron homeostasis and disease. Nat Chem Biol 2006, 2(8), 406–14. [Google Scholar] [CrossRef]
- Ueda, C.; Langton, M.; Chen, J.; Pandelia, M. E. The HBx protein from Hepatitis B Virus coordinates a redox-active Fe-S cluster. J Biol Chem 2022, 101698. [Google Scholar] [CrossRef] [PubMed]
- Monttinen, H. A. M.; Bicep, C.; Williams, T. A.; Hirt, R. P. The genomes of nucleocytoplasmic large DNA viruses: viral evolution writ large. Microb Genom 2021, 7 (9). [CrossRef]
- Villalta, A.; Srour, B.; Lartigue, A.; Clemancey, M.; Byrne, D.; Chaspoul, F.; Loquet, A.; Guigliarelli, B.; Blondin, G.; Abergel, C.; Burlat, B. Evidence for [2Fe-2S](2+) and Linear [3Fe-4S](1+) Clusters in a Unique Family of Glycine/Cysteine-Rich Fe-S Proteins from Megavirinae Giant Viruses. J Am Chem Soc 2023, 145(5), 2733–2738. [Google Scholar] [CrossRef]
- Zhang, B.; Bandyopadhyay, S.; Shakamuri, P.; Naik, S. G.; Huynh, B. H.; Couturier, J.; Rouhier, N.; Johnson, M. K. Monothiol glutaredoxins can bind linear [Fe3S4]+ and [Fe4S4]2+ clusters in addition to [Fe2S2]2+ clusters: spectroscopic characterization and functional implications. J Am Chem Soc 2013, 135(40), 15153–64. [Google Scholar] [CrossRef]
- Koonin, E. V.; Krupovic, M.; Agol, V. I. The Baltimore Classification of Viruses 50 Years Later: How Does It Stand in the Light of Virus Evolution? Microbiol Mol Biol Rev 2021, 85(3), e0005321. [Google Scholar] [CrossRef]
- Peersen, O. B. A Comprehensive Superposition of Viral Polymerase Structures. Viruses 2019, 11 (8). [CrossRef]
- de Farias, S. T.; Dos Santos Junior, A. P.; Rego, T. G.; Jose, M. V. Origin and Evolution of RNA-Dependent RNA Polymerase. Front Genet 2017, 8, 125. [Google Scholar] [CrossRef]
- Menendez-Arias, L.; Sebastian-Martin, A.; Alvarez, M. Viral reverse transcriptases. Virus Res 2017, 234, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Sessions, A. L.; Doughty, D. M.; Welander, P. V.; Summons, R. E.; Newman, D. K. The continuing puzzle of the great oxidation event. Curr Biol 2009, 19(14), R567–74. [Google Scholar] [CrossRef] [PubMed]
- Maio, N.; Cherry, S.; Schultz, D. C.; Hurst, B. L.; Linehan, W. M.; Rouault, T. A. TEMPOL inhibits SARS-CoV-2 replication and development of lung disease in the Syrian hamster model. iScience 2022, 25(10), 105074. [Google Scholar] [CrossRef] [PubMed]
- Long, S. SARS-CoV-2 Subgenomic RNAs: Characterization, Utility, and Perspectives. Viruses 2021, 13 (10). [CrossRef]
Figure 1.
Iron-sulfur clusters and their assembly. A. Biologically relevant FeS cluster geometries and possible interconversions.11 B. Iron sulfur cluster biogenesis is a complex highly regulated process that requires two basic steps: (I) de novo FeS cluster formation on the main scaffold ISCU16, 19, 21, 22 by the activity of a cysteine desulfurase, NFS1, which converts cysteine to alanine and generates a persulfide intermediate on ISCU in the process.16, 19, 21, 22 Donation of iron and reducing equivalents completes [2Fe-2S] cluster assembly on the main scaffold ISCU.30 (II) The newly assembled FeS cluster is delivered to the appropriate recipient proteins by the activity of an HSP70 chaperone and cognate J-domain HSP40 cochaperone system.22, 42, 44 In mammalian cells, the chaperone is HSPA9 and the cochaperone HSC20 (aka HSCB). HSC20 recognizes LYR-like motifs present in subsets of FeS recipient proteins.22, 42, 44 In the cytosol of mammalian cells, following cluster incorporation, a subset of proteins, such as NARFL and CIAPIN1, complete their maturation and become active.22, 42, 44 (III) Nucleic acid processing enzymes require a highly specialized cytoplasmic iron-sulfur assembly (CIA) complex to acquire their cofactors.22, 53 The CIA complex consists of CIAO1, MMS19, and FAM96B. CIAO1 was found to harbor a highly conserved LYR-motif that engages the cochaperone HSC20, thereby acting as a bridge between CIA and the de novo FeS assembly machinery.22, 53 A subset of FeS proteins that acquire their cofactors from the CIA complex, (e.g. DPYD and ABCE1) may function in the cytosol, while others are translocated to the nucleus (POLD1, PRIM2, FANCJ).4 Figure created with the help of BioRender.com.
Figure 1.
Iron-sulfur clusters and their assembly. A. Biologically relevant FeS cluster geometries and possible interconversions.11 B. Iron sulfur cluster biogenesis is a complex highly regulated process that requires two basic steps: (I) de novo FeS cluster formation on the main scaffold ISCU16, 19, 21, 22 by the activity of a cysteine desulfurase, NFS1, which converts cysteine to alanine and generates a persulfide intermediate on ISCU in the process.16, 19, 21, 22 Donation of iron and reducing equivalents completes [2Fe-2S] cluster assembly on the main scaffold ISCU.30 (II) The newly assembled FeS cluster is delivered to the appropriate recipient proteins by the activity of an HSP70 chaperone and cognate J-domain HSP40 cochaperone system.22, 42, 44 In mammalian cells, the chaperone is HSPA9 and the cochaperone HSC20 (aka HSCB). HSC20 recognizes LYR-like motifs present in subsets of FeS recipient proteins.22, 42, 44 In the cytosol of mammalian cells, following cluster incorporation, a subset of proteins, such as NARFL and CIAPIN1, complete their maturation and become active.22, 42, 44 (III) Nucleic acid processing enzymes require a highly specialized cytoplasmic iron-sulfur assembly (CIA) complex to acquire their cofactors.22, 53 The CIA complex consists of CIAO1, MMS19, and FAM96B. CIAO1 was found to harbor a highly conserved LYR-motif that engages the cochaperone HSC20, thereby acting as a bridge between CIA and the de novo FeS assembly machinery.22, 53 A subset of FeS proteins that acquire their cofactors from the CIA complex, (e.g. DPYD and ABCE1) may function in the cytosol, while others are translocated to the nucleus (POLD1, PRIM2, FANCJ).4 Figure created with the help of BioRender.com.
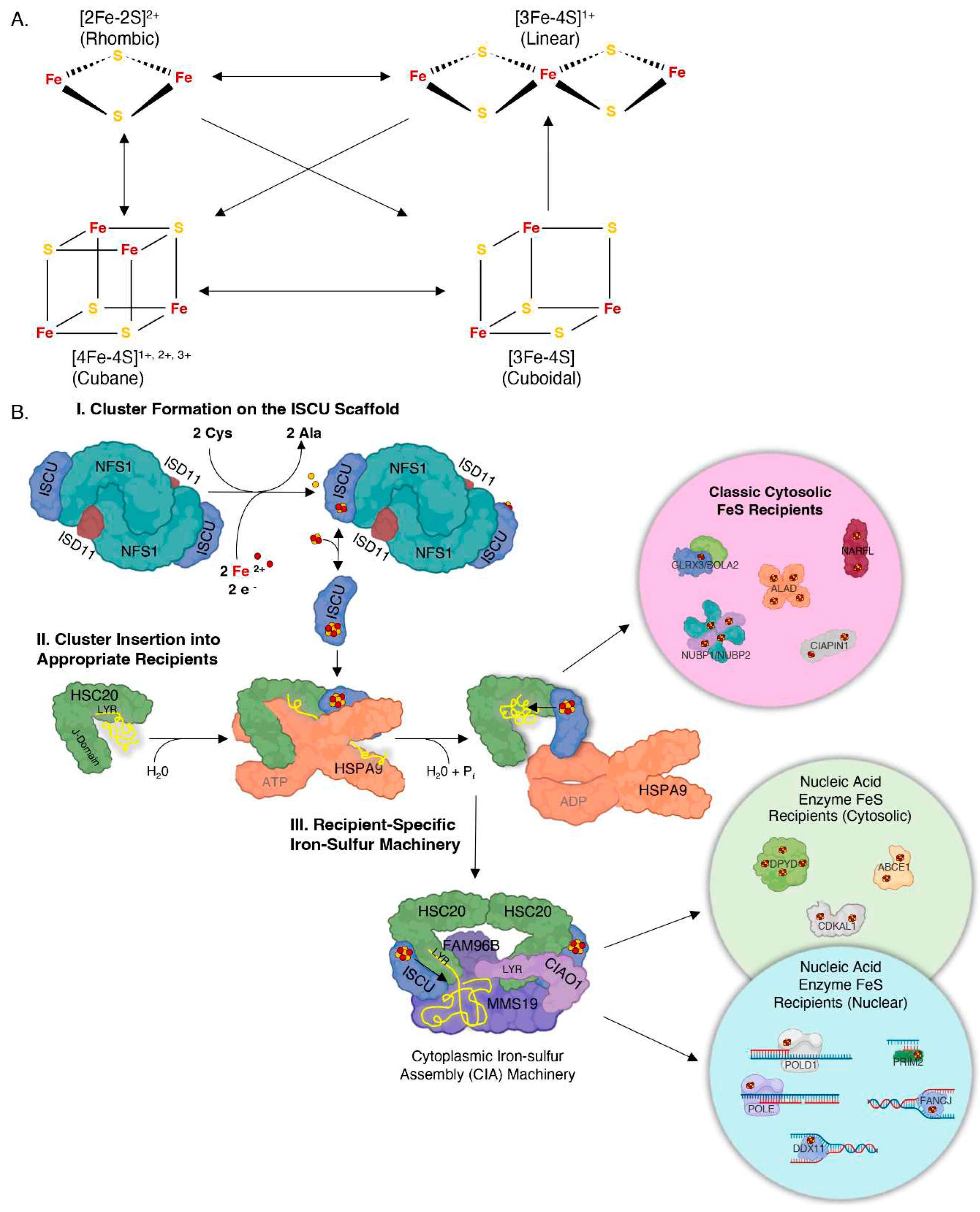
Figure 2.
The rotavirus infection cycle and its NSP5. Upon entering the cell, the rotavirus undergoes a process whereby its outer layer of triple-protein encasement is shed, allowing the virion to be released from the vesicle.63 Subsequently, within the cytosol, the double-stranded RNA (dsRNA) genome is transcribed by the polymerase VP1 while remaining confined within the virion, thereby preventing direct contact with the immune-ready cytosol.63 Positive-sense single-stranded RNAs ((+)ssRNAs) are extruded from the virion and are either translated or serve as templates for the synthesis of new virion dsRNA.63 This process involves NSP5 and NSP2. RNA transcripts forming RNA-protein condensates, known as viroplasms, serve as the location for the assembly of new virions.63 Within the viroplasms, the inner core of the virion is assembled, comprising eleven distinct strands of (+)ssRNA that act as templates for the dsRNA genome of the newly forming virion.63 Replication of these templates occurs within the core, which is subsequently encapsulated with the triple-protein layer, completing the formation of new virions.63 Figure created with the help of BioRender.com.
Figure 2.
The rotavirus infection cycle and its NSP5. Upon entering the cell, the rotavirus undergoes a process whereby its outer layer of triple-protein encasement is shed, allowing the virion to be released from the vesicle.63 Subsequently, within the cytosol, the double-stranded RNA (dsRNA) genome is transcribed by the polymerase VP1 while remaining confined within the virion, thereby preventing direct contact with the immune-ready cytosol.63 Positive-sense single-stranded RNAs ((+)ssRNAs) are extruded from the virion and are either translated or serve as templates for the synthesis of new virion dsRNA.63 This process involves NSP5 and NSP2. RNA transcripts forming RNA-protein condensates, known as viroplasms, serve as the location for the assembly of new virions.63 Within the viroplasms, the inner core of the virion is assembled, comprising eleven distinct strands of (+)ssRNA that act as templates for the dsRNA genome of the newly forming virion.63 Replication of these templates occurs within the core, which is subsequently encapsulated with the triple-protein layer, completing the formation of new virions.63 Figure created with the help of BioRender.com.
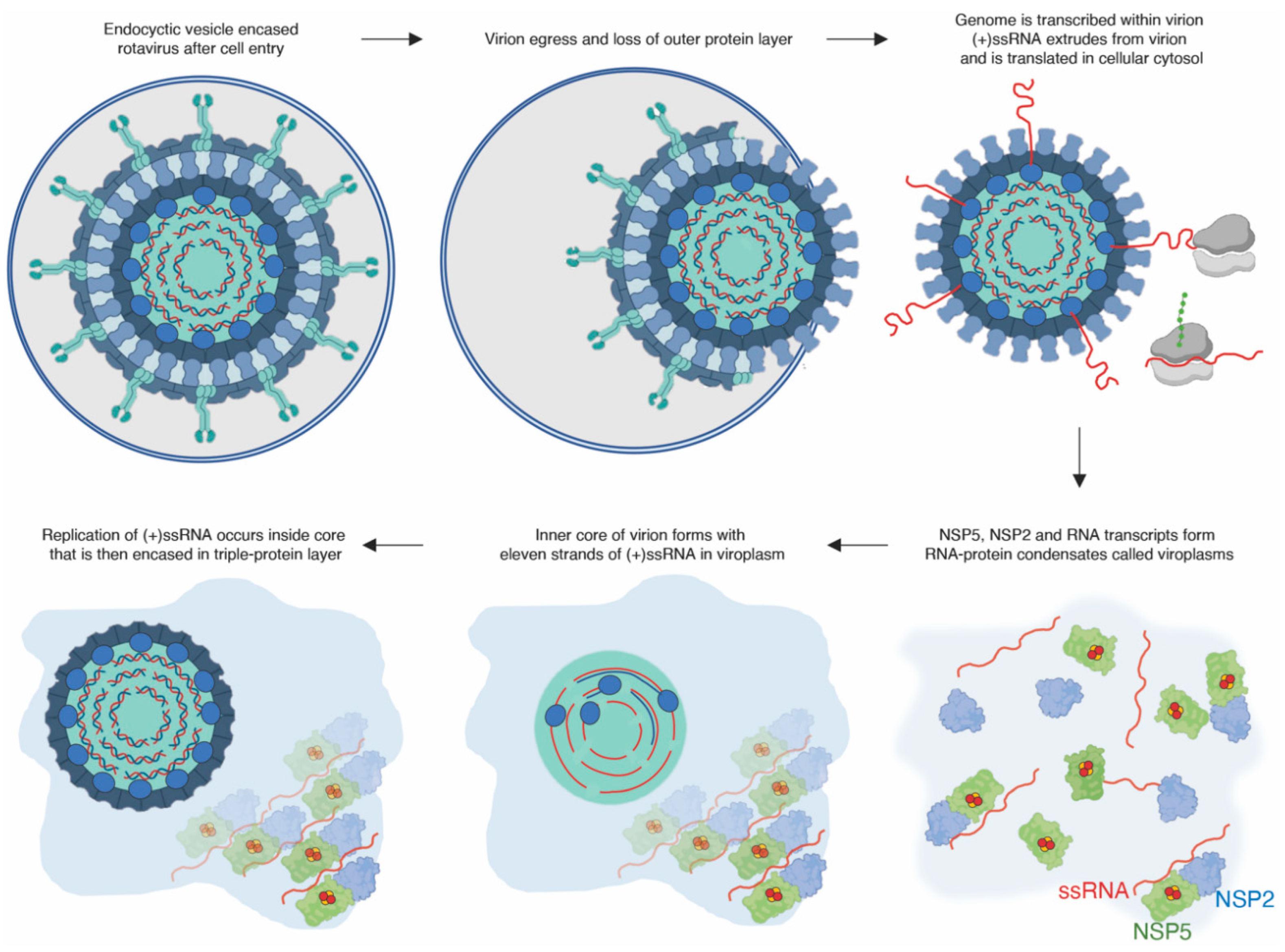
Figure 3.
Merkel-cell polyomavirus, sT. A. The circular double-stranded DNA (dsDNA) genome of Merkel cell polyomavirus undergoes replication within the cell nucleus through the involvement of cellular replication proteins.68 B. The early genes Large T-antigen (LT) and small-T antigen (sT) share N-terminal regions. LT comprises a C-terminal origin binding domain and a helicase domain, while sT contains an LT stabilizing domain (LSD) and protein phosphatase 2A (PP2A) binding domains.68 C. The structure of LT's origin binding domain interacts asymmetrically with dsDNA at the replication origin (PDB: 4FB3). D. The structure of LT-helicase assembles as a heptamer. Zinc-ligating residues are highlighted (PDB: 5J40). E. The structure of sT from a closely related virus SV40 exhibits similar metal-binding sites as sT of Merkel cell polyomavirus that are highlighted (PDB: 2PF4). The figure was created with the assistance of BioRender.com.
Figure 3.
Merkel-cell polyomavirus, sT. A. The circular double-stranded DNA (dsDNA) genome of Merkel cell polyomavirus undergoes replication within the cell nucleus through the involvement of cellular replication proteins.68 B. The early genes Large T-antigen (LT) and small-T antigen (sT) share N-terminal regions. LT comprises a C-terminal origin binding domain and a helicase domain, while sT contains an LT stabilizing domain (LSD) and protein phosphatase 2A (PP2A) binding domains.68 C. The structure of LT's origin binding domain interacts asymmetrically with dsDNA at the replication origin (PDB: 4FB3). D. The structure of LT-helicase assembles as a heptamer. Zinc-ligating residues are highlighted (PDB: 5J40). E. The structure of sT from a closely related virus SV40 exhibits similar metal-binding sites as sT of Merkel cell polyomavirus that are highlighted (PDB: 2PF4). The figure was created with the assistance of BioRender.com.
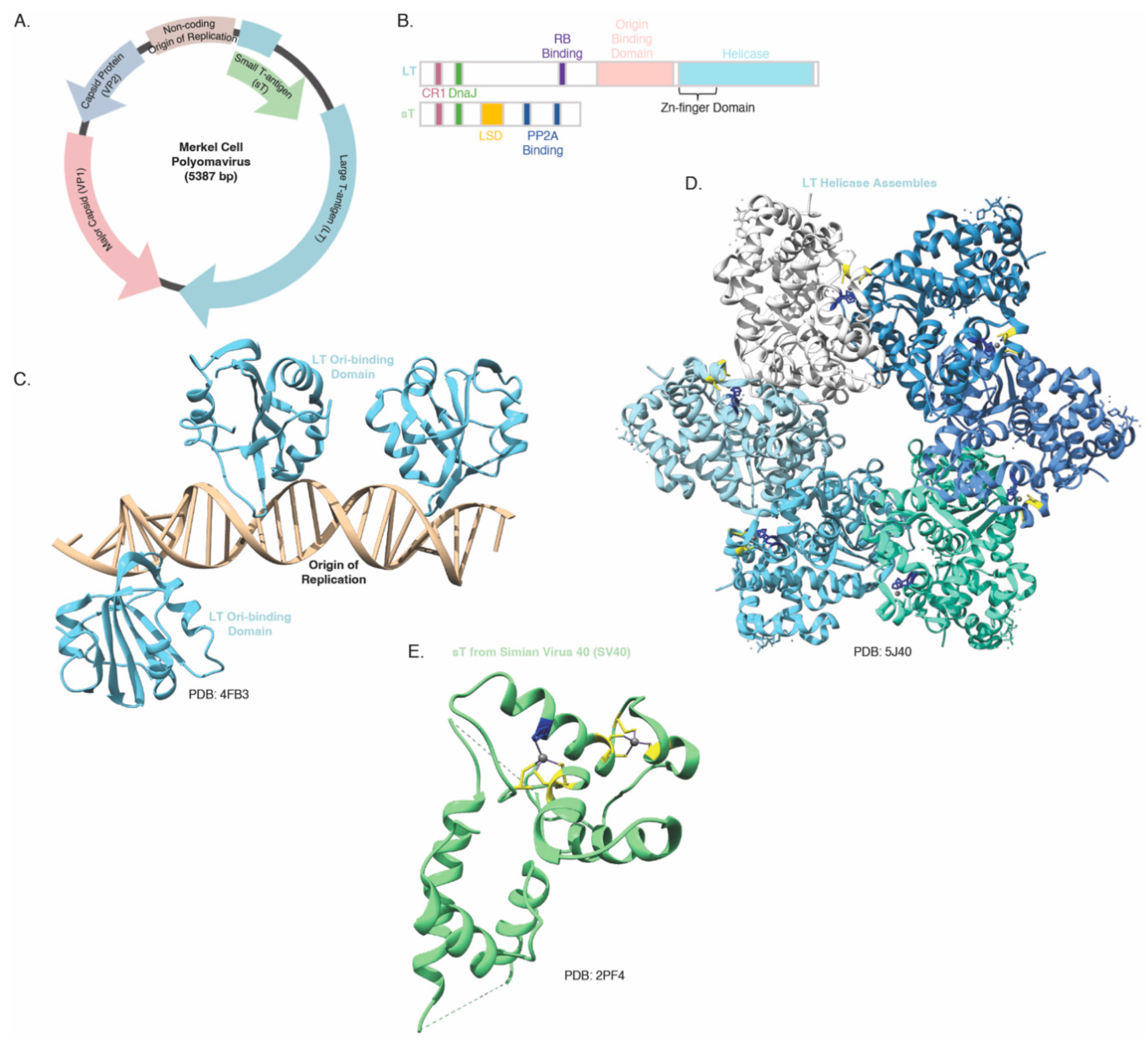
Figure 4.
SARS-CoV-2 RNA-dependent RNA polymerase, nsp12, and helicase, nsp13. A. Structure of the SARS-CoV-2 RNA-dependent RNA polymerase (RdRp) consisting of nsp12, a dimer of nsp8 and nsp7, along with a dimer of the helicase nsp13 and double-stranded primer-template RNA (PDB: 7CYQ). B. Domain organization of nsp12 and nsp13 with nidovirus specific domains highlighted.84, 80, 88, 89 C. Organization of the (+)ssRNA genome of SARS-CoV-2 and (+) subgenomic RNAs that encode structural and accessory proteins.80, 121 D. Schematic representation of continuous and discontinuous processes of SARS-CoV-2 replication. The continuous synthesis involves the replication of the (+)ssRNA genome into (-)ssRNA, forming double-stranded RNA as the product, with nsp13 unwinding the double-stranded RNA in the 5’ – 3’ direction. The (+)ssRNA strand is then synthesized using the (-)ssRNA as a template. On the other hand, discontinuous synthesis occurs at the 3’ end of the genome, where stalling during replication leads to template switching, allowing the 5’ leader sequence to serve as a template for the remaining (-) subgenomic RNA through base-pairing with the internal Transcription Regulatory Sequences (TRSs). This process results in the synthesis of the (+) subgenomic RNA using the (-) subgenomic RNA as a template.76, 121.
Figure 4.
SARS-CoV-2 RNA-dependent RNA polymerase, nsp12, and helicase, nsp13. A. Structure of the SARS-CoV-2 RNA-dependent RNA polymerase (RdRp) consisting of nsp12, a dimer of nsp8 and nsp7, along with a dimer of the helicase nsp13 and double-stranded primer-template RNA (PDB: 7CYQ). B. Domain organization of nsp12 and nsp13 with nidovirus specific domains highlighted.84, 80, 88, 89 C. Organization of the (+)ssRNA genome of SARS-CoV-2 and (+) subgenomic RNAs that encode structural and accessory proteins.80, 121 D. Schematic representation of continuous and discontinuous processes of SARS-CoV-2 replication. The continuous synthesis involves the replication of the (+)ssRNA genome into (-)ssRNA, forming double-stranded RNA as the product, with nsp13 unwinding the double-stranded RNA in the 5’ – 3’ direction. The (+)ssRNA strand is then synthesized using the (-)ssRNA as a template. On the other hand, discontinuous synthesis occurs at the 3’ end of the genome, where stalling during replication leads to template switching, allowing the 5’ leader sequence to serve as a template for the remaining (-) subgenomic RNA through base-pairing with the internal Transcription Regulatory Sequences (TRSs). This process results in the synthesis of the (+) subgenomic RNA using the (-) subgenomic RNA as a template.76, 121.
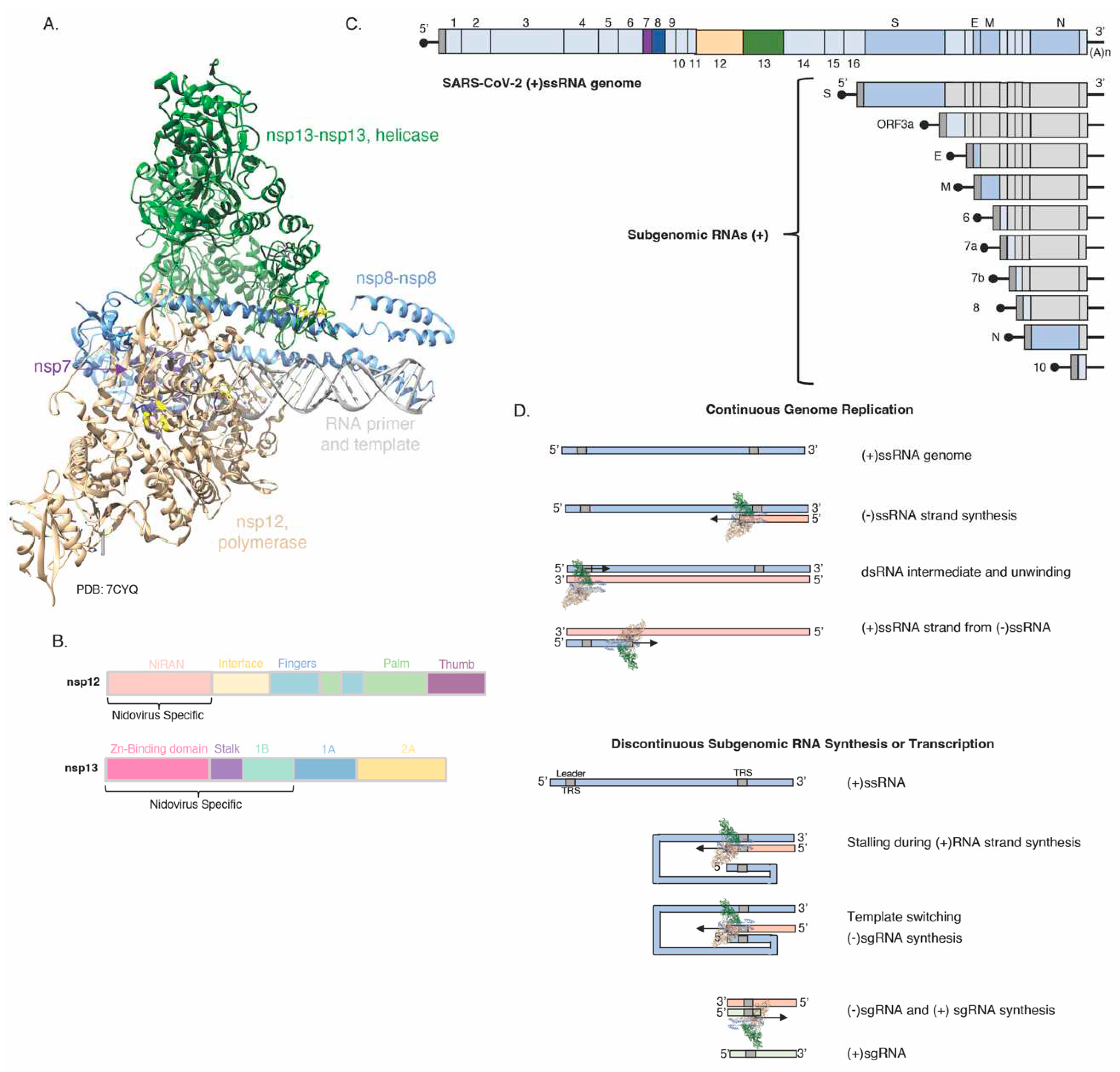
Figure 5.
Hepatitis B Virus, HBx. A. Computational model of HBx from van Hemert and colleagues, highlighting conserved histidines and cysteines marked in blue and yellow respectively.106 B. Depiction of HBx domain architecture indicating the role of the N terminus in signal transduction and of the C terminus as a nuclear transcriptional activator.99 C. Diagram of cluster conversion observed in HBx as monitored by Mossbauer spectroscopy.111.
Figure 5.
Hepatitis B Virus, HBx. A. Computational model of HBx from van Hemert and colleagues, highlighting conserved histidines and cysteines marked in blue and yellow respectively.106 B. Depiction of HBx domain architecture indicating the role of the N terminus in signal transduction and of the C terminus as a nuclear transcriptional activator.99 C. Diagram of cluster conversion observed in HBx as monitored by Mossbauer spectroscopy.111.
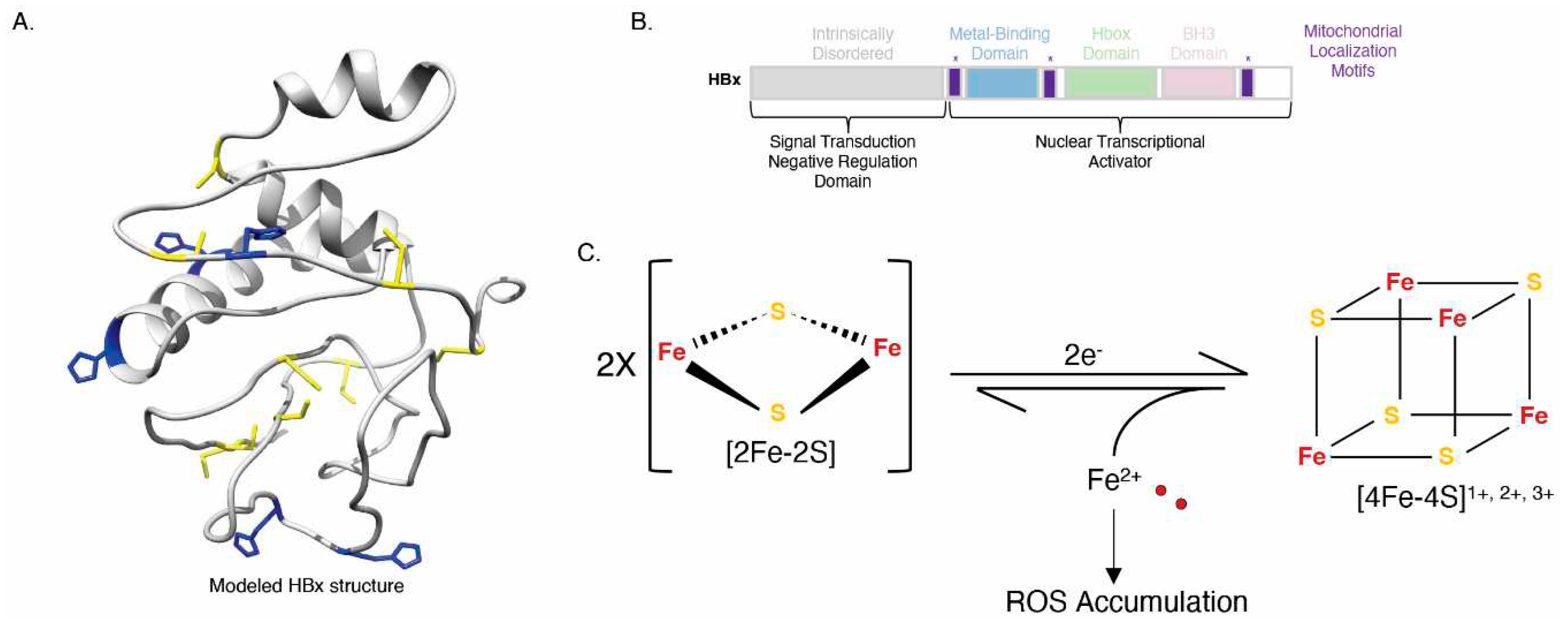
Figure 6.
Mimivirus, GciS. A. Sequence of GciS with cysteines highlighted in yellow.113 B. Diagram of cluster conversion observed in GciS by Mossbauer spectroscopy.113.
Figure 6.
Mimivirus, GciS. A. Sequence of GciS with cysteines highlighted in yellow.113 B. Diagram of cluster conversion observed in GciS by Mossbauer spectroscopy.113.
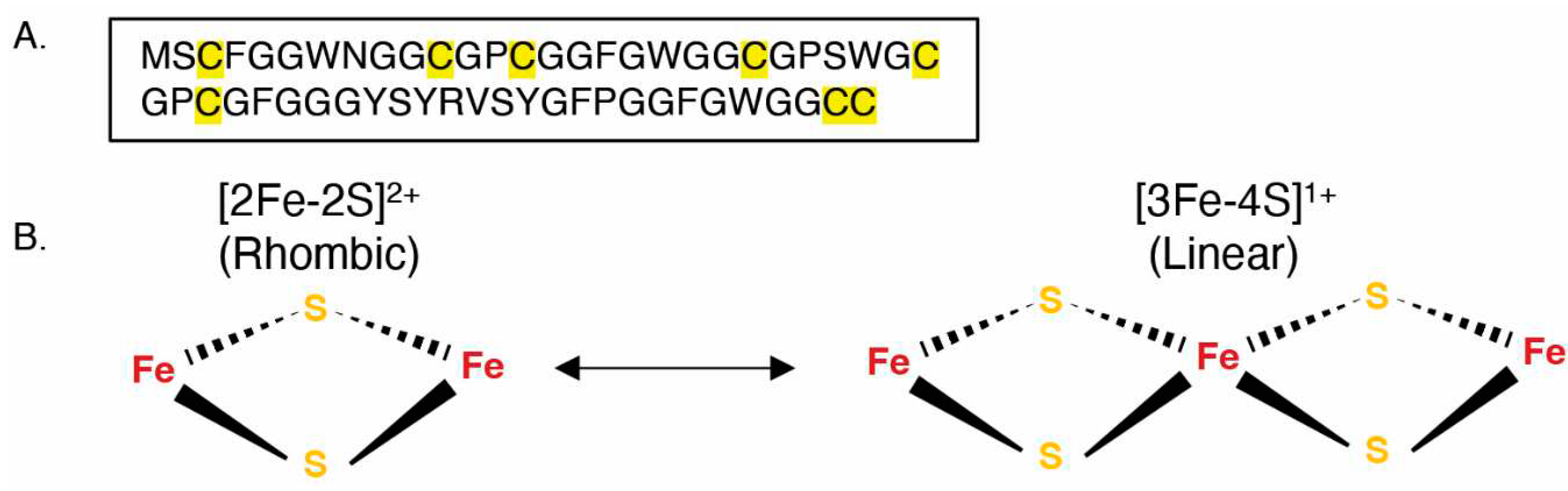

Table 1.
List of viruses thus far identified that encode FeS proteins.
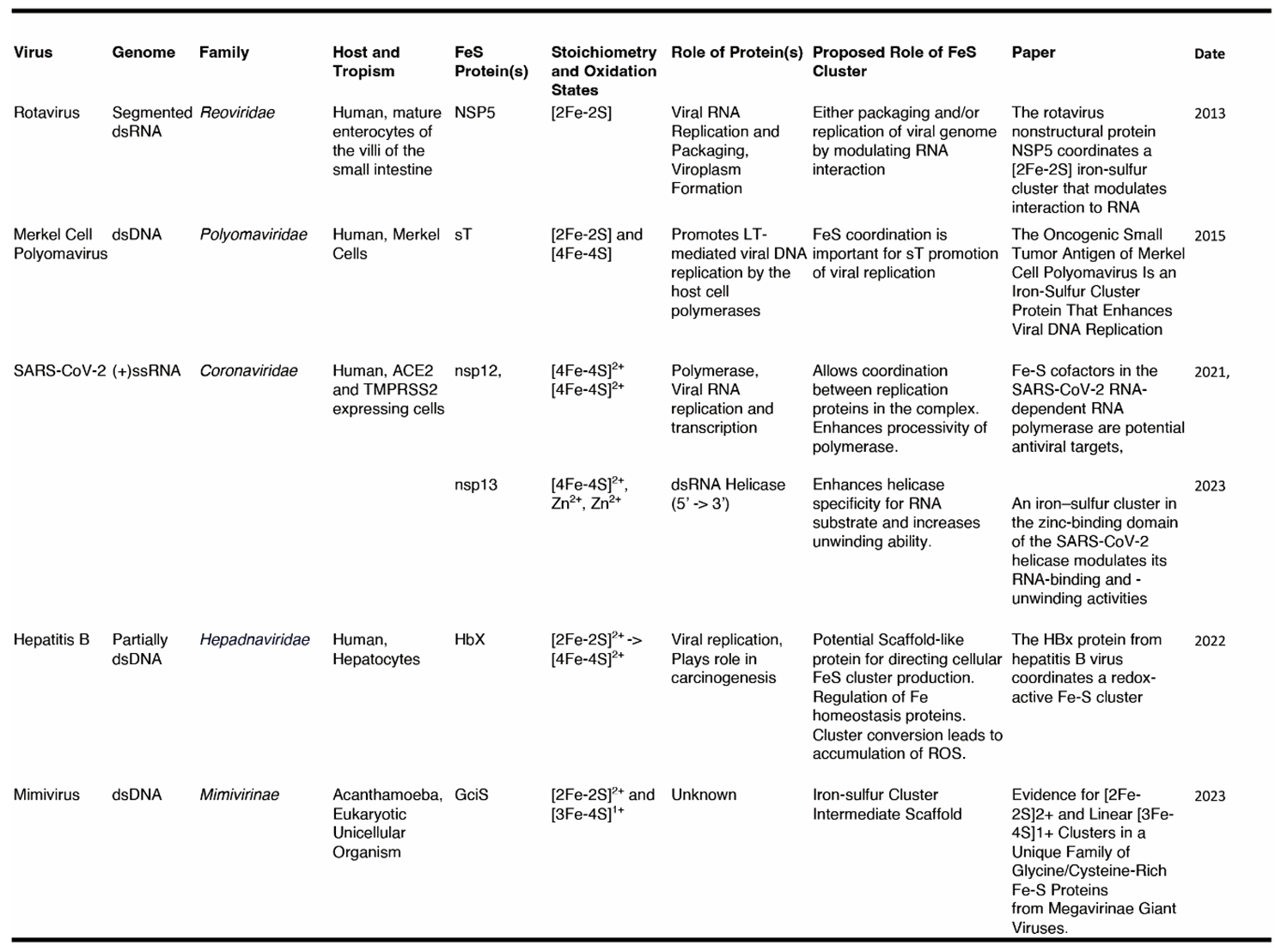
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



