
Submitted:
23 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Marine Protected Areas (MPAs) are vital for biodiversity conservation, yet their effectiveness in preserving foundation seaweeds remains understudied. This study investigates the diversity and distribution of Cystoseira sensu lato (including Cystoseira, Ericaria and Gongolaria) populations in the an MPA located in the central Mediterranean Sea, comparing them with those in two unprotected control sites. We hypothesize that MPA Cystoseira s.l. populations will display higher diversity and structure compared to outside control sites. Results reveal a total of 19 Cystoseira s.l. species at depths of 0-20 m, with the MPA exhibiting a higher diversity than control sites. Thus, MPAs can play a crucial role in fostering the diversity of Cystoseira s.l. populations. However, no significant differences were observed among the MPA's protection zones, raising questions about the zoning effectiveness. Additionally, our survey uncovered a substantial presence of non-indigenous seaweeds within the MPA. In conclusion, while MPAs improve Cystoseira s.l. diversity compared to unprotected sites, the varying efficacy of protection within MPA zones suggests a necessity for site-specific conservation strategies. The presence of non-indigenous seaweeds emphasizes ongoing challenges. This study provides a baseline for understanding Cystoseira s.l. population dynamics, crucial for future monitoring and conservation efforts in the face of climate change.
Keywords:
macroalgal distribution
; Cystoseira sensu lato
; Cystoseira
; Ericaria
; Gongolaria
; Biodiversity conservation
; Marine Protected Areas
; Mediterranean Sea
1. Introduction
Marine forest seaweeds, mainly brown algae of the orders Fucales and Laminariales, form some of the most productive and diverse habitats on the world’s cold-water and temperate rocky reefs [1,2,3,4]. In the Mediterranean Sea, these foundation seaweeds mainly comprise species of the genus Cystoseira C. Agardh (Fucales, Phaeophyceae), recently divided into three genera: Cystoseira, Gongolaria Boehmer and Ericaria Stackhouse [5,6] and hereafter referred to as Cystoseira s.l. (sensu lato). Cystoseira s.l. forests provide a wide range of ecosystem services, supporting a rich associated biota, including fish, invertebrates, and other algae [2,7,8]. They are essential foundation species, significantly enhancing the structural complexity and productivity of coastal communities from the surface down to the upper circalittoral zone [9,10,11,12]. Additionally, they are considered useful indicators of water and ecosystem quality according to the Water Framework Directive (2000/60/EC) and the Marine Strategy Framework Directive (2008/56/EC).
Like other coastal marine ecosystems, these long-lived primary producers are highly impacted, particularly close to urban areas, due to combined effects of anthropogenic impacts such as pollution, urbanization, the introduction of non-native species, overfishing, coastal aquaculture, and climate-change [2,13,14,15,16,17,18,19]. Consequently, the decline or loss of Cystoseria s.l. populations have been observed on many rocky coasts, leading to a shift from complex and productive benthic communities to less structured assemblages dominated by turf-forming algae, mussels, or sea urchin barrens [1,2,13,15,18,20,21,22,23]. This shift results in a decrease in essential ecosystem services [2,24] and economic value of coastal areas [25].
Marine Protected Areas (MPAs) have become highly important tools for biodiversity conservation and management worldwide [26]. In the Mediterranean Sea, a large part of the research concerning the effectiveness of MPAs has focused on fish assemblages [27], while few studies focused on foundation seaweeds [14,28,29]. MPAs can play a critical role in the conservation of Cystoseira s.l. forests, as they guarantee protection from various anthropogenic impacts [28]. Healthy Cystoseira s.l. forests in MPAs may also represent an important source of propagules for the restoration of lost or degraded forests outside MPA borders, a crucial aspect due to the low dispersal capacity of most of these species [14,21,30,31].
In this study, we investigate the diversity and distribution of Cystoseira s.l. populations within the “Capo Gallo - Isola delle Femmine” MPA in the central Mediterranean Sea. Moreover, we tested the effectiveness of the MPA for Cystoseira s.l. conservation by comparing these populations within the MPA to those in two unprotected control sites (Figure 1). We hypothesize that Cystoseira s.l. populations will be more diverse and structured within the MPA compared to outside control sites, and that the areas within the MPA characterized by higher levels of protection will host highly diverse populations.
2. Results
A total of 19 Cystoseira s.l. species were found between 0 to 20 m of depth in the surveyed sites (Table 1 and Table 2). The highest number (17 species) was found within the “Capo Gallo – Isola delle Femmine” MPA, while Monte Cofano and Punta Barcarello hosted 9 and 5 species, respectively (Table 1).
2.1. Cystoseira s.l. assemblage within MPA
Within the MPA, 10 Cystoseira s.l. species were found into zone A, while 14 were found in zone B and C (Table 1), with 6 species (Cystoseira compressa, Cystoseira humilis var. myriophilloides, Ericaria amentacea, Ericaria brachycarpa, Ericaria crinita, and Gongolaria montagnei var. compressa) consistently found across different levels of protection.
Whitin the MPA, zone A exhibited lover abundances (average cover percentage) and a lower number of Cystoseira s.l. species compared to the other two levels of protection (B and C) (Figure 2 a-b). The values for these two metrics differed between zone B and C, with zone B showing slightly lower average abundance but higher number of species compared to zone C (Figure 2 a-b). Shannon diversity and Pielou’s evenness were higher in zone A and B compared to zone C (Figure 2 c-d).
The Cystoseira s.l. assemblage differed significantly among the different MPA zones (PERMANOVA results model: F2,17 = 55.027 and 40.394 for structure and composition, respectively, p < 0.001; Table S1), explaining a substantial amount of the total variation (R2 = 70.73% and 64.27% for structure and composition, respectively). Additionally, there was a significant difference among sites within MPA zones (PERMANOVA results model: F3,17 = 10.706 and 10.711 for structure and composition, respectively, p < 0.001; Table S1), accounting for a moderate amount of the total variation (R2 = 20.64% and 25.56% for structure and composition, respectively; Table S1). No significant difference was found for transects within sites and MPA zones (Table S1). The Principal Coordinate Analysis (PCoA) ordination plot clearly separated the structure of the Cystoseira s.l. assemblage among MPA zones, with zone A at a far distance from zone B, while sites in zone C placed among the other two protection zones, with Punta Barcarello being closer to sites in zone B and Punta Matese to zone A (Figure 3 a). Cystoseira s.l. composition, instead, clearly separated zone A from B and C, with partial overlap of the Cystoseira s.l. composition of the site of La Cala (within zone B) with sites of zone B (Figure 3 b).
2.2. Comparison of Cystoseira s.l. assemblage between MPA and control sites
When comparing the MPA with control sites, urban and white sites exhibited higher average Cystoseira s.l. abundance, comparable to zones C and B within the MPA rather than zone A (Figure 4 a). The species richness of the white and urban sites was comparable to that within the MPA, with the white site being closer to zone C, while the urban site showed lower values more comparable to those of zone A (Figure 4 b). Shannon diversity and Pielou’s evenness of the white site were comparable to those of zone C, while those of the urban site showed lower values, although no significant differences were detected (Figure 4 c-d).
When comparing the Cystoseira s.l. assemblage between the MPA zone and control sites (Table S2), it was revealed that there were significant differences at the urban site compared to both the MPA’s and white sites (Figure 5, a-b), with an average dissimilarity of 71% and 60% for structure and composition, respectively (Table S3). On the other hand, the Cystoseira s.l. assemblage of the white site was more similar to the MPA’s sites (Figure 5 a-b) with an average similarity of 55% and 59% for structure and composition, respectively. It was closer to the Punta Matese site (zone C) with an average similarity of 64% and 67% for structure and composition, respectively (Table S3).
Here, we provide an overall description of Cystoseira s.l. populations across the investigated sites. Of the 19 species discovered, Cystoseira humilis showed the large depth adaptability (range depth = 0-20 m) followed by four species (Cystoseira foeniculacea f. tenuiramosa, Gongolaria montagnei, Gongolaria montagnei var. tenuior and Gongolaria sauvageauana) found between 5 and 20 meters depth (Table 2). At all investigated sites, E. amentacea grows on the outer margin of vermetid where forms dense and continuous belts across the MPA’s sites and the Monte Cofano (white) site; however, this species was not found at Punta Priola (urban) site. Sparse thalli of Cystoseira compressa f. rosetta were also found at the sites of Isola and Punta Barcarello, while C. compressa was present within both the MPA and the two control sites. Cystoseira compressa formed either small dense patches with thalli of about 10 cm high (without aerocystis) or individual thalli up to 40 cm high with branches provided by aerocystis. Between 1 and 5 meters depth, E. brachycarpa forms large and extended forests within the MPA at the sites of Punta Barcarello, La Cala, Isola, whereas at Punta Matese, Barcarello, and Capo Gallo, despite being dense, the populations assume a discontinuous pattern due to the presence of large rocky boulders placed on rocky carbonate platforms. Dense and continuous populations of E. brachycarpa were also found at Monte Cofano, whereas at Punta Priola E. brachycarpa stands were patchy and less dense, intercalated by thalli of Dictyopteris polypodioides (De Candolle) J. V. Lamouroux. Ericaria crinita was the second most common species discovered between 1 and 5 meters depth. At the sites of Barcarello, Punta Barcarello and Capo Gallo, this species forms dense patches covering big boulders or, as in the other sites, its thalli are widely spread on the substrates and surrounded by other macroalgae. Cystoseira humilis was found at the site of La Cala and Monte Cofano as individual sparse thalli or dense patches. Finally, only at the site of Monte Cofano sparse thalli of E. mediterranea were found.
Between 5 and 10 meters depth, G. montagnei var. tenuior was the most abundant which was found within the MPA at the sites of La Cala, Punta Matese, Punta Barcarello and Barcarello, and Monte Cofano. This species forms extensive dense forests, which can be seen in particular in La Cala and Punta Barcarello. In the other sites, G. montagnei var. tenuior grows in small patches or as individual thalli surrounded by other macroalgae. Cystoseira humilis var. myriophylloides on the other hand, was only found within the MPA at Isola, La Cala, Punta Matese, and Barcarello as isolated or groups of few thalli surrounded by other seaweeds, whereas C. foeniculacea was found only at the sites of Barcarello and Punta Priola.
The larger part of Cystoseira s.l. species (7 species) was found between 10 and 20 meters depth (Table 2). Ericaria funkii was only found within the MPA at the sites of La Cala, Punta Matese, Punta Barcarello and Barcarello. This species has a green iridescence and was found as groups of 2-3 individuals so close to be perceived as a single big individual. Cystoseira foeniculacea f. latiramosa and C. foeniculacea f. tenuiramosa were found as scattered individual thalli within the MPA, the first one at the sites of La Cala, Punta Matese, and Barcarello, and the second at La Cala, Punta Matese, Punta Barcarello and Barcarello. Cystoseira dubia was discovered in small patches within the MPA (Isola, La Cala and Punta Barcarello) in areas with high sedimentation, with flattened primary branches of light brown color emerging from the sediment. Finally, E. brachycarpa var. claudiae was found as isolated individual thalli within the MPA (Punta Barcarello, Barcarello and Capo Gallo) and at the controlled site of Punta Priola (Table 2).
2.3. Non-indigenous seaweeds
During data collection within both the MPA and control sites, the presence of non indigenous seaweeds was observed, regardless of the level of protection. In particular, four species were discovered: Asparagopsis taxiformis (Delile) Trevisan, Caulerpa cylindracea Sonder 1845, Caulerpa taxifolia var. distichophylla (Sonder) Verlaque, Huisman & Procaccini and Lophocladia lallemandii (Montagne) F. Schmitz (Figure 6). Thalli of A. taxiformis (Figure 6A) can be found across the entire “Capo Gallo - Isola delle Femmine” MPA, regardless of the protection zone and depth range. The species was discovered from 1m to 20 m depth with a particularly high density in the Isola delle Femmine site. Moreover, A. taxiformis was found as epiphyte on Cystoseira s.l. species, especially G. montagnei var. tenuior. Caulerpa cylindracea (Figure 6B) was also observed across the MPA and does not appear to have a preferred depth. It can be found from the mediolittoral zone (intertidal rocky pools, vermetid reef cuvettes) down to the 20 m. While stolons can attain enormous densities, the vertical frond of the alga is not always apparent. Caulerpa taxifolia var. distichophylla (Figure 6C) was found between 0 and 10 m at Punta Barcarello and Barcarello, particularly on rocky substrate covered with sediment. Finally, blooms of L. lallemandii (Figure 6D) were detected over the MPA during the summer, capable of completely covering vast areas of substrate and all the seaweeds inhabiting them. This species has also been found to epiphyte G. montagnei var. tenuior.
3. Discussion
According to our surveys, the MPA had more diverse Cystoseira s.l. populations in terms of number of species (Figure 7) than outside control sites (Figure 8), confirming the hypothesis that MPAs can be an effective strategy for preserving and restoring these important foundation seaweeds [14,28,29]. Although this observation remains true when comparing Cystoseira s.l. populations across the MPA and external sites, our findings demonstrate no variations among the various degrees of protection within the MPA. Then, our initial hypothesis posited that the effects of protection within the MPA would lead to significant differences in the patterns of structuring and abundance of these communities between areas with total protection and those with less protection and control, was thus unfounded.
Studies have shown that some Mediterranean MPAs are unable to protect or restore Fucalean algal forests, implying that some of them are merely “paper parks” where regulations are not enforced [29,33]. We believe that the lack of efficacy in protection observed within the MPA is due to the fact that the different zones within the MPA were designed primarily to protect fish stocks rather than Fucalean algal forests. Moreover, the observed differences between the zones can be explained by variations in seabed geomorphic features across MPA sites. In fact, our observations suggest that when the seabed consists of a gently sloping rocky carbonate platform, as seen at the MPA sites of La Cala and Punta Barcarello, we find more continuous and dense Cystoseira s.l. populations compared to other sites, such as Capo Gallo and Barcarello, where the presence of large scattered rocky boulders (Riggio and Raimondo, 1991; Lucido et al., 1992) creates a discontinuous environment with heterogeneous light conditions, where small patches and individual thalli of Cystoseira s.l. are most common.
Data on Cystoseira s.l. species distribution within the “Capo Gallo – Isola delle Femmine” MPA are scarce and date back at least thirty years, making them only partially helpful for comparison. Giaccone and Sortino (1964) reported the presence of C. compressa, E. mediterranea, E. crinita and Gongolaria barbata on the seabed of Isola. Of these, E. mediterranea and G. barbata were not found in our surveys. In 1985, data from G. barbata e G. montagnei were reported at Capo Gallo and Isola [36], while more recent data reported E. amentacea, Ericaria crinita and G. montagnei at the sites of Punta Barcarello and Capo Gallo [37,38]. The lack of historical data on Cystoseira s.l. species distribution within the MPA emphasizes the significance of our study as a baseline for understanding how these populations will change in the future.
The comparison with unprotected control sites revealed differences that were more evident when comparing the MPA sites with the Punta Priola site than with the site of Monte Cofano. The vegetation in Punta Priola was mainly characterized by high sedimentation rates, with Dictyopteris polypodioides being the main macroalgal species covering large part of the rocky substrate, which appears to inhibit the growth of other seaweeds (Figure 9B). The absence of E. amentacea, one of the most important Cystoseira s.l. species used to measure water quality, further revealed the site’s impacted state [39]. In contrast, our findings demonstrate that Cystoseira s.l. populations are comparable between the control site of Monte Cofano and the sites within the “Capo Gallo - Isola delle Femmine” MPA (Figure 9A). This indicates that anthropogenic disturbance, such as trampling, harvesting, pollution, and overgrazing, are fairly limited at Monte Cofano, and confirm that healthy dense forests can still be found in non-protected, naturally isolated, and lightly disturbed locations [29].
In addition to the above observations, it is important to mention the presence of four non-indigenous seaweeds (A. taxiformis , C. cylindracea, C. taxifolia var. distichophylla, and L. lallemandii) within the MPA and at the control sites. Non-indigenous species are one of the major threats to the Mediterranean Sea [40,41] Asparagopsis taxiformis has been named one of the top 100 invasive seaweeds in the Mediterranean Sea [41]. The presence of A. taxiformis need further investigation because it has negative effects on Cystoseira s.l. populations, eroding biomass of primary producers and the associated biodiversity [42]. In accordance with other research [43,44,45], C. cylindracea was largely observed within the MPA area across different habitats and types of substrate, regardless of the levels of protection. It was found across all the investigated depth range, but thalli were also found at depths of 35 m (Mancuso’s personal observations). This suggest that although MPAs are a useful management tool for the protection of biodiversity, they are still vulnerable to non-indigenous seaweeds [45,46]. The presence of Caulerpa cylindracea can have a negative impact on native seaweeds assemblages, also facilitating the subsequent invasion of a trophic specialist that takes advantage of niche opportunities created by the alga [47,48,49]. Finally, L. lallemandii can affect the structure of macrofauna associated with habitat forming seaweeds of Cystoseira s.l. [46], or cause the mortality of seagrasses [50].
4. Materials and Methods
The study was performed on the shallow rocky substrate (0 to 20 m depth) within the “Capo Gallo-Isola delle Femmine” MPA (Lat: 38.213961, Long: 13.277121) and two control sites, Monte Cofano (Lat: 38.114429, Long: 12.677827) and Punta Priola (Lat: 38.192074, Long: 13.358161), located in the northwestern coast of Sicily, Italy (Figure 1), which are not subjected to marine protection.
4.1. The “Capo Gallo – Isola delle Femmine” MPA
The “Capo Gallo-Isola delle Femmine” MPA, established in 2002 by the Italian Ministry of Environment and Protection of Land and Sea, affects the sea stretch between the towns of Palermo and Isola delle Femmine. Covering approximately 22 km2 of sea area and a coastline of about 16 km, it is bounded to the east by the gulf of Mondello and to the west by the bay of Carini. An imposing calcareous dolomitic mountain crest (Capo Gallo, 562 m a.s.l.) defines the coastal strip, resulting in a steep and rocky coastal morphology. Due to the limestone nature, flowing waters generate karst phenomena, leading to caves of high naturalistic value (Grotta dell’Olio and Grotta della Mazzara). Only towards the western part, the rocky coast assumes a flat conformation, enlivened by the presence, about 300 meters from the mainland, of the Isola delle Femmine (also known as Isola di Fuori), an isolated vestige of the aforementioned calcareous ridge.
The MPA is divided into three main zones (A, B and C), each with its own level of environmental protection (Figure 1). There are two no-take/no-access zones (zone A, total area of 1 km2, Figure 1), one in the north sector of Isola delle Femmine and the other in the stretch of sea at the west of Capo Gallo promontory, between the Puntazza and the Capo Gallo lighthouse. Zones B and C are buffer zones where human use restrictions, including fishing, become progressively lower. In particular, there are three general protection zones (zone B, total area of 2 km2, Figure 1), while the remaining sea within the MPA’s border includes a partial protection area (zone C, total area of 19 km2, Figure 1). The MPA area is also identified as Site of Community Importance (SCI, ITA020047 – Fondali di Isola delle Femmine Capo Gallo).
4.2. Control sites
To analyze the efficiency of MPA protection, we chose two unprotected marine regions: one natural and less impacted site named Monte Cofano and one highly impacted site close to the urban center called Punta Priola (Figure 1). The Monte Cofano site is located in front of the coast of the natural terrestrial reserve of the Monte Cofano promontory, near Custonaci and San Vito Lo Capo. Although it is not a marine reserve, its status as a terrestrial reserve offers some level of protection by restricting access from the land. Furthermore, there are no large urban areas nearby, and pollution is virtually absent. However, there are no restrictions on marine activities (like fishing or harvesting) in the area. The Monte Cofano site can then be considered an unaltered control site based on its characteristics (white site). The site of Punta Priola was located between Mondello and Palermo. This site is clearer influenced by several anthropogenic stressors (urban site). The shoreline is densely developed, with small untreated outfalls (particularly in summer), and the little Rousvelt harbor located approximately 300 meters west of the site. Moreover, the site attracts bathers who pour onto the seashore, resulting in an increase in stressors from trampling and harvesting activities.
4.3. Survey of Cystoseira s.l. populations
Scuba diving surveys were conducted at 6 sites within the “Capo Gallo - Isola delle Femmine” MPA, representing a large part of the MPA and the 3 levels of protection (2 sites for each level of protection), while surveys were carried out at 1 site in the control sites. The sites were mostly exposed to northwest winds and had a similar seabed environment with carbonate platforms and rocky substrates. At each site, 3 belt transects [51] from 0 to 20 meters depth were used to determine the distribution of Cystoseira s.l. species. Transect length changed according to seabed degradation, while width was 6 m (3 m left and right the transect). For each bathymetric range (0-5 m, 5-10 m, 10-15 m and 15-20 m), cover percentage of Cystoseira s.l. species were estimated in 4 quadrats (50 x 50 cm) haphazardly selected. During each dive, the water visibility was at least 10 meters, allowing easily identification of Cystoseira s.l. thalli. The depth range was chosen to allow safe scuba diving (diving constraints such as decompression schedules and air consumption normally limit depths to less than 20 m) and to give access to the majority of Cystoseira s.l. species.
All surveys were carried out in May, when the thalli of Cystoseira s.l. species in this area reach their maximum development [52]. Furthermore, pictures of the landscape were acquired to describe the status of the Cystoseira s.l. populations. Collection of thalli was limited to species that were difficult to identify in the field. Sampled thalli were deposited in the algological laboratory (Department STEBICEF - University of Palermo).
4.4. Data analysis
For each investigated depth range, the abundance (N, average percentage cover), frequency (F%, the percentage of samples in which a particular species is present), and dominance index (D%, the percentage ratio of individuals of a particular species to the total number of individuals within the sample) for each Cystoseira s.l. species were estimated [53]. Additionally, for each area investigated (MPA’s zone and control sites), Cystoseira s.l. species were characterized based on total abundance (N; expressed as cover percentage), rarefied species richness (S), Shannon-Wiener diversity index (H′), and Pielou’s Evenness index (J).
Analyses of variance (ANOVAs) were used to test: i) differences in the Cystoseira s.l. indices (N, S, H′, J) among the three protection zones (fixed factor with 3 levels: zone A, zone B and zone C) within the MPA; ii) differences among zones within the MPA and external control sites (fixed factor with 5 levels: zone A, zone B, zone C, white and urban). Besides the main factor in each analysis, ANOVAs included the factors site (random factor nested within zone) and transect (random factor nested within site and zone).
Louvain community detection [54] was performed to detect the Cystoseira s.l. set for each considered depth range. Differences in the Cystoseira s.l. structure (which takes into account species identity and relative abundance) and composition (presence/absence, which only takes into account species identity) among sites were assessed by Permutational Multivariate Analysis of Variance (PERMANOVA). The analyses were based on a Bray-Curtis distance matrix of square-root transformed cover percentage of Cystoseira s.l. using 9999 permutations [55]. A principal coordinate analysis (PCoA) plot was generated to visualize the variation in Cystoseira s.l. assemblage structure (based on a Bray-Curtis distance matrix) and composition (based on Jaccard distance matrix).
Statistical analyses were carried out in R open access statistical software version 4.1.2 [56].
5. Conclusions
Our findings emphasize the importance of the “Capo Gallo - Isola delle Femmine” MPA as a valuable tool for Fucalean forests of the genera Cystoseira, Ericaria and Gongolaria, as well as a good reference for monitoring the temporal evolution of these foundation seaweeds. Further research should be conducted to gain more insights into Cystoseria s.l. species present in the MPA, particularly at depths greater than 20 meters. Up until now, data on deeper Cystoseira s.l. species within the “Capo Gallo – Isola delle Femmine” MPA come from point-like observations. In particular, we have observed the presence of Ericaria zosteroides (C.Agardh) Molinari & Guiry and G. montagnei var. compressa were at a depth of 35 to 40 meters at the site of Isola and the presence of G. barbata (Stackhouse) Kuntze at a depth of around 1 meter in the port of Isola delle Femmine. This information remarks the MPA’s role in preserving diverse Cystoseira s.l. populations. It also emphasizes the importance of doing extensive habitat mapping of these key foundation seaweeds to monitor their range and health status. This is crucial in understanding how these valuable foundation species will respond to the effects of climate change.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: - PERMANOVAs results of the structure and composition of the Cystoseira s.l. assemblage among the MPA’s zones; Table S2: PERMANOVAs results of the structure and composition of the Cystoseira s.l. assemblage among all areas investigated (MPA’s zones and control sites).; Table S3: Average similarity of the Cystoseira s.l. assemblage between and within the different zones investigated.
Author Contributions
Conceptualization, F.P.M. and A.M.; Methodology, F.P.M. and A.M.M.; Data curation, F.P.M.; Investigation, F.P.M. and A.M.M; Visualization, F.P.M.; Laboratory Analyses, F.P.M. and A.M.M; Writing – original draft, F.P.M. and A.M.M; Writing – review & editing, F.P.M., A.M.M and G.S.;. Supervision, G.S. and A.M.M.; Funding acquisition, G.S. and A.M.M.
Funding
This research was funded by the Ministero dell’Ambiente e della Tutela del Territorio e del Mare through the project “AMPOCYS” (nota Prot. N. 29962 del 13/12/07), by the Ministero della transizione ecologica (MiTE) through the projects “Realizzazione di misure di conservazione del sito di interesse comunitario ITA 020047 “Fondali Isola delle Femmine – Capo Gallo” and “Valutazione dello stato di conservazione della vegetazione algale nell’AMP di Capo Gallo – Isola delle Femmine and Ministero delle politiche agricole alimentari, forestali e del turismo (Mipaaft) FEAMP PO 2014- 2020 Fondo europeo per gli affari marittimi e la pesca Misura: “1.40 - Protezione e ripristino della biodiversità e degli ecosistemi marini e dei regimi di compensazione nell’ambito di attività di pesca sostenibili.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global Patterns of Kelp Forest Change over the Past Half-Century. Proc. Natl. Acad. Sci. 2016, 113, 13785–13790. [Google Scholar] [CrossRef]
- Mineur, F.; Arenas, F.; Assis, J.; Davies, A.J.; Engelen, A.H.; Fernandes, F.; Malta, E.; Thibaut, T.; Van Nguyen, T.; Vaz-Pinto, F.; et al. European Seaweeds under Pressure: Consequences for Communities and Ecosystem Functioning. J. Sea Res. 2015, 98, 91–108. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J. a.; Tegner, M.J. Kelp Forest Ecosystems: Biodiversity, Stability, Resilience and Future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Coleman, M.A.; Wernberg, T. Forgotten Underwater Forests: The Key Role of Fucoids on Australian Temperate Reefs. Ecol. Evol. 2017, 7, 8406–8418. [Google Scholar] [CrossRef]
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira Sensu Lato (Fucales, Phaeophyceae) in the Eastern Atlantic and Mediterranean Based on Morphological and DNA Evidence, Including Carpodesmia Gen. Emend. and Treptacantha Gen. Emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Molinari Novoa, E.A.; Guiry, M.D. Reinstatement of the Genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Not. Algarum 2020, 172, 1–10. [Google Scholar]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.M.; Francour, P. Nursery Value of Cystoseira Forests for Mediterranean Rocky Reef Fishes. J. Exp. Mar. Bio. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Schiel, D.R.; Foster, M.S. The Population Biology of Large Brown Seaweeds: Ecological Consequences of Multiphase Life Histories in Dynamic Coastal Environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Giaccone, G.; Alongi, G.; Pizzuto, F.; Cossu, A.V.L. La Vegetazione Marina Bentonica Fotofila Del Mediterraneo: 2. Infralitorale e Circalitorale: Proposte Di Aggiornamento. Boll. dell’Accademia Gioenia di Sci. Nat. 1994, 27, 111–157. [Google Scholar]
- Falace, A.; Bressan, G. Seasonal Variations of Cystoseira Barbata (Stackhouse) C. Agardh Frond Architecture. Hydrobiologia 2006, 555, 193–206. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Acunto, S.; Cinelli, F.; Hawkins, S.J. The Influence of Canopy Algae on Vertical Patterns of Distribution of Low-Shore Assemblages on Rocky Coasts in the Northwest Mediterranean. J. Exp. Mar. Bio. Ecol. 2002, 267, 89–106. [Google Scholar] [CrossRef]
- Ballesteros, E.; Garrabou, J.; Hereu, B.; Zabala, M.; Cebrian, E.; Sala, E. Deep-Water Stands of Cystoseira Zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: Insights into Assemblage Structure and Population Dynamics. Estuar. Coast. Shelf Sci. 2009, 82, 477–484. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Pannacciulli, F.; Bulleri, F.; Moschella, P.S.; Airoldi, L.; Relini, G.; Cinelli, F. Predicting the Consequences of Anthropogenic Disturbance: Large-Scale Effects of Loss of Canopy Algae on Rocky Shores. Mar. Ecol. Prog. Ser. 2001, 214, 137–150. [Google Scholar] [CrossRef]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-Term Decline of the Populations of Fucales (Cystoseira Spp. and Sargassum Spp.) in the Albères Coast (France, North-Western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef] [PubMed]
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Loss of Fucoid Algae along a Gradient of Urbanisation, and Structure of Benthic Assemblages. Mar. Ecol. Prog. Ser. 2008, 358, 63–74. [Google Scholar] [CrossRef]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The Ups and Downs of a Canopy-Forming Seaweed over a Span of More than One Century. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of Vulnerable Cystoseira Populations along the Italian Infralittoral Fringe, and Relationships with Environmental and Anthropogenic Variables. Mar. Pollut. Bull. 2018, 129, 762–771. [Google Scholar] [CrossRef]
- Strain, E.M.A.; Thomson, R.J.; Micheli, F.; Mancuso, F.P.; Airoldi, L. Identifying the Interacting Roles of Stressors in Driving the Global Loss of Canopy-Forming to Mat-Forming Algae in Marine Ecosystems. Glob. Chang. Biol. 2014, 20, 3300–3312. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, R.; Pinedo, S.; Ballesteros, E.; Arévalo, R.; Pinedo, S.; Ballesteros, E. Changes in the Composition and Structure of Mediterranean Rocky-Shore Communities Following a Gradient of Nutrient Enrichment: Descriptive Study and Test of Proposed Methods to Assess Water Quality Regarding Macroalgae. Mar. Pollut. Bull. 2007, 55, 104–113. [Google Scholar] [CrossRef]
- Falace, A.; Alongi, G.; Cormaci, M.; Furnari, G.; Curiel, D.; Cecere, E.; Petrocelli, A. Changes in the Benthic Algae along the Adriatic Sea in the Last Three Decades. Chem. Ecol. 2010, 26, 77–90. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfune, A.; Boudouresque, C.F.; Verlaque, M. Decline and Local Extinction of Fucales in French Riviera: The Harbinger of Future Extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Connell, S.D.; Foster, M.S.; Airoldi, L. What Are Algal Turfs? Towards a Better Description of Turfs. Mar. Ecol. Prog. Ser. 2014, 495, 299–307. [Google Scholar] [CrossRef]
- Agnetta, D.; Badalamenti, F.; Ceccherelli, G.; Di Trapani, F.; Bonaviri, C.; Gianguzza, P. Role of Two Co-Occurring Mediterranean Sea Urchins in the Formation of Barren from Cystoseira Canopy. Estuar. Coast. Shelf Sci. 2015, 152, 73–77. [Google Scholar] [CrossRef]
- Buonomo, R.; Chefaoui, R.M.; Lacida, R.B.; Engelen, A.H.; Serrão, E.A.; Airoldi, L. Predicted Extinction of Unique Genetic Diversity in Marine Forests of Cystoseira Spp. Mar. Environ. Res. 2018, 138, 119–128. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, G.; Asnaghi, V.; Chiantore, M.; Thrush, S.; Povero, P.; Vassallo, P.; Petrillo, M.; Paoli, C. The Effect of Cystoseira Canopy on the Value of Midlittoral Habitats in NW Mediterranean, an Emergy Assessment. Ecol. Modell. 2019, 404, 1–11. [Google Scholar] [CrossRef]
- Lester, S.E.; Halpern, B.S.; Grorud-Colvert, K.; Lubchenco, J.; Ruttenberg, B.I.; Gaines, S.D.; Airamé, S.; Warner, R.R. Biological Effects within No-Take Marine Reserves: A Global Synthesis. Mar. Ecol. Prog. Ser. 2009, 384, 33–46. [Google Scholar] [CrossRef]
- Blowes, S.A.; Chase, J.M.; Di Franco, A.; Frid, O.; Gotelli, N.J.; Guidetti, P.; Knight, T.M.; May, F.; McGlinn, D.J.; Micheli, F.; et al. Mediterranean Marine Protected Areas Have Higher Biodiversity via Increased Evenness, Not Abundance. J. Appl. Ecol. 2020, 57, 578–589. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Cottalorda, J.M.; Hereu, B.; Susini, M.L.; Verlaque, M. Unexpected Temporal Stability of Cystoseira and Sargassum Forests in Port-Cros, One of the Oldest Mediterranean Marine National Parks. Cryptogam. Algol. 2016, 37, 61–90. [Google Scholar] [CrossRef]
- Sala, E.; Ballesteros, E.; Dendrinos, P.; Di Franco, A.; Ferretti, F.; Foley, D.; Fraschetti, S.; Friedlander, A.; Garrabou, J.; Güçlüsoy, H.; et al. The Structure of Mediterranean Rocky Reef Ecosystems across Environmental and Human Gradients, and Conservation Implications. PLoS One 2012, 7, e32742. [Google Scholar] [CrossRef]
- Falace, A.; Kaleb, S.; De La Fuente, G.; Asnaghi, V.; Chiantore, M. Ex Situ Cultivation Protocol for Cystoseira Amentacea Var. Stricta (Fucales, Phaeophyceae) from a Restoration Perspective. PLoS One 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Verdura, J.; Sales, M.; Ballesteros, E.; Cefalì, M.E.; Cebrian, E. Restoration of a Canopy-Forming Alga Based on Recruitment Enhancement: Methods and Long-Term Success Assessment. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 22 August 2022).
- Meinesz, A.; Blanfuné, A. 1983-2013: Development of Marine Protected Areas along the French Mediterranean Coasts and Perspectives for Achievement of the Aichi Target. Mar. Policy 2015, 54, 10–16. [Google Scholar] [CrossRef]
- Riggio, S.; Raimondo, F.M. Proposta Di Una Riserva Costiera per La Tutela e La Valorizzazione Dei Biotopi Di Isola Delle Femmine e Di Monte Gallo (Palermo). Quad. di Bot. Appl. 1991, 2, 59–96. [Google Scholar]
- Lucido, M. Geomorfologia Della Piattaforma Continentale Tra Torre Del Pozzillo e Torre Mondello (Sicilia Nord-Occidentale). Nat. Sicil. 1992, 91–107. [Google Scholar]
- Giaccone, G.; Colonna, P.; Graziano, C.; Mannino, A.M.; Tornatore, E.; Cormaci, M.; Furnari, G.; Scammacca, B. Revisione Della Flora Marina Di Sicilia e Isole Minori. Boll. dell’Accademia Gioenia di Sci. Nat. 1985, 18, 537–781. [Google Scholar]
- Mannino, A.M. Studio Fitosociologico Della Vegetazione Mesolitorale a Lithophyllum Lichenoides Philippi (Rhodophyceae, Corallinales). Nat. Sicil. 1992, 16, 3–25. [Google Scholar]
- Mannino, A.M.; Mancuso, F.P. Guida All’identificazione Delle Cistoseire (Area Marina Protetta “Capo Gallo-Isola Delle Femmine”), Palermo, 2009.
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A New Methodology Based on Littoral Community Cartography Dominated by Macroalgae for the Implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Zenetos, A.; Galanidi, M. Mediterranean Non Indigenous Species at the Start of the 2020s: Recent Changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean - the 100 “worst Invasives” and Their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Agostaro, R.D.; Milazzo, M.; Badalamenti, F. The Invasive Seaweed Asparagopsis Taxiformis Erodes the Habitat Structure and Biodiversity of Native Algal Forests in the Mediterranean Sea. Mar. Environ. Res. 2022, 173, 105515. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P. Citizen Science: A Successful Tool for Monitoring Invasive Alien Species (IAS) in Marine Protected Areas. The Case Study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 2018, 8386, 1–7. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P. Invasive Alien Species in Mediterranean Marine Protected Areas: The Egadi Islands (Italy) Case Study. Biodiversity 2021, 22, 13–23. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Issaris, Y.; Poursanidis, D.; Thessalou-Legaki, M. Vulnerability of Marine Habitats to the Invasive Green Alga Caulerpa Racemosa Var. Cylindracea within a Marine Protected Area. Mar. Environ. Res. 2010, 70, 210–218. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Chemello, R.; Mannino, A.M. The Effects of Non-Indigenous Macrophytes on Native Biodiversity: Case Studies from Sicily. J. Mar. Sci. Eng. 2023, 11, 1389. [Google Scholar] [CrossRef]
- Holmer, M.; Marbà, N.; Lamote, M.; Duarte, C.M. Deterioration of Sediment Quality in Seagrass Meadows (Posidonia Oceanica) Invaded by Macroalgae (Caulerpa Sp.). Estuaries and Coasts 2009, 32, 456–466. [Google Scholar] [CrossRef]
- Arias, A.; Giangrande, A.; Gambi, M.C.; Anadón, N. Biology and New Records of the Invasive Species Branchiomma Bairdi (Annelida: Sabellidae) in the Mediterranean Sea. Mediterr. Mar. Sci. 2013, 14, 162–171. [Google Scholar] [CrossRef]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Castelluccio, F.; Pozone, F.; Roussis, V.; Templado, J.; Ghiselin, M.T.; Cimino, G. Factors Promoting Marine Invasions: A Chemoecological Approach. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 4582–4586. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, E.; Cebrian, E.; Alcoverro, T. Mortality of Shoots of Posidonia Oceanica Following Meadow Invasion by the Red Alga Lophocladia Lallemandii. Bot. Mar. 2007, 50, 8–13. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti-Cecchi, L.; Morri, C.; Pasini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Hard Bottoms. In: GAMBI M.C., DAPPIANO M. (Eds), Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study. In Biol. Mar. Medit.; 2004; Vol. 11 (Suppl., pp. 185–215.
- Mancuso, F.P.; Milazzo, M.; Chemello, R. Decreasing in Patch-Size of Cystoseira Forests Reduces the Diversity of Their Associated Molluscan Assemblage in Mediterranean Rocky Reefs. Estuar. Coast. Shelf Sci. 2021, 250, 107163. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer Netherlands: Dordrecht, 1988; Volume 51, ISBN 978-94-015-7360-3. [Google Scholar]
- Blondel, V.D.; Guillaume, J.L.; Lambiotte, R.; Lefebvre, E. Fast Unfolding of Communities in Large Networks. J. Stat. Mech. Theory Exp. 2008, 2008. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, 2008; ISBN 0-12-044620-0. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing 2021.
Figure 1.
Study sites (red dots) along the northwester rocky-shore of Sicily: MPA = MPA of “Capo Gallo-Isola delle Femmine”, MC = Monte Cofano, PP = Punta Priola. The top panel displays information about the sites investigated within the MPA: IdF = Isola, LC = La Cala, PM = Punta Matese, PB = Punta Barcarello, B = Barcarello, CG = Capo Gallo. Within the MPA, red, yellow, and blue areas denote no-take, general, and limited protection zones, respectively.
Figure 1.
Study sites (red dots) along the northwester rocky-shore of Sicily: MPA = MPA of “Capo Gallo-Isola delle Femmine”, MC = Monte Cofano, PP = Punta Priola. The top panel displays information about the sites investigated within the MPA: IdF = Isola, LC = La Cala, PM = Punta Matese, PB = Punta Barcarello, B = Barcarello, CG = Capo Gallo. Within the MPA, red, yellow, and blue areas denote no-take, general, and limited protection zones, respectively.
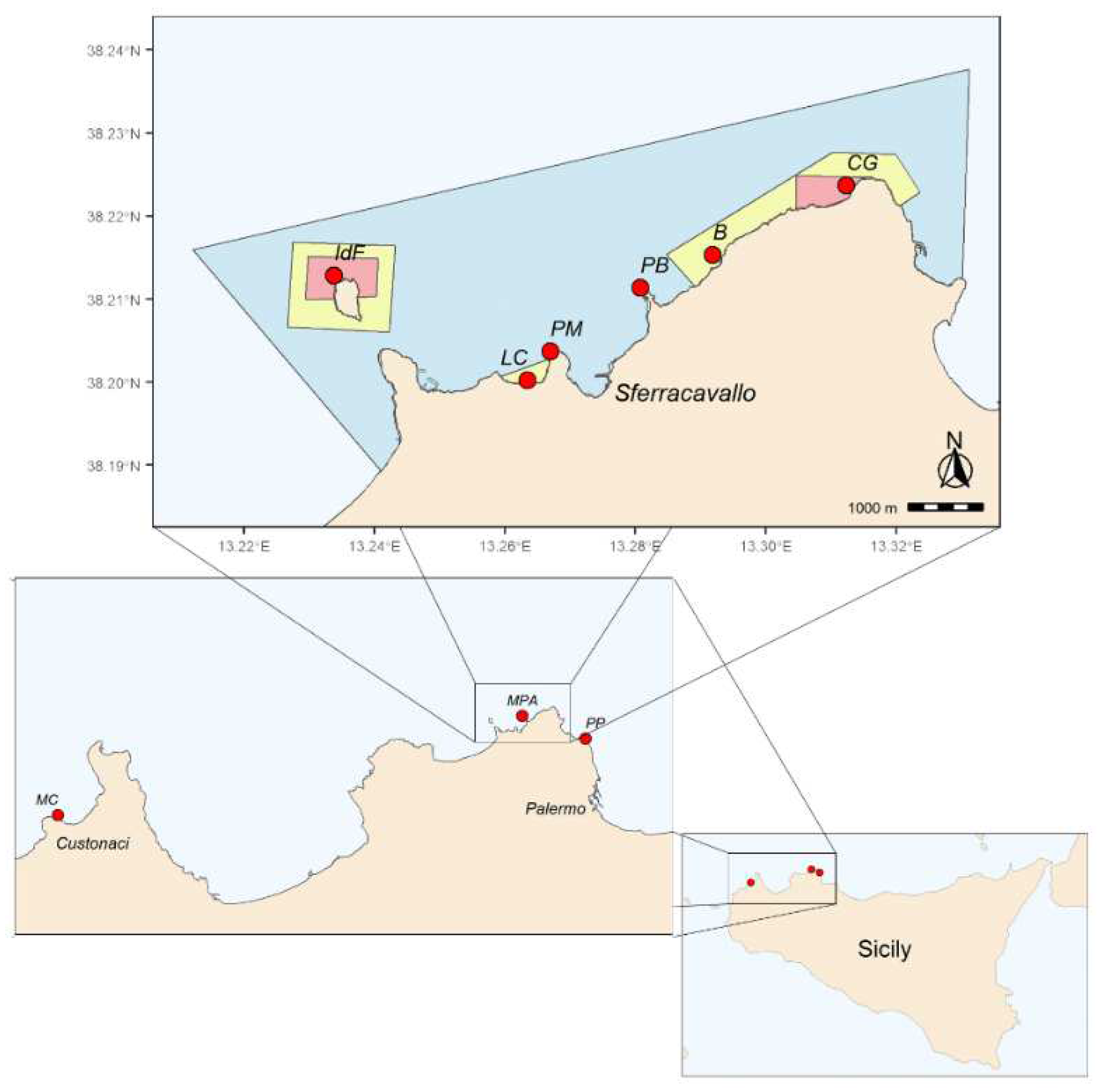
Figure 2.
Abundance (a, average cover percentage), species richness (b), Shannon-Wiener diversity (c), and Pielou’s evenness index (d) of the Cystoseira s.l. assemblage among the different MPA protection levels. Boxplots show extreme and lower whisker (vertical black line), lower and upper quartile (box), and median (horizontal black line). Density plot are shown beside each boxplot. Dots are raw data (n = 24). Signif. codes: * p < 0.01, . p < 0.05, ns p > 0.05.
Figure 2.
Abundance (a, average cover percentage), species richness (b), Shannon-Wiener diversity (c), and Pielou’s evenness index (d) of the Cystoseira s.l. assemblage among the different MPA protection levels. Boxplots show extreme and lower whisker (vertical black line), lower and upper quartile (box), and median (horizontal black line). Density plot are shown beside each boxplot. Dots are raw data (n = 24). Signif. codes: * p < 0.01, . p < 0.05, ns p > 0.05.
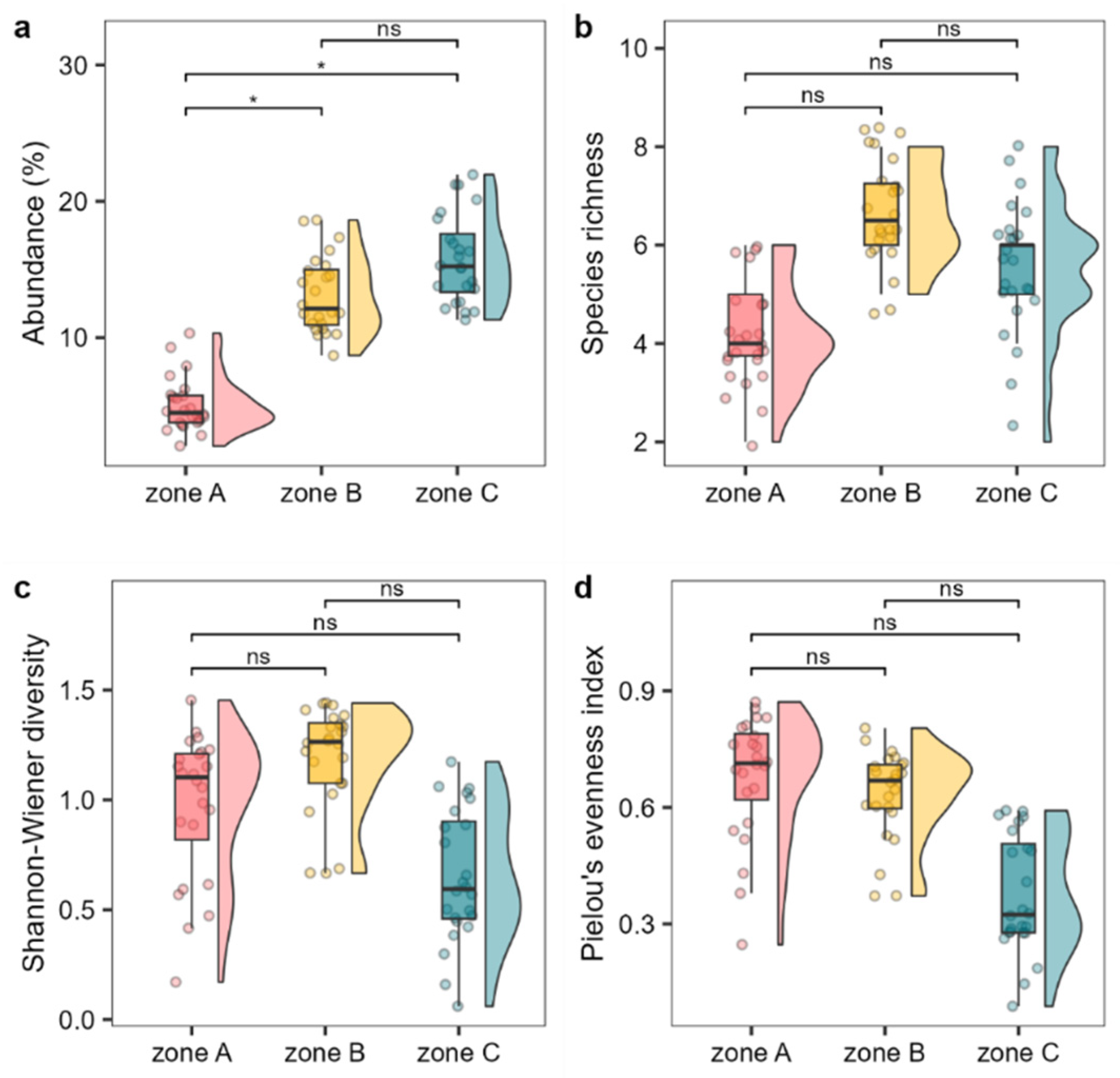
Figure 3.
Structure (a) and composition (b) of the Cystoseira s.l. assemblage founded. Circles show the 95% confidence of interval for each MPA zone (red = zone A, yellow = zone B and blue = zone C). Principal coordinate analysis plot (PCoA) based on Bray–Curtis measure of square-root transformed Cystoseira s.l. percentage cover (structure) or Jaccard measure (composition).
Figure 3.
Structure (a) and composition (b) of the Cystoseira s.l. assemblage founded. Circles show the 95% confidence of interval for each MPA zone (red = zone A, yellow = zone B and blue = zone C). Principal coordinate analysis plot (PCoA) based on Bray–Curtis measure of square-root transformed Cystoseira s.l. percentage cover (structure) or Jaccard measure (composition).
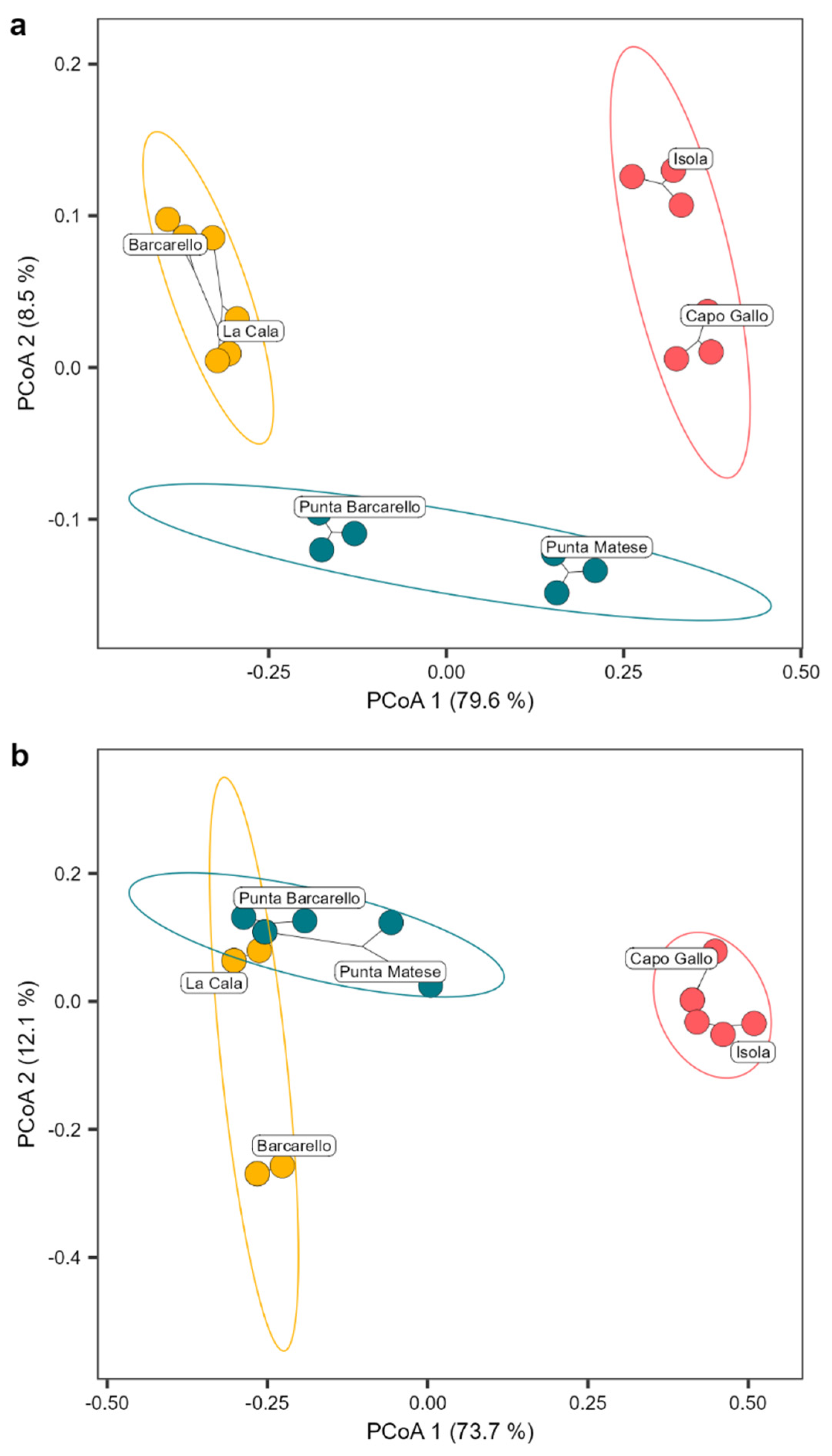
Figure 4.
Abundance (a, average cover percentage), species richness (b), Shannon-Wiener diversity (c), and Pielou’s evenness index (d) of the Cystoseira s.l. assemblage among the different MPA zones and outside control sites. Boxplots show extreme and lower whisker (vertical black line), lower and upper quartile (box), and median (horizontal black line). Density plot are shown beside each boxplot. Dots are raw data (n = 12 - 24). Signif. codes: * p < 0.01, . p < 0.05, ns p > 0.05.
Figure 4.
Abundance (a, average cover percentage), species richness (b), Shannon-Wiener diversity (c), and Pielou’s evenness index (d) of the Cystoseira s.l. assemblage among the different MPA zones and outside control sites. Boxplots show extreme and lower whisker (vertical black line), lower and upper quartile (box), and median (horizontal black line). Density plot are shown beside each boxplot. Dots are raw data (n = 12 - 24). Signif. codes: * p < 0.01, . p < 0.05, ns p > 0.05.
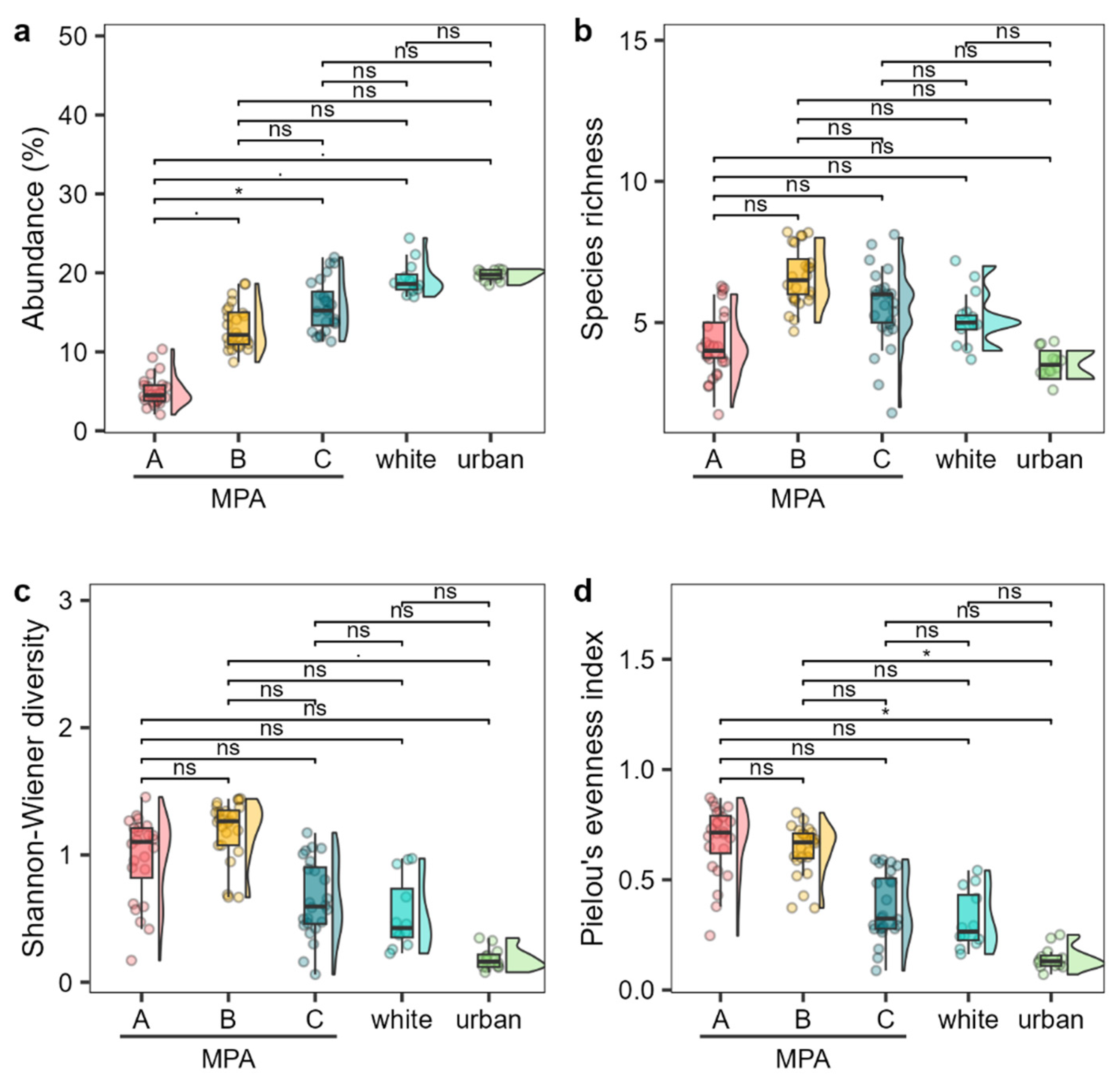
Figure 5.
Structure (a) and composition (b) of MPA and outside control sites Cystoseira s.l. assemblage. Circles show the 95% confidence of interval for each MPA zone (red = zone A, yellow = zone B, blue = zone C, light-blue = white site and green = urban site). Principal coordinate analysis plot (PCoA) based on Bray–Curtis measure of square-root transformed Cystoseira s.l. percentage cover (structure) or Jaccard measure (composition).
Figure 5.
Structure (a) and composition (b) of MPA and outside control sites Cystoseira s.l. assemblage. Circles show the 95% confidence of interval for each MPA zone (red = zone A, yellow = zone B, blue = zone C, light-blue = white site and green = urban site). Principal coordinate analysis plot (PCoA) based on Bray–Curtis measure of square-root transformed Cystoseira s.l. percentage cover (structure) or Jaccard measure (composition).
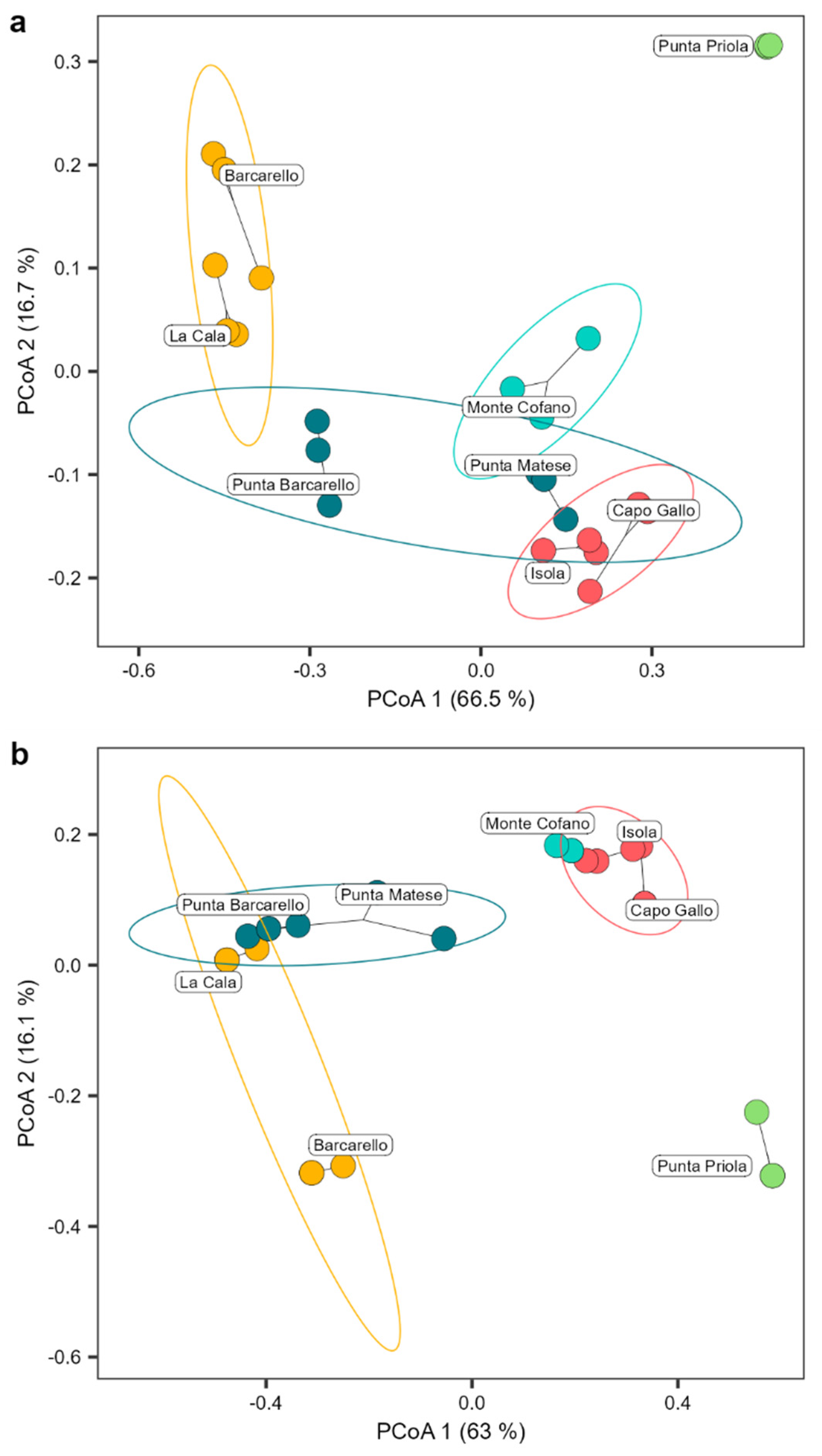
Figure 6.
Invasive species found within the MPA of Capo Gallo – Isola delle Femmine. A = Asparagopsis taxiformis; B = Caulerpa cylindracea; C = Caulerpa taxifolia var. distichophylla; D = Lophocladia lallemandii. Photos by Francesco Paolo Mancuso.
Figure 6.
Invasive species found within the MPA of Capo Gallo – Isola delle Femmine. A = Asparagopsis taxiformis; B = Caulerpa cylindracea; C = Caulerpa taxifolia var. distichophylla; D = Lophocladia lallemandii. Photos by Francesco Paolo Mancuso.

Figure 7.
Examples of benthic habitats with Cystoseira s.l. populations within the MPA of “Capo Gallo – Isola delle Femmine”: A = Isola (zone A); B = Barcarello (zone B); C = La Cala (zone B); D = Punta Barcarello (zone D). Photos by Francesco Paolo Mancuso.
Figure 7.
Examples of benthic habitats with Cystoseira s.l. populations within the MPA of “Capo Gallo – Isola delle Femmine”: A = Isola (zone A); B = Barcarello (zone B); C = La Cala (zone B); D = Punta Barcarello (zone D). Photos by Francesco Paolo Mancuso.

Figure 8.
Examples of benthic habitats at the two control sites. A = Monte Cofano characterized by the presence of well stated Cystoseira s.l. populations; B = Punta Priola, dominated by Dictyopteris polypodioides. Photos by Francesco Paolo Mancuso.
Figure 8.
Examples of benthic habitats at the two control sites. A = Monte Cofano characterized by the presence of well stated Cystoseira s.l. populations; B = Punta Priola, dominated by Dictyopteris polypodioides. Photos by Francesco Paolo Mancuso.


Table 1.
List of the recorded Cystoseira s.l. species. Taxonomy and nomenclature were updated according to the AlgaeBase [32] database.
Table 1.
List of the recorded Cystoseira s.l. species. Taxonomy and nomenclature were updated according to the AlgaeBase [32] database.
| MPA zones | Control sites | |||||
|---|---|---|---|---|---|---|
| A | B | C | white | urban | ||
| Species | ||||||
| Cystoseira compressa | (Esper) Gerloff & Nizamuddin 1975 | + | + | + | + | + |
| Cystoseira humilis var. myriophylloides | (Sauvageau) J.H.Price & D.M.John | + | + | + | ||
| Cystoseira compressa f. rosetta | (Ercegovic) Cormaci, G.Furnari, Giaccone, B.Scammacca & Serio | + | + | |||
| Cystoseira humilis | Schousboe ex Kützing | + | + | + | ||
| Cystoseira foeniculacea f. tenuiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.Rull Lluch | + | + | |||
| Cystoseira foeniculacea | (Linnaeus) Greville | + | + | |||
| Cystoseira foeniculacea f. latiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.R.Lluch 2001 | + | + | |||
| Cystoseira dubia | Valiante | + | + | |||
| Ericaria amentacea | (C.Agardh) Molinari & Guiry | + | + | + | + | |
| Ericaria brachycarpa | (J.Agardh) Molinari & Guiry | + | + | + | + | + |
| Ericaria crinita | (Duby) Molinari & Guiry | + | + | + | ||
| Ericaria brachycarpa var. claudiae | Boudouresque, Perret-Boudouresque & Blanfuné | + | + | + | ||
| Ericaria funkii | (Schiffner ex Gerloff & Nizamuddin) Molinari & Guiry | + | + | |||
| Ericaria mediterranea | (Sauvageau) Molinari & Guiry | + | ||||
| Gongolaria montagnei | (J.Agardh) Kuntze | + | + | + | + | |
| Gongolaria montagnei var. compressa | (Ercegovic) Verlaque, Blanfuné, Boudouresque & Thibaut | + | + | + | + | |
| Gongolaria squarrosa | (De Notaris) Kuntze | + | ||||
| Gongolaria montagnei var. tenuior | (Ercegovic) Molinari & Guiry | + | + | + | ||
| Gongolaria sauvageauana | (Hamel) Molinari & Guiry | + | ||||
Table 2.
Average abundance (N), frequency of occurrence (Fr %) and dominance (D %) of the Cystoseira sl. species identified on each site.
Table 2.
Average abundance (N), frequency of occurrence (Fr %) and dominance (D %) of the Cystoseira sl. species identified on each site.
| MPA zones | Control sites | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | white | urban | |||||||||||||
| Depth range | Species | N | F% | D% | N | F% | D% | N | F% | D% | N | F% | D% | N | F% | D% | |
| 0-5 m | Cystoseira compressa | (Esper) Gerloff & Nizamuddin 1975 | 3.5 | 41.7 | 3.8 | 8.3 | 50.0 | 4.7 | 5.4 | 37.5 | 5.1 | 7.9 | 50.0 | 6.7 | 11.7 | 50.0 | 51.9 |
| Cystoseira compressa f. rosetta | (Ercegovic) Cormaci, G.Furnari, Giaccone, B.Scammacca & Serio | 1.0 | 12.5 | 1.1 | 0.4 | 4.2 | 0.4 | ||||||||||
| Cystoseira humilis | Schousboe ex Kützing | 15.2 | 54.2 | 8.6 | 4.4 | 25.0 | 4.1 | 13.3 | 50.0 | 11.3 | |||||||
| Ericaria amentacea | (C.Agardh) Molinari & Guiry | 50.8 | 100.0 | 54.5 | 82.3 | 100.0 | 46.3 | 58.1 | 100.0 | 54.8 | 60.0 | 100.0 | 50.7 | ||||
| Ericaria brachycarpa | (J.Agardh) Molinari & Guiry | 37.9 | 95.8 | 40.6 | 71.9 | 100.0 | 40.4 | 37.7 | 87.5 | 35.6 | 31.7 | 83.3 | 26.8 | 10.8 | 50.0 | 48.1 | |
| Ericaria mediterranea | (Sauvageau) Molinari & Guiry | 5.4 | 25.0 | 4.6 | |||||||||||||
| 5-10 m | Cystoseira humilis var. myriophylloides | (Sauvageau) J.H.Price & D.M.John | 3.1 | 12.5 | 11.0 | 8.3 | 20.8 | 8.0 | 4.4 | 16.7 | 6.3 | ||||||
| Cystoseira humilis | Schousboe ex Kützing | 9.0 | 37.5 | 8.6 | 6.0 | 29.2 | 8.7 | 3.3 | 25.0 | 7.3 | |||||||
| Cystoseira foeniculacea f. tenuiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.Rull Lluch | 8.1 | 37.5 | 7.8 | 7.5 | 29.2 | 10.8 | ||||||||||
| Cystoseira foeniculacea | (Linnaeus) Greville | 3.3 | 16.7 | 3.2 | 5.0 | 33.3 | 54.5 | ||||||||||
| Ericaria crinita | (Duby) Molinari & Guiry | 19.2 | 70.8 | 67.6 | 34.8 | 79.2 | 33.5 | 23.5 | 58.3 | 34.1 | |||||||
| Gongolaria montagnei | (J.Agardh) Kuntze | 6.0 | 33.3 | 21.3 | 3.1 | 16.7 | 4.5 | 6.7 | 16.7 | 14.7 | 4.2 | 33.3 | 45.5 | ||||
| Gongolaria montagnei var. tenuior | (Ercegovic) Molinari & Guiry | 40.2 | 83.3 | 38.8 | 24.6 | 62.5 | 35.6 | 25.0 | 75.0 | 55.0 | |||||||
| Gongolaria sauvageauana | (Hamel) Molinari & Guiry | 10.4 | 41.7 | 22.9 | |||||||||||||
| 10-15 m | Cystoseira foeniculacea f. tenuiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.Rull Lluch | 11.2 | 50.0 | 12.3 | 5.2 | 25.0 | 14.5 | |||||||||
| Cystoseira humilis | Schousboe ex Kützing | 10.6 | 37.5 | 11.6 | |||||||||||||
| Cystoseira foeniculacea | (Linnaeus) Greville | 5.6 | 25.0 | 6.2 | 2.1 | 8.3 | 26.3 | ||||||||||
| Ericaria crinita | (Duby) Molinari & Guiry | 12.7 | 45.8 | 43.9 | 14.2 | 45.8 | 15.5 | 6.7 | 33.3 | 18.6 | |||||||
| Ericaria brachycarpa var. claudiae | Boudouresque, Perret-Boudouresque & Blanfuné | 1.2 | 8.3 | 4.3 | 6.0 | 16.7 | 6.6 | 4.2 | 16.7 | 52.6 | |||||||
| Ericaria funkii | (Schiffner ex Gerloff & Nizamuddin) Molinari & Guiry | 14.0 | 45.8 | 15.3 | 6.7 | 25.0 | 18.6 | ||||||||||
| Gongolaria montagnei var. compressa | (Ercegovic) Verlaque, Blanfuné, Boudouresque & Thibaut | 5.4 | 25.0 | 18.7 | 4.0 | 12.5 | 4.3 | 1.9 | 12.5 | 5.2 | 0.8 | 8.3 | 3.2 | ||||
| Gongolaria montagnei var. tenuior | (Ercegovic) Molinari & Guiry | 25.8 | 58.3 | 28.2 | 11.2 | 50.0 | 31.4 | 10.8 | 25.0 | 41.9 | |||||||
| 15-20 m | Cystoseira foeniculacea f. latiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.R.Lluch 2001 | 7.9 | 37.5 | 11.9 | 2.7 | 20.8 | 10.2 | |||||||||
| Cystoseira foeniculacea f. tenuiramosa | (Ercegovic) A.Gómez Garreta, M.C.Barceló, M.A.Ribera & J.Rull Lluch | 7.7 | 33.3 | 11.6 | 1.0 | 4.2 | 3.9 | ||||||||||
| Cystoseira dubia | Valiante | 4.6 | 25.0 | 6.9 | 1.7 | 12.5 | 6.2 | ||||||||||
| Cystoseira humilis | Schousboe ex Kützing | 3.5 | 16.7 | 5.3 | |||||||||||||
| Ericaria funkii | (Schiffner ex Gerloff & Nizamuddin) Molinari & Guiry | 14.8 | 37.5 | 22.3 | 4.2 | 16.7 | 15.6 | ||||||||||
| Gongolaria montagnei | (J.Agardh) Kuntze | 15.6 | 41.7 | 41.0 | 10.0 | 41.7 | 37.5 | 8.3 | 33.3 | 52.6 | 5.4 | 33.3 | 100.0 | ||||
| Gongolaria montagnei var. compressa | (Ercegovic) Verlaque, Blanfuné, Boudouresque & Thibaut | 14.8 | 70.8 | 38.8 | 5.0 | 16.7 | 7.5 | 3.5 | 20.8 | 13.3 | 4.2 | 33.3 | 26.3 | ||||
| Gongolaria squarrosa | (De Notaris) Kuntze | 7.7 | 29.2 | 20.2 | |||||||||||||
| Gongolaria montagnei var. tenuior | (Ercegovic) Molinari & Guiry | 22.9 | 54.2 | 34.5 | 3.5 | 12.5 | 13.3 | ||||||||||
| Gongolaria sauvageauana | (Hamel) Molinari & Guiry | 3.3 | 16.7 | 21.1 | |||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



