
Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
The morphological characters of adult females of Trionymus aberrans Goux 1938 that had been collected in post-industrial wastelands and other habitats in Poland and other countries were studied. A distinctive morphological variability was observed in the specimens that had been collected from post-industrial wastelands in Poland. SEM photos of the morphological characters of T. aberrans are provided. The occurrence of unusual pores with four loculi was demonstrated for the first time in a species of the family Pseudococcidae. The importance of introducing additional morphological characters into the species description is discussed. New data on the frequency and host preference of T. aberrans are also provided. Our research is the first long-time study on scale insect species in post-industrial wastelands. The second-instar nymph of T. aberrans is described. The presence of translucent pores on the hind coxae in second-instar nymph was revealed.
Keywords:
nymphal stages
; adult females
; morphology
; spoil heaps
; SEM
1. Introduction
The Pseudococcidae (mealybugs) comprises the second largest family within the scale insects (Coccoidea: Pseudococcidae) with 2034 species (Garcia Morales et al., 2016). Currently, the family Pseudococcidae is divided into two subfamilies - the Phenacoccinae and Pseudococcinae [1,2,3,4,5].
The genus Trionymus Berg 1899, which comprises 123 species, is one of the largest genera of the subfamily Pseudococcinae [6]. Adult females of the genus Trionymus are characterized by the presence of 6-8-segmented antennae, legs normally developed, claws without denticles, both pairs of dorsal ostioles frequently well developed, trilocular pores on both body surfaces, multilocular pores always present on the ventral side of the abdomen, and frequently occur on its dorsal side, tubular ducts on both body surfaces, cerarii reduced in number always including at least the anal lobe pair, all abdominal cerarii with two conical setae, circulus present or absent [3,7,8]. The genus is distributed all over the world. According to Kosztarab and Kozár [8], the genus Trionymus comprises a number of morphologically similar species and as the species are inadequately described, the genus needs a thorough revision.
In their comprehensive review of Palaearctic mealybug fauna, Danzig and Gavrilov-Zimin [3] distinguished a Trionymus group of genera comprising: Trionymus Berg 1899, Dysmicoccus Ferris 1950, Balanococcus Williams 1962, Kiritshenkella Borchsenius 1948 and Saccharicoccus Ferris 1950. The largest genera Trionymus and Dysmicoccus are perhaps the most primitive genera in this group [3].
Only eight species of the genus Trionymus occur in Poland, namely, T. aberrans Goux 1938, T. hamberdi (Borchsenius 1949), T. newsteadi (Green 1917), T. perrisii (Signoret 1875), T. phalaridis (Green 1925), T. placatus (Borchsenius 1949), T. radicum (Danzig 1986) and T. thulensis Green 1931 [9].
T. aberrans is distributed in almost all of southern Europe and extends northwards into Germany and Poland and eastwards into Ukraine and Georgia. This species is also found in southwest and central Asia and as far east as East Asia [6]. T. aberrans occurs in the leaf sheaths of different grass species [6,10].
Descriptions and illustrations of the adult female have been provided by e.g. Goux [11], Schmutterer [12], Kosztarab and Kozár [8] and Danzig and Gavrilov-Zimin [3].
Preliminary studies revealed an intraspecific variability in the specimens of Trionymus aberrans that had been found in spoil heaps in Poland [13]. These observations were confirmed during the present research and, moreover, additional features that are characteristic of this species were revealed.
The structure and arrangement of the wax glands (pores and ducts) are the main taxonomic characters that are used in the classification of mealybugs and all scale insects. The following types of pores occur in mealybugs: simple discoidal pores, trilocular pores, quinquelocular pores and multilocular pores [2,7,14]. The presence of quadrilocular pores (with four excretory openings (loculi)) has never been demonstrated in any species of the family Pseudococcidae.
A scanning electron microscope (SEM) was used to study the morphological features of females of T. aberrans. To the best of our knowledge, the cuticular structures of species of the genus Trionymus have never been examined using SEM before.
The classification and species identification of scale insects is based almost entirely on the morphology of adult females. In the Pseudococcidae, the female has three nymphal stages, whereas the male has four [14]. The morphology of the nymphs of most species of Palearctic Pseudococcidae is unknown [3]. Only the first-instar nymph of Trionymus aberrans was briefly described by Goux [11], but it was not illustrated. The other nymphal stages of this species are unknown [3,8].
Translucent pores are structures that are found in many mealybug species. The function of these pores is not clear, but they possibly release pheromones since they occur almost solely in adult females [14].
The aims of this study were to make the comparison of the morphology and morphological anomalies between female specimens of T. aberrans collected in post-industrial wastelands and those found in undegraded habitats using light and scanning electron microscope (SEM) and to describe the second-instar nymph of this species.
2. Materials and Methods
Adult females of Trionymus aberrans were mainly collected in degraded post-industrial wastelands including three zinc spoil heaps that are located in Ruda Śląska (site no. 1), Piekary Śląskie (no. 2) and Bolesław (no. 3), one coal heap in Ruda Śląska (no. 4) and a degraded area near a production plant in Piekary Śląskie (no. 5) between 2011 and 2021 (Figure 1).
All of the sites are located in the southern part of Poland. Specimens were also collected in undegraded habitats (unnumbered sites), primarily in psammophilous and xerothermic grasslands in Poland (also in Ruda Śląska, Piekary Śląskie and Bolesław), Austria and the Czech Republic. The post-industrial wastelands were visited regularly each year from April until October, while the unnumbered sites were visited frequently, but not regularly. The females were collected from the leaves or leaf sheaths of different grass species using the hand-collecting method and preserved in 70% ethanol and mounted. The material was identified on the basis of the following keys: Borchsenius [15], Kosztarab and Kozár [8] and Danzig and Gavrilov-Zimin [3].
Type material and other specimens of Trionymus aberrans deposited in the Muséum national d’Histoire naturelle (MNHN), Paris were examined.
T. aberrans specimens were identified using a Nikon Eclipse E-600 biological microscope with a Nikon DS-Fi2 digital camera and NIS Elements at the IBBEP. The females were examined under a dissecting microscope before they were prepared and identifying species was possible due to the presence of the horseshoe-shaped anal ring, which is a unique character of Trionymus aberrans.
First-instar nymphs of Trionymus aberrans were not collected. Second-instar nymphs were collected from the undegraded areas and the post-industrial wastelands, but the vast majority of these specimens were highly parasitised. Third-instar females were also collected, but they were not examined because the specimens were significantly damaged. Adult males and male immature instars (prepupa and pupa) were not collected.
For the SEM, the method of Kanturski et al. [16] was adapted and used to dehydrate and dry the specimens. The material was dehydrated in a graded ethanol/water series of 75%, 80%, 90%, 95% and 100% for 10 min at each concentration with three 100% ethanol changes. The samples were critical point dried in a Leica EM CPD300 automated critical point dryer (Leica Microsystems, Vienna, Austria), mounted on aluminium stubs with double-sided adhesive carbon tabs and sputter coated with gold (20-30 nm thick film) in a Pelco SC-6 sputter coater (Ted Pella, Inc., Redding, CA, USA).
The images were captured using a Hitachi SU8010 field emission scanning electron microscope (Hitachi High-Technologies Corporation, Tokyo, Japan) at 5.0, 7.0 and 10.0 kV accelerating voltages at the IBBEP.
The description of the second-instar nymph of T. aberrans is based on three specimens.
The description scheme was adapted from Kaydan et al. [17] and was slightly modified. The morphological terminology of the wax glands (pores and ducts) as well as the numeration of the abdominal segments followed those of Danzig and Gavrilov-Zimin [3]. Almost all of the measurements were performed in micrometers (μm); only the body length and width were measured in millimeters (mm).
The specimens of T. aberrans that were collected during the present studies are deposited in the IBBEP.
In total, 225 female specimens of Trionymus aberrans were examined, 75 were collected in post-industrial wastelands and 150 in undegraded habitats. Specimens collected in undegraded habitats including specimens collected in psammophilous grasslands located in Piekary Śląskie, Bolesław and Ruda Śląska. Fifteen specimens from each post-industrial wasteland were examined.
3. Results
General information
A total of about 900 adult females of Trionymus aberrans were collected, most of them (620) from the post-industrial wastelands. About 40% of all of the collected females were parasitised. The highest degree of parasitisation was characteristic of the females that had been collected from the different habitats between August and October.
The females had been found on the leaves or in the leaf sheaths of different grass species. In the post-industrial wastelands, about 90% of the specimens were collected from Deschampsia caespitosa (L.) Palisot de Beauvois. Other grass species, e.g., Agrostis capillaris L., Elymus repens (L.) Gould, Phleum pratense L. and Poa compressa L. were other common host plants in the degraded habitats. The adult females formed colonies consisting of four to ten specimens only on Deschampsia caespitosa in the post-industrial wastelands. In the present study, adult females of T. aberrans were often collected from Brachypodium pinnatum (L.) Palisot de Beauvois and Phleum sp. from the undegraded habitats. On the undegraded habitats, e.g., psammophilous or xerothermic grasslands, the females most often occurred singly on their host plants.
Intraspecific variability in adult females of Trionymus aberrans
Material examined:
Type material:
Trionymus aberrans n. sp. / Hem. Coccidae / ♀ / Avena / Marseille / 1.VIII.1936 / 806 /a / Type / Holotype (MNHN)
Trionymus aberrans ovalis/ Type / Colmars (Basses-Alpes) / L. Goux coll / 19/VII/1939/ 1013 (MNHN)
3 adult females deposited in MNHN
225 adult female specimens were involved in the morphological examination, out of the ca. 900 T. aberrans adult females that were collected during the present studies and deposited in the entomological collections of the Zoology Research Group, Institute of Biology, Biotechnology and Environmental Protection, Faculty of Life Sciences, University of Silesia, Poland (supplementary material, Table S1).
Instar diagnosis – Diagnosed by the following combination of features: (i) horseshoe-shaped anal ring; (ii) presence of simple tubular ducts of two types (dorsal ducts larger than those of venter); (iii) multilocular pores present on both body surfaces; (iv) presence of 2 pairs of cerarii; (v) each anal lobe cerarius (C18) with 2 conical setae plus 6–16 trilocular pores and 3–4 auxiliary hair-like setae; (vi) circulus absent or rarely present and (vii) antennae 8–or rarely 9–segmented.
Description of a slide-mounted adult female – Body elongate oval, 4.25–4.92 mm long, 2.74 –0.64 mm wide. Eye marginal, 31–32 μm wide. Antennae 8 (rarely 9) segmented, 560–610 μm long; first segment 58.13–64.20 μm long; second 50.05–55.8 μm; third 36.42–41.13 μm long; fourth 26.58–36.77 μm long; fifth 30.68–34.16 μm long; sixth 27.40–31.03 μm long; seventh 31.84–38.03 μm long and apical segment 81.77–93.18 μm long. Number of sensory setae on antennal segments: first segment: 3, second: 4, third: 4, fourth: 3, fifth: 5, sixth: 4–5, seventh: 4, apical segment: 16–17. Apical seta 30.71–32.1 μm long, subapical seta 24.17–26.15 μm long. Clypeolabral shield 180.22–196.61 μm long, wide 143.63–160.77 μm. Labium 3–segmented, 74.25–82.32 μm long, 79.78–80.75 μm wide. Stylet loop reaches line between anterior spiracles. Anterior spiracles 55.52–65.39 μm long, 28.90–32.58 μm wide across atrium with 3–4 associated trilocular pores and one multilocular pore located nearby; posterior spiracles 67.27–78.35 μm long, 42.53–44.23 μm wide across atrium with 5–6 associated trilocular pores and 3–4 multilocular pores located nearby. Circulus absent or rarely small oval circulus present. Legs well developed; hind trochanter + femur 230.55–239.72 μm long; hind tibia + tarsus 266.15–283.66 μm long; hind claw 27.81–29.06 μm long. Hind coxae with translucent pores. Tarsal digitules setose, subequal in length, each 46.45–48.33 μm long. Claw digitules subequal in length, each 27.29–28.46 μm long, both capitate and thicker than tarsal digitules. Ostioles: both pairs present; each anterior ostiole with 4–8 trilocular pores and 1 or 2 setae (total for both lips); each posterior ostiole with 8-12 trilocular pores and 2 setae (total for both lips). Horse-shoe-shaped anal ring 90–110 μm wide, with 6 anal-ring setae, each setae 76.87–91.22 μm long.
Venter. Body setae slender, each 52.8–61.14 μm long, longest setae present medially on the head; apical setae of anal lobe 147.95–181.34 μm long. Multilocular pores each 6–8 μm wide, numerous on the abdomen, forming transverse rows and bands on four last abdominal segments; 1–3 on the head; present irregularly along body margins. Trilocular pores each 4–5 μm in diameter, scattered on the entire surface. Unusual 4-locular pores are present on the three last abdominal segments in some specimens. Simple tubular ducts of two sizes. Larger tubular ducts each 10.4–12.2 μm long and 3.94–4.71 μm wide at distal form transverse bands on the four last abdominal segments, group on the body margin on the sixth abdominal segment, present occasionally on the cephalothorax. Smaller tubular ducts 4.95–6.56 μm long and 2.62–3.1 μm wide scattered on the entire surface.
Dorsum. Derm membranous, with 2 pairs of cerarii (C17 and C18) on the penultimate and last abdominal segments; C17 with 2 cerarian setae; 3–6 trilocular pores between cerarian setae, and 1–2 auxiliary setae; anal lobe cerarii (C18) each with 2 conical setae, 6–16 trilocular pores and 3–4 auxiliary hair-like setae. Dorsal body setae hair-like, each 25.2–32.46 μm long, scattered on the head and the thorax, in rows on abdominal segments. Multilocular pores each 6.68–8.22 μm wide, forming transverse rows on the last three abdominal tergites. Trilocular pores each 3.5–4.25 μm in diameter, scattered on the entire surface. Simple tubular ducts of two sizes are scattered throughout the body surface, present on the head and thorax. Ducts of two sizes are slightly larger than ventral ducts, each 10.7–12.8 μm long and 3.94–4.9 μm wide at distal.
Comments:
The holotype of Trionymus aberrans (MNHN) had the following characteristic features: horseshoe-shaped anal ring, hind coxae with many translucent pores, simple tubular ducts of two types on both body surfaces (dorsal ducts larger than those of venter), numerous trilocular pores on dorsal and ventral body surfaces, multilocular pores present on both body surfaces, 2 pairs of cerarii, each penultimate cerarius (C17) with 2 conical setae plus 2 trilocular pores and 1 auxiliary seta, each anal lobe cerarius (C18) with 2 conical setae plus 8 trilocular pores and 4 auxiliary hair-like setae, circulus absent and antennae 8-segmented.
Morphological intraspecific variability in adult females of Trionymus aberrans were observed. The vast majority of females had 8-segmented antennae (Figure 2 (a)), but 11 of the collected specimens had 9-segmented antennae. The presence of 8- or 9-segmented antennae was not associated with the habitat. The type specimen of Trionymus aberrans ovalis (MNHN) had 9-segmented antennae.
Trilocular pores were present on both body surfaces in all specimens (Figure 3a,c). Multilocular pores were observed on dorsal and ventral abdominal segments (Figure 3a,b).
Morphological anomalies were observed in females collected in four of the examined post-industrial wastelands, namely zinc heaps (site no. 1, 2 and 3) and degraded area near the production plant (site no. 5). Specimens collected in coal spoil heap (site no. 4) possess only fewer number of translucent pores in hind coxae compared to specimens found in undegraded habitats.
Unusual pores were observed in seven females out of fifteen examined specimens that had been collected from one of the post-industrial wastelands, namely, the degraded area near the production plant in Piekary Śląskie (site no.5). Females with unusual pores were collected in 2017, 2018 and 2020. The detailed structure of these unusual pores were accurately observed using a scanning electron microscope (SEM). These pores were characterised by the presence of four excretory openings (loculi) (Figure 4a,b). These unusual pores were scattered on the ventral surface of the last three abdominal segments. These pores were somewhat similar in appearance to the trilocular pores but had one more excretory opening. No pores with four loculi were observed in females that had been collected from the undegraded habitats or in the females that had been deposited in the collections in MNHN and IBBEP.
Five adult females out of fifteen examined specimens that had been collected in the degraded area near a production plant in Piekary Śląskie (site no. 5) had a small and oval circulus between the third and fourth abdominal segments (Figure 2b–d). The females with circulus were collected in 2015, 2017 and 2020. The circulus was observed in a type specimen of Trionymus aberrans ovalis. The circulus was absent in all of the females that had been collected from the undegraded habitats and in almost all of the females deposited in the examined collections in MNHN and IBBEP. The circulus was also absent in the vast majority of specimens that had been collected from post-industrial habitats.
Translucent pores were present on the hind coxae in all of the adult females, but there are differences in the number of these pores between the females that had been collected from the post-industrial wastelands and the undegraded habitats. Generally, 60 females collected in post-industrial wastelands (site no. 1-5) had fewer translucent pores than the specimens from the undegraded habitats.
All of the adult females, both those that had been collected during the present study and those that had been deposited in the collections in MNHN and IBBEP, had 2 pairs of abdominal cerarii (C17 and C18). There were differences in the morphology of the two pairs of cerarii in the examined specimens. The penultimate cerarii (C17) most often consisted of 2 conical setae, 1 or 2 auxiliary hair-like setae and a diverse number of trilocular pores that ranged from 3 to 8. Most of the specimens that had been collected from both the post-industrial wastelands and the other habitats had 5 trilocular pores in C17. Twelve females of fifteen examined specimens collected from the spoil heap in Bolesław (site no. 3) possessed 3 trilocular pores and two of them had 4 trilocular pores in C17. Ten specimens with 6 trilocular pores in C17 were collected in undegraded habitats.
Each of the anal lobe cerarius (C18) most often consisted of 2 conical setae, 2 auxiliary hair-like setae and 6-16 trilocular pores. The number of trilocular pores in C18 differed in the females living in different habitats. Most of the females that had been collected from three of the examined zinc heaps in Ruda Śląska (site no. 1), Piekary Śląskie (site no. 2) and Bolesław (site no. 3) and also in site no. 5 near production plant had 6-7 trilocular pores in each anal lobe cerarius. Ten females collected in site no. 1 had 6 trilocular pores in C18, whereas two of them possessed 7 trilocular pores in C18. Thirteen females collected in site no. 2 had 6 trilocular pores in C18. Eleven specimens collected in site no. 3 had 6 trilocular pores in C18 and three specimens had 7 trilocular pores in C18. Ten females collected in site no. 5 had 6 trilocular pores in C18 (these females did not have circulus). The females that were collected during the present study from coal spoil heap (site no. 4) and undegraded habitats had anal lobe cerarii with 8-14 trilocular pores each, most of them had 10 trilocular pores in C18. The females that were deposited in the collections in MNHN and IBBEP had 8-16 trilocular pores in C18, but 13 specimens with 15-16 pores were only collected in psammophilous grasslands in Austria. In four specimens from the post-industrial wasteland (Bolesław, site no.3), there was an additional third conical seta in C18.
In specimens collected in undegraded psammophilous grasslands in Ruda Śląska, Piekary Śląskie and Bolesław morphological anomalies were not observed.
Simple tubular ducts of two sizes were present in all of the examined specimens of Trionymus aberrans (Figure 4a,c).
Description of a second-instar nymph of Trionymus aberrans
Material examined:
2261 / Trionymus aberrans / ♀ / larva / Graminae vagina / det. Koteja // 5.9.1967 / 6 / Las Wolski Kraków / leg. Koteja // SC28-249-1-004-DZUS;
Trionymus aberrans / nymph / Phleum sp. / Olsztyn near Częstochowa / 12.09.2018 // leg. et det. M. Kalandyk-Kołodziejczyk;
Trionymus aberrans / nymph / Deschampsia caespitosa / Piekary Śląskie, Lotników / 12.09.2020 // leg. et det. M. Kalandyk-Kołodziejczyk.
Instar diagnosis – Diagnosed by the following combination of features: (i) horseshoe-shaped anal ring; (ii) presence of a few simple tubular ducts of one type only on the last segments of the abdomen (dorsal ducts larger than those on venter); (iii) absence of multilocular pores; (iv) presence of 2 pairs of cerarii; (v) each anal lobe cerarius (C18) with 2 conical setae, 5 trilocular pores and 2 auxiliary hair-like setae; (vii) circulus absent and (viii) antennae 6-segmented.
Description of slide-mounted second-instar (Figure 5) – Body elongate oval, 1.1–1.3 mm long, 0.4–0.5 mm wide.
Eyes present marginally, 20.18 μm wide. Antennae 6–segmented; first segment 33.94–38.37 μm long; second 35.40–36.48 μm long; third 39.83–41.80 μm long; fourth 21.17–21.80 μm long; fifth 25.16–25.68 μm long and apical segment 70.51–71.88 μm long. Number of sensory setae on antennal segments: first segment: 3, second: 3, third: 3, fourth: 3, fifth: 3, apical segment: 13. Apical seta 25.58–26.21 μm long, subapical seta 14.11–15.45 μm long. Clypeolabral shield 137.94–142.62 μm long and 99.53–100.79 μm wide. Labium 54.09–54.82 μm long and 57.51–56.58 μm wide. Stylet loop reaches line between anterior spiracles. Anterior spiracles 44.12–44.43 μm long, 16.32–18 μm wide across atrium; posterior spiracles 44.12–45.8 μm long, 20.18–21.5 μm wide across atrium. Both anterior and posterior spiracles associated with 2 trilocular pores each. Circulus absent. Legs well developed: hind trochanter + femur 148.11–151.65 μm long, hind tibia + tarsus 167.04–168.30 μm long, hind claw 14.74–16.80 μm long. Hind coxae with a few translucent pores. Claw without denticle. Tarsal digitules slightly capitate, subequal in length, each 30.87–31.64 μm long. Claw digitules clubbed, subequal in length, each 20.16 μm long, slightly thicker than tarsal digitules. Ostioles: both pairs present; each anterior ostiole with 4 trilocular pores and 1 seta (total for both lips); each posterior ostiole with 8 trilocular pores and 2 setae (total for both lips). Anal ring horseshoe-shaped, 42.46 μm 43.88 μm wide, with 6 anal-ring setae, each setae 64.05–67.63 μm long.
Venter. Body setae slender, hair-like, 21.53–25.96 μm long, longest setae present medially on the head; apical seta of anal lobe 107.15–109.14 μm long. Multilocular pores absent. Trilocular pores each scattered evenly on the entire surface. Very few simple tubular ducts of one type only on the lateral margin of the VII abdominal segment. Each duct 4.87¬–5.00 μm long and 3.65–3.85 μm wide at the distal end.
Dorsum. Derm membranous, with 2 pairs of cerarii on body margin, C17 with 1 conical seta, 2 trilocular pores and 2 hair-like setae. Anal lobe cerarii (C18) each with 2 conical setae, each seta 8.56–8.82 μm long, 5 trilocular pores and 2 auxiliary hair-like setae. Dorsal body setae hair-like, shorter than ventral setae, each 13.6–16.24 μm long. Trilocular pores evenly scattered on dorsum. A few simple tubular ducts of one type present only on VI-VIII abdominal segments; on VI and VII segment, very few ducts on lateral margins of each segment; on VIII abdominal segment, a few ducts on lateral margin and in the middle. Each duct longer than ventral ducts, 5.82–5.88 μm long and 3.5–3.66 μm wide at distal end.
Comments: Species identification of second-instar nymph was based on the presence of a horseshoe-shaped anal ring. According to Gullan et al. [18], the sex of the second-instar nymph can only be determined for specimens that have the pharate third-instar male or female. Several authors [17,19,20] report that the second instar males have more dorsal tubular ducts than the second instar females.
Translucent pores were observed on the hind coxae in all of the examined second-instar nymphs of Trionymus aberrans. These pores have so far been observed almost solely in adult females [14].
One of the examined nymphs had been collected from a post-industrial wasteland in Piekary Śląskie (site no. 5). Further studies with more specimens are necessary to investigate the existence of the intraspecific variability in the nymphs from post-industrial wastelands.
4. Discussion
Although studies on the occurrence of scale insects in anthropogenic habitats have been conducted in Poland and other countries [21,22,23], these studies did not include the coccoid fauna from post-industrial wastelands. We are not aware of any research that has been conducted on the scale insects from post-industrial wastelands other than those of Lubiarz and Golan [24], Lubiarz and Cichocka [25], Kalandyk-Kołodziejczyk et al. [26] and Mróz et al. [13]. Our study is the first long-time study conducted on scale insect species found in post-industrial wastelands.
Differences in the abundance and host-plant preferences have been observed between the adult females of Trionymus aberrans from the post-industrial wastelands and those living in undegraded habitats in Poland and other countries. In habitats such as xerothermic and psammophilous grasslands, one or two adult females have usually been observed on a single grass [27,28], while in the degraded areas that were examined in this study, from four to ten females were found on a single host plant.
A significantly higher number of individuals in heavily degraded areas compared to natural habitats have also been observed in Parthenolecanium rufulum (Cockerell 1903) [24,25]. The number of individuals of P. rufulum that were found on oaks in degraded areas in the neighborhood of a nitrogen fertiliser factory in Puławy, Poland were several times (more than 60) greater than in the natural habitats in Polesie National Park [25]. This may mean that difficult environmental conditions including soil contamination and low humidity in post-industrial areas are favorable for the occurrence of some scale insect species including T. aberrans.
Adult females of Trionymus aberrans have been found in both post-industrial wastelands and undegraded habitats in Poland and other countries from June until October, which is consistent with the results that were obtained by Schmutterer [12] in Germany. Many of the specimens that were collected in the present study were parasitised by a species of Hymenoptera, which Schmutterer [12] also observed.
Our research indicates that T. aberrans has different trophic preferences in degraded habitats than in other habitats.
Morphological variability in many species of the family Pseudococcidae has often been observed e.g. [2,3,13,20,29,30,31]. As was emphasized by Williams [14], the morphological differences in a single species can be quite striking. A considerable number of morphological variations in some mealybug species appear to be environmentally induced [20,30,32,33]. Mróz et al. [13] observed a smaller number of trilocular pores in anal lobe cerarii (C18) and a presence of very few translucent pores in hind coxae in specimens found in post-industrial wastelands in comparison to specimens collected in other habitats. Detailed morphological features of five examined specimens found in spoil heaps were not provided [13].
We observed unusual pores with four excretory openings (loculi) in seven specimens of Trionymus aberrans that had been collected from the degraded area in Piekary Śląskie (site no.5). To the best of our knowledge, the presence of such unusual pores in species of the genus Trionymus has never been described before. It is worth emphasizing that this type of pore has never been described in other species of the Pseudococcidae [3]. These unusual pores in Trionymus aberrans are not similar to the quadrilocular pores that are characteristic of the species that belong to other scale insect families, although four loculi are present therein. We named them “unusual pores with 4 loculi”. Perhaps these unusual pores are due to the transformation of the trilocular pores. It appears that the presence of these unusual pores might be associated with the environmental conditions in human-transformed and polluted habitats.
Although there are always two pairs of cerarii in the adult females of T. aberrans, our studies revealed that their morphology is variable.
Differences in other features were only observed in the adult females of Trionymus aberrans that had been found in post-industrial wastelands. In our opinion, the larger range of numbers of trilocular pores in the last pair of cerarii (C18), i.e., from 6-16 could be included in the species description. Moreover, four of the females that had been collected from the post-industrial wastelands had an additional thick conical seta in C18, which has not been observed by other authors e.g. [3,8,15,34]. Further research must be conducted to determine whether these features are characteristic for individuals from degraded areas.
Trionymus aberrans is characterized by the presence of hind coxae with translucent pores [3,8,15]. Although these pores were observed in all of the studied material, we observed differences in their number. The females that had been collected from the undegraded habitats generally had numerous translucent pores on their hind coxae, while those that had been collected from the post-industrial habitats had only few of them, which was observed earlier by Mróz et al. [13]. According to Williams [14], although the function of these pores is still not clear, because they are only present in the adult females, they probably emit pheromones. We also observed these pores in a second-instar nymph, so perhaps their function is different.
The adult females of Trionymus aberrans usually do not have a circulus [3,8,15], but a circulus is present in some specimens [34,35]. Only five females of T. aberrans from one post-industrial wasteland (site no.5) had a small circulus between abdominal segments 3 and 4. According to Danzig and Gavrilov-Zimin [2,3] mealybugs can have any number of circuli from one to five or may lack circulus, although the most commonly encountered condition is a single circulus that is situated on the abdomen. While there are species that generally lack a circulus, a circulus can be present in occasional specimens [14]. Females both with and without a circulus can be found in the same population [3].
Specimens collected in examined post-industrial wastelands differed in morphological anomalies. Unusual 4-locular pores and circulus were present only in females collected from the degraded area near the production plant in Piekary Śląskie (site no. 5). Smaller number of trilocular pores in C18 were observed in most females collected in this site and in three zinc heaps (sites no. 1-3). Females collected from coal spoil heap in Ruda Śląska did not possess morphological anomalies except for fewer number of translucent pores in hind coxae. This coal spoil heap was a subject of recultivation and covered entirely with vegetation, so the environmental conditions are different from those in other examined post-industrial wastelands.
What is worth emphasizing, the specimens collected from undegraded habitats in the same towns, where post-industrial wastelands are situated, did not possess morphological anomalies.
Our research enabled a significant morphological variability in adult females of Trionymus aberrans to be observed, especially in the specimens that had been collected from the post-industrial wastelands. Further studies are needed to investigate the morphological variability in other species of the family Pseudococcidae from highly contaminated habitats. This future research will help to clarify whether the variability in morphological features is a characteristic of post-industrial habitats.
The classification and species identification of scale insects is based almost entirely on the morphology of adult females [3,8]. It is worth emphasizing that often structures such as circuli, conical setae and ostioles are present in the nymphal stages but are absent in the imago and vice versa. Nymphal instars have not been sufficiently studied, despite the occurrence of the conserved morphological features and their potential to resolve evolutionary relationships [19]. Identifying species is often impossible for the immature stages and males, although the immature stages often make up a large proportion of a sample [36]. Little information is available that permits the species identification of the immature instars of the pseudococcid species [19]. We provide a description of the second-instar nymph, which is probably an immature female instar.
It is worth emphasizing that translucent pores were observed on the hind coxae in all of the examined second-instar nymphs. These pores have been observed almost exclusively in adult females. According to Williams [14], only the Australian species Sphaerococcus casuarinae has similar pores in its immatures.
According to Williams [14], there is a great need for taxonomic revisions of mealybugs that are based on the external morphological characters of the different instars and adult females.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of examined microscope slides of adult females of Trionymus aberrans Goux, 1938.
Author Contributions
Conceptualization, M.K.-K.; M.W.; J.B; methodology, M.K-K.; M.W.; B.F.-P; investigation M.K.-K.; M.W.; E.P; J.B.; resources, M.K.-K.; M.W.; data curation B.F.-P.; J.B.; writing—original draft preparation M.K.-K.; M.W.; E.P; B.F.-P.; J.B.; writing—review and editing M.K.-K.; B.F.-P.; J.B.; visualization, M.K.-K.; M.W.; B.F.-P.; project administration M.K-K.; funding acquisition M.K.-K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Ministry of Education and Science of Poland as a part of the statutory activities of the Faculty of Natural Sciences, University of Silesia in Katowice.
Acknowledgments
We are very grateful to Marzena Zmarzły MSc (University of Silesia, Faculty of Natural Sciences, Institute of Biology, Biotechnology and Environmental Protection–IBBEP) for making the drawing of the second-instar nymph. The authors would also like to thank Izabela Potocka PhD (IBBEP) for taking the SEM pictures. Our thanks are also extended to Professor Piotr Węgierek (IBBEP) for translating the publications in Russian. We also thank Krzysztof Kudła (IBBEP) for his technical support. We are very grateful to Mrs. Michele Simmons for checking the English version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hardy, N.B.; Gullan, P.J.; Hodgson C.J. A subfamily-level classification of mealybugs (Hemiptera: Pseudococcidae) based on integrated molecular and morphological data. Syst Entomol 2008, 33: 51–71. [CrossRef]
- Danzig, E.M.; Gavrilov-Zimin, I.A. Palaearctic mealybugs (Homoptera: Coccinea: Pseudococcidae), Part 1: Subfamily Phenacoccinae.- Zoological Institute of Russian Academy of Sciences, St. Petersburg, Russia, 2014; 1–678.
- Danzig, E.M.; Gavrilov-Zimin, I.A. Palaearctic mealybugs (Homoptera: Coccinea: Pseudococcidae), Part 2: Subfamily Pseudococcinae. Zoological Institute of Russian Academy of Sciences, St. Petersburg, Russia, 2015; 1–619.
- Gavrilov-Zimin, I.A. System of generic groups in mealybugs (Homoptera: Coccinea: Pseudococcidae).- Zoosystematica Ross. 2015, 24 (2): 237–260. [CrossRef]
- Kaydan, M.B.; Kozár, F.; Hodgson, C.J. A review of the phylogeny of Palaearctic mealybugs (Hemiptera: Coccomorpha: Pseudococcidae).- Arthropod Syst. Phylogeny 2015b, 73 (1): 175–195.
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database (Oxford). 2016 Feb 9;2016:bav118. [CrossRef]
- Ferris, G.F. Atlas of the Scale Insects of North America. (ser. 5). The Pseudococcidae (Part I). Stanford University Press Palo Alto, California, USA, 1950; 1–278.
- Kosztarab, M.P.; Kozár, F. Scale Insects of Central Europe. Akadémiai Kiadó Budapest, Hungary, 1988; 1–456.
- Łagowska, B.; Golan, K. An updated annotated checklist of scale insects (Hemiptera, Sternorrhyncha, Coccomorpha) of Poland. ZooKeys 2020, 918: 65–81.
- Bugaj-Nawrocka, A.; Junkiert, L.; Kalandyk-Kołodziejczyk, M.; Wieczorek, K. Scale insects (Hemiptera: Coccomorpha) in the entomological collection of the Zoology Research Group, University of Silesia in Katowice (DZUS) Poland. Bonn Zooll Bull 2021, 70 (2): 281–315.
- Goux, L. Notes sur les coccides [Hem.] de la France (23e note). Description d'un Trionymus nouveau. Bull Mens Soc Linn Lyon 1938, 7: 166–169. [CrossRef]
- Schmutterer, H. Über drei neue an Gräsern lebende Schildlausarten der deutschen Fauna. Beitr Entomol 1952, 2: 554–566.
- Mróz, E.; Kalandyk-Kołodziejczyk, M.; Simon, E. Preliminary morphological and molecular study of species of Saccharicoccus Ferris, 1950 and Trionymus Berg, 1899. Genus 2013, 24 (2): 117–130.
- Williams, D.J. Australian mealybugs. British Museum (Natural History), London, UK, 1985; 1–431.
- Borchsenius, N.S. Insects Homoptera. Suborders mealybugs and scales (Coccoidea). Family mealybugs (Pseudococcidae). Vol. VII. Fauna SSSR. Zoologicheskii Institut Akademii Nauk SSSR. N.S.. 1949; pp. 1-382.
- Kanturski, M.; Karcz, J.; Wieczorek, K. Morphology of the European species of the genus Eulachnus (Hemiptera: Aphididae: Lachninae). A SEM comparative and integrative study. Micron 2015, 76: 23–36.
- Kaydan, M.B.; Kilinçer, N.; Kondo, T. Descriptions of all Female Stages of the Maple Mealybug, Phenacoccus aceris (Hemiptera: Coccoidea: Pseudococcidae), with notes on its biology. Acta Zool Acad Sci Hungaricae 2015a, 61 (3): 255–277. [CrossRef]
- Gullan, P.J.; Downie, D.A.; Steffan, S.A.; A new pest species of the mealybug genus Ferrisia Fullaway (Hemiptera: Pseudococcidae) from the United States. Ann Entomol Soc Am 2003, 96 (6): 723–727. [CrossRef]
- Gullan, P.J. Identification of the immature instars of mealybugs (Hemiptera: Pseudococcidae) found on citrus in Australia.- Aust J Entomol 2000, 39: 160–166. [CrossRef]
- Hodgson, C.J.; Abbas, G.; Arif, M.J.; Saeed, S.; Karar, H. Phenacoccus solenopsis Tinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation. Zootaxa 2008, 1913: 1–35. [CrossRef]
- Łagowska, B. Występowanie czerwców (Homoptera, Coccinea) na drzewach i krzewach w środowisku miejskim,. In: Fauna miast. Urban fauna, Barczak, T., Indykiewicz P., Eds. Wydawnictwo ATR, Bydgoszcz, 1998; pp. 63–71.
- Kaydan, M.B.; Kilinçer, N.; Kozár, F. Studies on Pseudococcidae (Hemiptera Coccoidea) fauna of urban ecosystem of Ankara Province, Turkey. Boll Zool Agrar Bachicolt 2005, 37 (2): 85–95.
- Malumphy, C.P.; Kahrer, A. New data on the scale insects (Hemiptera: Coccoidea) of Vienna, including one invasive species new for Austria. Beiträge zur Entomofaunistik 2011,12: 47–60.
- Lubiarz, M.; Golan, K. Parthenolecanum rufulum (Cockerell) on pedunculate oak (Quercus robur L.) in the neighbourhood of the Nitrogen Fertilizer Factoring in Puławy and Poleski National Park. In: Aphids and other Hemipterous insects. Monograph 11, Wilkaniec, B., Ed. Agricultural University Publisher, Poznań, Poland, 2005; pp. 95–105.
- Lubiarz, M., Cichocka, E. Arthropods on pedunculate oak (Quercus robur L.) in Poleski National Park in the neighbourhood of a nitrogen fertilisers’ factory in Puławy (Zakłady Azotowe in Puławy). In: Aphids and other Hemipterous insects. Monograph 11, Wilkaniec B., Ed. Agricultural University Publisher, Poznań, Poland, 2005; pp. 85–94.
- Kalandyk-Kołodziejczyk, M.; Simon, E.; Herczek, A. Scale insects (Hemiptera, Coccoidea) of the post-industrial wastelands of Ruda Śląska (Poland). In: Urban Fauna. Studies of animal biology, ecology and conservation in European cities. Indykiewicz P., Ed. University of Technology and Life Sciences in Bydgoszcz, Poland, 2011; pp. 143–150.
- Kalandyk, M.; Węgierek, P. Scale insects (Hemiptera, Sternorrhyncha, Coccoidea) of selected plant communities in the eastern part of Garb Tarnogórski. Annals of the Upper Silesian Museum in Bytom. The Monograph, Entomology, 2010; 19: 1–116.
- Simon, E.; Herczek, A. Scale insects (Coccoidea) of the Landscape Park „Cistercian Landscape Compositions of Rudy Wielkie”. Katowice, Poland, 2010; 1-127.
- Danzig, E.M. Taxonomic status of Phenacoccus transcaucasicus Hadz. (=Ph. mespili sensu Borchsenius, 1949) and its intraspecific variability (Homoptera, Coccinea, Pseudococcidae).- Boll Zool Agrar Bachicolt 2002, 33 (3): 161–162.
- Cox, J.M. An experimental study of morphological variation in mealybugs (Homoptera: Coccoidea: Pseudococcidae). Syst Entomol 1983, 8: 361–382. [CrossRef]
- Simon, E.; Kalandyk-Kołodziejczyk, M. Morphological variability in Phenacoccus interruptus Green 1923 (Coccoidea: Pseudococcidae). Genus 2012, 23 (4): 585–590.
- Cox, J.M. Identification of Planococcus citri (Homoptera: Pseudococcidae) and the description of a new species.- Syst Entomol 1981, 6: 47–53. [CrossRef]
- Tanaka, H. A first record of Dysmicoccus walkeri (Newstead, 1891) (Hemiptera: Coccomorpha: Pseudococcidae) from Japan, with a key to Japanese species of the genus. J Asia Pac Entomol 2017, 20: 417–420. [CrossRef]
- Goux, L. Notes sur les Trionymus de la France et sur quelques espèces nouvelles pour la faune Francaise (Hem. Coccidae) (Notes sur les coccides (Hem. Coccidae) de la France (30e note). Bull Soc Hist Nat Afr N 1941, 32: 31–44.
- Gavrilov, I.A. Legless mealybugs (Homoptera: Pseudococcidae) from Russia and neighbouring countries.- Annal Zool 2003, 53 (3): 205–209.
- Assefa, Y.; Malindzisa, N. Molecular Identification of Mealybugs (Hemiptera: Pseudococcidae) on Cultivated, Ornamental and Wild Host Plants in Swaziland. Asian Res J Agric 2018, 9 (1): 1–10. [CrossRef]
Figure 1.
Sampling sites (post-industrial wastelands) in Upper Silesia, Poland. Map from https://pl.wikipedia.org/wiki/Konurbacja_górnośląska, modified with Adobe Photoshop.
Figure 1.
Sampling sites (post-industrial wastelands) in Upper Silesia, Poland. Map from https://pl.wikipedia.org/wiki/Konurbacja_górnośląska, modified with Adobe Photoshop.
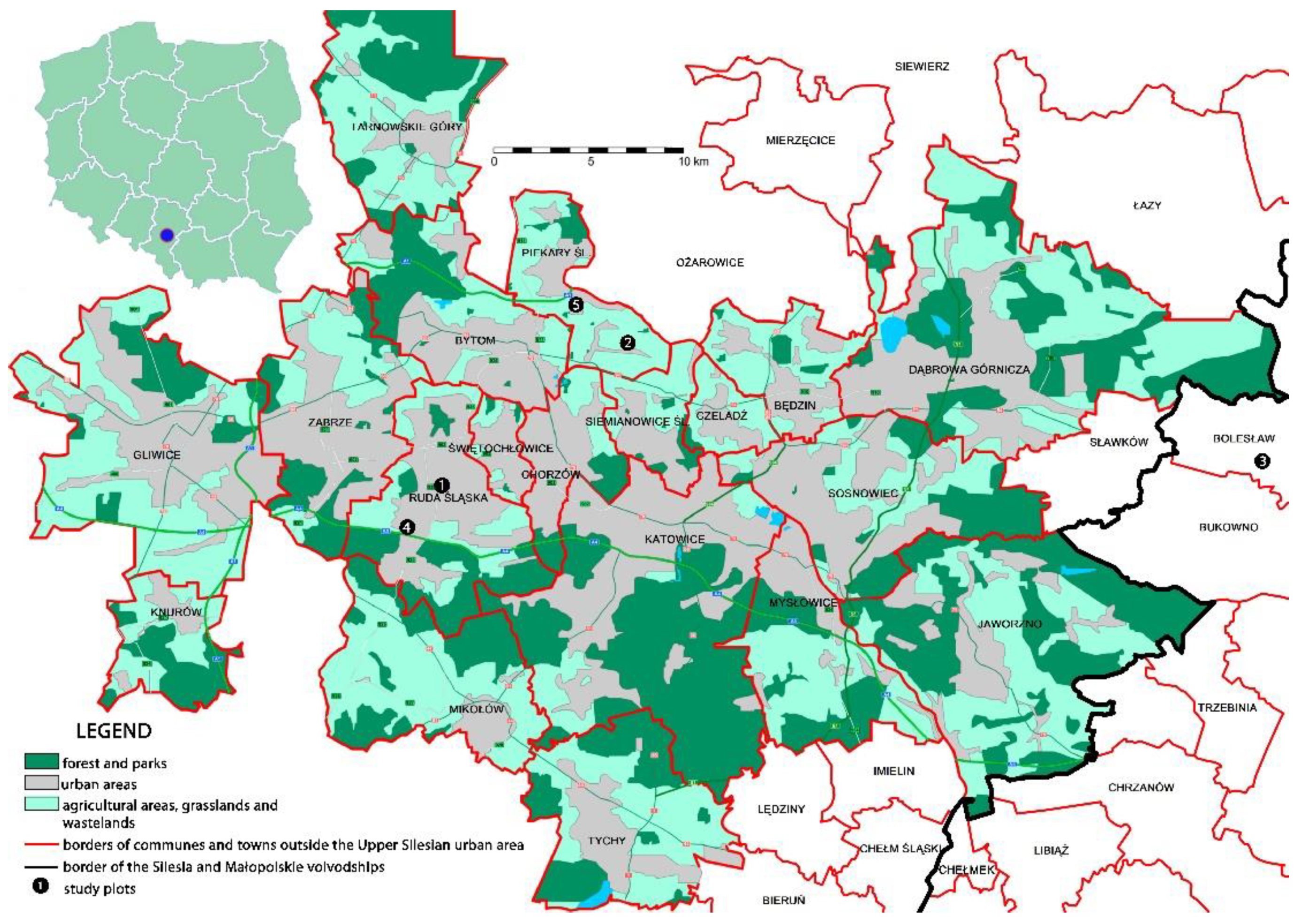
Figure 2.
Adult females of Trionymus aberrans Goux 1938 collected from post-industrial wastelands in Poland. (SEM). (a) female without circulus, ventral view; (b) female with circulus, ventral view; (c) and (d) circulus (marked with white arrows).
Figure 2.
Adult females of Trionymus aberrans Goux 1938 collected from post-industrial wastelands in Poland. (SEM). (a) female without circulus, ventral view; (b) female with circulus, ventral view; (c) and (d) circulus (marked with white arrows).

Figure 3.
Abdominal segment of adult female of Trionymus aberrans Goux 1938 collected from post-industrial wasteland in Poland. (SEM). (a) multilocular and trilocular pores (marked with white arrows), ventral view; (b) multilocular pore; (c) trilocular pore.
Figure 3.
Abdominal segment of adult female of Trionymus aberrans Goux 1938 collected from post-industrial wasteland in Poland. (SEM). (a) multilocular and trilocular pores (marked with white arrows), ventral view; (b) multilocular pore; (c) trilocular pore.
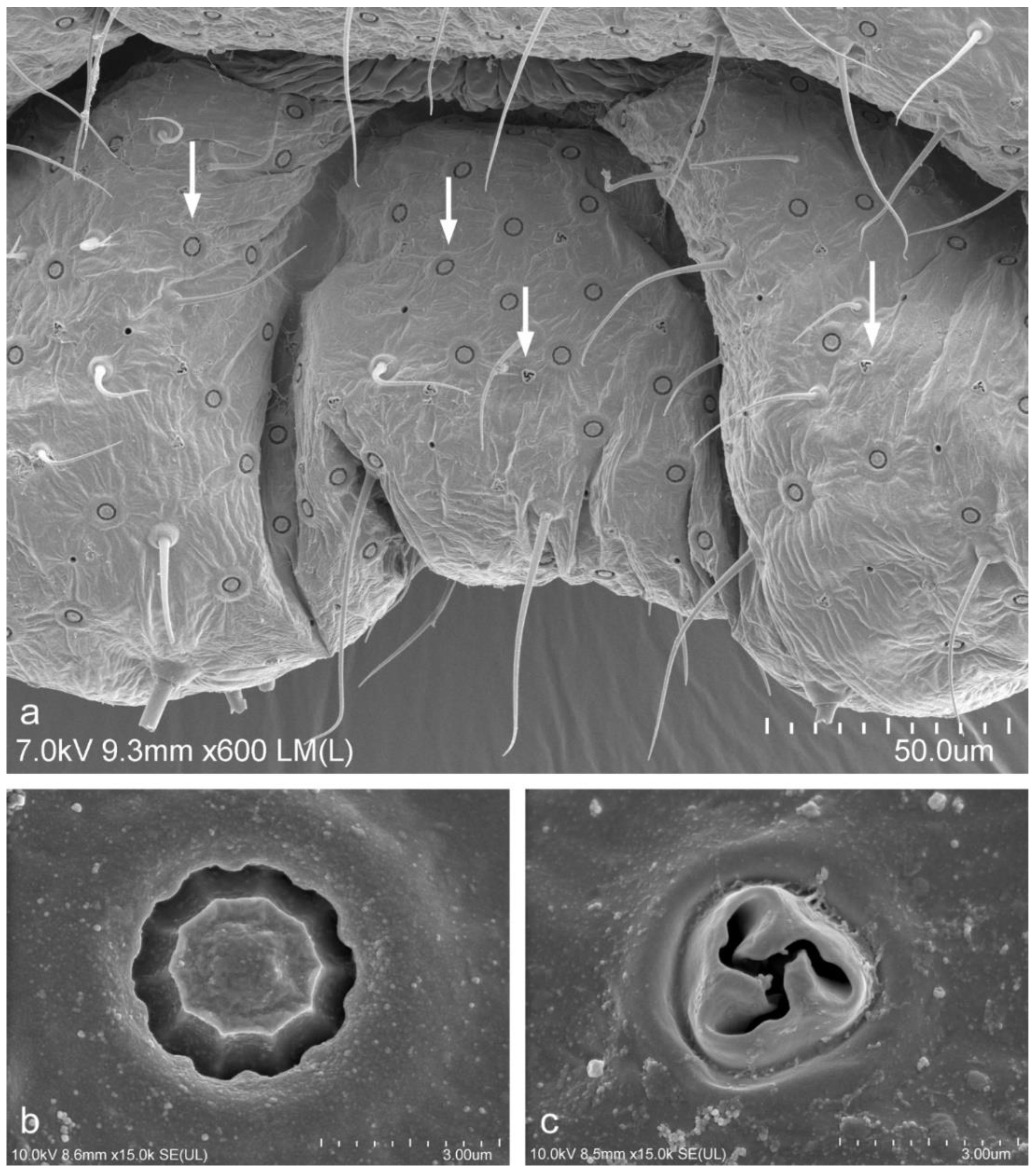
Figure 4.
Abdominal segment of adult female of Trionymus aberrans Goux 1938 (SEM) collected from post-industrial wasteland in Poland. (a) unusual pore with 4 loculi and simple tubular ducts, ventral view (marked with white arrows); (b) unusual pore with 4 loculi; (c) simple tubular duct.
Figure 4.
Abdominal segment of adult female of Trionymus aberrans Goux 1938 (SEM) collected from post-industrial wasteland in Poland. (a) unusual pore with 4 loculi and simple tubular ducts, ventral view (marked with white arrows); (b) unusual pore with 4 loculi; (c) simple tubular duct.

Figure 5.
Second-instar nymph of Trionymus aberrans Goux 1938; (a) antenna; (b) anterior spiracle; (c) tarsus of the hind leg; (d) tubular duct; (e) trilocular pore; (f) anal lobe cerarii (C18).
Figure 5.
Second-instar nymph of Trionymus aberrans Goux 1938; (a) antenna; (b) anterior spiracle; (c) tarsus of the hind leg; (d) tubular duct; (e) trilocular pore; (f) anal lobe cerarii (C18).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



