
Submitted:
08 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
The effect of Amaranthus cruentus L. seed oil (AmO) on collagen biosynthesis and wound healing was studied in cultured human dermal fibroblasts exposed to UVA radiation. It has been found that UVA radiation inhibited collagen biosynthesis, prolidase activity, expression of β1-integrin receptor, phosphorylated ERK1/2 and TGF-β, while increased the expression of p38 kinase. The AmO at 0.05-0.15% counteracted the above effects induced by UVA radiation in fibroblasts. UVA radiation also induced the expression and nuclear translocation of pro-inflammatory NF-κB fac-tor and enhanced the COX-2 expression. AmO effectively suppressed the expression of these pro-inflammatory factors induced by UVA radiation. Expressions of β1 integrin and IGF-I re-ceptors were decreased in the fibroblasts exposed to UVA radiation, while AmO counteracted the effects. Furthermore, AmO stimulated fibroblast’s migration in a wound healing model, thus facilitating the repair process following exposure of fibroblasts to UVA radiation. This data suggest the potential of AmO to counteract UVA-induced skin damage.
Keywords:
UVA radiation
; dermal fibroblasts
; sun-protective substance
; collagen biosynthesis
; wound healing
; Amaranthus cruentus L. seed oil
1. Introduction
Solar radiation is a significant factor contributing to the acceleration of skin aging through impairment of metabolism in the skin cells. Two mechanisms of this harmful effects are recognized, the first involves the production of reactive oxygen species (ROS), the second, UV radiation induced DNA damage. Both processes contribute to deregulation of metabolism in the skin cells, inhibition of collagen biosynthesis and induction of cell death through apoptosis [1,2]. As a result, collagen metabolism is impaired at transcriptional and post-transcriptional level. At transcriptional level UVA induces NF-kB expression, the inhibitor of collagen gene transcription [3]. Post-transcriptionally it impairs prolidase activity, the enzyme providing proline from imidodipeptides for collagen biosynthesis [4]) and activates matrix metalloproteinases (MMP) that degrade collagen extracellularly [5]. In addition, collagen also plays a critical role in regulating cellular metabolism as a ligand of integrin receptors. Integrin receptors participate in signaling that regulates collagen biosynthesis and prolidase activity [4]. Both processes are stimulated by the insulin-like growth factor-I (IGF-I), the most potent factor stimulating collagen biosynthesis [6]. Under physiological conditions, the activation of β1-integrin and IGF-I receptors initiate a cascade of mitogen-activated protein kinase (MAP-kinase) signaling pathway, including extracellular signal-regulated kinases (ERK-1 and ERK-2) [4]. It is well established that ROS down-regulate expression of MAP kinase ERK1/2 and support expression of stress-activated kinases, JNK (c-Jun N-terminal kinase) and p38. The effectors of MAP kinases are transcription factors, including c-Jun and c-Fos, which form the activating protein complex AP-1 (activating protein-1). This protein plays important role in regulation of collagen metabolism as inhibitor of pro-collagen type I gene expression and transforming growth factor beta (TGF-β) signaling pathway [7,8,9].
Chronic exposure to UVA induces inflammation in the skin, resulting in the release of cytokines and proinflammatory factors, including the nuclear transcription factor NF-κB (Nuclear factor kappa B) [5,10,11]. Activated NF-κB plays an important role in regulating the expression of various genes involved in immune and inflammatory responses, including the expression of cyclooxygenase COX-2, an enzyme associated with the inflammatory process [12].
Seed oil derived from Amaranthus cruentus L. (AmO) is considered a promising source of bioactive compounds of anti-oxidative potential [13]. Its use may contribute to various health benefits, including anti-oxidant, anti-cancer, anti-allergic and anti-hypertensive activity. The oil obtained from the cold-pressing of Amaranthus cruentus L. has a relatively low lipid content, approximately 7-8%. However, these lipids are valuable for health due to the presence of unsaturated fatty acids, tocopherols, tocotrienols, phytosterols, and squalene, which are not commonly found together in other oils.
The effect of AmO on collagen metabolism and wound healing in cells exposed to UVA radiation is currently unknown. Given the oil's rich composition of bioactive compounds, it has the potential to exhibit protective effects not only against oxidative stress but also other metabolic disturbances induced by UV radiation. This study aims to investigate the effect of AmO on collagen biosynthesis and wound healing process in human skin fibroblasts exposed to UVA radiation.
2. Results and Discussion
Prolonged exposure of skin to UV radiation leads to generation of reactive oxygen species (ROS) in the skin cells. They cause a redox imbalance, deregulation of collagen synthesis and damage to DNA, proteins and lipids. They also affect the integrity of biological membranes of skin cells, fibroblasts and keratinocytes [5]. In our previous study we found that exposure of fibroblasts to different doses of UVA radiation contributed to decrease in the cell viability in a UVA dose-dependent manner [13]. Based on these data, in present study we have selected a dosage of 10 J/cm2, which was IC50 value for cell viability in our experimental protocols [13] to evaluate the effect of Amaranthus cruentus L. seed oil (AmO) on collagen metabolism and wound healing.
Plant oils are among the cosmetic materials that demonstrate beneficial effects on the skin, due to their chemical composition, primarily the presence of Essential Polyunsaturated Fatty Acids (PUFAs), as well as other components of the unsaponifiable fraction, such as squalene, phytosterols, and tocopherols [14]. AmO, in this regard, is characterized by a favorable chemical composition, containing approximately 60% linoleic acid, 8- 10% squalene, and tocopherols, thus demonstrating protective and antioxidant potential against UV radiation. In our previous study we demonstrated that fibroblasts viability decreased to approximately 55% of the control value when exposed to 10 J/cm2 UVA radiation dose. We have found that in AmO at 0.05%, 0.1%, and 0.15% concentrations evoked protective effect on decreased viability of fibroblasts exposure to UVA irradiation [13]. These data led us to evaluate the effect of AmO on collagen metabolism in fibroblasts exposed to UVA radiation.
Collagen is the main structural protein of connective tissue, providing the skin with proper tension, elasticity, and flexibility. Excessive exposure to the sun, extreme temperatures, and certain compounds in cosmetics can affect the structure of collagen fibers, resulting in worsened skin condition [15]. Collagen biosynthesis was evaluated by measurement of L-5-[3H]-proline incorporation into collagen proteins. The results were expressed as dpm of L-5-[3H]-proline released from collagenase-sensitive proteins per milligram of total protein in the homogenate extract, and expressed as a percentage of the control value (100%) (Figure 1).
We have found that AmO at studied concentrations slightly inhibited collagen biosynthesis in fibroblasts, however, the inhibition was statistically insignificant. When the cells were exposed to UVA radiation, collagen biosynthesis was decreased to 58% of the control value (statistically significant at p < 0.05). AmO at concentrations 0.05%, 0.1% and 0.15% partially counteracted UVA-dependent inhibition of collagen biosynthesis to about 78%, 81%, and 85% of the control value, respectively. This data suggest that AmO has a protective effect on collagen biosynthesis in human skin fibroblasts exposed to UVA radiation.
Collagen content in the tissues depends on the balance between collagen biosynthesis and degradation. An important regulator of collagen turnover is prolidase, cytoplasmic enzyme catalyzing the final stage of collagen degradation. It cleaves di- and tri-peptides containing C-terminal proline. Released proline is reused for collagen biosynthesis and cell growth [16]. Therefore, we decided to study the prolidase activity in human skin fibroblasts exposed to UVA radiation and treated with AmO. We assessed this activity using a colorimetric method, based on the measurement of proline released from the substrate (Gly-Pro), using the Chinard's reagent. AmO did not affect significantly prolidase activity in the studied cells. In UVA-terated cells prolidase activity was decreased to 54% of the control value. AmO at concentrations 0.05%, 0.1% and 0.15% partially counteracted UVA-dependent inhibition of prolidase activity to about 75%, 80%, and 85% of the control value, respectively (Figure 2). These data shows the protective effect of the AmO on UVA-induced impairment of collagen biosynthesis and prolidase activity in fibroblasts.
Collagen plays a significant role in cellular metabolism as a ligand of integrin receptors containing β1 subunit. The β1-integrin receptor is involved in signaling pathways that regulate collagen biosynthesis and prolidase activity. Interactions between collagen and these receptors activate intracellular signaling pathways, contributing to the regulation of various cellular metabolic functions [4].
AmO at 0.05% and 0.1% concentrations did not affect the expression of β1-integrin receptor as presented in Figure 3A. Slight decrease in the expression of this receptor was found at 0.15% of AmO. In UVA-treated cells the expression of this receptor was drastically decreased, while the application of AmO counteracted this process in a dose-dependent manner. Regarding the results of collagen biosynthesis and prolidase activity, there is another level of correlation between these processes and β1-integrin receptor expression in UVA and AmO treated cells. It suggests that UVA radiation, by decreasing the expression of the β1-integrin receptor, down-regulate prolidase activity, leading to reduced collagen production. The protective effect of AmO on the the UVA-induced down-regulation of β1-integrin expression may represent a molecular mechanism for protection of collagen biosyntheis against deleterious effect of UV radiation on this process.
Prolidase activity and collagen biosynthesis are regulated also by insulin-like growth factor I receptor (IGF-IR) signaling. IGF-I belongs to a peptide growth factors that play a significant role in cell metabolism, proliferation, differentiation and it is the most potent stimulator of collagen biosynthesis [6]. The expression of IGF-I is elevated in skin cells under inflammatory conditions. For instance, in psoriatic skin, fibroblasts evoke high expression of IGF-I [17].
To date, the effect of UVA on IGF-IR expression in cells of normal skin is not well recognized. The study presented in this report shows that fibroblasts exposed to UVA radiation evoke decreased IGF-IR receptor expression, suggesting the mechanism for UVA-induced inhibition of collagen biosynthesis. The application of AmO at tested concentrations counteracted the deleterious effect of UVA on IGF-IR expression (Figure 3B). These findings suggest that down regulation of IGF-IR signaling may play important role in the mechanism of UVA-induced inhibition of collagen biosynthesis. They also suggest the mechanism for protective action of AmO on this process.
We have confirmed the potential involvement of the β1-integrin and IGF-I receptors in the molecular mechanism by which UVA affects collagen biosynthesis and prolidase activity. It is well established that stimulation of these receptors lead to the activation of mitogen-activated protein kinases (MAPK) and the transforming growth factor β (TGF-β) [8,9]. MAPK signaling pathways play a important role in cellular metabolism. For instance in skin cells ROS induce decrease in the expression of phosphorylated forms of ERK 1/2 kinase (p-ERK 1/2) and increase in the expression of stress-activated kinase p38, leading to the up-regulation of expression of transcription factors, including c-Jun and c-Fos, which form the activator protein-1 (AP-1) complex [7,8,9]. This protein is a key regulator in various skin processes, including photoaging, by affecting the TGF-β signaling pathway and inhibiting the expression of type I procollagen gene. In our previous studies, we demonstrated that UVA radiation induced an increase of ROS generation in fibroblasts [13]. In present studies we show that UVA induced a decrease in the expression of p-ERK 1/2 (Figure 4) and TGF-β (Figure 5A) and an increase in p38 protein expression (Figure 5B). This confirms the hypothesis on the correlation between UVA-induced ROS generation, deregulation of MAPK-dependent signaling pathways and collagen biosynthesis in fibroblasts. From several other studies, it is evident that the application of antioxidants from natural sources (e.g., green tea, pomegranate extract), reverses the negative effects (deregulation of the above-described cellular signaling pathways) of oxidative stress, including that induced by UVA [18,19]. Our research supports the data, showing that AmO, in a dose-dependent manner, restored the UVA-induced decrease in the expression of p-ERK 1/2 (Figure 4) and TGF-β (Figure 5A), and restored to the control value UVA-induced increase in the expression of p38 kinase (Figure 5B). This confirms the protective effect of AmO on the deleterious impact of UVA on the investigated cellular signaling pathways.
UVA-induced skin damage is preceded by tissue inflammation. Nuclear factor-kappa B (NF-κB) has been described as one of the pro-inflammatory factors activated by UV radiation. It serves as a mediator of cellular responses to inflammatory stimuli, pathogens, and cellular stressors. In the cytoplasm of cells, NF-κB is present in an inactive form bound to the inhibitory protein IκB. It is known that NF-κB activation is induced by inflammatory stimuli, and subsequently, NF-κB translocate to the cell nucleus [20]. Activated NF-κB plays a crucial role in regulating the expression of various genes involved in immune and inflammatory responses, including the expression of cyclooxygenase COX-2, an enzyme participating in the inflammatory process. UVA radiation activates COX-2, which induces inflammatory process. Vostálová et al. [12] observed an increase in the expression of COX-2 in the skin of mice exposed to a dose of 20 J/cm2 of UVA. In our experiments, we demonstrated that UVA irradiation of fibroblasts at a dose of 10 J/cm2 significantly increased the expression of NF-κB and its translocation to the cell nucleus (Figure 6A), along with increased COX-2 expression (Figure 6B). This confirms the pro-inflammatory mechanism of UVA in the studied cellular model. AmO by itself did not induce changes in the expression and translocation of NF-kB to the nucleus, however, in cells exposed to the UV radiation significantly inhibited the expression of this factor. These experiments show that UVA-induced increase in the expression of COX-2 associated with the translocation of NF-kB into the nucleus of human skin fibroblasts is counteracted by AmO (Figure 6A,B). This shows protective action of AmO against UVA-induced inflammation in fibroblasts and suggests that AmO preparation may be useful as a potential therapeutic agent.
Disruption of skin integrity by external factors (e.g., UV radiation) activate reparative processes. Fibroblasts play a crucial role in skin wound healing, from the early inflammatory phase to the final production of extracellular matrix components. Contemporary research is exploring substances, including natural compounds, that can impact tissue regeneration. Plant oils have shown promising results in both in vitro and in vivo studies. They influence various phases of the wound healing process through their antimicrobial, anti-inflammatory, and antioxidant properties, as well as by promoting cell proliferation, enhancing collagen synthesis, stimulating skin regeneration, and repairing the skin's lipid barrier function. It has been demonstrated that the fatty acids present in these oils also play a significant role in the wound healing process. Linolenic, linoleic, and oleic acids serve as precursors for the synthesis of inflammatory or anti-inflammatory mediators and are integral components of cell membrane phospholipids, ceramides, and sebum, all of which are vital constituents of the lipid barrier [21].
Plant-based oils are mixtures of glyceryl esters and higher fatty acids, and they also contain phytosterols, vitamin E and its derivatives, and squalene. It has been demonstrated that triglycerides, free fatty acids, glycerol, and nonsaponifiable compounds directly or indirectly influence the wound healing process. The biological effect of plant oils in wound healing is mainly attributed to their similarity to skin lipids. Numerous studies confirm that various natural oils, under different conditions, exhibit more or less effective actions in the wound healing process. For example, in vitro studies have shown that cold-pressed rapeseed oil stimulates fibroblast proliferation and promotes fibroblast migration to the wound area, indicating its wound-healing effects. Similar effects were observed with flaxseed oil [21]. Additionally, plant oils like borage oil, evening primrose oil, and avocado oil, containing gamma-linolenic acid (GLA), are beneficial for skin conditions, inflammation, and irritations. Borage oil, with its high GLA content, stimulates skin cell activity and regeneration, making it useful in treating skin conditions such as allergies and inflammations. Evening primrose oil also supports skin regeneration, eases skin issues, and reduces inflammation, making it a good choice for those with conditions like psoriasis. Avocado oil, rich in vitamin E, β-carotene, vitamin D, protein, lecithin, and fatty acids, offers substantial benefits when added to formulations [22]. In respect to AmO there is a lack of studies on its role in skin repair processes, so it was essential to conduct this study.
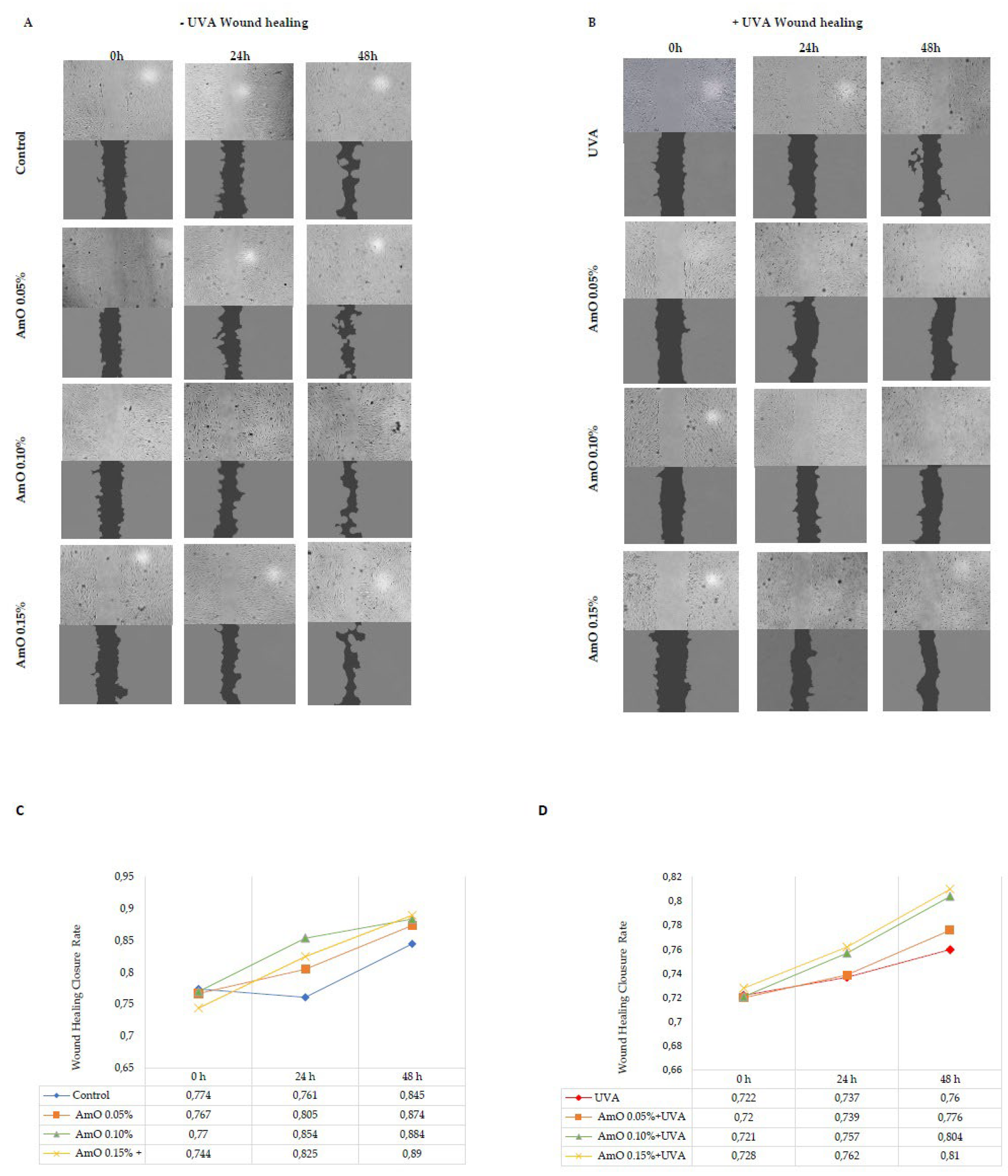
The results of wound closure are presented in images obtained from a fluorescence microscope and subjected to analysis using ImageJ® software with the Wound Healing Size Tool extension (Figure 7A,B). The wound healing closure rate was calculated and presented on a chart and in digital form (Figure 7C,D). The obtained results show that AmO stimulated fibroblast proliferation and promoted their migration in a model of wound healing under control conditions (without exposure to UVA radiation). All studied concentrations of AmO, (0.05%, 0.1%, and 0.15%) positively influenced the wound healing process in a time and dose dependent manner. Interestingly, the ability of AmO to stimulate fibroblast proliferation and migration was also enhanced after exposure to UVA, although less intensely than in non-UVA-irradiated cells. The effectiveness of AmO in the wound healing process may be attributed to the presence of components such as linoleic acid, squalene, derivatives of vitamin E, and phytosterols. These components not only scavenge free radicals, as demonstrated in our previous study [13], but also have a beneficial impact on the wound healing processes, as presented in this report.
It is known that migration, adhesion, proliferation, neovascularization, remodeling, and apoptosis are the key processes in wound healing, affected by ROS. An increased production of ROS can disrupt these processes [23]. Studies by Wlaschek and Scharffetter-Kochanek [24] have shown that changes in mitochondrial DNA induced by UV radiation can increase intracellular ROS levels in fibroblasts, leading to alterations in their migration and proliferation. In vitro wound healing tests have demonstrated a significantly reduced ability of fibroblasts to close the wound gap, suggesting that an excess of ROS in mitochondria negatively affects the wound healing process. This explains how the overlapping processes, resulting in increased ROS production due to cell damage in fibroblast exposed to UVA radiation limit the regenerative capacity of the compound studied in our study, in the wound healing process [24].
It has been demonstrated that therapies involving the application of plant-derived butters or oils are efficacious with limited adverse effects, rendering phytotherapy a compelling alternative for dermatological treatments. Fatty acids are recognized to play a significant role in the described wound healing processes, whereas non-saponifiable compounds can substantially contribute to antimicrobial, antioxidant, and anti-inflammatory effects [14]. Therefore, it is advisable, based on our research findings, to consider the use of unrefined plant-derived oils, including AmO in cosmetics, with a protective effect against solar radiation. Therefore, results of this study provide rational for applications of AmO in therapeutics of protective activity against UVA-induced skin damage
4. Materials and Methods
4.1. Materials
Plant Materials
The Amaranthus cruentus L. seed oil used in this research was a validated, commercially available product sourced from Szarłat M. and W. Lenkiewicz s.j. in Zawady, Poland. This oil was obtained through an unrefined seed cold-pressing process from locally cultivated Amaranthus cruentus plants. The fatty acid composition of the oil was as follows: palmitic acid accounted for 17.5–19.2% of the oil, oleic acid for 17.5–20.5%, linoleic acid for 52–55%, and stearic acid for 4.5–5.5%. Additionally, the oil contained approximately 6–8% of squalene and 8–10 mg of vitamin E per 100 grams. For detailed safety information regarding the investigated Amaranthus cruentus L. seed oil, refer to the comprehensive safety data sheet (SDS) provided by the manufacturer, Szarłat company. This document is available in the Supplementary Materials.
4.2.1. Cell Lines and Culture
Human dermal fibroblasts were obtained from the American Type Culture Collection (Manassas, VA, USA); maintained in Dulbecco’s Modified Eagle Medium (DMEM) (PANTM BIOTECH, Aidenbach, Germany); supplemented with 10% Fetal Bovine Serum (FBS) (Gibco, Waltham, MA, USA), 50 U/mL of Penicillin (Pen) (Gibco, Waltham, MA, USA) and 50 μg/mL of Streptomycin (Strep) (Gibco, Waltham, MA, USA); and incubated at 37 °C in 5% CO2. The cells were grown on 100 mm dishes in 10 mL of complete medium. The cell culture medium was changed 2–3 times per week. The cells were used after 8-10 passages. For the experiments with the investigated oil, we used DMEM without FBS and Pen/Strep. The AmO was used in the following concentrations: 0.05%, 0.1%, 0.15%. After 30 min of incubation, the cell culture media containing the studied oil were removed and the plates were washed with PBS. Subsequently, the cells were exposed to UVA radiation using a Bio-Link Crosslinker BLX 365 (Vilber Lourmat, Eberhardzell, Germany) at dose 10 J/cm2 in 1 mL of cold PBS (4 °C). After irradiation, PBS was exchanged for fresh DMEM. The cells were incubated for 24 h. The concentration of AmO and the 10 J/cm2 dose of UVA were selected based on cell viability measurements obtained from the MTT assay conducted in our previous study [13] (data not shown).
4.2.2. Collagen Biosynthesis Assay
In this experiment, cells were cultured in 100-mm plates, with approximately 1 × 10^6 cells per plate. The confluent fibroblast cells were treated as described in Section 4.2.1. Collagen biosynthesis in this research was evaluated using radioactive 5-[3H]-proline (5 μCi/mL; Hartmann Analytic, Germany) incorporated into proteins that are susceptible to bacterial collagenase. The Peterkofsky's method [25] was employed for this purpose. After a 24-hour incubation, the cells were washed with PBS (pH 7.4), collected in PBS containing 10 mM proline, and then stored at -80°C until the day of analysis. To perform collagen digestion, purified Clostridium histolyticum collagenase from Sigma Aldrich, Saint Louis, MO, USA, was used. Radiometric analysis was carried out using the Liquid Scintillation Analyzer Tri-Carb 2810 TR from PerkinElmer, Waltham, MA, USA. The results obtained from this analysis were standardized against total protein biosynthesis and presented as a percentage of the control value, providing valuable insights into collagen biosynthesis in the context of the study.
4.2.3. Prolidase Activity
In this experiment, cells were cultured in 100-mm plates, with approximately 1 × 10^6 cells per plate. The confluent fibroblast cells were treated as described in Section 4.2.1. After the 24-hour incubation, the cells were collected and subjected to a prolidase assay, following the method described by Myara [26]. To determine the total protein concentration in the samples, the Lowry method [27] was used. Prolidase activity was reported as the nanomoles of proline released from the synthetic substrate (Gly-Pro) within one minute, normalized per milligram of the supernatant protein in the cell homogenate.
4.2.4. Western Immunoblot Analysis
For the analysis of the protein expression via Western blotting, the cells were cultured in 100 mm plates at about 2.0 × 106 cells. The confluent fibroblast cells were treated as described in Section 4.2.1. After 24 h of incubation, the culture media were removed and the cells were harvested using a cell lysis buffer supplemented with a protease/phosphatase inhibitor cocktail. The protein concentrations of the samples were determined via the Lowry method [27]. Then, the proteins were separated using the SDS-PAGE method described by Laemmli [28]. After this step, the gels were washed in cold Towbin buffer (25 mM Tris, 192 mM glycine, 20% (v/v) methanol, 0.025–0.1% SDS, pH 8.3). The proteins in the gels were transferred onto the 0.2 µm nitrocellulose membranes using a Trans-Blot (BioRad, Hercules, CA, USA). The transfer conditions were 200 mA, 3 h in freshly prepared Towbin buffer and the temperature was maintained around 4–8 °C. The blocking of the membranes was performed using 5% NFDM for 1 h at RT. When the blocking was complete, the membranes were washed three times with 20 mL of TBS-T (20 mM Tris, 150 mM NaCl and 0.1% Tween® 20). After the washing step, the membranes were incubated with primary antibodies overnight at 4 °C. The concentration of the primary antibodies was 1:1000. Further-more, the membranes were washed three times with 20 mL of TBS-T and a secondary antibody conjugated with HRP solutions (1:3000) in 5% NFDM was used for 1 h at RT. Then, the membranes were washed 3 times with 20 mL of TBS-T and visualized using BioSpectrum Imaging System UVP (Ultra-Violet Products Ltd., Cambridge, UK).
4.2.5. Immunofluorescence Staining and Confocal Microscopy
The cells were cultured on a black 96-well plate. The confluent fibroblast cells were treated as described in Section 4.2.1. After 24 h, the culture media were removed and the cells were fixed with a 3.7% formaldehyde solution at room temperature for 10 min. Then, the plate was washed once with 100 µL/well of PBS. Later, the permeabilization with a 0.1% Triton X-100 solution and a 10 min step was performed. After the permeabilization the plate was washed twice with PBS and 3% FBS was used as a blocking agent at room temperature for 30 min. After the FBS removal, 50 µL of the primary antibody (1:50), diluted in 3% FBS, was added and the plate was incubated for 1 h at room temperature. After the incubation, the primary antibody plate was washed three times with PBS. Then, 50 µL per well of the secondary antibody (dilution 1:1000) was added for the next 1 h. During this step, the plate was covered from the light. When the secondary antibody solution was removed, the plate was washed 3 times with PBS and the wells were filled with 100 µL of PBS containing 2 µg/mL of Hoechst 33342 for the nuclei staining. The plate was visualized using a confocal laser scanning microscope BD Pathway 855 (Bioimager, Becton Dickinson, Franklin Lakes, NJ, USA) supported with AttoVisionTM 1.6 software.
4.2.6. Cell Migration Assay
Confluent fibroblasts plated in a 6-well plates were scratched with a sterile 200 µL pipette tip and treated as described in Section 4.2.1. The scratched area was monitored with an inverted optical microscope at 40× magnification (Nikon; Minato, Tokyo, Japan). Fibroblast migration was calculated using ImageJ® software and its rate was calculated according to the following formula.
5. Conclusions
UVA radiation impairs collagen biosynthesis, prolidase activity, the expression of β1-integrin receptor, IGF receptor, phosphorylated forms of ERK 1/2 kinases, and TGF-β factor, while it stimulates the expression of p38 kinase. The application of AmO reversed the adverse effects induced by UVA on these processes. UVA radiation induces the expression and nuclear translocation of the pro-inflammatory NF-κB factor and enhances COX-2 expression. The use of AmO significantly inhibited the expression of these factors. Amaranthus cruentus L. seed oil stimulate the proliferation and migration of fibroblasts in a wound healing model. After cell exposure to ultraviolet radiation, AmO promoted the wound healing process. The results of these studies allow us to assume that the presented properties of AmO may be used in preparations that protect the skin against the harmful effects of UV radiation.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Conception and design of the study, K.W. and A.S.; performance of experiments and tests, K.W., M.C. B.M. and A.S.; analysis and interpretation of the data, K.W. and A.S.; drafting of the article, K.W. and A.S.; final approval of the version to be submitted, A.S and J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the MUB grant number: B.SUB.23.399/01.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the main text of this article or on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yaar, M.; Gilchrest, B. A. Photoageing: Mechanism, Prevention and Therapy. Br. J. Dermatol. 2007, 157(5), 874–887. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R. M.; Gerberick, G. F.; Veres, D.; Irwin, C. J.; Kaidbey, K. H. Cumulative Effects from Repeated Exposures to Suberythemal Doses of UVB and UVA in Human Skin. J. Am. Acad. Dermatol. 1995, 32(1), 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kouba DJ, Chung KY, Nishiyama T, Vindevoghel L, Kon A, Klement JF, Uitto J, Mauviel A. Nuclear factor-kappa B mediates TNF-alpha inhibitory effect on alpha 2(I) collagen (COL1A2) gene transcription in human dermal fibroblasts. J Immunol. 1999, 1; 162(7), 4226–4234.
- Surazynski, A.; Miltyk, W.; Palka, J.; Phang, J. M. Prolidase-Dependent Regulation of Collagen Biosynthesis. Amino Acids 2008, 35(4), 731–738. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, H.; Yang, Y.; Zhang, S.; Wang, J.; Zhang, D.; Yu, H. Metformin Attenuates UVA-Induced Skin Photoaging by Suppressing Mitophagy and the PI3K/AKT/mTOR Pathway. Int. J. Mol. Sci. 2022, 23(13), 6960. [Google Scholar] [CrossRef] [PubMed]
- Baszanowska, W.; Misiura, M.; Oscilowska, I.; Palka, J.; Miltyk, W. Extracellular Prolidase (PEPD) Induces Anabolic Processes through EGFR, Β1-Integrin, and IGF-1R Signaling Pathways in an Experimental Model of Wounded Fibroblasts. Int. J. Mol. Sci. 2021, 22(2), 942. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R. J.; Greenberg, M. E. Opposing Effects of ERK and JNK-P38 MAP Kinases on Apoptosis. Science 1995, 270(5240), 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Verheij, M.; Bose, R.; Lin, X. H.; Yao, B.; Jarvis, W. D.; Grant, S.; Birrer, M. J.; Szabo, E.; Zon, L. I.; Kyriakis, J. M.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. N. Requirement for Ceramide-Initiated SAPK/JNK Signalling in Stress-Induced Apoptosis. Nature 1996, 380(6569), 75–79. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R. Skin Antioxidants: Their Role in Aging and in Oxidative Stress--New Approaches for Their Evaluation. Biomed. Pharmacother. Biomedecine Pharmacother. 1999, 53(4), 181–192. [Google Scholar] [CrossRef]
- Dadej, I.; Wołowiec, J. The Role of UVA of the Skin Pathology. Adv. Dermatol. Allergol. Dermatol. Alergol. 2003, 20(3), 170–175. [Google Scholar]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New Insights in Photoaging, UVA Induced Damage and Skin Types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Vostalova, J.; Rajnochova Svobodova, A.; Galandakova, A.; Sianska, J.; Dolezal, D.; Ulrichova, J. Differential Modulation of Inflammatory Markers in Plasma and Skin after Single Exposures to UVA or UVB Radiation in Vivo. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czechoslov. 2013, 157(2), 137–145. [Google Scholar] [CrossRef] [PubMed]
- Wolosik, K.; Chalecka, M.; Palka, J.; Surazynski, A. Protective Effect of Amaranthus Cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts. Int. J. Mol. Sci. 2023, 24(13), 10795. [Google Scholar] [CrossRef] [PubMed]
- Poljšak, N.; Kočevar Glavač, N. Vegetable Butters and Oils as Therapeutically and Cosmetically Active Ingredients for Dermal Use: A Review of Clinical Studies. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Jariashvili, K.; Madhan, B.; Brodsky, B.; Kuchava, A.; Namicheishvili, L.; Metreveli, N. Uv Damage of Collagen: Insights from Model Collagen Peptides. Biopolymers 2012, 97(3), 189–198. [Google Scholar] [CrossRef] [PubMed]
- Karna, E.; Szoka, L.; Huynh, T. Y. L.; Palka, J. A. Proline-Dependent Regulation of Collagen Metabolism. Cell. Mol. Life Sci. 2020, 77(10), 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Silesian College of Medicine in Katowice; Faruga-Lewicka, W.; Kardas, M.; Medical University of Silesia in Katowice. The Influence of Insulin-like Growth Factor IGF-1 on the Course of Acne Vulgaris. Aesthetic Cosmetol. Med. 2022, 11(3), 105–108. [Google Scholar] [CrossRef]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M. S.; Mukhtar, H. Pomegranate Fruit Extract Modulates UV-B-Mediated Phosphorylation of Mitogen-Activated Protein Kinases and Activation of Nuclear Factor Kappa B in Normal Human Epidermal Keratinocytes Paragraph Sign. Photochem. Photobiol. 2005, 81(1), 38–45. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S. K.; Matsui, M. S.; Elmets, C. A.; Mukhtar, H. Polyphenolic Antioxidant (-)-Epigallocatechin-3-Gallate from Green Tea Reduces UVB-Induced Inflammatory Responses and Infiltration of Leukocytes in Human Skin. Photochem. Photobiol. 1999, 69(2), 148–153. [Google Scholar] [CrossRef]
- O’Dea, E. L.; Kearns, J. D.; Hoffmann, A. UV as an Amplifier Rather than Inducer of NF-κB Activity. Mol. Cell 2008, 30(5), 632–641. [Google Scholar] [CrossRef]
- Poljšak, N.; Kreft, S.; Kočevar Glavač, N. Vegetable Butters and Oils in Skin Wound Healing: Scientific Evidence for New Opportunities in Dermatology. Phytother. Res. PTR 2020, 34(2), 254–269. [Google Scholar] [CrossRef]
- Korać, R. R.; Khambholja, K. M. Potential of Herbs in Skin Protection from Ultraviolet Radiation. Pharmacogn. Rev. 2011, 5(10), 164–173. [Google Scholar] [CrossRef]
- Janda, J.; Nfonsam, V.; Calienes, F.; Sligh, J. E.; Jandova, J. Modulation of ROS Levels in Fibroblasts by Altering Mitochondria Regulates the Process of Wound Healing. Arch. Dermatol. Res. 2016, 308(4), 239–248. [Google Scholar] [CrossRef] [PubMed]
- Wlaschek, M.; Scharffetter-Kochanek, K. Oxidative Stress in Chronic Venous Leg Ulcers. Wound Repair Regen. Off. Publ. Wound Heal. Soc. Eur. Tissue Repair Soc. 2005, 13(5), 452–461. [Google Scholar] [CrossRef] [PubMed]
- Peterkofsky, B.; Diegelmann, R. Use of a Mixture of Proteinase-Free Collagenases for the Specific Assay of Radioactive Collagen in the Presence of Other Proteins. Biochemistry 1971, 10(6), 988–994. [Google Scholar] [CrossRef] [PubMed]
- Myara, I.; Charpentier, C.; Lemonnier, A. Optimal Conditions for Prolidase Assay by Proline Colorimetric Determination: Application to Iminodipeptiduria. Clin. Chim. Acta 1982, 125(2), 193–205. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O. H.; Rosebrough, N. J.; Farr, A. L.; Randall, R. J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193(1), 265–275. [Google Scholar] [CrossRef]
- Laemmli, U. K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227(5259), 680–685. [Google Scholar] [CrossRef]
Figure 1.
Collagen biosynthesis in fibroblasts irradiated with UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. The mean ± standard error (SEM) values from the experiments performed in triplicates are presented. * Statistically significant differences at p < 0.05 compared to the control, ** Statistically significant difference at p<0.05 compared to UVA.
Figure 1.
Collagen biosynthesis in fibroblasts irradiated with UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. The mean ± standard error (SEM) values from the experiments performed in triplicates are presented. * Statistically significant differences at p < 0.05 compared to the control, ** Statistically significant difference at p<0.05 compared to UVA.
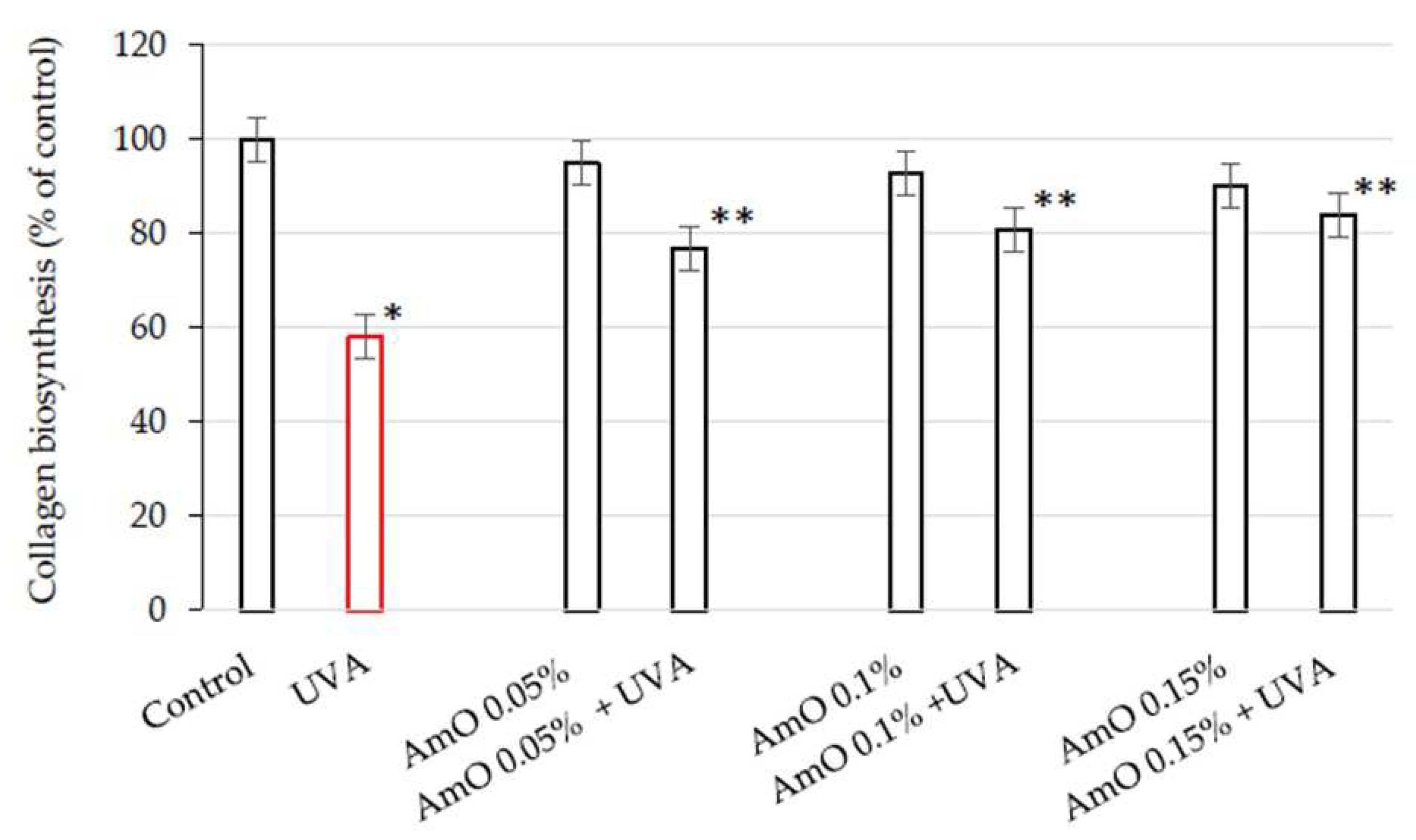
Figure 2.
Prolidase activity in fibroblasts irradiated with UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. The mean ± standard error (SEM) values from the experiments performed in triplicates are presented. * Statistically significant differences at p < 0.05 compared with the control, ** Statistically significant difference at p<0.05 compared to UVA.
Figure 2.
Prolidase activity in fibroblasts irradiated with UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. The mean ± standard error (SEM) values from the experiments performed in triplicates are presented. * Statistically significant differences at p < 0.05 compared with the control, ** Statistically significant difference at p<0.05 compared to UVA.
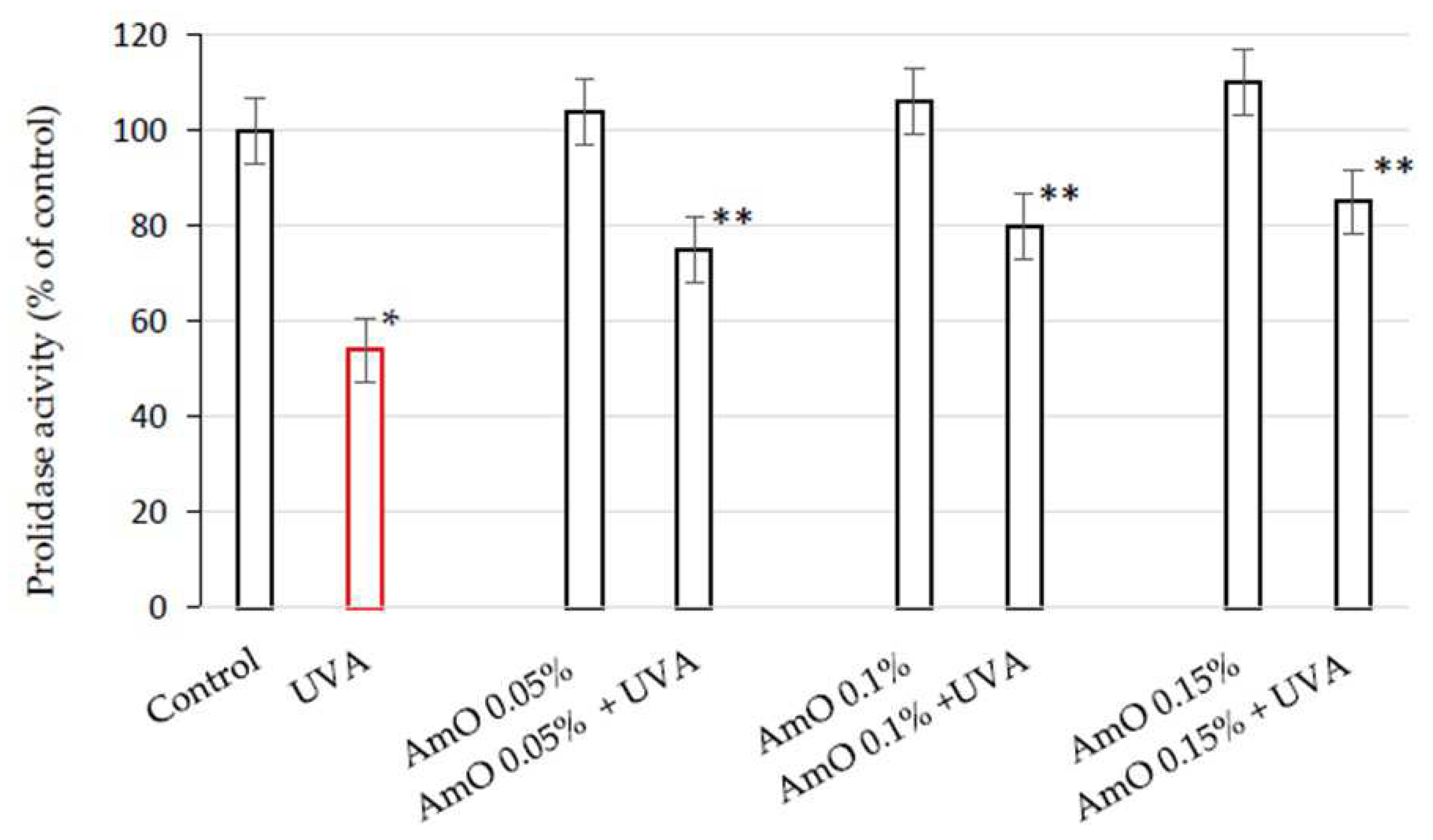
Figure 3.
Representative Western blot and densitometry analysis (done by ImageJ®) for β1- integrin receptor (A) and IGF-IR (B) expression in UVA-irradiated fibroblasts cultured in the presence of AmO at concentrations of 0.05%, 0.1% and 0.15% vs. control.
Figure 3.
Representative Western blot and densitometry analysis (done by ImageJ®) for β1- integrin receptor (A) and IGF-IR (B) expression in UVA-irradiated fibroblasts cultured in the presence of AmO at concentrations of 0.05%, 0.1% and 0.15% vs. control.
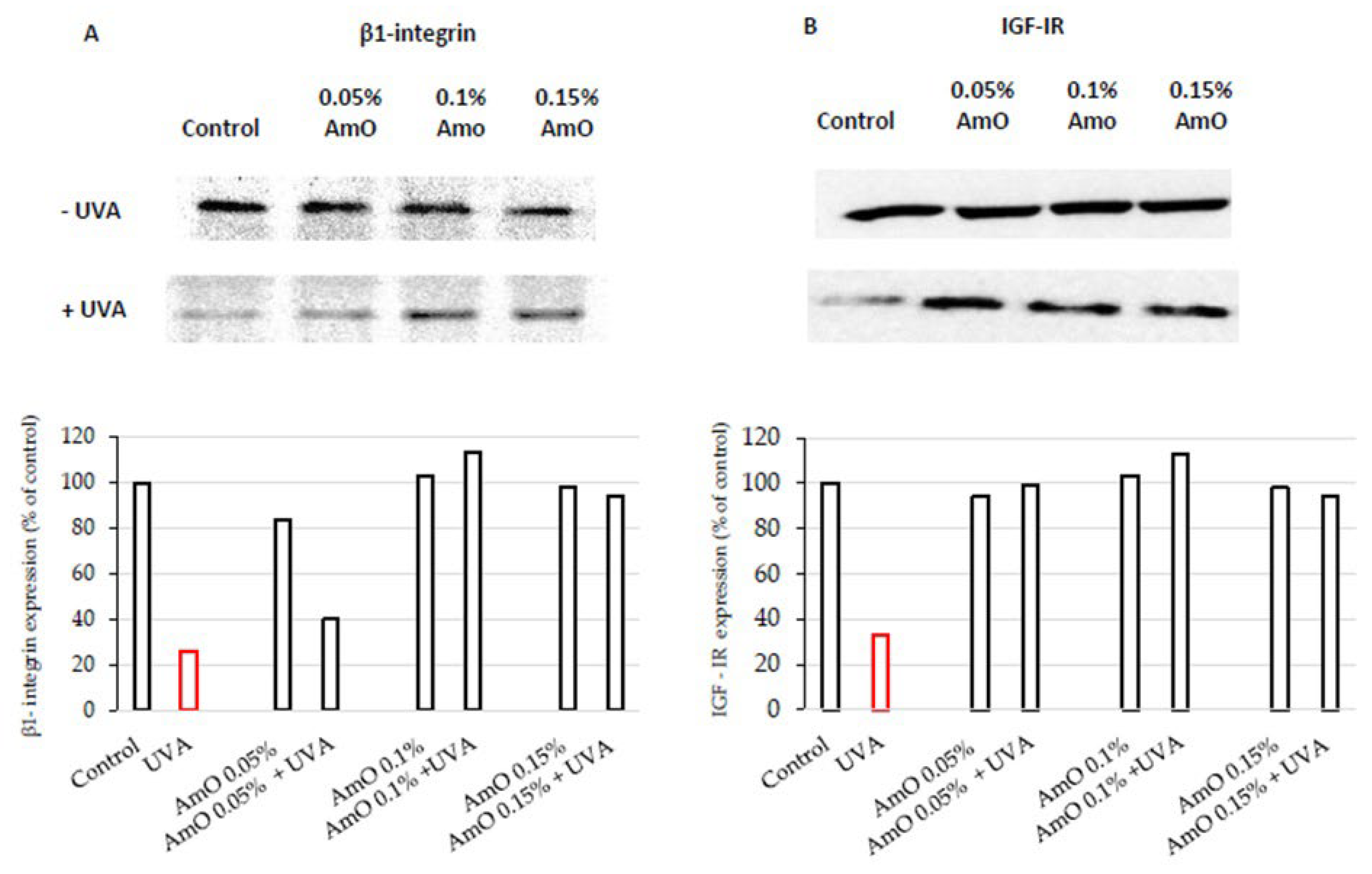
Figure 4.
Expression of p-ERK 1/2 kinase (visualized by immunofluorescence staining) in fibroblasts irradiated by UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. Blue staining indicates the nuclei and red staining represents ERK 1/2 expression. The images were obtained at a 20× magnification.
Figure 4.
Expression of p-ERK 1/2 kinase (visualized by immunofluorescence staining) in fibroblasts irradiated by UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. Blue staining indicates the nuclei and red staining represents ERK 1/2 expression. The images were obtained at a 20× magnification.
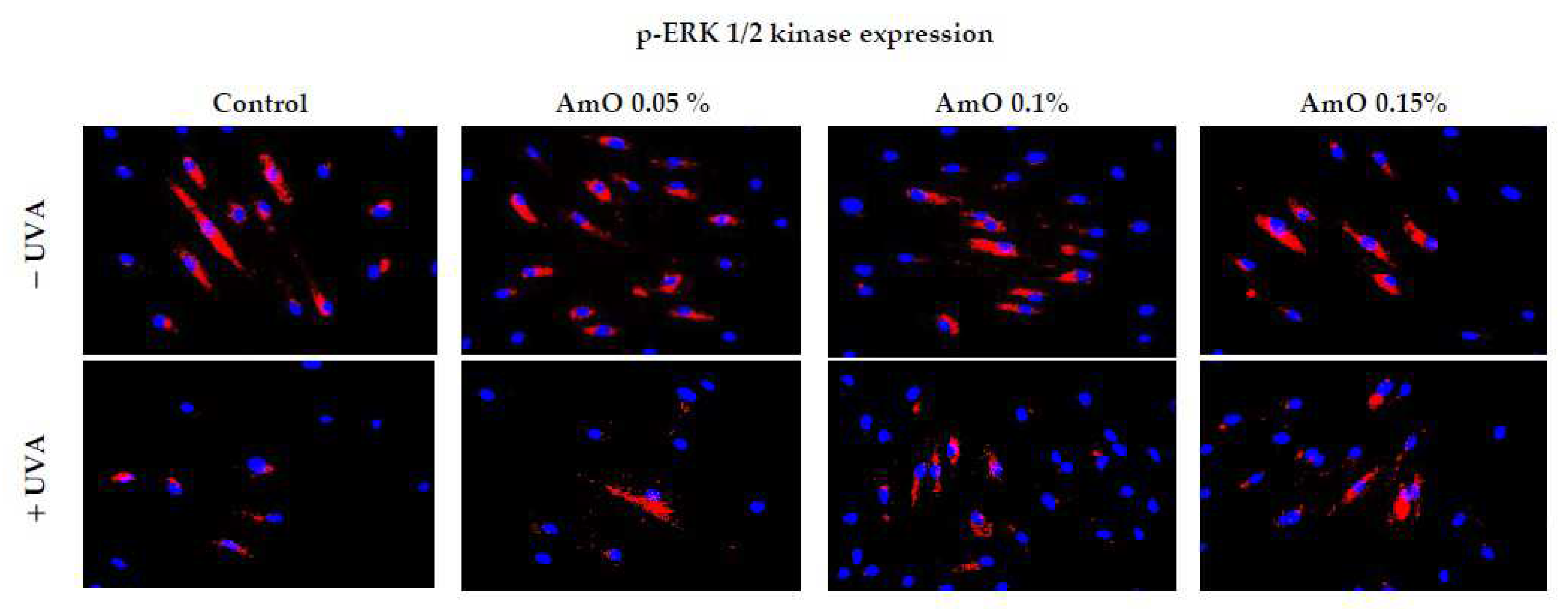
Figure 5.
Representative Western blot and densitometry analysis (done by ImageJ®) for TGF-β (A) and p38 (B) expression in UVA-irradiated fibroblasts cultured in the presence of AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control.
Figure 5.
Representative Western blot and densitometry analysis (done by ImageJ®) for TGF-β (A) and p38 (B) expression in UVA-irradiated fibroblasts cultured in the presence of AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control.
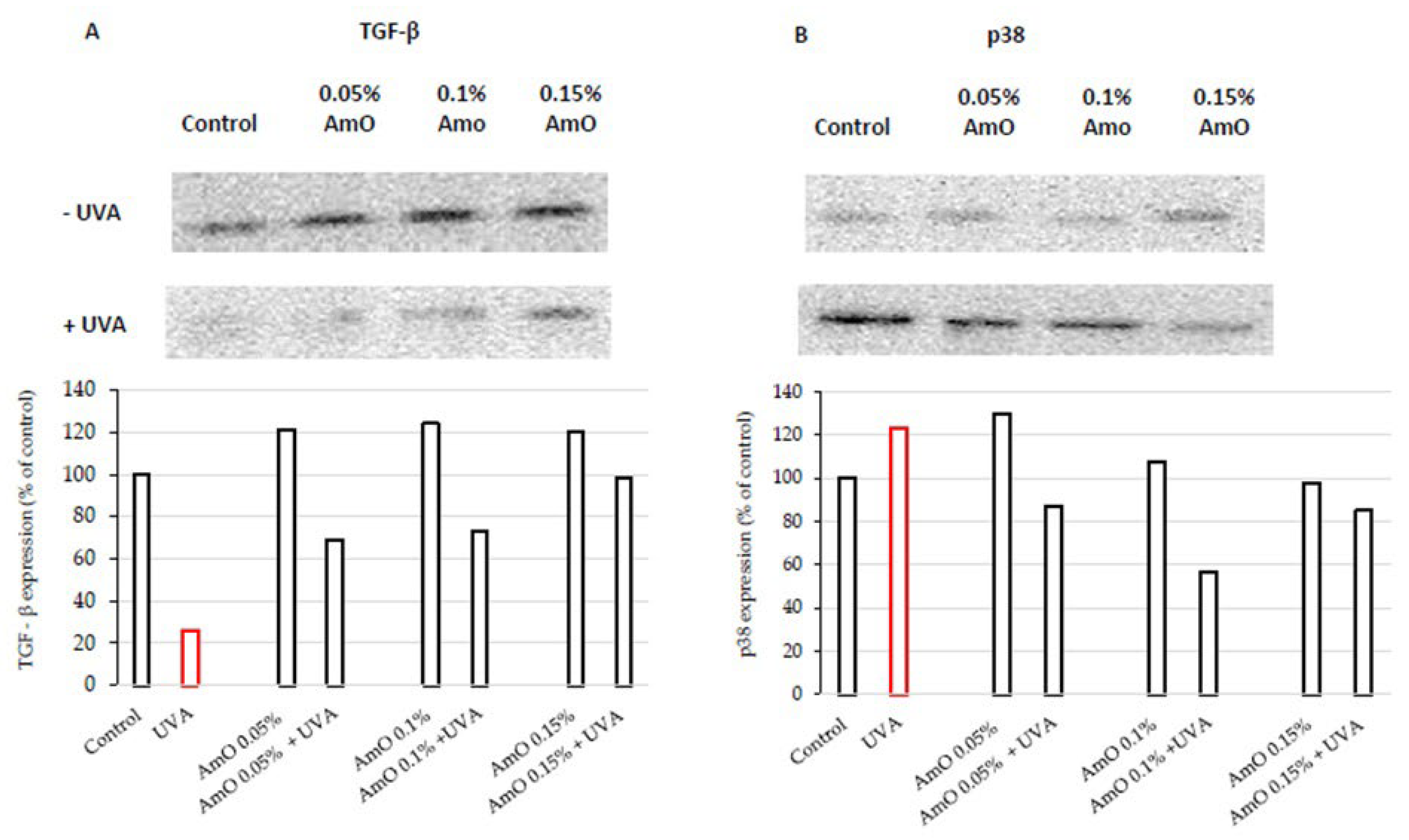
Figure 6.
(A) Expression of NFκB (visualized by immunofluorescence staining) in fibroblasts irradiated by UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. Blue staining indicates the nuclei and red staining represents NF-κB expression. The images were obtained at a 20× magnification. (B) Western blot and densitometry analysis (done by ImageJ®) for NFκB expression in UVA-irradiated fibroblasts and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control.
Figure 6.
(A) Expression of NFκB (visualized by immunofluorescence staining) in fibroblasts irradiated by UVA and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control. Blue staining indicates the nuclei and red staining represents NF-κB expression. The images were obtained at a 20× magnification. (B) Western blot and densitometry analysis (done by ImageJ®) for NFκB expression in UVA-irradiated fibroblasts and treated with AmO at concentrations of 0.05%, 0.1% and 0.15% vs. the control.
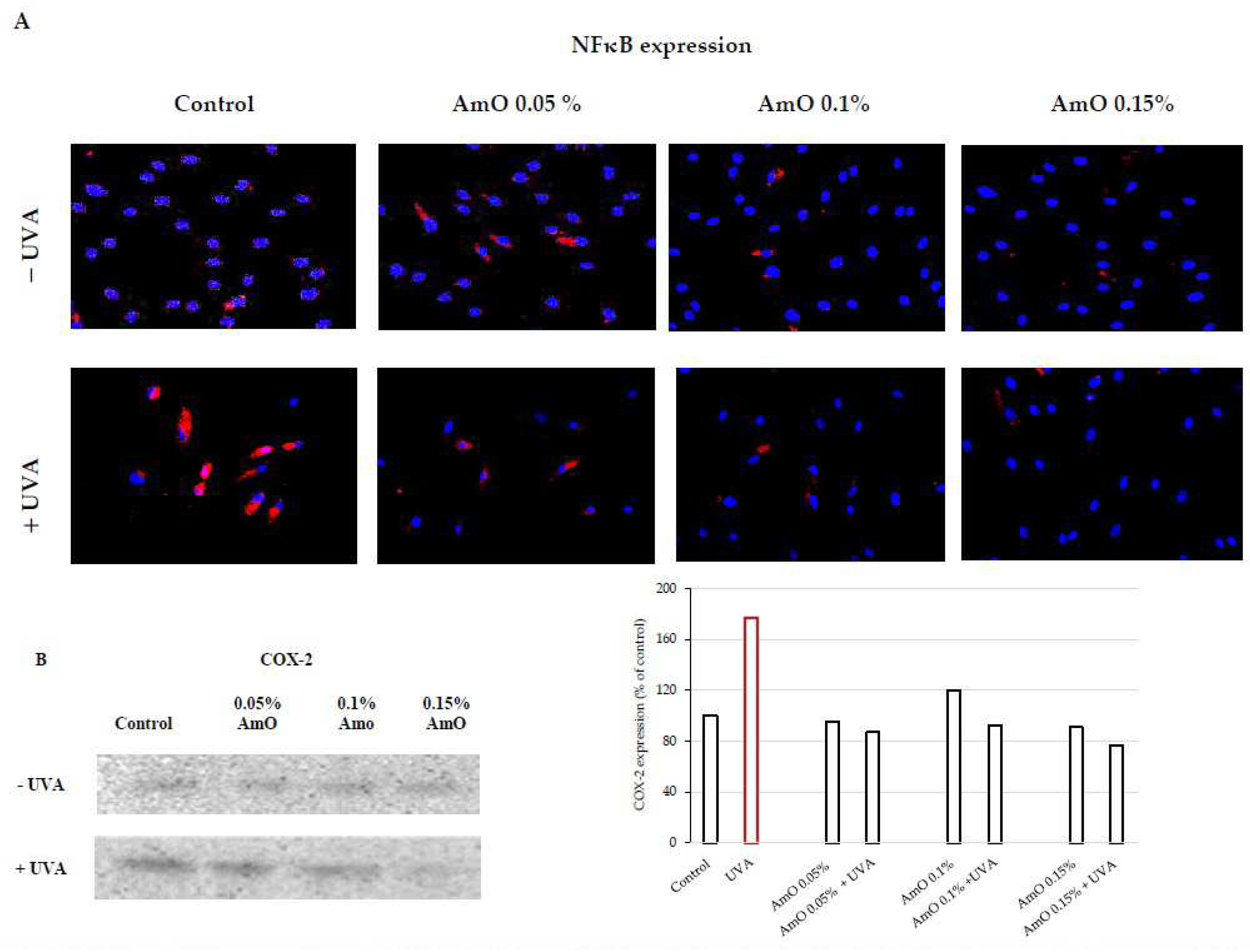
Figure 7.
Wound healing images in (A) non-UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15% (B) UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15%. The wound healing closure rate was calculated and analyzed by ImageJ software® in (C) non-UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15% (D) UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15%.
Figure 7.
Wound healing images in (A) non-UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15% (B) UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15%. The wound healing closure rate was calculated and analyzed by ImageJ software® in (C) non-UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15% (D) UVA-irradiated fibroblasts, and treated for 24 h and 48 h with AmO at concentrations of 0.05%, 0.1%, and 0.15%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



