Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Schizophrenia is a spectrum of neuropsychiatric deformities, characterized by hallucination, delusion, mood disorders, speech pathology, and neurocognitive deficits. Among various clinical manifestations, hallucination has been recognized as a core psychotic symptom that occurs more frequently in schizophrenia. A significant number of subjects with neurocognitive disorders like Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD), and other neurological diseases like stroke and epileptic seizure also experience hallucinations. While aberrant neurotransmission has been linked to the neuropathogenic events of schizophrenia, the specific cellular mechanism contributing to hallucinations remains ambiguous. Neurodegeneration in the hippocampus of the brain has been identified as a predominant pathogenic determinant of dementia. While the scientific proof for the neurodegeneration in schizophrenia is limited, the occurrence of dementia in schizophrenia has become increasingly evident. To note, both neurodegenerative, neurodevelopmental, and neuropsychiatric disorders display impaired neurogenesis in the brain. Impaired neurogenesis in the hippocampus has been linked to dementia and mood disorders. Notably, the early phase of many neurodegenerative disorders has been characterized by reactive neuroblostosis and aberrant cell cycle activation in mature neurons leading to the fluctuation in neurogenic processes leading to abnormal synaptogenesis and neurotransmission in the brain. Thus, this article emphasizes a hypothesis that aberrant neurogenic processes could be an underlying mechanism of hallucination in schizophrenia and other neurological diseases.
Keywords:
Schizophrenia
; hallucination
; neurogenesis
; reactive neuroblastosis
; dementia
Introduction
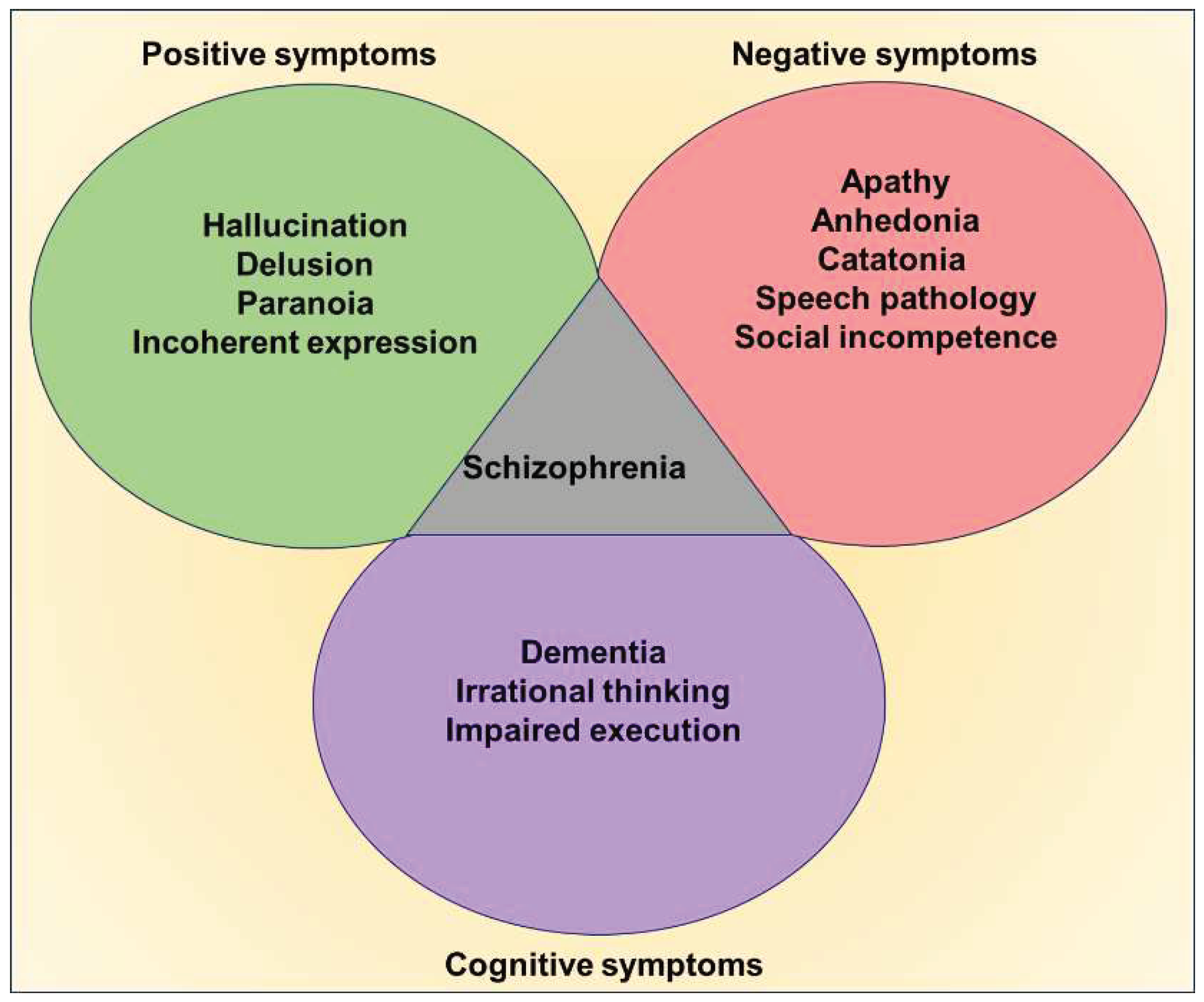
Schizophrenia is a debilitating neuropsychiatric disorder, characterized mainly by hallucination, delusions, mood disorders, and cognitive deficits[1,2,3]. Initially, Emil Kraepelin described the clinical features of this peculiar affective disorder as dementia praecox, and manic depression due to the admixer of behavioral deformities overlapping with various mental illnesses [4]. Later on, the term schizophrenia was suggested by Eugen Bleuler in 1908 and provided a further description of the different positive and negative psychotic symptoms [2,5]. While frequent episodes of hallucinations, delusions, paranoia, abnormal exhilaration, irrational thinking, and inexplicable behaviors are the positive symptoms of schizophrenia, the obvious negative symptoms comprise speech disorders, apathy, emotional blunting, catatonia, depression, and suicidal thoughts. Considerable degree of memory loss, deterioration of interpersonal skills, and attention deficits are the key cognitive deficits noticed in schizophrenia [1,2,4,6].
Figure 1.
Clinical symptoms of schizophrenia; The digital highlights the key clinical manifestations of schizophrenia, the positive, negative, and cognitive symptoms.
Figure 1.
Clinical symptoms of schizophrenia; The digital highlights the key clinical manifestations of schizophrenia, the positive, negative, and cognitive symptoms.
While the schizoid symptoms in childhood are rare, subtle prodrome signs appear to develop in the late teenage and thus marked degree of altered behavioral patterns is distinguishable in a significant percentage of individuals during their fourth decade of life[6,7]. The prevalence of schizophrenia is almost 1% of the global population regardless of gender and ethnicity [1,8]. Presently, there are no structured diagnostic strategies and confirmative biomarkers available to delineate the behavioral symptoms and pathogenesis of schizophrenia, because the etiology of the disease is not distinctive, the psychotic patterns are comorbid, vary among patients, and differ over time. The symptoms of schizophrenia appear to often be co-occurred with psychosis, BPD, OCD and MDD [9,10]. Despite the accumulation of enormous clinical reports and case studies, the precise etiology and distinguished pathogenic mechanisms accountable for the onset and progression of schizophrenia remain to be established. Also, the screening strategies for prenatal diagnosis and preventive measures for schizophrenia are highly limited. Thus, the therapeutic targets for establishing an effective curative regime remain ambiguous. The available interventional medications such as tailored combinations of antipsychotics, antidepressants, and anxiolytic agents are aimed at merely handling behavioral disorders through modulation of aberrant neurotransmission in the brain [1,11]. Moreover, many neuropharmacological approaches implemented for schizophrenia pose unforeseen adverse effects rather than a cure [1,12]. Therefore, there is a crucial need for scientific advancement in deciphering the underlying pathogenic determinants of schizophrenia which would help in identifying the ultimate therapeutic target. Reactive astrogliosis, an overpopulation of pathogenic astrocytes has widely been regarded as a non-neuronal pathogenic consequence leading to disruption of brain homeostasis and creating an imbalance in neurotransmitters during clinical episodes of various mental innless [13]. Though experimental evidence highlights the alterations in the expression of astrocyte-related genes, the reports on the abnormal astrogliogenic events in schizophrenia are inconsistent [13,14,15,16]. Thus, the involvement of astrocytes in the pathogenic events in schizophrenia remains to further be established. Besides, a prominent histopathological signature for the activation of microglial cells, in part responsible for neuroinflammation has also increasingly been evident in the brains of schizophrenic subjects [17,18]. The progressive neuroinflammatory process has been known to impair the ongoing neurogenic process in the brain [19,20,21]. Ample experimental evidence suggests dysregulation of adult neurogenesis, is a prominent pathogenic characteristic of various neurodegenerative, mood, and psychiatric disorders that include Alzheimer’s disease (AD), Huntington’s disease (HD), Parkinson’s disease (PD), stress, depression, anxiety, and schizophrenia [20,22,23,24,25,26]. While the progressive decline in hippocampal neurogenesis contributes to the pathomechanisms of dementia and mood disorders, ample experimental evidence demonstrated the occurrence of reactive neurogenesis in the early phases of many neurodegenerative and psychiatric disorders that include AD, PD, HD, and schizophrenia [27,28,29,30,31], Neuroblasts have been considered as immature neurons, derived from the neuronal stem cells of the developing as well as the adult brain [32,33]. The amount of neuroblasts produced in the brain determines the degree of terminal neurogenic process in the adult brain accounting for various neurogenerative plasticity and brain repair [32]. Recently, a new line of emerging scientific evidence underpins an abnormal activation profile of neural cells namely reactive neuroblostosis that depicts the overproduction of immature neurons leading to aberrant neurogenesis in the early phase of many neurodegenerative disorders that display movement disorders and dementia including AD and HD [27,33,34,35,36]. As these neurodegenerative disorders progress into the later stages, the neuroblastosis events appear to be diminished due to the depletion of neural precursor cells or degeneration of the neuroblasts [33,34,35,37]. Notably, traumatic brain injury, cerebral stroke, and epileptic seizure have also been characterized by reactive neuroblostosis and subsequent abnormal migration of mitotically active neuroblasts in the affected regions of the brain [36,38,39,40]. However, the role of reactive neuroblastosis in the pathogenic process or brain repair remains ambiguous. As the ultimate cell fate of reactive neuroblasts is uncertain, reports on their possible involvement in the disease progression, and cognitive, and psychiatric alterations are limited. To note, the fluctuation in the regulation of neurogenesis resulting from reactive neuroblasts in the adult brain could alter and deteriorate the neuroplasticity responsible for mental health and behavior including memory and mood.
Risk factors and etiopathological relevance of schizophrenia
The clinical manifestations of schizophrenia appear to originate from multifactorial elements including some definitive and mostly unknown gene mutations, copy number variations, epigenetic alterations, dysregulated transcriptomics, chromosomal abnormalities, metabolic defects, abnormal brain development, synaptic dysfunctions, aberrant neurotransmission, abnormal lifestyle, environmental factors [2,41]. Notably, maternal malnutrition, preeclampsia, gestational diabetes, prenatal viral infections, vitamin D deficiency, twin gestation, emergency cesarean section, childbirth complications, birth during the winter season, low birth weight, autoimmune diseases, chronic mood disorders, asphyxia, air pollution, illiteracy, and substance abuse, living in an urban area, immigration to the foreign country, living in unsuitable environment and transcultural influences have been identified as the key risk factors for schizophrenia [2,42,43,44,45]. Thus, the etiology and risk factors of schizophrenia are highly multifaceted. While the shreds of evidence for pathogenic signatures of neurodegeneration in schizophrenia-affected brains are indecisive, the neurodevelopmental hypothesis has been considered in which early neurogenic defects and aberrant migration of neuronal precursor cells have been emphasized as underlying psychotic symptoms and cognitive deficits at the later stage of life accounting for schizoaffective disorders [46]. Embryonic stem cells derived from early neurogenesis through the generation of neuroblast cells are an important aspect of brain development during embryogenesis [47]. The abnormal in-utero condition affecting brain development at the level of neuroblast formation and migration has been considered a prime risk factor for schizophrenia [48,49]. Eventually, several theories have been postulated for the neuropathogenic basis of schizophrenia. Disruption of glutamate transmission in the thalamocortical areas has been linked to the development of schizophrenia. Various experimental evidence gathered from the use of anesthetic agents namely phencyclidine and ketamine suggest that defects in the expression and function of glutamate decarboxylase (GAD)-1, hypofunction glutamate, and N-methyl-D-aspartate (NMDA) receptors are associated with the development of schizophrenia [50,51]. While glial cells are important for the neurotransmission of glutamate at the synapses, abnormal gliogenic events during brain development have also been proposed to be involved in the progression of schizophrenia [52]. Further, unusual flux in the release of dopamine and differential expression of its receptors in mesolimbic areas, nigrostriatal, and mesocortical tracts have also strongly been coupled to the symptomatic signature of schizophrenia [53]. As increased release and hyper-transmission of dopamine in the subcortical area of the brain is known to contribute to positive symptoms such as hallucinations and delusions in schizophrenia, its hypofunction resulting from decreased expression or inactivation of dopamine receptors in the prefrontal cortex and caudate nucleus appears to be associated with the development of negative symptoms like anhedonia, lack of motivation, and speech disorders [53,54]. Experimental studies established from the use of antipsychotic drugs that modulate the serotonergic and dopaminergic systems revealed impaired interaction between dopamine and serotonin could prime the abnormal neurochemical events accounting for schizophrenia [55,56,57]. Furthermore, recent evidence indicates the dysfunctions of GABAergic neurons in the cortex, altered levels of serotonin, and defects in the cholinergic system of the brain during the symptomatic phase of schizophrenia [11,41,58,59]. Recently, increased level of norepinephrine has also been suggested to play a role in the pathophysiology of schizophrenia [60].
Eventually, schizophrenia has a heritable nature as many genetic determinants have been linked to its pathogenesis [61]. The first-degree relatives and offspring of subjects with schizophrenia pose a considerable degree of risk of developing the clinical symptoms [42,61]. However, the genetic linkages and the mutations are not unique among schizophrenia patients. The clinical episodes of schizophrenia have been mapped to various polymorphisms or dysregulation of susceptibility genes such as 1) neuregulin (NGR)-1, a candidate gene involved in brain development, vesicular transport of glutamate and EGF signaling, 2) dystrobrevin-binding protein (DTNBP)-1 which aids in glutamate release, 3) catecholamine O-methyl transferase (COMT), important for signal transduction of dopamine, 4) dopamine beta-hydroxylase (DBH) that catalyzes the hydroxylation of dopamine and some phenylethylamine derivatives and 5) regulator of g-protein signaling (RGS)-9 responsible for various molecular pathways transduction in the brain and 6) the disrupted-in-schizophrenia 1 (DISC1) that is known to interact with factors responsible downstream dopamine signaling pathway and glycogen synthase kinase-3 (GSK-3) [62,63,64,65,66,67]. Eventually, the suicidal behaviors in schizophrenia have been attributed to defects in genes such as the corticotropin-releasing hormone receptor (CRHR)-1 and corticotropin-releasing hormone binding protein (CRHBP) that encode stress response elements involved in the regulation of the hypothalamic-pituitary-adrenal (HPA)-axis [68]. Notably, the aforementioned neurobiochemical and genetic determinants and risk factors appear to be associated with morphological differences and neuroanatomical abnormalities in schizophrenia.
Neuro morphological and pathological sequelae of schizophrenia
Owing to its obvious abnormalities in neurotransmission, there has been overwhelming data available for the description of the behavioral deformities and psychotic symptoms in schizophrenia [69]. However, the distinct neuropathological changes of schizophrenia arising from various idiopathic factors have long been refractory to the definitive diagnosis. Earlier radiology-based pneumoencephalography attempts revealed dilated lateral and third ventricles in the brains of subjects with schizophrenia [70,71]. Later on, Johnstone et al validated the enlarged brain ventricles using axial brain scans in schizophrenic brains [72]. As considerable scientific and technological advancements made in recent decades, neuroimaging techniques, and neuromorphometric assessments have revealed anatomical, cytoarchitectural alteration, and functional defects in the brains of subjects with and at risk of schizophrenia. In the quest to address the neuropathological changes in the brain of schizophrenia, ample neuroimaging evidence obtained from computed tomography (CT), magnetic resonance imaging (MRI) involved diffusion-tensor imaging (DTI) tractography, and functional imaging, magnetic resonance spectroscopy (MRS), magnetoencephalography (MEG) and positron emission tomography (PET)-based studies convincingly demonstrated and validated enlarged ventricles, grey matter loss, structural deformities and loss in corpus callosum, increased volume in basal ganglia, loss of myelination and dysconnectivity of neural network, differences in neurite curvature among key brain areas in schizophrenia [71,73,74,75]. Further, a series of MRI-based reports on gross brain morphometric and gyrification assessments unveiled obvious volumetric reduction in the frontal lobe and temporal lobe in schizophrenia subjects [76]. Concomitantly, decreased density of cortical regions and shrinkage in the amygdala, thalamus, nucleus accumbens, and hippocampus have been established as region-specific morphological defects in schizophrenia [78,79]. A surface-based MRI analysis by Sprooten et al., 2013 intended the widespread cortical thinning, more predominantly in superior frontal, medial parietal, and lateral occipital regions during the early stages of schizophrenia [80]. A resting-state functional magnetic resonance imaging (rs-fcMRI) based analytical study done by Sheffield et al., 2017 in schizophrenic individuals reported that cognitive impairment is linked to loss of functional connectivity within and between fronto-parietal lob and cingulo-opercular networks [81]. Studies from the magnetic resonance spectroscopy and PET on the brains of schizophrenia patients reported that dopamine and GABA systems primarily contribute to the pathophysiology and development of psychotic disorders, which has been attributed to an excitatory-inhibitory imbalance in the cerebello-thalamo-cortical and striato-thalamo-cortical loops, hyperfunction in the mesolimbic dopamine pathway and differences in dopaminergic content in the prefrontal cortex (PFC), anterior cingulate gyrus, and hippocampus [82,83].
The increased volume of cerebrospinal (CSF) and the possibilities of neurodegeneration have been proposed for the enlarged ventricles in schizophrenia [84]. To note, the enlarged ventricle has been established as a prominent neuropathological mark related to many neurodegenerative processes and tissue remodeling in many neurocognitive diseases as the neuroblasts migrate from the subventricular zone (SVZ) to the degenerating sites [85,86]. At the same time, the corpus callosum helps in connecting and coordinating the functions of two hemispheres, dysconnectivity between functional areas has been assumed to result in schizophrenia [87,88]. While the occurrence of neurodegeneration in schizophrenia has been a long-standing subject of examination, the studies of histopathological correlates of the postmortem brain samples from schizophrenia victims revealed synaptic loss rather than neurodegeneration can be responsible for the volume loss in many brain regions [89]. Eventually, demyelination resulting from the degeneration of oligodendrocytes has been predicted to be the reason for white matter lesions in the frontal cortex, hippocampus, and cerebellum in schizophrenia [56,90]. Among various brain regions, the hippocampus has been considered to play a crucial role in neurocognitive functions as it holds a niche for neural stem cells (NSCs) and regenerative capacity [91,92] . Atrophy or dysfunction of the hippocampus has been linked to dementia, mood, and psychotic disorders [20,33,34,35,91,92]. Notably, schizophrenia has been characterized by neuroanatomical, cytoarchitectural, synaptic dissociation, demyelination, microglial activation, and functional abnormalities in association with neuroinflammation in the hippocampus [93,94]. Distinctly, the ongoing neurogenesis mediated by NSCs-derived neuroblasts has been reported to play a key role in regenerative plasticity, neurocognitive regulation, memory, and mood functions [92]. Defects in the hippocampal neurogenic process have been linked to dementia, stress, and depression-related symptoms in various neurological illnesses and traumatic brain injuries [20,34,35,39,40,85,93,95]. Eventually, abnormal neurogenesis appears to be associated with neurodevelopmental disorders like autism [96]. Therefore, the possibilities for the involvement of aberrant neurogenesis in the hippocampus in the establishment of psychotic disorders cannot be excluded. Among various, maladaptive disorders, hallucination appears to be a district psychotic symptom predominantly observed in schizophrenia [97]. While the aging-related progressive cognitive decline has been correlated with a steady decline in hippocampal neurogenesis, recent evidence suggests varying degrees of neurogenesis upon the pathogenic progression in mental deterioration and neurodegenerative illnesses that not only display dementia [98]. Notably, schizophrenia has been characterized by arrest in the maturation of the hippocampus due to elevated levels of cellular and molecular signatures of immature neurons [99,100]. Considering the aforementioned facts, the possibilities for the involvement of the regulation of neurogenesis in the hippocampus along with the disease progression could be highly relevant in schizophrenia. Differential regulation, prolonged dysregulation, and impaired neurogenesis may significantly contribute to dysfunctional information processing leading to both cognitive deficits and hallucinations in schizophrenia. Thus, insight into mechanisms associated with hallucinations and impaired neurogenesis might aid in the emergence of future therapeutics to mitigate the disease progression.
Figure 2.
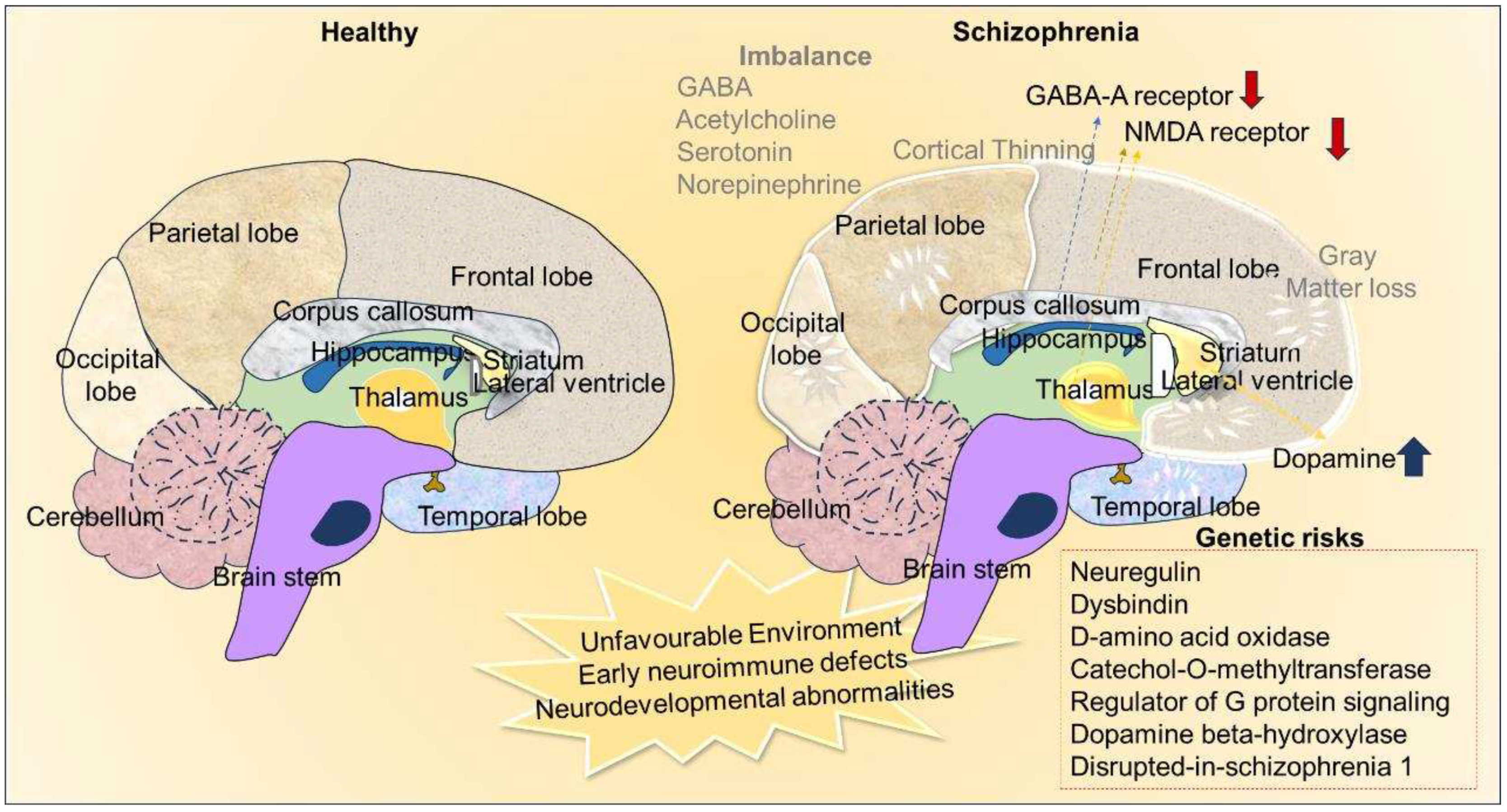
Neurochemical and anatomical differences in healthy and schizophrenia. The image depicts the environmental impact, variations in neurotransmission and hereditary factors as the etiological causes of schizophrenia. Nneuroanatomical alterations such as cortical thinning and loss of grey matter in the schizophrenic brain compare to healthy brains.
Figure 2.
Neurochemical and anatomical differences in healthy and schizophrenia. The image depicts the environmental impact, variations in neurotransmission and hereditary factors as the etiological causes of schizophrenia. Nneuroanatomical alterations such as cortical thinning and loss of grey matter in the schizophrenic brain compare to healthy brains.
Reactive neuroblastosis as an underlying mechanism of hallucination in schizophrenia and other neurological disease
Hallucination is the involuntary generation of illusory, perceptual, and mystical experiences of the brain that occur devoid of external stimuli from the sensory organs and environment [101]. Hallucination can occur at the level of auditory, visual, tactile, olfactory, and gustatory functions during conscious as well as in paradoxical sleeping states. Ample functional neuroimaging studies suggest that the generation of abrupted neural impulses in the key brain areas including the sensory cortex, insula, putamen, and hippocampus could be an underlying cause of hallucination [102,103,104]. However, the origin of the spontaneous neural activity in the brain that synthesizes substrate for the generation of hallucination remains obscure. Thus far, numerous theories have been postulated for the underlying basis of hallucination [105,106]. For example, Lopez Ibor proposed that abnormal activation of engram, a hypothetical form of substratum essential for the cognitive process in ideational centers of the brain, can be an underlying basis of hallucination [105,107]. According to Hughlings Jackson, the upper motor centers of the brain are involved in intellectual ability when spontaneous activities of the foremost motor centers of the brain independent of peripheral reactions or activation of mid-level motor centers upon deactivation of the upper motor center could result in hallucination [108,109]. As per Wilder Penfield’s experimental findings, electrical stimulation in certain cortical or subcortical structures can cause different forms of hallucinations [110,111]. Notably, the occurrence of hallucination is associated with changes in neuroplasticity closely related to key brain regions like primary and secondary sensory cortices, basal ganglia, and limbic system including the hippocampus [112]. Among different neurotransmitter-based hypotheses, varying levels of dopamine in the limbic system have been considered to actively contribute to the development of hallucinations [113]. Recently, abnormal immune activation, increased cerebral blood flow, circulating metabolites, and energy metabolism in the brain have been linked to hallucinations [114,115,116,117]. However, most of the theories on hallucinations appear to be merely paradoxical and none of the concepts has been proven to delineate the definite mechanism of hallucination. Therefore, advancement in the understanding of the root cause of hallucination from different perspectives including at the cellular level has become important.
Though hallucination has been ascertained as a foremost psychotic symptom of schizophrenia, it also occurs in many other diseases and arises in response to some substance abuse [113]. Indeed, hallucination is a key clinical problem in diverged medical conditions including psychiatric complications and neurodegenerative diseases [118]. Notably, various forms of dementia, bipolar disorder, obsessive-compulsive disorder (OCD), epilepsy, cerebral stroke, brain trauma, migraine, brain lesions, brain tumors, metabolic disorders, and Charles bonnet syndrome have been known to be associated with a considerable degree of incidence with hallucination [119]. Mood disorders like anxiety, stress, depression, and post-traumatic stress disorder (PTSD) have also been linked to hallucinations [120]. In AD, reduced acetylcholinesterase activity in the brain has been predicted as a biochemical cause of hallucinations in a significant percentage of subjects with progressive memory loss [121,122]. Eventually, PD and HD patients have also been reported to experience hallucinations due to an imbalance in dopamine and GABA in the limbic system of the brain [123,124]. However, the occurrence of hallucinations in neuropsychiatric and neurodegenerative disorders is a subject of debate as many drugs that are used for the management therapy of neurological deficits, psychotic problems, sleeping abnormalities, and mood disorders have also been known to induce hallucinations [125]. Eventually, prolonged intake of antiepileptic drugs, antidepressants, anticancer medications, and sleeping pills such as narcotics, steroids, pentoxifylline, tramadol, bromocriptine, sertraline, trazodone, appears to cause hallucinations [126,127]. There is no defined biochemical or molecular pathway ascertained for the incidence of hallucinations. Therefore, understanding a specific pathogenic signature in schizophrenia that overlaps with other neurological diseases and the mode of action of hallucinating drugs could provide a valid clue in understanding the underlying basis of the hallucinations.
During embryogenesis, the generation of neurons from embryonic NSCs-derived neuroblasts plays a key role in neurodevelopmental, whereas pathogenic processes in neurodevelopmental disorders and mental disturbances have been attributed to defects in the early neurogenic process [128,129]. Among various predictions, a potential link between schizophrenia and aberrant neurogenic events responsible for unrecognizable abnormal brain development in early life has been widely considered to have a negative impact in the latter adulthood stage leading to psychiatric disturbance and neurocognitive impairments [31,130,131]. Thus, there has been considerable scientific interest in exploring the alteration of neurogenesis in schizophrenia [132]. In the healthy brain, the degree of neuroblasts generation accounting for the neuroregenerative characteristics has been directly correlated with mental health, intellectual ability, sexual drive, pattern separation, and cognitive functions, including learning and memory [91,133,134,135]. The immature and differentiating state of neuroblasts has been characterized by the expression of PSA-NCAM, doublecortin, and calretinin in the adult brain [32,136,137]. Though turnover of neurogenesis in the human brain and animal brain has been reported to be dissimilar at the level of neuroblasts generation, occurrence of neurogenesis in adulthood has been demonstrated in human brains [134,138,139].
In general, understanding the regulation of neurogenesis in the human brain has some drawbacks due to the unavailability of healthy brain samples during the critical period and technical disadvantages [140]. However, the persistence and regulation of neuroblasts in the healthy human brain and aberrant levels of neurogenic process in disease conditions have been unequivocally demonstrated [22,138,140,141]. While impaired neurogenesis during fetal development has been linked to intellectual disability disorders, the progressive decline in the neuroblast population followed by the loss of new neurons in the hippocampus has been established as a brain aging and distinct neuropathogenic event along with progressive memory loss [35,91,98,134,135,142,143]. However, the role of aberrant neurogenesis in pathogenic mechanism in neuropsychiatric and neurodegenderative disorders has been less explored. Notably, neurological diseases like epilepsy and cerebral stroke are associated with increased neurogenesis in the hippocampus, unusual migration of neuroblasts in the cortex and striatum, and altered cell fate events in the neurogenic and non-neurogenic areas [36,39]. Notably, a significant percentage of subjects with epileptic seizures and cerebral ischemia have been reported to experience hallucination [144,145,146]. Besides, physical exercise in a physiological state has been known to enhance cognitive ability via the brain-derived neurotrophic factor (BDNF) mediated neurogenesis [147]. However, aggressive physical exercise appears to exacerbate the disease progression in some neurodegenerative diseases and some individuals experience hallucination after vigorous physical activities [148]. Interestingly, elevated levels of BDNF induced by physical exercise, enriched environment, and thorough supplement of neuroprotective agents have been established to facilitate hippocampal neurogenesis as it promotes the proliferation and survival of neuroblasts. Moreover, schizophrenic brains have been characterized by increased level of BDNF which might in part contribute to increased neurogenic events accounting for hallucinations [149].
Interestingly, increased level of neuroblasts proliferation and their ectopic migration has also become increasingly evident in the early phase of neurodegenerative disorders, while the late phase of these disorders with the prominent sign of dementia and depression-related disease have been characterized by diminished level of neuroblasts in the brain [20,33,35,37,96,150]. Notably, the ongoing turnover of neuroblasts has generally been confined to the hippocampus and SVZ-OB system, several reports indicate the possibility for the occurrence and migration of neuroblasts in the other brain regions including the cortex, striatum, and hypothalamus [33,37,151,152,153]. The presence of astroglial cells in the non-neurogenic areas of the brain has been regarded as an alternate source of NSCs responsible for neurogenic events in different areas of the brain [154]. Recently, the sustaining of non-newly generated terminally undifferentiated neuroblasts with differential neuroplastic nature has also been reported in the brains of some mammals such as dolphins and sheep [155,156]. These quiescent neuroblasts are likely to be activated upon environmental stimuli or during disease progression [20,157]. In disease conditions, the abnormal discharge of proinflammatory cytokine from activated immune cells leads to a cell cycle arrest in NSCs which can cause the stimulation of proliferation in neuroblasts as a compensatory cellular effect [20,35]. Recently, based on the immunohistological findings of the DCX staining in brains of the transgenic animal models of HD, Kandasamy and Aigner proposed a concept known as reactive neruoblastosis which describes an abnormal cellular condition of the brain in which neuroblasts undergo enormous proliferation and ectopic migration in the non-classical neurogenic areas [33,34]. These abnormal cell cycle events have been demonstrated to occur as a result of elevated levels of TGF-β mediated induced quiescent of NSCs as well as neuronal differentiation, owing to its pleiotropic properties [20,35,158]. Besides, Kandasamy and colleagues conceptualized an alternative hypothesis that reactive neuroblastosis might also occur due to the dedifferentiation of neurons in neurodegenerative conditions [150]. Likewise, the level of TGF-β has been reported to be increased during the disease course of schizophrenia [159]. This indicates the possibility of the occurrence of reactive neuroblastosis in schizophrenia. Notably, postmortem studies have revealed reduced cell proliferation by means of a lesser count of Ki 67 and PCNA immunopositive cells in the hippocampus of schizophrenia victims [160]. However, the reduction in the overall cell proliferation did not reflect the number of neuroblasts in the neurogenic area of the brain, as D Barbeau et al., reported that reduction in the number of PSA-NCAM-immunoreactive neuroblasts was confined in the hilar region but not in the DG of brains of schizophrenic subjects [161]. Notably, N M Walton et al in 2012 reported the transition state of immature neurons in the hippocampal DG of CaMKIIα-hKO mice that display behavior abnormalities similar to schizophrenia and bipolar disorder [162,163]. This report also points towards the possibilities for the reactive neuroblastosis in schizophrenia. Interestingly, a recent report from Joen-Rong Sheu et al in 2019 revealed an increased number of DCX-expressing neuroblasts with enhanced dendritic arborization, indicating the surplus amount of interacting neuroblasts in the circuit of the hippocampus in the brains of the maternal immune-activated rodent model of schizophrenia [31]. In corroboration with experimental animal study, they demonstrated an increased level of calretinin-positive cells in the hippocampal DG of postmortem brains of schizophrenia [31]. Eventually, a large number of postmortem data indicated an increased density of interstitial white matter neurons (IWMNs) in the brains with schizophrenia, which could have been derived from the enhanced production of neuroblasts [164]. However, Feng et al indicated the there is no change mRNA expression of neuroblast marker in the schizophrenic brains compared to age matched controls that indicates a compensatory cellular mechanism upon ageing process accounting for interstitial white matter neurons enhanced [165].
Moreover, the treatment of experimental animals with M108, a potent halogenic compound resulted in increased levels of hippocampal neurogenesis and aggravated the symptoms of schizophrenia [31]. In contrast, suppression of the neurogenic process during the adolescent period delayed the onset and progression of schizophrenia-like symptoms in experimental rats [31]. Also, Ketamine and phencyclidine have been considered as potent hallucinating drugs [166]. Multiple lines of experimental evidence suggest that these hallucination drugs increase hippocampal neurogenesis in the hippocampus. Experimental studies revealed that ketamine treatment increases the number of neuroblasts in the hippocampus [167]. Though repeated phencyclidine treatment has been reported to decrease the survival of neurons, a week later repopulation of the compensatory neurons was evident [168,169]. Besides, ample evidence suggested increased proliferative events in the neuroepithelium of schizophrenia patients, compared to explants from healthy controls [170]. Hong, S. et al. provide experimental evidence that PARP-1 is required for the differentiation of NSCs, while its absence results in defective neurogenesis and behavioral impairments leading to schizophrenia-like phenotype in experimental animal models [171]. Eventually, altered levels of Wnt signaling components that induce cell cycle events are evident in schizophrenia [172]. A recent study conducted using iPSC reported altered Wnt-1 signaling activity, a potent inducer of cell cycle in association with abnormal NPC proliferation and imbalanced differentiation of excitatory and inhibitory neurons leading to neuronal circuit miswiring as the developmental origin of schizophrenia [173]. Moreover, subjects with brain metastatic conditions, a state with the malignant proliferation of neuroblasts and glial cells have been reported to display hallucinations [174]. Therefore. it can be strongly argued that reactive proliferation and dendritic growth of neuroblasts contribute to the pathophysiological of hallucination in schizophrenia. Arnold et al, 2001, conducted a study using olfactory epithelium (OE), a structure that continues to undergo neurogenesis and regulation throughout adult life. In his study, he reported the presence of neurons at different stages of development in the post-mortem OE samples, and an increase in the number of immature neurons was evident in schizophrenic patients [175]. In order to survive, the generation of neuroblasts has to be integrated into existing circuits as failure in the integration of neuroblasts would result in their extinction [176]. Therefore, it can be expected that an increased amount of neuroblast generation would form redundant synapses with existing neurons in a competitive nature. Strikingly, neuroblasts have been considered a potential source of the engram, possess electrical properties, and exhibit synoptical activities even in an immature state [33,34,177]. A surplus number of immature neurons resulting from reactive neuroblostosis may tend to integrate into the existing neuronal circuit or may form an extra neural connection in a provocative manner leading to abnormal synoptical, and metabolic activation in the brain thereby drastically misshaping the synapse and electrical events, and creating dysregulation in the release of neurotransmission in the cognitive centers of the diseased brain. Therefore, ultrastructural findings reported the abnormal formation of synapses in schizophrenia patients could be a result of neuroblastosis events leading to miswiring or abnormal patterns of synaptic connections in the brain. The abnormal neurogenic process at the level of integrating an exceeding number of neuroblasts can be proposed to induce abnormal maladaptive psychotic behaviors specifically hallucination. Whereas failure or defects in the integration of a subset of neuroblasts and their consequent loss could be associated with dementia regardless of neurodegeneration of existing mature neurons in schizophrenia.
Figure 3.
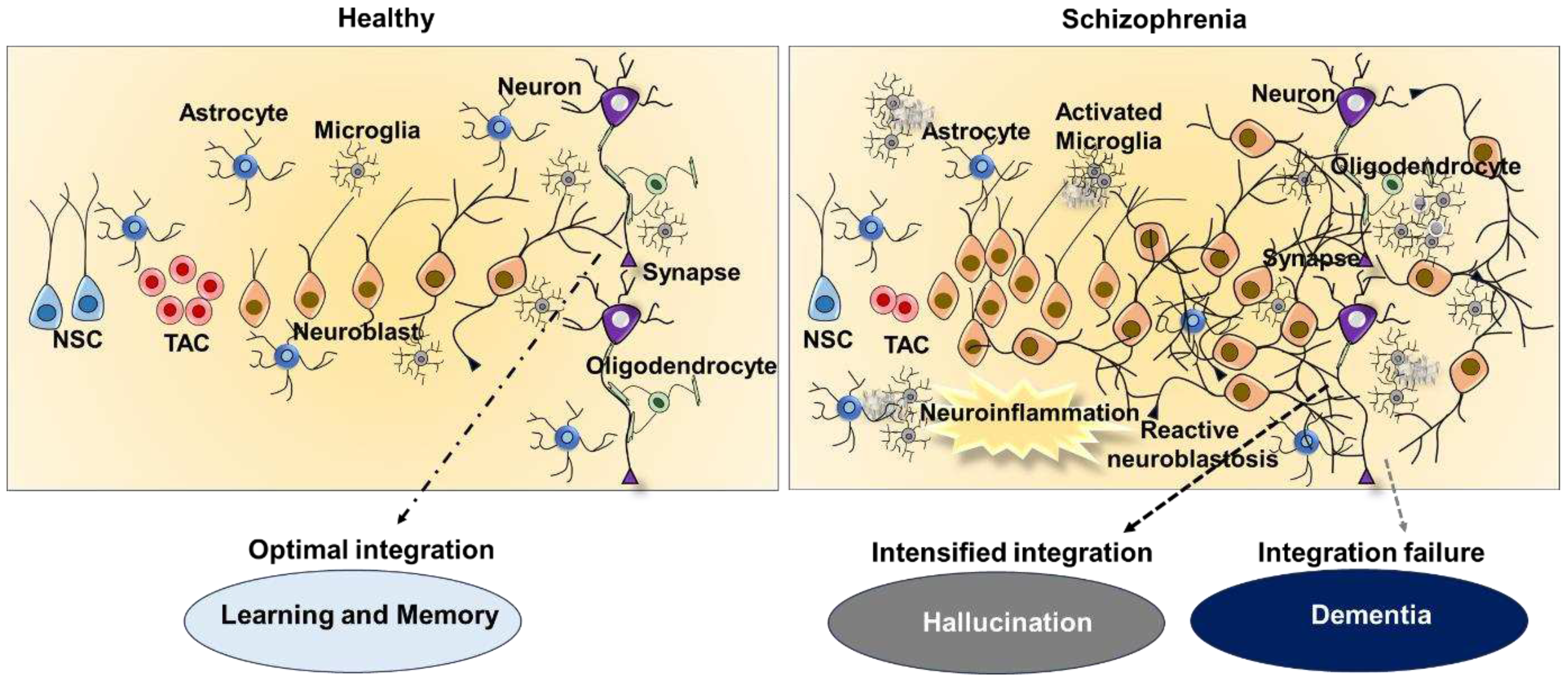
Regulation of adult neurogenesis and reactive neuroblastosis in heathy and schizophrenia brain. The picture provides an overview on the regulation of adult neurogenesis and neuronal integrity in a healthy brain, as well as abnormal neurogenesis and neural integration in schizophrenia. While physiological adult neurogenesis contributes to brain plasticity responsible for cognition function, defective neurogenesis brought on by reactive neuroblastosis causes disruption in neuronal integration and neurotransmission in schizophrenia. Increased neurotransmission from enhanced integration of reactive neuroblasts from could be an underling cause of hallucinations, while decreased neural integration could results in dementia.
Figure 3.
Regulation of adult neurogenesis and reactive neuroblastosis in heathy and schizophrenia brain. The picture provides an overview on the regulation of adult neurogenesis and neuronal integrity in a healthy brain, as well as abnormal neurogenesis and neural integration in schizophrenia. While physiological adult neurogenesis contributes to brain plasticity responsible for cognition function, defective neurogenesis brought on by reactive neuroblastosis causes disruption in neuronal integration and neurotransmission in schizophrenia. Increased neurotransmission from enhanced integration of reactive neuroblasts from could be an underling cause of hallucinations, while decreased neural integration could results in dementia.
Conclusions
The brain is an irreplaceable organ of complex structure that synthesizes dreams, imagination, innovative ideas, and various forms of experiences. Highly structured and regulated forms of neuroplasticity resulting from voluntary and involuntary signals are crucial for motor, physiological, and cognitive performance. Various functional regions of the brain have a great capacity to spontaneously produce neuroplasticity independent of internal and external stimuli. The key functional areas of the brain have the ability to regenerate throughout life. Regulation of NSCs-mediated adult neurogenesis through the generation of neuroblasts has been known to be an integral mechanism for various forms of cognitive functions including intellectual measures, learning, and memory. While various factors can modulate neurogenesis from development to the adult stages, defects in neurogenesis at the level of NSC proliferation and differentiation are associated with various diseases. While known or unknown mutations may alter the cell cycle parameters of the NSC and neuroblasts prolonged activation of immune cells can interfere with their neuronal differentiation and integration process through the proinflammatory and neurotropic factors. Notably, schizophrenia has been linked to an aberrant neurogenic process in the brain. Hallucination is the prime behavioral pathology of schizophrenia and subjects with neurological diseases and intake of some drugs also experience Hallucination for which no confined mechanism has been established. This article emphasizes that reactive neuroblastosis might be responsible for the occurrence of hallucinations as it can abruptly strengthen the synapses and induce abnormal neurotransmission. Besides, failure in the integration of newly generated neuroblasts could contribute to dementia as evidence for neurodegeneration is limited in schizophrenia. Therefore, the futuristic therapeutic targets aimed at harnessing reactive neuroblastosis and correcting the functional integration might be highly beneficial against hallucination and dementia in schizophrenia and other diseases.
Author Contributions
M.K. conceived the idea, and hypothesis and contributed to the framework of the manuscript and illustrations. M.P.B.D.I, J.H.M.J and M.K wrote the initial draft. All contributed to the revision of the article and made critical comments and suggestions. All authors have read and agreed with this version of the manuscript for submission.
Acknowledgment
MK would like to acknowledge and dedicate this article to the late Palani Murugan Rangasamy, senior scientist, centre for cellular and molecular biology (CCMB), Hyderabad, Telangana, India for inspiration. MK has been supported by the University Grants Commission-Faculty Recharge Programme (UGC-FRP), New Delhi, India. The authors also thank RUSA 2.0, Biological Sciences, Bharathidasan University, for their financial support (TN RUSA: 311/RUSA (2.0)/2018 dt. 2 December 2020).
References
- Patel, K.R.; Cherian, J.; Gohil, K.; Atkinson, D. Schizophrenia: Overview and Treatment Options. P T 2014, 39, 638–645. [Google Scholar]
- Hany, M.; Rehman, B.; Azhar, Y.; Chapman, J. Schizophrenia. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2023. [Google Scholar]
- Zubin, J.; Spring, B. Vulnerability: A New View of Schizophrenia. Journal of Abnormal Psychology 1977, 86, 103–126. [Google Scholar] [CrossRef]
- Kendler, K.S. Kraepelin’s Final Views on Dementia Praecox. Schizophr Bull 2020, 47, 635–643. [Google Scholar] [CrossRef]
- Ashok, A.H.; Baugh, J.; Yeragani, V.K. Paul Eugen Bleuler and the Origin of the Term Schizophrenia (SCHIZOPRENIEGRUPPE). Indian J Psychiatry 2012, 54, 95–96. [Google Scholar] [CrossRef]
- Schultz, S.H.; North, S.W.; Shields, C.G. Schizophrenia: A Review. Am Fam Physician 2007, 75, 1821–1829. [Google Scholar]
- George, M.; Maheshwari, S.; Chandran, S.; Manohar, J.S.; Sathyanarayana Rao, T.S. Understanding the Schizophrenia Prodrome. Indian J Psychiatry 2017, 59, 505–509. [Google Scholar] [CrossRef]
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. A Systematic Review of the Prevalence of Schizophrenia. PLoS Med 2005, 2, e141. [Google Scholar] [CrossRef]
- Cederlöf, M.; Lichtenstein, P.; Larsson, H.; Boman, M.; Rück, C.; Landén, M.; Mataix-Cols, D. Obsessive-Compulsive Disorder, Psychosis, and Bipolarity: A Longitudinal Cohort and Multigenerational Family Study. Schizophr Bull 2015, 41, 1076–1083. [Google Scholar] [CrossRef]
- Etchecopar-Etchart, D.; Korchia, T.; Loundou, A.; Llorca, P.-M.; Auquier, P.; Lançon, C.; Boyer, L.; Fond, G. Comorbid Major Depressive Disorder in Schizophrenia: A Systematic Review and Meta-Analysis. Schizophr Bull 2020, 47, 298–308. [Google Scholar] [CrossRef]
- Stępnicki, P.; Kondej, M.; Kaczor, A.A. Current Concepts and Treatments of Schizophrenia. Molecules 2018, 23, 2087. [Google Scholar] [CrossRef]
- Stroup, T.S.; Gray, N. Management of Common Adverse Effects of Antipsychotic Medications. World Psychiatry 2018, 17, 341–356. [Google Scholar] [CrossRef]
- Kim, R.; Healey, K.L.; Sepulveda-Orengo, M.T.; Reissner, K.J. Astroglial Correlates of Neuropsychiatric Disease: From Astrocytopathy to Astrogliosis. Prog Neuropsychopharmacol Biol Psychiatry 2018, 87, 126–146. [Google Scholar] [CrossRef]
- Abazyan, S.; Yang, E.J.; Abazyan, B.; Xia, M.; Yang, C.; Rojas, C.; Slusher, B.; Sattler, R.; Pletnikov, M. Mutant Disrupted-In-Schizophrenia 1 in Astrocytes: Focus on Glutamate Metabolism. J Neurosci Res 2014, 92, 1659–1668. [Google Scholar] [CrossRef]
- Kolomeets, N.S. [Astroglia of the hippocampus in schizophrenia]. Zh Nevrol Psikhiatr Im S S Korsakova 2008, 108, 70–76. [Google Scholar]
- Feresten, A.H.; Barakauskas, V.; Ypsilanti, A.; Barr, A.M.; Beasley, C.L. Increased Expression of Glial Fibrillary Acidic Protein in Prefrontal Cortex in Psychotic Illness. Schizophr Res 2013, 150, 252–257. [Google Scholar] [CrossRef]
- Laskaris, L.E.; Di Biase, M.A.; Everall, I.; Chana, G.; Christopoulos, A.; Skafidas, E.; Cropley, V.L.; Pantelis, C. Microglial Activation and Progressive Brain Changes in Schizophrenia. Br J Pharmacol 2016, 173, 666–680. [Google Scholar] [CrossRef]
- Vallée, A. Neuroinflammation in Schizophrenia: The Key Role of the WNT/β-Catenin Pathway. Int J Mol Sci 2022, 23, 2810. [Google Scholar] [CrossRef]
- Radhakrishnan, R.K.; Kandasamy, M. SARS-CoV-2-Mediated Neuropathogenesis, Deterioration of Hippocampal Neurogenesis and Dementia. Am J Alzheimers Dis Other Demen 2022, 37, 15333175221078418. [Google Scholar] [CrossRef]
- Kandasamy, M.; Couillard-Despres, S.; Raber, K.A.; Stephan, M.; Lehner, B.; Winner, B.; Kohl, Z.; Rivera, F.J.; Nguyen, H.P.; Riess, O.; et al. Stem Cell Quiescence in the Hippocampal Neurogenic Niche Is Associated with Elevated Transforming Growth Factor-Beta Signaling in an Animal Model of Huntington Disease. J Neuropathol Exp Neurol 2010, 69, 717–728. [Google Scholar] [CrossRef]
- Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Brain Inflammation and Adult Neurogenesis: The Dual Role of Microglia. Neuroscience 2009, 158, 1021–1029. [Google Scholar] [CrossRef]
- Moreno-Jiménez, E.P.; Flor-García, M.; Terreros-Roncal, J.; Rábano, A.; Cafini, F.; Pallas-Bazarra, N.; Ávila, J.; Llorens-Martín, M. Adult Hippocampal Neurogenesis Is Abundant in Neurologically Healthy Subjects and Drops Sharply in Patients with Alzheimer’s Disease. Nat Med 2019, 25, 554–560. [Google Scholar] [CrossRef]
- Marxreiter, F.; Regensburger, M.; Winkler, J. Adult Neurogenesis in Parkinson’s Disease. Cell Mol Life Sci 2013, 70, 459–473. [Google Scholar] [CrossRef]
- Winner, B.; Kohl, Z.; Gage, F.H. Neurodegenerative Disease and Adult Neurogenesis. Eur J Neurosci 2011, 33, 1139–1151. [Google Scholar] [CrossRef]
- Eisch, A.J.; Cameron, H.A.; Encinas, J.M.; Meltzer, L.A.; Ming, G.-L.; Overstreet-Wadiche, L.S. Adult Neurogenesis, Mental Health, and Mental Illness: Hope or Hype? J Neurosci 2008, 28, 11785–11791. [Google Scholar] [CrossRef]
- Apple, D.M.; Fonseca, R.S.; Kokovay, E. The Role of Adult Neurogenesis in Psychiatric and Cognitive Disorders. Brain Res 2017, 1655, 270–276. [Google Scholar] [CrossRef]
- Jin, K.; Peel, A.L.; Mao, X.O.; Xie, L.; Cottrell, B.A.; Henshall, D.C.; Greenberg, D.A. Increased Hippocampal Neurogenesis in Alzheimer’s Disease. Proc Natl Acad Sci U S A 2004, 101, 343–347. [Google Scholar] [CrossRef]
- Boekhoorn, K.; Joels, M.; Lucassen, P.J. Increased Proliferation Reflects Glial and Vascular-Associated Changes, but Not Neurogenesis in the Presenile Alzheimer Hippocampus. Neurobiol Dis 2006, 24, 1–14. [Google Scholar] [CrossRef]
- Terreros-Roncal, J.; Moreno-Jiménez, E.P.; Flor-García, M.; Rodríguez-Moreno, C.B.; Trinchero, M.F.; Cafini, F.; Rábano, A.; Llorens-Martín, M. Impact of Neurodegenerative Diseases on Human Adult Hippocampal Neurogenesis. Science 2021, 374, 1106–1113. [Google Scholar] [CrossRef]
- Ruzo, A.; Croft, G.F.; Metzger, J.J.; Galgoczi, S.; Gerber, L.J.; Pellegrini, C.; Wang, H.; Fenner, M.; Tse, S.; Marks, A.; et al. Chromosomal Instability during Neurogenesis in Huntington’s Disease. Development 2018, 145, dev156844. [Google Scholar] [CrossRef]
- Sheu, J.-R.; Hsieh, C.-Y.; Jayakumar, T.; Tseng, M.-F.; Lee, H.-N.; Huang, S.-W.; Manubolu, M.; Yang, C.-H. A Critical Period for the Development of Schizophrenia-Like Pathology by Aberrant Postnatal Neurogenesis. Front Neurosci 2019, 13, 635. [Google Scholar] [CrossRef]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.-G.; Aigner, L. Doublecortin Expression Levels in Adult Brain Reflect Neurogenesis. Eur J Neurosci 2005, 21, 1–14. [Google Scholar] [CrossRef]
- Kandasamy, M.; Aigner, L. Neuroplasticity, Limbic Neuroblastosis and Neuro-Regenerative Disorders. Neural Regen Res 2018, 13, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.; Aigner, L. Reactive Neuroblastosis in Huntington’s Disease: A Putative Therapeutic Target for Striatal Regeneration in the Adult Brain. Front Cell Neurosci 2018, 12, 37. [Google Scholar] [CrossRef]
- Kandasamy, M.; Anusuyadevi, M.; Aigner, K.M.; Unger, M.S.; Kniewallner, K.M.; de Sousa, D.M.B.; Altendorfer, B.; Mrowetz, H.; Bogdahn, U.; Aigner, L. TGF-β Signaling: A Therapeutic Target to Reinstate Regenerative Plasticity in Vascular Dementia? Aging Dis 2020, 11, 828–850. [Google Scholar] [CrossRef] [PubMed]
- Lindvall, O.; Kokaia, Z. Neurogenesis Following Stroke Affecting the Adult Brain. Cold Spring Harb Perspect Biol 2015, 7, a019034. [Google Scholar] [CrossRef]
- Kandasamy, M.; Rosskopf, M.; Wagner, K.; Klein, B.; Couillard-Despres, S.; Reitsamer, H.A.; Stephan, M.; Nguyen, H.P.; Riess, O.; Bogdahn, U.; et al. Reduction in Subventricular Zone-Derived Olfactory Bulb Neurogenesis in a Rat Model of Huntington’s Disease Is Accompanied by Striatal Invasion of Neuroblasts. PLoS One 2015, 10, e0116069. [Google Scholar] [CrossRef]
- Zheng, W.; ZhuGe, Q.; Zhong, M.; Chen, G.; Shao, B.; Wang, H.; Mao, X.; Xie, L.; Jin, K. Neurogenesis in Adult Human Brain after Traumatic Brain Injury. J Neurotrauma 2013, 30, 1872–1880. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, S.; Parent, J.M. Epilepsy and Adult Neurogenesis. Cold Spring Harb Perspect Biol 2015, 7, a020677. [Google Scholar] [CrossRef]
- Roshan, S.A.; Elangovan, G.; Gunaseelan, D.; Jayachandran, S.K.; Kandasamy, M.; Anusuyadevi, M. Pathogenomic Signature and Aberrant Neurogenic Events in Experimental Cerebral Ischemic Stroke: A Neurotranscriptomic-Based Implication for Dementia. J Alzheimers Dis 2023, 94, S289–S308. [Google Scholar] [CrossRef]
- Walker, E.; Kestler, L.; Bollini, A.; Hochman, K.M. Schizophrenia: Etiology and Course. Annu Rev Psychol 2004, 55, 401–430. [Google Scholar] [CrossRef]
- Gejman, P.V.; Sanders, A.R.; Duan, J. The Role of Genetics in the Etiology of Schizophrenia. Psychiatr Clin North Am 2010, 33, 35–66. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, R.A.; Reis Marques, T.; Howes, O.D. Schizophrenia-An Overview. JAMA Psychiatry 2020, 77, 201–210. [Google Scholar] [CrossRef]
- Tandon, R.; Keshavan, M.S.; Nasrallah, H.A. Schizophrenia, “Just the Facts” What We Know in 2008. 2. Epidemiology and Etiology. Schizophr Res 2008, 102, 1–18. [Google Scholar] [CrossRef]
- Stilo, S.A.; Murray, R.M. Non-Genetic Factors in Schizophrenia. Curr Psychiatry Rep 2019, 21, 100. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kulhara, P. What Is Schizophrenia: A Neurodevelopmental or Neurodegenerative Disorder or a Combination of Both? A Critical Analysis. Indian J Psychiatry 2010, 52, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hartenstein, V.; Stollewerk, A. The Evolution of Early Neurogenesis. Dev Cell 2015, 32, 390–407. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Derkits, E.J. Prenatal Infection and Schizophrenia: A Review of Epidemiologic and Translational Studies. Am J Psychiatry 2010, 167, 261–280. [Google Scholar] [CrossRef]
- Muraki, K.; Tanigaki, K. Neuronal Migration Abnormalities and Its Possible Implications for Schizophrenia. Front Neurosci 2015, 9, 74. [Google Scholar] [CrossRef]
- Javitt, D.C. Glutamate and Schizophrenia: Phencyclidine, N-Methyl-D-Aspartate Receptors, and Dopamine-Glutamate Interactions. Int Rev Neurobiol 2007, 78, 69–108. [Google Scholar] [CrossRef]
- Beck, K.; Hindley, G.; Borgan, F.; Ginestet, C.; McCutcheon, R.; Brugger, S.; Driesen, N.; Ranganathan, M.; D’Souza, D.C.; Taylor, M.; et al. Association of Ketamine With Psychiatric Symptoms and Implications for Its Therapeutic Use and for Understanding Schizophrenia: A Systematic Review and Meta-Analysis. JAMA Netw Open 2020, 3, e204693. [Google Scholar] [CrossRef]
- Dietz, A.G.; Goldman, S.A.; Nedergaard, M. Glial Cells in Schizophrenia: A Unified Hypothesis. Lancet Psychiatry 2020, 7, 272–281. [Google Scholar] [CrossRef]
- Brisch, R.; Saniotis, A.; Wolf, R.; Bielau, H.; Bernstein, H.-G.; Steiner, J.; Bogerts, B.; Braun, K.; Jankowski, Z.; Kumaratilake, J.; et al. The Role of Dopamine in Schizophrenia from a Neurobiological and Evolutionary Perspective: Old Fashioned, but Still in Vogue. Front Psychiatry 2014, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.L.; Kahn, R.S.; Ko, G.; Davidson, M. Dopamine in Schizophrenia: A Review and Reconceptualization. Am J Psychiatry 1991, 148, 1474–1486. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.P.; Bozzi, Y. The Role of Dopaminergic and Serotonergic Systems in Neurodevelopmental Disorders: A Focus on Epilepsy and Seizure Susceptibility. Bioimpacts 2015, 5, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Kokkinou, M.; Ashok, A.H.; Howes, O.D. The Effects of Ketamine on Dopaminergic Function: Meta-Analysis and Review of the Implications for Neuropsychiatric Disorders. Mol Psychiatry 2018, 23, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, J.D.; Roth, R.H. The Neuropsychopharmacology of Phencyclidine: From NMDA Receptor Hypofunction to the Dopamine Hypothesis of Schizophrenia. Neuropsychopharmacology 1999, 20, 201–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wong, A.H.C. GABAergic Inhibitory Neurons as Therapeutic Targets for Cognitive Impairment in Schizophrenia. Acta Pharmacol Sin 2018, 39, 733–753. [Google Scholar] [CrossRef] [PubMed]
- Scarr, E.; Gibbons, A.S.; Neo, J.; Udawela, M.; Dean, B. Cholinergic Connectivity: It’s Implications for Psychiatric Disorders. Front Cell Neurosci 2013, 7, 55. [Google Scholar] [CrossRef]
- Mäki-Marttunen, V.; Andreassen, O.A.; Espeseth, T. The Role of Norepinephrine in the Pathophysiology of Schizophrenia. Neurosci Biobehav Rev 2020, 118, 298–314. [Google Scholar] [CrossRef]
- Salleh, M.R. The Genetics of Schizophrenia. Malays J Med Sci 2004, 11, 3–11. [Google Scholar]
- Stefansson, H.; Sigurdsson, E.; Steinthorsdottir, V.; Bjornsdottir, S.; Sigmundsson, T.; Ghosh, S.; Brynjolfsson, J.; Gunnarsdottir, S.; Ivarsson, O.; Chou, T.T.; et al. Neuregulin 1 and Susceptibility to Schizophrenia. Am J Hum Genet 2002, 71, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.M.; Preece, A.; Morris, D.W.; Spurlock, G.; Bray, N.J.; Stephens, M.; Norton, N.; Williams, H.; Clement, M.; Dwyer, S.; et al. Identification in 2 Independent Samples of a Novel Schizophrenia Risk Haplotype of the Dystrobrevin Binding Protein Gene (DTNBP1). Arch Gen Psychiatry 2004, 61, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.-C.; Wassink, T.H.; O’Leary, D.S.; Sheffield, V.C.; Andreasen, N.C. Catechol-O-Methyl Transferase Val158Met Gene Polymorphism in Schizophrenia: Working Memory, Frontal Lobe MRI Morphology and Frontal Cerebral Blood Flow. Mol Psychiatry 2005, 10, 229, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Wise, C.D.; Stein, L. Dopamine-Beta-Hydroxylase Deficits in the Brains of Schizophrenic Patients. Science 1973, 181, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Mirnics, K.; Middleton, F.A.; Stanwood, G.D.; Lewis, D.A.; Levitt, P. Disease-Specific Changes in Regulator of G-Protein Signaling 4 (RGS4) Expression in Schizophrenia. Mol Psychiatry 2001, 6, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, C.A.; Goldman, D.; Jaeger, J.; Persaud, S.; Kane, J.M.; Lipsky, R.H.; Malhotra, A.K. Disrupted in Schizophrenia 1 (DISC1): Association with Schizophrenia, Schizoaffective Disorder, and Bipolar Disorder. The American Journal of Human Genetics 2004, 75, 862–872. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Tharmalingam, S.; Zai, C.; Potapova, N.; Strauss, J.; Vincent, J.; Kennedy, J.L. Association of HPA Axis Genes with Suicidal Behaviour in Schizophrenia. J Psychopharmacol 2010, 24, 677–682. [Google Scholar] [CrossRef]
- Matthysse, S.; Sugarman, J. Neurotransmitter Theories of Schizophrenia. In Handbook of Psychopharmacology: Volume 10: Neuroleptics and Schizophrenia; Iversen, L.L., Iversen, S.D., Snyder, S.H., Eds.; Springer US: Boston, MA, 1978; pp. 221–242. ISBN 978-1-4613-4042-3. [Google Scholar]
- Jacobi, W.; Winkler, H. Encephalographische Studien an chronisch Schizophrenen. Archiv f. Psychiatrie 1927, 81, 299–332. [Google Scholar] [CrossRef]
- Ellis, J.K.; Walker, E.F.; Goldsmith, D.R. Selective Review of Neuroimaging Findings in Youth at Clinical High Risk for Psychosis: On the Path to Biomarkers for Conversion. Front Psychiatry 2020, 11, 567534. [Google Scholar] [CrossRef]
- Johnstone, E.C.; Crow, T.J.; Frith, C.D.; Husband, J.; Kreel, L. Cerebral Ventricular Size and Cognitive Impairment in Chronic Schizophrenia. Lancet 1976, 2, 924–926. [Google Scholar] [CrossRef]
- Vita, A.; De Peri, L.; Deste, G.; Sacchetti, E. Progressive Loss of Cortical Gray Matter in Schizophrenia: A Meta-Analysis and Meta-Regression of Longitudinal MRI Studies. Transl Psychiatry 2012, 2, e190. [Google Scholar] [CrossRef]
- Cropley, V.L.; Klauser, P.; Lenroot, R.K.; Bruggemann, J.; Sundram, S.; Bousman, C.; Pereira, A.; Di Biase, M.A.; Weickert, T.W.; Weickert, C.S.; et al. Accelerated Gray and White Matter Deterioration With Age in Schizophrenia. Am J Psychiatry 2017, 174, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Tovar, M.; Rodríguez-Ramírez, A.M.; Rodríguez-Cárdenas, L.; Sotelo-Ramírez, C.E.; Camarena, B.; Sanabrais-Jiménez, M.A.; Solís-Chagoyán, H.; Argueta, J.; López-Riquelme, G.O. Insights into Myelin Dysfunction in Schizophrenia and Bipolar Disorder. World J Psychiatry 2022, 12, 264–285. [Google Scholar] [CrossRef]
- Turetsky, B.; Cowell, P.E.; Gur, R.C.; Grossman, R.I.; Shtasel, D.L.; Gur, R.E. Frontal and Temporal Lobe Brain Volumes in Schizophrenia. Relationship to Symptoms and Clinical Subtype. Arch Gen Psychiatry 1995, 52, 1061–1070. [Google Scholar] [CrossRef]
- Kaur, A.; Basavanagowda, D.M.; Rathod, B.; Mishra, N.; Fuad, S.; Nosher, S.; Alrashid, Z.A.; Mohan, D.; Heindl, S.E. Structural and Functional Alterations of the Temporal Lobe in Schizophrenia: A Literature Review. Cureus 12, e11177. [CrossRef]
- Jaaro-Peled, H.; Ayhan, Y.; Pletnikov, M.V.; Sawa, A. Review of Pathological Hallmarks of Schizophrenia: Comparison of Genetic Models With Patients and Nongenetic Models. Schizophr Bull 2010, 36, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Glausier, J.R.; Lewis, D.A. Dendritic Spine Pathology in Schizophrenia. Neuroscience 2013, 251, 90–107. [Google Scholar] [CrossRef]
- Sprooten, E.; Papmeyer, M.; Smyth, A.M.; Vincenz, D.; Honold, S.; Conlon, G.A.; Moorhead, T.W.J.; Job, D.; Whalley, H.C.; Hall, J.; et al. Cortical Thickness in First-Episode Schizophrenia Patients and Individuals at High Familial Risk: A Cross-Sectional Comparison. Schizophr Res 2013, 151, 259–264. [Google Scholar] [CrossRef]
- Sheffield, J.M.; Barch, D.M. Cognition and Resting-State Functional Connectivity in Schizophrenia. Neuroscience & Biobehavioral Reviews 2016, 61, 108–120. [Google Scholar] [CrossRef]
- Jiang, S.; Huang, H.; Zhou, J.; Li, H.; Duan, M.; Yao, D.; Luo, C. Progressive Trajectories of Schizophrenia across Symptoms, Genes, and the Brain. BMC Med 2023, 21, 237. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ouyang, P.; Zheng, Y.; Mi, L.; Zhao, J.; Ning, Y.; Guo, W. A Selective Review of the Excitatory-Inhibitory Imbalance in Schizophrenia: Underlying Biology, Genetics, Microcircuits, and Symptoms. Front Cell Dev Biol 2021, 9, 664535. [Google Scholar] [CrossRef]
- Horga, G.; Bernacer, J.; Dusi, N.; Entis, J.; Chu, K.; Hazlett, E.A.; Mehmet Haznedar, M.; Kemether, E.; Byne, W.; Buchsbaum, M.S. Correlations between Ventricular Enlargement and Gray and White Matter Volumes of Cortex, Thalamus, Striatum, and Internal Capsule in Schizophrenia. Eur Arch Psychiatry Clin Neurosci 2011, 261, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, L.G.; Green, A.E.; Babakchanian, S.; Hwang, K.S.; Chou, Y.-Y.; Toga, A.W.; Thompson, P.M. Hippocampal Atrophy and Ventricular Enlargement in Normal Aging, Mild Cognitive Impairment and Alzheimer’s Disease. Alzheimer Dis Assoc Disord 2012, 26, 17–27. [Google Scholar] [CrossRef]
- Mak, E.; Su, L.; Williams, G.B.; Firbank, M.J.; Lawson, R.A.; Yarnall, A.J.; Duncan, G.W.; Mollenhauer, B.; Owen, A.M.; Khoo, T.K.; et al. Longitudinal Whole-Brain Atrophy and Ventricular Enlargement in Nondemented Parkinson’s Disease. Neurobiol Aging 2017, 55, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Stephan, K.E.; Friston, K.J.; Frith, C.D. Dysconnection in Schizophrenia: From Abnormal Synaptic Plasticity to Failures of Self-Monitoring. Schizophr Bull 2009, 35, 509–527. [Google Scholar] [CrossRef]
- Crow, T.J. Schizophrenia as a Transcallosal Misconnection Syndrome. Schizophr Res 1998, 30, 111–114. [Google Scholar] [CrossRef]
- Osimo, E.F.; Beck, K.; Reis Marques, T.; Howes, O.D. Synaptic Loss in Schizophrenia: A Meta-Analysis and Systematic Review of Synaptic Protein and mRNA Measures. Mol Psychiatry 2019, 24, 549–561. [Google Scholar] [CrossRef]
- Najjar, S.; Pearlman, D.M. Neuroinflammation and White Matter Pathology in Schizophrenia: Systematic Review. Schizophrenia Research 2015, 161, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The Role of Adult Hippocampal Neurogenesis in Brain Health and Disease. Mol Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the Adult Hippocampus. Cold Spring Harb Perspect Biol 2015, 7, a018812. [Google Scholar] [CrossRef]
- Lieberman, J.A.; Girgis, R.R.; Brucato, G.; Moore, H.; Provenzano, F.; Kegeles, L.; Javitt, D.; Kantrowitz, J.; Wall, M.M.; Corcoran, C.M.; et al. Hippocampal Dysfunction in the Pathophysiology of Schizophrenia: A Selective Review and Hypothesis for Early Detection and Intervention. Mol Psychiatry 2018, 23, 1764–1772. [Google Scholar] [CrossRef]
- Pujol, N.; Penadés, R.; Junqué, C.; Dinov, I.; Fu, C.H.Y.; Catalán, R.; Ibarretxe-Bilbao, N.; Bargalló, N.; Bernardo, M.; Toga, A.; et al. Hippocampal Abnormalities and Age in Chronic Schizophrenia: Morphometric Study across the Adult Lifespan. Br J Psychiatry 2014, 205, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Heckers, S.; Konradi, C. Hippocampal Neurons in Schizophrenia. J Neural Transm 2002, 109, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Poornimai Abirami, G.P.; Radhakrishnan, R.K.; Johnson, E.; Roshan, S.A.; Yesudhas, A.; Parveen, S.; Biswas, A.; Ravichandran, V.R.; Muthuswamy, A.; Kandasamy, M. The Regulation of Reactive Neuroblastosis, Neuroplasticity, and Nutraceuticals for Effective Management of Autism Spectrum Disorder. Adv Neurobiol 2020, 24, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Polner, B.; Hupuczi, E.; Kéri, S.; Kállai, J. Adaptive and Maladaptive Features of Schizotypy Clusters in a Community Sample. Sci Rep 2021, 11, 16653. [Google Scholar] [CrossRef]
- Babcock, K.R.; Page, J.S.; Fallon, J.R.; Webb, A.E. Adult Hippocampal Neurogenesis in Aging and Alzheimer’s Disease. Stem Cell Reports 2021, 16, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, H.; Takao, K.; Walton, N.M.; Matsumoto, M.; Miyakawa, T. Immature Dentate Gyrus: An Endophenotype of Neuropsychiatric Disorders. Neural Plast 2013, 2013, 318596. [Google Scholar] [CrossRef] [PubMed]
- Tavitian, A.; Song, W.; Schipper, H.M. Dentate Gyrus Immaturity in Schizophrenia. Neuroscientist 2019, 25, 528–547. [Google Scholar] [CrossRef] [PubMed]
- Boksa, P. On the Neurobiology of Hallucinations. J Psychiatry Neurosci 2009, 34, 260–262. [Google Scholar] [PubMed]
- Hare, S.M. Hallucinations: A Functional Network Model of How Sensory Representations Become Selected for Conscious Awareness in Schizophrenia. Front Neurosci 2021, 15, 733038. [Google Scholar] [CrossRef]
- Jardri, R.; Pouchet, A.; Pins, D.; Thomas, P. Cortical Activations during Auditory Verbal Hallucinations in Schizophrenia: A Coordinate-Based Meta-Analysis. Am J Psychiatry 2011, 168, 73–81. [Google Scholar] [CrossRef]
- Wu, J.L.; Haberman, R.P.; Gallagher, M.; Koh, M.T. Probing for Conditioned Hallucinations Through Neural Activation in a Ketamine Mouse Model of Schizophrenia. Neurosci Bull 2020, 36, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Telles-Correia, D.; Moreira, A.L.; Gonçalves, J.S. Hallucinations and Related Concepts—Their Conceptual Background. Front Psychol 2015, 6, 991. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, N.C. Concept of Schizophrenia: Past, Present, and Future. In Schizophrenia; John Wiley & Sons, Ltd, 2010; pp. 1–8. ISBN 978-1-4443-2729-8.
- Ibor, J.J.L. Lecciones de psicologia medica: según apuntes tomados en la cátedra. In Lecciones de psicologia medica: según apuntes tomados en la cátedra; 1964; pp. 427–427.
- Feinberg, I. Corollary Discharge, Hallucinations, and Dreaming. Schizophr Bull 2011, 37, 1–3. [Google Scholar] [CrossRef]
- Jackson, H. The Selected Writings of John Hughlings Jackson. Volume 1. On Epilepsy and Epileptiform Convulsions. Archives of Neurology & Psychiatry 1932, 27, 757. [Google Scholar] [CrossRef]
- Penfield, W.; Boldrey, E. SOMATIC MOTOR AND SENSORY REPRESENTATION IN THE CEREBRAL CORTEX OF MAN AS STUDIED BY ELECTRICAL STIMULATION. Brain 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Catani, M. A Little Man of Some Importance. Brain 2017, 140, 3055–3061. [Google Scholar] [CrossRef] [PubMed]
- Perez-Costas, E.; Melendez-Ferro, M.; Roberts, R.C. BASAL GANGLIA PATHOLOGY IN SCHIZOPHRENIA: DOPAMINE CONNECTIONS and ANOMALIES. J Neurochem 2010, 113, 287–302. [Google Scholar] [CrossRef]
- Kumar, S.; Soren, S.; Chaudhury, S. Hallucinations: Etiology and Clinical Implications. Ind Psychiatry J 2009, 18, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Khandaker, G.M.; Cousins, L.; Deakin, J.; Lennox, B.R.; Yolken, R.; Jones, P.B. Inflammation and Immunity in Schizophrenia: Implications for Pathophysiology and Treatment. Lancet Psychiatry 2015, 2, 258–270. [Google Scholar] [CrossRef] [PubMed]
- McGuire, P.K.; Shah, G.M.; Murray, R.M. Increased Blood Flow in Broca’s Area during Auditory Hallucinations in Schizophrenia. Lancet 1993, 342, 703–706. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, H.; Li, C.; Li, Z.; Li, J.; Li, H.; Dai, L.; Dong, M.; Zhou, J.; He, J.; et al. Metabolite Differences in the Medial Prefrontal Cortex in Schizophrenia Patients with and without Persistent Auditory Verbal Hallucinations: A 1H MRS Study. Transl Psychiatry 2022, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cleghorn, J.M.; Garnett, E.S.; Nahmias, C.; Brown, G.M.; Kaplan, R.D.; Szechtman, H.; Szechtman, B.; Franco, S.; Dermer, S.W.; Cook, P. Regional Brain Metabolism during Auditory Hallucinations in Chronic Schizophrenia. Br J Psychiatry 1990, 157, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S. Hallucinations: Clinical Aspects and Management. Ind Psychiatry J 2010, 19, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Degueure, A.; Fontenot, A.; Husan, A.; Khan, M.W. An Unusual Presentation of Vivid Hallucinations. Cureus 14, e25441. [CrossRef]
- Gottlieb, J.D.; Mueser, K.T.; Rosenberg, S.D.; Xie, H.; Wolfe, R.S. Psychotic Depression, Posttraumatic Stress Disorder, and Engagement in Cognitive-Behavioral Therapy within an Outpatient Sample of Adults with Serious Mental Illness. Compr Psychiatry 2011, 52, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bassiony, M.M.; Lyketsos, C.G. Delusions and Hallucinations in Alzheimer’s Disease: Review of the Brain Decade. Psychosomatics 2003, 44, 388–401. [Google Scholar] [CrossRef]
- Patel, S.S.; Attard, A.; Jacobsen, P.; Shergill, S. Acetylcholinesterase Inhibitors (AChEI’s) for the Treatment of Visual Hallucinations in Schizophrenia: A Review of the Literature. BMC Psychiatry 2010, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, A. Neuropsychiatry of Huntington’s Disease. Dialogues Clin Neurosci 2007, 9, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.; David, A.S. Visual Hallucinations in Parkinson’s Disease: A Review and Phenomenological Survey. J Neurol Neurosurg Psychiatry 2001, 70, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Lafay-Chebassier, C.; Chavant, F.; Favrelière, S.; Pizzoglio, V.; Pérault-Pochat, M.-C. French Association of Regional Pharmacovigilance Centers Drug-Induced Depression: A Case/Non Case Study in the French Pharmacovigilance Database. Therapie 2015, 70, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Niebrzydowska, A.; Grabowski, J. Medication-Induced Psychotic Disorder. A Review of Selected Drugs Side Effects. Psychiatr Danub 2022, 34, 11–18. [Google Scholar] [CrossRef]
- Rolland, B.; Jardri, R.; Amad, A.; Thomas, P.; Cottencin, O.; Bordet, R. Pharmacology of Hallucinations: Several Mechanisms for One Single Symptom? Biomed Res Int 2014, 2014, 307106. [Google Scholar] [CrossRef] [PubMed]
- Taoufik, E.; Kouroupi, G.; Zygogianni, O.; Matsas, R. Synaptic Dysfunction in Neurodegenerative and Neurodevelopmental Diseases: An Overview of Induced Pluripotent Stem-Cell-Based Disease Models. Open Biol 2018, 8, 180138. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.; Iyer, S.; Chatterjee, S.; Kumar, M. In Vivo Models to Study Neurogenesis and Associated Neurodevelopmental Disorders—Microcephaly and Autism Spectrum Disorder. WIREs Mechanisms of Disease 2023, 15, e1603. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Sportelli, V.; Ziller, M.; Spengler, D.; Hoffmann, A. Tracing Early Neurodevelopment in Schizophrenia with Induced Pluripotent Stem Cells. Cells 2018, 7, 140. [Google Scholar] [CrossRef] [PubMed]
- Toro, C.T.; Deakin, J.F.W. Adult Neurogenesis and Schizophrenia: A Window on Abnormal Early Brain Development? Schizophrenia Research 2007, 90, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Iannitelli, A.; Quartini, A.; Tirassa, P.; Bersani, G. Schizophrenia and Neurogenesis: A Stem Cell Approach. Neurosci Biobehav Rev 2017, 80, 414–442. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Drew, M.R. Functional Neurogenesis Over the Years. Behav Brain Res 2020, 382, 112470. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.; Song, H. Adult Neurogenesis in the Mammalian Brain: Significant Answers and Significant Questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef]
- Kempermann, G.; Wiskott, L.; Gage, F.H. Functional Significance of Adult Neurogenesis. Curr Opin Neurobiol 2004, 14, 186–191. [Google Scholar] [CrossRef]
- Bonfanti, L.; Seki, T. The PSA-NCAM-Positive “Immature” Neurons: An Old Discovery Providing New Vistas on Brain Structural Plasticity. Cells 2021, 10, 2542. [Google Scholar] [CrossRef]
- Kumar, A.; Pareek, V.; Faiq, M.A.; Kumar, P.; Kumari, C.; Singh, H.N.; Ghosh, S.K. Transcriptomic Analysis of the Signature of Neurogenesis in Human Hippocampus Suggests Restricted Progenitor Cell Progression Post-Childhood. IBRO Reports 2020, 9, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pareek, V.; Faiq, M.A.; Ghosh, S.K.; Kumari, C. ADULT NEUROGENESIS IN HUMANS: A Review of Basic Concepts, History, Current Research, and Clinical Implications. Innov Clin Neurosci 2019, 16, 30–37. [Google Scholar] [PubMed]
- Amrein, I. Adult Hippocampal Neurogenesis in Natural Populations of Mammals. Cold Spring Harb Perspect Biol 2015, 7, a021295. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Leal-Galicia, P.; Chávez-Hernández, M.E.; Mata, F.; Mata-Luévanos, J.; Rodríguez-Serrano, L.M.; Tapia-de-Jesús, A.; Buenrostro-Jáuregui, M.H. Adult Neurogenesis: A Story Ranging from Controversial New Neurogenic Areas and Human Adult Neurogenesis to Molecular Regulation. Int J Mol Sci 2021, 22, 11489. [Google Scholar] [CrossRef]
- Stagni, F.; Giacomini, A.; Emili, M.; Guidi, S.; Bartesaghi, R. Neurogenesis Impairment: An Early Developmental Defect in Down Syndrome. Free Radic Biol Med 2018, 114, 15–32. [Google Scholar] [CrossRef]
- Guarnieri, F.C.; de Chevigny, A.; Falace, A.; Cardoso, C. Disorders of Neurogenesis and Cortical Development. Dialogues Clin Neurosci 2018, 20, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Stangeland, H.; Orgeta, V.; Bell, V. Poststroke Psychosis: A Systematic Review. J Neurol Neurosurg Psychiatry 2018, 89, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Cascella, N.G.; Schretlen, D.J.; Sawa, A. SCHIZOPHRENIA AND EPILEPSY: IS THERE A SHARED SUSCEPTIBILITY? Neurosci Res 2009, 63, 227–235. [Google Scholar] [CrossRef]
- Kasper, B.S.; Kasper, E.M.; Pauli, E.; Stefan, H. Phenomenology of Hallucinations, Illusions, and Delusions as Part of Seizure Semiology. Epilepsy Behav 2010, 18, 13–23. [Google Scholar] [CrossRef]
- Liu, P.Z.; Nusslock, R. Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front Neurosci 2018, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Lessell, S. Exercise-Induced Visual Hallucinations A Symptom of Occipital Lobe Tumors. Journal of Neuro-Ophthalmology 1988, 8, 81. [Google Scholar]
- Nieto, R.; Kukuljan, M.; Silva, H. BDNF and Schizophrenia: From Neurodevelopment to Neuronal Plasticity, Learning, and Memory. Frontiers in Psychiatry 2013, 4. [Google Scholar] [CrossRef]
- Manickam, N.; Radhakrishnan, R.K.; Vergil Andrews, J.F.; Selvaraj, D.B.; Kandasamy, M. Cell Cycle Re-Entry of Neurons and Reactive Neuroblastosis in Huntington’s Disease: Possibilities for Neural-Glial Transition in the Brain. Life Sci 2020, 263, 118569. [Google Scholar] [CrossRef] [PubMed]
- Bartkowska, K.; Turlejski, K.; Koguc-Sobolewska, P.; Djavadian, R. Adult Neurogenesis in the Mammalian Hypothalamus: Impact of Newly Generated Neurons on Hypothalamic Function. Neuroscience 2023, 515, 83–92. [Google Scholar] [CrossRef]
- Kandasamy, M.; Radhakrishnan, R.K.; Poornimai Abirami, G.P.; Roshan, S.A.; Yesudhas, A.; Balamuthu, K.; Prahalathan, C.; Shanmugaapriya, S.; Moorthy, A.; Essa, M.M.; et al. Possible Existence of the Hypothalamic-Pituitary-Hippocampal (HPH) Axis: A Reciprocal Relationship Between Hippocampal Specific Neuroestradiol Synthesis and Neuroblastosis in Ageing Brains with Special Reference to Menopause and Neurocognitive Disorders. Neurochem Res 2019, 44, 1781–1795. [Google Scholar] [CrossRef]
- Jurkowski, M.P.; Bettio, L.; K. Woo, E.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front Cell Neurosci 2020, 14, 576444. [Google Scholar] [CrossRef]
- Gonzalez-Perez, O.; Quiñones-Hinojosa, A. Astrocytes as Neural Stem Cells in the Adult Brain. J Stem Cells 2012, 7, 181–188. [Google Scholar]
- Parolisi, R.; Cozzi, B.; Bonfanti, L. Humans and Dolphins: Decline and Fall of Adult Neurogenesis. Front Neurosci 2018, 12, 497. [Google Scholar] [CrossRef]
- Piumatti, M.; Palazzo, O.; La Rosa, C.; Crociara, P.; Parolisi, R.; Luzzati, F.; Lévy, F.; Bonfanti, L. Non-Newly Generated, “Immature” Neurons in the Sheep Brain Are Not Restricted to Cerebral Cortex. J Neurosci 2018, 38, 826–842. [Google Scholar] [CrossRef]
- Urbán, N.; Cheung, T.H. Stem Cell Quiescence: The Challenging Path to Activation. Development 2021, 148, dev165084. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.; Lehner, B.; Kraus, S.; Sander, P.R.; Marschallinger, J.; Rivera, F.J.; Trümbach, D.; Ueberham, U.; Reitsamer, H.A.; Strauss, O.; et al. TGF-Beta Signalling in the Adult Neurogenic Niche Promotes Stem Cell Quiescence as Well as Generation of New Neurons. J Cell Mol Med 2014, 18, 1444–1459. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Zhou, Y.; Yan, L.; Xuan, F.; Tong, J.; Li, Y.; Huang, J.; Feng, W.; Chen, S.; Cui, Y.; et al. TGF-Β1 Is Associated with Deficits in Cognition and Cerebral Cortical Thickness in First-Episode Schizophrenia. J Psychiatry Neurosci 2022, 47, E86–E98. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.M.; Fung, S.J.; Shannon Weickert, C. Cell Proliferation Is Reduced in the Hippocampus in Schizophrenia. Aust N Z J Psychiatry 2016, 50, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Barbeau, D.; Liang, J.J.; Robitalille, Y.; Quirion, R.; Srivastava, L.K. Decreased Expression of the Embryonic Form of the Neural Cell Adhesion Molecule in Schizophrenic Brains. Proc Natl Acad Sci U S A 1995, 92, 2785–2789. [Google Scholar] [CrossRef] [PubMed]
- Walton, N.M.; Shin, R.; Tajinda, K.; Heusner, C.L.; Kogan, J.H.; Miyake, S.; Chen, Q.; Tamura, K.; Matsumoto, M. Adult Neurogenesis Transiently Generates Oxidative Stress. PLoS One 2012, 7, e35264. [Google Scholar] [CrossRef] [PubMed]
- Walton, N.M.; Zhou, Y.; Kogan, J.H.; Shin, R.; Webster, M.; Gross, A.K.; Heusner, C.L.; Chen, Q.; Miyake, S.; Tajinda, K.; et al. Detection of an Immature Dentate Gyrus Feature in Human Schizophrenia/Bipolar Patients. Transl Psychiatry 2012, 2, e135. [Google Scholar] [CrossRef] [PubMed]
- Duchatel, R.J.; Weickert, C.S.; Tooney, P.A. White Matter Neuron Biology and Neuropathology in Schizophrenia. NPJ Schizophrenia 2019, 5. [Google Scholar] [CrossRef]
- Fung, S.J.; Joshi, D.; Allen, K.M.; Sivagnanasundaram, S.; Rothmond, D.A.; Saunders, R.; Noble, P.L.; Webster, M.J.; Shannon Weickert, C. Developmental Patterns of Doublecortin Expression and White Matter Neuron Density in the Postnatal Primate Prefrontal Cortex and Schizophrenia. PLoS One 2011, 6, e25194. [Google Scholar] [CrossRef]
- Lodge, D.; Mercier, M.S. Ketamine and Phencyclidine: The Good, the Bad and the Unexpected. Br J Pharmacol 2015, 172, 4254–4276. [Google Scholar] [CrossRef]
- Rawat, R.; Tunc-Ozcan, E.; McGuire, T.L.; Peng, C.-Y.; Kessler, J.A. Ketamine Activates Adult-Born Immature Granule Neurons to Rapidly Alleviate Depression-like Behaviors in Mice. Nat Commun 2022, 13, 2650. [Google Scholar] [CrossRef]
- Wang, C.Z.; Yang, S.F.; Xia, Y.; Johnson, K.M. Postnatal Phencyclidine Administration Selectively Reduces Adult Cortical Parvalbumin-Containing Interneurons. Neuropsychopharmacol 2008, 33, 2442–2455. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Suzuki, T.; Seki, T.; Namba, T.; Tanimura, A.; Arai, H. Effects of Repeated Phencyclidine Administration on Adult Hippocampal Neurogenesis in the Rat. Synapse 2006, 60, 56–68. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, R.D.; Féron, F.; Perry, C.; Chant, D.C.; McLean, D.; Matigian, N.; Hayward, N.K.; McGrath, J.J.; Mackay-Sim, A. Cell Cycle Alterations in Biopsied Olfactory Neuroepithelium in Schizophrenia and Bipolar I Disorder Using Cell Culture and Gene Expression Analyses. Schizophrenia Research 2006, 82, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yi, J.H.; Lee, S.; Park, C.-H.; Ryu, J.H.; Shin, K.S.; Kang, S.J. Defective Neurogenesis and Schizophrenia-like Behavior in PARP-1-Deficient Mice. Cell Death Dis 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hoseth, E.Z.; Krull, F.; Dieset, I.; Mørch, R.H.; Hope, S.; Gardsjord, E.S.; Steen, N.E.; Melle, I.; Brattbakk, H.-R.; Steen, V.M.; et al. Exploring the Wnt Signaling Pathway in Schizophrenia and Bipolar Disorder. Transl Psychiatry 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Topol, A.; Zhu, S.; Tran, N.; Simone, A.; Fang, G.; Brennand, K.J. Altered WNT Signaling in hiPSC NPCs Derived from Four Schizophrenia Patients. Biol Psychiatry 2015, 78, e29–e34. [Google Scholar] [CrossRef]
- Romero-Luna, G.; Mejía-Pérez, S.I.; Ramírez-Cruz, J.; Aguilar-Hidalgo, K.M.; Ocampo-Díaz, K.M.; Moscardini-Martelli, J.; Ramírez-Stubbe, V.; Santellán-Hernández, J.O. Schizophrenia-Like Psychosis Presented in a Patient With a Temporal Lobe Tumor: A Case Report. Cureus 14, e29034. [CrossRef]
- Arnold, S.E.; Han, L.Y.; Moberg, P.J.; Turetsky, B.I.; Gur, R.E.; Trojanowski, J.Q.; Hahn, C.G. Dysregulation of Olfactory Receptor Neuron Lineage in Schizophrenia. Arch Gen Psychiatry 2001, 58, 829–835. [Google Scholar] [CrossRef]
- Shors, T.J.; Anderson, M.L.; Curlik, D.M.; Nokia, M.S. Use It or Lose It: How Neurogenesis Keeps the Brain Fit for Learning. Behav Brain Res 2012, 227, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Baptista, P.; Andrade, J.P. Adult Hippocampal Neurogenesis: Regulation and Possible Functional and Clinical Correlates. Frontiers in Neuroanatomy 2018, 12. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



