
Submitted:
13 November 2023
Posted:
13 November 2023
You are already at the latest version
Abstract
In this study, the potential of whey protein hydrolysate (WPH) containing LDIQK peptide and treadmill exercise to prevent cognitive decline was investigated along with the neuroprotective mechanisms. Cognitive dysfunction was induced in mice by 1 mg/kg scopolamine, followed by administration of WPH at 100 and 200 mg/kg and/or treadmill exercise at 15 m/min for 30 min five days per week. Both WPH administration and treadmill exercise significantly improved memory in mice with scopolamine-induced cognitive impairment, which was attributed to several key mechanisms, including a reduction in oxidative stress based on decreased levels of reactive oxygen species and malondialdehyde in the brain tissue and an increase in acetylcholine by increasing choline acyltransferase and decreasing acetylcholine esterase levels. Exercise and WPH also exerted neuroprotective effects by inhibiting the hyperphosphorylation of tau protein, enhancing the expression of brain-derived neurotrophic factor, and inhibiting apoptosis by reducing the Bax/Bcl2 ratio in conjunction with downregulation of the mitogen-activated protein kinase pathway. Moreover, the impact of WPH and treadmill exercise extended to the gut microbiome, suggesting a potential link with cognitive improvement. These findings suggest that both WPH intake and treadmill exercise are effective strategies for mitigating cognitive impairment, providing promising avenues for treating neurodegenerative diseases.
Keywords:
whey protein hydrolysate
; LDIQK
; neuroprotective effect
; treadmill exercise
; scopolamine
1. Introduction
Dementia can be broadly classified into Alzheimer’s disease (AD) and vascular dementia based on its mechanism of occurrence. AD is the most prevalent and rapidly progressive type of dementia, mainly affecting individuals over 60 years of age, accounting for 50–60% of all dementia cases [1]. AD can be further subdivided into subtypes triggered by environmental and physiological factors. External factors, including age, genetic determinants, and environmental factors, can lead to toxic effects, whereas internal factors involve a decline in neurotransmitters due to reduced acetylcholine (ACh) levels, neuroinflammation initiated by cytokines, cytotoxicity due to the aggregation of beta-amyloid (Aβ) protein and hyperphosphorylated tau protein, and neuronal demise induced by oxidative stress-generated free radicals. Accumulation of Aβ, an aberrant protein within nerve cells, in the brain results in synaptic damage and activation of peripheral glial cells, ultimately culminating in brain cell death mediated by various inflammatory, apoptotic, and oxidative stress responses. ACh is a neurotransmitter required for cognitive functions such as learning and memory [2]. Acetylcholinesterase (AChE) inhibitors serve as therapeutic agents that alleviate the symptoms of cognitive decline in dementia by enhancing cholinergic neurotransmitters [3]. Recent studies have investigated the potential inhibitory effects of substances derived from natural products and food materials against ACh-degrading enzymes. Scopolamine, a muscarinic cholinergic receptor antagonist, inhibits cholinergic neuronal function in the central nervous system, leading to oxidative stress and subsequent cognitive impairment [4], offering a useful drug to stimulate cholinergic memory loss in animal models, thereby facilitating studies on the efficacy of various drugs and natural compounds on protecting memory and cognitive functions [5].
Exercise has been shown to have a positive effect on neurodegenerative diseases through the release of brain-derived neurotrophic factor (BDNF), inhibition of neuronal cell death, and promotion of neurogenesis, collectively enhancing cognitive functions related to memory and learning [6,7,8]. Among various exercises, treadmill exercise has been shown to enhance mitochondrial function and promote autophagy, contributing to the breakdown of abnormal proteins and the maintenance of cellular homeostasis, ultimately fostering positive effects on learning and memory [9,10]. In particular, one study indicated that 4 weeks of endurance exercise had a positive impact on adaptation of the skeletal muscles to physical activity, thereby increasing the expression of proteins involved in mitochondrial biosynthesis and mitophagy in mice [11].
Whey protein is a valuable by-product of the cheese-making process accounting for approximately 20% of total milk proteins. Whey protein is quickly absorbed by the body and contains a large number of branched-chain amino acids that are widely distributed in the muscles [12]. Enzymatic hydrolysis is a common method utilized to improve the functional and nutritional properties of proteins [13,14]. The peptides produced by protein hydrolysis have smaller molecular weights and altered secondary structures compared to those of the intact proteins, with potential to enhance physiological functions, including digestive ability, and to mitigate allergic reactions [15]. Whey protein hydrolysate (WPH) has been demonstrated to have a range of beneficial effects, including increasing the body’s metabolic rate, promoting intestinal health, regulating blood sugar levels, and managing blood pressure. A recent study demonstrated that WPH containing a glycine-threonine-tryptophan-tyrosine peptide improved cognitive function in mice [16]. Nevertheless, there is a lack of research on the cognitive function improvement resulting from the combination of peptides derived from WPH and aerobic exercise.
To address this question, in this study, we evaluated the individual and synergistic effects of administration of WPH containing the LDIQK peptide and treadmill exercise on cognitive function in mice with scopolamine-induced cognitive decline and further explored the underlying mechanism.
2. Materials and Methods
2.1. Experimental Animals and Design
Six-week-old male specific pathogen-free C57BL6 mice weighing 20–25 g were purchased from OrientBio (Seongnam, Korea). The animals were housed in an environment with a temperature of 23 ± 3°C, relative humidity of 50 ± 10%, ventilation frequency of 10–15 times/h, 12-h light/dark cycle (08:00 to 20:00), and illuminance of 150–300 Lux. During the 1-week acclimatization period, the animals were allowed to consume solid laboratory-grade food (Cargil Agri Purina, Inc., Seongnam, Korea) and drinking water ad libitum. After the acclimatization period, the mice were divided into seven experimental groups with eight mice per group: the normal (NOR), scopolamine control (CON), scopolamine + exercise (EXR), scopolamine + WPH (WPH_L, WPH_H), and scopolamine + exercise + WPH groups (EWPH_L, EWPH_H). The mice in the WPH groups were orally administered 100 (WPH_L/EWPH_L) or 200 mg/kg (WPH_H/EWPH_H) WPH per day based on body weight. Mice in the exercise groups were subjected to treadmill exercise five times per week for 30 min at a speed of 15 m/min during the four-week experimental period. To induce cognitive impairment, 1 mg/kg scopolamine (Sigma-Aldrich Co., St. Louis, MO, USA) was administered intraperitoneally 30 min before the cognitive tests.
All animal experiments were approved by the Institutional Animal Care and Use Committee of Korea University (KIACUC-2022-0076).
2.2. Y-Maze Test
The Y-maze test is a behavioral test commonly used to assess short-term spatial cognition [17]. This assessment involves the use of a Y-shaped maze constructed of white plastic material. The maze used in this study comprised three arms, each measuring 50 cm in length, 20 cm in height, and 10 cm in width. These arms were folded at an angle of 120° to each other. After designating each branch as A, B, or C, the mouse was placed at the beginning of the maze and allowed to move freely through the maze for 60 s. The number and order of entries into each part of the maze were measured to evaluate the change in behavior. The scoring system for this assessment involved awarding one point when the mouse sequentially entered all three main areas (A, B, and C) in the correct order. However, no points were awarded if entries occurred in a non-sequential manner. The test conductor established the basic conditions for learning and memory assessment criteria and validated the test method in terms of rationality, accuracy, and reproducibility.
2.3. Novel Object Recognition Test
The novel object recognition test was performed in a white polyvinyl plastic box (30 × 30 × 30 cm). The mice were allowed to explore the box freely for 10 min for the first 2 days for acclimation [18]. On the third day of the experiment, the mice were injected with WPH at 100 or 200 mg/kg 1 h before placing them in the box. All experimental groups except the NOR group were administered an intraperitoneal dose of scopolamine (1 mg/kg) dissolved in a 0.9% saline solution 30 min before the start of the experiment, placed in the front center of the box, and then presented with identical objects at equal distances (5 cm) from both diagonal vertices of the box and allowed to explore the objects freely for 5 min. After 24 h of exploration, one of the objects in the box was replaced with a new object of a different shape and the mouse was allowed to explore the object freely for another 5 min. During these 5 min, we measured the time that the mouse exhibited exploratory behaviors such as touching, sniffing, and licking familiar and novel objects. The object preference ratio and discrimination indices were then calculated by recording the number of touches between the familiar object and the novel object and comparing the total number of touches of the objects. A higher discrimination index indicates greater recognition of novel objects.
2.4. Measurement of ACh Content
The ACh content in the brain tissue was measured using the modified method of Vincent and Newsom-Davis [19]. In brief, the harvested brain tissue was homogenized in 1 mL of phosphate-buffered saline (PBS), centrifuged (10,000 ×g, 10 min), and the supernatant was obtained for analysis. Alkaline hydroxylamine reagent (3.5 N sodium hydroxide and 2 M hydroxylamine in HCl) was added to the supernatant and left to react at room temperature for 1 min, followed by the addition of 0.1 N HCl in 0.5 N HCl (pH 1.2) and 0.37 M FeCl3. The absorbance was then measured at a wavelength of 540 nm.
2.5. Measurement of AChE Activity
AChE activity was measured using an AChE activity assay kit (BM-ACH-100; BIOMAX, SEOUL, South Korea) according to the manufacturer instructions. The brain tissue was homogenized in 1 mL of PBS containing 1% Triton X-100 (Sigma Aldrich) and centrifuged (12,000 ×g 10 min) to obtain the supernatant. Subsequently, 50 μL of the supernatant was mixed with the reaction mixture containing assay buffer, enzyme mix, substrate solution, and the probe, and the absorbance was measured at 570 nm with a microplate reader in kinetic mode at 37°C for 30 min in the dark.
2.6. Determination of Reactive Oxygen Species (ROS) and Malondialdehyde (MDA) Contents
To measure ROS levels, 1 mL of 40 mM Tris–HCl buffer was added to 50 mg of brain tissue. The tissues were homogenized and centrifuged to collect the supernatant. Subsequently, 500 μL of 40 mM Tris–HCl buffer and 10 μM of the fluorescent probe 2′,7′-dichlorofluorescein diacetate (DCF-DA) were added to 50 μL of the supernatant and reacted for 30 min at 37°C. Fluorescence was measured at an excitation wavelength of 482 nm and an emission wavelength of 535 nm to quantify the ROS content in the tissue by comparison with an ROS standard curve.
The MDA content in the brain tissue was analyzed using the OxitecTM TBARS Assay kit (BIOMAX Co, Ltd., UK) following the manufacturer’s protocol. Briefly, 1 mL of PBS was added to 100 mg of brain tissue, which was homogenized and centrifuged to collect the supernatant. Subsequently, 200 μL of the indicator solution was added to 200 μL of the supernatant and reacted at 65°C for 45 min. The absorbance was measured at 450 nm and compared to the MDA standard curve to quantify the MDA content in the tissue.
2.7. Western Blotting
Approximately 50 mg of brain tissue was homogenized in 1000 μL of lysis buffer [200 mM Tris (pH 8.0), 150 mM NaCl, 2 mM EDTA, 1 mM NaF, 1% NP40, 1 mM phenylmethanesulfonyl fluoride, 1 mM Na3VO4, and protease inhibitor cocktail] and centrifuged at 12,000 ×g for 5 min at 4°C to recover the supernatant.
The sample was subjected to protein quantification by the bicinchoninic acid method and 30 μg of protein was subjected to electrophoresis on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gel. After transfer to a polyvinylidene fluoride membrane, the blots were blocked with 5% skim milk and bovine serum albumin for 1 h and probed with the following primary antibodies for 16 h at 4°C: alpha-tubulin (Cell Signaling Technology, Inc., Cat. #2144), Bax (Cell Signaling Technology, Inc., Cat. #2772), Bcl-2 (Cell Signaling Technology, Inc., Cat. #2876), PARP (Cell Signaling Technology, Inc., Cat. #9553), BDNF (Cell Signaling Technology, Inc., Cat. #47808), phosphor (p)-tau (Cell Signaling Technology, Inc., Cat. #29957), tau (Cell Signaling Technology, Inc., Cat. #4019), and choline acyltransferase (ChAT; abcam, ab183591) antibodies. After washing three times with Tris-buffered saline with Tween 20, the blot was treated with horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody (Cell Signaling Technology, Inc., Cat. #7074) and reacted for 2 h at room temperature, followed by dispensing of SuperSignal™ Western Blot Enhancer (Thermo Scientific, Cat. #46641) and protein bands were identified using the FluorChem M Fluorescent Western Imaging System (Protein Simple, California, USA). The antibodies used were diluted in 5% skim milk and bovine serum albumin, according to the manufacturer’s instructions.
2.8. Gut Microbiome Analysis
DNA extraction and 16S rRNA gene sequencing of double-stranded DNA were performed from 100 mg of cecum samples using the QIAamp Power Fecal Pro DNA Kit (QIAGEN, Frederick, MD, USA) following the manufacturer’s protocol. The DNA concentration of all samples was adjusted to 5 ng/µL, and the uniformly concentrated DNA was subjected to two-step polymerase chain reaction using the primer set 341F and 806R to amplify the V3–V4 variable region of the 16S rRNA gene. Illumina MiSeq (Illumina) library construction was performed according to the manufacturer’s protocol and sequencing was performed by Macrogen (Seoul, Korea).
2.9. Statistical Analysis
SPSS software (version 12.0; SPSS Inc., Chicago, IL, USA) was used to analyze the experimental data. The data of each experiment are expressed as percentages or mean ± standard error of the mean as appropriate, and all measurements were subjected to one-way analysis of variance followed by the post-hoc Tukey test to evaluate significance. Statistical significance was judged at a threshold of p < 0.05.
3. Results
3.1. Effect of Treadmill Exercise and WPH on Scopolamine-Induced Cognitive Decline
Y-maze and novel object recognition tests were used to measure memory improvement conferred by WPH administration and treadmill exercise in mice with scopolamine-induced cognitive impairment (Figure 1). In the Y-maze, the spontaneous alternation in the NOR group (no treatment) was 79.99%, which was higher than that of the CON group (group administered scopolamine and saline) of 42.25% (Figure 1A). Spontaneous alternation increased to 54.1% and 62.7% in a concentration-dependent manner following the administration of 100 and 200 mg/kg WPH, respectively. The frequency of spontaneous alternations in the EXR (treadmill exercise), EWPH_L (treadmill exercise with low-concentration WPH administration), and EWPH_H (treadmill exercise with high-concentration WPH administration) groups was 61.4%, 66.1%, and 64.7%, respectively, which were all significantly higher than those in the CON group (p < 0.001, Figure 1A). The total number of entries into each arm did not show significant differences among any of the experimental groups (Figure 1B), even though spontaneous alternations tended to increase when WPH was combined with exercise.
In the novel object recognition test, which evaluates the ability of mice to explore novelty, the discrimination index (Figure 1C) was calculated by analyzing the time taken to explore new and existing objects. The discrimination index for novel objects in the CON group was significantly reduced compared to that of the NOR group (p < 0.05), indicating that scopolamine induced a cognitive impairment associated with new object recognition. The discrimination index for novel objects significantly increased in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups compared with that of the CON group (Figure 1C).
By contrast, the discrimination index for familiar objects significantly increased in the CON group compared to that of the NOR group, but decreased to the level of the NOR group in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups compared to the CON group. Unlike the NOR, EXR, and WPH groups, the CON group showed similar indices for familiar and novel objects (Figure 1C). This result indicated that both exercise and WPH administration improved the ability of mice to explore novel objects (p < 0.001, Figure 1C). No synergistic effect was observed with the combination of treadmill exercise and WPH.
3.2. Effects of Treadmill Exercise and WPH on ROS and MDA Levels in Scopolamine-Administered Mice
The ROS content in the CON group was significantly increased by 185.6% compared to that in the NOR group, indicating that scopolamine induced an increase in ROS production. The administration of 100 and 200 mg/kg WPH significantly reduced ROS levels by 132.1% and 129.4%, respectively, compared to those of the CON group (Figure 2A). In the EXR group, which only performed treadmill exercise, and the EWPH_L and EWPH_H groups, which combined WPH administration and exercise, ROS levels were significantly reduced by 124.0%, 129.6%, and 109.8%, respectively, compared to those in the CON group (p < 0.001, Figure 2B). These results showed that treadmill exercise and WPH administration inhibited the scopolamine-induced increase in ROS levels responsible for oxidative stress. The combination of exercise and WPH administration had a synergistic effect on ROS removal from the mouse brain.
The MDA content of the CON group was 52.6 μM, which was significantly higher than that of the NOR group at 12.1 μM (p < 0.001). The MDA content of the EXR and WPH_H groups was significantly reduced to 27.1 and 34.6, μM, respectively, compared to that of the CON group (p < 0.001, Figure 2B). The MDA content of the groups that combined exercise and WPH administration (EWPH_L and EWPH_H) was 29.9 and 24.8 μM, which was significantly decreased compared to that of the CON group (p < 0.001, Figure 2B). Therefore, the combined use of treadmill exercise and high-dose WPH had a synergistic effect on MDA reduction. This result showed that exercise and WPH administration effectively reduced the scopolamine-induced formation of MDA in the mouse brain.
3.3. Effects of Treadmill Exercise and WPH on ChAT protein abundance, ACh Content, and AChE Activity in Scopolamine-Administered Mice
In the mouse brain tissue, the ACh concentration of the CON group was significantly reduced by 0.6 times compared to that of the NOR group (Figure 3A). In addition, the activity of AChE, an ACh-degrading enzyme, was significantly increased by 2.7 times, and the protein level of ChAT was significantly decreased by 3.3 times in the NOR group compared with those of the CON group (Figure 3). These results indicated that the administration of scopolamine reduced ACh levels by increasing AChE activity and decreasing ChAT protein levels, consequently leading to cognitive dysfunction. Treadmill exercise and WPH administration restored ACh levels reduced by scopolamine administration (Figure 3A). The ACh concentration significantly increased by 1.3 times in the EXR, WPH_H, and EWPH_H groups compared to that in the CON group (p < 0.01, Figure 3A). AChE activity significantly decreased by 0.8-fold, 0.8-fold, 0.5-fold, 0.5-fold, and 0.4-fold in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups, respectively (p < 0.001; Figure 3B). The expression level of ChAT protein was significantly increased by 309.12% in the EWPH_H group compared to that in the CON group (p < 0.05; Figure 3C). The combination of treadmill exercise and high-dose WPH administration synergistically reduced the ACh concentration and increased ChAT protein levels. These results showed that both exercise and WPH administration effectively restored ACh levels, which are essential for neurotransmission and cognitive function.
3.4. Effects of Treadmill Exercise and WPH on Neuronal Apoptosis and Tau Phosphorylation in Scopolamine-Administered Mice
The Bax/Bcl-2 ratio was significantly increased in the CON group following scopolamine treatment compared to that in the NOR group (Figure 4A,B). However, the Bax/Bcl-2 ratio significantly decreased in the EXR group subjected to treadmill exercise; the WPH_L and WPH_H groups treated with WPH at low and high dose, respectively; and the EWPH_L and EWPH_H groups receiving both exercise and WPH at low and high doses, respectively (31.6%, 32.7%, 31.3%, 41.6%, and 27.6%, respectively; p < 0.001, Figure 4B). Therefore, exercise and WPH administration effectively inhibited scopolamine-induced apoptosis. The combination of treadmill exercise and WPH administration synergistically reduced the Bax/Bcl-2 ratio. These results demonstrated that both exercise and WPH administration effectively inhibited apoptotic damage to neurons by reducing the Bax/Bcl-2 ratio. In addition, scopolamine significantly decreased the expression level of BDNF and impaired the expression of neuroplasticity factors; however, WPH treatment (100 and 200 mg/kg) and co-treatment with WPH and treadmill exercise significantly increased BDNF expression (150.8%, 151.1%, 153.3%, and 168.3%, respectively; p < 0.05, Figure 4A and C). The combination of treadmill exercise and high-dose WPH administration synergistically increased BDNF protein levels.
Furthermore, WPH and treadmill exercise modulated the phosphorylation of tau proteins associated with neuronal function. Scopolamine administration significantly increased the p-tau/tau protein ratio, a marker of neurodegeneration, in the brain by 3.4-fold compared to that in the NOR group. However, the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups showed concentration-dependent reductions in the p-tau/tau ratio of 33.4%, 32.5%, 24.6%, 41.9%, and 28.9%, respectively (Figure 4A,D). These results indicate that treadmill exercise and WPH administration regulate neurogenesis and the neurodegenerative factors p-tau/tau and BDNF.
3.5. Effects of Treadmill Exercise and WPH on Mitogen-Activated Protein Kinase (MAPK) Signaling in Scopolamine-Administered Mice
The effects of WPH and treadmill exercise on the MAPK signaling pathway were evaluated in the brain tissue. Scopolamine administration significantly increased the protein expression levels of p-c-Jun NH2-terminal kinase (JNK), p-extracellular signal-regulated kinase 1/2 (ERK1/2), and p-p38, key components of the MAPK signaling pathway, compared to those in the NOR group by 2.3-, 2.8-, and 1.9-fold, respectively (p < 0.01, Figure 5A–E). However, in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups, p-JNK expression was significantly reduced compared to that in the CON group by 53.3%, 50.9%, 42.4%, 41.1%, and 39.8%, respectively; p-ERK expression was reduced by 50.7%, 71.8%, 50.3%, 57.2%, and 51.1%, respectively; and p-P38 expression was reduced by 70.2%, 78.2%, 75.3%, 59.5%, and 47.3%, respectively (Figure 5B–E). These results indicated that exercise and WPH administration effectively inhibited the MAPK signaling pathway activated by scopolamine. Additionally, inhibition of the MAPK signaling pathway was most pronounced when treadmill exercise and WPH administration were concomitantly performed.
3.6. Effects of Treadmill Exercise and WPH on Intestinal Microbiota
Cecal microbiota were measured to evaluate changes in the intestinal microbiota following WPH administration and treadmill exercise. The Shannon and Chao indices, which represent the diversity and abundance of the microbiome, respectively, tended to decrease in the CON group compared to those of the NOR group (Supplementary Figure S1). Treadmill exercise, WPH (100 and 200 mg/kg), and combined treatment with exercise and WPH (100 and 200 mg/kg) increased the Shannon index by 112%, 112%, 114%, 108%, and 110%, respectively, and the Chao index by 123%, 114%, 120%, 113%, and 116%, respectively, compared with that of the CON group (Supplementary Figure S1A,B). Principal coordinates analysis, which represents beta diversity, confirmed that the microbiome community changed between the NOR group and CON group administered scopolamine. Treadmill exercise, WPH administration, and their combination resulted in a different microbiome community than that of the CON group (Supplementary Figure S1C).
Changes in the cecal microbiota at the phylum level showed that treadmill exercise (EXR), WPH administration (WPH_L and WPH_H), and combined exercise and WPH administration (EWPH_L and EWPH_H) significantly reduced the relative abundance of Verrucomicrobiota that was increased in the CON group following scopolamine administration (Figure 6A). The exercise and high-dose WPH group (EWPH_H) had a significantly higher relative abundance of Bacillota than that of the CON group (Figure 6B). The Bacillota/Bacteroidota ratio increased after scopolamine administration. At the phylum level, Bacillota and Bacteroidota were the main taxa with high relative abundances, and there was no significant difference in their abundance among the groups (Figure 6B-D). The relative abundance of Actinomycetota in the EWPH_H group was significantly higher than that in the CON group (p < 0.01; Figure 6E), but did not differ significantly from that of the other groups. The combination of treadmill exercise and high-dose administration of WPH showed a synergistic effect in increasing the relative abundances of Bacillota (Figure 6B) and Actinomycetota (Figure 6E) and in decreasing the relative abundance of Verrucomicrobiota (Figure 6F).
At the class and order levels, Verrucomicrobiae and Verrucomicrobiales had significantly decreased relative abundances in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups compared to those of the CON group (p < 0.05, Supplementary Figure S2). The relative abundances of Bacteroidia and Bacteroidales were significantly higher in the NOR group than in the scopolamine-treated CON group. Furthermore, the relative abundance of Actinomycetes in the EWPH_H group was significantly higher than that in the CON group.
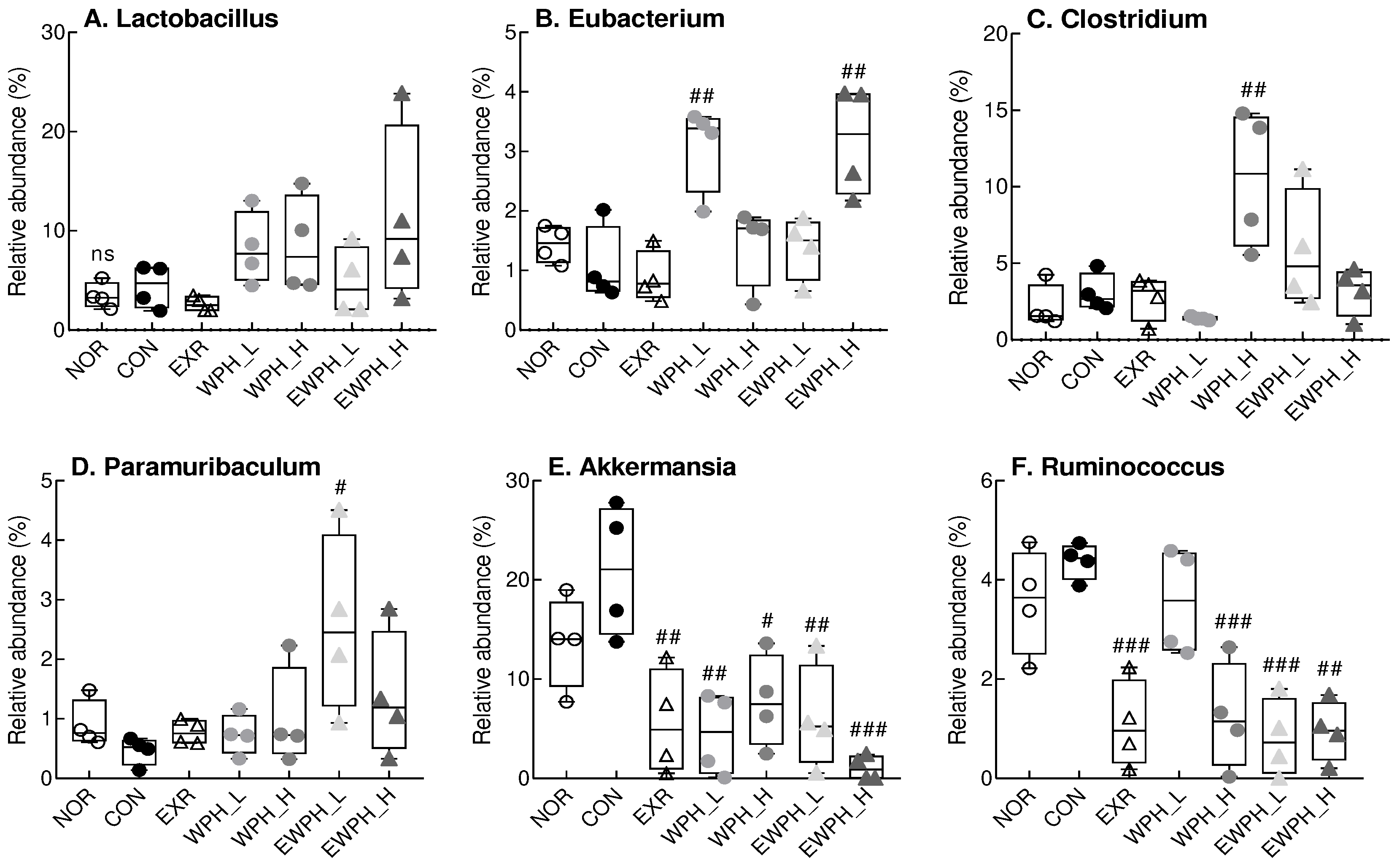
Changes in the cecal microbiota were also detected at the family level. The relative abundance of Akkermansiaceae was significantly lower in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups than in the CON group (p < 0.05, Supplementary Figure S2). The relative abundances of Rikenellaceae and Clostridiaceae were significantly higher in the WPH_H group than in the CON group (p < 0.05; Supplementary Figure S2). In addition, the relative abundance of Eubacteriaceae was significantly higher in the WPH_L and EWPH_H groups than in the CON group, whereas the relative abundance of Lachnospiraceae was significantly higher in the EXR group than in the CON group (p < 0.05, Supplementary Figure S2).
At the genus level, WPH administration alone and in combination with treadmill exercise tended to increase the relative abundance of Lactobacillus compared to that of the CON group, but the difference was not statistically significant (Figure 7). The relative abundance of Eubacterium in the WPH_L and EWPH_H groups was significantly higher than that in the CON group (p < 0.01; Figure 7). Furthermore, the relative abundance of Clostridium was significantly increased in the WPH_H group compared to that of the CON group (p < 0.01, Figure 7). The relative abundance of Paramuribaculum, which decreased in the CON group, increased significantly in the EWPH_L group (p < 0.05; Figure 7). The relative abundances of Akkermansia and Ruminococcus were significantly lower in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups than in the CON group. These results indicated that treadmill exercise and WPH protected against the scopolamine-induced alterations of the gut microbiota.
4. Discussion
In the present study, we investigated the effects of treadmill exercise and WPH intake on scopolamine-induced cognitive dysfunction in mice. Scopolamine, a muscarinic cholinergic receptor antagonist, induces cognitive impairment through cholinergic dysfunction and oxidative stress in the brain [20]. Therefore, animal models treated with scopolamine serve as reliable representations of cognitive impairment, and play a crucial role in investigations aimed at assessing the effectiveness of substances for the prevention and treatment of AD and uncovering their underlying mechanisms [21].
We found that WPH administration and treadmill exercise improved scopolamine-induced memory impairment in behavioral tests, including the Y-maze and novel object recognition tests. The Y-maze test is a straightforward method for assessing short-term spatial memory in experimental animals, which involves placing animals in the Y-maze and quantifying the number of sequential entries they make [22]. We found that scopolamine treatment significantly decreased spontaneous alternation behavior compared to that of the untreated NOR group. In addition, the EXR group, which received treadmill exercise; the WPH_L and WPH_H groups, which were supplemented with WPH; and the EWPH_L and EWPH_H groups, which received both exercise and WPH, showed recovery of alternation behavior at levels similar to those in the NOR group, confirming that memory was improved. The novel object recognition test has been widely used to study behavior and brain function in rats and mice, as well as in memory research. There was no significant difference in the time required to explore the novel and familiar objects after scopolamine administration [23]. However, the time spent exploring novel objects was significantly longer than the time spent exploring familiar objects in the EXR, WPH_L, WPH_H, EWPH_L, and EWPH_H groups. This suggests that exercise and WPH administration mitigate scopolamine-induced cognitive impairment. In our previous work (revision in progress), WPH has demonstrated its ability to protect neuronal cells from oxidative stress conditions, a finding that aligns with the current study’s data showcasing improved cognitive function. LDIQK, a WPH-derived pentapeptide (leucine-aspartate-isoleucine-glutamine-lysine), has been isolated and confirmed as an active principle for neuroprotective effect (revision in progress). The present study substantiates the positive impact of WPH containing LDIQK on cognitive function in an animal model and introduces an additional intervention: exercise. This dual approach is believed to create a synergistic effect, further enhancing ability. Exercise, a well-established contributor to overall health, is known to deliver a fresh supply of oxygen and nutrients to the brain by boosting blood flow. This physiological boost acts as a power-up for cognitive function, supporting optimal brain performance [24]. Furthermore, exercise has been reported to promote the release of neurotransmitters such as dopamine and serotonin, influencing mood and cognitive process [25]. By comparing the neuroprotective effects of WPH with the cognitive benefits of exercise, our study suggests a synergistic relationship that could lead to a more significant improvement in cognitive ability. This dual intervention approach provides a comprehensive strategy of enhancing brain health and function.
The cholinergic nervous system plays a major role in cognition and memory, and scopolamine has been shown to increase AChE levels and decrease ACh and ChAT levels [26]. ACh is a major neurotransmitter and regulator in the nervous system, which is considered to play an important role in cognitive functions such as learning and memory at the neuromuscular junction and in the parasympathetic nervous system [27]. Increased activity of enzymes such as AChE and butyrylcholinesterase in the brain leads to cholinergic dysfunction by breaking down the neurotransmitter ACh into choline and acetyl-coenzyme A, which contributes to impaired memory and cognitive function [28]. In this study, we found that treadmill exercise and WPH administration significantly decreased the activity of AChE and increased the protein expression of ChAT, leading to an increase in the ACh concentration. These effects of exercise and WPH administration on ACh, AChE, and ChAT correlated with the results of the behavioral tests (Y-maze and novel object recognition), showing an increase in spontaneous alternation and the novel object discrimination index.
Along with its involvement in cognitive function, ACh is closely associated with oxidative stress in the brain. Given the abundance of unsaturated fatty acids in the brain tissue and their susceptibility to oxidative stress, excessive production of free radicals leads to the accumulation of lipid peroxides, protein denaturation, DNA oxidation, and subsequent cell damage, thereby hindering physiological activities [28]. In particular, oxidative stress leads to the peroxidation of unsaturated fatty acids surrounding nerve cells and an increase in the activity of AChE, which promotes the degradation of ACh, resulting in impaired neurotransmission and, ultimately, a decrease in memory and learning ability. The findings of this study indicated that both exercise and WPH administration significantly reduced scopolamine-induced ROS and MDA levels. These results suggest that treadmill exercise and WPH improve cognitive function by inhibiting ACh degradation through a reduction of oxidative stress.
The Bax/Bcl-2 ratio, which is increased by oxidative stress, is also an important factor in the regulation of apoptosis [29]. Bcl-2, Bcl-xL, and related factors inhibit apoptosis, whereas Bax, Blk, and Bad promote apoptosis. Bax translocates to the mitochondria, where it becomes an activated monodimer and promotes apoptosis; this Bax-induced cell death pathway is inhibited by its heterodimerization with Bcl-2. Previous studies linking exercise to synaptic plasticity and cell proliferation have shown that exercise increases the expression of Bcl-2, an anti-apoptotic marker, and proteins involved in synaptic efficacy and learning in key brain regions [30]. By contrast, exercise inhibits the expression of Bax, a member of the caspase family acting downstream of the proapoptotic pathway and a major promoter of cell death, thereby alleviating cognitive dysfunction. The results of this study demonstrated that treadmill exercise and WPH administration (100 and 200 mg/kg) significantly reduced the Bax/Bcl-2 ratio. These results implied that treadmill exercise and WPH inhibited apoptosis to exert positive effects on cognitive function.
One of the most prominent pathological features of AD is the accumulation of neurofibrillary tangles due to hyperphosphorylated tau protein in the brain, which is toxic to neurons [31]. Additionally, the deposition of tau protein leads to oxidative stress, resulting in extensive neuronal destruction. The destruction of nerve cells in turn causes changes in brain structure and function, leading to cognitive decline [32,33,34]. In particular, pronounced brain atrophy has been observed in the brains of patients with AD, and the administration of scopolamine has been reported to cause a decrease in brain weight along with a decline in overall cognitive function [35]. A recent study showed that whey protein administration reduced the high hyperphosphorylation levels of tau protein in aged rats [35], and another study demonstrated that treadmill exercise inhibited tau protein hyperphosphorylation in tau transgenic mouse models [36]. The results of the present study showed that four weeks of moderate-intensity treadmill exercise and WPH administration at concentrations of 100 and 200 mg/kg significantly inhibited tau protein hyperphosphorylation. The inhibition of p-tau by exercise and WPH administration may contribute to a reduction in neuronal apoptosis.
BDNF is a neurotrophic factor that regulates neuronal growth, increases the activity of acetylcholine synthase in the central nervous system, enhances synaptic plasticity, and is directly involved in memory storage and utilization [38,39,40]. Notably, transgenic mice with reduced BDNF expression showed impaired synaptic function, long-term memory, and learning, and BDNF expression in the brain was found to be reduced in patients with AD [41]. Consistently, in the present study, we found that BDNF protein expression was significantly reduced in the CON group following scopolamine administration. However, when WPH was administered in combination with treadmill exercise, BDNF expression significantly increased. This increase in BDNF levels after exercise and WPH administration may also be associated with a reduction in apoptosis and p-tau.
Under oxidative stress, MAPKs are signal transducers that regulate cell death [42]. Phosphorylation of MAPK proteins by scopolamine leads to an elevated Bax/Bcl-2 ratio [42]. Activation of the MAPK pathway (including ERK1/2, JNK, and p38 MAPK) is one of the markers observed in patients with AD [43]. Moderate-intensity treadmill exercise and WPH administration significantly reduced MAPKs activation in scopolamine-induced mice. These results suggest that whey protein supplementation and treadmill exercise are effective in improving cognitive function by inhibiting the scopolamine-induced activation of MAPKs.
Food ingredients can enhance cognitive function by influencing the gut bacterial diversity. The gut microbiome, constituting 95% of the human microbiome, plays a vital role in the gut–brain axis [44]. This axis links the gut microbiome to the neurological, immune, endocrine, and metabolic systems of the host, thereby playing a role in various neurological and cognitive disorders [45]. Actinomycetes are significant contributors to these pathologies by inhibiting the activity of AChE, which breaks down ACh [46]. We found that high doses of WPH and treadmill exercise increased the relative abundance of Actinomycetes and reduced AChE activity. WPH administration and treadmill exercise also decreased and increased the relative abundance of Verrucomicrobiota and Lachnospiraceae, respectively. The relative abundance of Lachnospiraceae has been reported to be higher in healthy populations than in patients with AD [47].
Treadmill exercise, WPH administration, and their combined treatment decreased the relative abundances of Ruminococcus and Akkermansia at the genus level. A decrease in Ruminococcus abundance was also observed in the normal group during a clinical trial investigating the interaction between cognitive function and the gut flora, whereas an increase in Ruminococcus gnavus relative abundance was reported in a scopolamine-treated model of cognitive dysfunction [48]. These results suggest that cognitive dysfunction may be ameliorated through interactions with other microbes.
Changes in the relative abundance of Akkermansia, a bacterial genus belonging to the phylum Verrucomicrobiota, were similar to those of Verrucomicrobiota detected at the phylum level. Akkermansia was previously shown to ameliorate impaired glucose, fat metabolism, and intestinal epithelial cell damage in AD models [49]. However, an excessive increase in Akkermansia muciniphila did not effectively enhance memory in cognitively impaired mice. The relative abundance of Clostridium tended to increase in all groups in this study, except for the WPH_L group, and the relative abundance increased with higher concentrations of WPH administered. An increase in Clostridium was associated with enhanced synthesis of 3-indolepropionic acid, which scavenges free radicals, reduces neuronal cell death, and alters blood–brain barrier permeability to improve cognitive function [50]. These results suggest that treadmill exercise and WPH improve cognitive function by modulating the gut microbiota associated with cognitive function.
5. Conclusion
This study demonstrates that WPH intake and treadmill exercise prevent scopolamine-induced cognitive decline. WPH and treadmill exercise improved cognitive function by reducing ROS and MDA levels in the brain tissue and by increasing the concentration of ACh. WPH and exercise also protected neurons by inhibiting the hyperphosphorylation of tau protein, increasing the expression of BDNF, and inhibiting apoptosis by downregulating the MAPK pathway. Furthermore, we found that WPH and treadmill exercise improved cognitive function by altering the composition of the gut microbiome, with an increase in the Chao and Shannon indices, suggesting that treadmill exercise and WPH are effective in ameliorating cognitive impairment, providing a potential strategy for addressing neurodegenerative diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Effects of WPH and treadmill exercise on gut microbiome diversity; Figure S2: Effect of WPH and treadmill exercise on gut microbiome composition at the class, order, and family levels.
Author Contributions
Conceptualization, H.J.S. and H.-S.C.; methodology, Y.B.C.; software, Y.B.C. and E.-J.J.; validation, Y.B.C. and E.-J.J.; formal analysis, Y.B.C and E.-J.J.; investigation, Y.B.C.; resources, H.J.S.; data curation, Y.B.C. and E.-J.J.; writing—original draft preparation, Y.B.C., H.J.S., and H.-S.C.; writing—review and editing, H.J.S. and H.-S.C.; visualization, Y.B.C.; supervision, H.J.S. and H.-S.C.; project administration, H.J.S. and H.-S.C.; funding acquisition, H.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through the High Value-added Food Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) (No. 322011-4).
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: a review of progress. Journal of Neurology, Neurosurgery & Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- H Ferreira-Vieira, T.; M Guimaraes, I.; R Silva, F.; M Ribeiro, F. Alzheimer's disease: targeting the cholinergic system. Current neuropharmacology 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Tinker, J.P.; Menzaghi, F.; Lloyd, G.K. The subtype-selective nicotinic acetylcholine receptor agonist SIB-1553A improves both attention and memory components of a spatial working memory task in chronic low dose 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-treated monkeys. Journal of Pharmacology and Experimental Therapeutics 2003, 306, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; He, Z.; Peng, Y.; Jin, W.; Wang, Z.; Mu, L.; Chang, M.; Wang, R. Phoenixin-14 enhances memory and mitigates memory impairment induced by Aβ1-42 and scopolamine in mice. Brain research 2015, 1629, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, M.V.; Frozza, R.L.; de Freitas, G.B.; Zhang, H.; Kincheski, G.C.; Ribeiro, F.C.; Gonçalves, R.A.; Clarke, J.R.; Beckman, D.; Staniszewski, A. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nature medicine 2019, 25, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361, eaan8821. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Magnus, T. Ageing and neuronal vulnerability. Nature reviews neuroscience 2006, 7, 278–294. [Google Scholar] [CrossRef]
- Vainshtein, A.; Tryon, L.D.; Pauly, M.; Hood, D.A. Role of PGC-1α during acute exercise-induced autophagy and mitophagy in skeletal muscle. American Journal of Physiology-Cell Physiology 2015, 308, C710–C719. [Google Scholar] [CrossRef]
- Marques-Aleixo, I.; Santos-Alves, E.; Balça, M.; Rizo-Roca, D.; Moreira, P.; Oliveira, P.; Magalhães, J.; Ascensão, A. Physical exercise improves brain cortex and cerebellum mitochondrial bioenergetics and alters apoptotic, dynamic and auto (mito) phagy markers. Neuroscience 2015, 301, 480–495. [Google Scholar] [CrossRef]
- Lira, V.A.; Okutsu, M.; Zhang, M.; Greene, N.P.; Laker, R.C.; Breen, D.S.; Hoehn, K.L.; Yan, Z. Autophagy is required for exercise training-induced skeletal muscle adaptation and improvement of physical performance. The FASEB Journal 2013, 27, 4184. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, L.S.; Kimball, S.R. Amino acid regulation of gene expression. The Journal of nutrition 2001, 131, 2460S–2466S. [Google Scholar] [CrossRef] [PubMed]
- Doucet, D.; Gauthier, S.; Foegeding, E. Rheological characterization of a gel formed during extensive enzymatic hydrolysis. Journal of Food Science 2001, 66, 711–715. [Google Scholar] [CrossRef]
- Ortiz, S.E.M.; Wagner, J.R. Hydrolysates of native and modified soy protein isolates: structural characteristics, solubility and foaming properties. Food research international 2002, 35, 511–518. [Google Scholar] [CrossRef]
- Yang, H.; Lee, S. Emulsifying properties of whey protein hydrolysates. Korean J Food Sci 2003, 23, 63–69. [Google Scholar]
- Ding, N.; Meng, H.; Wu, C.; Yokoyama, W.; Hong, H.; Luo, Y.; Tan, Y. Whey Protein Hydrolysate Renovates Age-Related and Scopolamine-Induced Cognitive Impairment. Nutrients 2023, 15, 1228. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.L.; Conrad, C.D. Short CommunicationChronic stress leaves novelty-seeking behavior intact while impairing spatial recognition memory in the Y-maze. Stress 2005, 8, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The novel object recognition memory: neurobiology, test procedure, and its modifications. Cognitive processing 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Newsom-Davis, J. Acetylcholine receptor antibody as a diagnostic test for myasthenia gravis: results in 153 validated cases and 2967 diagnostic assays. Journal of Neurology, Neurosurgery & Psychiatry 1985, 48, 1246–1252. [Google Scholar] [CrossRef]
- Rahnama, S.; Rabiei, Z.; Alibabaei, Z.; Mokhtari, S.; Rafieian-Kopaei, M.; Deris, F. Anti-amnesic activity of Citrus aurantium flowers extract against scopolamine-induced memory impairments in rats. Neurological Sciences 2015, 36, 553–560. [Google Scholar] [CrossRef]
- Rabiei, Z.; Mokhtari, S.; Asgharzade, S.; Gholami, M.; Rahnama, S.; Rafieian-kopaei, M. Inhibitory effect of Thymus vulgaris extract on memory impairment induced by scopolamine in rat. Asian Pacific Journal of Tropical Biomedicine 2015, 5, 845–851. [Google Scholar] [CrossRef]
- Todirascu-Ciornea, E.; El-Nashar, H.A.; Mostafa, N.M.; Eldahshan, O.A.; Boiangiu, R.S.; Dumitru, G.; Hritcu, L.; Singab, A.N.B. Schinus terebinthifolius essential oil attenuates scopolamine-induced memory deficits via cholinergic modulation and antioxidant properties in a zebrafish model. Evidence-Based Complementary and Alternative Medicine 2019, 2019. [Google Scholar] [CrossRef]
- Pahaye, D.B.; Bum, E.N.; Taïwé, G.S.; Ngoupaye, G.T.; Sidiki, N.; Moto, F.C.O.; Kouemou, N.; Njapdounke, S.J.K.; Nkantchoua, G.; Kandeda, A. Neuroprotective and antiamnesic effects of Mitragyna inermis willd (Rubiaceae) on scopolamine-induced memory impairment in mice. Behavioural Neurology 2017, 2017. [Google Scholar] [CrossRef]
- Mulser, L.; Moreau, D. Effect of acute cardiovascular exercise on cerebral blood flow: A systematic review. Brain Research 2023, 1809, 148355. [Google Scholar] [CrossRef]
- Basso, J.C.; Suzuki, W.A. The Effects of Acute Exercise on Mood, Cognition, Neurophysiology, and Neurochemical Pathways: A Review. Brain Plasticity, 2017; 2, 127–152. [Google Scholar] [CrossRef]
- Ago, Y.; Koda, K.; Takuma, K.; Matsuda, T. Pharmacological aspects of the acetylcholinesterase inhibitor galantamine. Journal of pharmacological sciences 2011, 116, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Kopanska, M.; Muchacka, R.; Czech, J.; Batoryna, M.; Formicki, G. Acrylamide toxicity and cholinergic nervous system. J Physiol Pharmacol 2018, 69, 847–858. [Google Scholar] [CrossRef]
- Ha, Z.Y.; Mathew, S.; Yeong, K.Y. Butyrylcholinesterase: a multifaceted pharmacological target and tool. Current Protein and Peptide Science 2020, 21, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, J.; Miao, Y.; Cui, Q.; Zhao, W.; Zhang, J.; Wang, H. Pinocembrin protects SH-SY5Y cells against MPP+-induced neurotoxicity through the mitochondrial apoptotic pathway. Journal of Molecular Neuroscience 2014, 53, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Khanzadeh, T.; Hagh, M.F.; Talebi, M.; Yousefi, B.; Azimi, A.; Baradaran, B. Investigation of BAX and BCL2 expression and apoptosis in a resveratrol-and prednisolone-treated human T-ALL cell line, CCRF-CEM. Blood research 2018, 53, 53. [Google Scholar] [CrossRef] [PubMed]
- Chong, F.P.; Ng, K.Y.; Koh, R.Y.; Chye, S.M. Tau proteins and tauopathies in Alzheimer’s disease. Cellular and molecular neurobiology 2018, 38, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Manna, J.; Veleri, S.; Frautschy, S. Molecular chaperone dysfunction in neurodegenerative diseases and effects of curcumin. BioMed research international 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Henneman, W.; Sluimer, J.; Barnes, J.; Van Der Flier, W.; Sluimer, I.; Fox, N.; Scheltens, P.; Vrenken, H.; Barkhof, F. Hippocampal atrophy rates in Alzheimer disease: added value over whole brain volume measures. Neurology 2009, 72, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Westman, E.; Simmons, A.; Muehlboeck, J.-S.; Mecocci, P.; Vellas, B.; Tsolaki, M.; Kłoszewska, I.; Soininen, H.; Weiner, M.W.; Lovestone, S. AddNeuroMed and ADNI: similar patterns of Alzheimer's atrophy and automated MRI classification accuracy in Europe and North America. Neuroimage 2011, 58, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Memudu, A.E.; Adewumi, A.E. Alpha lipoic acid ameliorates scopolamine induced memory deficit and neurodegeneration in the cerebello-hippocampal cortex. Metabolic Brain Disease 2021, 36, 1729–1745. [Google Scholar] [CrossRef] [PubMed]
- El-Beeh, M.E.; El-Badawi, A.A.; Amin, A.H.; Qari, S.H.; Ramadan, M.F.; Filfilan, W.M.; El-Sayyad, H.I. Anti-aging trait of whey protein against brain damage of senile rats. Journal of Umm Al-Qura University for Applied Sciences 2022, 8, 8–20. [Google Scholar] [CrossRef]
- Ohia-Nwoko, O.; Montazari, S.; Lau, Y.-S.; Eriksen, J.L. Long-term treadmill exercise attenuates tau pathology in P301S tau transgenic mice. Molecular neurodegeneration 2014, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.-K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer's disease. Life sciences 2020, 257, 118020. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, C.K.; Kelly, A.M. Differential BDNF signaling in dentate gyrus and perirhinal cortex during consolidation of recognition memory in the rat. Hippocampus 2012, 22, 2127–2135. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Kwon, M.Y.; Pak, M.E.; Park, S.H.; Baek, J.U.; Choi, B.T. Beneficial effects of gagam-palmultang on scopolamine-induced memory deficits in mice. Evidence-Based Complementary and Alternative Medicine 2018, 2018. [Google Scholar] [CrossRef]
- Tanila, H. The role of BDNF in Alzheimer's disease. Neurobiology of disease 2017, 97, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nature Reviews Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, M.; SoukhakLari, R.; Moezi, L.; Pirsalami, F. Scopolamine-induced passive avoidance memory retrieval deficit is accompanied with hippocampal MMP2, MMP-9 and MAPKs alteration. European Journal of Pharmacology 2018, 819, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O'Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V. The microbiota-gut-brain axis. Physiological reviews 2019. [CrossRef] [PubMed]
- Gareau, M. Cognitive function and the microbiome. International review of neurobiology 2016, 131, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Almasi, F.; Mohammadipanah, F.; Adhami, H.R.; Hamedi, J. Introduction of marine-derived Streptomyces sp. UTMC 1334 as a source of pyrrole derivatives with anti-acetylcholinesterase activity. Journal of applied microbiology 2018, 125, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Bairamian, D.; Sha, S.; Rolhion, N.; Sokol, H.; Dorothée, G.; Lemere, C.A.; Krantic, S. Microbiota in neuroinflammation and synaptic dysfunction: a focus on Alzheimer’s disease. Molecular neurodegeneration 2022, 17, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Wu, X. Modulation of the Gut Microbiota in Memory Impairment and Alzheimer’s Disease via the Inhibition of the Parasympathetic Nervous System. International Journal of Molecular Sciences 2022, 23, 13574. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Deng, L.; Lu, Z.; Wu, F.; Liu, W.; Huang, D.; Peng, Y. Protective effects of Akkermansia muciniphila on cognitive deficits and amyloid pathology in a mouse model of Alzheimer’s disease. Nutrition & Diabetes 2020, 10, 12. [Google Scholar] [CrossRef]
- Liu, Z.; Qiao, K.; Tian, L.; Zhang, Q.; Liu, Z.-Y.; Li, F.-L. Spontaneous large-scale autolysis in Clostridium acetobutylicum contributes to generation of more spores. Frontiers in Microbiology 2015, 6, 950. [Google Scholar] [CrossRef]
Figure 1.
Effect of WPH and treadmill exercise on cognitive behaviors in mice in the Y-maze and novel object recognition test. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, not significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
Figure 1.
Effect of WPH and treadmill exercise on cognitive behaviors in mice in the Y-maze and novel object recognition test. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, not significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
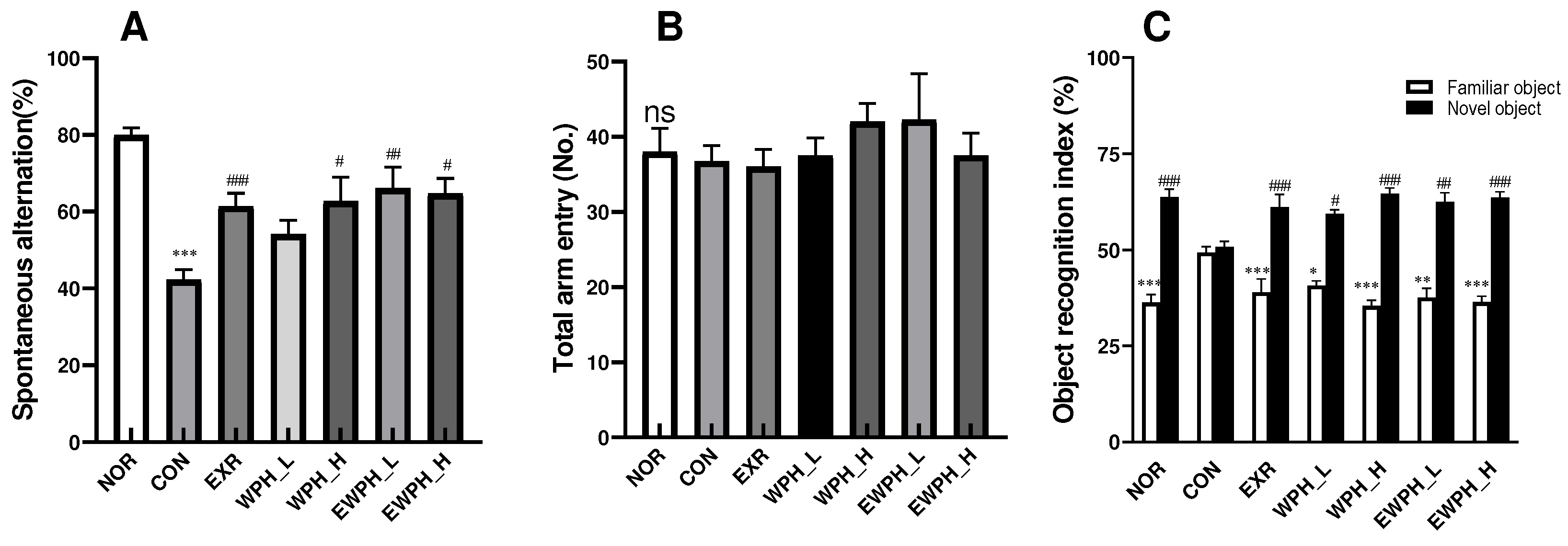
Figure 2.
Effect of WPH and treadmill exercise on reactive oxygen species (ROS) and malondialdehyde (MDA) production. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The levels of ROS (A) and MDA (B) were determined using DCFH-DA and TBRARS assays, respectively. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
Figure 2.
Effect of WPH and treadmill exercise on reactive oxygen species (ROS) and malondialdehyde (MDA) production. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The levels of ROS (A) and MDA (B) were determined using DCFH-DA and TBRARS assays, respectively. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
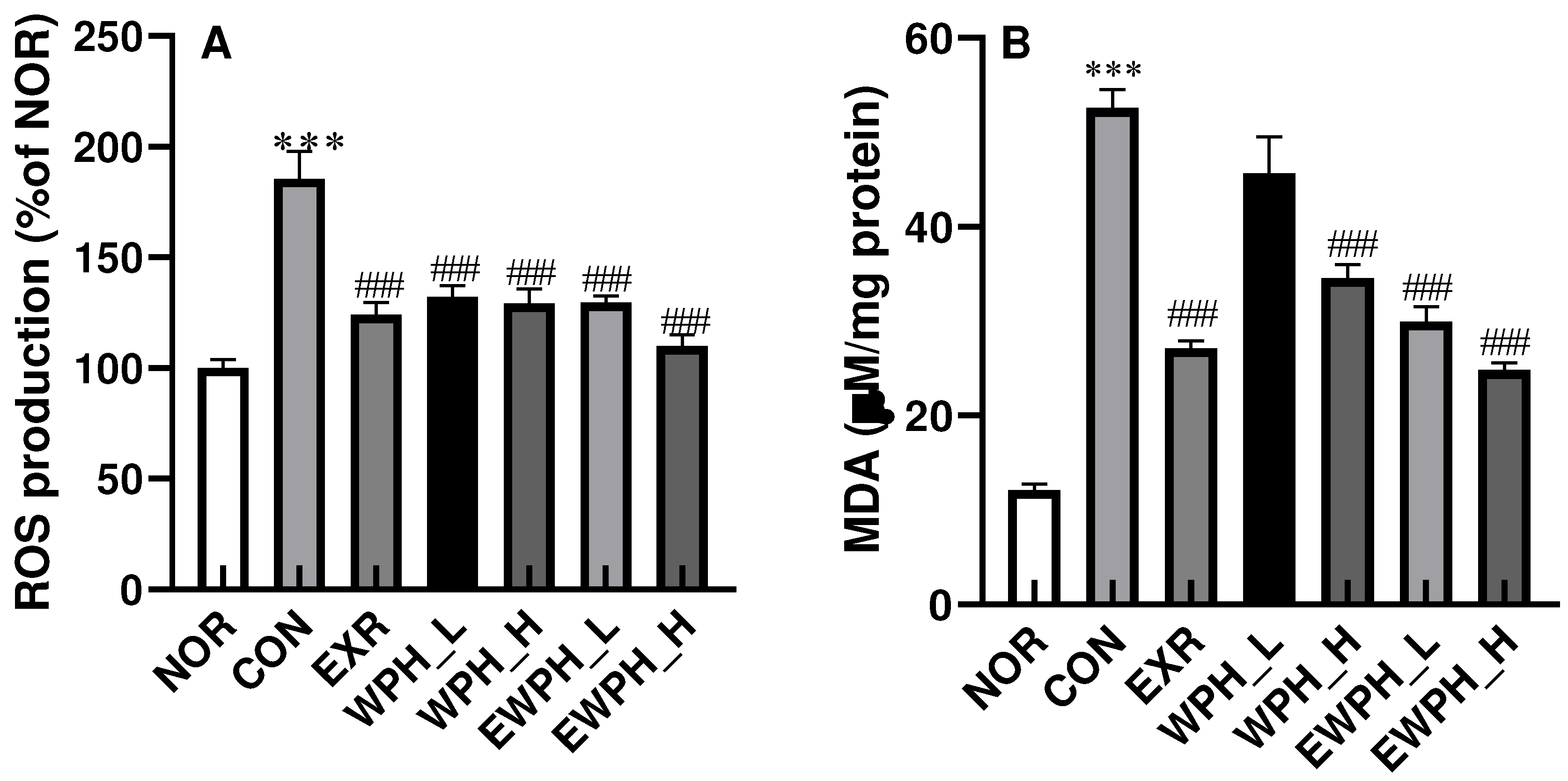
Figure 3.
Effect of WPH and treadmill exercise on ACh, AChE, and ChAT. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate; ACh, acetylcholine; AChE, acetylcholine esterase; ChAT, choline acetyltransferase. The ACh content in the brain was measured using a colorimetric assay at 540 nm (A). AChE activity was examined using a specific assay kit (B). ChAT protein levels from the brain were determined using western blot (C) and quantified by ImageJ (D). Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
Figure 3.
Effect of WPH and treadmill exercise on ACh, AChE, and ChAT. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate; ACh, acetylcholine; AChE, acetylcholine esterase; ChAT, choline acetyltransferase. The ACh content in the brain was measured using a colorimetric assay at 540 nm (A). AChE activity was examined using a specific assay kit (B). ChAT protein levels from the brain were determined using western blot (C) and quantified by ImageJ (D). Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
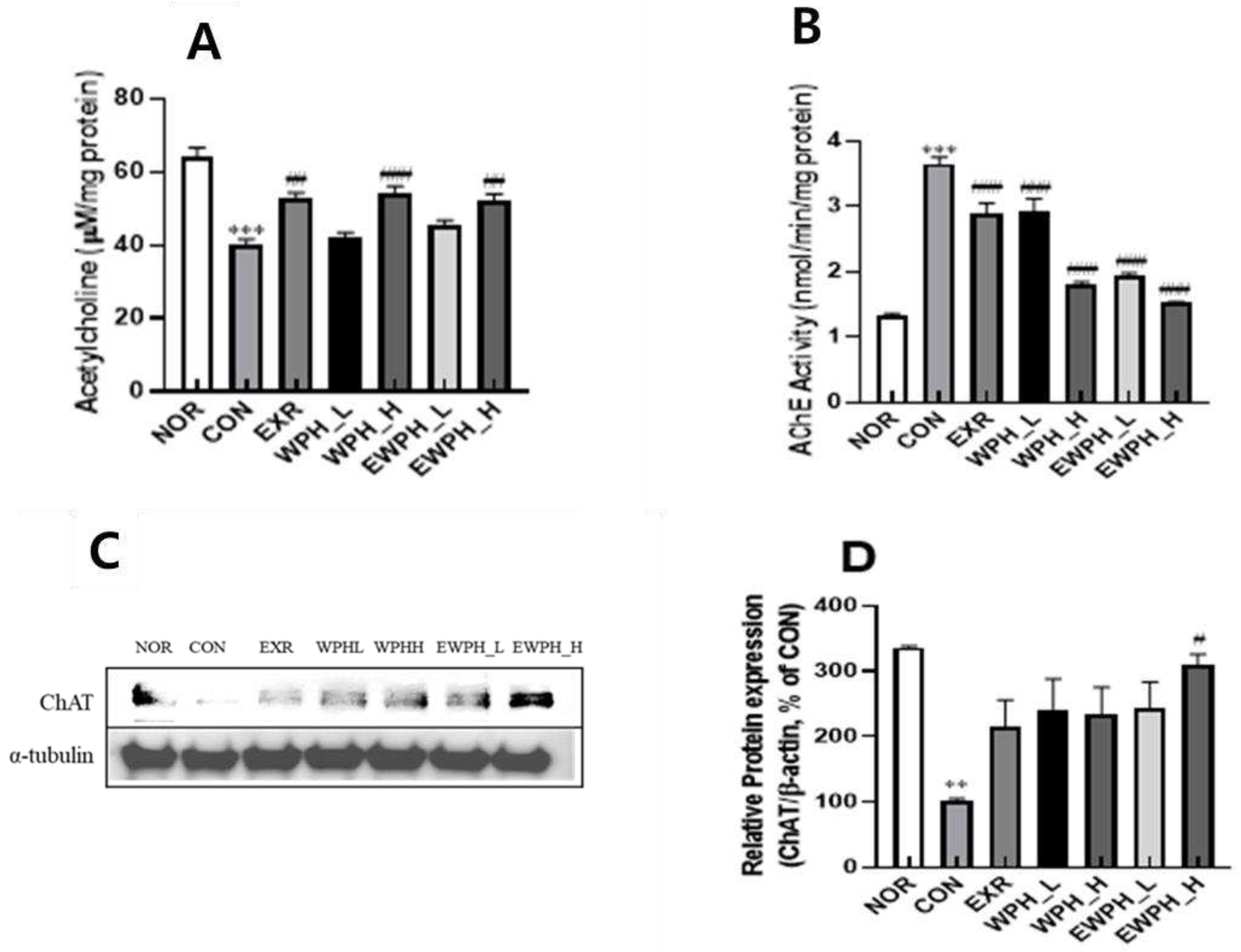
Figure 4.
Effect of WPH and treadmill exercise on the relative protein ratios of Bax/Bcl-2, BDNF, and p-tau/tau. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Bax, Bcl-2, BDNF, and p-tau, and tau protein levels were determined using western blot (A), and the Bax/Bcl2 (B), BDNF (C), and p-tau/tau (C) ratios were measured by ImageJ. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
Figure 4.
Effect of WPH and treadmill exercise on the relative protein ratios of Bax/Bcl-2, BDNF, and p-tau/tau. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Bax, Bcl-2, BDNF, and p-tau, and tau protein levels were determined using western blot (A), and the Bax/Bcl2 (B), BDNF (C), and p-tau/tau (C) ratios were measured by ImageJ. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
Figure 5.
Effect of WPH and treadmill exercise on relative protein expression levels of the mitogen-activated protein kinase (MAPK) signaling pathway. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Relative protein levels of p-JNK/JNK (B), p-ERK/ERK (C), and p-p38/p38 (D) were determined using western blot. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
Figure 5.
Effect of WPH and treadmill exercise on relative protein expression levels of the mitogen-activated protein kinase (MAPK) signaling pathway. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. Relative protein levels of p-JNK/JNK (B), p-ERK/ERK (C), and p-p38/p38 (D) were determined using western blot. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group (analysis of variance followed by post-hoc Tukey’s test).
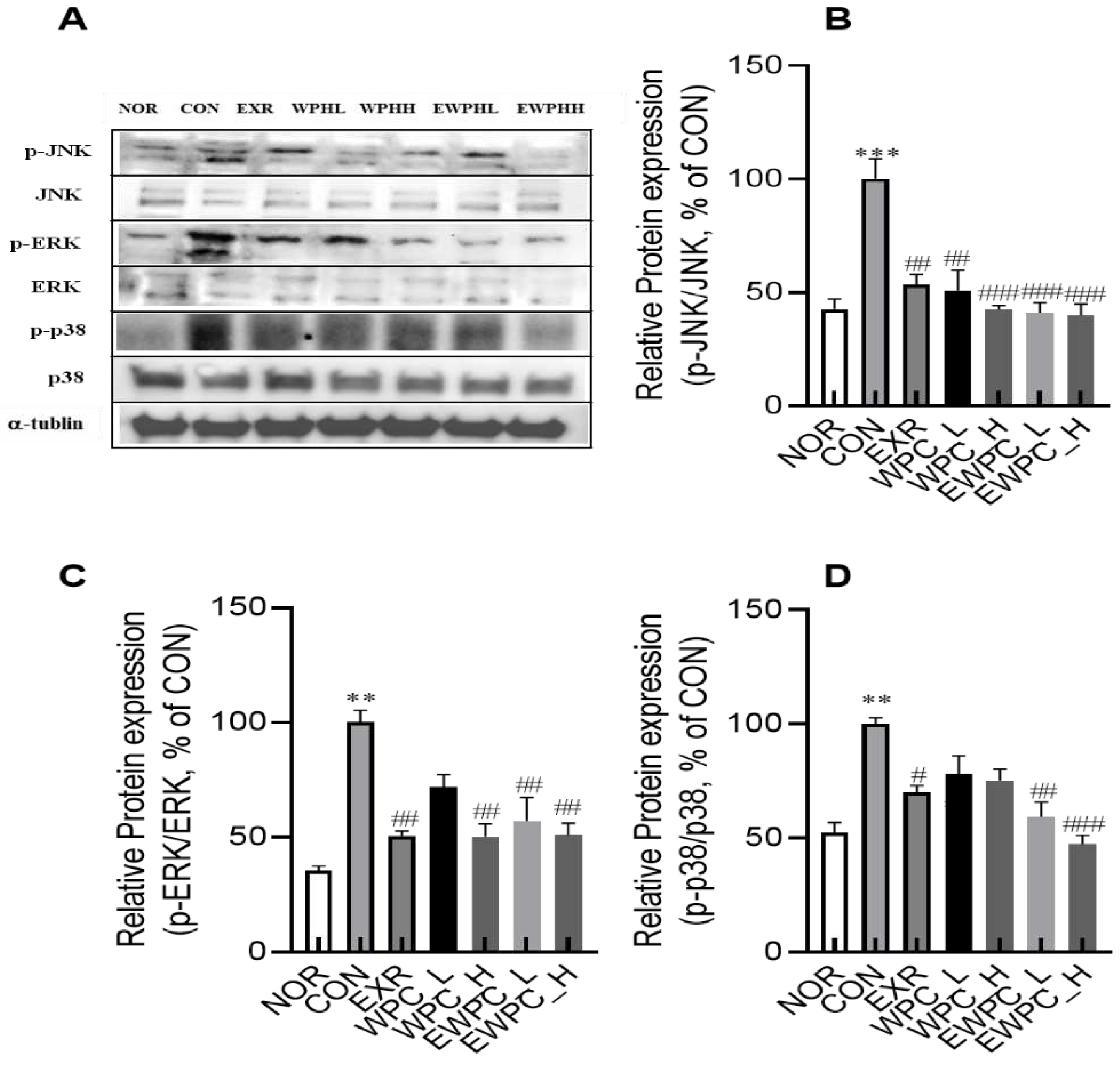
Figure 6.
Effect of WPH and treadmill exercise on the gut microbiome composition at the phylum level. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The gut microbiome at the phylum level was analyzed using rRNA gene sequencing. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, non-significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
Figure 6.
Effect of WPH and treadmill exercise on the gut microbiome composition at the phylum level. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The gut microbiome at the phylum level was analyzed using rRNA gene sequencing. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, non-significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
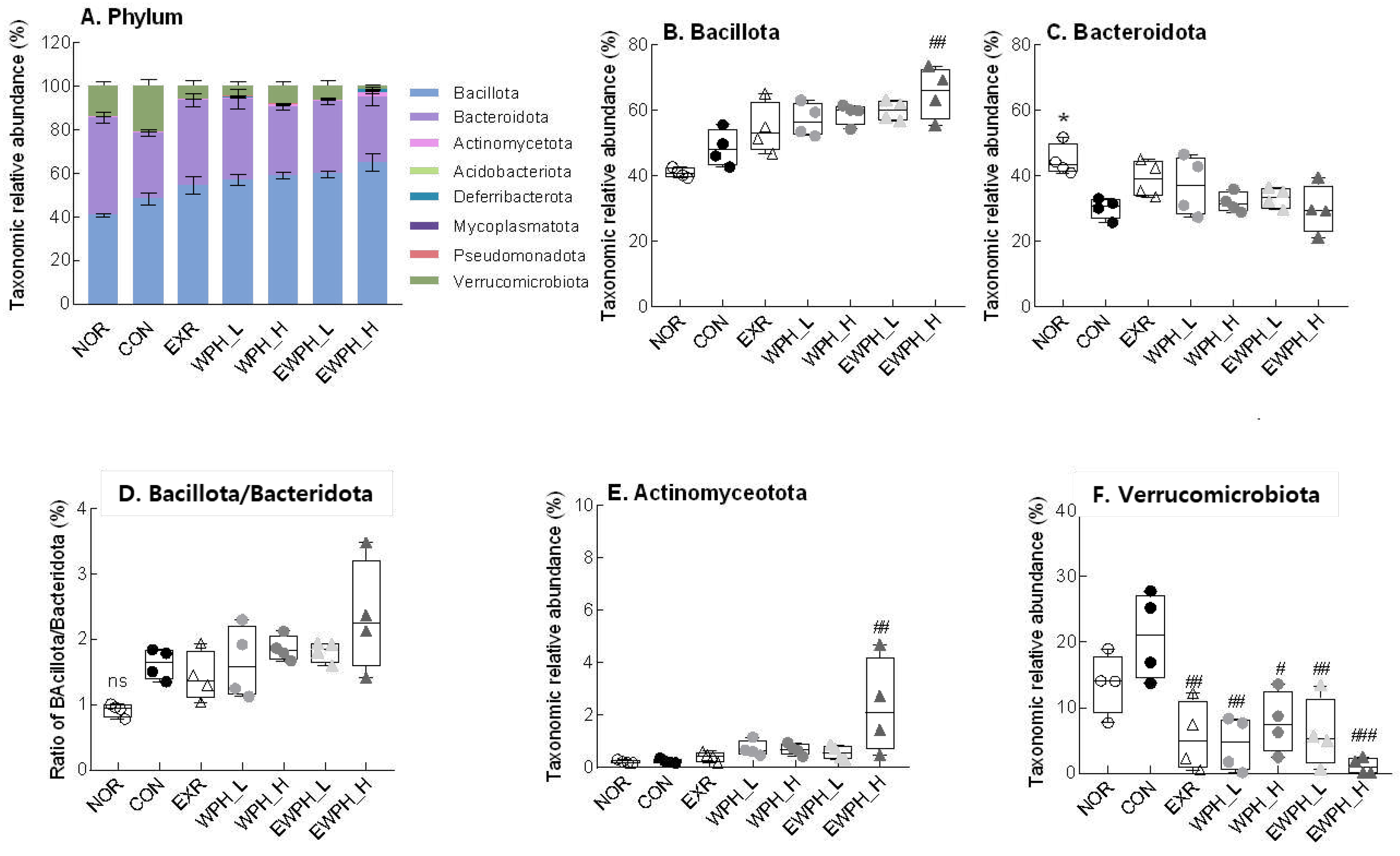
Figure 7.
Effect of WPH and treadmill exercise on gut microbiome composition at the genus level. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The gut microbiome at the genus level was analyzed using rRNA gene sequencing. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, non-significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
Figure 7.
Effect of WPH and treadmill exercise on gut microbiome composition at the genus level. NOR, normal; CON, control; EXR, exercise; WPH-L/H, low-dose/high-dose whey protein hydrolysate; EWPH_L/H, exercise + low-dose/high-dose whey protein hydrolysate. The gut microbiome at the genus level was analyzed using rRNA gene sequencing. Data are presented as mean ± standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001 vs. NOR group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. CON group; ns, non-significant (p > 0.05) (analysis of variance followed by post-hoc Tukey’s test).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



