
Submitted:
02 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
The present review critically examines the advancements in the past 5 years concerning the re-search on the bioavailability of the nanoplastics (NPLs) to freshwater plankton. With a specific emphasis on two bioavailability components: uptake availability and toxico-availability, we dis-cuss the recent progress in the understanding of the adsorption, absorption, trophic transfer and biological effects in phyto- and zooplankton induced by NPLs exposure. The influence of the plankton on the NPLs bioavailability via excretion of the biomolecules and formation of eco-corona is also examined. In spite of the important research developments, there are still con-siderable knowledge gaps with respect to NPLs biouptake and trophic transfer by plankton, as well as a potential adverse effect in natural aquatic systems. As plankton play a critical role in primary production, nutrient cycling and food web structure, understanding the interaction be-tween NPLs and plankton is essential in assessing the potential implications of the nanoplastics pollution for aquatic ecosystem biodiversity and services.
Keywords:
nanoplastics
; uptake
; toxicity
; trophic transfer
; algae
; cyanobacteria
; Daphnia
; EPS
; ecocorona
1. Introduction
The use of various plastic materials is continuously growing leading to an increase of plastic waste. Consequently, the concentrations of micro- and nano-plastics (NPLs1) in the environment are increasing [1,2,3]. In parallel, the concerns about the environmental implications of NPLs are rising [4,5]. Currently, the NPLs, originating from primary or secondary sources, are the least studied area of plastic pollution, however potentially the most hazardous, given their small size and high surface-specific area [4,5]. Indeed, under environmental conditions, various plastic debris can release a large amount of NPLs through various physical and chemical degradation pathways, as comprehensively reviewed by[6]. NPLs are characterized by very small size and high reactivity, which distinguish them from microplastics [7]. NPLs interact with different abiotic and biotic components in aquatic environment, and thus they are transformed by various interconnected processes involving aggregation, sedimentation, chemical and physical alterations etc. which greatly affect the particles’ properties, their reactivity, fate and impact [3,5,6,8,9].
Given the limitations of the existing analytical techniques, the concentrations of the NPLs in the environment are not yet measured. However, the estimates show that more than 5 trillions of pieces with the size between 300 μm and 5 mm are floating in the ocean [12]. As the smaller particles are usually more abundant in number, it can be assumed that comparable or larger number of NPLs will be present in the aquatic environment. Multi-media model estimations give average concentrations of NPLs for surface water of 280 µg L-1 [13]. More recently, the NPLs abundance have been estimated to be in the range 0.3 – 488 μg L-1 in freshwater environments, values higher than those for marine environment (2.7–67 μg L-1)[14]. These concentrations are lower as compared with the predicted no-effect concentrations (PNECs) derived from the probabilistic species sensitivity distributions, resulting in values of 99 μg L–1 and 72 μg L–1 for the freshwater and marine datasets, [15]. They are also below the estimated hazard concentration affecting 5% of the species (HC5) of 410 μg L-1 for marine plankton for two types of materials polymethylmethacrylate (PMMA) and polystyrene (PS) [16], as well as HC5 for NPLs with a size of 50 nm in freshwater 187.9 (8.0–2978.3) μg L-1 [17]. Nevertheless, some hotspots of NPLs pollution be of risk for aquatic biota. Critical gaps in NPLs research and their connection to risk assessment show the pattern of the work that does not answer a number of key factors vital to calculating risk that takes into account the smallest plastic particles; namely, sources, fate and transport, exposure measures, toxicity and effects [18].
A lot of attention is paid to the NPLs bioaccumulation and adverse effects at the high level of the trophic webs [14]. Advances in the understanding of their absorption, distribution, metabolisation, and excretion within organisms’ bodies have been recently put in perspective and revealed many challenges [19]. The accumulation of NPLs in aquatic organisms results in adverse effects on different freshwater organisms [3,4,5,6,8,14,20,21,22,23,24,25,26]. A plethora of the such effects, including oxidative stress and damage, inflammation, altered development, reduced growth, energy and movement, genotoxicity etc. have been reported as recently reviewed [14,22,25,27,28,29,30]. Different factors, such as particle nature, concentration and size, exposure time and co-factors such as contaminants, food availability, species, developmental stage and environmental conditions were also thoroughly discussed [22,28,30,31].
In the present review paper, we comprehensively reviewed the progress in the past 5 years concerning the bioavailability of the NPLs to the freshwater plankton. The biological availability (or bioavailability) is defined as “the extent of absorption of a substance by a living organism compared to a standard system” [32]. Bioavailability is considered a key concept allowing to quantitatively relate the changes in the (dissolved or particulate) pollutant concentrations, including nanoparticles, with the intensity of the biological response in biota [33,34]. It was proposed that the concept of bioavailability contains three main components: "exposure availability," "actual or potential uptake availability," and "toxico-availability" [34]. Furthermore, different biological endpoints in addition to the concentration of bioaccumulated pollutant, can be considered as an indication of bioavailability, including effects on the growth rates, reproduction, oxidative stress or photosynthesis etc.
Plankton, consisting of free-floating phytoplankton and zooplankton, represent a critical component of aquatic ecosystems, playing a central role in driving primary production, shaping food webs, and influencing nutrient cycling [35]. Therefore, understanding the interaction between NPLs and plankton is essential in assessing the potential consequences for aquatic ecosystems. Their contributions to primary production, nutrient cycling, and trophic interactions are pivotal in maintaining biodiversity and ecosystem services.
In this paper we specially focuss on the two key components of bioavailability: uptake availability and toxico-availability, which can be considered as a result of several key processes (Figure 1a): (i) Adsorption on plankton: NPLs diffuse towards and can adsorb onto the surfaces of plankton, including both phytoplankton and zooplankton.; (ii) Absorption by plankton NPLs can penetrate (or not) the cell wall and membrane of phytoplankton species. Zooplankton can ingest NPLs along with their regular diet of suspended particles. Once ingested, NPLs may be retained within the digestive tracts or tissues; (iii) Trophic transfer from phytoplankton to zooplankton: NPLs enter the food web at lower trophic levels, such as phytoplankton, and subsequently be consumed by higher trophic level organisms, including zooplankton. (iv) Biological effects and NPLs transformation. NPLs can degrade within the bodies of planktonic organisms, releasing smaller plastic fragments or chemical components that can be further absorbed or retained by the organisms. (v) Excretion of the NPLs. In addition to the above-mentioned processes, in the present review we will discuss how the plankton species can affect the bioavailability of the NPLs by secreting different biomolecules.
Similarly to the other nano-sized materials, the bioavailability of the NPLs to plankton depends on many factors, including (Figure 1b) (i) the type and characteristics of the NPLs, such as its chemical composition, size, shape, surface functionalization etc.; (ii) the physicochemical parameters of the aquatic environment, including pH, water hardness and alkalinity; (iii) the presence and concentrations of other dissolved, nanoparticulate and colloidal forms of contaminants; (iv) the presence and concentrations of different ligands from natural and anthropogenic origins, which could absorb on the NPLs and influence their fate and impact; (v) the plankton characteristics, such as type, cell wall, differentiation stage and its pathways of particle uptake and cellular processing.
2. NPLs Interaction with Freshwater Phytoplankton Species
Table 1 summarizes the most recent studies published in the last 5 years dealing with the interactions of NPLs with microalgae and cyanobacteria and the progress is discuss below.
2.1. Advances in Uptake Availability of NPLs to Phytoplankton
The uptake and effects of NPLs on microalgae have been recently reviewed [27]. By analogy with engineered nanoparticles, potential mechanisms governing the bioavailability could include absorption on the surface of microalgae, penetration into the cell either by endocytosis or physical damage, shading and blocking the exchange of substances and energy with ambient medium [27]. However, up to now the scientific evidences concerning uptake availability of NPLs to phytoplankton are still very limited. Both amidine and carboxyl functionalized polystyrene NPLs (PS-COOH) adsorbed on marine diatom Dunaliella tertiolecta, however only amidine PS induced inhibition of algal growth (concentration inducing effect in 50% of algal population, EC50 12.97 μg mL-1) [36]. Fluorescent-blue 50 nm polystyrene amino NPLs (PS-NH2) adsorbed on the diatom Chaetoceros neogracile impaired photosynthetic machinery, increased of ROS production and alter the transparent exopolymeric particles at low (0.05 μg mL−1) and high (5 μg mL−1) exposure concentrations [37]. The adsorption onto the cells of green alga Pseudokirchneriella subcapitata was higher for 100 mg L-1 neutral and positively charged PS-NH2. Negatively charged PS-COOH adsorption onto the algal cell wall was very low but increased with water hardness [38]. These examples demonstrated the important role of surface charge of primary NPLs particles and the algal species specificity of these interactions. The PS NPLs ad/absorbed rapidly onto/in Phaeodactylum tricornutum as revealed by an increase in cell complexity, cell size and microalgae fluorescence caused by 100 nm fluoresbrite [39]. Fluorescent PS of 51 nm attached to the surface and penetrated the outer layer of Chlamydomonas reinhardtii during cell division [40]. More recently, a study with metal doped PS, revealed that more than 60% of Fe-PS or Eu-PS for 72 h were associated with algal cells of P. subcapitata [41]. A pioneer study has revealed that the fluorescent aggregation-induced emission fluorogens-incorporated nanoparticles (AIE-NPs) of 40, 70, and 85 nm were internalized intracellularly via clathrine - dependent endocytosis in C. reinhardtii, while the 140 nm AIE-NPs were attached to the surface [42]. Importantly the authors have demonstrated that the endocytosis, as well as algal cell membrane permeability, and exopolymer substance thickness playing the role in internalization processes are cell cycle - dependent [42].
2.2. Advances in Toxico-Availability of NPLs to Phytoplankton
Recent review paper systematically presented what influence toxicological effects of MPLs/NPLs and other contaminants could have on phytoplankton and aquatic environment [43]. Behavior and major known adverse effects of PS NPLs having positive and negative surface charge to marine plankton have been also reviewed [44]. For example, exposure of marine green microalga Platymonas helgolandica to 20, 200, and 2000 μg L-1 70 nm PS beads demonstrated inhibition of the algal growth during the 3 days, but stimulated after 5 days of exposure. High concentrations of 200 and 2000 μg L-1 PS increased the membrane permeability and mitochondrial membrane potential, and decreased light energy used in photochemical processes of microalgae, as well as the damage on cell morphology and organelle function [45]. The exposure to 10-200 mg L-1 100 nm PS of green alga Scenedesmus quadricauda resulted even in the stimulation of the growth. At high concentration of 200 mg L-1 PS induced increased the contents of chlorophyll, soluble protein, and polysaccharide and activated the antioxidant enzyme activities [46]. Exposure to 10-100 mg L-1 100 nm PS caused dose-dependent adverse effects on Chlorella pyrenoidosa growth from the lag to the earlier logarithmic phases. However, from the end of the logarithmic to the stationary phase, C. pyrenoidosa could cope with the adverse effects of microplastics jointly through cell wall thickening, algae homo-aggregation and algae-microplastics hetero-aggregation, hence triggering an increase of algal photosynthetic activity and its growth, and cell structures turned to normal [47].
Further studies have reported that the exposure of various alga and cyanobacteria to different NPLs resulted in the oxidative stress, membrane damages, and alteration of the photosynthesis. PS beads of 50 and 100 nm after 24 h induced an increase in oxidative stress biomarkers, damage to the photosynthetic apparatus, DNA damage and depolarization of mitochondrial and cell membrane in the marine diatom P. tricornutum at concentrations from 5 mg L−1 [39]. Exposure of C. pyrenoidosa to 5 -50 mg L−1 80 nm PS for 24-48 h, resulted in a decrease of algal growth, chlorophyll a, and Fv/Fm by 27.73%, 29.64%, and 11.76%, however, these effects decreased with exposure time at 96 h. The transcriptomics revealed that the inhibition effect of NPLs was due to the blockage of the gene expression of aminoacyl tRNA synthetase and the synthesis of related enzymes and proteins at low concentrations (10 mg L−1) effect on DNA damage repair and hindered photosynthesis at high concentrations (50 mg L−1) [48]. 28 - day exposure of Chlorella vulgaris to 250 mg L-1 carboxyl-functionalized and non-functionalized PS of 20 and 50 nm revealed a reduction in algal cell viability and chlorophyll a concentration an increase of the lactate dehydrogenase activity and reactive oxygen species concentration, together with increased cell size, deformed cell wall and increased volume of starch grains [49]. 50 mg L-1, 100 nm PS inhibited the growth of Euglena gracilis by 35.5%, which was significantly higher than the effect of 5 μm PS-MPs (27.9%) in 96h exposure. PS of both sizes significantly reduced pigment contents, altered SOD and POD activities, dysregulated the expression of genes involved in cellular processes, genetic information processing, organismal systems, and metabolisms [50]. By contrast 1 mg L-1, 5 μm PS exhibited stronger growth inhibition and physiological toxicity than 100 nm PS in 96h exposure of Euglena gracilis [51]. Positively charged amino-modified polystyrene nanoparticle (PS-NH2) of 50 nm induced growth inhibition of cyanobacterium Synechococcus elongatus (48h EC50 of 3.81 mg L−1). The oxidative stress, destruction, disruption of glutathione metabolism and damage to membrane integrity were confirmed as main toxicity mechanisms [52]. Negatively charged amino-modified PS with size of 500 nm at 2.5 μg mL−1were not attached to the microalgal cell and do not induced changes in the growth, morphology, photosynthesis, reactive oxygen species levels and membrane potential however, significantly decreased the cellular esterase activity and the neutral lipid content suggesting that the cells modulate their energy metabolism in response to stress [53]. Exposure of C. reinhardtii to increasing concentrations (5, 25, 50 and 100 mg L-1) of 300–600 nm PS reduces chlorophyll a fluorescence yields and photosynthetic activities. The PS beads wrapped the surface of microalgae and damaged their membrane[54]. Pd-doped PS-NPLs affected the growth of both filamentous cyanobacterium Anabaena sp. (72hEC50 151.3 ± 22.5 mg L-1) and the green alga C. reinhardtii (72hEC50 of 247.8 ± 32.7 mg L-1), but the cyanobacterium was more sensitive than the green alga. In addition, a dose-dependent reactive oxygen species overproduction, membrane damage and metabolic alterations in both algae, but ROS overproduction and damages were less pronounced in C. reinhardtii [55].

Table 1.
Selected examples of the most recent studies (<5 years) researching phytoplankton interactions with NPLs.
Table 1.
Selected examples of the most recent studies (<5 years) researching phytoplankton interactions with NPLs.
| Species | Type of NPLs | Size of NPLs | Concentration | Duration | Observed effects | Reference |
|---|---|---|---|---|---|---|
|
Alexandrium tamarense (marine dinoflagellate) |
PS (plain) | 100 nm 1 µm |
0, 0.1, 1, 5, 10, 50, 100 mg L-1 | 4 days | Inhibition of growth, photosynthetic production and extracellular carbonic anhydrase activities stronger in MPLs than in NPLs. Intracellular paralytic shellfish toxins production stimulated by NPLs and decreased by MPLs. | [56] |
|
Anabaena sp. (freshwater cyanobacteria) C. reinhardtii (freshwater green algae) |
PHB (polyhydroxybutyrate, mechanically broken-down) | 200 nm | 0, 50 mg L-1 | 3 days | Decrease in growth, increase in ROS production and membrane damage, secondary NPLs may be more toxic than primary. Biodegradable plastics show the same toxic effects to organisms as non-biodegradable. | [57] |
|
Chlorella sp. (freshwater green algae) |
PS (plain) PS-NH2 PS-COOH |
200 nm | 0, 1 mg L-1 | 3 days | EPS aged NPLs significantly lowered the oxidative stress and cytotoxic impact, eco-corona may change the way NPLs interact with the organisms. | [58] |
|
Chlorella vulgaris (freshwater green algae) |
PS (plain) PS-COOH |
20, 50, 500 nm |
0, 250 mg L-1 | 28 days | Smaller NPLs have a higher impact - decrease in algal viability and pigments; increase in ROS, lactate dehydrogenase activity and starch grains content; shrinkage in cell wall. Bigger PS could aggregate and sediment making them non-bioavailable. | [49] |
|
Chlorella pyrenoidosa (freshwater green algae) |
PS (plain) | 100 nm 1 µm |
0, 10, 50, 100 mg L-1 |
30 days | Hetero- and homoaggregation observed, EPS production increased, during the first phase growth rate and photosynthesis decreased, while in the second phase growth and photosynthesis recovered. | [47] |
| PS (plain) | 80 nm | 0, 5, 10, 20, 30, 40, 50 mg L-1 | 4 days | Strong inhibition of growth, photosynthetic pigments and efficiency after 24–48 h, after 96 h inhibition lowered. Heteroaggregation, ROS production, gene expression changes, membrane and DNA damage observed. | [48] | |
|
Chlamydomonas reinhardtii (freshwater green algae) |
PS (plane) | 300–600 nm | 0, 5, 25, 50, 100 mg L-1 |
10 days | A decrease in growth, photosinthetic activity and EPS follows an increase in concentration, observed higher soluble proteins and membrane damage. | [54] |
| PS (fluorescent) | 51 nm | 0, 20, 40, 60, 80, 100 mg L-1 | 2 days | Adsorped to the surface of algae, passing into the outer layer when the cell is dividing. | [40] | |
|
Cocconeis placentula var lineata (freshwater diatom) |
P(Sco-MMA) (poly(styrene-co-methyl methacrylate) ) | 100 - 2800 nm |
0, 0.0001, 0.001, 0.1,10 mg L-1 |
28 days | Significant increase in teratogenic effects in the lowest concentration (deformed valve outline, changes in characteristics of longitudinal and central area, and mixed type). | [59] |
|
Dunaliella tertiolecta (marine green algae) |
PS-COOH (fluorescent) | 40 nm | 0, 0.5, 1, 5, 10, 25, 50 mg L-1 | 3 days | Aggregation, adsorbed on the surface of algae, potential trophic transfer. | [36] |
| PS-NH2 | 50 nm | Aggregation, inhibition of algal growth. | ||||
|
Euglena gracilis (freshwater euglena) |
PS (fluorescent) | 100 nm, 5 µm |
1 mg L-1 (NPLs or MPLs) +/- 0.5 mg L-1 (Cd2+) |
4 days | MPLs alone inhibits the growth while mixture with Cd2+ increases it. NPLs shows lower toxicity than MPLs while in mixture with Cd2+ acts synergistically and exceed toxic effects. | [51] |
|
Microcystis aeruginosa (freshwater cyanobacteria) |
PS (plain) | 60 nm | 0, 25, 50, 100 mg L-1 |
30 days | Growth inhibited at the beginning while aggregation rates were high. After 10 days growth increases, while aggregation decreases, connection between growth rate and aggregation. Negative effect on photosynthetic activity, SOD and MDA affected in the beginning, then mitigated. Production of microcystin increased with the concentration increase. | [60] |
|
Phaeodactylum tricornutum (marine diatom) |
PS (plain and fluorescent) | 50, 100 nm | 0, 0.1, 1, 5, 10, 20, 50 mg L-1 | 3 days | Hetero- and homoaggregation observed, during the first 24 hours changes in oxidative stress, photosynthesis, membrane integrity and DNA damage, while after 48 h these responses were mitigated. Growth, Chlat levels and fluorescence and protein content negatively influenced after 72 h. | [39] |
| PS-COOH | 60 nm | 0, 1, 5, 50, 100 mg L-1 |
3 days | EPS reduces aggregation and ROS production, toxicity of NPLs not observed with or without EPS. | [61] | |
|
Platymonas helgolandica0 (marine green algae) |
PS (plain) | 70 nm | 0, 0.02, 0.2, 2 mg L-1 |
6 days | Observed morphological changes, inhibition of growth during the first 4 days, increase in growth (after 5 days) and membrane permeability, disturbance in mitochondrial and chloroplast functions. | [45] |
|
Rhodomonas baltica (marine red algae) |
PMMA PMMA-COOH |
50 nm | 0, 0.5, 1, 5, 10, 25, 50, 100 mg L-1 |
3 days | PMMA aggregated, impacted cell viability and size, pigments, membrane integrity, ROS formation, lipid peroxidation, DNA content and photosynthetic capacity, while PMMA-COOH influenced viability, metabolic activity, photosynthetic performance and algal growth changes. PMMA physico-chemical charactaristics important in response to interaction with cells. | [62] |
|
Scenedesmus subspicatus (freshwater green algae) |
PE (plain) PE (from Atlantic Gyre, mechanically broken-down) |
<450 nm | 0, 0.001, 0.01, 0.1, 1, 10 mg L-1 |
2 days | PE from the Atlantic gyre negatively influencing algal growth more than plain PE, may be due to presence of other contaminants like metals. | [63] |
|
Scenedesmus quadricauda (freshwater green algae) |
PS (plane) | 100 nm | 0, 10, 25, 50, 100, 200 mg L-1 | 14 days | Increase in growth, antioxidant enzyme activity, pigments, soluble proteins and soluble polysaccharides. Observed strong defensive and recovery response to stress. | [46] |
|
Synechococcus elongatus (freshwater cyanobacteria) |
PS-NH2 | 50 nm | 2 – 9 mg L-1 | 2 days | PS-NH2 negatively impacted growth rate, PS-SO3H had no effect. PS-NH2 induced oxidative stress and membrane permeability which lead to damage. | [52] |
| PS-SO3H | 52.03 nm | |||||
|
Tetraselmis chuii, Nannochloropsis gaditana, Isochrysis galbana, Thalassiosira weissflogii (marine algae) |
PMMA | 40 nm | 0 - 304.1 mg L-1 | 3 days | Growth rates inhibited at higher concentrations with T. weissflogii being the most affected. Big aggregates observed which could explain higher tolerance to PMMA. | [16] |
A 96h-exposure to PMMA resulted in species-specific reduction of the growth rate of several marine microalgae Tetraselmis chuii (EC50 132.5 mgL-1), Nannochloropsis gaditana (EC50 116.5 mg L-1), Isochrysis galbana (EC50 123.8 mg L-1) and Thalassiosira weissflogii (EC50 83.4 mg L-1) [16]. PMMA and PMMA-COOH induced overproduction of pigments, loss of membrane integrity, hyperpolarization of the mitochondrial membrane, increased production of ROS and lipid peroxidation, decrease in DNA content and reduced photosynthetic capacity in red marine alga Rhodomonas baltica. It was suggested that PMMA interaction with cell walls inducing the formation of small holes in the lipid layer, that would later lead to permeabilization and internalization of small PMMA aggregates. The exposure to PMMA-COOH reduced the algal growth that was ascertained to an alteration of cell cycle and consequent decrease in cell viability, metabolic activity and photosynthetic performance [62], however, no results about the possible uptake and cell association to the algae were provided. PS-NPs as well as their combined effect with generation 7 PAMAM dendrimers (G7) on the filamentous cyanobacterium Anabaena sp. PCC7120, a relevant aquatic primary producer. PS-NPs were internalized in cyanobacterium Anabaena sp. and induced excessive generation of ROS, lipid peroxidation, membrane disruptions, intracellular acidification and a decrease in photosynthetic activity. Combined exposure to PS and poly(amidoamine) dendrimers of generation 7 (G7) significantly decreased their cellular internalization and thus lowered PS-NPs adverse effects [64]. Recently, a significant increase in the teratological frequency in diatom Cocconeis placentula 0.1 μg L-1 of poly(styrene-co-methyl methacrylate) (P(S-co-MMA)) with average size of 425.70 ± 175.02 nm was observed , but no effects on diatom growth was found in the concentration range 0.1, 1, 100, 10,000 μg L-1 for 28 days [59].
Novel evidences have been obtained demonstrating the effects of the secondary NPLs to phytoplankton species. The exposure to 48 h at 1 μg L-1 - 10 mg L-1 of reference polyethylene (PE) or NPLs made from PE collected in the North Atlantic gyre (PEN) of two microalgae, green alga Scenedesmus subspicatus and diatom Thalassiosira weissflogii revealed no influence on the cell growth of T. weissflogii but the PEN exposure causes growth inhibition of S. subspicatus [63]. The nano-sized fraction (< 100 nm) of the degraded polycaprolactone (PCL-plastics + PCL oligomers) induced overproduction of the ROS, alteration of the intracellular pH and metabolic activity in filamentous nitrogen-fixing cyanobacterium Anabaena sp., and the unicellular cyanobacterium Synechococcus sp., as well as inhibited nitrogen fixation in Anabaena sp.[65]. Secondary NPLs from degradation of polyhydroxybutyrate (PHB) significantly decreased the growth of both Anabaena and C. reinhardtii by 90 and 95%, respectively; increased intracellular ROS levels and induced membrane damage which were more pronounced in Anabaena [57].
Exposure to NPLs has been shown to alter the production of some toxins by algae as well as the release of the extracellular polymeric substances (EPS). For example, both 100 nm and 1 μm PS inhibited the growth, photosynthetic parameters, nutrients uptake and extracellular carbonic anhydrase activities (CAext) of harmful algal blooms-causing dinoflagellate Alexandrium tamarense, and the inhibition effects were more severe under 1 μm sized NPs than 100nm PS [56]. Interestingly, 100 nm PS increased while 1 μm PS decreased the concentrations of the intracellular paralytic shellfish toxins contents in this alga [56]. Increase in PS concentration from 25, 50, and 100 mg L-1 promoted the production and release of microcystine in M. aeruginosa, which reached values of 199.1 and 166.5 μg L-1 for the intracellular and extracellular microcystine at 100 mg L-1 PS [60]. Finally, exposure to NPLs could affect the production of the EPS (e.g. increase the release of EPS and protein-to-carbohydrate ratio) as shown for marine species T. pseudonana, Skeletonema grethae, P. tricornutum, and D. tertiolecta exposed to 1 mg L-1 PS [66]. Carboxyl-functionalized and non-functionalized PS of 20, 50 nm adsorbed onto the cell wall to C. vulgaris and induced generation of great amount of EPS material [49].
PS particles tend to be the most toxic while PE and PMMA have less effects [28]. Size of the NPLs is another key parameter affecting the effects of the NPLs to phytoplankton species. Indeed, the central role of the particle size and its role in biological interactions and physicochemical behavior of plastics in the environment is thoroughly discussed [67]. However, probabilistic species sensitivity distributions analysis of the available data did not reveal any substantial variance in ecotoxicity among NPLs of different sizes [15].
2.3. Phytoplankton Feedback on NPLs Uptake and Toxico-Availability
The phytoplankton have been shown to affect the bioavailability of the NPLs through the excretion of different extracellular polymeric substances and forming the eco-corona as recently reviewed [68,69]. EPS are ubiquitous in the environment [70], e.g. can represent up to 25% of natural organic matter in freshwaters, especially during algal blooms [71]. Polysaccharides and proteins are often the dominant components of the EPS, however their composition and properties varied with the species and environmental conditions [72]. The EPS produced by marine phytoplankton have been shown to form eco-corona on different NPLs which affects their reactivity [68]. For example, EPS of diatom Phaeodactylum tricornutum containing proteins of 30-100 kDa and high molecular weight carbohydrates, formed eco-corona on COOH-PS with a size of 60 nm, leading to reduced NPLs aggregation [61]. However, the corona formed on 23 nm PS by EPS of P. tricornutum, Ankistrodesmus angustus, and Amphora sp. resulted in a formation of gel like micrometer aggregates, presumably via hydrophobic interactions [73]. The eco-corona formation has been found to depend on NPLs size, charges, and incubation time [68]. Alginate, a model polysaccharide, formed eco-corona on amidine functionalized PS and modulated the surface charge, but the aggregation was negligible [74,75]. Aminated, carboxyl and plain NPLs aged in the EPS reduced the oxidative stress and lessen the toxic effects in marine alga Chlorella sp. [58].
3. NPLs Interactions with Freshwater Zooplankton Species
Numerous research papers have shown that the NPLs and their aggregates can be taken up by different zooplankton species through waterborne and foodborne exposure [76,77,78,79,80]. Among different zooplankton species, daphnids are the species being most studied model system [81]. Indeed, most of the studies used Daphnia magna (79%), followed by Daphnia pulex (18%), and Daphnia galeata (3%) [82]. Daphnia sp. are filter feeders and representative primary consumers linking primary producers to higher trophic levels in freshwaters [83]. Both “apical endpoints” (mortality/immobility, growth, feeding and egestion, swimming behavior, reproduction, embryonic development, body adsorption) and mechanistic endpoints (oxidative stress, detoxification, immune-related processes, neurotoxicity, energy metabolism, heart rate, changes in gut epithelium, moulting related processes) have been reported [82]. Based on the meta-analysis of the effect of the micro- and nano-plastics on the mortality and immobilization rates the influence of the particle properties (size, density, shape, surface coating, additives) and the Daphnia species characteristics (body size, clone, and the chosen brood), as well as the impacts of different qualities/quantities of food supply and the temperature on the toxicity outcome have been thoroughly discussed [84]. A bioconcentration factor (BCF) of 12 to 363 was calculated for the fluorescent PS of 1000 and 20 nm primary sizes in D. magna, but the values were lower in comparison with the BCF estimated for other carbonaceous nanoparticles, such as fullerene carbon nanotubes and graphene [85].
Below we will present only the recent advances obtained in the last 5 years (Table 2) concerning the uptake and toxico-availability of NPLs.
3.1. Advances in Uptake Availability via Direct Exposure
Further advances in the understanding and quantification of the uptake availability were obtained by the employment of the fluorescent, luminescent and metal-doped NPLs. For example, the exposure of D. magna to 100 nm fluorescent PS resulted in lower egestion and decreased feeding rates by 21%, however no effect on the reproduction was found over 21d period [86]. Fluorescent PS entered the inner gut of D. magna and induced histological damage to the intestinal walls (squashed and torn-out microvilli), but little or no acute toxicity was observed in the tested concentration range of 1 to 10 mg L-1 [40]. Fluorescently labeled 75 nm PS were found in the digestive organs of the Daphnia pulex [87]. Employing the fluorescent PS and palmitic acid-functionalized PS F-PA) and Al2O3 metal-core PS NPLs with size 90-95 nm their ingestion by D. magna was quantified to be higher by factors of 2.8 and 3.0 for PSNP/F-PA and 1.9 and 1.7 for PSNP/F when compared to PS [88].
Very recent study has demonstrated that D. magna rapidly ingested aggregated-induced emission (AIE) microplastic fluorogens plastics with size of 20 μm and 200 nm (50% of steady-state amount within 1 h), with particles of larger size and positive charge being ingested in higher amounts but egested in lower amounts. The presence of algae had a significant negative impact on NPLs uptake and deputation [89]. Comparing the positively charged AIE-NH2-NPs and negatively charged AIE-COOH-NPs of apprx. 230 nm and 21 μm demonstrated that the size and the surface charge of the AIE have altered the selectivity of D. magna. The daphnids selectively ingested larger and positively charged plastics which accumulated in the middle and posterior gut [90].
Fluorescent PS-COOH accumulated and retained in the gut of other zooplankton species rotifer Brachionus plicatilis, a retention was still present after the recovery period suggesting that PS-COOH can be retained in the gut of the larvae for long time [91]. Amidine- and carboxyl- PS were ingested by the three zooplankton species and accumulated mainly in the gut of D. magna and Thamnocephalus platyurus, and the stomach of Brachionus. calyciflorus [74]. The accumulation was dependent on the exposure concentration and the PS surface functionalization, however further studies are necessary to explore if NPLs could pass through the gut epithelial cells or not.
Very recently, the uptake and excretion kinetics during the direct exposure to polystyrene NPs labelled with lanthanide up-conversion luminescence (UCNPs@PS) of 49-58 nm with concentration of 500 μg L-1 have been followed in D. magna over 24 h period. A bi-phasic uptake dependence including rapid uptake by filter-feeding and quick diffusion in the intestine tract of D. magna in few minutes and deposition in the carapace and body tissue during several hours. Similarly a bi-phase excretion was also found, but UCNPs@PS were still be retained after 48 h excretion [92]. Pd-doped PS were accumulated in the digestive tract and deposited over the carapace surface forming aggregates attached to the chitin that covered the entire D. magna [55].
Novel insights have been obtained concerning the transgenerational transfer of the NPLs. A study has shown that the Eu-doped PS of 640 nm can be transferred from parents to offspring [41]. This finding is consistent with the transgenerational effects observed on D. magna indicated only negligible effects on the first generation but when neonates were exposed to same concentration of MPs they went extinct after 2 generations [93]. Parental exposure to NPLs can induce transgenerational transfer and toxicity. For example, the exposure of the parental rotifer Brachionus koreanus to fluorescently labeled nonfunctionalized 50 nm PS resulted to the transfer in offspring, and induced adverse effects on life-cycle parameters, such as development and reproduction, in offspring rotifers, and was associated with oxidative stress [94].
3.2. Advances in Uptake Availability via Trophic Transfer
Important advances and key research gaps in the understanding about the exposure, uptake and propagation of microplastics in aquatic food webs have been comprehensively discussed [95,96]. NPLs trophic transfer along the food web have also been recently reviewed [27]. Nevertheless, only few laboratory studies with the simplified food webs have clearly demonstrated that NPLs can be readily transferred along different trophic levels [40,41,97]. In a four trophic level chain experiments it was shown that 51 nm green fluorescence PS are transferred from alga C. reinhardtii through each trophic level until the fish Zacco temminckii [40]. Similarly, a study of two-trophic level and three-trophic level food chain including alga Chlorella pyrenoidosa, crustacean D. magna and fish Micropterus salmoides (algae-crustacean-fish) revealed that fluorescent PS of 80 nm accumulated in C. pyrenoidosa or D. magna and transferred to the M. salmoides, illustrating trophical amplification by the food chain. Th trophic transfer resulted in antioxidant response and histopathological damage and disturbance of lipid metabolism in M. salmoides [98]. In another study the 90-nm-sized PS-NH2 were shown to adsorb to the cell walls of microalga Dunaliella salina and to transfer to crustaceans Artemia franciscana inducing an alteration of gut permeability. In addition, PS-NPs were gradually transferred to small yellow croakers; Larimichthys polyactis through the three-level food chain inducing the inhibition of digestive enzyme activity [99].
Using Fe-PS and Eu-PS NPLs it has been shown quantitatively for a first time that these NPLs were transferred from alga P. subcapitata to D. magna and that the transfer was higher for particles with lower size [41]. This first finding suggests that NPLs of lower size could have a higher probability to travel along the food webs. Another study has highlighted the importance of the feeding strategy and algal food type (Nannochloropsis gaditana and Tetraselmis chuii) in addition to waterborne exposure to the effects of NPLs on rotifer Brachionus plicatilis exposed to PMMA [97].
3.3. Advances in "Toxico-Availability” Research
Chronic exposures of D.magna to 0.1 mg L-1 and 50 mg L-1 of 20 nm PS resulted in an impacted growth, molting and reproduction, but not the survival at both tested sizes, and at particle concentrations of 0.1 mg L-1 [100]. In the 21-day chronic toxicity test, dose- and time-dependent relationships were observed for D. pulex body length, time to clutch was delayed, and total offspring per female and number of clutches and the offspring per clutch were significantly decreased in the 0.1 mg L-1 [87]. The importance of the inter-clonal variability together with the PS size have been explored demonstrating that a twofold decrease in PS particle size from 100 to 50 nm resulted to up to 100 s-fold increase in toxicity 48hEC(10), while the inter-clonal variability of the three genotypically different clones of the D. longispina was about tenfold [101].
Further evidences from biochemical, genomic and transcriptomic studies have been obtained comforting the idea that the oxidative stress is a major toxicity mechanism of NPLs. For example, the expression of stress defense genes (SOD, GST, GPx, and CAT) was induced and then inhibited by the exposure of D. magna to 75 nm PS of 0.1.-2 mg L-1 [87]. The activities of other antioxidant enzymes (CAT, total SOD, and CuZn SOD) were significantly decreased together with the overproduction of ROS in D. pulex [102]. Exposure to 0.1 and/or 0.5 mg L-1 PS significantly increased the expressions of genes of the MAPK pathway, HIF-1 pathway, superoxide dismutase (SOD) and glutathione-S-transferase, which decreased at 2 mg L-1 PS [102] However, the protein expression ratio of ERK, JNK, AKT, HIF1α, and NFkBp65 (nuclear transcription factor-kB p65) as well as the phosphorylation of ERK and NFkBp65 were increased in a dose-dependent manner. These results were further underpinned by the RNASeq study showing that the exposure of the neonates of D. pulex to 71 nm PS induced the oxidative stress, immune suppression, and affected glycometabolism [103]. ROS overproduction and alterations in cellular membrane integrity was observed in D. magna at 80 mg L-1 Pd-doped nanoplastics but a small increase in mitochondrial membrane depolarization in the gut was observed at 10 mg L-1 [55]. 48h exposure of D. magna to secondary NPLs of PHB (< 100 nm) induced excessive ROS and severe membrane damage [57]. The age of the daphnids have been shown to alter their sensitivity to PS, as demonstrated by the expression levels of genes encoding key stress defense enzymes and proteins (SOD, CAT, GST, GPx, HSP70, and HSP90) and energy-sensing enzyme AMPK (adenosine monophosphate-activated protein kinase) in 7- and 21-day-old D. pulex [104].
Table 2.
Selected examples of the most recent studies (<5 years) researching zooplankton interactions with NPLs.
Table 2.
Selected examples of the most recent studies (<5 years) researching zooplankton interactions with NPLs.
| Species | Type of NP | Size of NP | Concentration | Duration | Path of NPLs entrance | Observed effects | Reference |
|---|---|---|---|---|---|---|---|
|
Artemia franciscana (marine) |
PS-COOH (fluorescent) | 40 nm | 0.5, 1, 1.5, 2.5, 5 mg L-1 |
14 days | waterborne | Aggregation, accumulation and excretion noticed, potential trophic transfer. | [36] |
| PS-NH2 | 190 nm | 0, 1 mg L-1 0 - 200 mg L-1 |
14 days 2 days |
waterborne and foodborne - D. salina | Found in the gut, higher levels by direct uptake than through trophic transfer, observed damage to the digestive tract, no difference in mortality and immobilization in short-term exposure. | [99] | |
| PS (amine) PS (sulfate) |
100 nm | 0, 1, 10, 100 mg L-1 |
2 days | waterborne, different levels of temperature, salinity and humic acid and bentonite |
Amine NPLs produced additional toxic effects at high salinity, while at low temperatures HA and bentonite reduced toxicity. Multi-stressor experiment showed that toxicity depends on the physico-chemical characteristics of the water. | [105] | |
|
Brachionus koreanus (freshwater) |
PS (plain) | 50 nm | 10 mg L-1 | 1 day | pre-exposed to NPLs, waterborne to POPs | Pre-exposure to NPLs leads to oxidative damage of membranes and disruption of multixenobiotic resistance (MXR) functions, NPLs subsequently enhanced the toxicity of persistent organic pollutants (POPs). | [106] |
| PS (plain and fluorescent) |
50 nm | 0, 1, 10 mg L-1 | ~1,5 days | maternal transfer to unexposed neonats | Maturation time and reproduction negatively impacted at higher concentration. Bioaccumulated maternally transfered NPLs in offspring. Parent exposures induces an increase in ROS production in offspring. | [94] | |
|
Brachionus plicatilis (freshwater) |
PMMA | 40 nm | 4.7, 9.4, 18.9, 37.5, 75.0 mg L-1 |
2 days | waterborne | Mortality increased after exposure, especially in higher concentrations. | [16] |
|
Daphnia galeata × longispina (freshwater) |
PS (fluorescent) | 100 nm | 0, 5, 20 mg L-1 | 29 days | waterborne with/without inoculated spores of parasite Metschnikowia bicuspidata | Increased number of infected hosts in the presence of NPLs, lifespan and reproduction ability are reduced. Parasite reproduction is three times lower in high NPLs concentration. NPLs have a hormetic effect on the host, increasing its fitness. | [107] |
|
Daphnia longispina (freshwater) |
PS (fluorescent) | 50 nm, 100 nm |
0, 0.01, 0.1, 1, 2, 10, 20, 100 mg L-1 |
4 days | waterborne | Smaller NPLs may be more toxic due to higher bioavailability and particle toxicity. | [101] |
|
Daphnia magna (freshwater) |
PS (plain) | 50 nm | 0.05, 0.5 mg L-1 |
21 days | waterborne | Increase in energy reserves, no changes in oxidative stress and swimming activity. | [108] |
| HDPE - (mechanically broken-down) | 90 - 200 nm | High / low mix of fractions | 98 / 134 days | waterborne and in mixture with smaller fractions | HDPE nanoplastic not toxic but the fraction of leached additives and short chain HDPE cause toxicity. | [109] | |
| PS (fluorescent) | 51 nm | 0, 20, 40, 60, 80, 100 mg L-1 | 3 days | foodborne - C. reinhardtii | Presence in the gut and damage to the intestinal walls, trophic transfer detected. | [40] | |
| PS (plain) | 100 nm | 1 mg L-1 | 2 days | waterborne | Plain PS had the highest acute toxicity and ROS production, activated MAPKs but didn't influence AChE changes, while PS-COOH, PS-n-NH2 and PS-p-NH2 activated antioxidant system and lowered ROS production. | [78] | |
| PS-p-NH2 | 50-100 nm | ||||||
| PS-COOH | 300 nm | ||||||
| PS-n-NH2 | 110 nm | ||||||
| PS-NH2 | 53 nm | 0, 0.0032, 0.032. 0.32 mg L-1 |
64.3 ± 32.5 days | waterborne | Highest concentration increased mortality, long term exposure to low concentrations leads to a decrease in survival, offspring and delay in first brood. | [77] | |
| PS-COOH | 26nm, 62 nm |
||||||
| Eu-PS NPD (NPLs debrise) Fe-PS NPD | 640 nm | 0, 1, 7 mg L-1 | 21 days | foodborne - Pseudokirchinella subcapitata | Fe-PS-NPD impacted reproduction time, increased mortality and decreased the number of neonats. Eu-PS-NPD lowered number of neonats per brood. Smaller NPD (Fe-PS-NPD) have a higher impact on the reproduction than the larger NPD (Eu-PS-NPD). | [41] | |
| PS-COOH (fluorescent) | 20 nm, 200 nm | 0, 0.1, 50 mg L-1 |
21 days | waterborne | Molting and time to first brood prolonged, changes in the body length, neonat production in 200 nm may be higher because of hormesis. | [100] | |
| PS (fluorescent) | 80 nm | 0, 5 mg L-1 | 28 days | foodborne - Chlorella pyrenoidosa | Trophic transfer observed, higher accumulation through direct exposure than foodborne. Histopathological damages in the intestinal. | [98] | |
| Amidine PS | 20, 40, 60, and 100 nm | 0.5 to 30 mg L-1 (0.5 to 100 mg L-1 for 100nm NPLs) | 2 days | waterborne | Exposure in lake water. The effect depended on the primary size of PS, with 20 and 40-nm-size PS NPLs inducing a stronger effect. | [75] | |
|
Daphnia pulex (freshwater) |
PS (fluorescent) | 75 nm | 0, 0.1, 0.5, 1 and 2 mg L-1 |
21 days | waterborne | Growth inhibition, reproduction time longer while number of neonats reduced, heat shot proteins (HSP70 and HSP90) increased in the higher concentrations. | [87] |
| PS (plain) | 75 nm | 0, 0.1, 0.5, 1, 2 mg L-1 |
21 days | waterborne | Increase in concentration of NPLs stimulates increase in ROS production which leads to increase in antioxidative gene expression and enzyme activity, possible negative effects on cell survival and proliferation via MAPK pathways. | [102] | |
| PS (plain) | 71.18 ± 6.03 nm | 0, 1 mg L-1 | 4 days | waterborne | 208 differentially expressed genes analysed - changes in the expression for oxidative stress, immune defense and glycometabolism pathways. | [103] | |
|
Daphnia magna ,larvae Thamnocephalus platyurus, and rotifer Brachionus calyciflorus (freshwater) |
Amidine PS | 226.0 ± 8.6 nm |
0 to 400 mg L−1 | 1 day and 2 days | waterborne | The toxicity decreased in the order D. magna (48 h -immobilization) > B. calyciflorus (24 h - lethality) > T. platyurus (24 h - lethality). Amidine PS were more toxic than carboxyl PS. Alginate and humic acid formed eco-corona on amidine PS nanospheres and reduced toxicity to zooplankton. | [74] |
| Carboxyl PS | 220.1 ± 9.1 nm |
0 to 400 mgL-1 |
Multi-generational effects of low concentrations of NPL have been also observed. For example, D. pulex reproduction was affected in offspring from exposed parents. Exposure of F0 and F1 generations of D. pulex to 75 nm PS at 1 μg L-1 resulted in significant increase in the expression of antioxidant genes coding for Mn SOD, CuZn SOD, GCL, HO1, CYP4C33, and CYP4C34 and the enzyme activity of GST, CAT, but were inhibited in F2 generation. By contrast, the AMPK was further increased in F2 generation[110]. The exposure of D. magna to Pd-doped 200 nm PS, resulted in no impact at 0.1 mg L-1. However, 1 mg L-1 significantly increased the fertility in the F3 generation and decreased the size and lipid content in F3 offspring [111]. The authors have pointed out that the NPLs “effects to D. magna adults and offspring began only after multigenerational exposure despite similar body burden values between the adults and offspring of different generations”[111].
3.4. Effect of Zooplankton on NPLs Biouptake and Toxico Availability
Zooplankton species, for example D. magna release biological material into their surroundings, including kairomones, enzymes and proteins from expelled gut, chitin-based carbohydrate from molting, digestive enzymes and undigested or partially digested matter [76,112,113]. The possible consequences of the formed eco-corona as a modulator of the bioavailability and toxicity of NPLs to D. magna have been thoroughly reviewed [112] and include: (i) altering the NPLs stability and uptake; (ii) influencing residency time and absorption of nutrients; (iii) influencing biomolecules released by gut bacteria; (iv) affecting signaling molecules by binding to key signaling molecules such as kairomones. These findings confirm the importance of eco-corona formation in modulating NPLs uptake and effect, as well as the necessity to perform the bioassays under more environmentally relevant conditions for example with NPLs coated by EPS.
4. Conclusions and Perspectives
The increasing evidences demonstrate that the NPLs are bioavailable and can cause harm to the planktonic organisms if present at concentrations much higher than the expected one in the aquatic environment. Recent advances in toxico-availability from biochemical, genomic and transcriptomic studies with model NPls and plankton organisms demonstrated that high concentrations of NPLs could induce oxidative stress and damage, DNA damage and depolarization of mitochondrial and cell membrane in various plankton species. Alteration of the photosynthetic activity in phytoplankton species has been also found as a major response to NPls induced stress. Multi-generational effects of low concentrations of NPLs have been also observed in zooplankton.
Despite of these recent advances there is a controversy in the discussion concerning the toxicity of the NPLs in aquatic environment. Therefore, the assessment of the biological responses of more environmentally relevant materials, such as secondary NPLs and aged NPLs at concentrations which are closer to those expected to the aquatic environment could provide further insights on the responses in reals environment. On the other hand, the lack of reference materials, standardized testing procedures limit the comparability of the results and repeatability of the bioassays used to dose-response relationships obtained in different laboratories. Very recently a procedure short- and long-term toxicity tests for aquatic organisms together with the top-down production of more realistic NPLs and their characterization were proposed [114].
The most of the toxicity data are obtained with commercially available NPLs, mainly PS, and should be considered with cation since (i) they do not represent the diversity of the NPLs (in size, shapes and composition)[15] and (ii) some preservative present in these products could affect the toxicity. For example, PS used as purchased were toxic to Vibrio fischeri, Raphidocelis subcapitata, Daphnia magna (EC50 ranging from 13 to 71 mg L-1), however, after sample dialysis neither NPLs induced toxicity (L(E)C50 > 100 mg L-1)[115]. Therefore, the assessment of the responses of planktonic species to NPLs under environmentally realistic conditions is an important research priority. Indeed, most of the studies are using virgin and/or surface-modified NPLs in the same shape, while in the aquatic environment we are finding a mix of different types, sizes and shapes of polymers with diverse additives added to improve their performances. In addition, when assessing the effects of NPLs the presence of other environmental pollutants which can adsorb to NPLs and possible synergistic, additive of antagonistic responses in aquatic plankton need to consider [116].
Regardless of the existing progress, the understanding of the accumulation patterns and translocation mechanisms of NPLs within planktonic organisms remains incomplete. The bioaccumulation and trophic transfer factors, two key ecotoxicological parameters, needed to accurately assess the potential impacts of NPLs on aquatic organisms are not yet quantified. Since many food webs are based on phytoplankton species, the association of the NPLs on the cell surface or their cell penetration by endocytosis will inevitably introduce them into the food webs. However, most of the existing papers consider the toxicity of NPLs exposure to individual zooplankton species rarely considered the trophic transfer at the bottom of the food webs as well as between the waterborne and foodborne exposures. Further studies are thus necessary to address the uptake and clearance rate and mechanisms and to distinguish the contribution of the waterborne and foodborne exposure to NPLs accumulation in freshwater zooplankton. It is worth noting that such studies are quite challenging due to the limitations of the current analytical techniques in terms of detection and quantification of NPLs in complex biological samples.
Most of the current knowledge concerning the uptake availability and toxico-availability of NPLs originate from studies performed with model NPLs, mainly PS nanospheres, which are different and cannot be expected to behave in a similar manner to secondary, heterogeneous NPLs [117,118]. Incidentally produced NPLs exhibit a diversity of compositions and morphologies and a heterogeneity that is typically absent from model NPLs and engineered nanomaterials [7]. Due to the challenge of quantifying NPLs in complex environmental and biological samples [119], bioaccumulation and trophic transfer of NPLs was only qualitatively observed with fluorescent-labelled NPLs or metal-doped particles [120]. Indeed, even if they are not representative of the diversity of incidental NPLs in the environment, their use will provide a critical new information about the interactions of NPLs with phyto- and zooplankton and possible trophic transfer. Bioassays with the secondary NPLs will provide an insight into effects inflicted by naturally fragmentated and aged NPLs from the environment. Their potentially higher toxicity, because of the contaminants from the environment, eco-corona formation and acute exposure of organisms, must be assessed.
Furthermore, aquatic plankton is expected to contribute to the NPLs transformations. For example, different marine diatoms and cyanobacteria have been shown to biodegrade various plastic materials via fouling, corrosion, hydrolysis and penetration, degradation of leaching components as well as pigment coloration via diffusion into the polymers [121,122]. However, it is unclear, if this is more general phenomena and if it is of relevance to the natural environment. The plankton species can affect the NPLs, by release of the biomolecules capable to could modify the surface of the NPLs; affect their stability and thus need to be considered for improved understanding their interaction with aquatic organisms. Despite the advances concerning marine phytoplankton EPS, no similar studies deal with EPS released by freshwater phytoplankton have been yet published. Very few studies are available for the zooplankton species. Given the existing knowledge about the role of freshwater EPS in the fate and transformations of metal-containing nanoparticles [123], it is expected that they affect significantly the bioreactivity of NPLs. Indeed, the need to incorporate the concept of biomolecular corona within more general framework considering the interactions and feedback of phyto-and zooplankton to nanoparticles exposure was highlighted [69]
The trophic transfer of NPLs in aquatic food chains appears possible as demonstrated by the study with model NPLs and artificial two- to four-level trophic chains. However, it is essential to confirm this trophic transfer through comprehensive in situ surveys of trophic chains. Additionally, we must carefully evaluate the transport and destiny of biologically ingested plastics within the food chain. In this relation, the detection, quantification and characterization of key characteristics of the secondary NPLs, as generated in the environment, represent a cornerstone for understanding their fate and impact, and for enabling scientifically sound and quantitative risk assessment, environmental quality standard definition, monitoring and management.
Author Contributions
VIS conceived the concept, participated in the manuscript writing and overviewed all the work; MM participated in manuscript writing and made conceptual Figure 1 and Table 1 and Table 2. All the authors critically commented and revised the manuscript. All the authors have approved the paper submission.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thompson, R.C.; Moore, C.J.; vom Saal, F.S.; Swan, S.H. Plastics, the environment and human health: Current consensus and future trends. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364, 2153–2166. [Google Scholar] [CrossRef]
- Zhang, B.; Chao, J.Y.; Chen, L.; Liu, L.C.; Yang, X.; Wang, Q. Research progress of nanoplastics in freshwater. Sci. Total Environ. 2021, 757, 12. [Google Scholar] [CrossRef]
- Alimi, O.S.; Farner Budarz, J.; Hernandez, L.M.; Tufenkji, N. Microplastics and nanoplastics in aquatic environments: Aggregation, deposition, and enhanced contaminant transport. Environmental Science & Technology 2018, 52, 1704–1724. [Google Scholar] [CrossRef]
- Kukkola, A.; Krause, S.; Lynch, I.; Sambrook Smith, G.H.; Nel, H. Nano and microplastic interactions with freshwater biota – current knowledge, challenges and future solutions. Environment International 2021, 152, 106504. [Google Scholar] [CrossRef]
- Gaylarde, C.C.; Baptista Neto, J.A.; da Fonseca, E.M. Nanoplastics in aquatic systems - are they more hazardous than microplastics? Environmental Pollution 2021, 272, 115950. [Google Scholar] [CrossRef] [PubMed]
- Atugoda, T.; Piyumali, H.; Wijesekara, H.; Sonne, C.; Lam, S.S.; Mahatantila, K.; Vithanage, M. Nanoplastic occurrence, transformation and toxicity: A review. Environmental Chemistry Letters 2023, 21, 363–381. [Google Scholar] [CrossRef]
- Gigault, J.; El Hadri, H.; Nguyen, B.; Grassl, B.; Rowenczyk, L.; Tufenkji, N.; Feng, S.; Wiesner, M. Nanoplastics are neither microplastics nor engineered nanoparticles. Nature Nanotechnology 2021, 16, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.; An, Y.J. Effects of micro- and nanoplastics on aquatic ecosystems: Current research trends and perspectives. Marine Pollution Bulletin 2017, 124, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Fan, X.; Zou, H.; Guo, R.-B.; Fu, S.-F. Effects of size and surface charge on the sedimentation of nanoplastics in freshwater. Chemosphere 2023, 336, 139194. [Google Scholar] [CrossRef] [PubMed]
- Commission, E. Chemicals strategy for sustainability. Commission, E., Ed. 2022.
- Gigault, J.; Halle, A.t.; Baudrimont, M.; Pascal, P.-Y.; Gauffre, F.; Phi, T.-L.; El Hadri, H.; Grassl, B.; Reynaud, S. Current opinion: What is a nanoplastic? Environmental Pollution 2018, 235, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, M.; Lebreton, L.; Carson, H.; Thiel, M.; Moore, C.; Borerro, J.; Galgani, F.; Ryan, P.; Reisser, J. Plastic pollution in the world’s oceans: More than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Besseling, E.; Redondo-Hasselerharm, P.; Foekema, E.M.; Koelmans, A.A. Quantifying ecological risks of aquatic micro- and nanoplastic. Critical Reviews in Environmental Science and Technology 2019, 49, 32–80. [Google Scholar] [CrossRef]
- Shi, C.; Liu, Z.; Yu, B.; Zhang, Y.; Yang, H.; Han, Y.; Wang, B.; Liu, Z.; Zhang, H. Emergence of nanoplastics in the aquatic environment and possible impacts on aquatic organisms. Sci. Total Environ. 2024, 906, 167404. [Google Scholar] [CrossRef]
- Yang, T.; Nowack, B. A meta-analysis of ecotoxicological hazard data for nanoplastics in marine and freshwater systems. Environmental Toxicology and Chemistry 2020, 39, 2588–2598. [Google Scholar] [CrossRef]
- Venâncio, C.; Ferreira, I.; Martins, M.A.; Soares, A.M.V.M.; Lopes, I.; Oliveira, M. The effects of nanoplastics on marine plankton: A case study with polymethylmethacrylate. Ecotoxicology and Environmental Safety 2019, 184, 109632. [Google Scholar] [CrossRef]
- Takeshita, K.M.; Iwasaki, Y.; Sinclair, T.M.; Hayashi, T.I.; Naito, W. Illustrating a species sensitivity distribution for nano- and microplastic particles using bayesian hierarchical modeling. Environmental Toxicology and Chemistry 2022, 41, 954–960. [Google Scholar] [CrossRef]
- Cunningham, B.E.; Sharpe, E.E.; Brander, S.M.; Landis, W.G.; Harper, S.L. Critical gaps in nanoplastics research and their connection to risk assessment. Frontiers in Toxicology 2023, 5. [Google Scholar] [CrossRef]
- Abdolahpur Monikh, F.; Vijver, M.G.; Kortet, R.; Lynch, I.; Peijnenburg, W. Emerging investigator series: Perspectives on toxicokinetics of nanoscale plastic debris in organisms. Environmental Science: Nano 2022. [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Research 2015, 75, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Bakir, A.; Burton, G.A.; Janssen, C.R. Microplastic as a vector for chemicals in the aquatic environment: Critical review and model-supported reinterpretation of empirical studies. Environmental Science & Technology 2016, 50, 3315–3326. [Google Scholar] [CrossRef]
- Kogel, T.; Bjoroy, O.; Toto, B.; Bienfait, A.M.; Sanden, M. Micro- and nanoplastic toxicity on aquatic life: Determining factors. Sci. Total Environ. 2020, 709. [Google Scholar] [CrossRef]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.-J. Microplastics in aquatic environments: Occurrence, accumulation, and biological effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef]
- Huang, D.; Tao, J.; Cheng, M.; Deng, R.; Chen, S.; Yin, L.; Li, R. Microplastics and nanoplastics in the environment: Macroscopic transport and effects on creatures. Journal of Hazardous Materials 2021, 407, 124399. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.R.; Lian, F.; Xiao, Z.G.; Gu, S.G.; Cao, X.S.; Wang, Z.Y.; Xing, B.S. Potential toxicity of nanoplastics to fish and aquatic invertebrates: Current understanding, mechanistic interpretation, and meta-analysis. Journal of Hazardous Materials 2022, 427. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhang, Y.; Zhu, Y.; Song, B.; Zeng, G.; Hu, D.; Wen, X.; Ren, X. Recent advances in toxicological research of nanoplastics in the environment: A review. Environmental Pollution 2019, 252, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Fu, S.F.; Zou, H.; Su, Y.Y.; Zhang, Y.F. Effects of nanoplastics on microalgae and their trophic transfer along the food chain: Recent advances and perspectives. Environmental Science-Processes & Impacts 2021, 23, 1873–1883. [Google Scholar] [CrossRef]
- Larue, C.; Sarret, G.; Castillo-Michel, H.; del Real, A.E.P. A critical review on the impacts of nanoplastics and microplastics on aquatic and terrestrial photosynthetic organisms. Small 2021, 17. [Google Scholar] [CrossRef]
- Nguyen, M.-K.; Lin, C.; Nguyen, H.-L.; Le, V.-G.; Haddout, S.; Um, M.-J.; Chang, S.W.; Nguyen, D.D. Ecotoxicity of micro- and nanoplastics on aquatic algae: Facts, challenges, and future opportunities. Journal of Environmental Management 2023, 346, 118982. [Google Scholar] [CrossRef]
- Gong, H.; Li, R.X.; Li, F.; Guo, X.W.; Xu, L.J.; Gan, L.; Yan, M.T.; Wang, J. Toxicity of nanoplastics to aquatic organisms: Genotoxicity, cytotoxicity, individual level and beyond individual level. Journal of Hazardous Materials 2023, 443. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Iavicoli, I.; Barcelo, D.; Calabrese, E.J. Micro/nanoplastics effects on organisms: A review focusing on 'dose'. Journal of Hazardous Materials 2021, 417. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, M.; Duffus, J.H.; Templeton, D.M. Explanatory dictionary of key terms in toxicology: Part ii (iupac recommendations 2010). Pure and Applied Chemistry 2010, 82, 679–751. [Google Scholar] [CrossRef]
- Slaveykova, V.I.; Li, M.; Worms, I.A.; Liu, W. When environmental chemistry meets ecotoxicology: Bioavailability of inorganic nanoparticles to phytoplankton. CHIMIA 2020, 74, 115. [Google Scholar] [CrossRef]
- Vijver, M.G.; Zhai, Y.; Wang, Z.; Peijnenburg, W.J.G.M. Emerging investigator series: The dynamics of particle size distributions need to be accounted for in bioavailability modelling of nanoparticles. Environmental Science: Nano 2018, 5, 2473–2481. [Google Scholar] [CrossRef]
- Karpowicz, M.; Zieliński, P.; Grabowska, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I. Effect of eutrophication and humification on nutrient cycles and transfer efficiency of matter in freshwater food webs. Hydrobiologia 2020, 847, 2521–2540. [Google Scholar] [CrossRef]
- Bergami, E.; Pugnalini, S.; Vannuccini, M.L.; Manfra, L.; Faleri, C.; Savorelli, F.; Dawson, K.A.; Corsi, I. Long-term toxicity of surface-charged polystyrene nanoplastics to marine planktonic species dunaliella tertiolecta and artemia franciscana. Aquatic Toxicology 2017, 189, 159–169. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Toullec, J.; Lambert, C.; Le Goïc, N.; Seoane, M.; Moriceau, B.; Huvet, A.; Berchel, M.; Vincent, D.; Courcot, L. , et al. Do transparent exopolymeric particles (tep) affect the toxicity of nanoplastics on chaetoceros neogracile? Environmental Pollution 2019, 250, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Nolte, T.M.; Hartmann, N.B.; Kleijn, J.M.; Garnæs, J.; van de Meent, D.; Jan Hendriks, A.; Baun, A. The toxicity of plastic nanoparticles to green algae as influenced by surface modification, medium hardness and cellular adsorption. Aquatic Toxicology 2017, 183, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sendra, M.; Staffieri, E.; Yeste, M.P.; Moreno-Garrido, I.; Gatica, J.M.; Corsi, I.; Blasco, J. Are the primary characteristics of polystyrene nanoplastics responsible for toxicity and ad/absorption in the marine diatom phaeodactylum tricornutum? Environmental Pollution 2019, 249, 610–619. [Google Scholar] [CrossRef]
- Chae, Y.; Kim, D.; Kim, S.W.; An, Y.J. Trophic transfer and individual impact of nano-sized polystyrene in a four-species freshwater food chain. Scientific Reports 2018, 8. [Google Scholar] [CrossRef]
- Monikh, F.A.; Chupani, L.; Vijver, M.G.; Peijnenburg, W. Parental and trophic transfer of nanoscale plastic debris in an assembled aquatic food chain as a function of particle size. Environmental Pollution 2021, 269. [Google Scholar] [CrossRef]
- Yan, N.; Tang, B.Z.; Wang, W.X. Cell cycle control of nanoplastics internalization in phytoplankton. Acs Nano 2021, 15, 12237–12248. [Google Scholar] [CrossRef]
- Zhao, Y.; Tao, S.; Liu, S.; Hu, T.; Zheng, K.; Shen, M.; Meng, G. Research advances on impacts micro/nanoplastics and their carried pollutants on algae in aquatic ecosystems: A review. Aquat Toxicol 2023, 264, 106725. [Google Scholar] [CrossRef]
- Corsi, I.; Bellingeri, A.; Eliso, M.C.; Grassi, G.; Liberatori, G.; Murano, C.; Sturba, L.; Vannuccini, M.L.; Bergami, E. Eco-interactions of engineered nanomaterials in the marine environment: Towards an eco-design framework. Nanomaterials 2021, 11, 1903. [Google Scholar] [CrossRef]
- Wang, S.; Liu, M.; Wang, J.; Huang, J.; Wang, J. Polystyrene nanoplastics cause growth inhibition, morphological damage and physiological disturbance in the marine microalga platymonas helgolandica. Marine Pollution Bulletin 2020, 158, 111403. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-r.; Wang, B.-l.; Nan, F.-r.; Lv, J.-p.; Liu, X.-d.; Liu, Q.; Feng, J.; Xie, S.-l. Effects of polystyrene nanoplastics on the physiological and biochemical characteristics of microalga scenedesmus quadricauda. Environmental Pollution 2023, 319, 120987. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Ai, H.; Chen, Y.; Zhang, Z.; Zeng, P.; Kang, L.; Li, W.; Gu, W.; He, Q.; Li, H. Phytoplankton response to polystyrene microplastics: Perspective from an entire growth period. Chemosphere 2018, 208, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gao, P.; Li, H.; Huang, J.; Zhang, Y.; Ding, H.; Zhang, W. Mechanism of the inhibition and detoxification effects of the interaction between nanoplastics and microalgae chlorella pyrenoidosa. Sci. Total Environ. 2021, 783, 146919. [Google Scholar] [CrossRef] [PubMed]
- Hazeem, L.J.; Yesilay, G.; Bououdina, M.; Perna, S.; Cetin, D.; Suludere, Z.; Barras, A.; Boukherroub, R. Investigation of the toxic effects of different polystyrene micro-and nanoplastics on microalgae chlorella vulgaris by analysis of cell viability, pigment content, oxidative stress and ultrastructural changes. Marine Pollution Bulletin 2020, 156, 111278. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, X.; Liao, Y.; Zhao, W.; Zhao, P.; Li, M. Adverse physiological and molecular level effects of polystyrene microplastics on freshwater microalgae. Chemosphere 2020, 255, 126914. [Google Scholar] [CrossRef]
- Liao, Y.; Jiang, X.; Xiao, Y.; Li, M. Exposure of microalgae euglena gracilis to polystyrene microbeads and cadmium: Perspective from the physiological and transcriptional responses. Aquatic Toxicology 2020, 228, 105650. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-J.; Li, J.-W.; Xu, E.G.; Sun, X.-D.; Zhu, F.-P.; Ding, Z.; Tian, H.; Dong, S.-S.; Xia, P.-F.; Yuan, X.-Z. Short-term exposure to positively charged polystyrene nanoparticles causes oxidative stress and membrane destruction in cyanobacteria. Environmental Science: Nano 2019, 6, 3072–3079. [Google Scholar] [CrossRef]
- Seoane, M.; González-Fernández, C.; Soudant, P.; Huvet, A.; Esperanza, M.; Cid, Á.; Paul-Pont, I. Polystyrene microbeads modulate the energy metabolism of the marine diatom chaetoceros neogracile. Environmental Pollution 2019, 251, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, P.; Zhang, C.; Zhou, X.; Yin, Z.; Hu, T.; Hu, D.; Liu, C.; Zhu, L. Influence of polystyrene microplastics on the growth, photosynthetic efficiency and aggregation of freshwater microalgae chlamydomonas reinhardtii. Sci. Total Environ. 2020, 714, 136767. [Google Scholar] [CrossRef] [PubMed]
- Tamayo-Belda, M.; Pérez-Olivares, A.V.; Pulido-Reyes, G.; Martin-Betancor, K.; González-Pleiter, M.; Leganés, F.; Mitrano, D.M.; Rosal, R.; Fernández-Piñas, F. Tracking nanoplastics in freshwater microcosms and their impacts to aquatic organisms. Journal of Hazardous Materials 2023, 445, 130625. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tang, X.X.; Zhang, B.H.; Li, L.Y.; Zhao, Y.R.; Lv, M.C.; Li, J.; Kan, C.X.; Zhao, Y. The effects of two sized polystyrene nanoplastics on the growth, physiological functions, and toxin production of alexandrium tamarense. Chemosphere 2022, 291. [Google Scholar] [CrossRef] [PubMed]
- González-Pleiter, M.; Tamayo-Belda, M.; Pulido-Reyes, G.; Amariei, G.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Secondary nanoplastics released from a biodegradable microplastic severely impact freshwater environments. Environmental Science: Nano 2019, 6, 1382–1392. [Google Scholar] [CrossRef]
- Natarajan, L.; Omer, S.; Jetly, N.; Jenifer, M.A.; Chandrasekaran, N.; Suraishkumar, G.K.; Mukherjee, A. Eco-corona formation lessens the toxic effects of polystyrene nanoplastics towards marine microalgae chlorella sp. Environmental Research 2020, 188, 109842. [Google Scholar] [CrossRef] [PubMed]
- Cesarini, G.; Secco, S.; Taurozzi, D.; Venditti, I.; Battocchio, C.; Marcheggiani, S.; Mancini, L.; Fratoddi, I.; Scalici, M.; Puccinelli, C. Teratogenic effects of environmental concentration of plastic particles on freshwater organisms. Sci. Total Environ. 2023, 898, 165564. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yuan, Y.; Li, Y.; Liu, X.; Wang, X.; Fan, Z. Polystyrene nanoplastics affect growth and microcystin production of microcystis aeruginosa. Environmental Science and Pollution Research 2021, 28, 13394–13403. [Google Scholar] [CrossRef]
- Grassi, G.; Gabellieri, E.; Cioni, P.; Paccagnini, E.; Faleri, C.; Lupetti, P.; Corsi, I.; Morelli, E. Interplay between extracellular polymeric substances (eps) from a marine diatom and model nanoplastic through eco-corona formation. Sci. Total Environ. 2020, 725, 138457. [Google Scholar] [CrossRef]
- Gomes, T.; Almeida, A.C.; Georgantzopoulou, A. Characterization of cell responses in rhodomonas baltica exposed to pmma nanoplastics. Sci. Total Environ. 2020, 726, 138547. [Google Scholar] [CrossRef]
- Baudrimont, M.; Arini, A.; Guégan, C.; Venel, Z.; Gigault, J.; Pedrono, B.; Prunier, J.; Maurice, L.; Ter Halle, A.; Feurtet-Mazel, A. Ecotoxicity of polyethylene nanoplastics from the north atlantic oceanic gyre on freshwater and marine organisms (microalgae and filter-feeding bivalves). Environmental Science and Pollution Research 2020, 27, 3746–3755. [Google Scholar] [CrossRef]
- Tamayo-Belda, M.; Vargas-Guerrero, J.J.; Martín-Betancor, K.; Pulido-Reyes, G.; González-Pleiter, M.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Understanding nanoplastic toxicity and their interaction with engineered cationic nanopolymers in microalgae by physiological and proteomic approaches. Environmental Science: Nano 2021, 8, 2277–2296. [Google Scholar] [CrossRef]
- Tamayo-Belda, M.; Pulido-Reyes, G.; González-Pleiter, M.; Martín-Betancor, K.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Identification and toxicity towards aquatic primary producers of the smallest fractions released from hydrolytic degradation of polycaprolactone microplastics. Chemosphere 2022, 303, 134966. [Google Scholar] [CrossRef] [PubMed]
- Shiu, R.-F.; Vazquez, C.I.; Chiang, C.-Y.; Chiu, M.-H.; Chen, C.-S.; Ni, C.-W.; Gong, G.-C.; Quigg, A.; Santschi, P.H.; Chin, W.-C. Nano- and microplastics trigger secretion of protein-rich extracellular polymeric substances from phytoplankton. Sci. Total Environ. 2020, 748, 141469. [Google Scholar] [CrossRef]
- Mitrano, D.M.; Wick, P.; Nowack, B. Placing nanoplastics in the context of global plastic pollution. Nature Nanotechnology 2021, 16, 491–500. [Google Scholar] [CrossRef]
- Junaid, M.; Wang, J. Interaction of nanoplastics with extracellular polymeric substances (eps) in the aquatic environment: A special reference to eco-corona formation and associated impacts. Water Research 2021, 201. [Google Scholar] [CrossRef]
- Liu, W.; Worms, I.A.M.; Jakšić, Ž.; Slaveykova, V.I. Aquatic organisms modulate the bioreactivity of engineered nanoparticles: Focus on biomolecular corona. Frontiers in Toxicology 2022, 4. [Google Scholar] [CrossRef]
- Shou, W.; Kang, F.; Lu, J. Nature and value of freely dissolved eps ecosystem services: Insight into molecular coupling mechanisms for regulating metal toxicity. Environmental Science & Technology 2018, 52, 457–466. [Google Scholar] [CrossRef]
- Wilkinson, K.J.; Joz-Roland, A.; Buffle, J. Different roles of pedogenic fulvic acids and aquagenic biopolymers on colloid aggregation and stability in freshwaters. Limnology and Oceanography 1997, 42, 1714–1724. [Google Scholar] [CrossRef]
- Naveed, S.; Li, C.H.; Lu, X.D.; Chen, S.S.; Yin, B.; Zhang, C.H.; Ge, Y. Microalgal extracellular polymeric substances and their interactions with metal(loid)s: A review. Critical Reviews in Environmental Science and Technology 2019, 49, 1769–1802. [Google Scholar] [CrossRef]
- Chen, C.-S.; Anaya, J.M.; Zhang, S.; Spurgin, J.; Chuang, C.-Y.; Xu, C.; Miao, A.-J.; Chen, E.Y.; Schwehr, K.A.; Jiang, Y. Effects of engineered nanoparticles on the assembly of exopolymeric substances from phytoplankton. PLoS One 2011, 6, e21865. [Google Scholar] [CrossRef]
- Saavedra, J.; Stoll, S.; Slaveykova, V.I. Influence of nanoplastic surface charge on eco-corona formation, aggregation and toxicity to freshwater zooplankton. Environmental Pollution 2019, 252, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Pochelon, A.; Stoll, S.; Slaveykova, V.I. Polystyrene nanoplastic behavior and toxicity on crustacean daphnia magna: Media composition, size, and surface charge effects. Environments 2021, 8, 101. [Google Scholar] [CrossRef]
- Fadare, O.O.; Wan, B.; Liu, K.; Yang, Y.; Zhao, L.; Guo, L.-H. Eco-corona vs protein corona: Effects of humic substances on corona formation and nanoplastic particle toxicity in daphnia magna. Environmental Science & Technology 2020, 54, 8001–8009. [Google Scholar] [CrossRef]
- Kelpsiene, E.; Torstensson, O.; Ekvall, M.T.; Hansson, L.-A.; Cedervall, T. Long-term exposure to nanoplastics reduces life-time in daphnia magna. Scientific Reports 2020, 10, 5979. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Jiang, R.; Hu, S.; Xiao, X.; Wu, J.; Wei, S.; Xiong, Y.; Ouyang, G. Investigating the toxicities of different functionalized polystyrene nanoplastics on daphnia magna. Ecotoxicology and Environmental Safety 2019, 180, 509–516. [Google Scholar] [CrossRef]
- Allen, S.; Allen, D.; Karbalaei, S.; Maselli, V.; Walker, T.R. Micro(nano)plastics sources, fate, and effects: What we know after ten years of research. Journal of Hazardous Materials Advances 2022, 6, 100057. [Google Scholar] [CrossRef]
- Latchere, O.; Audroin, T.; Hetier, J.; Metais, I.; Chatel, A. The need to investigate continuums of plastic particle diversity, brackish environments and trophic transfer to assess the risk of micro and nanoplastics on aquatic organisms. Environmental Pollution 2021, 273. [Google Scholar] [CrossRef] [PubMed]
- Schröter, L.; Ventura, N. Nanoplastic toxicity: Insights and challenges from experimental model systems. Small 2022, 18, 2201680. [Google Scholar] [CrossRef]
- Jemec Kokalj, A.; Heinlaan, M.; Novak, S.; Drobne, D.; Kühnel, D. Defining quality criteria for nanoplastic hazard evaluation: The case of polystyrene nanoplastics and aquatic invertebrate daphnia spp. Nanomaterials 2023, 13, 536. [Google Scholar] [CrossRef]
- Seda, J.; Petrusek, A. Daphnia as a model organism in limnology and aquatic biology: Introductory remarks. Journal of Limnology 2011, 70, 337–344. [Google Scholar] [CrossRef]
- Brehm, J.; Ritschar, S.; Laforsch, C.; Mair, M.M. The complexity of micro- and nanoplastic research in the genus daphnia – a systematic review of study variability and a meta-analysis of immobilization rates. Journal of Hazardous Materials 2023, 458, 131839. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Nowack, B. Meta-analysis of bioaccumulation data for nondissolvable engineered nanomaterials in freshwater aquatic organisms. Environmental Toxicology and Chemistry 2022, 41, 1202–1214. [Google Scholar] [CrossRef]
- Rist, S.; Baun, A.; Hartmann, N.B. Ingestion of micro- and nanoplastics in daphnia magna – quantification of body burdens and assessment of feeding rates and reproduction. Environmental Pollution 2017, 228, 398–407. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, P.; Cai, M.; Wu, D.; Zhang, M.; Huang, Y.; Zhao, Y. Polystyrene nanoplastic exposure induces immobilization, reproduction, and stress defense in the freshwater cladoceran daphnia pulex. Chemosphere 2019, 215, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, D.S.; Nogueira, D.J.; Melegari, S.P.; Arl, M.; Köerich, J.S.; Cruz, L.; Justino, N.M.; Oscar, B.V.; Puerari, R.C.; da Silva, M.L.N. , et al. Toxicological evaluation and quantification of ingested metal-core nanoplastic by daphnia magna through fluorescence and inductively coupled plasma-mass spectrometric methods. Environmental Toxicology and Chemistry 2019, 38, 2101–2110. [Google Scholar] [CrossRef]
- Wang, M.J.; Wang, W.X. Accumulation kinetics and gut microenvironment responses to environmentally relevant doses of micro/nanoplastics by zooplankton daphnia magna. Environmental Science & Technology 2023, 57, 5611–5620. [Google Scholar] [CrossRef]
- Wang, M.J.; Wang, W.X. Selective ingestion and response by daphnia magna to environmental challenges of microplastics. Journal of Hazardous Materials 2023, 458. [Google Scholar] [CrossRef]
- Manfra, L.; Rotini, A.; Bergami, E.; Grassi, G.; Faleri, C.; Corsi, I. Comparative ecotoxicity of polystyrene nanoparticles in natural seawater and reconstituted seawater using the rotifer brachionus plicatilis. Ecotoxicology and Environmental Safety 2017, 145, 557–563. [Google Scholar] [CrossRef]
- Zhang, Y.-H.; Gao, R.-Y.; Wang, Z.-J.; Shao, Q.-Q.; Hu, Y.-W.; Jia, H.-B.; Liu, X.-J.; Dong, F.-Q.; Fu, L.-M.; Zhang, J.-P. Daphnia magna uptake and excretion of luminescence-labelled polystyrene nanoparticle as visualized by high sensitivity real-time optical imaging. Chemosphere 2023, 326, 138341. [Google Scholar] [CrossRef]
- Martins, A.; Guilhermino, L. Transgenerational effects and recovery of microplastics exposure in model populations of the freshwater cladoceran daphnia magna straus. Sci. Total Environ. 2018, 631-632, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Yeo, I.-C.; Shim, K.-Y.; Kim, K.; Jeong, C.-B. Maternal exposure to nanoplastic induces transgenerational toxicity in the offspring of rotifer brachionus koreanus. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2023, 269, 109635. [Google Scholar] [CrossRef]
- Krause, S.; Baranov, V.; Nel, H.A.; Drummond, J.D.; Kukkola, A.; Hoellein, T.; Sambrook Smith, G.H.; Lewandowski, J.; Bonet, B.; Packman, A.I. , et al. Gathering at the top? Environmental controls of microplastic uptake and biomagnification in freshwater food webs. Environmental Pollution 2021, 268, 115750. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Chen, H.; Shen, M.; Tao, J.; Chen, S.; Yin, L.; Zhou, W.; Wang, X.; Xiao, R.; Li, R. Recent advances on the transport of microplastics/nanoplastics in abiotic and biotic compartments. Journal of Hazardous Materials 2022, 438, 129515. [Google Scholar] [CrossRef]
- Venancio, C.; Ciubotariu, A.; Lopes, I.; Martins, M.A.; Oliveira, M. Is the toxicity of nanosized polymethylmethacrylate particles dependent on the exposure route and food items? Journal of Hazardous Materials 2021, 413. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Gao, D.; Kong, C.; Junaid, M.; Li, Y.; Chen, X.; Zheng, Q.; Chen, G.; Wang, J. Trophic transfer of nanoplastics and di(2-ethylhexyl) phthalate in a freshwater food chain (chlorella pyrenoidosa-daphnia magna-micropterus salmoides) induced disturbance of lipid metabolism in fish. Journal of Hazardous Materials 2023, 459, 132294. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Cui, R.; Il Kwak, J.; An, Y.-J. Trophic transfer of nanoplastics through a microalgae–crustacean–small yellow croaker food chain: Inhibition of digestive enzyme activity in fish. Journal of Hazardous Materials 2022, 440, 129715. [Google Scholar] [CrossRef]
- Pikuda, O.; Dumont, E.R.; Matthews, S.; Xu, E.G.; Berk, D.; Tufenkji, N. Sub-lethal effects of nanoplastics upon chronic exposure to daphnia magna. Journal of Hazardous Materials Advances 2022, 7, 100136. [Google Scholar] [CrossRef]
- De Souza Machado, A.A.; Ghadernezhad, N.; Wolinska, J. Potential for high toxicity of polystyrene nanoplastics to the european daphnia longispina. Environmental Sciences Europe 2023, 35, 78. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Huang, Y.H.; Jiao, Y.; Chen, Q.; Wu, D.L.; Yu, P.; Li, Y.M.; Cai, M.Q.; Zhao, Y.L. Polystyrene nanoplastic induces ros production and affects the mapk-hif-1/nfκb-mediated antioxidant system in daphnia pulex. Aquatic Toxicology 2020, 220. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Pérez, E.; Jiang, Q.; Chen, Q.; Jiao, Y.; Huang, Y.; Yang, Y.; Zhao, Y. Polystyrene nanoplastic induces oxidative stress, immune defense, and glycometabolism change in daphnia pulex: Application of transcriptome profiling in risk assessment of nanoplastics. Journal of Hazardous Materials 2021, 402, 123778. [Google Scholar] [CrossRef]
- Liu, Z.; Cai, M.; Yu, P.; Chen, M.; Wu, D.; Zhang, M.; Zhao, Y. Age-dependent survival, stress defense, and ampk in daphnia pulex after short-term exposure to a polystyrene nanoplastic. Aquatic Toxicology 2018, 204, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lins, T.F.; O'Brien, A.M.; Kose, T.; Rochman, C.M.; Sinton, D. Toxicity of nanoplastics to zooplankton is influenced by temperature, salinity, and natural particulate matter. Environmental Science-Nano 2022, 9, 2678–2690. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Lee, Y.H.; Kim, M.S.; Lee, J.S.; Seo, J.S.; Wang, M.; Lee, J.S. Nanoplastic ingestion enhances toxicity of persistent organic pollutants (pops) in the monogonont rotifer brachionus koreanus via multixenobiotic resistance (mxr) disruption. Environ Sci Technol 2018, 52, 11411–11418. [Google Scholar] [CrossRef] [PubMed]
- Mavrianos, S.; Manzi, F.; Agha, R.; Azoubib, N.; Schampera, C.; Wolinska, J. Nanoplastics modulate the outcome of a zooplankton-microparasite interaction. Freshwater Biology 2023, 68, 847–859. [Google Scholar] [CrossRef]
- De Felice, B.; Sugni, M.; Casati, L.; Parolini, M. Molecular, biochemical and behavioral responses of daphnia magna under long-term exposure to polystyrene nanoplastics. Environment International 2022, 164, 107264. [Google Scholar] [CrossRef]
- Ekvall, M.T.; Gimskog, I.; Hua, J.; Kelpsiene, E.; Lundqvist, M.; Cedervall, T. Size fractionation of high-density polyethylene breakdown nanoplastics reveals different toxic response in daphnia magna. Scientific Reports 2022, 12. [Google Scholar] [CrossRef]
- Liu, Z.; Cai, M.; Wu, D.; Yu, P.; Jiao, Y.; Jiang, Q.; Zhao, Y. Effects of nanoplastics at predicted environmental concentration on daphnia pulex after exposure through multiple generations. Environmental Pollution 2020, 256, 113506. [Google Scholar] [CrossRef]
- Heinlaan, M.; Viljalo, K.; Richter, J.; Ingwersen, A.; Vija, H.; Mitrano, D.M. Multi-generation exposure to polystyrene nanoplastics showed no major adverse effects in daphnia magna. Environmental Pollution 2023, 323, 121213. [Google Scholar] [CrossRef]
- Nasser, F.; Constantinou, J.; Lynch, I. Nanomaterials in the environment acquire an “eco-corona” impacting their toxicity to daphnia magna—a call for updating toxicity testing policies. PROTEOMICS 2020, 20, 1800412. [Google Scholar] [CrossRef]
- Nasser, F.; Lynch, I. Secreted protein eco-corona mediates uptake and impacts of polystyrene nanoparticles on daphnia magna. Journal of Proteomics 2016, 137, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Abdolahpur Monikh, F.; Baun, A.; Hartmann, N.B.; Kortet, R.; Akkanen, J.; Lee, J.-S.; Shi, H.; Lahive, E.; Uurasjärvi, E.; Tufenkji, N. , et al. Exposure protocol for ecotoxicity testing of microplastics and nanoplastics. Nature Protocols 2023. [Google Scholar] [CrossRef] [PubMed]
- Heinlaan, M.; Kasemets, K.; Aruoja, V.; Blinova, I.; Bondarenko, O.; Lukjanova, A.; Khosrovyan, A.; Kurvet, I.; Pullerits, M.; Sihtmäe, M. , et al. Hazard evaluation of polystyrene nanoplastic with nine bioassays did not show particle-specific acute toxicity. Sci. Total Environ. 2020, 707, 136073. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Shi, H.; Li, X.; Gao, C.; Liu, R. Combined toxicity of micro/nanoplastics loaded with environmental pollutants to organisms and cells: Role, effects, and mechanism. Environment International 2023, 171, 107711. [Google Scholar] [CrossRef]
- Li, P.; Li, Q.; Hao, Z.; Yu, S.; Liu, J. Analytical methods and environmental processes of nanoplastics. Journal of Environmental Sciences 2020, 94, 88–99. [Google Scholar] [CrossRef]
- El Hadri, H.; Gigault, J.; Maxit, B.; Grassl, B.; Reynaud, S. Nanoplastic from mechanically degraded primary and secondary microplastics for environmental assessments. NanoImpact 2020, 17, 100206. [Google Scholar] [CrossRef]
- Catarino, A.I.; Patsiou, D.; Summers, S.; Everaert, G.; Henry, T.B.; Gutierrez, T. Challenges and recommendations in experimentation and risk assessment of nanoplastics in aquatic organisms. TrAC Trends in Analytical Chemistry 2023, 167, 117262. [Google Scholar] [CrossRef]
- Mitrano, D.M.; Beltzung, A.; Frehland, S.; Schmiedgruber, M.; Cingolani, A.; Schmidt, F. Synthesis of metal-doped nanoplastics and their utility to investigate fate and behaviour in complex environmental systems. Nature Nanotechnology 2019, 14, 362–368. [Google Scholar] [CrossRef]
- Chia, W.Y.; Ying Tang, D.Y.; Khoo, K.S.; Kay Lup, A.N.; Chew, K.W. Nature’s fight against plastic pollution: Algae for plastic biodegradation and bioplastics production. Environmental Science and Ecotechnology 2020, 4, 100065. [Google Scholar] [CrossRef]
- He, Y.; Deng, X.; Jiang, L.; Hao, L.; Shi, Y.; Lyu, M.; Zhang, L.; Wang, S. Current advances, challenges and strategies for enhancing the biodegradation of plastic waste. Sci. Total Environ. 2024, 906, 167850. [Google Scholar] [CrossRef]
- Slaveykova, V. Phytoplankton controls on the transformations of metal-containing nanoparticles in aquatic environmen. In Environmental nanoparzicles: Sources, occurrence, analysis and fate, Jimenez Lamana, J., Szpunar J.,, Ed. Royal Society of Chemistry,: 2022; Vol. in press. [CrossRef]
According to the European Commission, plastic particles would be defined as nanomaterials if 50% or more of the plastic particles “in an unbound state or as an aggregate or as an agglomerate and where, for 50 % or more of the particles in the number size distribution, one or more external dimensions is in the size range 1 nm-100 nm. In specific cases and where warranted by concerns for the environment, health, safety or competitiveness the number size distribution threshold of 50 % may be replaced by a threshold between 1 and 50 %” [10]. However, most frequently the term NPLs is used for materials within the size range 1–1000 nm [11]. |
Figure 1.
(a) Key processes involving uptake- and toxico-availability of NPLs by phyto- and zooplankton. (b) Factors influencing the bioavailability of NPLs in aquatic environment.
Figure 1.
(a) Key processes involving uptake- and toxico-availability of NPLs by phyto- and zooplankton. (b) Factors influencing the bioavailability of NPLs in aquatic environment.
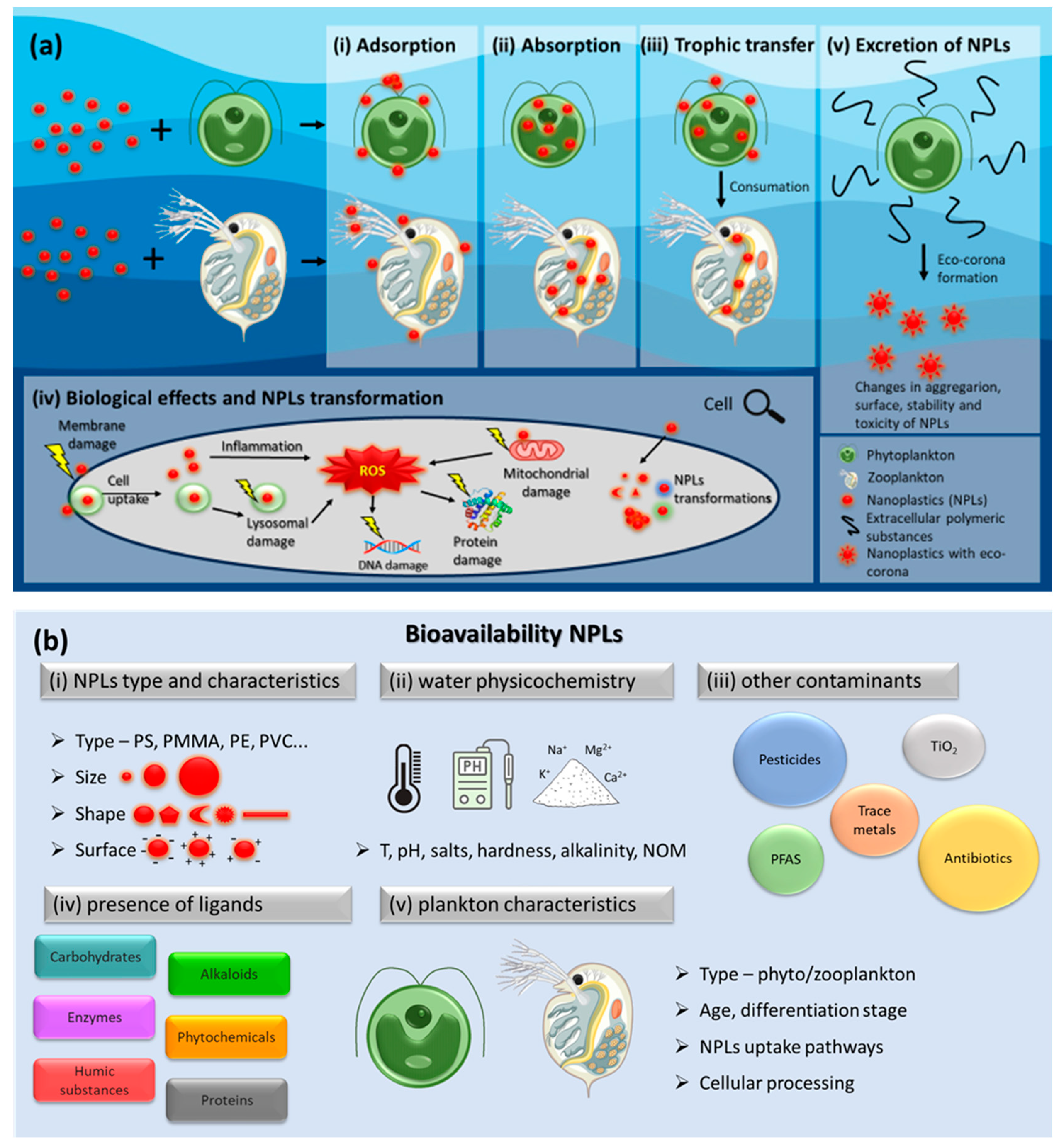
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



