Submitted:
12 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
A single type I RC (Fe-S type, ferredoxin reducing) is found in several of the phyla of anoxygenic phototrophic bacteria. They include the heliobacteria (HB), the green sulfur bacteria (GSB) and the chloroacidobacteria (CAB), for which high-resolution homodimeric RC-PS structures have recently appeared [1-4]. The 2.2-Å Xray structure of the RC-PS of Heliomicrobium modesticaldum [1] revealed that the core PshA apoprotein (PshA1 and PshA2 homodimeric pair) exhibits a structurally conserved PSI arrangement comprised of five C-terminal transmembrane helices (TMHs) forming the RC domain and six N-terminal TMHs coordinating the light-harvesting (LH) pigments. The Hm. modesticaldum structure lacked quinones, indicating that electrons were transferred directly from the A0 (81-OH-chlorophyl (Chl) a) primary acceptor to the FX [4Fe4S] component, serving as the terminal RC acceptor. A pair of additional TMHs designated as Psh X were found, that function as a low-energy antennae. The 2.5Å resolution cryo-electron microscopy (cryo-EM) structure for the RC-PS of the GSB Chlorobaculum tepidum [2] included two Fenna-Matthews-Olson protein (FMO) antennae, which transfer excitations from the chlorosomes to the RC-PS (PscA1 and PscA2) core. Two cytochromes c (PscC) molecules were revealed, which act in series as electron donors to the photooxidized RC BChl a’ special pair, as well as PscB, housing the [4Fe-4S] cluster FA and FB, and the associated PscD protein [3]. Although their FMO components were missing from the 2.6-Å cyro-EM structure of the Zn-BChl a’ special pair containing RC-PS of Chloroacidobacterium thermophilum [4], a unique architecture was revealed that consisted of seven additional subunits including PscZ in place of PscD, the PscX and PscY cytochrome c serial electron donors and four low mol. wt. subunits of unknown function. Overall, these diverse structures have revealed that: (i) the HB RC-PS is the simplest light-energy transducing complex yet isolated, and represents the closest known homolog to a common homodimeric RC-PS ancestor; (ii) the symmetrically localized Ca2+binding sites found in each of the Type I homodimeric RC-PSs likely gave rise to the analogously positioned Mn4CaO5 cluster of the PSII RC and the TyrZ RC donor site; (iii) a close relationship between the GSB RC-PS and Chl protein (CP)43 and CP47 of PSII was demonstrated by the strongly conserved locations of their LH-(B)Chls; (iv) LH-BChls of the GSB-RC-PS are also localized in the conserved RC-associated positions of the PSII ChlZ-D1 and ChlZ-D2 sites; (v) glycosylated carotenoids of the GSB RC-PS are located in the homologous carotenoid-containing positions of PSII, reflecting an O2-tolerance mechanism capable of sustaining early stages in the evolution of oxygenic photosynthesis. In addition to the close relationships found between the homodimeric RC-PS and PSII, duplication of the gene encoding the ancestral Type I RC apoprotein, followed by genetic divergence my well account for the appearance of the heterodimeric Type I and Type II RC found in extant oxygenic phototrophs. Accordingly, the long-held view that PSII arose from the anoxygenic Type II RC is now supplanted by the new evidence provided by Type I RC-PS homodimer structures, indicating that the evolutionary origins of anoxygenic Type II RCs along with their distinct antenna rings are likely to have occured later than the events which gave rise to their oxygenic counterparts.
Keywords:
chloroacetobacteria
; green sulfur bacteria
; heliobacteria
; membrane proteins
; molecular evolution
; photosystems
; protein structure
; reaction centers

1. Introduction
In contrast to the heterodimeric Type II reaction centers (RCs) (pheophytin-quinone (Q) type, Q reducing) found in the well-studied anoxygenic purple chlorophototrophic bacteria, homodimeric Type I RCs ([4Fe-4S] cluster type, ferredoxin reducing) catalyze the primary photochemical reactions in the heliobacteria (HB), green sulfur bacteria (GSB) and chloroacidobacteria (CAB)[5]. These anoxygenic Type I structures are believed to have arisen from the primordial homodimeric RC (the last common RC ancestor) that subsequently gave rise to both the heterodimeric photosystem (PS) I and PSII RCs of the oxygenic phototrophs [6], which catalyze transmembrane charge separations resulting in the respective reduction of [4Fe-4S] centers and Q molecules, as accompanied by O2 evolution by PSII. TheType I bacterial RC-PS protomers [1,2,3,4], designated as PshA in HB and as PscA in GSB and CAB, contain 11 transmembrane α-helices (TMHs), with the six N-terminal helices binding the light-harvesting (LH) BChls that transfer excitation energy to the RC cofactors housed within the five C-terminal helices. This structural organization is conserved in the multifunctional PsaA and PsaB proteins forming PSI, which also possess fused antenna and RC domains [7]. By contrast, the structurally conserved Type II RCs of the purple bacteria are surrounded by unique elliptical assemblies of LH1 proteins consisting of up to 17 subunits to form RC-LH1 complexes (the structures of these RC-LH1 complexes have been recently reviewed in detail [8,9,10,11]).
The electron transferring cofactors of the homodimeric Type I RCs include the P800/P840 special pair BChls, two accessory BChls (ACC) and two Chl (AO) acceptors, which together with the [4Fe-4S] cluster (FX), form a bifurcated chain capable of functioning in the absence of the A1 menaquinone (MQ) component which transfers electrons between the AO and FX electron carriers of PSI [5]. The light-driven charge separations created by the homodimeric Type I RC powers cyclic electron transfer pathways creating a transmembrane electrochemical proton gradient which drives adenosine 5’-triphosphate synthesis. The remaining BChl molecules, which include 56, 26 and 24 BChls in the HB, GSB and CAB, respectively [1,2,3,4], function in LH capacities, with the additional HB BChls representing an expanded PshA associated antenna, in part compensating for the lack of chlorosomes and the Fenna-Matthews-Olson protein (FMO) antennae present in the GSB and CAB.
In this review, I will describe the recently elucidated structures of the homodimeric Type I RC-PSs from HB [1], GSB [2,3] and CAB [4], with emphasis on their structural diversity and the manner in which they provide an improved understanding of both the LH and primary photochemical events catalyzed by these unique pigment-protein complexes. Moreover, I will detail how these structures have contributed to our current understanding of the evolutionary origins of RC-PSs, especially with regard to the relationships which the observed homodimeric architectures have to the emergence of the extant heterodimeric PSI and PSII complexes of the oxygenic phototrophs.
2. Type I Homodimeric Reaction Center-Photosystems of Anoxygenic Phototrophs
Unlike higher phototrophs, in which separate psaA and psaB genes encode the PsaA and PsaB apoproteins that form heterodimeric Type I RCs, a single pshA gene encodes the the HB RC-PS homodimer, while the RC-PSs of GSB and Cab are encoded by single pscA genes. These unique homodimeric RCs apparently arose from the last common PSI ancestor that served as the evolutionary precursor of the heterodimeric Type I RC [12] in which duplication of the gene encoding an ancestral core homodimeric RC apoprotein, followed by genetic divergence likely accounts for the appearance of the Type I RC in their heterodimeric form [13]. While high-resolution structures have long been available for the heterodimeric RCs of the purple bacteria [14,15] as well as for both PSI [7] and PSII [16], elucidation of the structures of Type I RC-PS complexes of the HB, GSB and CAB have proved to be of considerable importance in obtaining a more complete picture of these early photosynthetic proteins and pigments, with their apparent close relationships to the ancestral RC protein.
2.1. Structure of the Reaction Center-Photosystem of the Heliobacteria
The first structure of a Type I homodimeric RC-PS was determined for Heliomicrobium modesticaldum [1], a thermophilic anaerobe isolated from hot spring and volcanic soil environments [17]. Hm. modesticaldum, as a member the family Firmicutes, is among the simplest known complexes capable of catalyzing primary photochemical reactions. In addition, this RC-PS is considered most closely related to the common RC-PS progenitor [1,12]. Figure 1A,B display the 2.2-Å resolution X-ray crystallography structure determined for the Hm. modesticaldum RC-PS by Gisriel et al. [1]. The structure exhibits perfect C2 symmetry, suggesting that it is a likely close homolog of the homodimeric ancestral RC, as also supported by the anoxic niches in which HB are found, comparable to that of the early earth. Hm. modesticaldum is further unique in lacking the peripheral antenna components found in the GSB and CAB, as well as in containing a RC-PS largely utilizing BChl g, an isomer of Chl a [18].
The overall molecular architecture is structurally homologous to known Type I RCs which is confirmed in Figure 1B,C, which shows that the PshA apoprotein forms 11 TMH, with the first six containing the antenna BChls and the remaining five comprising the RC domain. Two additional α-helices labeled “X” are also found, located in approximately the same positions as the PsaI and PsaJ proteins in the heterodimeric PSI. Recent spectroscopic evidence demonstrates that protein X, which binds two BChl g molecules, serves as low energy LH component involved in the uphill transfer of excitations to the RC [19]. While both the X protein and PsaJ bind antenna (B)Chl pigments, their lack of significant sequence identity suggests the common position occupied by these subunits reflects convergent evolution. A total of 54 BChl g function as LH-BChls , while four BChl g’, two 81-hydroxy-Chl a, two 4,4’-di-aponeurosporenes, two lipids and one [4Fe-4S] cluster are also associated with the Hm. Modesticaldum RC-PS.
The C2 symmetry axis assuming cofactors of the RC electron transfer chain give rise to two essentially identical cofactor branches, as governed by the homodimeric nature of the HB RC (Figure 2A). Residues coordinating the cofactors consist of His 537 for the BChl g’ dimer special pair, together with Cys432 and Cys441 for the FX terminal acceptor as provided by the individual PshA protomers which give rise to the symmetrical arrangement. However, the A0 81-OH-Chl a primary acceptor and the ACC accessory Bchl g’molecules (Recent 2-D hyperfine sublevel correlation spectroscopy and density function theory studies of PSI [20] demonstrated that Chl2A and Chl3A located in the respective positions of the heliobacterial ACC and A0, form a Chl dimer that functions as the primary electron acceptor in which extensive delocalization promotes a prolonged charge separation, resulting in the high quantum efficiency of PSI. This mechanism may apply to the HB RC, since ACC (BChl g) and A0 (81-OH-Chl a) possess the conserved orientation similar to the homologous PS I cofactors and the ~9.0 Å Mg → Mg distances between them [1,7].) are likely coordinated by respective water molecules and an unidentified molecule of a size similar to a chloride ion, respectively, which are in turn H-bonded to the respective Ser545 and Gln458 side chains.
Up to two MQ molecules have been found in purified HB RCs [22], despite the absence of a conserved quinone binding site. The MQ molecules are loosely bound and were lost during purification of the crystalline RC-PS [1]. Recently, a model was constructed by Kashey et al. [21] (Figure 2B) in which MQ molecules were placed in potential locations between A0 and FX. This was based upon unassigned electron density in the crystal structure sufficient for placement of an isoprenyl tail (Figure 2C). A comparison of the phylloquinone binding site of PSI with the potential quinone site for PsaA (Figure 2D) revealed conservation of only a Leu residue homologous to the PsaA-Leu722 on one sides of the naphthoquinone head group, while the Trp697 on the opposite side and the nearby Phe689 are replaced by Arg554 and Met546 residues, respectively, in the HB RC. Since the distance between A0 and FX is shorter than in PSI where the electron carrier phylloquinone is positioned between them, electron transfer in the HB is capable of occurring between A0 and FX at an intrinsic slowed rate without the need for an intermediate quinone. Nevertheless, MQ was shown to function as an alternative electron carrier in HB RC under high-light conditions, with the final soluble acceptor pool replete [21]. This ability to alternatively engage MQ as the terminal RC electron acceptor points to the inadequacy in the classification of the HB RC as solely belonging to Type I (Fe-S type, ferredoxin reducing) [12].
The loosely bound PshB subunit lost during purification of the HB RC-PS core complex is considered to function as a ferredoxin electron acceptor [23], rather than as the bound FA and FB terminal electron acceptors of the PsaC subunit of PSI. This requires that FX functions as the bound terminal electron acceptor [23]. Patches of positive charge on the cytoplasmic surface of the HB RC provided by Lys584 and Lys587 of PshA, as well as twin Lys423 residues extending out on either side of the FX site have been modeled as a potential interaction site for the negatively charged surface of PshB [1].
It is generally accepted that photochemical RCs arose only once in the form of a homodimeric complex more than 3 billion years ago and ultimately evolved into currently existing heterodimeric versions. While the homodimeric Hm. modesticaldum PS represents the simplest such complex isolated thus far, it is not thought to represents the actual ancestral form, but rather a close homolog of the RC common ancestor [24]. This is based upon the likelihood that the presence of this PS protein in Hm. modesticaldum originated from horizontal gene transfer, a possibility supported by the presence of pshA in a single gene cluster along with the pigment synthesis genes [25].
A comparison of structural details from the homodimeric HB RC with all known RC structures has led to proposals on the structural and functional properties of a common RC ancestor as related to the evolutionary origin of the resulting complexes [12]. Emphasis was placed upon the loose Q-binding capacity of the HB RC thought also to be a property of the homodimeric ancestral complex, purported to bind two mobile Q molecules that reduce Q through an inefficient disproportionation reaction involving simultaneous oxidation and reduction of Q redox species. Optimization of this reaction through subsequent evolutionary changes led to a divergence into the two types of extant RCs. For Type I RCs, a [4Fe-4S] cluster was added that enables double Q reduction, while heterodimerization led to affixing a Q molecule into the QA site on the D1 protomer of PSII within electron transferring distance from the second (QB) binding site on D2, where a gated double Q reduction to a hydroquinone is catalyzed via a radical anion Q intermediate in Type II RC. In Type I RCs, subsequent fixation of the Q sites was followed by heterodimerization to permit association of the PsaC subunit housing the FA and FB [4Fe-4S] clusters, in response to O2 levels arising after the oxygen-evolving complex had appeared. Further possible evolutionary scenarios will be discussed in Section 3, following presentation of the GSB and CAB RC-PS structures (Section 2.3 and Section 2.4, respectively).
2.2. Structure of the Reaction Center-Photosystem of the Green Sulfur Bacteria
The green sulfur bacterium Chlorobaculum tepidum, like Hm. modesticaldum is a thermophilic obligate anaerobe that also contains a Type I homodimeric RC with the RC protein (PscA)2 as a core. The LH capabilities of Cba. tepidum are considerably enhanced by a chlorosome peripheral antenna system containing self-assembled Chl c molecules that exist as a lamellar structure lacking a protein scaffold [26]. The chlorosomes is connected by a baseplate to two copies of the trimeric FMO protein, in which each protomer contains eight BChls a from which excitons are passed onto the RC-core of the RC-PS complex. While energy transfer efficiencies between the Chls c within chlorosomes to the FMO BChls a, and within FMO approach the 100% levels found for transfer from LH Chls to the PSI RC, the efficiencies in the GSB range from only 35 to 75% for the FMO trimer to the RC-PS [27,28,29,30]
In an effort to elucidate the basis for the reduction in energy transfer efficiency and to obtain an improved understandings of the overall excitation energy and electron transfer pathways, a complete 2.5 Å cryo-EM structure of the Cba. tepidum RC-PS was recently determined by Xie et al. [2](Figure 3A,B). This was accomplished with a complex purified after solubilization with n-dodecly-β-D-maltoside and detergent exchange to lauryl maltoside neopentyglycol; the resulting structural model follows an earlier 2.7 Å cryo-EM structure [3] which lacked the cytochromes cZ (PscC-1 and PscC-2), as well as a second FMO trimer subunit.
The core molecular architecture of the GSB complex is also homologous to the known Type I RC structures (Figure 3C) in which the PscA protomer forms 11 transmembrane α-helices, with the first six binding the antenna BChls and the remaining five associated with the RC cofactors [2,3]. Along with the 82-KDa core PscA-1 and PsaA-2 protomers and the two accompanying, asymmetrically positioned FMO trimers, four additional subunits were assigned [2]: two identical 23-kDa cytochromes cZ (PscC-1 and PscC-2); the 24-KDa PscB housing the FA and FB [4Fe-4S] clusters; and the accompanying 17-KDa PscD subunit (the PscC subunits as well as the second FMO trimer were absent from the initial structure [3], reflecting loss during purification following Triton X-100 solubilization). The cofactor assignments included: 78 (B)Chls (48 BChls a bound to the two FMO trimers and 26 BChls a and 4 Chls a bound to the RC-PS); five lipids (two monogalactosyl digylerides and two phosphatidylglycerol (PG) molecules [2,3], as well an additional PG modeled adjacent to the A-1 site [2] not seen in initial structure [3]); four carotenoids (two OH-chlorobactene glucoside laurates in contact with both the cytoplasmic and periplasmic BChl layers and two chlorobactenes associated with the cytoplasmic layer [3]); and 2 Ca2+ ions and 3 [4Fe-4S] clusters (FX, FA and FB)[2,3].
The C2 symmetry axis as assumed by the GSB RC cofactors is shown in Figure 3D, together with the coordinating residues provided by the homodimeric (PscA)2 protein [3]. The essentially identical distances between the cofactors of each branch suggests that electrons are capable of flowing down both sides. Note that the distances between A0 and FX of 18.1 and 18.2 Å [2] (reported as ~17.5 and 17.6 Å in [3]) which is shorter than the ~23 Å observed in PSI [7], and similar to the 17.9 Å for the HB RC [1]. Thus, the A0 and FX components of the GSB RCs are sufficiently close to promote the observed slowed electron transfer rate [31]. While a phylloquinone molecule intervenes in sustaining electron transfer between A0 and FX in PSI, and an inserted MQ was shown to play such a role in the HB RC under special high-light intensity conditions [21], only trace levels of Q were found in the GSB core RC-PS complex [3]. Moreover, it was not possible to model an MQ into an appropriate binding site in the GSB complex, and therefore general agreement exists that electrons are transferred directly from A0 and FX (see, however [32,33]).
The residues serving as the coordinating ligands for the FX cluster were identified as Cys527 of PscA-1 and Cys 536 of PscA-2, located at the interface between the two PscA protomers [3] as seen in other Type I RCs [1,4,7]. The PscB-associated FA and FB clusters are located at the cytoplasmic surface along with the PscD subunit (Figure 3A,B), in positions similar to the FA and FB containing PsaC and the associated PsaD in the PSI complex. While PscB has a similar structure and electron transfer role as PsaC, they show little sequence homology, as also observed between PsaD and PscD, suggesting that the PsaC and PsaD components of the heterodimeric PSI RC arose by convergent evolution. The associated PscB and PscD proteins also interact with the FMO and RC core and are in positions to contribute to the stability of the FMO-RC-PS complex [3].
A comparison of the Type I GSB RC-PS with the PSII complex (Figure 4A,B) shows that the majority of the 24 BChls a serving as antenna pigments, exhibit binding sites that are largely conserved in each of the 12 Chls a associated with the CP43 and CP47 PSII core antennae, among their respective 13 and 16 Chl a molecules [16]. Moreover, an essentially complete overlap occurs in the locations of the six GSB RC (B)Chl cofactors with those of the HB and PSI RCs (Figures4C,D), together with good agreement in the positioning of the 24 common antenna (B)Chls, with only a few minor differences in locations. The additional (B)Chls found in the HB RC-PS and in PSI are mainly located at the peripheries and gaps in each pigment cluster.
The similar arrangement of pigments in Type I and II RCs (Figure 4), is also seen in the RC of the purple chlorotrophic bacteria, together with conservation in their accompanying structural architecture. This reflects common evolutionary histories, whereas the differences in the types of both additional core and peripheral antennae reveal distinct events in evolution. This is seen in the RC-PS for both the HB and the GSB, as well as in PSI and PSII, where such differences signify their distinct phylogenic status. Thus, early in the course of evolution, the lack of pigments associated with the GSB RC-PS was compensated for by the presence of the FMO and chlorosomes, acting as peripheral antennae. Moreover, additional pigments are present in the Type I RCs of HB and PSI, while for PSII, the core LH pigments are contained within the CP43 and CP47 proteins. While the LH1 antenna rings of purple bacteria house the core LH pigments, distinct peripheral LH complexes designated as LH2, LH3 and LH4 function as accessory antennae in many anoxygenic phototrophs [34,35]. For the oxygenic phototrophs, the phycobilisomes in cyanobacteria and red algae and the LHCII and LHCI complexes of higher forms, such as land plants and green algae, also function in these accessory roles.
It is also noteworthy that when viewed parallel to the membrane plane, a similar layering of (B)Chl distributions was seen for the GSB RC-PS as observed for the HB complex, as well as in PSI and PSII [1,2,3,4,7,16]. Since the pigment distances observed between the two layers of these antenna components is longer than those within each layer, energy transfer would be slightly less efficient in the former circumstances. This problem is overcome in the HB RC-PS and in PSI and PSII by (B)Chls that act as linkers between the two pigment layers having shorter Mg-to-Mg distances (9.7-11.7 Å), allowing more rapid interlayer energy transfer. Since the GSB complex lacks such linker BChls, the energy transfer rate between the two layers is consequently slowed. Nevertheless, a carotenoid derivative modeled as OH-chlorobactene glucoside laurate was shown to connect the two pigment layers [2,3], and is capable of transferring energy to the BChls, while also functioning in their quenching, but is unable to transfer energy between the BChls in the two layers [3].
The structure obtained for the GSB RC-PS complex allows an assessment of the low efficiency energy transfer pathway from the FMO BChls to the GSB antenna assembly [27,28,29,30]. BChl-3 molecules provided by the FMO-1 and FMO-2 are located in positions to pass excitons across 21.5 and 21.7 Å gaps to respective core PS LH-BChls designated as BChls A808 and A810 of PscA-1 [2,3], located near the outer surface of the cytoplasmic pigment layer (Figure 5A). On the PscA-2 side, the BChl-3 provided by FMO-4 is positioned to pass excitons across a 27.0 Å gap onto BChl A807, while for the BChl-3 molecules provided by FMO-5, paths of 23.4 and 22.5 Å to BChl A807 and A808, respectively were seen [2](Figure 5B). In contrast, while efficient energy transfer occurs between a gap of 8 to 13 Å between LHCI-Chls and PSI antenna pigments [36,37] as aided by numerous Chls occupying the gap space, in the case of the GSB RC-PS no such intragap BChls were found, and these much longer distances clearly account for the low efficiency for energy transfer from the FMO to the core RC-PS. Since BChls A811 and A815 of the GSB photosystem are located closest to the RC P840 BChl special pair (respective edge-to-edge distances of 12.0 and 13.4 Å [3]; respective Mg-to-Mg distances of 20.4 and 18.9 Å [3](Figure 5C)), they are in a position to perform the final energy transfer step. The energy transfer distances provided by these structural considerations provide a blueprint for future theoretical studies aimed at further elucidating relevant pathways culminating in exciton transfer to P840, facilitating the initiation of the RC charge separated state.
Two copies of the unique cytochrome cZ (PscC-1 and -2) are associated with the GSB RC (Figure 3A,C). They exist as integral membrane proteins each with two TMHs, and function in the re-reduction of the photooxidzed P840+ BChl special pair, using electrons obtained from the oxidation of reduced sulfur compounds [38]. Due to the flexibility of the heme c-containing C-terminal portion of PscC which during electron transfer, fluctuates between the cytochrome bc1 complex and P840 [39], it was not possible to model this domain into the RC structure [2]. Instead, the crystal structure of the Cba. tepidum cytochrome cZ electron carrier domain [40] was utilized in a molecular docking analysis providing a detailed picture of P840 BChl special pair re-reduction site (Figure 6A). Although both PscC-1 and PscC-2 are associated with the GSB RC, only a single heme c bind-site was revealed [2]. The parallel PscA-1 and PscA-2 Trp residues (Trp601) located in the PscC crevices, participate in the formation of the heme docking site, in which they are positioned at a distance of 2.4 Å from the heme c propionate group of PscC (Figure 6A). A similar set of twin Trp residues, separately provided by PsaA and PsaB in the cyanobacterium Thermosynechococcus elongatus, participates in the docking between PSI and cytochrome c6 [41]. In the PscC docking site, the exposed K168 and R181 residues at the heme edges interact with the negatively charged PscA surface. In addition, the shortest observed center-center distance between the cytochrome cZ heme and P840 of 21.9 Å (Figure 6A) compares favorably with the distances observed for the hemes of T. elongatus cytochrome c6 [41] and the Cereibacter sphaeroides (formerly Rhodobacter sphaeroides) cytochrome c2 [42] and their primary RC donors (21.1 Å and 21.3 Å respectively). Despite these advances in elucidation of the mechanism of GSB RC special-pair reduction by cytochrome cZ , the presence of parallel pathways of energy transfer to P840, while only a single site exists for the re-reduction this donor site, leaves open the question of how the two cytochromes communicate in supplying necessary electrons [2].
As first observed for the ferredoxin binding site of PSI, which includes basic residues provided by both PsaA and PsaC [43], participating residues provided by the in GSB-RC (Figure 6B) include K149 and K153 of PscB, as well K45, R47 and R48 of PscA; each of these residues are grouped in the vicinity of FB [4Fe-4S] cluster. While an insertional mutant of the Cba. tepidum pscD showed a much slower rate of energy transfer to P840 from the antenna pigments, the photoreduction of NADP+ by ferredoxin was only slightly diminished [44], apparently reflecting an additive relationship of PscD to the PscB catalyzed ferredoxin reduction. Nevertheless, interactions between PscB and PscD are thought to promote the docking of ferredoxin, while also contributing to the electron exit site [2].
Overall, these crucial structural studies have revealed the surprising finding that the positioning of the LH-BChls in the Type I GSB RC-PS homodimer most closely resembles that of the PSII antenna, in which the core GSB LH-Chls occupy two separate clusters similar to that provided in PSII by the CP43 and CP47 proteins located one either side of the electron transferring cofactors associated with the respective D1 and D2 proteins. Notably, both LH domain provided by PscA of the GSB-RC and the PSII core LH proteins contain fewer core antenna pigments than the HB RC-PS and PSI. Lastly, the recent recognition that the PSII water-oxidizing Mn4CaO5 cluster is located in a homologous position to the Ca2+-binding site associated with the bacterial Type I RCs, and also possesses a number of structural similarities, further supports a close relationship between these homodimeric structures and PSII [6,61]. These key findings [2,3] provide further examples of how the elucidation of new structural details of the Type I homodimeric RC-PS complexes aid in our understanding of the origins of heterodimeric photosystem. Moreover, the detailed molecular architecture of the entire GSB RC-PS [2] provides an incentive for further experimental and theoretical examination of the energy and electron transfer steps catalyzed by this photosynthetic prokaryotic system.
2.3. Structure of the Reaction Center-Photosystem of the Chloroacidobacteria
Chloroacidobacterium thermophilum was first recognized in a metagenomic analysis of microbial mats retrieved from an alkaline hot spring, representing the earliest phototrophic member of the phylum Acidobacteria to be identified [45]. The Type I homodimeric RC-PS isolated from Cab. thermophilum [46,47] has a unique pigment content consisting of three forms of (B)Chl, including Zn-BChl a’ [48]. FMO and chlorosomes are also present as peripheral antennae [49], as seen for the GSB [26,27,28].
The structure of the Type I RC-PS of Cab. thermophilum, recently determined by cyro-EM at 2.6 Å resolution [4], is shown in Figure 7A,B. Besides the homodimeric PscA
(PscA-1 and PscA-2) protomers, eight additional subunits were found which included PscB, housing the terminal [4Fe-4S] cluster electron acceptors FA and FB , which is accompanied by PscZ, which has an apparent PscB stabilizing role (Table 1). Also included are the distinct membrane-associated monoheme cytochrome c subunits PscX and PscY, which form a heterodimer in which the hemes c are located within deep protein grooves where they serve as the respective secondary and primary electron donors to the RC P840 special pair. In addition, newly identified low mol. wt. subunits Psc U, PscV, PscW and UPP of unknown function were identified. The RC pigments and cofactors consist of the unique P840 (Zn-BChl a’ special pair), ACC (Chl a), A0 (Chl a), and the terminal [4Fe4S] electron acceptors (FX, FA, and FB)(Figure 7C). Two phosphatidyl-N-methylethanolamine (PME), and notably, 2 Ca2+ ions were also associated with the RC. Possible roles of the requisite prokaryotic Ca2+-Type I RC binding sites in the evolution of the PSII H2O oxidizing Mn4CaO5 cluster are discussed in detail below. Additional molecules identified in other regions of the structure included 16 BChls a, 6 Chl a, 10 lipids (4 PMEs, 4 diacylglycerylhydroxymethyl-N,N,N-trimethylalanines, plus 2 phosphatidylethanolamines), 2 lycopenes, and 6 H20 molecules.
The LH-(B)Chl a content consisting of 6 LH-Chls a and 16 LH-BChls a provides the Cab. themophilum RC-PS with the ability to harvest both visible and far-red light, giving rise to an excitonically coupled “hybrid” LH system [4]. The boxes in Figure 8A demarcate pigment distribution in the cytoplasmic and periplasmic bilayer leaflets, with the PscA-2 cytoplasmic leaflet harboring one Chl a (913) and five Bchls a (902-905 and 907) (Figure 8B), while two Chls a (912 and 914) together with three BChls a (906, 908 and 909) are found in the corresponding periplasmic leaflet (Figure 6C). The Mg2+-to-Mg2+ distance of 15.0 Å between BChl a902 and BChl a909 represents the closest inter-bilayer connection and serves as a common energy transfer point for the distinct hybrid pigment arrays (Figure 6D)[4]. Based upon the Mg2+-to-Mg2+ distances between BChl a902 and the cytoplasmic array of BChls a ranging from 12.8-17.9 Å and 9.6 Å to Chl a913 (Figure 6B), a pathway of energy transfer was posited between these pigments that culminates at BChl a909 within the periplasmic layer (Figure 6D). Once energy reaches BChl a908, transfer occurs directly to P840, representing the PscA-1 half of the Zn-BChl a’ special pair, but it also expected to occur simultaneously from Chl a914 (Figure 6C), thereby completing this efficient excitation energy scheme within the bilayer leaflets.
Unlike PSII and the HB-Type I RC-PS, the (B)Chl molecules located in the positions of the PSII ChlZ-D1 and ChlZ-D2, serving as excitation donors to the RC, are absent from both the GSB and CAB RC-PS. This raises the possibility that their absence represented a driving force for the appearance of a peripheral FMO-chlorosome antenna. In addition to individual lycopene molecules associated with the LH-BChls a in each CAB PscA protomer (Figure 8A), a 22-kDa carotenoid-binding protein is also present in Cab. thermophilum [46]. Although absent from the RC-PS structure [4], the carotenoid-binding protein may serve as both a peripheral antenna, passing visible excitations onto LH-BChls, and in a photoprotective role exerted against reactive oxygen species, maintaining survival when exposed to high O2 levels in the microbial mat environment.
The membrane anchored PscX and PscY subunits of the CAB RC, which function in the serial donation of electrons to photooxidized P840, are positioned with edge-edge heme distances of 2.3 Å, while the PscY heme 2 is located at a distance of 10.4 Å from the Zn-BChl a P840 special pair [4]. Distinct from the GSB-Type I RC, in which the immediate RC-donor c-type cytochrome (cytochrome cZ) consists of a homodimer of the integral-membrane PscC-1 and PscC-2 protomers, with each providing two RC-associated TMHs (Figure 3C), PscX and PscY form a heterodimer associated with the RC-PS through both intramembrane regions and the periplasmic membrane surface. Associations within the membrane occur through hydrophobic interactions involving loop domains of PscX (residues 16-25) and TMH1 of PscA-1, and through PscY (residues 15-24) involving both TMH1 and TMH11 of Psc-A2. In addition, N-terminal Cys residues (PscX-C21 and PscY-C20) are subjected to posttranslational modifications that form linkages modeled as hydrocarbon chains (designated as ChainCys21 and ChainCys20) that assist in anchoring the cytochromes to the RC. At the peripheral membrane region, PscX and PscY are associated with PscA-1 and PscA-2 through numerous H-bonds linkages as well as salt bridges in which the Asp55 and Lys346 of each PscA protomer play crucial roles in spite of the asymmetry of the PscX-PscY heterodimer [4].
In contrast to the above associations existing between monoheme cytochromes c and homodimeric Type-1 RC, the RCs in many purple chlorophototrophic bacteria are directly reduced by tetraheme cytochromes c attached to the membrane by means of lipid anchors covalently linked to N-terminal Cys residues [9,10,11], which arise after removal of N-terminal signal sequences by the post-translational truncation process. These lipid anchors were first identified as a thioester-linked diacylglycerol molecule in Blastochloris viridis [50,51] and subsequently, a triacyl structure was found in Thermochromatium tepidum [52] consisting of an N-acyl chain and an S-diacylglycerol linked to the N-terminal Cys. However, the purple chlorophototrophic extremophile Rhodopila globiformis, capable of growth at ≥pH 3.0, lacks this Cys within the N-terminal region. Consequently, the unprocessed, full-length tetraheme cytochrome c is attached to the membrane through a membrane-spanning N-terminal ΤΜH [53]. Interestingly, this membrane anchoring helix shows a significant structural similarity with the N-terminal domain of the RC-associated PufX polypeptide, responsible for establishing the LH1 ring gap, facilitating ubiquinone exchange between the RC and the cytochrome bc1 complex of Cba. sphaeroides [54] and other Rba. and related species [9,10,11]. Comparative sequence analyses of the N-terminal regions of the Rpi. globiformis pufC gene (encoding the tetraheme c-type cytochrome) and the Cba. sphaeroides pufX gene, which share a common puf operon location immediately downstream from pufM encoding the RC-M subunit, demonstrated a phylogenetic relationship with the PufX protein as well as with pufC genes of related species forming tetraheme cytochromes in which N-terminal signal peptides are replaced with a lipid N-terminal membrane anchor [55]. Accordingly, a full-length tetraheme cytochrome, through partial deletions within an ancestral pufC gene, may represent the original archetypes for both the truncated cytochrome forms and the PufX protein [55]. It should also be noted that in the filamentous anoxygenic phototroph Roseiflexus castenholzii [56], attachment of the tetraheme c-type cytochrome component is also through a single N-terminal TMH anchor, albeit with additional associations at the periplasmic surface with both RC and LH proteins [57,58]. Importantly, Rfl. castenholzii, a green-nonsulfur bacterium lacking chlorosomes, is among the earliest anoxygenic phototrophs to emerge based upon a position in a deep eubacterial branch of the phylogenetic tree of 16S rRNA gene sequences [59].
Among the numerous genes encoding c-type cytochromes in the Cab. thermophilum genome [60], Cabther_A2183 and Cabther_A2184 were assigned to the respective PscX and PscY cytochrome c subunits, which correspond to the respective 189 and 221 amino acid (AA) gene products (GenBank Accession Nos. AEP129221.1 and AEP12922.1), exhibiting a ~30%. AA sequence identities (SI). This dicistronic operon is situated immediately downstream from the pscAB-fmo operon and was expected to be the source of the RC-associated cytochromes c [60], as ultimately confirmed in the RC-PS structure [4]. In addition, a second dicistronic operon (Cabther_A1370 and Cabther_A1371 was found whose respective gene products AEP12121.1, (158 AA, ~34% SI to PscY) and AEP1212.1 (119 AA, ~42% SI to PscY,) also form putative membrane-associated proteins. Other located genes include Cabther_A0280, which encodes a soluble 117 AA protein (AEP11047.1, ~30% SI to PscY), along with a protein annotated as the 190 AA subunit III of cytochrome c oxidase cbb3 ABV27349.1, 84% SI to PscX) in which a c-type heme CXXCH binding motif was found.
Further BLAST searches of the Cab. thermophilum Type B laboratory isolate genome revealed an apparent c-type cytochrome ortholog with Accession No. 7VZR_c (145 AA, 100% SI with PscX, while lacking the 15 N-terminal and the 29 C-terminal residues). It is not known whether this putative truncated protein undergoes specific N- and C-terminal processing, or if it represents a degradation product, and this puzzling sequence requires further investigation. Other identified cytochromes c included the putative monoheme c cytochrome subunit of a quinol-cytochrome c oxidase (AEP13240.1, 213 AA, ~32% SI to PscY). Also, a petAB operon is present in which a downstream gene (Cabther_A 0912) encodes a multifunctional cytochrome cyd-type quinol oxidase protein (AEP11669.1, 483 AA) containing a C-terminal c-type cytochrome region, apparently fulfilling the role of cytochrome c1 which is missing from the petAB operons. This cytochrome c region is attached to the N-terminal membrane cyd-integral domain by a 70-residue linker. Moreover, a second petAB operon is present that contains a petB gene whose multifunctional product contains a C-terminal cytochrome c domain with a similar unusual membrane linkage.
The possibility has been proposed that these multiple oxidoreductases, together with an Alternate Complex III, function in accommodating the wide-ranging environmental conditions existing during the diel cycle in the microbial mat community [60]. These include extensive changes in O2 availability ranging from O2 supersaturation under intense sunlight to the anoxic conditions that prevail during the late afternoon and evenings. Clearly, as a homodimeric Type I-RC containing aerobic anoxygenic phototroph, Cab. thermophilum clearly provides a unique system for obtaining a more detailed picture of the evolution of oxygenic phototrophs.
3. Further Insights into the Evolutionary Origins of Oxygenic Photosynthesis Provided by Structures of Homodimeric Type I Reaction Center-Photosystems
A variety of structural and functional features of the Type I-homodimeric RC-PSs of the HB, GSB and CAB have provided valuable insights into the evolutionary origins of both the PSI and PSII heterodimeric RCs of higher phototrophs. The major relevant discoveries as discussed recently by Gisriel et al. [61] include: (i) structural similarities between the Ca2+-binding site of the HB RC and the Mn4CaO5 cluster and TyrZ regions of PS II; (ii) the presence of a PG molecule involved in the ligation of the AO cofactor of the GSB electron transfer chain, associated with conserved Arg residues in both the HB and GSB RCs which has possible implications for the development of Q exchange and reduction mechanisms; (iii) the similarities in the arrangement of antenna (B)Chls within the PS II and GSB RC-PS; and (iv) the presence of a GSB excitation energy quenching mechanism promoted by a glycosylated carotenoid which has implications for the role played by O2 tolerance processes in supporting the evolutionary origins of oxygenic photosynthesis. These findings hold promise for guiding further investigations into both the evolutionary diversity of the RC and the relationships that led to the emergence H2O oxidation in higher phototrophs, largely responsible for the origin of an O2-containing atmosphere that facilitated the subsequent evolution of further life on earth.
Structural similarities between the sites on the homodimeric HB RC and the PSII Mn4CaO5 cluster, together with the respective TyrZ and TyrD donor sites of the D1 and D2 polypeptides of PSII, are shown in Figure 9A,B. It is seen that the recently recognized Ca2+ binding sites of (PshA)2 [61] and (PscA)2 of the GSB [2,3] and Cab [4] RCs overlap (Figure 9C) and are located in structurally homologous positions to the Mn4CaO5 cluster of the D1 RC polypeptide. In addition, the TyrZ donor site of D1 (Tyr161) is H-bonded to His190, that has a counterpart in TyrD of the D2 polypeptide, where an H-bond exists between Tyr160 and His189 (Figure 9A), considered as evidence for a homodimeric origin of the two PSII regions [6]. This scenario holds that H2O oxidation was initiated in two symmetrically placed catalytic cluster sites on each side of the RC (see below) as supported by ligands found in both the D1 and D2 polypeptides and their respective CP43 and CP47 antennae, and ultimately culminating in the assembly of the active Mn4CaO5 cluster on the D1 side.
Parallels are shown in Figure 9D between the position of the PshA Ca2+-binding site and the D1 Mn4CaO5 cluster in which Asp468 in the former is seen coordinated to the Ca2+ atom located in a region comparable to the D1 Tyr-His redox pair and adjacent to the D1 Mn4CaO5 cluster. Similarities in locations are also observed between the C-termini of the PshA-1 and D1 polypeptides, where the PshA1 C-terminal Val608 residue is liganded to the Ca2+ atom (Figure 9E,F), while for D1, the Ca2+ atom within the Mn4CaO5 cluster is liganded to the C-terminal Ala344 (Figure 9G). Furthermore, N263 positioned within the peripheral domain of the PshA-1 antenna region between the 5th and 6th TMH (Figure 9F), is associated with the Ca2+-binding site via two H2O molecules. Likewise, E354 is also located between the 5th and 6th TMH of the CP43 peripheral domain and is linked to the D1 Mn4CaO5 cluster (Figure 9G). It is also noteworthy that PSI lacks a Ca2+ binding site (Figure 9E), as the C-terminal region of helix 11 of PsaB is two turns greater in length than in the PshA counterpart. While these structural comparisons have contributed to our understanding of the origins of the H2O oxidation process (see below), they do not explain the origin of the Pheo-Q Type II electron transfer chain of PSII, which has instead been suggested to have occurred after the heterodimerization process [12], as previously discussed in section 2.1. It is therefore likely that PSII arose from a chimera of an H2O-oxidizing complex and the RC-electron transfer chain [62]. Such an ancestor would have served in the origin of extant RCs that evolved into Type I RCs of the oxygenic phototrophs by incorporating more pigments, and into Type II RCs by losing the Fe-S cluster which was replaced by quinone as the terminal electron acceptor.
Cardona and Rutherford [6] additionally posited that the increased redox potentials (“oxidative jumps”) attained on the donor and acceptor sides leading to functional PSII electron transfer and H2O oxidation were achieved by means of crucial mutations which occurred in a common homodimeric ancestor resulting in the pronounced structural changes responsible for simultaneous oxidative jumps. Following these large structural adaptations, a Tyr residue adjacent to the Ca2+ binding site, located in the position near the D1 YZ donor (Figure 9F,G), was able to generate a tyrosyl radical capable of oxidizing a number of aqueous Mn2+ cations. The bound Ca2+ together with the C-terminal carboxyl group and a ligand formed with the antenna domain, already existing in the HB RC-PS structure (Figure 9F), stabilized the oxidized Mn. Subsequently, protons released from the bound H2O molecules by the Mn oxidation process promoted μ-oxo-bridge formation between adjoining Mn cations as reflected in the assembly factor-free photoactivation process realized in the case of the PSII Mn4CaO5 cluster [61]. Moreover, the ability to oxidize H2O is thought to have occurred prior to the emergence of both PSI and the Type I homodimeric RCs, which required the presence of a system for the avoidance of reactive oxygen species [6].
It is important to note that for the GSB, each PscA protomer contains 12 LH BChls situated in positions conserved in the HB Type I photosystem, as is also largely seen for the LH Chls found in both PSI and PSII (Figure 4). This is consistent with the possibility that these GSB antenna BChls occupy sites found in the ancestral RC-PS complex. The HB complex possesses an additional 13 LH BChl g molecules associated with both PshA protomers, compensating for the lack of peripheral antennae, as exhibited by the presence of chlorosomes and FMO in the GSB. Two of the additional BChls g in the HB complex are located closest to electron transfer chain (Figure 4C); the absence of BChls in these sites accounts in part for the poor coupling of the bulk GSB antenna to the RC. Notably, the shared features of the antennae of the GSB core PS and PSII [61] imply an early origin for the water oxidation process. Moreover, the fact that the GSB RC-PS possesses features found in both Types I and II RCs likely reflects an origin for the GSB complex from a common homodimeric RC-PS ancestor.
The GSB RC-PS associated chlorobactene carotenoid designated as F26, bound in a position adjacent to the core complex interface with FMO [3], was shown to be involved in both energy quenching and transfer processes [63]. Accordingly, a slowing of fluorescence quenching of the core LH BChls was demonstrated in a Cba. tepidum mutant unable to glycosylate carotenoids, despite no apparent effects on photosynthetic apparatus structure. In addition to inhabiting anaerobic environments, Cba. tepidum is microaerophilic, which is supported by the ability of the glycosylated carotenoid components to facilitate tolerance to low O2 levels. An additional glycosylated carotenoid glycoside, F39 (hydroxy-γ- chlorobactene glucoside laurate), specific to the core complex, is located in a region that is structurally conserved in PSII and binds both carotenoids and lipids, while also participating in the quenching of Chl excitation energy. Importantly, since the quenching of excitation energy by F39 has provided a basis for O2-tolerance in GSB, the structural conservation found between regions inhabiting GSB F39 and PSII has suggested that the development of such energy quenching process served as a potent driving force in the very early stages RC evolution [61].
Overall, these important findings point to the origins of an O2 evolving complex, as well as a functional PSII early in the history of the evolution of life on earth, and further suggest that both current RC types arose from a primordial ancestor that already contained Types I and II RCs [6]. Moreover, Cyanobacteria likely diverged early and prior to extant phyla which include anoxygenic phototrophs [64,65]. Consequently, anoxygenic Type II RCs may have evolved independently subsequent to their oxygenic counterparts, with the Type I/Type II split occurring prior to the diversification of anoxygenic phototrophic bacteria [66]. This runs counter to the earlier, but largely unsupported views that anoxygenic Type II RCs are more ancient than the Type II RCs of their oxygenic counterparts [67] and that the anoxygenic to oxygenic photosynthetic transition occurred when a PSI containing cyanobacterium formed PSII with the ability to oxidize H2O (summarized by Olson and Blankenship [68]). Instead, the early origins of both PSI and II are now supported by a considerable body of evidence [6,12,13,61,62,63,64,65,66,69,70,71] forming the basis for the proposal that the gene duplication event, resulting in the heterodimerization of a homo-dimeric ancestor of PSII into distinct D1 and D2 polypeptides likely predated the subsequent convergent evolution of the well-studied L and M polypeptides of anoxygenic Type II RC [69,71].
References
- Gisriel, C.; Sarrou, I.; Ferlez, B.; Golbeck, J.H.; Redding, K.E.; Fromme, R. Structure of a Symmetric Photosynthetic Reaction Center-Photosystem, Science 2017, 357, 1021–1025. [CrossRef]
- Xie, H.; Lyratzakis, A.; Khera, R.; Koutantou, M.; Welsch, S.; Michel, H.; Tsiotis, G. Cryo-EM Structure of the Whole Photosynthetic Reaction Center Apparatus from the Green Sulfur Bacterium Chlorobaculum tepidum. Proc. Natl. Acad. Sci. USA 2023, 120, e2216734120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Wu, H.; Xu, C.; Liu, X.-C.; Huang, Z.; Chang, S.; Wang, W.; Han, G.; Kuang, T.; Shen, J.-R.; et al. Architecture of the Photosynthetic Complex from a Green Sulfur Bacterium. Science 2020, 370, eabb6350. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Huang, G.; Wang, C.; Wang, J.; Sui, S.-F.; Qin, X. Structure of the Acidobacteria Homodimeric Reaction Center Bound with Cytochrome c. Nat. Commun. 2022, 13, 7745. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis, 3rd ed.; John Wiley & Sons: Hoboken, NJ USA, 2021; p. 352p. ISBN 978-1-119-80001-9. [Google Scholar]
- Cardona, T.; Rutherford, A.W. Evolution of Photochemical Reaction Centres: More Twists. Trends Plant Sci. 2019, 24, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-Dimensional Structure of Cyanobacterial Photosystem I at 2.5 Å Resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, A.T.; Nguyen-Phan, T.C.; Cogdell, R.J. A Comparative Look at Structural Variation Among RC-LH1 'Core' Complexes Present in Anoxygenic Phototrophic Bacteria. Photosynth. Res. 2020, 145, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Niederman, R.A. The Structural Diversity of Bacterial Reaction Center-Light Harvesting 1 Complexes and Their Role in Developing Biohybrid Photoelectrochemical Cells. In Photosynthesis: From Plants to Nanomaterials, Hou, H.J.M.; Allakhverdiev, S.I. Eds, Ed.; .Elsevier: Amsterdam, NL, 2023; Chapter 12; pp. 239–291. [Google Scholar] [CrossRef]
- Swainsbury, D.J.K.; Qian, P.; Hitchcock, A.; Hunter, C.N. The Structure and Assembly of Reaction Centre-Light-Harvesting 1 Complexes in Photosynthetic Bacteria. Biosci. Rep. 2023, 43, BSR20220089. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Bracun, L.; Li, M. Structural Diversity and Modularity of Photosynthetic RC-LH1 Complexes. Trends Microbiol. 2023; 26, S0966-842X(23)00173-7. [Google Scholar] [CrossRef]
- Orf, G.S.; Gisriel, C.J.; Redding, K.E. Evolution of Photosynthetic Reaction Centers: Insights from the Structure of the Heliobacterial Reaction Center. Photosynth. Res. 2018, 138, 11–37. [Google Scholar] [CrossRef]
- Cardona, T. Early Archean Origin of Heterodimeric Photosystem I. Heliyon 2018, 4, e00548. [Google Scholar] [CrossRef]
- Deisenhofer, J.; Epp, O.; Miki, K.; Huber, R.; Michel, H. Structure of the Protein Subunits in the Photosynthetic Reaction Centre of Rhodopseudomonas viridis at 3Å Resolution. Nature 1985, 318, 618–624. [Google Scholar] [CrossRef]
- Allen, J.P.; Feher, G.; Yeates, T.O.; Komiya, H.; Rees, D.C. Structure of the Reaction Center from Rhodobacter sphaeroides R-26: The Protein Subunits. Proc. Natl. Acad. Sci. USA 1987, 84, 6162–6166. [Google Scholar] [CrossRef] [PubMed]
- Umena, Y.; Kawakami, K.; Shen, J.-R.; Kamiya, N. Crystal Structure of Oxygen-Evolving Photosystem II at a Resolution of 1.9 Å. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Kimble, L.K.; Mandelco, L.; Woese, C.R.; Madigan, M.T. Heliobacterium modesticaldum, sp. nov., a Thermophilic Heliobacterium of Hot Springs and Volcanic Soils. Arch. Microbiol. 1983, 136, 17–19. [Google Scholar] [CrossRef]
- Brockmann, H.; Lipinski, A. Bacteriochlorophyll g. A New Bacteriochlorophyll from Heliobacterium chlorum. Arch. Microbiol. 1983, 136, 17–19. [Google Scholar] [CrossRef]
- Orf, G.S.; Gisriel, C.J.; Granstrom, J.; Baker, P.L.; Redding, K.E. The PshX Subunit of the Photochemical Reaction Center from Heliobacterium modesticaldum Acts as a Low-Energy Antenna. Photosynth. Res. 2022, 151, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Gorka, M.; Charles, P.; Kalendra, V.; Baldansuren, A.; Lakshmi, K.V.; Golbeck, J.H. A Dimeric Chlorophyll Electron Acceptor Differentiates Type I from Type II Photosynthetic Reaction Centers. iScience 2021, 24, 102719. [Google Scholar] [CrossRef]
- Kashey, T.S.; Luu, D.D.; Cowgill, J.C.; Baker, P.L.; Redding, K.E. Light-Driven Quinone Reduction in Heliobacterial Membranes. Photosynth. Res. 2018, 138, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sarrou, I.; Khan, Z.; Cowgill, J.; Lin, S.; Brune, D.; Romberger, S.; Golbeck, J.H.; Redding, K.E. Purification of the Photosynthetic Reaction Center from Heliobacterium modesticaldum. Photosynth. Res. 2012, 111, 291–302. [Google Scholar] [CrossRef]
- Romberger, S.P.; Golbeck, J.H. The FX Iron–Sulfur Cluster Serves as the Terminal Bound Electron Acceptor in Heliobacterial Reaction Centers. Photosynth. Res. 2012, 111, 285–290. [Google Scholar] [CrossRef]
- Khadka, B.; Adeolu, M.; Blankenship, R.E.; Gupta, R.S. Novel Insights into the Origin and Diversification of Photosynthesis Based on Analyses of Conserved Indels in the Core Reaction Center Proteins. Photosynth. Res. 2017, 131, 159–171. [Google Scholar] [CrossRef]
- Sattley, W.M.; Madigan, M.T.; Swingley, W.D.; Cheung, P.C.; Clocksin, K.M.; Conrad, A.L.; Dejesa, L.C.; Honchak, B.M.; Jung, D.O.; Karbach, L.E.; et al. The Genome of Heliobacterium modesticaldum, a Phototrophic Representative of the Firmicutes Containing the Simplest Photosynthetic Apparatus. J. Bacteriol. 2008, 190, 4687–4696. [Google Scholar] [CrossRef]
- Psencík, J.; Ikonen, T.P.; Laurinmäki, P.; Merckel, M., C.; Butcher, S.J.; Serimaa, R., E.; Tuma, R. Lamellar Organization of Pigments in Chlorosomes, the Light Harvesting Complexes of Green Photosynthetic Bacteria. Biophys. J. 2004, 87, 1165–1172. [Google Scholar] [CrossRef]
- Francke, C.; Otte, S.C.M.; Miller, M.; Amesz, J.; Olson, J.M. Energy Transfer from Carotenoid and FMO-protein in Subcellular Preparations from Green Sulfur Bacteria. Spectroscopic Characterization of an FMO-Reaction Center Core Complex at Low Temperature. Photosynth. Res. 1996, 50, 71–77. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Niedzwiedzki, D.M.; Orf, G.S.; Zhang, H.; Blankenship, R.E. Dynamics of Energy and Electron Transfer in the FMO-Reaction Center Core Complex from the Phototrophic Green Sulfur Bacterium Chlorobaculum tepidum. J. Phys. Chem. B. 2015, 119, 8321–8329. [Google Scholar] [CrossRef] [PubMed]
- Magdaong, N.C.M.; Niedzwiedzki, D.M.; Saer, R.G.; Goodson, C.; Blankenship, R.E. Excitation Energy Transfer Kinetics and Efficiency in Phototrophic Green Sulfur Bacteria. Biochim. Biophys. Acta 2018, 1859, 1180–1190. [Google Scholar] [CrossRef]
- Dostál, J.; Pšenčík, J.; Zigmantas, D. In Situ Mapping of the Energy Flow Through the Entire Photosynthetic Apparatus. Nat. Chem. 2016, 8, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, M.; Itoh, S.; Kamei, S.; Matsubara, H.; Oh-oka, H. Time-Resolved Spectroscopy of Chlorophyll-a like Electron Acceptor in the Reaction Center Complex of the Green Sulfur Bacterium Chlorobium tepidum. Plant Cell Physiol., 1999, 40, 1021–1028. [CrossRef]
- Kjaer, B.; Frigaard, N.-U.; Yang, F.; Zybailov, B.; Miller, M.; Golbeck, J.H.; Scheller, H.V. Menaquinone-7 in the Reaction Center Complex of the Green Sulfur Bacterium Chlorobium vibrioforme Functions as the Electron Acceptor A1. Biochemistry 1998, 37, 3237–3242. [Google Scholar] [CrossRef]
- Van der Est, A.; Scheller, H.; Hager-Braun, C.; Hauska, G.; Stehlik, D. Forward Electron Transfer in Chlorobium Reaction Centres Studied by Transient EPR. In Photosynthesis: Mechanisms and Effects, Garab, G. Ed.; Springer: Berlin, DE, 1998; pp. 547–550. [Google Scholar]
- Fixen, K.R.; Oda, Y.; Harwood, C.S. Clades of Photosynthetic Bacteria Belonging to the Genus Rhodopseudomonas Show Marked Diversity in Light-Harvesting Antenna Complex Gene Composition and Expression. mSystems 2015, 1, e00006-15. [Google Scholar] [CrossRef]
- Southall, J.; Henry, S.L.; Gardiner, A.T.; Roszak, A.W.; Mullen, W.; Carey, A.M.; Kelly, S.M.; de Percin Northumberland, C.O.; Cogdell, R.J. Characterisation of a pucBA Deletion Mutant from Rhodopseudomonas palustris Lacking All But the pucBAd Genes. Photosynth. Res. 2018, 135, 9–21. [Google Scholar] [CrossRef]
- Ben-Shem, A.; Frolow, F.; Nelson, N. Crystal Structure of Plant Photosystem I. Nature 2003, 426, 630–635. [Google Scholar] [CrossRef]
- Qin, X.; Suga, M.; Kuang, T.; Shen, J.-R. Photosynthesis. Structural Basis for Energy Transfer Pathways in the Plant PSI-LHCI Supercomplex. Science 2015, 348, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Tsukatani, Y.; Azai, C.; Kondo, T.; Itoh, S.; Oh-Oka, H. Parallel Electron Donation Pathways to Cytochrome cZ in The Type I Homodimeric Photosynthetic Reaction Center Complex of Chlorobium tepidum. Biochim. Biophys. Acta 2008, 1777, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Oh-oka, H.; Kamei, S.; Matsubara, H.; Iwaki, M.; Itoh, S. Viscosity Dependence of the Electron Transfer Rate from Bound Cytochrome c to P840 in the Photosynthetic Reaction Center of the Green Sulfur Bacterium Chlorobium tepidum. Biochemistry 1997, 36, 9267–9272. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Higuchi, M.; Azai, C.; Oh-Oka, H.; Miki, K.; Wang, Z.-Y. Crystal Structure of the Electron Carrier Domain of the Reaction Center Cytochrome cZ Subunit from Green Photosynthetic Bacterium Chlorobium tepidum. J. Mol. Biol. 2010, 397, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Kölsch, A.; Madhi, H.; Stieger, K.R.; Feifel, S.C.; Kern, J.F.; Müh, F.; Lisdat, F.; Lokstein, H.; Zouni, A. Insights into the Binding Behavior of Native and Non-Native Cytochromes to Photosystem I from Thermosynechococcus elongatus. J. Biol. Chem. 2018, 293, 9090–9100. [Google Scholar] [CrossRef]
- Axelrod, H., L.; Abresch, E.C.; Okamura, M.Y.; Yeh, A.P.; Rees, D.C.; Feher, G. X-ray Structure Determination of the Cytochrome c2: Reaction Center Electron Transfer Complex from Rhodobacter sphaeroides. J. Mol. Biol. 2002, 319, 501–515. [Google Scholar] [CrossRef]
- Kubota-Kawai, H.; Mutoh, R.; Shinmura, K.; Sétif, P.; Nowaczyk, M.M.; Rögner, M.; Ikegami, T.; Tanaka, H.; Kurisu, G. X-ray Structure of an Asymmetrical Trimeric Ferredoxin-Photosystem I Complex. Nat. Plants 2018, 4, 218–224. [Google Scholar] [CrossRef]
- Tsukatani, Y.; Miyamoto, R.; Itoh, S.; Oh-Oka, H. Function of a PscD Subunit in a Homodimeric Reaction Center Complex of the Photosynthetic Green Sulfur Bacterium Chlorobium tepidum Studied by Insertional Gene Inactivation. Regulation of Energy Transfer and Ferredoxin-Mediated NADP+ Reduction on the Cytoplasmic Side. J. Biol. Chem. 2004, 279, 51122–51130. [Google Scholar] [CrossRef]
- Bryant, D.A.; Costas, A.; Maresca, J.; Chew, A.; Klatt, C.G.; Bateson, M.M.; Tallon, L.J.; Hostetler, J.; Nelson, W.C.; Heidelberg, J.F. et al.; et al. Candidatus Chloracidobacterium thermophilum: an Aerobic Phototrophic Acidobacterium. Science 2007, 317, 523–526. [Google Scholar] [CrossRef]
- Tsukatani, Y.; Romberger, S.P.; Golbeck, J.H.; Bryant, D.A. Isolation and Characterization of Homodimeric Type-I Reaction Center Complex from Candidatus Chloracidobacterium thermophilum, an Aerobic Chlorophototroph. J. Biol. Chem. 2012, 287, 5720–5732. [Google Scholar] [CrossRef]
- He, Z.; Ferlez, B.; Kurashov, V.; Tank, M.; Golbeck, J.H.; Bryant, D.A. Reaction Centers of the Thermophilic Microaerophile, Chloracidobacterium thermophilum (Acidobacteria) I: Biochemical and Biophysical Characterization. Photosynth. Res. 2019, 142, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Garcia Costas, A.M.; Tsukatani, Y.; Rijpstra, W.I.; Schouten, S.; Welander, P.V.; Summons, R.E.; Bryant, D.A. Identification of the Bacteriochlorophylls, Carotenoids, Quinones, Lipids, and Hopanoids of "Candidatus Chloracidobacterium thermophilum." J. Bacteriol. 2012 194,1158-1168. 194. [CrossRef]
- Tsukatani, Y.; Wen, J.; Blankenship, R.E.; Bryant, D.A. Characterization of the FMO Protein from the Aerobic Chlorophototroph, Candidatus Chloracidobacterium thermophilum. Photosynth. Res. 2010, 104, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Weyer, K.A.; Schaefer, W.; Lottspeich, F.; Michel, H. Cytochrome Subunit of the Photosynthetic Reaction Center from Rhodopseudomonas viridis is a Lipoprotein, Biochemistry 1987, 26, 2909–2914. 26. [CrossRef]
- Qian, P.; Siebert, C.A.; Wang, P.; Canniffe, D.P.; Hunter, C.N. Cryo-EM Structure of the Blastochloris viridis LH1-RC Complex at 2.9 Å. Nature 2018, 556, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.J.; Suga, M.; Wang-Otomo, Z.Y.; Shen, J.R. Structure of Photosynthetic LH1-RC Supercomplex At 1.9 A Resolution. Nature 2018, 556, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Tani, K.; Kanno, R.; Kurosawa, K.; Takaichi, S.; Nagashima, K.V.P.; Hall, M.; Yu, L.J.; Kimura, Y.; Madigan, M.T.; Mizoguchi, A.; et al. An LH1-RC Photocomplex from an Extremophilic Phototroph Provides Insight into Origins of Two Photosynthesis Proteins. Commun. Biol. 2022, 5, 1197. [Google Scholar] [CrossRef] [PubMed]
- Barz, W.P.; Vermeglio, A.; Francia, F.; Venturoli, G.; Melandri, B.A.; Oesterhelt, D. Role of the PufX Protein in Photosynthetic Growth of Rhodobacter sphaeroides. 2. PufX is Required for Efficient Ubiquinone/Ubiquinol Exchange Between the Reaction Center QB Site and the Cytochrome bc1 Complex. Biochemistry 1995, 34, 15248–15258. [Google Scholar] [CrossRef]
- Hucke, O.; Schiltz, E.; Drews, G.; Labahn, A. Sequence Analysis Reveals New Membrane Anchor of Reaction Center-Bound Cytochromes Possibly Related to PufX. FEBS Lett, 2003; 535, 166–170. [Google Scholar] [CrossRef]
- Hanada, S.; Takaichi, S.; Matsuura, K.; Nakamura, K. Roseiflexus castenholzii gen. nov., sp. nov., a Thermophilic, Filamentous, Photosynthetic Bacterium that Lacks Chlorosomes. Int. J. Syst. Evol. Microbiol. 2002, 52, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Shi, Y.; Niu, T.; Wang, Q.; Niu, W.; Huang, X.; Ding, W.; Yang, L.; Blankenship, R.E.; Xu, X. Cryo-EM Structure of the RC-LH Core Complex from an Early Branching Photosynthetic Prokaryote. Nature Commun. 2018, 9, 1568. [Google Scholar] [CrossRef]
- Qi, C.H.; Wang, G.L.; Wang, F.F.; Xin, Y.; Zou, M.J.; Madigan, M.T.; Wang-Otomo, Z.Y.; Ma, F.; Yu, L.J. New Insights on the Photocomplex of Roseiflexus castenholzii Revealed from Comparisons of Native and Carotenoid-Depleted Complexes. J. Biol. Chem. 2023, 299, 105057. [Google Scholar] [CrossRef]
- Woese, C.R. Bacterial Evolution. Microbiol. Rev. 1987, 51, 221–271. [Google Scholar] [CrossRef] [PubMed]
- Garcia Costas, A.M.; Liu, Z.; Tomsho, L.P.; Schuster, S.C.; Ward, D.M.; Bryant, D.A. Complete Genome of Candidatus Chloracidobacterium thermophilum, a Chlorophyll-Based Photoheterotroph Belonging to the Phylum Acidobacteria. Environ. Microbiol. 2012, 14, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Gisriel, C.J.; Azai, C.; Cardona, T. Recent Advances in the Structural Diversity of Reaction Centers. Photosynth. Res. 2021, 149, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T. Photosystem II is a Chimera of Reaction Centers. J. Mol. Evol. 2017, 84, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Azai, C.; Harada, J.; Fujimoto, S.; Masuda, S.; Kosumi, D. Anaerobic Energy Dissipation by Glycosylated Carotenoids in the Green Sulfur Bacterium Chlorobaculum tepidum. J. Photochem. Photobiol. A: Chemistry 2020, 403, 112828. [Google Scholar] [CrossRef]
- Cardona, T. A Fresh Look at the Evolution and Diversification of Photochemical Reaction Centers. Photosynth. Res. 2015, 126, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F.; Bryant, D.A.; Beatty, JT. A Physiological Perspective on the Origin and Evolution of Photosynthesis. FEMS Microbiol. Rev. 2018, 42, 205–231. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T. Thinking Twice About the Evolution of Photosynthesis. Open Biol. 2019, 9, 180246. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Bauer, C.E. Complex Evolution of Photosynthesis. Annu. Rev. Plant Biol. 2002, 53, 503–5321. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.M.; Blankenship, R.E. Thinking About the Evolution of Photosynthesis. Photosynth. Res. 2004;80, 373-386. [CrossRef]
- Cardona, T. Reconstructing the Origin of Oxygenic Photosynthesis: Do Assembly and Photoactivation Recapitulate Evolution? Front. Plant Sci. 2016, 7, 257. [Google Scholar] [CrossRef]
- Cardona, T.; Sánchez-Baracaldo, P.; Rutherford, A.W.; Larkum, A.W. Early Archean Origin of Photosystem II. Geobiol. 2019, 17, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Oliver, T.; Kim, T.D.; Trinugroho, J.P.; Cordón-Preciado, V.; Wijayatilake, N.; Bhatia, A.; Rutherford, A.W.; Cardona, T. The Evolution and Evolvability of Photosystem II. Annu. Rev. Plant Biol. 2023, 74, 225–257. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of the Type I homodimeric RC-PS complex of Hm. modesticaldum as determined by X-ray crystallography at 2.2 Å resolution [1]( Protein Data Bank (PDB) accession number: 5V8K). View parallel (A) and perpendicular (B) to membrane plane with later showing cytoplasmic surface: PshA, red, pink; PshX, orange; cofactors (electron transferring), teal; antenna, blue; carotenoids, lime; [4Fe-4S], red and yellow spheres. BChl and Chl tails are not shown. (C, D) Respective cytoplasmic views of TMH and cofactor arrangements. PshA helices are numbered 1-11, with transparent gray helices corresponding to heterodimeric PSI helical arrangement and with [4Fe-4S] FX component at center. The PS pigments in panel D are shown superimposed upon the gray PsaA-PsaB heterodimeric core-associated PSI cofactors. HB electron transferring BChls and Chls, brown; bulk antenna pigments, blue; BChl g molecules flanking electron transport chain, teal.
Figure 1.
Structure of the Type I homodimeric RC-PS complex of Hm. modesticaldum as determined by X-ray crystallography at 2.2 Å resolution [1]( Protein Data Bank (PDB) accession number: 5V8K). View parallel (A) and perpendicular (B) to membrane plane with later showing cytoplasmic surface: PshA, red, pink; PshX, orange; cofactors (electron transferring), teal; antenna, blue; carotenoids, lime; [4Fe-4S], red and yellow spheres. BChl and Chl tails are not shown. (C, D) Respective cytoplasmic views of TMH and cofactor arrangements. PshA helices are numbered 1-11, with transparent gray helices corresponding to heterodimeric PSI helical arrangement and with [4Fe-4S] FX component at center. The PS pigments in panel D are shown superimposed upon the gray PsaA-PsaB heterodimeric core-associated PSI cofactors. HB electron transferring BChls and Chls, brown; bulk antenna pigments, blue; BChl g molecules flanking electron transport chain, teal.
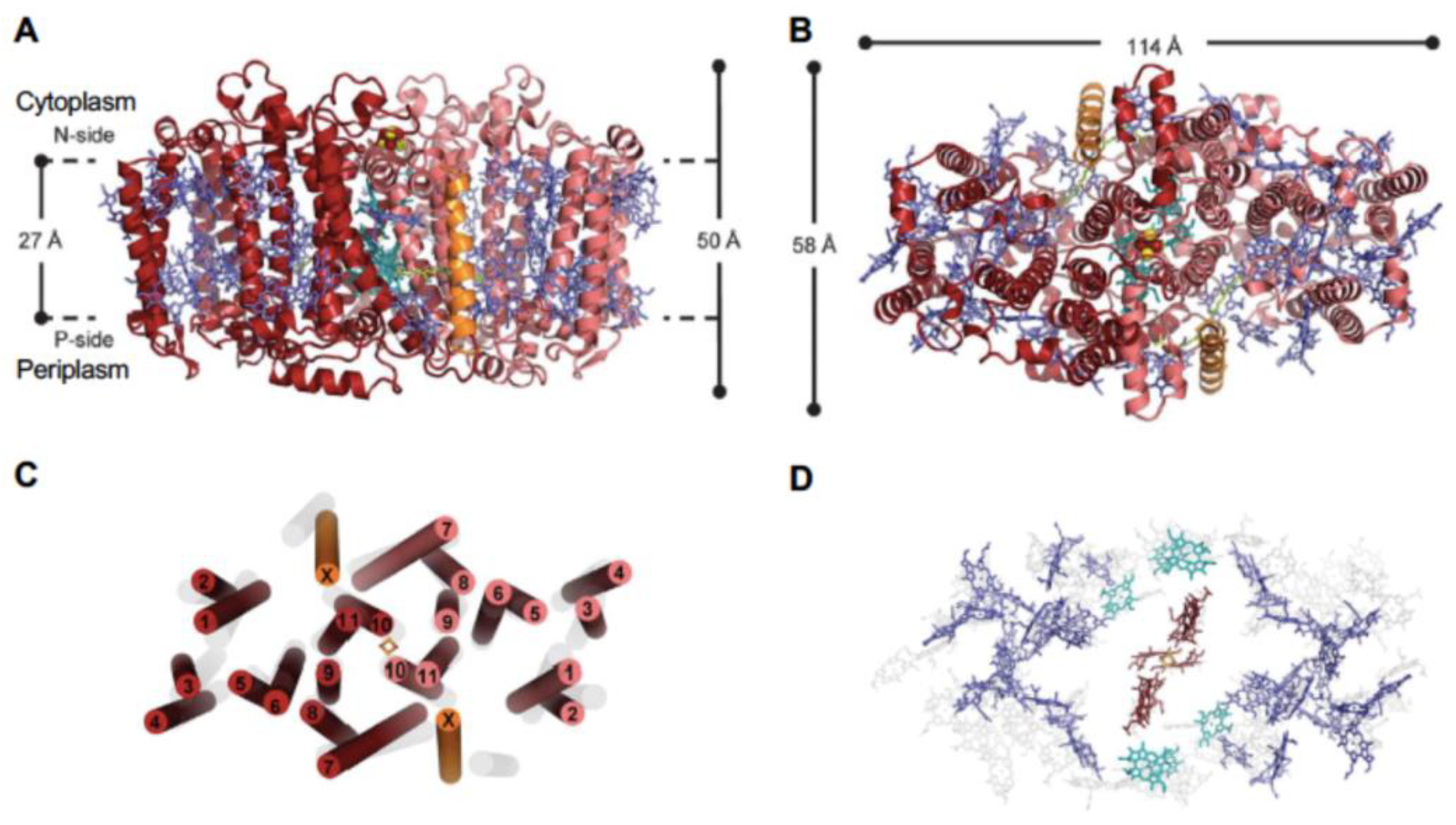
Figure 2.
Cofactor arrangement of the HB RC electron transfer chain and modeled menaquinone (MQ) molecule inserted into a potential quinone binding site [3]. (A) Cofactors showing coordinating residues provided by the homodimeric (PshA)2. Water molecules (small red balls) serve as axail ligands of the A0 primary acceptor, while the purple spheres which ligate the ACC accessory BChls are possible chloride ions and the green balls at the centers of the (B)chlorin rings represent magnesium atoms. BChl and Chl tails are not shown. Note that no carotenoid was found within the RC and only two are seen in the (PshA)2 complex, a reflection of the anaerobic environment in which Hm. modesticaldum is found which lacks reactive oxygen species. (B) Placement of MQ molecules in potential binding sites located with isoprenyl tails in positions similar to unassigned electron densities in the crystal structure [21]. (C) Unassigned electron density map in the vicinity of A0 of the HB RC. (D) Residues (light gray carbons) provided by PsaA coordinating the phylloquinone molecule of PSI (dark gray carbons) together with the residues of PshA (dark red carbons) forming an analogous binding site for MQ.
Figure 2.
Cofactor arrangement of the HB RC electron transfer chain and modeled menaquinone (MQ) molecule inserted into a potential quinone binding site [3]. (A) Cofactors showing coordinating residues provided by the homodimeric (PshA)2. Water molecules (small red balls) serve as axail ligands of the A0 primary acceptor, while the purple spheres which ligate the ACC accessory BChls are possible chloride ions and the green balls at the centers of the (B)chlorin rings represent magnesium atoms. BChl and Chl tails are not shown. Note that no carotenoid was found within the RC and only two are seen in the (PshA)2 complex, a reflection of the anaerobic environment in which Hm. modesticaldum is found which lacks reactive oxygen species. (B) Placement of MQ molecules in potential binding sites located with isoprenyl tails in positions similar to unassigned electron densities in the crystal structure [21]. (C) Unassigned electron density map in the vicinity of A0 of the HB RC. (D) Residues (light gray carbons) provided by PsaA coordinating the phylloquinone molecule of PSI (dark gray carbons) together with the residues of PshA (dark red carbons) forming an analogous binding site for MQ.
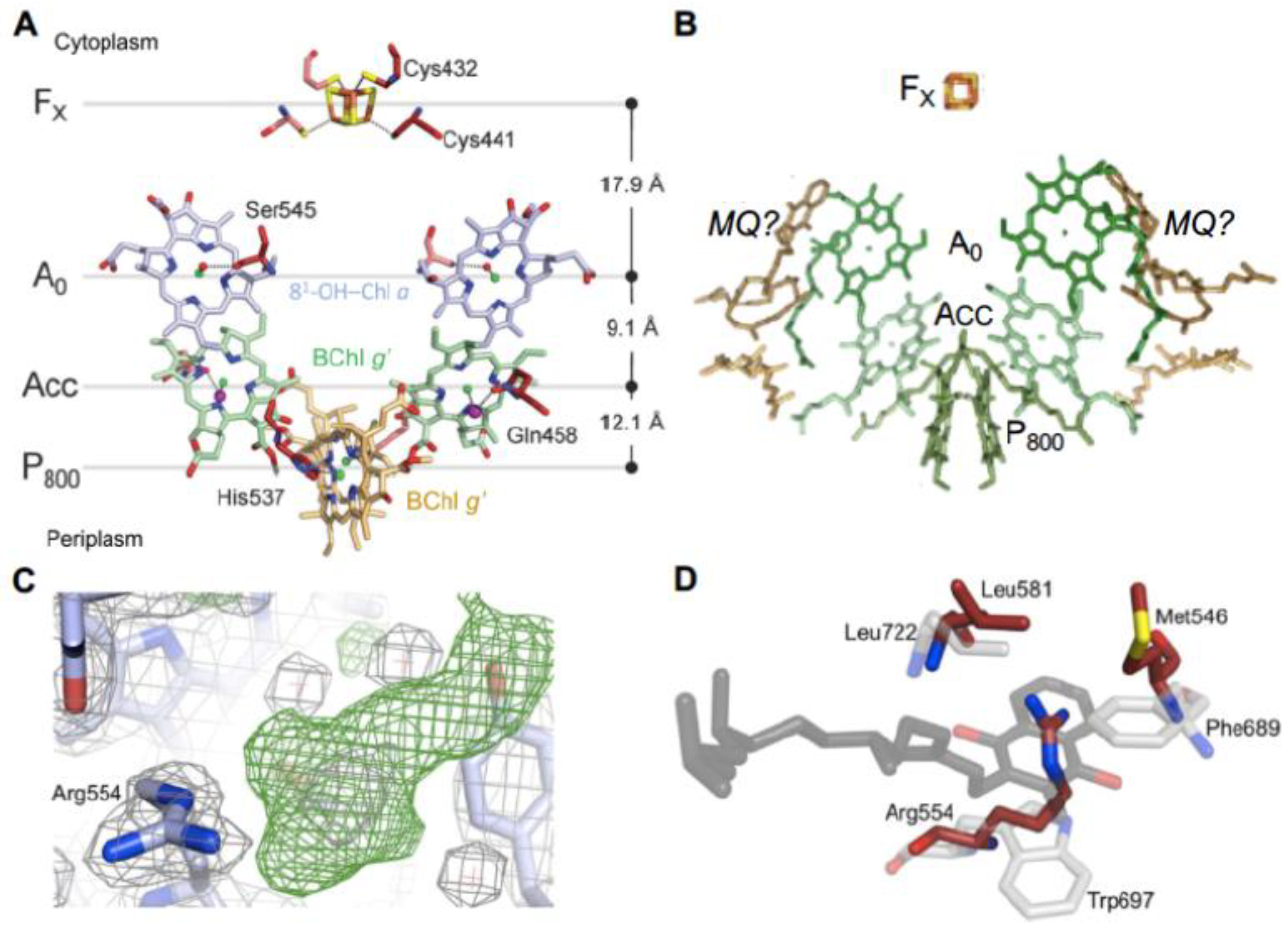
Figure 3.
Overall asymmetric structure of the complete Type I homodimeric RC-PS complex of Cba. tepidum with associated trimeric FMO, as determined by cryo-EM at 2.5 Å resolution [2] (PDB accession number: 7Z6Q). View seen parallel to membrane plane, showing C2 symmetry of PscA components (A) and perpendicular (B) to membrane plane, revealing the asymmetric arrangement FMO components at cytoplasmic surface: BChls, dark green; carotenoids, orange; lipids, black; [4Fe-4S], dark orange/yellow. The distinct FMO-1 to FMO-3 and FMO-4-to FMO-6 components forming the respective FMO trimers are shown. (C) Periplasmic surface view of arrangement of PscA and PscC helices, together with the RC-PS pigments. Helices are numbered starting from N-terminal position: (B)Chls, in stick representations, gray; central Mg atoms, pink. (D) Cofactor arrangement of the Cba. tepidum RC electron transfer chain [3]. Residues coordinating the (B)Chl cofactors are shown at right: A0, Chl a; ACC, Chl a; and P840 special pair, BChl a’; PscA-1 residues, gray; PscA-2 residues, pink.
Figure 3.
Overall asymmetric structure of the complete Type I homodimeric RC-PS complex of Cba. tepidum with associated trimeric FMO, as determined by cryo-EM at 2.5 Å resolution [2] (PDB accession number: 7Z6Q). View seen parallel to membrane plane, showing C2 symmetry of PscA components (A) and perpendicular (B) to membrane plane, revealing the asymmetric arrangement FMO components at cytoplasmic surface: BChls, dark green; carotenoids, orange; lipids, black; [4Fe-4S], dark orange/yellow. The distinct FMO-1 to FMO-3 and FMO-4-to FMO-6 components forming the respective FMO trimers are shown. (C) Periplasmic surface view of arrangement of PscA and PscC helices, together with the RC-PS pigments. Helices are numbered starting from N-terminal position: (B)Chls, in stick representations, gray; central Mg atoms, pink. (D) Cofactor arrangement of the Cba. tepidum RC electron transfer chain [3]. Residues coordinating the (B)Chl cofactors are shown at right: A0, Chl a; ACC, Chl a; and P840 special pair, BChl a’; PscA-1 residues, gray; PscA-2 residues, pink.
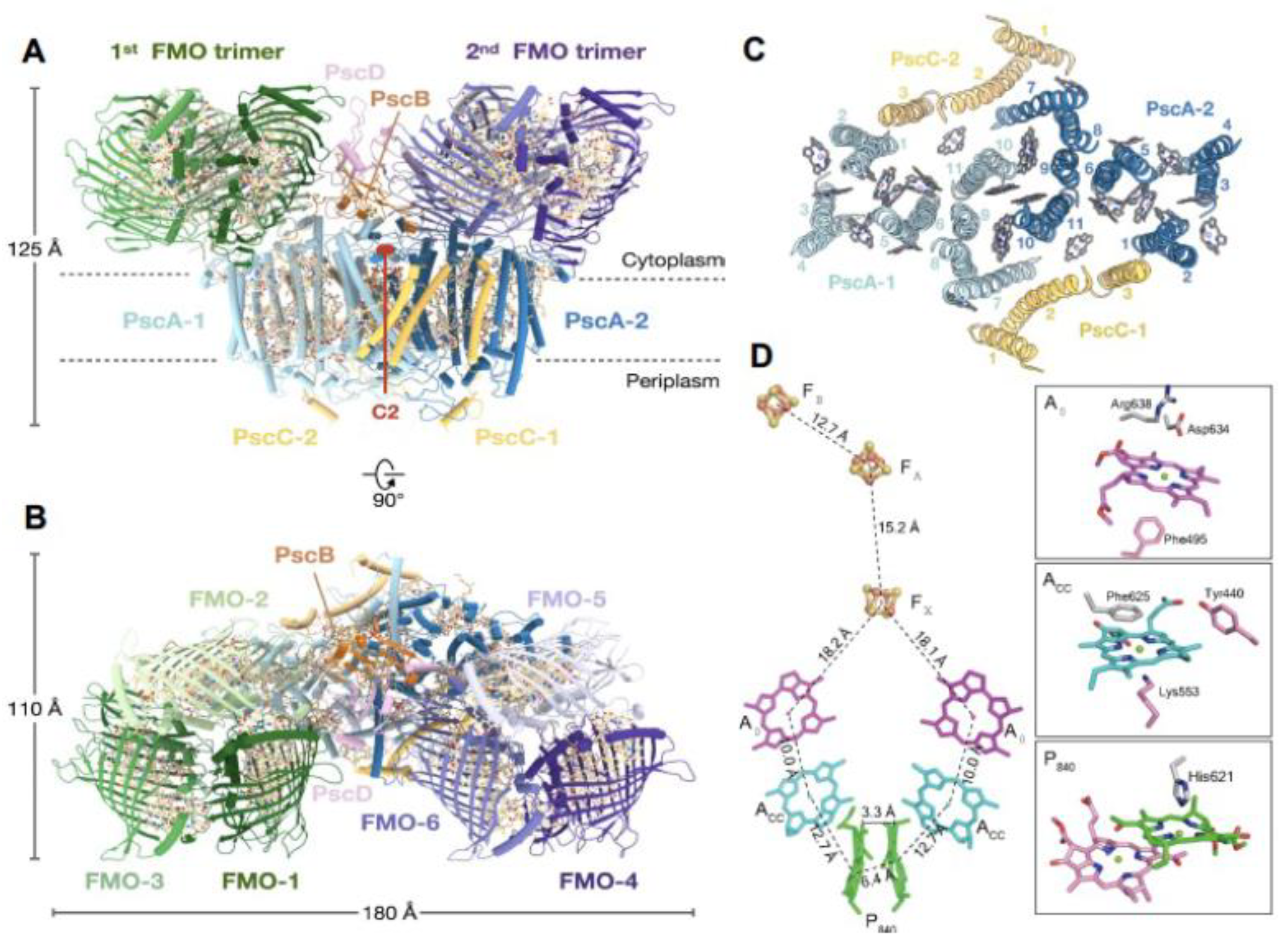
Figure 4.
The common arrangement of (B)Chls in Type I and II RC complexes. (A,B) Distributions and superpositioning of (B)Chls in the GSB RC-PS complex (dark green) and in PSII (yellow) shown perpendicular to membrane plane [3]. Dashed circles demarcate the clustering of antenna (B)Chls around the RC electron transferring cofactors. (C,D) Distributions and superpositioning of (B)Chls in GSB RC-PS (dark green) with those of the HB RC-PS (purple). and PSI (blue). Dashed ellipses demarcate the closed rings of (B)Chls distributed across the two RC-photosystem subunits. Note that the (B)Chl distribution in the GSB RC-PS more closely resembles that of PSII than PSI.
Figure 4.
The common arrangement of (B)Chls in Type I and II RC complexes. (A,B) Distributions and superpositioning of (B)Chls in the GSB RC-PS complex (dark green) and in PSII (yellow) shown perpendicular to membrane plane [3]. Dashed circles demarcate the clustering of antenna (B)Chls around the RC electron transferring cofactors. (C,D) Distributions and superpositioning of (B)Chls in GSB RC-PS (dark green) with those of the HB RC-PS (purple). and PSI (blue). Dashed ellipses demarcate the closed rings of (B)Chls distributed across the two RC-photosystem subunits. Note that the (B)Chl distribution in the GSB RC-PS more closely resembles that of PSII than PSI.
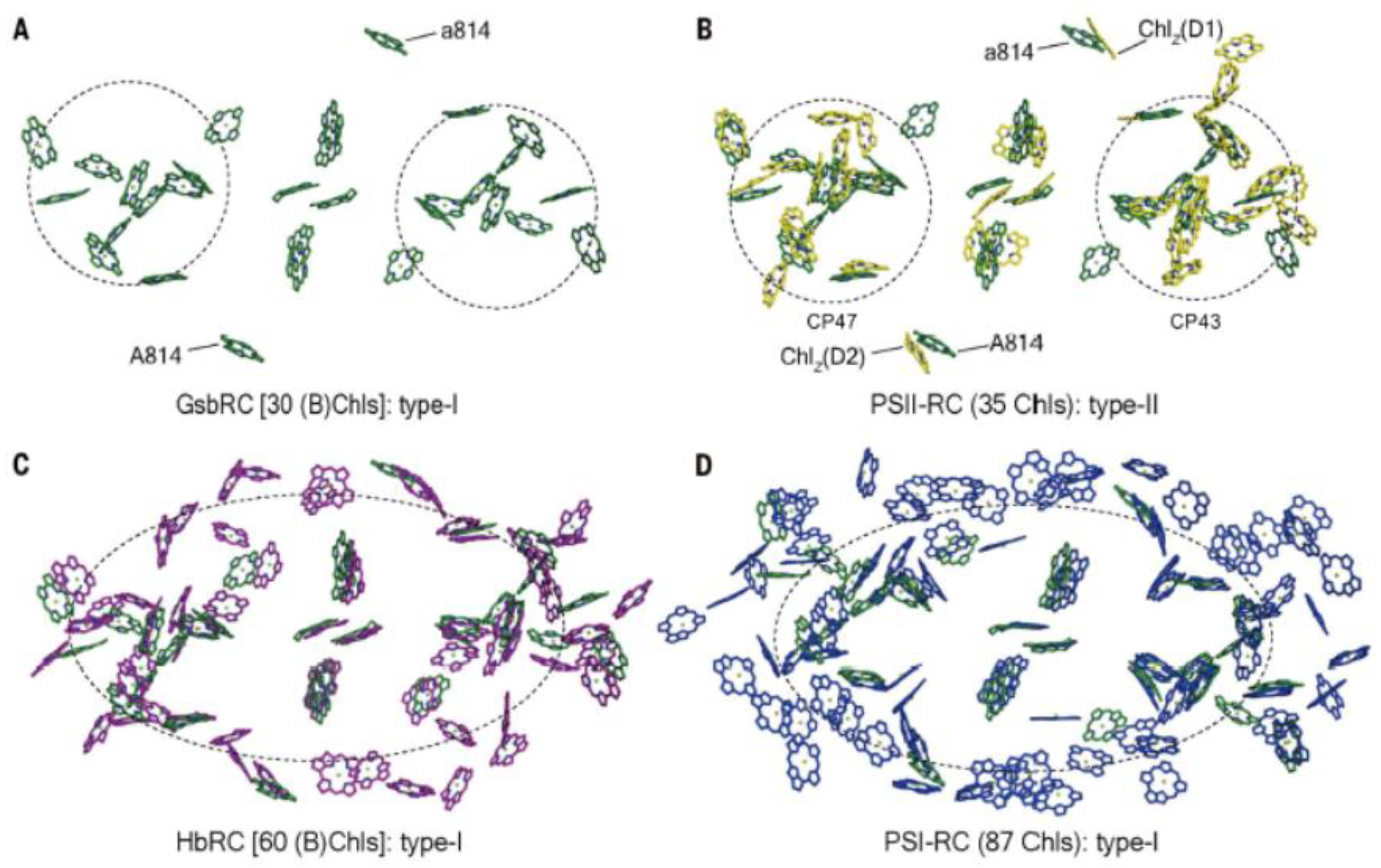
Figure 5.
Potential pathways of excitation energy transfer revealed by the structure of GSB FMO-RC-PS complex [2]. (A) Interface region between FMO-1 and PscA-1 protomers at cytoplasmic pigment surface. Inter-pigment distances (Å) represent BChl edge-to-edge distances. (B) Interface region between FMO-4 and PscA-2 protomers showing BChl edge-to-edge distances. (C) Potential pathways of energy transfer from nearby LH-BChls to P840 special pair located in periplasmic bilayer. BChl edge-to-edge distances are shown for PsaA-2-associated BChls, while their Mg-to-Mg distances are shown for their PsaA-1 associated counterparts. Phytyl tails of (B)Chls have been removed for clarity.
Figure 5.
Potential pathways of excitation energy transfer revealed by the structure of GSB FMO-RC-PS complex [2]. (A) Interface region between FMO-1 and PscA-1 protomers at cytoplasmic pigment surface. Inter-pigment distances (Å) represent BChl edge-to-edge distances. (B) Interface region between FMO-4 and PscA-2 protomers showing BChl edge-to-edge distances. (C) Potential pathways of energy transfer from nearby LH-BChls to P840 special pair located in periplasmic bilayer. BChl edge-to-edge distances are shown for PsaA-2-associated BChls, while their Mg-to-Mg distances are shown for their PsaA-1 associated counterparts. Phytyl tails of (B)Chls have been removed for clarity.
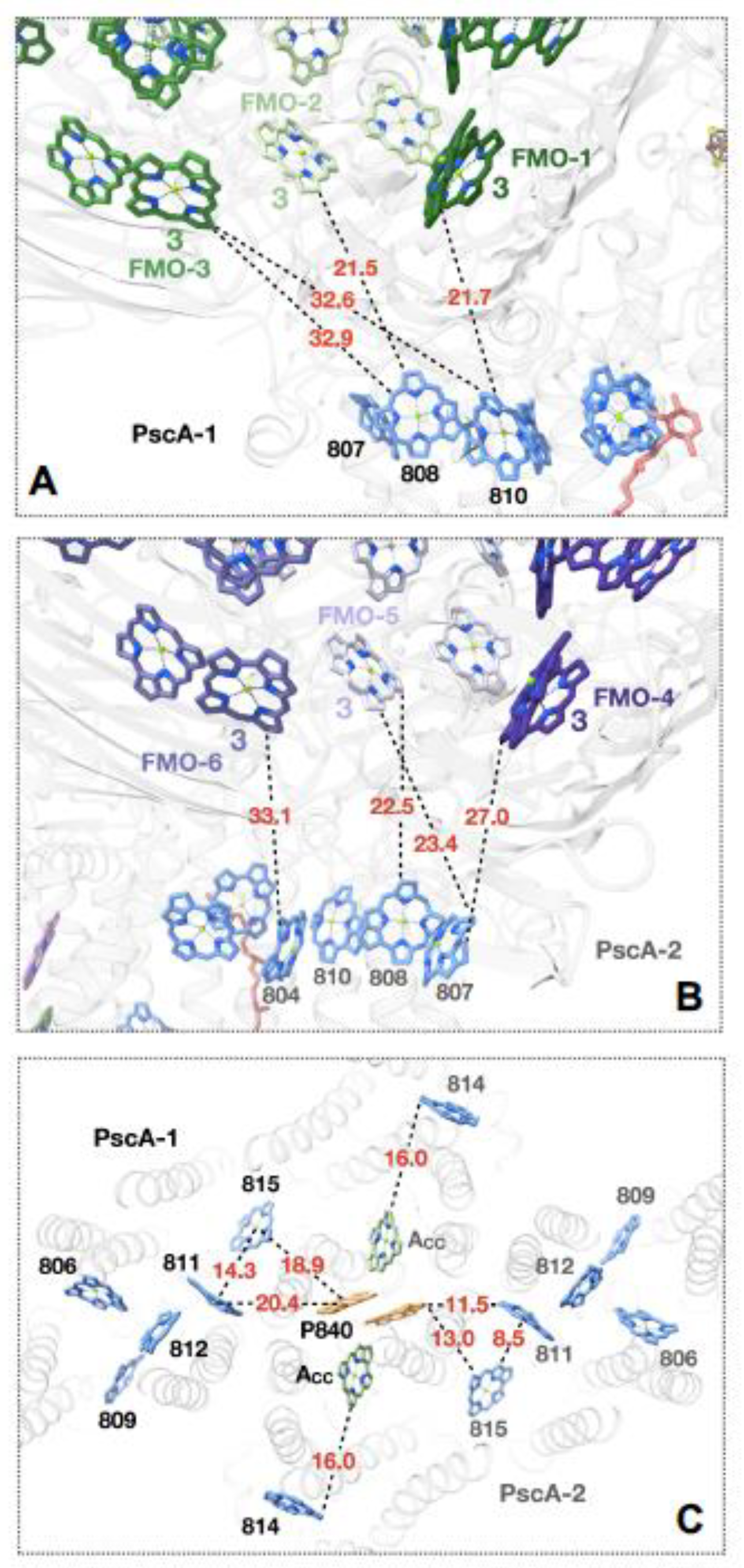
Figure 6.
(A). Potential Cba. tepidum cytochrome cZ binding site. This site was predicted from protein-protein docking analysis [2] using the crystal structure of C-terminal electron carrier domain [40] with enlarged view at right. Red dotted lines in right panel show respective distances from the heme Fe to the Mg ions of each of the P840 special pair BChls. At right, the distance between the heme-associated propionate group and the neighboring PscA Trp106 residues is also shown. (B). Potential ferredoxin-PscB binding site [5] shown in electrostatic surface representation of the Cba. tepidum FMO-RC-PS. Protein surface coloring represents electrostatic surface potential ( -10 kT, red; +10 kT, blue). An enlarged detailed view of putative ferredoxin-PscB docking site is seen at right along with a number of adjacent positively charged residues provided by both PscA-1 and PscB.
Figure 6.
(A). Potential Cba. tepidum cytochrome cZ binding site. This site was predicted from protein-protein docking analysis [2] using the crystal structure of C-terminal electron carrier domain [40] with enlarged view at right. Red dotted lines in right panel show respective distances from the heme Fe to the Mg ions of each of the P840 special pair BChls. At right, the distance between the heme-associated propionate group and the neighboring PscA Trp106 residues is also shown. (B). Potential ferredoxin-PscB binding site [5] shown in electrostatic surface representation of the Cba. tepidum FMO-RC-PS. Protein surface coloring represents electrostatic surface potential ( -10 kT, red; +10 kT, blue). An enlarged detailed view of putative ferredoxin-PscB docking site is seen at right along with a number of adjacent positively charged residues provided by both PscA-1 and PscB.
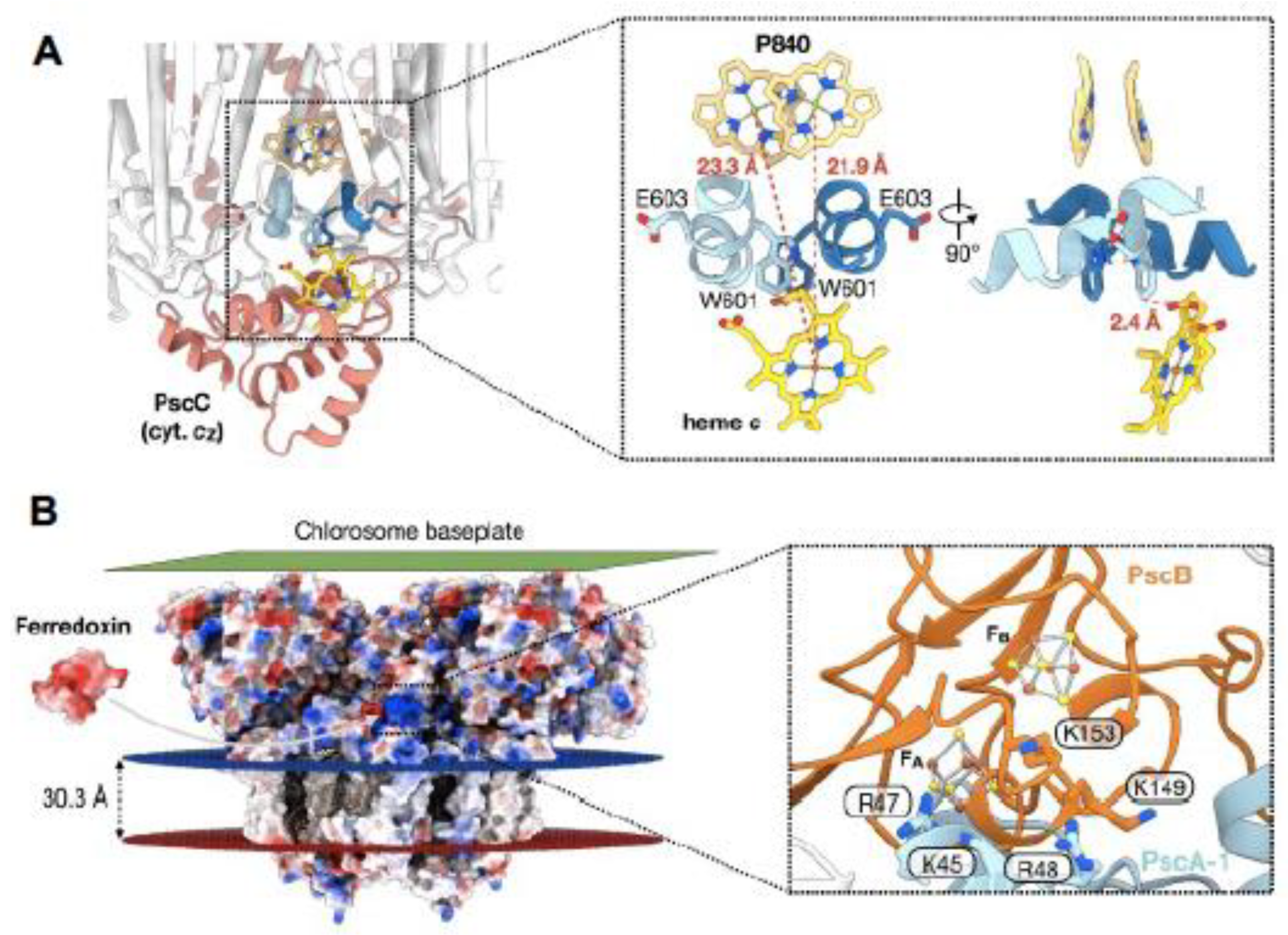
Figure 7.
Structure of the Type I homodimeric RC-PS complex of Cab. thermophilum as determined by cryo-EM at 2.6 Å resolution [5](PDB accession number: 7VZR). (A) View parallel to membrane plane. (B) Side view along membrane plane. Main tetrapyrroles of (B)Chls and Heme groups, green and hot pink, respectively; carotenoids, lipids and unidentified molecules are shown as line models in light blue, [4Fe4S] clusters FX, FA and FB, red and yellow spheres. While PscA has a conserved Type I PS arrangement of 11 TMHs, a unique extramembrane loop region protrudes between helices 7 and 8. (C) Cofactor arrangement of the Cab. themophilum RC electron transfer chain as viewed from membrane plain (left), periplasmic surface (center) and cytoplasmic surface (right). Center-to-center distances between cofactors are presented on the ordinates. PME, phosphatidyl-N-methylethanolamine.
Figure 7.
Structure of the Type I homodimeric RC-PS complex of Cab. thermophilum as determined by cryo-EM at 2.6 Å resolution [5](PDB accession number: 7VZR). (A) View parallel to membrane plane. (B) Side view along membrane plane. Main tetrapyrroles of (B)Chls and Heme groups, green and hot pink, respectively; carotenoids, lipids and unidentified molecules are shown as line models in light blue, [4Fe4S] clusters FX, FA and FB, red and yellow spheres. While PscA has a conserved Type I PS arrangement of 11 TMHs, a unique extramembrane loop region protrudes between helices 7 and 8. (C) Cofactor arrangement of the Cab. themophilum RC electron transfer chain as viewed from membrane plain (left), periplasmic surface (center) and cytoplasmic surface (right). Center-to-center distances between cofactors are presented on the ordinates. PME, phosphatidyl-N-methylethanolamine.
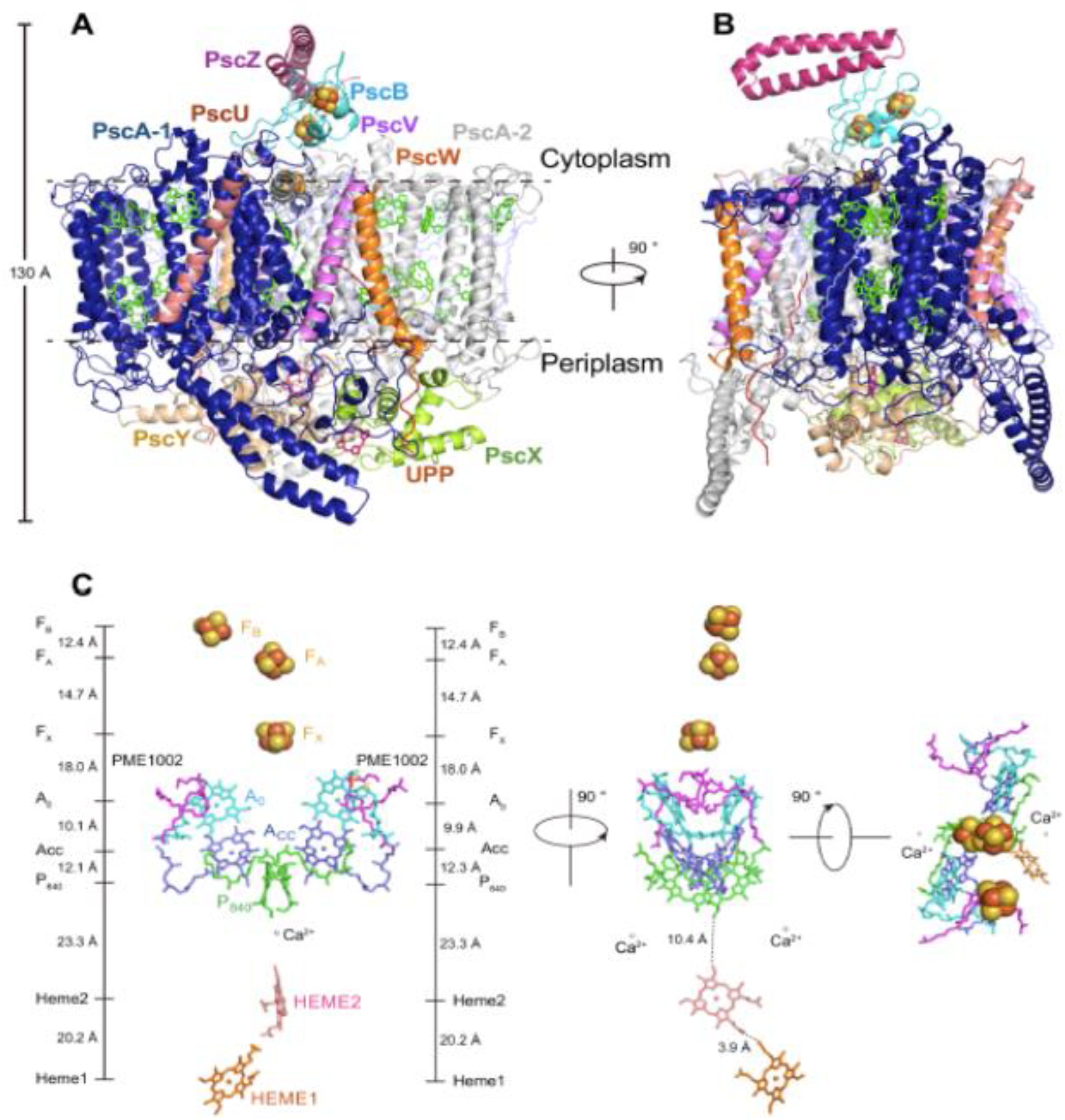
Figure 8.
Pigment distribution and proposed excitation energy transfer pathways in the Cab. thermophilus RC-PS [5]. (A) Layering of pigments in cytoplasmic and periplasmic membrane bilayer leaflets. Chl a, dark blue; BChl, green; lycopene, brown. (B) Mg2+ → Mg2+ distances (Å) for PsaA1 (B)Chls in cytoplasmic leaflet (C) Mg2+ → Mg2+ distances (Å) for PsaA1 (B)Chls in periplasmic leaflet.(D) Proposed pathways of PsaA1 antenna (B)Chl a → RC excitation energy transfer as denoted by the dotted red arrows.
Figure 8.
Pigment distribution and proposed excitation energy transfer pathways in the Cab. thermophilus RC-PS [5]. (A) Layering of pigments in cytoplasmic and periplasmic membrane bilayer leaflets. Chl a, dark blue; BChl, green; lycopene, brown. (B) Mg2+ → Mg2+ distances (Å) for PsaA1 (B)Chls in cytoplasmic leaflet (C) Mg2+ → Mg2+ distances (Å) for PsaA1 (B)Chls in periplasmic leaflet.(D) Proposed pathways of PsaA1 antenna (B)Chl a → RC excitation energy transfer as denoted by the dotted red arrows.
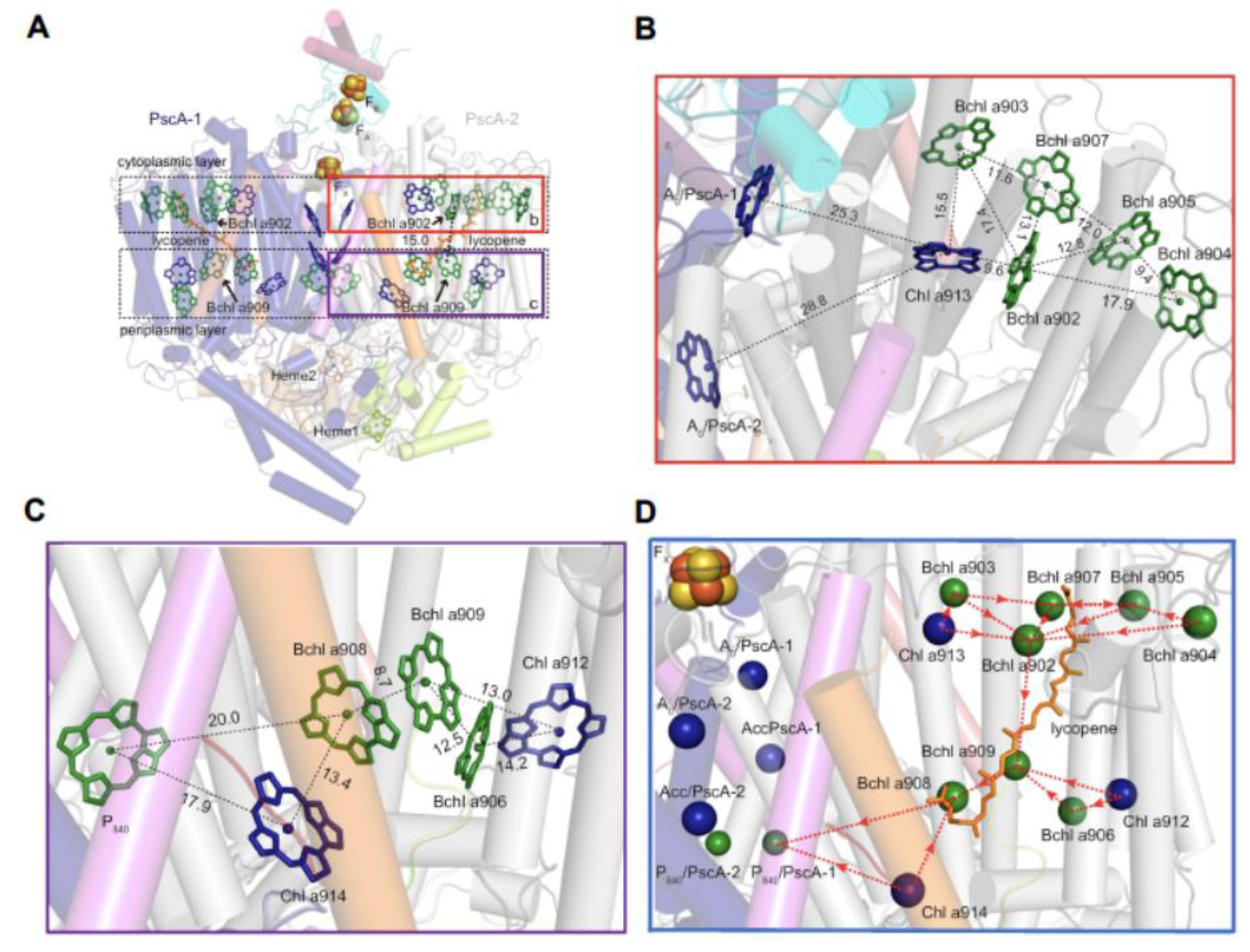
Figure 9.
Structural evidence supporting the role for the Ca2+-binding site of the HB RC as a possible evolutionary precursor of the PSII water-oxidizing Mn4CaO5 cluster region [6]. (A) PSII RC heterodimeric D1 (left) and D2 (right) RC core polypeptides (gray structures); CP43 (left) and CP47 (right) antenna proteins (orange structures); cofactors are shown as stick structures. Boxed regions show enhanced view of homology between the D1 YZ donor site and YD of the D2 polypeptide (B) The HB (PshA)2 homodimeric antenna (orange structures comprising α-helices 1-6) and RC core regions (gray structure comprising α-helices 7-11). Spheres in panels A and B represent Fe (orange), S (yellow), Mn (purple), O (red), Ca (green) atoms. (C) Superpositioning of homodimeric Type I RC Ca2+-binding sites within RC-PS structures [6]. (D) Enhanced view of conserved regions of TMH 3 and 4 of D1 polypeptide (orange) and helices 9 and 10 of the PshA RC domain (gray). (E) Enhanced view of matching regions of PsaB of cyanobacterial PSI RC (orange) and PshA RC (gray), demonstrating that helix 11 is two turns greater in length than PshA counterpart (as demarcated by the black and red arrowheads), as well as the lack of Ca2+ binding site in PSI. (F) Enhanced view of PshA Ca2+-binding site illustrating connection of ligand of N263 of the antenna domain of the PshA protein. (G) Enhanced view of the Mn4CaO5 cluster and YZ regions of PSII illustrating how the C-terminal residues E354 of CP43 and A344 of D1 provide direct ligands to the Ca2+ atom within the Mn4CaO5 cluster.
Figure 9.
Structural evidence supporting the role for the Ca2+-binding site of the HB RC as a possible evolutionary precursor of the PSII water-oxidizing Mn4CaO5 cluster region [6]. (A) PSII RC heterodimeric D1 (left) and D2 (right) RC core polypeptides (gray structures); CP43 (left) and CP47 (right) antenna proteins (orange structures); cofactors are shown as stick structures. Boxed regions show enhanced view of homology between the D1 YZ donor site and YD of the D2 polypeptide (B) The HB (PshA)2 homodimeric antenna (orange structures comprising α-helices 1-6) and RC core regions (gray structure comprising α-helices 7-11). Spheres in panels A and B represent Fe (orange), S (yellow), Mn (purple), O (red), Ca (green) atoms. (C) Superpositioning of homodimeric Type I RC Ca2+-binding sites within RC-PS structures [6]. (D) Enhanced view of conserved regions of TMH 3 and 4 of D1 polypeptide (orange) and helices 9 and 10 of the PshA RC domain (gray). (E) Enhanced view of matching regions of PsaB of cyanobacterial PSI RC (orange) and PshA RC (gray), demonstrating that helix 11 is two turns greater in length than PshA counterpart (as demarcated by the black and red arrowheads), as well as the lack of Ca2+ binding site in PSI. (F) Enhanced view of PshA Ca2+-binding site illustrating connection of ligand of N263 of the antenna domain of the PshA protein. (G) Enhanced view of the Mn4CaO5 cluster and YZ regions of PSII illustrating how the C-terminal residues E354 of CP43 and A344 of D1 provide direct ligands to the Ca2+ atom within the Mn4CaO5 cluster.
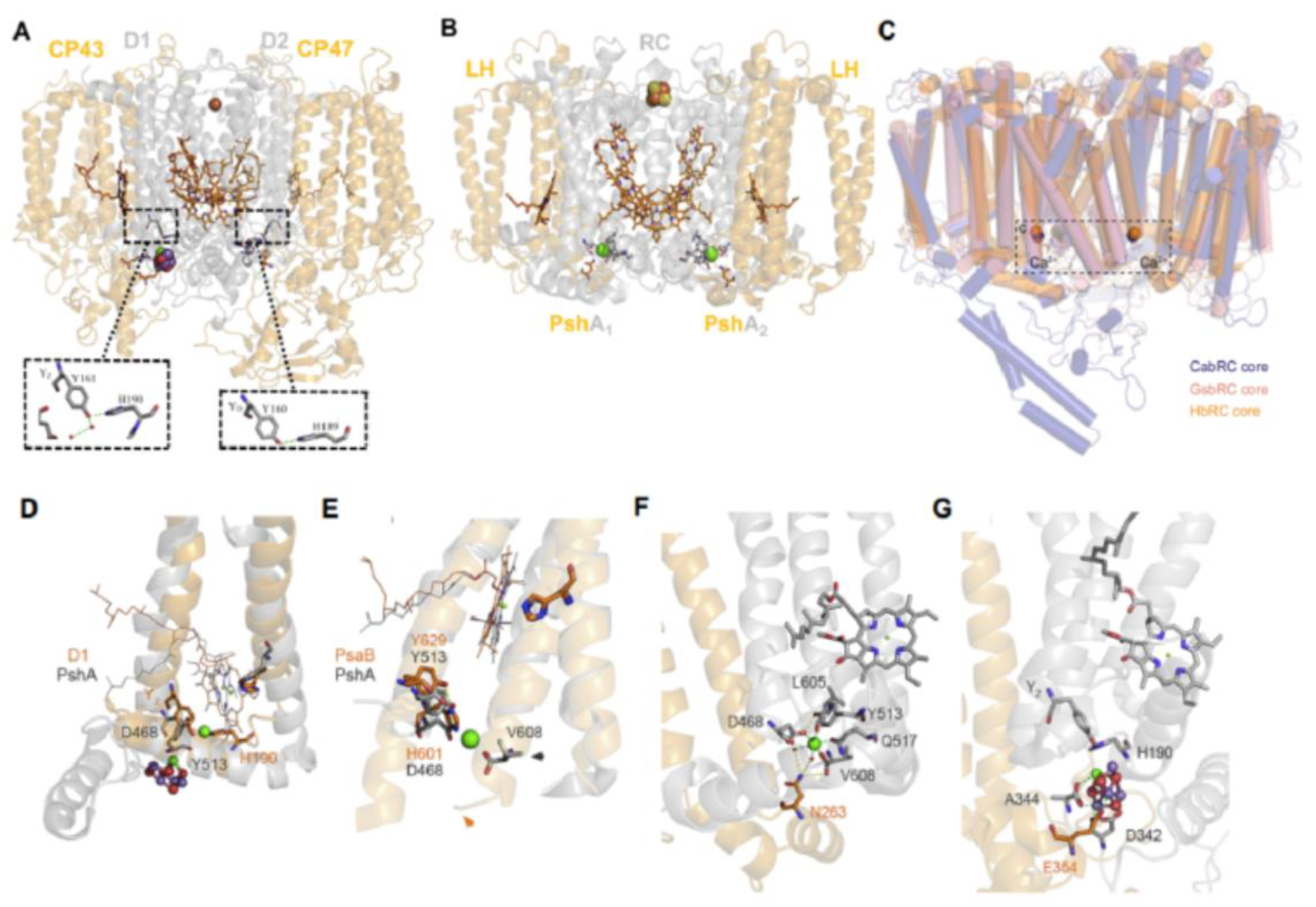

Table 1.
Resolved Subunits of the Cab. thermophilum RC-PS Structure [5].
Table 1.
Resolved Subunits of the Cab. thermophilum RC-PS Structure [5].
| Amino | |||
| Designation Acids | TMHs[2] | Functional assignment | |
| PscA-1 | 865 | 11 | Homodimeric RC-PS core, LH and RC charge separation roles |
| PscA-2 | 865 | 11 | Homodimeric RC-PS core, LH and RC charge separation roles |
| PscB | 179 | None | Houses terminal RC [4Fe-4S] electron acceptors FA and FB |
| PscU | 35 | 1 | Newly identified, unknown function |
| PscV | 36 | 1 | Newly identified, unknown function |
| PscW | 45 | 1 | Newly identified, unknown function |
| PscX | 189 | None | Cytochrome c electron donor 1 to RC, forms heterodimer with PscY |
| PscY | 221 | None | Cytochrome c electron donor 2 to RC, forms heterodimer with PscX |
| PscZ | 70 | None | Putative PscB stabilizer, may play PsaD role in enhancing PSI e- transfer |
| UPP | 19 | 1 | Unidentified polypeptide, non-helical |
1Transmembrane α-helical spans.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



