Submitted:
04 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
GvHD still remains, despite the continuous improvement of transplantation platforms, a fearful complication of transplantation from allogeneic donors. Being able to separate GvHD from GvL represents the greatest challenge in the allogeneic transplant setting. This may be possible through continuous improvement of cell therapy techniques. In this review, current cell therapies are taken into consideration which are based on the use of TCR alpha/beta depletion, CD45 RA depletion, T regulatory cells enrichment, NK cell-based immunotherapies and suicide gene therapies in order to prevent GvHD and maximally amplify the GvL effect in the setting of haploidentical transplantation.
Keywords:
GvHD
; GvL
; TCR alpha-beta
; CD45RA
; Tregs
; NK
; Suicide gene
; immunotherapies
1. Introduction
Graft-versus-host disease (GvHD) is one of the most serious complications of allogeneic stem cell transplantation [1] and it’s tied up to the action of the T lymphocytes present in the inoculum of stem cells which attack the host's tissues, giving an extremely polymorphic and potentially fatal clinical complication [2]. On the other hand, the T lymphocytes themselves are mainly responsible for the antileukemic effect (i.e. Graft-versus-Leukemia, GvL) associated with the transplant procedure [3]. In the setting of allogeneic transplantation from a haploidentical donor, the first platform for GvHD prevention was based on the profound non-selective T depletion, where all T cells were removed from the hematopoietic stem cell (HSC) inoculum and which, however, compromised the immunological reconstitution and the same GvL effect [4,5]. Since then, different approaches have been proposed, but the separation between GvHD and GvL still remains an open challenge [6,7]. In this review we will summarize the more recent cellular strategies aimed at separating GvHD from GvL by using selective T cell targeting for patients undergoing stem cell transplantation for haematological malignancies.
2. Removal Of αβ T-Cells
The T cell receptor (TCR) is made up of two different chains. The majority (95%) of circulating peripheral blood T lymphocytes expresses αβ chains while the remaining has γδ chains. In the setting of allogeneic HSCT, experimental models have clearly demonstrated how αβ T lymphocytes are implicated in the generation of GvHD. Conversely, γδ T lymphocytes that recognize antigens in an MHC-independent manner are involved in facilitating engraftment, driving immune reconstitution, reducing the risk of opportunistic infections and potentially exerting anti-leukemic effects [8]. Clinical studies have clearly demonstrated that higher number of γδ lymphocytes in the graft correlates with better disease free survival [9,10]. Hence the rational for removal of αβ T cells from the graft with the aim of limiting GvHD and preserving at the same time immunological reconstitution and GvL, thus providing a valid alternative for those patients requiring an urgent allogeneic HSCT but lacking full-matched donors. A large-scale and efficient method was developed for the selective ex vivo depletion of alloreactive αβ T cells and CD19+ B cells from mobilized peripheral blood stem cells and generation of an allogeneic graft enriched for CD34+ stem cells and γδ T lymphocytes [11,12,13]. Moreover, differently from the positive selection of CD34+ cells, procedures of αβ T-cells depletion allowed the sparing of donor-derived NK cells, a subset of cells fundamental in both GVL and control of opportunistic infections [14,15,16,17]. The first applications of αβ T-cell depletion involved pediatric haploidentical HSCT for both malignant and nonmalignant diseases. After removal of αβ T-cells and CD19+ B-cells, haploidentical HSCT proved feasible and effective in children with life-threatening nonmalignant disorders. Four out of 23 patients experienced graft failure that was successfully resolved with re-transplantation, while the remaining children had a rapid hematopoietic recovery. Notably, only 3 children suffered from skin-limited and grade I-II acute GVHD, whereas none of them had neither severe acute nor chronic GvHD. After a median follow up of 18 months, cumulative incidence of transplantation-related mortality was 9.3% and 2-year disease-free survival was 91.1%, comparing favorably with outcomes of HSCT from HLA-matched donors and cord blood unit [18,19]. Reduced incidence of graft failure (14%) was observed in patients with hemoglobinopathies, a setting where engraftment historically represented a relevant obstacle to HSCT [20]. Comparable results between αβ T-cells and CD19+ B-cells depleted MUD and mismatched related donors were also confirmed in a prospective trial enrolling pediatric patients with primary immunodeficiencies [21]. In children with either high-risk or relapsed acute leukemia lacking suitable full-matched donors, an αβ T-cells and B-cells depleted haploidentical HSCT was given following myeloablative conditioning regimen. Anti–T-lymphocyte globulin was given for preventing graft rejection and GvHD, whereas no further GvHD prophylaxis was given post infusion. Among 80 enrolled patients, primary graft failure occurred in only 2 children. At day 100 skin-only and grade I/II acute GvHD occurred in 24 patients (30%), whereas no case of severe and gut/liver acute GvHD was reported. Among patients surviving >100 days after HSCT, CI of chronic GVHD was 5% and all cases were limited in severity. After a median follow up of 46 months, CI of relapse and non-relapse mortality was 24% and 5%, respectively. The 5-year OS and GvDH-free relapse-free survival (GRFS) was 72% and 71%, respectively, comparing favorably with outcomes of HSCT from both MRD and MUD [22]. Similar results following a treosulfan-based conditioning regimen were obtained in patients receiving αβ T-cells and B-cells depleted unrelated as well as haploidentical HSCT [23]. Among pediatric patients with both malignant and nonmalignant disease, αβ T-cells and CD19+ B-cells depletion granted efficient control of opportunistic infections. Although detection of CMV and EBV viremia occurred in 51% and 33% of patients, respectively, the incidence of CMV-associated disease was 6% whereas EBV-related disease involved 0.5% of patients [24]. Moreover, compared to CD34+ selected grafts, those patients who received αβ T-cells and CD19+ B-cells depleted haploidentical HSCT also showed more rapid immune recovery in terms of CD3+, CD19+ and CD56+ counts, with γδ T cells representing the prevalent T-cell subset in the early post-transplant period [8,25]. Notably, a robust recovery of γδ T cells at early timepoints appeared to correlate with decreased risk of CMV infection and leukemia relapse [26]. Following results in children, αβ T-cells and CD19+ B-cells depletion was then employed in adults. Following a conditioning regimen including ATG, treosulfan, fludarabine and thiotepa without further GvHD prophylaxis post transplantation, 59 adult patients with acute leukemia underwent haploidentical HSCT. Only 3 patient (95%) experienced graft failure, whereas rapid full-donor hematological engraftment as well as sustained immune reconstitution were observed among the remaining patients. Grade II/IV acute GvHD was limited to two cases, while two patients developed chronic GvHD. Disease relapse and non-relapse mortality remained the main causes of treatment failure [27]. Similar results were reported in a Turkish experience [28]. More recently, αβ T-cells depletion has been tested even in the setting of HSCT from matched related (MRD) and matched unrelated donor (MUD). In a phase 1/2 prospective trial, 35 adult patients with hematological malignancies underwent peripheral blood derived αβ T-cells depleted allogeneic HSCT. At day 100, cumulative incidence (CI) of grade II-IV and grade III-IV acute GvHD was 26% and 14%, respectively. Rapid immunological reconstitution by NK and γδ T-cells was observed and the majority of patients could also receive DLI. At 2 years, CI of moderate and severe chronic GvHD was 17% and 0%, respectively, while incidence of relapse and non-relapse mortality was 29% and 32% respectively [29]. Procedures of αβ T-cells depletion have been applied for the manipulation of DLI and stem-cell booster for the treatment of poor graft function, mixed chimerism and opportunistic infections after HSCT with limited GvHD and promising results [30]. The clinical trial are summarized in Table 1.
2.1. Removal Of Naive T-Cells (CD45RA T-Cell Subset)
CD45RA, an isoform of the well-known common leukocyte antigen CD45, identifies human naïve T (TN) which are a subtype of T cells that have yet to encounter their antigen, while T cells that previously responded to their antigen, called memory T (TM) cells became CD54RA negative.In preclinical models, it has been observed that TN are responsible for a more severe GVHD than TM , which retain more antipathogen immunity with a graft versus leukemia (GVL) activity [31,32,33,34]. These findings supported the hypothesis that eliminating TN cells (CD45RA+) from the graft could be a potential weapon for dissecting GVHD from GVL and enhance immune reconstitution. Teschner et al. [35] firstly described the depletion of CD45RA+ cells from leukapheresis product of six donors using immunomagnetic beads. Post CD45RA+ depletion, the targeted T cell content was 1 x107 cells/Kg in the graft. This tecnique allowed a TN depletion of median of 4 log. Based on these data, depleted CD45RA+ cells were investigated in the clinical setting either as progenitor cell grafts that in post-transplant setting as donor lymphocyte infusions (DLIs) to enhance immune reconstitution. Different groups reported outcomes of CD45RA+ depleted grafts in matched related, unrelated and haploidentical HCT. In 2015 Bleakley et al. published the results of the first pivotal single arm phase II clinical trial (NCT00914940) [36] including 35 adults with high risk acute leukemia transplanted from a matched sibling donor. Conditioning regimen was myeloablative and GVHD prophylaxis was based on tacrolimus alone. CI of II-IV aGVHD was high resulting 66% but no steroid-refractory aGVHD was observed; only 9% of patients developed cGVHD. 2 years OS was 78% and 2 years DFS was 70%. The 2 years probability of relapse was 21%. Immune reconstitution was rapid and sustained resulting in 2 y NRM of 9%. EBV reactivation and post-HCT lymphoproliferative disease were not observed. More recently, the same group [37] reported the outcomes of 138 (adult and pediatric) patients with acute leukemia and myelodisplastic syndrome treated on 3 different prospective phase II single arm trials (NCT00914940, NCT01858740, NCT02220985) receiving TN-depleted peripheral graft from HLA matched related or unrelated donors. Conditioning was of high intensity for 100 patients and of intermediate intensity for 38 patients with age >50 or comorbidities. GVHD prophylaxis was based on tacrolimus alone for patients that received matched related donor (n= 41) and on tacrolimus plus methotrexate (n=59) or tacrolimus plus mycophenolate mofetil (n=38) in matched unrelated donors receiving high and intermediate conditioning intensity, respectively. CI of grade II aGVHD was 71% and mostly was stage I upper gastrointestinal and steroid responsive; CI of III-IV aGVHD was 4%. 3 years CI of cGVHD occurred in only 7% of patients, but was mostly mild and steroid-responsive. No differences in acute and cGVHD were found according to the donor type. 3 years OS was 77%, cGVHD free and relapse free survival (GRFS) was 68%, CI of relapse was 28% and NRM was 8%.
Overall, these results showed a low incidence in severe acute and cGVHD, without apparent risk of relapse and NRM. One possible explanation of these clinical results is that TN includes a greater frequency minor histocompatibility (H) antigen-reactive T cells, while the TM remaining after the depletion of CD45RA +cells, have a limited TCR repertoire and could potentially recognize minor H antigens to a lower extent [38]. This might generate a sufficient alloresponse to induce limited aGVHD and some GVL but insufficient to initiate or sustain cGVHD. This hypothesis could support the fact that TM favor an alloresponse which lead to limited aGVHD and is not sufficient to generate and or sustain cGVHD [39]. In haploidentical HCT, the adoptive transfer of diverse memory T cell population from the CD45RA+ depleted grafts have been reported by some investigators [40,41] mostly focused on the pediatric population. Naik S et al. [42] reported results of a prospective clinical trial using CD45RA+ depleted haplo transplant followed by donor NK cell addback in 72 pediatric patients with hematological malignancies. All patients received submyeloablative conditioning and GVHD prophylaxis consisted of a short course of Mycophenolate mofetile and/or sirolimus. Patients received CD34+ selected graft at day 0 and a second progenitor graft depleted of CD45RA+ cells; NK cells were infused at day +6. CI of overall aGVHD was 36.1% and cGVHD was 20.8%. 3 years CI of relapse and NRM were 26.5% and 11.5%, despite the majority of patients were transplanted in relapsed refractory setting. Sisinni et al [43] reported outcomes of 25 pediatric patients with acute leukemia who received CD45RA+ deplete T cell grafts after submyeloablative conditioning. GVHD prophylaxis was based on short course of cyclosporine and CI of II-IV aGVHD was 39% and at 30 months CI of cGVHD was 22%. Immune reconstitution was rapid but there was an unexpectedly high rate of HHV6 encephalitis (34% of patients) at a median 35 days after transplant. Some in vitro experiments showed that NK could eliminate HHV6 CD4+ T cells [44], so the same group [45] reported outcomes of 18 patients, incorporating NK cell infusion within 10 days after CD45RA+ depleted transplant. 8 of 18 patients had HHV6 reactivation but none of patients developed HHV6 encephalitis. Despite CD45RA+ deplete T cell grafts provided a feasible transplant platform with a reliable cell processing using widely available commercial technology, some points need to be elucidated especially in the haploidentical setting, as the optimal T cell dose and the GVHD prophylaxis. To date there are two randomized clinical trial ongoing comparing outcomes of naive T cell depleted HCT to T replete transplant platform (NCT03970096 NCT03779854) both in adult and in the pediatric setting. Results will determine whether this approach could really improve the risk of acute and chronic GVHD and survival outcomes compared to the standard HCT platforms. Furthermore, CD45RA+ T cell depletion was also reported in a small series of non-malignant disorders [45,46] but further data are needed in this setting. Importantly, CD45RA+ depleted T cells have also been used in the post-transplant setting as modified donor lymphocyte infusions (DLIs) with the main objective to reduce relapse incidence and to enhance immune reconstitution while preventing GVHD. The use of DLI to treat relapse was investigated by Muffly et al. [47]. They evaluated the feasibility and safety of infusing freshly isolated and purified donor-derived phenotypic CD8+ TM cells into 15 adults with disease relapse after allo-HCT. DLI were given at escalated doses (from 1 × 106 /kg to 10 × 106/Kg) and the majority received chemotherapy before infusion. aGVHD grade II occurred in 1 patient. 67% achieved response for at least 3 months after infusion. Median LFS was 4.9 months and OS was 19 months. Other groups focused the attention mainly on the role of DLI to enhance immune reconstitution and prevent GVHD. Dunaikina et al. [48] evaluated the safety and efficacy of CD45RA+ depleted prophylactic DLI given early after haplo-HCT with αβ T-cell depletion in pediatric patients with acute hematological malignancies. A cohort of 149 children, 76 patients were randomized to receive scheduled DLI and 73 received standard care. The median number of DLI was 4 and the dose was escalated from 25x103/kg up to 50x103/kg. The CI of grade II-IV aGVHD, the incidence of CMV viremia, and survival outcomes were similar in the two groups. The use of DLI was associated with improved recovery of CMV T cell responses in a sub cohort of CMV IgG seropositive recipients. In the same year, Naik et al. [49] reported an interim analysis of a prospective clincal trial (NCT03849651) utilizing escalating doses of CD45RA-depeleted T-cell as addback following TCRαβ/CD19-depleted haplo-HCT to improve immune recovery in 30 children with acute leukemia. Patients with acute lymphoblastic leukemia (ALL) also received prophylactic Blinatumomab following infusion of CD45RA-depeleted T-cell to overcome the risk immune escape secondary to HLA-loss and relapse. Two weeks after engraftment patients received CD45RA+ depleted cell in three escalating doses (starting dose 1 x105/Kg increasing of 1 log for each infusion). At 1 month post infusion, authors described significant expansion of virus specific T cells and and at 6 months TCR repertoire was broad and comparable to that of the donor. The cumulative incidence of aGVHD and grade III-IV aGVHD for the entire cohort was 26.7% and 13.3% respectively, there was no chronic GVHD. In adult setting, Castagna et al. evaluated the role of CD45RA+ depleted DLI after haplo-HCT with post-transplant Cyclophosphamide for patients with hematological malignancies [50]. DLI was delivered in three escalating doses, the median first dose was given at 55 days post-transplant. 16 of 19 patients received all the three planned infusions (starting dose was 5x105/kg up to 5x106/kg). Only 1 patient had development of grade II acute GVHD, and 2 patients had moderate chronic GVHD. 100-day CI of viral infection was reduced (53% vs 32%) from previously published data of the same group. Maung et al. [51] assessed the safety of prophylactic CD45RA+ depleted DLI after reduced intensity conditioning transplant from T replete matched related or unrelated donors in 16 patients with hematological malignancies. The first dose was given at a median of 112 days (starting dose was 1x105/kg increasing 1 log each administration up to 1x107). No dose limiting grade III-IV aGVHD was observed suggesting that prophylactic modified DLI is safe and not associated with increased risk of acute and chronic GVHD. Taken together, this findings are promising but future strategies could include more harmonized procedures and randomized clinical trials to determine whether prophylactic CD45RA+ depleted DLIs can improve immune recovery and reduce infectious complications without increasing the risk of GVHD. The clinical trials are summarized in Table 2.
2.2. Tregs Selection
Regulatory T cells (Tregs) play a critical role in regulating adaptive immunity and maintaining tolerance [52]. Tregs exhibit a CD4+/CD25+ phenotype accounting for 5-10% of the circulating T cells [53] and up to 10% of peripheral blood CD4+ T cells express the CD25 antigen [54]. However, only 1–2% express high levels of CD25 (CD25hi) and have suppressor activity [55]. Tregs can be easily separated from leukapheresis of the donor by immunomagnetic separation (CD19 w/o CD8 depletion followed by CD25 enrichment) resulting in a population ranging from 200 to 400 million cells with the CD4+/CD25+ phenotype. These cells contain a variable amount of CD127+ cells (5-20%) and are strongly enriched in FoxP3 (about 90%) [56,57,58,59]. Another approach used in the setting of Cord Blood transplantation is based on obtaining Tregs from Umbelical Cord Blood (UCB) by CD25 enrichment with a final purity of CD4/CD25 ≥ 60% [60]. In addition to immunomagnetic selection, some groups have also developed selection by GMP-grade cell sorter [61,62], with, however, clinical applications different than haploidentical transplantation. The main transplantation platform based on the use of Tregs cells in haploidentical setting represents the evolution of the T-depleted haploidentical transplantation [5], in which, after a myeloablative conditioning regimen, in addition to the megadose of CD34+ stem cells, 1 million CD3+ cells/kg are infused under the protection of 2 million/kg Tregs [63,64,65] in the absence of post-transplant immunosuppression. Furthermore, a characteristic of this procedure is that the Tregs are infused 4 days before the conventional T cells, on the basis of the experimental model [66] in which it is clearly demonstrated that the prevention of GvHD correlates with the early administration of Tregs. With such a strategy the probability of moderate/severe cGVHD/relapse-free survival was 75% [65]. The mechanisms of action of Tregs in terms of immunosuppression and therefore inhibition of GvHD are multiple and include: -cytokine production such as interleuchin (IL)-10 [67], IL-35 [68] and TGF-beta [69]; -direct killing of Tcons by perforine/granzyme mechanism [67]; -IL-2 competition that cause IL-2 starvation on Tcons [67]; -Tregs/DC interaction with downregulation of CD80/CD86 on DCs via CTLA-4 [70], interference with DC maturation through the LAG-3 molecule highly expressed on Tregs [71], Treg-mediated enhanced expression of indoleamine 2, 3-dioxygenase (IDO) by DCs, which accelerates tryptophan distruption fundamental for the survival of Tcons [72]; adenosine triphosphate (ATP) cleavage by CD39/73 expressed on Tregs wich transform ATP to adenosine, an anti-inflammatory factor [73]. However, how Tregs inhibit GvHD but not GvL has not yet beenfully clarified. Mouse models have demonstrated that Tregs are able to inhibit the onset of GvHD but do not hinder the GvL effect of simultaneously co-infused T lymphocytes [64,74]. The explanation given was that Tregs inhibited the proliferation of T lymphocytes but not their activation, thus resulting capable of lysing the leukemic target [74]. This observation has recently been supported by TCR receptor and transcriptome analyses which confirmed that Tregs do not alter the activation of Tcons thus guaranteeing the GvL effect and revealed as potential GvHD modulating molecules IL-10 and IL35 [75]. Interestingly, both molecules are found significantly increased in GMP expanded Tregs [76]. Tregs cells are also implicated in the mechanisms of inhibition of NOTCH1, which represents a key regulator of alloreactivity [77,78], and in this regard it has been demonstrated that NOTCH1 is down-regulated in Tcons in presence of Tregs through a CD39 dependent mechanism both in vitro and in vivo model [79]. Interestingly, NOTCH1 inhibition, on the one hand blocks GvHD [80], and, on the other hand, does not alter the T-cell-mediated GvL effect [81]. STAT3 deficiency and PD1 signaling are both important for the prevention of GvHD in target organs while, where this axis functions less, i.e. in lympho-hematopoietic tissues, T cell proliferation is maintained with a consequent powerful GvL effect [82]. The analysis of the interaction between the STAT3/PD1 axis and Tregs have yielded conflicting results and further studies are needed to establish the exact role of Tregs in the above tight regulation [82,83,84,85]. The presence of an environment more prone to inflammatory activity in the bone marrow and instead of suppression of proliferation in the target organs of GvHD, is confirmed in a recent study which demonstrates the presence of a population of the less suppressive CD161+ Tregs electively localized in the bone marrow of patients who have undergone a transplant with regulatory T cells [86]. Figure 1 illustrates the main mechanisms involved in controlling GvHD while sparing GvL effect. Currently the most used protocols are based on the use of Tregs cells selected immunomagnetically but not expanded in vitro. However the number of Tregs that can be collected from a donor is relatively low (1 million/Kg). One possibility to increase the number of infused Tregs is to use ex vivo expanded Tregs with the advantage of having large numbers of cGMPgrade Tregs [60,62,76,87,88,89] . The major obstacle is represented by the requirement of GMP manufacture which is expensive, not always available and requires expert, dedicated laboratory staff [90].
The success of CAR T-cell therapy in hematological cancers has sparked interest in redirecting the specificity of regulatory T cells (Tregs) towards antigens responsible for autoimmunity and transplant rejection [91,92]. In recent studies, CAR Tregs were specifically designed to address alloimmunity, focusing on the human leukocyte antigen A2 (HLA-A2) present in transplanted tissues but absent in recipients. The primary objective was to reorient the Tregs to enhance tolerance for transplanted grafts and reduce the incidence of GVHD [93,94,95,96]. Notably, anti-HLA-A2 CAR Tregs demonstrated superior performance compared to polyclonal Tregs. They were more effective in suppressing xenogeneic GVHD and significantly reducing the rejection of skin allografts [93,94,95,96]. In an early phase I clinical trial (NCT05993611) for the treatment of cGvHD, a different antigen is under investigation to redirect CAR Tregs [97]. This antigen is known as CD6 and is found primarily on the patient's T-lymphocytes. CD6 binds to activated leukocyte cell adhesion molecule (ALCAM) expressed on antigen-presenting cells (APCs) [97]. CD6 is crucial in the activation, growth, differentiation, and movement of T-lymphocytes. Another antigen that has been recently examined in preclinical trials for the redirection of CAR Tregs is the CD19 antigen found on B cells [98]. The evaluation of the best CAR design for Tregs is still ongoing since different costimulatory domains could have an impact on the phenotype, function, and cytokine secretion of Tregs [99,100]. According to a study conducted by Boroughs and colleagues, using a 4-1BB-based CAR in Tregs had a detrimental impact on their ability to carry out their regulatory functions [99]. Another study by Dawson et al. conducted an extensive investigation into how various co-stimulatory domains influence the function of an anti-HLA-A2 CAR in an allotransplantation model [100,101]. Their data revealed that the CAR encoding CD28 was more effective both in vitro and in vivo concerning proliferation, suppression, and the delay of GVHD symptoms while the presence of 4-1BB-CAR had a negative impact on Treg function and stability [100]. In contrast to Dawson and colleagues' findings, Koristka et al. employed a modular CAR technology known as UniCAR and demonstrated that CD28-based CARs might exhibit off-target effects and enhanced cytolytic activity when compared to CARs based on 4-1BB [102].
2.3. NK
The therapeutic potential of donor NK cells was studied in both haploidentical transplantation and also in the nontransplantation setting; 3 main sources for allogenic NK cells are available actually, each presenting advantages and disadvantages: 1) donor peripheral blood, cord blood (CB) or differentiation from CB hematopoietic stem and progenitor cells HSPC or induced pluripotent stem cells (IPSc), though these latter cells have a limited use because of difficult sourcing and their use is challenging at the moment [103,104]. Several trials showed feasibility and safety of infusing high doses of NK cells after haploidentical HSCT with relevant benefit for hematologic malignancies. In 2014 Choi et al. [105] published the results of the first trial of donor NK cells administered at high dose after haploidentical allotransplant, this latter performed as salvage treatment in n 41 patients with previous diagnosis of active/refractory hematological malignancies, mostly AML. In this study donor-derived NK cells from a mobilized leukapheresis were infused at median dose of 2.0x108/kg. Side effects were not appreciable in any patient. Acute GVHD was reported in 22% of cases at a median of 8 months after transplant, chronic GVHD occurred in 24% of cases at a median of 3.3 months after HCT. As compared to patients who underwent HLA-haploidentical HCT investigated patients obtained a significant reduction in leukemia progression (74% to 46%), providing evidence of enhanced antileukemia effect of donor NK cells, possibly with direct action on leukemia cells or action as enhancers of a T cell mediated antileukemia effect [106]. Notably, more NK cells expressing activating receptors were detectable early after infusion in the peripheral blood of patients who received NK infusion [106]. NK cells require homeostatic cytokine support after transfer. Intermediate or low dose rhIL2 for 2 weeks after transfer has been tested in multiple trials and they are generally considered safe with evidence of in vivo function persistence [103]. In a first trial after infusion of donor mature NK cells followed by administration of IL-2 daily for 14 days a successful donor NK-cell expansion was observed for patients previously treated with cyclophosphamide and fludarabine [107,108], thus allowing the achievement of complete remission in 30% of poor prognosis AML patients. Furthermore, in order to increase the antileukemic action of transplant without worsening the risk of GVHD one can decide to infuse NK cells from an HLA haploidentical donor, chosen for its alloreactivity, and distinct from a separate HLA identical donor, chosen for allotransplantation. In this study NK cells obtained from PBMCs of an HLA haploidentical related donor, after an overnight incubation with IL-2, were infused at escalating doses (dose ranging from 0.02 to 8.32 ×106 /kg) in 21 high-risk myeloid malignancies. After infusion, subcutaneous IL-2 was administered daily for 5 days. 100% engraftment and a rate of ≥ grade 3 aGvHD (10%) was observed. According to this study relapse-free, overall and GvHD/relapse free survival were 102, 233 and 89 days respectively [109]. IL15R agonists may represent a possible alternative to IL2 [110,111] in spite of the risk of an induced cytokine-exhausted state [112]. The use of membrane-bound interleukin 21 expressing cells led to an impressive 35,000-fold expansion of natural killer (NK) cells within 21 days [113]. In a clinical study involving 13 individuals with high-risk myeloid malignancies, increasing doses of donor NK cells, expanded using these interleukin 21 expressing cells, were administered before and after haploidentical HCT (on days −2, +7, and +28). Doses ranged from 1×105/kg to 1×108/kg per dose, escalating up to 3×108/kg, followed by post-transplant cyclophosphamide to prevent severe GVHD. Approximately half of the cases experienced mild to moderate acute GVHD (Grade 1-2), while severe acute GVHD (Grade III-IV) or chronic GVHD were notably absent. Relapse rate and overall mortality was not different than in conventional transplants, without NK cell infusion [113]. The only case of relapse happened at day +120 posttransplant and it was observed in a patient who received the lowest investigated dose (1×105/kg per dose). 1 year OS and DFS are 92% and 85%, respectively [114]. Memory NK cells are being used as an adjunct to haploidentical transplants for patients with advanced AML [115]. Human memory-like (CIML) NK cells, previously preactivated with a combination of IL-12, IL-15, and IL-18 [116,117] could represent another population able to induce a powerful GvL effect [118] in the absence of GvHD. 15 adult patients with high risk relapsed/refractory AML were infused with a range of 0.5 ×106 to 10 × 106 cells/kg after a RIC conditioning. The infusion was performed on day + 7 after transplant and supported by an IL-15 administered subcutaneously at 10 μg/kg starting on day +7 and over 3 weeks. Tacrolimus and mycophenolate mofetil were administered as GVHD prophylaxis Patients showed a good tolerance profile to NK cell transfer, only limited CRS happened and only transient reactions in the site of injection were observed. Acute GvHD occurred in 10 patients (grade 1: 4, grade 2: 6), comparable to expected rates with RIC haplo-HCT, as well as graft failure. Chronic GVHD occurred in nearly 20% of patients. The clearance of high-risk mutations, including TP53 variants was obtained in 87% of patients. 80% of patients were alive at day +100, four patients were still in CR at that time. 1 year OS was 29% [110]. Taking in consideration their rapid expansion and long-term persistence, cytokine-induced memory-like (CIML) NK cells could represent a plausible platform for the treatment of post-transplant relapse of myeloid disease. In a Phase I trial, the recurrence of myeloid malignancies (AML, MDS, MDS/MPN, or blastic plasmacytoid dendritic cell neoplasm (BPDC)) post-haploidentical HCT was addressed with lymphodepleting chemotherapy followed by the infusion of donor-derived CIML NK cells at a dosage ranging from 5 to 10 million cells/kg, along with IL-2 administration. This approach led to a rapid and sustained in vivo expansion of NK cells. The CIML NK cells were generated from non-mobilized apheresis products using a two-step process involving CD3+ depletion followed by CD56+ selection. Among the initial six enrolled patients, by day +28, a favorable disease response was observed in 4 out of 6 individuals, with three out of 6 patients demonstrating a complete response (CR). Notably, neither acute nor chronic graft-versus-host disease (GVHD) was evidenced in any patient [119]. The clinical trials are summarized in Table 3.
2.4. Suicide Gene Therapy
The engineering of polyclonal donor T cells with the insertion of suicide genes capable of limiting their expansion and activity is a strategy that has been adopted in the setting of allogeneic HSCT to separate GvL and GvHD. The major experience was carried out by the San Raffaele group in Milan with the use of the herpes simplex thymidine kinase (HSV-TK) gene. The HSV-TK gene is involved in the metabolism of antiviral ganciclovir and causes selective death of transfected cells when exposed to ganciclovir. First experience with TK lymphocytes were conducted in the context of hematological disease relapse and EBV+ PTLD as DLI after HLA-identical allogeneic HSCT. Survival of TK lymphocytes after infusion, GvL activity directly correlating with in vivo expansion of TK lymphocytes and effective control of GvHD with ganciclovir were demonstrated [120,121]. Based on these results, TK lymphocytes were then tested in the haploidentical context with the aim of improving immune reconstitution and GvL as well as controlling GvHD. The phase I – II multicenter TK007 trial enrolled patients with hematological malignancies undergoing haploidentical HSCT with positive selection of CD34+ cells and no further GvHD prophylaxis after infusion. In absence of valid immune reconstitution, 28 out of 50 enrolled patients received TK lymphocytes at escalating doses. Engraftment of TK lymphocytes was reported in 22 patients short after the first infusion. All grade acute GvHD occurred in 10 patients while 1 patient developed chronic extensive GvHD, both effectively controlled with ganciclovir. At 3-year NRM and OS were 40% and 49%, respectively, in patients with acute leukemia in complete remission at HSCT [122]. Notably, TK lymphocytes have been proven to support also the long-term reconstitution of polyclonal unmanipulated lymphocytes by positively modulation of thymic functions [123]. A similar approach was carried out in the setting of haploidentical HSCT with donor T lymphocytes engineered with the inducible caspase 9 suicide gene (iC9-T cells). The iC9-T cells could be eliminated by administration of a chemical inducer of dimerization (AP1903). Compared to TK lymphocytes, this different mechanism allowed a more rapid inactivation of engineered cells and the possibility to receive antiviral drugs without T-cell damage. All 12 enrolled patients who underwent haploidentical HSCT and received iC9-T cells obtained robust immune reconstitution against viral and opportunistic infections. The administration of AP1903 in 4 patients with GvHD granted rapid and highly effective clearance of iC9-T cells from both peripheral blood and the central nervous system [124]. Long-term persistence of iC9-T as well as a positive immunological effect on polyclonal unmanipulated T lymphocytes were observed also with this approach, reflecting in sustained protection from infectious complications [125]. The clinical trials are summarized in Table 4.
3. Conclusions
The cell therapy platforms associated with stem cell transplantation discussed here have been shown to be able to successfully prevent GvHD and at the same time allow a potent antileukemic effect (Figure 2). The proposed immunotherapies could represent the solution capable of reducing the incidence of post-transplant relapses.
Author Contributions
MDI, CL, NS, PR, FG, GC, IV, BDF, SL, CP, PL and PA wrote and prepared the draft manuscript; MDI review and edited the final version. All authors have read and agreed to the submitted version of the manuscript.
Acknowledgments
This work was supported by Italian Ministry of Health (PNC-E3-2022-23683269 PNC-HLS-TA) and by the Associazione Italiana contro le Leucemie-Linfomi e Mieloma (AIL), L’Aquila Section, Italy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jagasia, M.; Arora, M.; Flowers, M.E.; Chao, N.J.; McCarthy, P.L.; Cutler, C.S.; Urbano-Ispizua, A.; Pavletic, S.Z.; Haagenson, M.D.; Zhang, M.J.; et al. Risk factors for acute GVHD and survival after hematopoietic cell transplantation. Blood 2012, 119, 296–307. [Google Scholar] [CrossRef]
- Zeiser, R.; Blazar, B.R. Acute Graft-versus-Host Disease - Biologic Process, Prevention, and Therapy. N Engl J Med 2017, 377, 2167–2179. [Google Scholar] [CrossRef]
- Chiad, Z.; Chojecki, A. Graft versus Leukemia in 2023. Best Pract Res Clin Haematol 2023, 36, 101476. [Google Scholar] [CrossRef]
- Aversa, F.; Tabilio, A.; Velardi, A.; Cunningham, I.; Terenzi, A.; Falzetti, F.; Ruggeri, L.; Barbabietola, G.; Aristei, C.; Latini, P.; et al. Treatment of high-risk acute leukemia with T-cell-depleted stem cells from related donors with one fully mismatched HLA haplotype. N Engl J Med 1998, 339, 1186–1193. [Google Scholar] [CrossRef]
- Aversa, F.; Terenzi, A.; Tabilio, A.; Falzetti, F.; Carotti, A.; Ballanti, S.; Felicini, R.; Falcinelli, F.; Velardi, A.; Ruggeri, L.; et al. Full haplotype-mismatched hematopoietic stem-cell transplantation: a phase II study in patients with acute leukemia at high risk of relapse. J Clin Oncol 2005, 23, 3447–3454. [Google Scholar] [CrossRef]
- Harris, R.; Karimi, M. Dissecting the regulatory network of transcription factors in T cell phenotype/functioning during GVHD and GVT. Front Immunol 2023, 14, 1194984. [Google Scholar] [CrossRef]
- Blazar, B.R.; Murphy, W.J.; Abedi, M. Advances in graft-versus-host disease biology and therapy. Nat Rev Immunol 2012, 12, 443–458. [Google Scholar] [CrossRef]
- Airoldi, I.; Bertaina, A.; Prigione, I.; Zorzoli, A.; Pagliara, D.; Cocco, C.; Meazza, R.; Loiacono, F.; Lucarelli, B.; Bernardo, M.E.; et al. gammadelta T-cell reconstitution after HLA-haploidentical hematopoietic transplantation depleted of TCR-alphabeta+/CD19+ lymphocytes. Blood 2015, 125, 2349–2358. [Google Scholar] [CrossRef]
- Godder, K.T.; Henslee-Downey, P.J.; Mehta, J.; Park, B.S.; Chiang, K.Y.; Abhyankar, S.; Lamb, L.S. Long term disease-free survival in acute leukemia patients recovering with increased gammadelta T cells after partially mismatched related donor bone marrow transplantation. Bone Marrow Transplant 2007, 39, 751–757. [Google Scholar] [CrossRef]
- Lamb, L.S., Jr.; Henslee-Downey, P.J.; Parrish, R.S.; Godder, K.; Thompson, J.; Lee, C.; Gee, A.P. Increased frequency of TCR gamma delta + T cells in disease-free survivors following T cell-depleted, partially mismatched, related donor bone marrow transplantation for leukemia. J Hematother 1996, 5, 503–509. [Google Scholar] [CrossRef]
- Chaleff, S.; Otto, M.; Barfield, R.C.; Leimig, T.; Iyengar, R.; Martin, J.; Holiday, M.; Houston, J.; Geiger, T.; Huppert, V.; et al. A large-scale method for the selective depletion of alphabeta T lymphocytes from PBSC for allogeneic transplantation. Cytotherapy 2007, 9, 746–754. [Google Scholar] [CrossRef]
- Handgretinger, R. New approaches to graft engineering for haploidentical bone marrow transplantation. Semin Oncol 2012, 39, 664–673. [Google Scholar] [CrossRef]
- Li Pira, G.; Malaspina, D.; Girolami, E.; Biagini, S.; Cicchetti, E.; Conflitti, G.; Broglia, M.; Ceccarelli, S.; Lazzaro, S.; Pagliara, D.; et al. Selective Depletion of alphabeta T Cells and B Cells for Human Leukocyte Antigen-Haploidentical Hematopoietic Stem Cell Transplantation. A Three-Year Follow-Up of Procedure Efficiency. Biol Blood Marrow Transplant 2016, 22, 2056–2064. [Google Scholar] [CrossRef]
- Locatelli, F.; Pende, D.; Maccario, R.; Mingari, M.C.; Moretta, A.; Moretta, L. Haploidentical hemopoietic stem cell transplantation for the treatment of high-risk leukemias: how NK cells make the difference. Clin Immunol 2009, 133, 171–178. [Google Scholar] [CrossRef]
- Moretta, L.; Locatelli, F.; Pende, D.; Marcenaro, E.; Mingari, M.C.; Moretta, A. Killer Ig-like receptor-mediated control of natural killer cell alloreactivity in haploidentical hematopoietic stem cell transplantation. Blood 2011, 117, 764–771. [Google Scholar] [CrossRef]
- Leung, W.; Iyengar, R.; Turner, V.; Lang, P.; Bader, P.; Conn, P.; Niethammer, D.; Handgretinger, R. Determinants of antileukemia effects of allogeneic NK cells. J Immunol 2004, 172, 644–650. [Google Scholar] [CrossRef]
- Muccio, L.; Bertaina, A.; Falco, M.; Pende, D.; Meazza, R.; Lopez-Botet, M.; Moretta, L.; Locatelli, F.; Moretta, A.; Della Chiesa, M. Analysis of memory-like natural killer cells in human cytomegalovirus-infected children undergoing alphabeta+T and B cell-depleted hematopoietic stem cell transplantation for hematological malignancies. Haematologica 2016, 101, 371–381. [Google Scholar] [CrossRef]
- Bertaina, A.; Merli, P.; Rutella, S.; Pagliara, D.; Bernardo, M.E.; Masetti, R.; Pende, D.; Falco, M.; Handgretinger, R.; Moretta, F.; et al. HLA-haploidentical stem cell transplantation after removal of alphabeta+ T and B cells in children with nonmalignant disorders. Blood 2014, 124, 822–826. [Google Scholar] [CrossRef]
- Merli, P.; Pagliara, D.; Galaverna, F.; Li Pira, G.; Andreani, M.; Leone, G.; Amodio, D.; Pinto, R.M.; Bertaina, A.; Bertaina, V.; et al. TCRalphabeta/CD19 depleted HSCT from an HLA-haploidentical relative to treat children with different nonmalignant disorders. Blood Adv 2022, 6, 281–292. [Google Scholar] [CrossRef]
- Gaziev, J.; Isgro, A.; Sodani, P.; Paciaroni, K.; De Angelis, G.; Marziali, M.; Ribersani, M.; Alfieri, C.; Lanti, A.; Galluccio, T.; et al. Haploidentical HSCT for hemoglobinopathies: improved outcomes with TCRalphabeta(+)/CD19(+)-depleted grafts. Blood Adv 2018, 2, 263–270. [Google Scholar] [CrossRef]
- Laberko, A.; Sultanova, E.; Gutovskaya, E.; Shipitsina, I.; Shelikhova, L.; Kurnikova, E.; Muzalevskii, Y.; Kazachenok, A.; Pershin, D.; Voronin, K.; et al. Mismatched related vs matched unrelated donors in TCRalphabeta/CD19-depleted HSCT for primary immunodeficiencies. Blood 2019, 134, 1755–1763. [Google Scholar] [CrossRef]
- Locatelli, F.; Merli, P.; Pagliara, D.; Li Pira, G.; Falco, M.; Pende, D.; Rondelli, R.; Lucarelli, B.; Brescia, L.P.; Masetti, R.; et al. Outcome of children with acute leukemia given HLA-haploidentical HSCT after alphabeta T-cell and B-cell depletion. Blood 2017, 130, 677–685. [Google Scholar] [CrossRef]
- Maschan, M.; Shelikhova, L.; Ilushina, M.; Kurnikova, E.; Boyakova, E.; Balashov, D.; Persiantseva, M.; Skvortsova, Y.; Laberko, A.; Muzalevskii, Y.; et al. TCR-alpha/beta and CD19 depletion and treosulfan-based conditioning regimen in unrelated and haploidentical transplantation in children with acute myeloid leukemia. Bone Marrow Transplant 2016, 51, 668–674. [Google Scholar] [CrossRef]
- Laberko, A.; Bogoyavlenskaya, A.; Shelikhova, L.; Shekhovtsova, Z.; Balashov, D.; Voronin, K.; Kurnikova, E.; Boyakova, E.; Raykina, E.; Brilliantova, V.; et al. Risk Factors for and the Clinical Impact of Cytomegalovirus and Epstein-Barr Virus Infections in Pediatric Recipients of TCR-alpha/beta- and CD19-Depleted Grafts. Biol Blood Marrow Transplant 2017, 23, 483–490. [Google Scholar] [CrossRef]
- Lang, P.; Feuchtinger, T.; Teltschik, H.M.; Schwinger, W.; Schlegel, P.; Pfeiffer, M.; Schumm, M.; Lang, A.M.; Lang, B.; Schwarze, C.P.; et al. Improved immune recovery after transplantation of TCRalphabeta/CD19-depleted allografts from haploidentical donors in pediatric patients. Bone Marrow Transplant 2015, 50 (Suppl 2), S6-10. [Google Scholar] [CrossRef]
- Park, M.; Im, H.J.; Lee, Y.J.; Park, N.; Jang, S.; Kwon, S.W.; Park, C.J.; Choi, E.S.; Koh, K.N.; Seo, J.J. Reconstitution of T and NK cells after haploidentical hematopoietic cell transplantation using alphabeta T cell-depleted grafts and the clinical implication of gammadelta T cells. Clin Transplant 2018, 32. [Google Scholar] [CrossRef]
- Prezioso, L.; Manfra, I.; Bonomini, S.; Schifano, C.; Segreto, R.; Monti, A.; Sammarelli, G.; Todaro, G.; Sassi, M.; Bertaggia, I.; et al. Haploidentical hematopoietic stem cell transplantation in adults using the alphabetaTCR/CD19-based depletion of G-CSF-mobilized peripheral blood progenitor cells. Bone Marrow Transplant 2019, 54, 698–702. [Google Scholar] [CrossRef]
- Kaynar, L.; Demir, K.; Turak, E.E.; Ozturk, C.P.; Zararsiz, G.; Gonen, Z.B.; Gokahmetoglu, S.; Sivgin, S.; Eser, B.; Koker, Y.; et al. TcRalphabeta-depleted haploidentical transplantation results in adult acute leukemia patients. Hematology 2017, 22, 136–144. [Google Scholar] [CrossRef]
- de Witte, M.A.; Janssen, A.; Nijssen, K.; Karaiskaki, F.; Swanenberg, L.; van Rhenen, A.; Admiraal, R.; van der Wagen, L.; Minnema, M.C.; Petersen, E.; et al. alphabeta T-cell graft depletion for allogeneic HSCT in adults with hematological malignancies. Blood Adv 2021, 5, 240–249. [Google Scholar] [CrossRef]
- Radestad, E.; Sundin, M.; Torlen, J.; Thunberg, S.; Onfelt, B.; Ljungman, P.; Watz, E.; Mattsson, J.; Uhlin, M. Individualization of Hematopoietic Stem Cell Transplantation Using Alpha/Beta T-Cell Depletion. Front Immunol 2019, 10, 189. [Google Scholar] [CrossRef]
- Anderson, B.E.; McNiff, J.; Yan, J.; Doyle, H.; Mamula, M.; Shlomchik, M.J.; Shlomchik, W.D. Memory CD4+ T cells do not induce graft-versus-host disease. J Clin Invest 2003, 112, 101–108. [Google Scholar] [CrossRef]
- Anderson, B.E.; Tang, A.L.; Wang, Y.; Froicu, M.; Rothstein, D.; McNiff, J.M.; Jain, D.; Demetris, A.J.; Farber, D.L.; Shlomchik, W.D.; et al. Enhancing alloreactivity does not restore GVHD induction but augments skin graft rejection by CD4(+) effector memory T cells. Eur J Immunol 2011, 41, 2782–2792. [Google Scholar] [CrossRef]
- Li, N.; Matte-Martone, C.; Zheng, H.; Cui, W.; Venkatesan, S.; Tan, H.S.; McNiff, J.; Demetris, A.J.; Roopenian, D.; Kaech, S.; et al. Memory T cells from minor histocompatibility antigen-vaccinated and virus-immune donors improve GVL and immune reconstitution. Blood 2011, 118, 5965–5976. [Google Scholar] [CrossRef]
- Zheng, H.; Matte-Martone, C.; Li, H.; Anderson, B.E.; Venketesan, S.; Sheng Tan, H.; Jain, D.; McNiff, J.; Shlomchik, W.D. Effector memory CD4+ T cells mediate graft-versus-leukemia without inducing graft-versus-host disease. Blood 2008, 111, 2476–2484. [Google Scholar] [CrossRef]
- Teschner, D.; Distler, E.; Wehler, D.; Frey, M.; Marandiuc, D.; Langeveld, K.; Theobald, M.; Thomas, S.; Herr, W. Depletion of naive T cells using clinical grade magnetic CD45RA beads: a new approach for GVHD prophylaxis. Bone Marrow Transplant 2014, 49, 138–144. [Google Scholar] [CrossRef]
- Bleakley, M.; Heimfeld, S.; Loeb, K.R.; Jones, L.A.; Chaney, C.; Seropian, S.; Gooley, T.A.; Sommermeyer, F.; Riddell, S.R.; Shlomchik, W.D. Outcomes of acute leukemia patients transplanted with naive T cell-depleted stem cell grafts. J Clin Invest 2015, 125, 2677–2689. [Google Scholar] [CrossRef]
- Bleakley, M.; Sehgal, A.; Seropian, S.; Biernacki, M.A.; Krakow, E.F.; Dahlberg, A.; Persinger, H.; Hilzinger, B.; Martin, P.J.; Carpenter, P.A.; et al. Naive T-Cell Depletion to Prevent Chronic Graft-Versus-Host Disease. J Clin Oncol 2022, 40, 1174–1185. [Google Scholar] [CrossRef]
- Bleakley, M.; Otterud, B.E.; Richardt, J.L.; Mollerup, A.D.; Hudecek, M.; Nishida, T.; Chaney, C.N.; Warren, E.H.; Leppert, M.F.; Riddell, S.R. Leukemia-associated minor histocompatibility antigen discovery using T-cell clones isolated by in vitro stimulation of naive CD8+ T cells. Blood 2010, 115, 4923–4933. [Google Scholar] [CrossRef]
- van Bergen, C.A.; van Luxemburg-Heijs, S.A.; de Wreede, L.C.; Eefting, M.; von dem Borne, P.A.; van Balen, P.; Heemskerk, M.H.; Mulder, A.; Claas, F.H.; Navarrete, M.A.; et al. Selective graft-versus-leukemia depends on magnitude and diversity of the alloreactive T cell response. J Clin Invest 2017, 127, 517–529. [Google Scholar] [CrossRef]
- Mamcarz, E.; Madden, R.; Qudeimat, A.; Srinivasan, A.; Talleur, A.; Sharma, A.; Suliman, A.; Maron, G.; Sunkara, A.; Kang, G.; et al. Improved survival rate in T-cell depleted haploidentical hematopoietic cell transplantation over the last 15 years at a single institution. Bone Marrow Transplant 2020, 55, 929–938. [Google Scholar] [CrossRef]
- Triplett, B.M.; Shook, D.R.; Eldridge, P.; Li, Y.; Kang, G.; Dallas, M.; Hartford, C.; Srinivasan, A.; Chan, W.K.; Suwannasaen, D.; et al. Rapid memory T-cell reconstitution recapitulating CD45RA-depleted haploidentical transplant graft content in patients with hematologic malignancies. Bone Marrow Transplant 2015, 50, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Talleur, A.C.; Li, Y.; Madden, R.M.; Mamcarz, E.; Qudeimat, A.; Sharma, A.; Srinivasan, A.; Suliman, A.Y.; Epperly, R.; et al. CD45RA-Depleted Haploidentical Transplantation Combined with NK Cell Addback Results in Promising Long-Term Outcomes in Pediatric Patients with High-Risk Hematologic Malignancies. Blood 2021, 138, 172. [Google Scholar] [CrossRef]
- Sisinni, L.; Gasior, M.; de Paz, R.; Querol, S.; Bueno, D.; Fernandez, L.; Marsal, J.; Sastre, A.; Gimeno, R.; Alonso, L.; et al. Unexpected High Incidence of Human Herpesvirus-6 Encephalitis after Naive T Cell-Depleted Graft of Haploidentical Stem Cell Transplantation in Pediatric Patients. Biol Blood Marrow Transplant 2018, 24, 2316–2323. [Google Scholar] [CrossRef] [PubMed]
- Perruccio, K.; Sisinni, L.; Perez-Martinez, A.; Valentin, J.; Capolsini, I.; Massei, M.S.; Caniglia, M.; Cesaro, S. High Incidence of Early Human Herpesvirus-6 Infection in Children Undergoing Haploidentical Manipulated Stem Cell Transplantation for Hematologic Malignancies. Biol Blood Marrow Transplant 2018, 24, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Gasior Kabat, M.; Bueno, D.; Sisinni, L.; De Paz, R.; Mozo, Y.; Perona, R.; Arias-Salgado, E.G.; Rosich, B.; Marcos, A.; Romero, A.B.; et al. Selective T-cell depletion targeting CD45RA as a novel approach for HLA-mismatched hematopoietic stem cell transplantation in pediatric nonmalignant hematological diseases. Int J Hematol 2021, 114, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Touzot, F.; Neven, B.; Dal-Cortivo, L.; Gabrion, A.; Moshous, D.; Cros, G.; Chomton, M.; Luby, J.M.; Terniaux, B.; Magalon, J.; et al. CD45RA depletion in HLA-mismatched allogeneic hematopoietic stem cell transplantation for primary combined immunodeficiency: A preliminary study. J Allergy Clin Immunol 2015, 135, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Muffly, L.; Sheehan, K.; Armstrong, R.; Jensen, K.; Tate, K.; Rezvani, A.R.; Miklos, D.; Arai, S.; Shizuru, J.; Johnston, L.; et al. Infusion of donor-derived CD8(+) memory T cells for relapse following allogeneic hematopoietic cell transplantation. Blood Adv 2018, 2, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Dunaikina, M.; Zhekhovtsova, Z.; Shelikhova, L.; Glushkova, S.; Nikolaev, R.; Blagov, S.; Khismatullina, R.; Balashov, D.; Kurnikova, E.; Pershin, D.; et al. Safety and efficacy of the low-dose memory (CD45RA-depleted) donor lymphocyte infusion in recipients of alphabeta T cell-depleted haploidentical grafts: results of a prospective randomized trial in high-risk childhood leukemia. Bone Marrow Transplant 2021, 56, 1614–1624. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Madden, R.M.; Mamcarz, E.; Srinivasan, A.; Sharma, A.; Talleur, A.C.; Epperly, R.; Qudeimat, A.; Suliman, A.Y.; Obeng, E.A.; et al. CD45RO+ T-Cell Add Back and Prophylactic Blinatumomab Administration Post Tcrαβ/CD19-Depleted Haploidentical Transplantation in Pediatric Patients with High Risk Acute Leukemia. Blood 2021, 138, 2897. [Google Scholar] [CrossRef]
- Castagna, L.; Valli, V.; Timofeeva, I.; Capizzuto, R.; Bramanti, S.; Mariotti, J.; De Philippis, C.; Sarina, B.; Mannina, D.; Giordano, L.; et al. Feasibility and Efficacy of CD45RA+ Depleted Donor Lymphocytes Infusion After Haploidentical Transplantation With Post-Transplantation Cyclophosphamide in Patients With Hematological Malignancies. Transplant Cell Ther 2021, 27, 478.e471–478.e475. [Google Scholar] [CrossRef]
- Maung, K.K.; Chen, B.J.; Barak, I.; Li, Z.; Rizzieri, D.A.; Gasparetto, C.; Sullivan, K.M.; Long, G.D.; Engemann, A.M.; Waters-Pick, B.; et al. Phase I dose escalation study of naive T-cell depleted donor lymphocyte infusion following allogeneic stem cell transplantation. Bone Marrow Transplant 2021, 56, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol 2004, 22, 531–562. [Google Scholar] [CrossRef]
- Itoh, M.; Takahashi, T.; Sakaguchi, N.; Kuniyasu, Y.; Shimizu, J.; Otsuka, F.; Sakaguchi, S. Thymus and autoimmunity: production of CD25+CD4+ naturally anergic and suppressive T cells as a key function of the thymus in maintaining immunologic self-tolerance. J Immunol 1999, 162, 5317–5326. [Google Scholar] [CrossRef]
- Stephens, L.A.; Mottet, C.; Mason, D.; Powrie, F. Human CD4(+)CD25(+) thymocytes and peripheral T cells have immune suppressive activity in vitro. Eur J Immunol 2001, 31, 1247–1254. [Google Scholar] [CrossRef]
- Baecher-Allan, C.; Brown, J.A.; Freeman, G.J.; Hafler, D.A. CD4+CD25high regulatory cells in human peripheral blood. J Immunol 2001, 167, 1245–1253. [Google Scholar] [CrossRef]
- Di Ianni, M.; Del Papa, B.; Cecchini, D.; Bonifacio, E.; Moretti, L.; Zei, T.; Ostini, R.I.; Falzetti, F.; Fontana, L.; Tagliapietra, G.; et al. Immunomagnetic isolation of CD4+CD25+FoxP3+ natural T regulatory lymphocytes for clinical applications. Clin Exp Immunol 2009, 156, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Di Ianni, M.; Del Papa, B.; Zei, T.; Iacucci Ostini, R.; Cecchini, D.; Cantelmi, M.G.; Baldoni, S.; Sportoletti, P.; Cavalli, L.; Carotti, A.; et al. T regulatory cell separation for clinical application. Transfus Apher Sci 2012, 47, 213–216. [Google Scholar] [CrossRef]
- Hoffmann, P.; Boeld, T.J.; Eder, R.; Albrecht, J.; Doser, K.; Piseshka, B.; Dada, A.; Niemand, C.; Assenmacher, M.; Orso, E.; et al. Isolation of CD4+CD25+ regulatory T cells for clinical trials. Biol Blood Marrow Transplant 2006, 12, 267–274. [Google Scholar] [CrossRef]
- Safinia, N.; Vaikunthanathan, T.; Fraser, H.; Thirkell, S.; Lowe, K.; Blackmore, L.; Whitehouse, G.; Martinez-Llordella, M.; Jassem, W.; Sanchez-Fueyo, A.; et al. Successful expansion of functional and stable regulatory T cells for immunotherapy in liver transplantation. Oncotarget 2016, 7, 7563–7577. [Google Scholar] [CrossRef]
- Brunstein, C.G.; Fuchs, E.J.; Carter, S.L.; Karanes, C.; Costa, L.J.; Wu, J.; Devine, S.M.; Wingard, J.R.; Aljitawi, O.S.; Cutler, C.S.; et al. Alternative donor transplantation after reduced intensity conditioning: results of parallel phase 2 trials using partially HLA-mismatched related bone marrow or unrelated double umbilical cord blood grafts. Blood 2011, 118, 282–288. [Google Scholar] [CrossRef]
- Chandran, S.; Tang, Q.; Sarwal, M.; Laszik, Z.G.; Putnam, A.L.; Lee, K.; Leung, J.; Nguyen, V.; Sigdel, T.; Tavares, E.C.; et al. Polyclonal Regulatory T Cell Therapy for Control of Inflammation in Kidney Transplants. Am J Transplant 2017, 17, 2945–2954. [Google Scholar] [CrossRef]
- Bluestone, J.A.; Buckner, J.H.; Fitch, M.; Gitelman, S.E.; Gupta, S.; Hellerstein, M.K.; Herold, K.C.; Lares, A.; Lee, M.R.; Li, K.; et al. Type 1 diabetes immunotherapy using polyclonal regulatory T cells. Sci Transl Med 2015, 7, 315ra189. [Google Scholar] [CrossRef] [PubMed]
- Di Ianni, M.; Falzetti, F.; Carotti, A.; Terenzi, A.; Castellino, F.; Bonifacio, E.; Del Papa, B.; Zei, T.; Ostini, R.I.; Cecchini, D.; et al. Tregs prevent GVHD and promote immune reconstitution in HLA-haploidentical transplantation. Blood 2011, 117, 3921–3928. [Google Scholar] [CrossRef] [PubMed]
- Martelli, M.F.; Di Ianni, M.; Ruggeri, L.; Falzetti, F.; Carotti, A.; Terenzi, A.; Pierini, A.; Massei, M.S.; Amico, L.; Urbani, E.; et al. HLA-haploidentical transplantation with regulatory and conventional T-cell adoptive immunotherapy prevents acute leukemia relapse. Blood 2014, 124, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Pierini, A.; Ruggeri, L.; Carotti, A.; Falzetti, F.; Saldi, S.; Terenzi, A.; Zucchetti, C.; Ingrosso, G.; Zei, T.; Iacucci Ostini, R.; et al. Haploidentical age-adapted myeloablative transplant and regulatory and effector T cells for acute myeloid leukemia. Blood Adv 2021, 5, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Zeiser, R.; Dasilva, D.L.; Chang, D.S.; Beilhack, A.; Contag, C.H.; Negrin, R.S. In vivo dynamics of regulatory T-cell trafficking and survival predict effective strategies to control graft-versus-host disease following allogeneic transplantation. Blood 2007, 109, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat Rev Immunol 2008, 8, 523–532. [Google Scholar] [CrossRef]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef]
- Nakamura, K.; Kitani, A.; Strober, W. Cell contact-dependent immunosuppression by CD4(+)CD25(+) regulatory T cells is mediated by cell surface-bound transforming growth factor beta. J Exp Med 2001, 194, 629–644. [Google Scholar] [CrossRef]
- Bolton, H.A.; Zhu, E.; Terry, A.M.; Guy, T.V.; Koh, W.P.; Tan, S.Y.; Power, C.A.; Bertolino, P.; Lahl, K.; Sparwasser, T.; et al. Selective Treg reconstitution during lymphopenia normalizes DC costimulation and prevents graft-versus-host disease. J Clin Invest 2015, 125, 3627–3641. [Google Scholar] [CrossRef]
- Liang, B.; Workman, C.; Lee, J.; Chew, C.; Dale, B.M.; Colonna, L.; Flores, M.; Li, N.; Schweighoffer, E.; Greenberg, S.; et al. Regulatory T cells inhibit dendritic cells by lymphocyte activation gene-3 engagement of MHC class II. J Immunol 2008, 180, 5916–5926. [Google Scholar] [CrossRef]
- Yan, N.; Regalado-Magdos, A.D.; Stiggelbout, B.; Lee-Kirsch, M.A.; Lieberman, J. The cytosolic exonuclease TREX1 inhibits the innate immune response to human immunodeficiency virus type 1. Nat Immunol 2010, 11, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Borsellino, G.; Kleinewietfeld, M.; Di Mitri, D.; Sternjak, A.; Diamantini, A.; Giometto, R.; Hopner, S.; Centonze, D.; Bernardi, G.; Dell'Acqua, M.L.; et al. Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: hydrolysis of extracellular ATP and immune suppression. Blood 2007, 110, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Edinger, M.; Hoffmann, P.; Ermann, J.; Drago, K.; Fathman, C.G.; Strober, S.; Negrin, R.S. CD4+CD25+ regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation. Nat Med 2003, 9, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Lohmeyer, J.K.; Hirai, T.; Turkoz, M.; Buhler, S.; Lopes Ramos, T.; Kohler, N.; Baker, J.; Melotti, A.; Wagner, I.; Pradier, A.; et al. Analysis of the T-cell repertoire and transcriptome identifies mechanisms of regulatory T-cell suppression of GVHD. Blood 2023, 141, 1755–1767. [Google Scholar] [CrossRef]
- Ulbar, F.; Villanova, I.; Giancola, R.; Baldoni, S.; Guardalupi, F.; Fabi, B.; Olioso, P.; Capone, A.; Sola, R.; Ciardelli, S.; et al. Clinical-Grade Expanded Regulatory T Cells Are Enriched with Highly Suppressive Cells Producing IL-10, Granzyme B, and IL-35. Biol Blood Marrow Transplant 2020, 26, 2204–2210. [Google Scholar] [CrossRef]
- Zhang, Y.; Sandy, A.R.; Wang, J.; Radojcic, V.; Shan, G.T.; Tran, I.T.; Friedman, A.; Kato, K.; He, S.; Cui, S.; et al. Notch signaling is a critical regulator of allogeneic CD4+ T-cell responses mediating graft-versus-host disease. Blood 2011, 117, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Hippen, K.L.; Aguilar, E.G.; Rhee, S.Y.; Bolivar-Wagers, S.; Blazar, B.R. Distinct Regulatory and Effector T Cell Metabolic Demands during Graft-Versus-Host Disease. Trends Immunol 2020, 41, 77–91. [Google Scholar] [CrossRef]
- Del Papa, B.; Pierini, A.; Sportoletti, P.; Baldoni, S.; Cecchini, D.; Rosati, E.; Dorillo, E.; Aureli, P.; Zei, T.; Iacucci Ostini, R.; et al. The NOTCH1/CD39 axis: a Treg trip-switch for GvHD. Leukemia 2016, 30, 1931–1934. [Google Scholar] [CrossRef]
- Tkachev, V.; Vanderbeck, A.; Perkey, E.; Furlan, S.N.; McGuckin, C.; Gomez Atria, D.; Gerdemann, U.; Rui, X.; Lane, J.; Hunt, D.J.; et al. Notch signaling drives intestinal graft-versus-host disease in mice and nonhuman primates. Sci Transl Med 2023, 15, eadd1175. [Google Scholar] [CrossRef]
- Baldoni, S.; Ruggeri, L.; Del Papa, B.; Sorcini, D.; Guardalupi, F.; Ulbar, F.; Marra, A.; Dorillo, E.; Stella, A.; Giancola, R.; et al. NOTCH1 inhibition prevents GvHD and maintains GvL effect in murine models. Bone Marrow Transplant 2021, 56, 2019–2023. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Song, Q.; Yang, S.; Wu, X.; Yang, D.; Marie, I.J.; Qin, H.; Zheng, M.; Nasri, U.; et al. Donor T cell STAT3 deficiency enables tissue PD-L1-dependent prevention of graft-versus-host disease while preserving graft-versus-leukemia activity. J Clin Invest 2023, 133. [Google Scholar] [CrossRef] [PubMed]
- Radojcic, V.; Pletneva, M.A.; Yen, H.R.; Ivcevic, S.; Panoskaltsis-Mortari, A.; Gilliam, A.C.; Drake, C.G.; Blazar, B.R.; Luznik, L. STAT3 signaling in CD4+ T cells is critical for the pathogenesis of chronic sclerodermatous graft-versus-host disease in a murine model. J Immunol 2010, 184, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Laurence, A.; Amarnath, S.; Mariotti, J.; Kim, Y.C.; Foley, J.; Eckhaus, M.; O'Shea, J.J.; Fowler, D.H. STAT3 transcription factor promotes instability of nTreg cells and limits generation of iTreg cells during acute murine graft-versus-host disease. Immunity 2012, 37, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.; Fernandez, M.R.; Sagatys, E.M.; Reff, J.; Kim, J.; Lee, M.C.; Kiluk, J.V.; Hui, J.Y.C.; McKenna, D., Jr.; Hupp, M.; et al. Metabolic reprogramming augments potency of human pSTAT3-inhibited iTregs to suppress alloreactivity. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Guardalupi, F.; Sorrentino, C.; Corradi, G.; Giancola, R.; Baldoni, S.; Ulbar, F.; Fabi, B.; Andres Ejarque, R.; Timms, J.; Restuccia, F.; et al. A pro-inflammatory environment in bone marrow of Treg transplanted patients matches with graft-versus-leukemia effect. Leukemia 2023, 37, 1572–1575. [Google Scholar] [CrossRef]
- Issa, F.; Strober, S.; Leventhal, J.R.; Kawai, T.; Kaufman, D.B.; Levitsky, J.; Sykes, M.; Mas, V.; Wood, K.J.; Bridges, N.; et al. The Fourth International Workshop on Clinical Transplant Tolerance. Am J Transplant 2021, 21, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Raffin, C.; Vo, L.T.; Bluestone, J.A. T(reg) cell-based therapies: challenges and perspectives. Nat Rev Immunol 2020, 20, 158–172. [Google Scholar] [CrossRef] [PubMed]
- O'Neil, A.; Brook, M.; Abdul-Wahab, S.; Hester, J.; Lombardi, G.; Issa, F. A GMP Protocol for the Manufacture of Tregs for Clinical Application. Methods Mol Biol 2023, 2559, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, C.; Deptula, M.; Hester, J.; Issa, F. Barriers to Treg therapy in Europe: From production to regulation. Front Med (Lausanne) 2023, 10, 1090721. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, W.; Liang, C.L.; Chen, Y.; Liu, H.; Qiu, F.; Dai, Z. Chimeric Antigen Receptor (CAR) Treg: A Promising Approach to Inducing Immunological Tolerance. Front Immunol 2018, 9, 2359. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, Y.R.; Tung, S.L.; Dudreuilh, C.; Lechler, R.I.; Fruhwirth, G.O.; Lombardi, G. The Future of Regulatory T Cell Therapy: Promises and Challenges of Implementing CAR Technology. Front Immunol 2020, 11, 1608. [Google Scholar] [CrossRef] [PubMed]
- Noyan, F.; Zimmermann, K.; Hardtke-Wolenski, M.; Knoefel, A.; Schulde, E.; Geffers, R.; Hust, M.; Huehn, J.; Galla, M.; Morgan, M.; et al. Prevention of Allograft Rejection by Use of Regulatory T Cells With an MHC-Specific Chimeric Antigen Receptor. Am J Transplant 2017, 17, 917–930. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, K.G.; Hoeppli, R.E.; Huang, Q.; Gillies, J.; Luciani, D.S.; Orban, P.C.; Broady, R.; Levings, M.K. Alloantigen-specific regulatory T cells generated with a chimeric antigen receptor. J Clin Invest 2016, 126, 1413–1424. [Google Scholar] [CrossRef] [PubMed]
- Boardman, D.A.; Philippeos, C.; Fruhwirth, G.O.; Ibrahim, M.A.; Hannen, R.F.; Cooper, D.; Marelli-Berg, F.M.; Watt, F.M.; Lechler, R.I.; Maher, J.; et al. Expression of a Chimeric Antigen Receptor Specific for Donor HLA Class I Enhances the Potency of Human Regulatory T Cells in Preventing Human Skin Transplant Rejection. Am J Transplant 2017, 17, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Dawson, N.A.; Lamarche, C.; Hoeppli, R.E.; Bergqvist, P.; Fung, V.C.; McIver, E.; Huang, Q.; Gillies, J.; Speck, M.; Orban, P.C.; et al. Systematic testing and specificity mapping of alloantigen-specific chimeric antigen receptors in regulatory T cells. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Ampudia, J.; Chu, D.; Connelly, S.; Ng, C. CD6 Is a Modulator of Treg Differentiation and Activity. Blood 2022, 140, 12653–12654. [Google Scholar] [CrossRef]
- Imura, Y.; Ando, M.; Kondo, T.; Ito, M.; Yoshimura, A. CD19-targeted CAR regulatory T cells suppress B cell pathology without GvHD. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Boroughs, A.C.; Larson, R.C.; Choi, B.D.; Bouffard, A.A.; Riley, L.S.; Schiferle, E.; Kulkarni, A.S.; Cetrulo, C.L.; Ting, D.; Blazar, B.R.; et al. Chimeric antigen receptor costimulation domains modulate human regulatory T cell function. JCI Insight 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Dawson, N.A.J.; Rosado-Sanchez, I.; Novakovsky, G.E.; Fung, V.C.W.; Huang, Q.; McIver, E.; Sun, G.; Gillies, J.; Speck, M.; Orban, P.C.; et al. Functional effects of chimeric antigen receptor co-receptor signaling domains in human regulatory T cells. Sci Transl Med 2020, 12. [Google Scholar] [CrossRef]
- Rosado-Sanchez, I.; Haque, M.; Salim, K.; Speck, M.; Fung, V.C.; Boardman, D.A.; Mojibian, M.; Raimondi, G.; Levings, M.K. Tregs integrate native and CAR-mediated costimulatory signals for control of allograft rejection. JCI Insight 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Koristka, S.; Kegler, A.; Bergmann, R.; Arndt, C.; Feldmann, A.; Albert, S.; Cartellieri, M.; Ehninger, A.; Ehninger, G.; Middeke, J.M.; et al. Engrafting human regulatory T cells with a flexible modular chimeric antigen receptor technology. J Autoimmun 2018, 90, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Berrien-Elliott, M.M.; Jacobs, M.T.; Fehniger, T.A. Allogeneic natural killer cell therapy. Blood 2023, 141, 856–868. [Google Scholar] [CrossRef] [PubMed]
- Lupo, K.B.; Matosevic, S. Natural Killer Cells as Allogeneic Effectors in Adoptive Cancer Immunotherapy. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Yoon, S.R.; Park, S.Y.; Kim, H.; Jung, S.J.; Jang, Y.J.; Kang, M.; Yeom, Y.I.; Lee, J.L.; Kim, D.Y.; et al. Donor-derived natural killer cells infused after human leukocyte antigen-haploidentical hematopoietic cell transplantation: a dose-escalation study. Biol Blood Marrow Transplant 2014, 20, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Krebs, P.; Barnes, M.J.; Lampe, K.; Whitley, K.; Bahjat, K.S.; Beutler, B.; Janssen, E.; Hoebe, K. NK-cell-mediated killing of target cells triggers robust antigen-specific T-cell-mediated and humoral responses. Blood 2009, 113, 6593–6602. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, C.; Bear, J.; Rosati, M.; Beach, R.K.; Alicea, C.; Sowder, R.; Chertova, E.; Rosenberg, S.A.; Felber, B.K.; Pavlakis, G.N. Circulating IL-15 exists as heterodimeric complex with soluble IL-15Ralpha in human and mouse serum. Blood 2012, 120, e1-8. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.A.; Denman, C.J.; Rondon, G.; Woodworth, G.; Chen, J.; Fisher, T.; Kaur, I.; Fernandez-Vina, M.; Cao, K.; Ciurea, S.; et al. Haploidentical Natural Killer Cells Infused before Allogeneic Stem Cell Transplantation for Myeloid Malignancies: A Phase I Trial. Biol Blood Marrow Transplant 2016, 22, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Berrien-Elliott, M.M.; Foltz, J.A.; Russler-Germain, D.A.; Neal, C.C.; Tran, J.; Gang, M.; Wong, P.; Fisk, B.; Cubitt, C.C.; Marin, N.D.; et al. Hematopoietic cell transplantation donor-derived memory-like NK cells functionally persist after transfer into patients with leukemia. Sci Transl Med 2022, 14, eabm1375. [Google Scholar] [CrossRef]
- Berrien-Elliott, M.M.; Becker-Hapak, M.; Cashen, A.F.; Jacobs, M.; Wong, P.; Foster, M.; McClain, E.; Desai, S.; Pence, P.; Cooley, S.; et al. Systemic IL-15 promotes allogeneic cell rejection in patients treated with natural killer cell adoptive therapy. Blood 2022, 139, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Merino, A.; Zhang, B.; Dougherty, P.; Luo, X.; Wang, J.; Blazar, B.R.; Miller, J.S.; Cichocki, F. Chronic stimulation drives human NK cell dysfunction and epigenetic reprograming. J Clin Invest 2019, 129, 3770–3785. [Google Scholar] [CrossRef] [PubMed]
- Denman, C.J.; Senyukov, V.V.; Somanchi, S.S.; Phatarpekar, P.V.; Kopp, L.M.; Johnson, J.L.; Singh, H.; Hurton, L.; Maiti, S.N.; Huls, M.H.; et al. Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells. PLoS One 2012, 7, e30264. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, S.O.; Schafer, J.R.; Bassett, R.; Denman, C.J.; Cao, K.; Willis, D.; Rondon, G.; Chen, J.; Soebbing, D.; Kaur, I.; et al. Phase 1 clinical trial using mbIL21 ex vivo-expanded donor-derived NK cells after haploidentical transplantation. Blood 2017, 130, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Terren, I.; Orrantia, A.; Astarloa-Pando, G.; Amarilla-Irusta, A.; Zenarruzabeitia, O.; Borrego, F. Cytokine-Induced Memory-Like NK Cells: From the Basics to Clinical Applications. Front Immunol 2022, 13, 884648. [Google Scholar] [CrossRef]
- Cooper, M.A.; Elliott, J.M.; Keyel, P.A.; Yang, L.; Carrero, J.A.; Yokoyama, W.M. Cytokine-induced memory-like natural killer cells. Proc Natl Acad Sci U S A 2009, 106, 1915–1919. [Google Scholar] [CrossRef] [PubMed]
- Romee, R.; Schneider, S.E.; Leong, J.W.; Chase, J.M.; Keppel, C.R.; Sullivan, R.P.; Cooper, M.A.; Fehniger, T.A. Cytokine activation induces human memory-like NK cells. Blood 2012, 120, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Romee, R.; Rosario, M.; Berrien-Elliott, M.M.; Wagner, J.A.; Jewell, B.A.; Schappe, T.; Leong, J.W.; Abdel-Latif, S.; Schneider, S.E.; Willey, S.; et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci Transl Med 2016, 8, 357ra123. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R.M.; Birch, G.C.; Hu, G.; Vergara Cadavid, J.; Nikiforow, S.; Baginska, J.; Ali, A.K.; Tarannum, M.; Sheffer, M.; Abdulhamid, Y.Z.; et al. Expansion, persistence, and efficacy of donor memory-like NK cells infused for posttransplant relapse. J Clin Invest 2022, 132. [Google Scholar] [CrossRef]
- Bonini, C.; Ferrari, G.; Verzeletti, S.; Servida, P.; Zappone, E.; Ruggieri, L.; Ponzoni, M.; Rossini, S.; Mavilio, F.; Traversari, C.; et al. HSV-TK gene transfer into donor lymphocytes for control of allogeneic graft-versus-leukemia. Science 1997, 276, 1719–1724. [Google Scholar] [CrossRef]
- Ciceri, F.; Bonini, C.; Marktel, S.; Zappone, E.; Servida, P.; Bernardi, M.; Pescarollo, A.; Bondanza, A.; Peccatori, J.; Rossini, S.; et al. Antitumor effects of HSV-TK-engineered donor lymphocytes after allogeneic stem-cell transplantation. Blood 2007, 109, 4698–4707. [Google Scholar] [CrossRef] [PubMed]
- Ciceri, F.; Bonini, C.; Stanghellini, M.T.; Bondanza, A.; Traversari, C.; Salomoni, M.; Turchetto, L.; Colombi, S.; Bernardi, M.; Peccatori, J.; et al. Infusion of suicide-gene-engineered donor lymphocytes after family haploidentical haemopoietic stem-cell transplantation for leukaemia (the TK007 trial): a non-randomised phase I-II study. Lancet Oncol 2009, 10, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Vago, L.; Oliveira, G.; Bondanza, A.; Noviello, M.; Soldati, C.; Ghio, D.; Brigida, I.; Greco, R.; Lupo Stanghellini, M.T.; Peccatori, J.; et al. T-cell suicide gene therapy prompts thymic renewal in adults after hematopoietic stem cell transplantation. Blood 2012, 120, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Dotti, G.; Krance, R.A.; Martinez, C.A.; Naik, S.; Kamble, R.T.; Durett, A.G.; Dakhova, O.; Savoldo, B.; Di Stasi, A.; et al. Inducible caspase-9 suicide gene controls adverse effects from alloreplete T cells after haploidentical stem cell transplantation. Blood 2015, 125, 4103–4113. [Google Scholar] [CrossRef]
- Zhou, X.; Di Stasi, A.; Tey, S.K.; Krance, R.A.; Martinez, C.; Leung, K.S.; Durett, A.G.; Wu, M.F.; Liu, H.; Leen, A.M.; et al. Long-term outcome after haploidentical stem cell transplant and infusion of T cells expressing the inducible caspase 9 safety transgene. Blood 2014, 123, 3895–3905. [Google Scholar] [CrossRef]
Figure 1.
Main mechanisms of action of Tregs cells involve cytokine production such as IL-10, IL-35 and TGF-beta; direct killing of Tcons by perforine/granzyme mechanism; IL-2 competition; Tregs/DC interaction with downregulation of CD80/CD86 on DCs via CTLA-4; interference with DC maturation through the LAG-3; production of IDO in DC through CTLA-4 and CD80/86 interaction; ATP cleavage by CD39/73; NOTCH1 downregulation in Tcons through CD39. Tregs CD161+ are involved in the same mechanisms with lower suppression activity.
Figure 1.
Main mechanisms of action of Tregs cells involve cytokine production such as IL-10, IL-35 and TGF-beta; direct killing of Tcons by perforine/granzyme mechanism; IL-2 competition; Tregs/DC interaction with downregulation of CD80/CD86 on DCs via CTLA-4; interference with DC maturation through the LAG-3; production of IDO in DC through CTLA-4 and CD80/86 interaction; ATP cleavage by CD39/73; NOTCH1 downregulation in Tcons through CD39. Tregs CD161+ are involved in the same mechanisms with lower suppression activity.
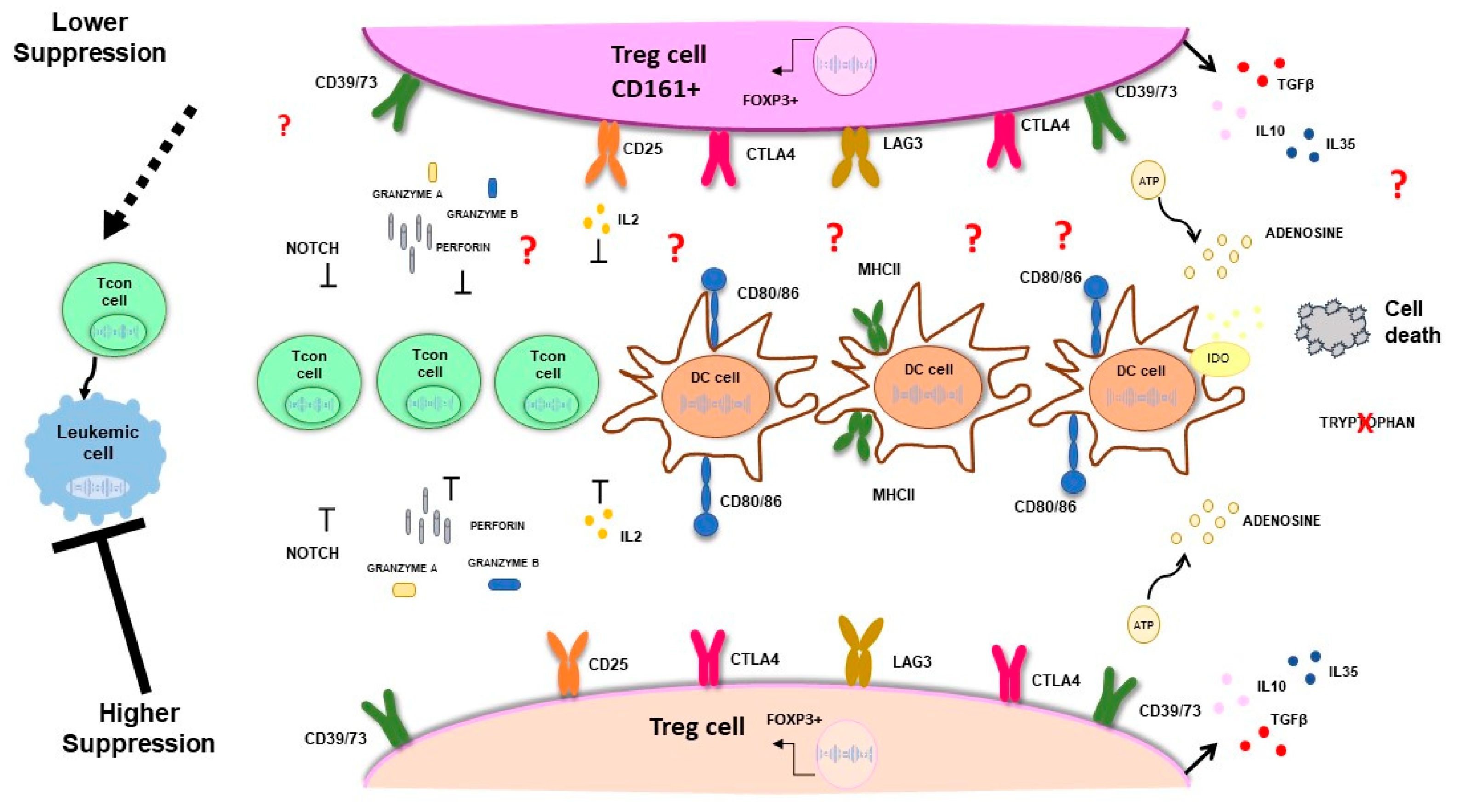
Figure 2.
Current strategies to prevent GvHD and GvL include: a) the depletion of αβ T and CD19+ B cells from mobilized peripheral blood stem cells produces the generation of an allogeneic graft enriched for CD34+ stem cells and γδ T lymphocytes, preserving the NK compartment; b) the depletion of naive T-cells (CD45RA+) is depicted; c) purified Tregs are separated, expanded, and infused together with CD34+ hematopoietic stem cells, as well as with conventional T cells; d) NK cells are selected and infused; e) the suicide gene therapy is represented. The engineering of polyclonal donor T cells with the insertion of suicide genes capable of limiting their expansion and activity is a strategy that has been adopted in the setting of allogeneic HSCT to separate GvL and GvHD. The major experience was carried out using the herpes simplex thymidine kinase (HSV-TK) gene. The HSV-TK gene is involved in the metabolism of antiviral ganciclovir and causes the selective death of transfected cells when exposed to ganciclovir.
Figure 2.
Current strategies to prevent GvHD and GvL include: a) the depletion of αβ T and CD19+ B cells from mobilized peripheral blood stem cells produces the generation of an allogeneic graft enriched for CD34+ stem cells and γδ T lymphocytes, preserving the NK compartment; b) the depletion of naive T-cells (CD45RA+) is depicted; c) purified Tregs are separated, expanded, and infused together with CD34+ hematopoietic stem cells, as well as with conventional T cells; d) NK cells are selected and infused; e) the suicide gene therapy is represented. The engineering of polyclonal donor T cells with the insertion of suicide genes capable of limiting their expansion and activity is a strategy that has been adopted in the setting of allogeneic HSCT to separate GvL and GvHD. The major experience was carried out using the herpes simplex thymidine kinase (HSV-TK) gene. The HSV-TK gene is involved in the metabolism of antiviral ganciclovir and causes the selective death of transfected cells when exposed to ganciclovir.
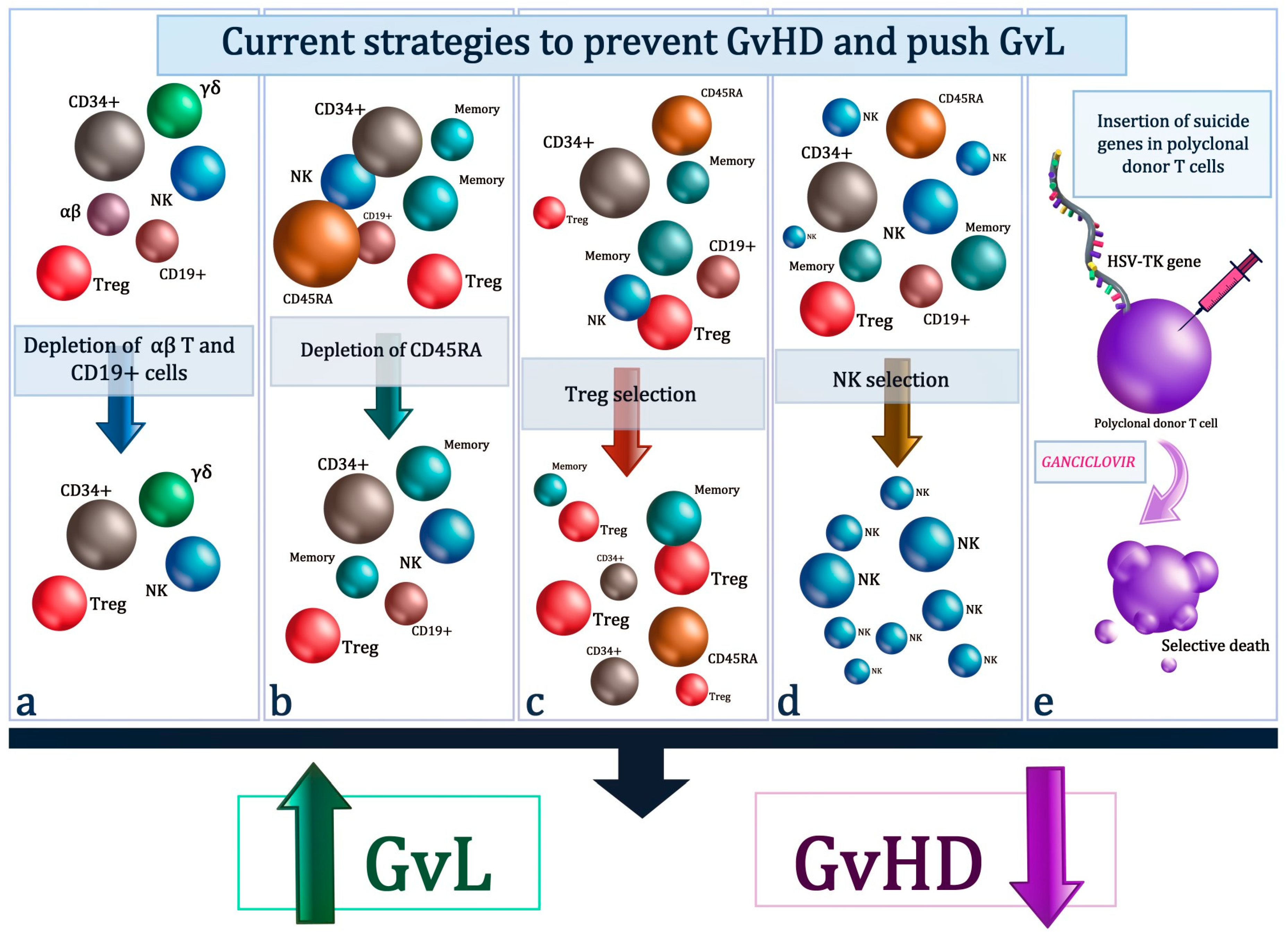

Table 1.
Clinical trials and outcomes after allo-HSCT with depletion of alloreactive αβ T cells and CD19+ B cells.
Table 1.
Clinical trials and outcomes after allo-HSCT with depletion of alloreactive αβ T cells and CD19+ B cells.
| Author | Diagnosis Patient number |
Type of donor | Conditioning | GVHD prophylaxis |
Graft composition | Survival | CI of aGVHD | CI of cGVHD |
| Bertaina A et al. Blood. 2014 | Children with both malignant and nonmalignant diseases (n=23) | haplo | MAC 30%, NMA 70%, according to the original disorder |
ATG (n=23) | CD34+ cells/kg: 15.8 × 106 (range: 10.4 × 106 to 40 × 106) TCR-αβ+CD3+ cells/kg: 4 × 104 (range: 1 × 104 to 9.5 × 104) |
2y TRM 9.3% (SE ±6.1); 2y DFS 91.1% (SE ±6.2); 2y OS 91.1% (SE ±6.2). |
Grade I–II: 13% Grade III–IV: 0% |
18 months: 0% |
| Gaziev J et al. Blood Adv. 2018 | Children with nonmalignant diseases (n=14) | haplo | MAC 100% | CSA + steroids (n=12); CSA + MMF (n=2) |
CD34+ cells/kg: 15.7 × 106 (range: 8.1 × 106 to 39.2 × 106) TCR-αβ+CD3+ cells/kg: 4 × 104 (range: 1 × 104 to 10 × 104) |
5y DFS 69% (95% CI: 37%–87%) 5y OS 84% (95% CI: 49%–96%) |
Grade II–III: 28% (95% CI, 1%–49%) |
extensive cGVHD: 21% (95% CI, 0% to 40%) |
| Laberko A et al. Blood. 2019 | Children with nonmalignant diseases (n=98) | MUD, haplo | MAC 74%, NMA 26%, |
ATG + CSA ± MTX/MMF (n=96) | CD34+ cells/kg: 10.5 × 106 (range: 6.3 × 106 to 14.9 × 106) in haplo TCR-αβ+CD3+ cells/kg: 1.4 × 104 (range: 0.5 × 104 to 13 × 104) in haplo |
5y OS 86% (95% CI: 76%–94%) in MUD; 5y OS 87% (95% CI: 73%–100%) in haplo |
Grade II–IV 17% (95% CI, 10%–28%) in MUD; Grade II–IV 22% (95% CI, 10%–47%) in haplo |
limited cGVHD 9% (95% CI, 4%–19%) in MUD; chronic GVHD 13% (95% CI, 5%–37%) in haplo |
| Locatelli F et al, Blood 2017 | Children with malignant diseases (n=80) | haplo | MAC 100% |
ATG (n=80) | CD34+ cells/kg: 13.9 × 106 (range: 6 × 106 to 40.4 × 106) TCR-αβ+CD3+ cells/kg: 4.7 × 104 (range: 0.2 × 104 to 9.9 × 104) |
5y NRM 5% (95% CI, 2%–13%); 5y DFS 71% (95% CI, 61%–81%); 5y OS 72% (95% CI, 62%–82%) |
Grade I–II 30% | limited cGVHD 5% |
| Maschan M et al. Bone Marrow Transplant. 2016 | Children with malignant diseases (n=33) |
MUD, haplo | MAC 100% |
ATG + tacrolimus + MTX (n=21), ATG + tacrolimus (n=5), ATG + MTX (n=2), ATG (n=5) |
CD34+ cells/kg: 9 × 106 (range: 3.8 × 106 to 21 × 106) TCR-αβ+CD3+ cells/kg: 2 × 104 (range: 0.29 × 104 to 12.6 × 104) |
2y TRM 10% (95% CI, 4%–26%); 2y DFS 60% (95% CI, 43%–76%); 2y OS 67% (95% CI, 50%–84%) |
Grade III–IV: 39% (95% CI, 26%–60%) |
cGvHD 30% (95% CI, 18–50)% |
| Prezioso L et al, 2019 | Adult with malignant diseases (n=59) |
haplo | MAC 100% |
ATG (n=59) | CD34+ cells/kg: 11 × 106 (range: 5 × 106 to 19 × 106) TCR-αβ+CD3+ cells/kg: 8.4 × 104 (range: 0.4 × 104 to 62 × 104) |
2y TRM 25%; 2y OS 51% |
Grade II–IV: 17% Grade III–IV: 3% |
limited cGvHD 3% |
| Kaynar L et al. Hematology. 2017 | Adult with malignant diseases (n=34) |
haplo | MAC 100% |
ATG (n=34) | CD34+ cells/kg: 12.7 × 106 (range: 10.3 × 106 to 16 × 106) TCR-αβ+CD3+ cells/kg: 1.8 × 104 (range: 0.7 × 104 to 2.5 × 104) |
2y DFS 33%; 2y OS 36% |
Grade I–IV: 30.3% Grade III–IV: 6.1% |
cGvHD 6.1% |
| de Witte MA et al, Blood Advances 2021 | Adult with malignant diseases (n=35) |
MRD, MUD | MAC 100% |
ATG + MMF (n=35) | CD34+ cells/kg: 6.1 × 106 (range: 2.8 × 106 to 19 × 106) TCR-αβ+CD3+ cells/kg: 2 × 104 (range: 0.0 × 104 to 9.0 × 104) |
2y NRM 32% (95% CI, 17%–48%); 2y DFS 40% (95% CI, 24%–56%); 2y OS 54% (95% CI, 37%–67%) |
Grade II–IV 26% (95% CI, 13%–41%); Grade III–IV 14% (95% CI, 5%–28%) |
2y cGvHD 23% (95% CI, 1%–38%) |
MAC: myeloablative conditioning regimen; NMA: non-myeloablative conditioning regimen; TRM: treatment-related mortality; DFS: disease-free survival; OS: overall survival; SE: standard error; NRM: non-relapse mortality.
Table 2.
Clinical trials and outcomes after haplo-HCT with CD45RA depleted progenitor cell grafts.
| Author | Diagnosis Patient number |
Conditioning | GVHD prophylaxis |
Graft composition | Survival | CI of aGVHD | CI of cGVHD | ||||||
| Naik S et al Blood 2021 |
Children with hematologic malignancies (n = 72) |
Submyeloablative conditioning |
MMF (n = 61) and/or sirolimus (n = 8) |
Day 0: CD34+ cells/kg: 9.85 × 106 (range: 1.96 × 106 to 44.64 × 106 ) Day +1: CD45RA-depleted graft: CD34+ cells/kg: 5.82 × 106 (range: 0.58 × 106 to 39.43 × 106 ) CD3+ T cells/kg: 60.1 × 106 (range: 16.08 × 106 to 528.43 × 106 ) CD3+CD45RA+ cells/kg: median 0, range 0-0.2 x10 6cells/kg). day+6: NK cells (median: 11.7 x10 6cells/kg; range: 1.65-99.2) |
3-y OS: 68.9% (95%CI: 58.9%– 80.6%) 3-y LFS: 62.2% (95% CI 51.9%– 74.6%) |
Grade II–IV: 36.1% (95% CI: 25.1%– 47.2%) Grade III or IV: 29.2% (95% CI: 19.1%–39.9%) | 3y: 20.8% (95% CI: 12.3%-30.9%) |
||||||
| Sisinni et al. Biology of BMT 2018 |
Children with acute leukemias (n = 25) |
Submyeloablative conditioning | CSA (n= 3), CSA+ MTX (n= 1), MMF (n = 21) |
CD34+ cells/kg: 6.29 × 106 (range: 4.04 × 106 to 18.1 × 106) CD45RA+ cells/kg: 0.6 × 104 (range: 0.2 × 104 to 1 × 104 ) |
30 months OS: 58% | Grade II–IV: 39% Grade III or IV: 33% | 30 months: 22% | ||||||
| Gasior et al. Transfusion 2021 |
Children with hematologic malignancies (n = 17) Severe aplastic anemia (n = 1) | Submyeloablative conditioning | MMF (n = 18) | CD34+ cells/kg: 6.5 × 106 (range: 5 × 106 to 11.2 × 106) CD3+CD45RA+ cells/kg: 3.6 × 104 (range: 0 to 23 × 104 ) Day +7: NK cells/kg: 12.6 × 106 (range: 3.9 × 106 to 100 × 106) |
2yOS: 87.2% (95% CI: 78.6%–95.8%) 2y EFS: 67.3% (95% CI: 53.1%–81.5%) 2-year GRFS: 64% (95% CI: 50.5%–78.1%) | At day +180: Gr III or IV: 34.8% (95% CI: 21.8%–47.8%) | At 1 year: 23.1% (95% CI: 11.4%–34.1%) | ||||||
| Clinical trials and outcomes after CD45RA-depleted Donor Lymphocyte Infusions after haploidentical HCT | |||||||||||||
| Authors | Diagnosis | Transplant platform | Indication, timing,cell dose used | survival | Acute GVHD | Chronic GVHD | |||||||
| Dunkaina et al. BMT 2021 | Children with hematologic malignancies (Total = 143. experimental arm, memory DLI, n = 76; control arm, n = 73 | TCR αβ depletion Myeloablative conditioning Haplo (n = 69) MUD (n = 6) MSD (n = 1) |
Prophylactic - day 0: 25 × 103 cell/kg, day 30,60,90,120 50 ×103 cell/kg Median number of DLI given = 4 (range:1-5) |
2y OS: 79% (95% CI: 69%–88%) 2y EFS: 72% (95% CI: 62%–83%) | Grade II–IV: 14.5% (95% CI:8%–25%) Grade III or IV: 8% (95% CI: 4%– 17%) |
2y: 6% (95% CI: 2%– 16%) | |||||||
| Naik et al. Blood 2021 | Children with acute leukemia (n = 30) |
TCR αβ depletion Submyeloablative conditioning | Prophylactic two weeks following engraftment: DL1: 1 × 105 cells/kg DL2: 1 × 106 cells/kg DL3: 1 × 107 cells/kg |
1y OS: 86.3% (95% CI: 74.6%–99.7%) 1-year EFS: 69.8% (95% CI: 55.2%– 88.4%) |
Grade II–IV: 26.7% (95% CI: 2.4%– 43.3%) Grade III or IV: 13.3% (95% CI: 4.1%–28.1%) | None | |||||||
| Castagna et al. Transplant Cell Ther. 2021 | Adults with hematologic malignancies (n = 19) | Post-transplant cyclophosphamide Myeloablative, reduced-intensity conditioning | Prophylactic DLI of 3 infusions each 4-6 weeks apart First dose given at median of 55 days (range, 46-63) post HCT DL1 5 × 105 cells/kg DL2 1 × 106 cells/kg DL3 5 × 106 cells/kg |
1y OS: 79% (95% CI: 45%–93%) 1y EFS: 75% (95% CI: 46%–90%) |
aGVHD I-IV 6% (95% CI: 0%–17%) | 1y: 15% (95% CI: 0%– 35%). | |||||||
Table 3.
Clinical trials and outcomes with NK cells.
| Author | Diagnosis Patient number |
Type of donor | Conditioning | GVHD prophylaxis |
NK Source and Timing of administration | Survival | CI of aGVHD | CI of cGVHD |
| Choi I et al. (2014) |
Adults with hematological malignancies, mostly AML (n =41) |
haplo | reduced-intensity conditioning |
Cyclosporin alone (n 13); Methotrexate + cyclosporin (n 28) |
Donor-derived NK cells Infusion 2 and 3 weeks after HCT Escalating doses (median dose of 2.0x108/kg) |
31.5 months EFS: 31% OS: 35% (refractory AML) EFS: 0% OS: 0% (refractory ALL/lymphoma) leukemia progression vs historical control cohort (46% vs 74%) |
22% (8 months after HCT) | 24% (3.3 months after HCT) |
| Lee DA et al. (2016) |
Children and adults with high-risk myeloid malignancies (AML, MDS, or CML refractory or beyond first remission)) (n=21) |
MSD: 13 (62%); MUD: 8 (38%) | regimen with busulfan and fludarabine | Thymoglobulin 1.5 mg/kg/day was given on days -3 to -1 (n 21); Tacrolimus started from day -2 and for 3 months; Methotrexate 5 mg/m2 was given on days 1, 2, 6, and 11. |
NK cells from an HLA haploidentical related donor (distinct from transplantation donor) Dose ranging from 0.02 to 8.32 ×106 /kg) Daily infusions for five days after conditioning |
relapse-free survival: 102 days; OS: 233 days; GVH- and Relapse-free: 89 days |
Grade I/II: n 5/21, Grade 3: n 2/21 (10%) |
n 6/21 |
| Ciurea SO et al. (2017) | Adults with high-risk myeloid malignancies (AML, MDS or CML with ≤ 5% bone marrow blasts) (n 13) |
haplo | RIC (melphalan-based) |
cyclophosphamide (50 mg/kg per day on days 13 and 14) + tacrolimus from day 15 and for 6 months + mycophenolate mofetil from day 15 and for 3 months | Membrane-bound IL21 expanded donor NK cells Doses ranging form 1 × 105/kg to to 3 × 1087kg) n 3 infusions before and after haploidentical HSCT (on days −2, +7, and +28). |
1 year OS: 92% 1 year DFS: 85% Relapse rate and overall mortality was not different than in conventional transplants |
Grade I/II: n 7/13 (54%) Grade III-IV: 0 |
0 |
| Berrien Elliot MM et al (2022) |
adult patients with high risk relapsed/refractory AML (n 15) |
haplo | RIC |
tacrolimus and mycophenolate mofetil starting on day +5 (until days +180 and +35, respectively) |
Same haploidentical donor memory like NK cells. Doses ranging from 0.5 ×106 to 10 × 106 cells/kg Infusion on day + 7 after transplant conditioned with a RIC regimen. Posttransplant cyclophosphamide on days +3 and + 4 IL 15 agonist administered subcutaneously on day +7 and over 3 weeks. |
1 year OS: 29% | grade I: 4/15 grade 2: 6/15 |
All grade:2/15 |
| Shapiro RM et al. (2022) |
relapsed myeloid malignancies relapsed after haploidentical HCT (AML, MDS, MDS/MPN, BPDC) (n 6) |
haplo | RIC | ATG + tacrolimus + MTX (n=21), ATG + tacrolimus (n=5), ATG + MTX (n=2), ATG (n=5) |
Donor-derived memory like NK cells Dose ranging from 5 to 10 million cells/kg Infusion after lymphodepleting chemotherapy Administration of IL2 (n 7 doses) |
All grade: 0 |
All grade: 0 |
Table 4.
Clinical trials and outcomes after haplo-HSCT with suicide gene therapy.
| Author | Diagnosis Patient number |
Type of donor | Conditioning | GVHD prophylaxis |
Graft composition | Survival | CI of aGVHD | CI of cGVHD |
| Ciceri F et al., Lancet Oncol 2009 | Adult with malignant diseases (n=50) 22 received TK cells |
haplo | MAC 100% | ATG (n=45) | CD34+ cells/kg: 11.6 × 106 (range: 4.6 × 106 to 16.8 × 106) CD3+ cells/kg: 1 × 104 (range: 0.26 × 104 to 10 × 104) |
3y NRM 49% (95% CI, 17%–63%); 3y OS 49% (95% CI, 25%–73%) |
Grade I-IV 45% (n=10/22) | Extensive cGvHD 4% (n=1/22) |
MAC: myeloablative conditioning regimen; NMA: non-myeloablative conditioning regimen; TRM: treatment-related mortality; DFS: disease-free survival; OS: overall survival; SE: standard error; NRM: non-relapse mortality.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



