Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
. Shrub patches influence soil fertility and vegetation, impacting species composition and diversity. The Eastern Qilian Mountains' unique context provides insights into alpine ecosystems' response to environmental challenges. This study aimed to evaluate the physical characteristics and soil nutrient contents of shrub patches for four different shrub species. Four alpine shrub species (Salix oritrepha, Spiraea alpina, Rhododendron capitatum, and Potentilla fruticose) were studied, assessing their patch characteristics and soil nutrients at different depths within three patch microsites. Soil samples were collected and analyzed for organic matter, nitrogen, phosphorus, and potassium content. Statistical analyses were conducted to evaluate differences among shrub species and locations within the shrub patches. Our results showed that the shrub species exhibited variations in patch characteristics and soil nutrient distribution. Soil nutrient content varied by depth and location within the shrub patches, with higher concentrations at the center. The relative interaction intensity (RII) showed nutrient aggregation or dispersion trends. The study highlighted the complex interactions between shrub characteristics and soil nutrients, emphasizing their influence on nutrient cycling, vegetation dynamics, and soil properties. These findings contribute to our understanding of alpine ecosystem dynamics and inform conservation and management strategies.
Keywords:
Shrub patch
; Alpine shrub plant
; Fertility Island
; Relative interaction intensity (RII)
1. Introduction
Shrub patch configuration can be defined as spatial plant characteristics such as cover, size, density, height, shape, biomass distribution, and ratios to adjacent bare ground or grass patches [1]. Globally, shrublands are widespread, covering an area of 8.5 × 106 km2, and consist of a mosaic of shrub patches interspersed with grass patches [2]. Global change and human interventions and their complex interactions have caused the encroachment of indigenous shrubs in arid and semiarid grassland regions, resulting in complex interactions with vegetation dynamics [3,4,5]. These shrub patches and their associated fertility islands are important self-organized systems and serve as "indicators" of ecosystem response to climate change or human activities. The presence of shrub patches acts as an early signal of ecosystem changes in response to these factors [6]. Many studies have investigated the characteristics of shrub patterns and their underlying mechanisms. It was found that the characteristics of patches, labyrinths, gaps, and stripes (among others) are due to the redistribution of soil water and competition for limiting nutrients [7,8]. The "fertility island effect" describes the competitive advantage that shrub patches have in relation to nutrient availability. Compared to adjacent grass patches, shrub patches exhibit significantly higher soil nutrient content, particularly in organic matter, nitrogen, phosphorus, and potassium [9]. This phenomenon highlights the capacity of shrubs to influence biogeochemical cycles, with soil inside the patches becoming enriched compared to the surrounding areas [9]. Shrub patches not only influence soil fertility but also have a substantial impact on vegetation dynamics. They often support different plant communities, influencing species composition, diversity, and distribution [10]. The spatial distribution of shrubs in the Eastern Qilian Mountains can vary according to species and local soil characteristics, further emphasizing the complex relationships between shrubs, soil, and vegetation [11]. Soil characteristics also influence shrub plants, and the shrub patches they form. That is, soil nutrients, temperature, and moisture control the processes of shrub invasion, plant diversity, and the formation of shrub patches [12]. According to Berdugo et al. (2019), the effects and impacts of different shrublands and grassland ecosystems on soil properties were different due to differences in habitat, vegetation, and soil properties [13].
The Eastern Qilian Mountains present a unique context for studying fertility islands in alpine ecosystems. This high-altitude region is characterized by harsh environmental conditions, including low temperatures and limited water availability. Various alpine shrub species, such as S. oritrepha, S. alpina, R. capitatum, and P. fruticosa, have adapted to these challenging circumstances. Each of these species exhibits distinct characteristics that allow them to thrive in this unique environment [14,15]. The study of fertility islands provides insights into how alpine ecosystems function in response to these environmental challenges [16]. Therefore, the analysis of shrubland characteristics, soil properties, and relationships among different vegetation types plays an important role in better analyzing the vegetation structure, landscape pattern, and feedback relationships among alpine grassland shrubs in the alpine vegetation ecosystem [17]. Therefore, this study aimed to evaluate the physical characteristics and soil nutrient contents of shrub patches at different depths within three patch microsites (center, edge, and mid-point between the two) for four different species of shrubs.[ (a) we determined the shrub patch characteristics and soil nutrient distribution of various shrub types, and (b) we analyzed the relationship between the shrub patch characteristics and soil nutrients to reveal the soil nutrient enrichment characteristics among shrub species].
2. Materials and Methods
2.1. Study area
The study conducted field experiments in the alpine grasslands of the eastern Qilian Mountains in Daiqian village (37° 14' 34" N, 102° 35' 38" E) in the Tianzhu Tibetan Autonomous County of Gansu Province of China. The area has a typical alpine climate and is usually cold and wet for most parts of the year. It also has weather conditions such as thin air with low oxygen concentrations, high solar, and high ultraviolet radiation. The area has an average annual temperature of -0.1 °C, an accumulated temperature of 1380 ℃ (average monthly minimum and maximum temperature of –18.3 °C and 12.7 °C, respectively), annual sunshine hours of 2600 h, annual precipitation of 416 mm (which mostly topographic rain concentrated in July, August, and September), and annual evaporation of 1592 mm. The planting season is about 120 d, ranging from May to September. The soil is a typical alpine Chernozem with about 40–80 cm thick, bulk density of about 0.73 g cm-3, organic C of about 138.45 g kg-1, TN of about 4.31 g kg-1, and TP of about 0.65 g kg-1. Figure 1 presents a map showing the description and altitude of the study area. The elevation ranges from 2900 to 4300 m. SO, SA, RC, and PF dominate the shrubby grassland ecosystem, with heights of 50–120cm [18]. All these alpine shrub species play important roles in maintaining the biodiversity and stability of their respective ecosystems. They provide food and shelter for various animal species and help to prevent soil erosion and retain moisture in the soil.
2.2. Field investigation
This study was conducted from July 2021 to August 2021 and was based on the principles of uniform distribution of shrub patches, elevation, and slope to avoid biases from site heterogeneity. Four shrublands were formed by four typical alpine shrubs: Salix oritrepha (SO), Spiraea alpina (SA), Rhododendron capitatum (RC), and Potentilla fruticosa (PF), which were widely distributed in the eastern Qilian Mountain. Quadrats of the herbaceous community under the shrub plots and grass plots adjacent to shrubland were randomly selected to survey the plant community with three 10 m × 10 m shrub patches set up in each sample to investigate the sample plot. Plant species, height, shrub area, litter depth, and biomass were measured in all patches in the sample plots. In each quadrant, all aboveground shrub parts were cut, collected, and put into separate labeled envelopes. To measure the belowground biomass, we used the complete excavation method (getting the entire root from the soil) to collect the entire root. The roots and the aboveground shrub parts were washed and oven-dried at 65 °C for 24 h to a constant weight for the determination of biomass. Six measurements at each quadrant were taken to determine the average shrub height [19].
2.3. Sampling and measurement of soil properties
Each shrub patch was approximated as a circle, and the center of the circle is the center of the shrub patches. Three patch microsites were set in each of the four directions of the patches: east, south, west, and north, respectively. The three sites were the center of the shrub patch (CS), the edge of the shrub patch (ES), and the midpoint between the center and the edge of the shrub patch (BC). Soil samples at each site were collected from beneath the organic layer after litter removal. They were kept separately in aluminum boxes. At the same time, the undisturbed core samples were taken at different depths using standard core steel samplers (55-mm internal diameter and 45-mm height). The soil was sampled at a time interval of 10 days at three depths 0–20 cm, 20–40 cm, and 40–60 cm at CS, ES, and BC and samples were taken at the same layer and same location points were then mixed to make a single sample. Soil samples in three positions of each site (CS, ES, and BC) at five depths were collected from beneath the organic layer after litter removal. They were kept separately in plastic bags. The soil samples were air-dried at room temperature for one week and sieved through a 2-mm sieve to remove stones roots and coarse debris. A soil passing sieve was used to determine the soil organic matter content (SOM). Soil organic matter was measured using the dichromate oxidation method [20]. Total nitrogen (TN) was determined using Kjeldahl analysis [21]. Total phosphorus (TP) and total potassium (TK) were fused by NaOH and measured by molybdenum antimony anti-colorimetric and flame photometry, respectively.
2.4. Data analysis
One-way ANOVA and the least significant difference (LSD) test were used to analyze the shrub characteristics and soil chemical properties of different types of shrub-encroached grassland. Furthermore, the Pearson correlation was used to examine the correlation between shrub characteristics and soil chemical properties. The relative interaction intensity (RII) was used to estimate the magnitude of the shrub patch effect on soil organic matter (SOM), total nitrogen (TN), total potassium (TK), and total phosphorus (TP)[22].
The relative interaction intensity (RII) was calculated as:
where and represent values for soil nutrient content at different position points and depths of shrub patch type, respectively. The index ranged from 1 to -1, and with RII values > 0, the positive and negative values indicate an increasing and decreasing effect on soil nutrients, respectively.
3. Results
3.1. Characteristics of shrub patch
We conducted measurements on four patches characterized by a single species of shrubs (Table 1). The presence of shrubs encroaching into grassland areas results in the formation of distinct shrub patches, which are regions predominantly covered by shrub vegetation. The features of these patches exhibit variations contingent on the specific shrub species and the extent of their encroachment. Among the species investigated, S. oritrepha displayed the most substantial values for patch area, shrub height, litter depth, and shoot biomass; however, it exhibited the lowest root biomass. In contrast, R. capitatum showcased the highest root biomass. P. fruticosa, on the other hand, displayed the lowest shrub height and shoot biomass, while S. alpina exhibited the smallest patch area and least litter depth. A noteworthy distinction is observed between S. oritrepha and R. capitatum, as they tend to form dense patches characterized by a high canopy cover and stem density. In contrast, S. alpina and P. fruticosa tend to create more dispersed and less densely populated patches.
3.2. Heterogeneity of soil nutrient distribution and the shrub types
We analyzed the soil nutrient profiles at a soil depth of 60 cm within various shrub patches situated at microsites within the patches (Table 2). Across the 0-60 cm soil layer, which includes the edge of the shrub patch (ES) and the center of the shrub patch (CS), we observed that Soil Organic Matter (SOM) reached its highest concentration in the PF shrub patch and its lowest in the SA shrub patch. In contrast, below the crown (BC), the highest SOM content was observed in the RC shrub patch, while the SA shrub patch displayed the least. Concerning total potassium levels in the three different microsites within the shrub plot, we found the highest concentration in the SO shrub patch and the lowest in the RC patch. In the 60 cm soil depth, we also assessed total nitrogen levels within the four different shrub types across the three patch microsites. The trend observed across this 60 cm range was consistent, with the highest concentration in the SO shrub patch, followed by SA, PF, and finally RC, in descending order. Total phosphorus levels followed a different pattern, with the highest concentration occurring in the PF shrub patch, followed by RC, SO, and SA in that order.
3.3. Distribution characteristics of shrub patch soil nutrients in different soil layers
The levels of soil organic matter (SOM), total nitrogen (TN), total potassium (TK), and total phosphorus (TP) within the various shrub patches and the different locations within the shrub plot were consistently higher in the 40–60 cm soil layer compared to the 0–20 cm soil layer (Figure 2). Through one-way analysis of variance (ANOVA), it was determined that the SOM and TN values within the SO, SA, and RC shrub patches exhibited significant differences between the 0–20 cm and 40–60 cm soil layers (p < 0.05). In contrast, the SOM and TN values of PF shrub patches, as well as the TK and TP values within all shrub patches, did not display significant differences between the 0–20 cm and 40–60 cm soil layers (p > 0.05).
3.4. Characteristics of soil nutrient distribution in different locations of shrub patches
A total of 36 soil samples were collected for the analysis of soil properties across various shrub types, encompassing soil depths from 0 to 60 cm. The summary of these soil properties, including their probability distribution and notable differences observed among them in different patch microsites, is presented in Table 2. The findings revealed noteworthy changes in Soil Organic Matter (SOM), Total Nitrogen (TN), Total Potassium (TK), and Total Phosphorus (TP), with respective increases of 8.74%, 9.21%, and 17.12%, and a decrease of 7.04%. This alteration in soil nutrient distribution was notably dependent on the location of the shrub patch (Table 2). At the edge of the shrub patches (ES), the mean values for SOM, TN, TK, and TP were recorded as 67.51 g/kg, 0.99 g/kg, 1.06 g/kg, and 0.73 g/kg, respectively. Conversely, at the center of the shrub patch (CS), the mean values for SOM, TN, TK, and TP were notably higher, at 73.97 g/kg, 1.09 g/kg, 1.28 g/kg, and 0.68 g/kg, respectively. This analysis indicates a tendency for SOM, TN, and TK to exhibit an increase at the center of the shrub patches, while TP tended to decrease in concentration at these locations.

Table 2.
The characteristics of soil available nutrients in the 0–60 cm soil profile of patch microsites under four shrub patch types.
Table 2.
The characteristics of soil available nutrients in the 0–60 cm soil profile of patch microsites under four shrub patch types.
| Shrub patch type | Patch microsites | soil depth (cm) | SOM (g/kg) | TN (g/kg) | TK (g/kg) | TP (g/kg) |
| SO | ES | 0—20 | 91.603 | 2.224 | 1.872 | 0.749 |
| 20—40 | 52.842 | 2.163 | 1.444 | 0.490 | ||
| 40—60 | 35.822 | 1.681 | 1.411 | 0.451 | ||
| mean value | 60.09±65a | 2.02±0.17a | 1.58±0.15b | 0.56±0.09a | ||
| BC | 0—20 | 101.541 | 2.666 | 1.993 | 0.610 | |
| 20—40 | 61.453 | 2.147 | 1.703 | 0.515 | ||
| 40—60 | 49.071 | 1.556 | 1.595 | 0.440 | ||
| mean value | 70.69±5.83a | 2.12±0.32a | 1.76±0.12ab | 0.52±0.05a | ||
| CS | 0—20 | 138.960 | 3.182 | 2.188 | 0.526 | |
| 20—40 | 97.297 | 1.929 | 2.010 | 0.411 | ||
| 40—60 | 54.224 | 1.774 | 1.927 | 0.407 | ||
| mean value | 96.83±4.46a | 2.29±0.45a | 2.04±0.08a | 0.45±0.04a | ||
| SA | ES | 0—20 | 45.132 | 1.378 | 1.225 | 0.647 |
| 20—40 | 35.180 | 1.028 | 1.193 | 0.591 | ||
| 40—60 | 21.960 | 0.530 | 0.954 | 0.417 | ||
| mean value | 34.09±6.71a | 0.98±0.24a | 1.12±0.09a | 0.55±0.07a | ||
| BC | 0—20 | 37.154 | 1.338 | 1.143 | 0.558 | |
| 20—40 | 29.853 | 1.276 | 1.122 | 0.358 | ||
| 40—60 | 27.948 | 0.841 | 0.978 | 0.389 | ||
| mean value | 31.65±2.81a | 1.15±0.16a | 1.08±0.05a | 0.43±0.06a | ||
| CS | 0—20 | 53.197 | 1.875 | 1.674 | 0.441 | |
| 20—40 | 37.099 | 1.027 | 1.146 | 0.379 | ||
| 40—60 | 32.680 | 0.639 | 0.894 | 0.191 | ||
| mean value | 40.99±6.23a | 1.18±0.36a | 1.24±0.23a | 0.34±0.08a | ||
| RC | ES | 0—20 | 96.349 | 0.450 | 0.703 | 0.653 |
| 20—40 | 88.560 | 0.555 | 0.748 | 0.670 | ||
| 40—60 | 58.997 | 0.446 | 0.716 | 0.663 | ||
| mean value | 81.30±11.38a | 0.48±0.03a | 0.72±0.01b | 0.66±0.00b | ||
| BC | 0—20 | 113.684 | 0.520 | 0.820 | 0.938 | |
| 20—40 | 100.827 | 0.466 | 0.809 | 0.886 | ||
| 40—60 | 79.332 | 0.497 | 0.864 | 0.794 | ||
| mean value | 97.95±10.02a | 0.49±0.02a | 0.83±0.02a | 0.87±0.04a | ||
| CS | 0—20 | 120.596 | 0.474 | 0.914 | 0.836 | |
| 20—40 | 60.498 | 0.317 | 0.832 | 0.727 | ||
| 40—60 | 51.402 | 0.284 | 0.834 | 0.842 | ||
| mean value | 77.50±7.08a | 0.36±0.06a | 0.86±0.03a | 0.80±0.04a | ||
| PF | ES | 0—20 | 103.082 | 0.579 | 0.900 | 1.319 |
| 20—40 | 92.046 | 0.451 | 0.797 | 0.938 | ||
| 40—60 | 88.549 | 0.448 | 0.784 | 1.159 | ||
| mean value | 94.56±4.38a | 0.49±0.04b | 0.83±0.04b | 1.14±0.11a | ||
| BC | 0—20 | 86.040 | 0.396 | 0.982 | 1.148 | |
| 20—40 | 81.743 | 0.280 | 0.885 | 1.091 | ||
| 40—60 | 73.026 | 0.419 | 0.845 | 0.928 | ||
| mean value | 80.27±3.83a | 0.36±0.04ab | 0.90±0.04ab | 1.06±0.07a | ||
| CS | 0—20 | 97.748 | 0.623 | 1.041 | 1.124 | |
| 20—40 | 70.697 | 0.431 | 1.017 | 1.206 | ||
| 40—60 | 73.282 | 0.588 | 0.902 | 1.082 | ||
| mean value | 73.97±9.57a | 1.09±0.26a | 1.28±0.15a | 0.68±0.10a |
3.5. Characteristics of soil nutrient enrichment rate in shrub patches
We examined the soil nutrient enrichment characteristics across various shrub patches, spanning from the edge of the shrub patch (ES) to the center of the shrub patch (CS), utilizing the RII calculation method (as presented in Table 3). Here's a summary of the observed phenomena: In the SO shrub patch, soil nutrients such as SOM, TN, and TK exhibited an aggregation effect, with SOM demonstrating the highest aggregation effect (0.235) and TN displaying the lowest (0.049). Within the SA shrub patch, SOM, TN, and TK exhibited aggregation effects as well, with the RII values ranking as SOM > TN > TK. In the case of the RC shrub patch, the TK and TP demonstrated clustering effects, with respective clustering indices (RII) of 0.087 and 0.094 as one moves from the edge to the center of the patch. For the PF shrub patch, the TN, TK, and TP displayed aggregation characteristics, with corresponding aggregation indices (RII) of 0.050, 0.088, and 0.004, respectively. Interestingly, an aggregation effect contrary to the general trend was also noted, where soil exhibited an aggregation effect from the edge to the center of the shrub patches. The average RII values from the edge of the shrub to the center of the shrub were found to be -0.105 and -0.260 in the SO and SA shrub patch types, respectively, for TP. Meanwhile, RC displayed average RII values of -0.048 and -0.156 for SOM and TN, respectively. PF in SOM exhibited an average RII value of -0.084. Negative RII values indicated a gradual decrease in soil nutrient content from the edge of the shrub to the center of the shrub patches, signifying a lack of nutrient aggregation in the soil. Conversely, soil nutrients increased from the center of the shrub patch to the edge, reflecting a pattern of soil nutrient accumulation.
We employed the RII calculation method to assess the accumulation patterns of soil nutrients across different soil layers, extending from the edge of the shrub to the center of the shrub (as outlined in Table 3). Our findings revealed variations in RII values for soil nutrients within the same shrub, signifying differences in the soil's nutrient aggregation potential. In the SO shrub patch, both SOM and TK displayed positive RII values across the 0–60 cm soil layer. Notably, the highest RII values for both SOM and TK were observed within the 20–40 cm soil layer. This pattern suggests that the 0–60 cm soil layer, extending from the edge of the SO shrub patch to the center of the patch, exhibited significant nutrient enrichment, particularly within the 20–40 cm soil layer. Across the soil nutrients of the SO, SA, and RC shrub patches, there was an observable trend of aggregation from the edge of the shrub patch to the center. However, for TP in the SO and SA shrub patches, negative RII values indicated an absence of nutrient enrichment from the edge to the center of these shrub patches. This phenomenon was most pronounced within the 0–20 cm and 40–60 cm soil layers. In the SA, RC, and PF shrub patches, the RII values exhibited both positive and negative values within the 0–60 cm range. This indicated that soil nutrients aggregated in the soil layers with positive RII values, while the opposite was true for those with negative RII values, signifying a lack of aggregation within these layers in the research direction.
3.6. The correlation between shrub patch characteristics and soil nutrients
The correlation analysis revealed highly significant relationships between the characteristics of shrub patches and soil nutrients, as depicted in Figure 3. The intensity of the color signifies the strength of the correlation, with darker shades indicating stronger associations and lighter shades suggesting weaker ones. Significant correlations were observed between the characteristics of the shrub patch and the following soil nutrient parameters: SOM, TN, and TP within the center of the shrub patch (CS); SOM beneath the crown (BC); and TK and TP at the edge of the shrub patch (ES) (p < 0.01). Additionally, a significant correlation was identified between the height of the shrub patches and TK beneath the crown (BC), as well as TP within the center of the shrub patches (CS) (p < 0.05). The shoot biomass of the shrub patches exhibited a significant correlation with TN and TP beneath the crown (BC), and TN at the edge of the shrub patches (ES) (p < 0.05). A significant correlation was also established between the root biomass of the shrub patches and TK beneath the crown (BC), and TP within the center of the shrub patches (CS) (p < 0.05). These findings underscore the intricate relationships between shrub patch characteristics and soil nutrient parameters, shedding light on the interconnected dynamics in this ecosystem.
4. Discussion
According to the findings of this study, it can reflect the effect on shrub growth status or properties. The patchy distribution of soil resources generally leads to increased productivity and diversity in water-limited environments [23]. Changes in plant distribution characteristics reflect the effects of environmental factors on plant growth [24]. The species composition and distributional characteristics of alpine shrub plants in the study area reflect the eastern Qilian Mountains' long-term adaptation to a variety of factors, including climate, soil, water, topography, and plant biological characteristics [25,26]. Fan et al. describe the spread of shrubby plant aggregates with shrubby patch characteristics as a long-term evolutionary response to environmental factors [27]. The shrub patches in the study area are mainly a group of shade-tolerant and cold-tolerant shrubs adapted to the high-altitude environment, and their vegetation is mainly characterized by tussocks, mats, and low plants [28]. Under the long-term impact of the environmental conditions of the study area and the biological characteristics of the plants, different shrub patch characteristics have been formed. The result showed that the shrub patch characteristics of the four shrub species (Salix oritrepha, Spiraea alpina, Rhododendron capitatum, and Potentilla fruticosa) were different. S. oritrepha had the highest patch area, height, litter depth, and shoot biomass, and R. capitatum had the highest root biomass (table 1). Such a result suggests that the growth of S. oritrepha was also influenced by patch microsite conditions. The reasons for this may come from several sources. First, the poor palatability of shrubs increases the difficulty of livestock foraging and provides shelter for shrub understory herbs [29]. Second, the shrub canopy can have a significant positive effect on understory herbaceous vegetation by improving the understory microenvironment, cover, biomass, and species diversity by improving the understory microenvironment [30].
Furthermore, shrub patches have higher biomass and litter than their surrounding grassland patches, providing higher soil nutrient inputs [31]. Also, this may potentially be due to their elevation differences, which may lead to differentiation in phenology and growth [32]. S. oritrepha possesses certain traits or adaptations that give it a competitive advantage over other plant species in the ecosystem [15]. These traits could include efficient resource utilization, superior growth rates, or the ability to outcompete other species for light, nutrients, and water. S. oritrepha is well suited to the specific environmental conditions of the study area. It could have adaptations that allow it to thrive in the prevailing climate, soil conditions, or other ecological factors. These adaptations could enhance its growth, reproduction, and overall biomass production. S. oritrepha may have a high reproductive capacity, allowing it to produce abundant offspring and establish larger patches. This could result in a higher overall patch area compared to other species. S. oritrepha has efficient mechanisms for nutrient acquisition and utilization [33]. It could have deep root systems that access nutrients from lower soil layers or symbiotic relationships with beneficial microorganisms that enhance nutrient uptake. This improved nutrient acquisition could contribute to its increased shoot biomass and overall growth. S. oritrepha may be adapted to respond positively to disturbances or changes in the ecosystem [34]. If the study area experiences disturbances such as fire or grazing, S. oritrepha has mechanisms to recover and grow rapidly, leading to increased patch area, height, and biomass. July temperature is a common climate factor driving the growth of S. oritrepha across the latitudinal gradient of the eastern Qilian Mountains. It is assumed that low temperatures drive the onset of alpine shrub growth [35]. Furthermore, warm summer temperatures could benefit new wood tissue formation, and more than half of the ring width was completed in July [35]. This phenomenon reflects the strongest suitability of the S. oritrepha shrub patch type to the natural environment and climatic and soil conditions. In a recent synthesis, summer temperature was found to be the dominant climate variable controlling shrub growth across different taxa and locations in the tundra biome [36]. R. capitatum possesses a well-developed and extensive root system that allows for efficient nutrient and water uptake [37]. Its root architecture includes deep penetrating roots, numerous lateral roots, or a dense root network that facilitates resource acquisition and storage. This robust root system enables R. capitatum to accumulate higher biomass compared to other plant species [38]. R. capitatum has efficient mechanisms for nutrient acquisition and utilization. It possesses traits such as enhanced nutrient absorption capacity, increased root exudation, or symbiotic associations with beneficial microorganisms that enhance nutrient uptake [39]. It out-competes other species for limited resources, including nutrients, water, and space which can cause shrub encroachment. Shrub encroachment can alter the composition and structure of plant communities, potentially leading to changes in biodiversity. Some shrub species may outcompete or shade out other plant species, reducing plant diversity. On the other hand, shrubs can provide new habitat niches for different wildlife species, contributing to increased biodiversity at the animal level. The impacts of shrub encroachment can be both positive and negative. On one hand, shrubs can provide habitat and food sources for wildlife, stabilize slopes, and contribute to soil fertility. However, excessive shrub encroachment can have negative effects on the ecosystem. It may reduce the availability of resources for other plant species, alter water and nutrient cycles, increase the risk of wildfires, and affect the biodiversity of the region. The ability of R. capitatum to effectively utilize resources could result in increased root growth and biomass. These adaptations would enable R. capitatum to access and utilize available nutrients more effectively, leading to increased root biomass. In the Qilian Mountains, summer temperature is also a primary limiting factor for the radial growth of alpine rhododendron shrubs [40]. Shrubs formed larger patches coupled with higher shoot biomass. The interaction of grasses and shrubs may shape the configuration of shrub patches. For example, native grasses could control shrub demography by competing with juvenile shrubs for overlapped water resources in the upper soil layer [41]. Root competition for water in xeric regions might also affect patch size and spacing between them [27].
Erfanzadeh found that shrub type was an essential factor affecting shrub understory herbaceous communities’ productivity and species diversity in semiarid regions [42]. For example, shrub height, leaf area, and canopy structure affect rainfall redistribution [43]. P. fruticosa had the lowest shrub height and shoot biomass. Studies in the northern grasslands of China have shown that shrub encroachment reduces herbaceous abundance and aboveground biomass and that shrub type and climate are dominant factors [44]. According to research, the early growth season soil moisture content influences willow shrub growth variation [45]. P. fruticosa naturally exhibits a compact or dwarf growth habit [46]. Some plant species, including certain shrubs, are genetically predisposed to have shorter heights and smaller overall sizes. This growth habit may result in lower shrub height and shoot biomass compared to species with more vigorous growth patterns. P. fruticosa experiences limitations in accessing essential resources such as light, nutrients, or water. Eastern Qilian Mountain has resource-poor conditions, P. fruticosa must allocate limited resources to various physiological processes, resulting in reduced growth and biomass production. P. fruticosa faces intense competition from other plant species in the ecosystem. The other three species are more efficient at resource acquisition or have traits that provide them with a competitive advantage, resulting in P. fruticosa experiencing restricted access to vital resources. This competition can limit its growth and biomass accumulation. P. fruticosa is more susceptible to environmental stress factors such as drought, extreme temperatures, or poor soil conditions [47]. These stressors can negatively impact its growth and development, leading to reduced shrub height and shoot biomass. Also, P. fruticosa is prone to herbivory or grazing by animals, which hinders its growth and biomass production [48]. Continuous browsing or grazing pressure can limit its ability to recover and allocate resources toward shoot growth. Within the species, there can be natural phenotypic variation. Some individuals or populations of P. fruticosa might inherently exhibit lower heights and shoot biomass compared to others. This variation could be a result of genetic factors, local adaptation, or historical ecological interactions.
Shrub cover often results in soil heterogeneity, especially in nutrient accumulation in shrubland. Many studies have shown that shrubs can form ‘fertile islands’ by accumulating C and N [49]. The effects of shrub encroachment on soil nutrients vary at the landscape level, ranging from positive to negative or neutral [50]. In contrast, some shrubs positively affect their understory soil conditions at the patch level [51]. This study determined the distribution of soil nutrients in different soil depths and the position points of different shrub species. Our study showed that soil nutrients in the shallow (0–20 cm) soil depths were higher than those in the deeper (20–40 cm and 40–60 cm) soil depths (table 2). This may result from organic matter accumulation, biological activity, root activity, leaching and erosion, weathering, and mineralization: organic matter, including decomposed plant and animal material, accumulates near the soil surface. As organic matter decomposes, it releases nutrients into the soil, enriching the shallow soil layers with higher concentrations of nutrients [52]. Over time, the accumulation of organic matter in the topsoil contributes to higher nutrient levels in the shallow soil depths. Most soil microorganisms, including bacteria and fungi, are concentrated in the upper soil layers due to the availability of organic matter and aeration. These microorganisms play a crucial role in nutrient cycling and decomposition processes, releasing nutrients from organic matter and making them available for plant uptake. Consequently, their activity is more prominent in shallow soil depths, leading to higher nutrient concentrations. The majority of plant roots are concentrated in the upper soil layers, primarily in shallow depths of 0–20 cm [29].
Shrubs only develop tap roots in the upper layers of the xeric site and tap roots, lateral roots, and fine roots simultaneously in the upper soil profile under better moisture conditions [53]. So shrubs uptake nutrients from the soil through their roots, and their activity in the shallow soil layers promotes nutrient uptake and accumulation in this region [54]. As a result, nutrient concentrations tend to be higher in the topsoil where root density is greatest. Nutrients are subject to leaching, where water carries dissolved nutrients from the topsoil down into the deeper soil layers. Leaching can lead to lower nutrient concentrations in the deeper soil depths [55]. Additionally, erosion processes can remove nutrient-rich topsoil, further depleting the nutrient content in the deeper soil layers [56]. Soil weathering processes and mineralization of parent materials occur primarily in the upper soil layers [57]. These processes release nutrients from minerals and rocks, contributing to higher nutrient concentrations in shallow soil depths. The differentiation of litter between the shrubs indicates that the additional input of nitrogen led to an intensified microbial turnover of organic matter [58]. The mineralized nitrogen is quickly incorporated by plants thereafter [59]. This model of decomposition is consistent with the desert model of Barnes et al. (2015), who proposed that recently senesced plant material is initially subject to high rates of photodegradation while it is standing dead. At this initial stage, litter decomposes slowly. The soil-litter matrix forms when standing dead plant material falls to the soil surface. Thereafter, the effect of photodegradation declines, and microbial decomposition increases, due at least in part to the erosive effect of wind. Overall rates of decomposition peak due to rapid losses of easily decomposable chemical constituents in the litter [60]. Soil organic matter and plant litter with low carbon and nitrogen ratios tend to decompose faster than those with higher carbon and nitrogen ratios [61]. Another potential influence of shrubs on soils is the effect of size: several studies have shown that large shrubs have much greater effects on the soil than small shrubs due to their greater biomass that returns to the soil [62,63]. The larger shrub may also exude more root exudates than smaller plants [64].
R. capitatum had the largest SOM, while S. alpina had the lowest SOM. The order of content is R. capitatum > P. fruticosa > S. oritrepha > S. alpina. This may be attributed to litter quality, rhizosphere effects, mycorrhizal associations, and microbial activity: R. capitatum produces leaf litter with higher organic matter content or slower decomposition rates compared to the other plant species in the Eastern Qilian Mountains [65]. Leaf litter with higher organic matter content takes longer to decompose, resulting in a greater accumulation of SOM in the soil [66]. The rhizosphere, the soil region influenced by plant roots, plays a significant role in SOM dynamics. R. capitatum has a more extensive root system, higher root exudation rates, or a stronger influence on the soil microbial community compared to the other species [67]. These factors can enhance the microbial decomposition of organic matter, leading to increased SOM levels. It is known to form mycorrhizal associations with beneficial soil fungi. Mycorrhizal fungi contribute to nutrient cycling and organic matter decomposition, potentially leading to higher SOM levels in the presence of Rhododendron capitatum [68]. R. capitatum supports higher microbial biomass and activity in the rhizosphere, resulting in increased decomposition of organic matter and subsequent SOM accumulation [69].
Characteristics of soil nutrient distribution in different locations of shrub patches were analyzed. The result showed that soil organic matter, total nitrogen, and total potassium tended to increase by 8.74%, 9.21%, and 17.12%, respectively, across the three position points (ES, BC, and CS) of the shrub species, ' total phosphorus tended to decrease with 7.04%. The result indicated that shrub species influenced the distribution pattern of soil nutrients, which is in line with the results of Tuomisto et al [70]. There was also an assessment of soil nutrient enrichment from various study area position points. The analysis suggested that the causes of this phenomenon are closely related to the distribution, composition, and biomass of shrubs, nutrient uptake through the root, and the redeposition of plant debris on the soil because of organic residues. Due to the differences in the patch area, height, litter depth, and shoot biomass (Table 1), the soil nutrient enrichment characteristics and the degree of absorption and utilization of nutrients will also differ, indicating an inconsistency in the soil nutrient enrichment rate. One of the most interesting aspects to consider is whether there are positive effects of encroaching shrubs on soil fertility that may lead to the formation of "islands of fertility" [71,72]. The study also found that there were differences in the area, height, litter depth, and shoot and root biomass of shrub patches (Table 1). Therefore, the characteristics of soil nutrient enrichment in different shrub patches are not the same, and the degree of absorption and utilization of different nutrients and nutrient return by shrub vegetation to the soil are also different, leading to the phenomenon that the enrichment rate of different soil nutrients is inconsistent. There may be positive effects of the presence of shrubs that lead to positive feedback utilizing further deposition of nutrients in these shrub islands of fertility, leading to increased shrub growth and reproduction [73].
Soil nutrients in the shrub patches across the position points had enrichment characteristics, i.e., a "fertility island" effect, while some aggregation effects had no "fertility island" effect [74]. This showed that some soil nutrients gradually accumulated in the central position point of the shrub patches, while others had no or were less disturbed by the shrub, which cannot affect the distribution of soil elements and develop aggregation characteristics. This was due to shrub patch characteristics having different degrees of influence on soil nutrients due to the different shrub species, resulting in the formation of soil nutrient aggregation or non-aggregation characteristics. Correlation analysis showed that shrub patch characteristics were closely related to soil nutrient factors (Figure 4). The area, height, litter depth, and biomass of shrub patches had a positive correlation with SOM, TN, and TP in the center of the patch (CS), while SOM below the crown (BC), TK, and TP at the edge of the patch (ES) had a very significant positive correlation (p< 0.01). This phenomenon showed that shrub patch characteristics had the closest interaction between soil nutrients in the center of the shrub patch (CS) and further explained that the mutual adaptation characteristics between shrub plants and soil nutrients were formed under long-term interaction.
5. Conclusions
The concept of fertility islands in shrub patch characteristics and soil nutrient distribution patterns within the Eastern Qilian Mountains highlights the complex interactions between shrubs and their surrounding environments. Our study showed that different shrub patches in the study area were significantly affected by the plant species in the shrub patches and showed great differences. Overall, the patches of the four different species exhibited differing above- and below-ground morphological characteristics (e.g., height, shoot, and root biomass). Soil organic matter (SOM) and macronutrient (N, P, and K) concentrations varied by shrub patch species, soil depth, and distance from the patch center. The soil nutrient content of the shrub patches decreased gradually with the deepening of the soil layer, while the soil nutrient content was significantly higher in the center of the shrub, showing a more significant soil nutrient aggregation phenomenon. The complex interactions between the shrub patch characteristics and soil nutrients investigated in this study make it difficult to determine the level of influence of each factor. A 'relative interaction intensity' variable that compared SOM and macronutrient concentrations from the center to the edges of shrub patches was calculated for each shrub species and at different depths, and the results revealed that the phenomenon of the index of soil nutrient aggregation differs across soil depths of the same shrub patch type. Finally, correlations between shrub patch characteristics and soil nutrients were calculated across all shrub patch types, and the results revealed that there was a highly significant correlation between patch characteristics and soil nutrients at the three patch microsites of the patch types. Their impact on nutrient cycling, vegetation dynamics, and soil properties underscores their significance in understanding ecological processes in these unique landscapes. This study explored the intricate relationships between soil properties, shrub characteristics, and nutrient cycling in the context of alpine ecosystems in the Eastern Qilian Mountains, contributing to our broader understanding of ecosystem dynamics and the development of effective conservation and management strategies.
Author Contributions
Z. J: Conceptualization, Investigation, Methodology, Visualization, Writing – original draft. B. A: Writing – review & editing. W. J: Investigation, Visualization. F. Y: Investigation, Visualization. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by 1. Gansu Province Higher Education Institutions Industrial Support Program Project (2023CYZC-45), 2. National Natural Science Foundation of China (32360378).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ravolainen, V.T.; Bråthen, K.A.; Ims, R.A.; Yoccoz, N.G.; Soininen, E.M. Shrub patch configuration at the landscape scale is related to the diversity of adjacent herbaceous vegetation. Plant Ecology & Diversity 2013, 6, 257–268. [Google Scholar]
- Whitford, W.G.; Duval, B.D. Ecology of desert systems; Academic Press, 2019. [Google Scholar]
- D'Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecology letters 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Y.; Yu, Q.; Lü, X.-T.; Trumbore, S.E.; Yang, J.-J.; Han, X.-G. Impacts of leguminous shrub encroachment on neighboring grasses include transfer of fixed nitrogen. Oecologia 2016, 180, 1213–1222. [Google Scholar] [CrossRef]
- Zhao, L.-X.; Zhang, K.; Siteur, K.; Li, X.-Z.; Liu, Q.-X.; van de Koppel, J. Fairy circles reveal the resilience of self-organized salt marshes. Science Advances 2021, 7, eabe1100. [Google Scholar] [CrossRef] [PubMed]
- Sheffer, E.; von Hardenberg, J.; Yizhaq, H.; Shachak, M.; Meron, E. Emerged or imposed: a theory on the role of physical templates and self-organisation for vegetation patchiness. Ecology letters 2013, 16, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.-Q.; Wang, C.-H.; Chang, L.-L.; Wu, Y.-P.; Li, L.; Jin, Z. Effects of feedback regulation on vegetation patterns in semi-arid environments. Applied Mathematical Modelling 2018, 61, 200–215. [Google Scholar] [CrossRef]
- Vogel, B.; Rostagno, C.M.; Molina, L.; Antilef, M.; La Manna, L. Cushion shrubs encroach subhumid rangelands and form fertility islands along a grazing gradient in Patagonia. Plant and Soil 2022, 475, 623–643. [Google Scholar] [CrossRef]
- Chen, L.; Li, H.; Zhang, P.; Zhao, X.; Zhou, L.; Liu, T.; Hu, H.; Bai, Y.; Shen, H.; Fang, J. Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China. Landscape Ecology 2015, 30, 1627–1641. [Google Scholar] [CrossRef]
- Soliveres, S.; Eldridge, D.J. Do changes in grazing pressure and the degree of shrub encroachment alter the effects of individual shrubs on understorey plant communities and soil function? Functional Ecology 2014, 28, 530–537. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Eldridge, D.J.; Delgado-Baquerizo, M.; Soliveres, S.; Bowker, M.A.; Gross, N.; Le Bagousse-Pinguet, Y.; Quero, J.L.; García-Gómez, M.; Valencia, E. Soil fungal abundance and plant functional traits drive fertile island formation in global drylands. Journal of Ecology 2018, 106, 242–253. [Google Scholar] [CrossRef]
- Berdugo, M.; Soliveres, S.; Kéfi, S.; Maestre, F.T. The interplay between facilitation and habitat type drives spatial vegetation patterns in global drylands. Ecography 2019, 42, 755–767. [Google Scholar] [CrossRef]
- Liu, Z.-w.; Chen, R.-s.; Song, Y.-x.; Han, C.-t. Aboveground biomass and water storage allocation in alpine willow shrubs in the Qilian Mountains in China. Journal of Mountain Science 2015, 12, 207–217. [Google Scholar] [CrossRef]
- Zhang, F.; Jia, W.; Zhu, G.; Zhang, Z.; Shi, Y.; Yang, L.; Xiong, H.; Zhang, M. Using stable isotopes to investigate differences of plant water sources in subalpine habitats. Hydrological Processes 2022, 36, e14518. [Google Scholar] [CrossRef]
- Niu, H.; Lu, X.; Zhang, G.; Sarangi, C. Investigation of water-soluble organic constituents and their spatio-temporal heterogeneity over the Tibetan Plateau. Environmental Pollution 2022, 302, 119093. [Google Scholar] [CrossRef] [PubMed]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Abalori, T.A.; Cao, W.; Weobong, C.A.-A.; Wang, S.; Anning, D.K.; Sam, F.E.; Liu, W.; Wang, W. Spatial variability of soil organic carbon fractions and aggregate stability along an elevation gradient in the alpine meadow grasslands of the Qilian Mountains, China. Chilean journal of agricultural research 2022, 82, 52–64. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, C.; Xu, Z.; Wang, Y.; Peng, H. Effect of vegetation on soil water retention and storage in a semi-arid alpine forest catchment. Journal of arid land 2013, 5, 207–219. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. Methods of soil analysis: Part 3 Chemical methods 1996, 5, 961–1010. [Google Scholar]
- Lynch, J.M.; Barbano, D.M. Kjeldahl nitrogen analysis as a reference method for protein determination in dairy products. Journal of AOAC international 1999, 82, 1389–1398. [Google Scholar] [CrossRef]
- Armas, C.; Ordiales, R.; Pugnaire, F.I. Measuring plant interactions: a new comparative index. Ecology 2004, 85, 2682–2686. [Google Scholar] [CrossRef]
- Titus, J.H.; Nowak, R.S.; Smith, S.D. Soil resource heterogeneity in the Mojave Desert. Journal of Arid Environments 2002, 52, 269–292. [Google Scholar] [CrossRef]
- Zhang, Q.-p.; Wang, J.; Wang, Q. Effects of abiotic factors on plant diversity and species distribution of alpine meadow plants. Ecological Informatics 2021, 61, 101210. [Google Scholar] [CrossRef]
- Chen, L.; Li, H.; Zhang, P.; Zhao, X.; Zhou, L.; Liu, T.; Hu, H.; Bai, Y.; Shen, H.; Fang, J. Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China. Landscape Ecology 2015, 30, 1627–1641. [Google Scholar] [CrossRef]
- Hao, H.-M.; Lu, R.; Liu, Y.; Fang, N.-F.; Wu, G.-L.; Shi, Z.-H. Effects of shrub patch size succession on plant diversity and soil water content in the water-wind erosion crisscross region on the Loess Plateau. Catena 2016, 144, 177–183. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.Y.; Huang, Y.M.; Li, L.; Zhang, J.H.; Liu, Q.; Jiang, Z.Y. Shrub patch configuration in relation to precipitation and soil properties in Northwest China. Ecohydrology 2018, 11, e1916. [Google Scholar] [CrossRef]
- Shi, Y.; Jia, W.; Zhu, G.; Ding, D.; Yuan, R.; Xu, X.; Zhang, Z.; Yang, L.; Xiong, H. Hydrogen and Oxygen Isotope Characteristics of Water and the Recharge Sources in Subalpine of Qilian Mountains, China. Polish Journal of Environmental Studies 2021, 30. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cui, H.-Y.; Jia, B.; Gang, S.; Li, Y.; Li, F.-C.; Mou, X.M.; Li, X.G. Soil sampling depth matters in assessing the impact of shrubification on soil organic carbon storage in grazed alpine meadows. Geoderma 2022, 426, 116119. [Google Scholar] [CrossRef]
- Howard, K.S.; Eldridge, D.J.; Soliveres, S. Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic and Applied Ecology 2012, 13, 159–168. [Google Scholar] [CrossRef]
- Aguirre, D.; Benhumea, A.E.; McLaren, J.R. Shrub encroachment affects tundra ecosystem properties through their living canopy rather than increased litter inputs. Soil Biology and Biochemistry 2021, 153, 108121. [Google Scholar] [CrossRef]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Canadian Journal of Forest Research 2009, 39, 1259–1269. [Google Scholar] [CrossRef]
- Cui, G.; Wei, X.; Degen, A.A.; Wei, X.; Zhou, J.; Ding, L.; Shang, Z.; Liu, S.; Long, R. Trolox-equivalent antioxidant capacity and composition of five alpine plant species growing at different elevations on the Qinghai–Tibetan Plateau. Plant ecology & diversity 2016, 9, 387–396. [Google Scholar]
- Wang, J.; Li, W.; Cao, W.; Abalori, T.A.; Liu, Y.; Xin, Y.; Wang, S.; Zhang, D. Soil bacterial community responses to short-term grazing exclusion in a degraded alpine shrubland–grassland ecotone. Ecological Indicators 2021, 130, 108043. [Google Scholar] [CrossRef]
- Li, X.; Rossi, S.; Liang, E.; Julio Camarero, J. Temperature thresholds for the onset of xylogenesis in alpine shrubs on the Tibetan Plateau. Trees 2016, 30, 2091–2099. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Elmendorf, S.C.; Beck, P.S.; Wilmking, M.; Hallinger, M.; Blok, D.; Tape, K.D.; Rayback, S.A.; Macias-Fauria, M.; Forbes, B.C. Climate sensitivity of shrub growth across the tundra biome. Nature climate change 2015, 5, 887–891. [Google Scholar] [CrossRef]
- Wu, B.; Peng, H.; Sheng, M.; Luo, H.; Wang, X.; Zhang, R.; Xu, F.; Xu, H. Evaluation of phytoremediation potential of native dominant plants and spatial distribution of heavy metals in abandoned mining area in Southwest China. Ecotoxicology and Environmental Safety 2021, 220, 112368. [Google Scholar] [CrossRef] [PubMed]
- Makoto, K.; Kudo, G. Intraspecific differentiation in the root system of Potentilla matsumurae along a snow accumulation gradient in mid-altitude alpine environment. Arctic, Antarctic, and Alpine Research 2020, 52, 408–415. [Google Scholar] [CrossRef]
- Hales, T.C. Modelling biome-scale root reinforcement and slope stability. Earth Surface Processes and Landforms 2018, 43, 2157–2166. [Google Scholar] [CrossRef]
- Lu, X.; Camarero, J.J.; Wang, Y.; Liang, E.; Eckstein, D. Up to 400-year-old Rhododendron shrubs on the southeastern Tibetan Plateau: prospects for shrub-based dendrochronology. Boreas 2015, 44, 760–768. [Google Scholar] [CrossRef]
- d'Onofrio, D.; Baudena, M.; d'Andrea, F.; Rietkerk, M.; Provenzale, A. Tree-grass competition for soil water in arid and semiarid savannas: The role of rainfall intermittency. Water Resources Research 2015, 51, 169–181. [Google Scholar] [CrossRef]
- Erfanzadeh, R.; Yazdani, M.; Arani, A.M. Effect of different shrub species on their sub-canopy soil and vegetation properties in semiarid regions. Land Degradation & Development 2021, 32, 3236–3247. [Google Scholar]
- Yue, K.; De Frenne, P.; Fornara, D.A.; Van Meerbeek, K.; Li, W.; Peng, X.; Ni, X.; Peng, Y.; Wu, F.; Yang, Y. Global patterns and drivers of rainfall partitioning by trees and shrubs. Global change biology 2021, 27, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Shen, H.; Chen, L.; Li, H.; Zhang, P.; Zhao, X.; Liu, T.; Liu, S.; Xing, A.; Hu, H. Ecological consequences of shrub encroachment in the grasslands of northern China. Landscape Ecology 2019, 34, 119–130. [Google Scholar] [CrossRef]
- Dearborn, K.D.; Danby, R.K. Topographic influences on ring widths of trees and shrubs across alpine treelines in southwest Yukon. Arctic, Antarctic, and Alpine Research 2018, 50, e1495445. [Google Scholar] [CrossRef]
- Hrotkó, K. Potentials in Prunus mahaleb L. for cherry rootstock breeding. Scientia Horticulturae 2016, 205, 70–78. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, Z.-h.; Yao, B.-q.; Ma, Z.; Huang, X.-t.; Zhou, B.-r.; Xu, M.-h.; Guo, J.; Zhou, H.-k. Effects of drought and heat on the productivity and photosynthetic characteristics of alpine meadow plants on the Qinghai-Tibetan Plateau. Journal of Mountain Science 2021, 18, 2079–2093. [Google Scholar] [CrossRef]
- Midoko-Iponga, D.; Krug, C.B.; Milton, S.J. Competition and herbivory influence growth and survival of shrubs on old fields: Implications for restoration of renosterveld shrubland. Journal of Vegetation Science 2005, 16, 685–692. [Google Scholar] [CrossRef]
- Li, H.; Shen, H.; Zhou, L.; Zhu, Y.; Chen, L.; Hu, H.; Zhang, P.; Fang, J. Shrub encroachment increases soil carbon and nitrogen stocks in temperate grasslands in China. Land Degradation & Development 2019, 30, 756–767. [Google Scholar]
- Du, B.; Ji, H.; Peng, C.; Liu, X.; Liu, C. Altitudinal patterns of leaf stoichiometry and nutrient resorption in Quercus variabilis in the Baotianman Mountains, China. Plant and Soil 2017, 413, 193–202. [Google Scholar] [CrossRef]
- Ding, L.; Wang, P.; Zhang, W.; Zhang, Y.; Li, S.; Wei, X.; Chen, X.; Zhang, Y.; Yang, F. Shrub encroachment shapes soil nutrient concentration, stoichiometry and carbon storage in an abandoned subalpine Grassland. Sustainability 2019, 11, 1732. [Google Scholar] [CrossRef]
- Armas, C.; Kim, J.H.; Bleby, T.M.; Jackson, R.B. The effect of hydraulic lift on organic matter decomposition, soil nitrogen cycling, and nitrogen acquisition by a grass species. Oecologia 2012, 168, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Mazza, G. Mediterranean pine root systems under drought. Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin 2021, 129–140. [Google Scholar]
- Li, S.-X.; Wang, Z.-H.; Malhi, S.; Li, S.-Q.; Gao, Y.-J.; Tian, X.-H. Nutrient and water management effects on crop production, and nutrient and water use efficiency in dryland areas of China. Advances in agronomy 2009, 102, 223–265. [Google Scholar]
- Lehmann, J.; Schroth, G. Nutrient leaching. In Trees, crops and soil fertility: Concepts and research methods; CABI publishing Wallingford UK, 2002; pp. 151–166. [Google Scholar]
- Uhlig, D.; Amelung, W.; Von Blanckenburg, F. Mineral nutrients sourced in deep regolith sustain long-term nutrition of mountainous temperate forest ecosystems. Global Biogeochemical Cycles 2020, 34, e2019GB006513. [Google Scholar] [CrossRef]
- Kowalska, J.B.; Zaleski, T.; Józefowska, A.; Mazurek, R. Soil formation on calcium carbonate-rich parent material in the outer Carpathian Mountains–A case study. Catena 2019, 174, 436–451. [Google Scholar] [CrossRef]
- Wutzler, T.; Zaehle, S.; Schrumpf, M.; Ahrens, B.; Reichstein, M. Adaptation of microbial resource allocation affects modelled long term soil organic matter and nutrient cycling. Soil Biology and Biochemistry 2017, 115, 322–336. [Google Scholar] [CrossRef]
- Radicetti, E.; Mancinelli, R.; Moscetti, R.; Campiglia, E. Management of winter cover crop residues under different tillage conditions affects nitrogen utilization efficiency and yield of eggplant (Solanum melanogena L.) in Mediterranean environment. Soil and Tillage Research 2016, 155, 329–338. [Google Scholar] [CrossRef]
- Barnes, P.W.; Throop, H.L.; Archer, S.R.; Breshears, D.D.; McCulley, R.L.; Tobler, M.A. Sunlight and soil–litter mixing: drivers of litter decomposition in drylands. Progress in Botany 2015, 76, 273–302. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The M icrobial E fficiency-M atrix S tabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Global change biology 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Thomas, A.D.; Elliott, D.R.; Dougill, A.J.; Stringer, L.C.; Hoon, S.R.; Sen, R. The influence of trees, shrubs, and grasses on microclimate, soil carbon, nitrogen, and CO2 efflux: Potential implications of shrub encroachment for Kalahari rangelands. Land Degradation & Development 2018, 29, 1306–1316. [Google Scholar]
- Ward, D.; Trinogga, J.; Wiegand, K.; du Toit, J.; Okubamichael, D.; Reinsch, S.; Schleicher, J. Large shrubs increase soil nutrients in a semi-arid savanna. Geoderma 2018, 310, 153–162. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: feedback effect of root exudates-rhizobiome interactions. Applied microbiology and biotechnology 2019, 103, 1155–1166. [Google Scholar] [CrossRef]
- Wang, J.; Xu, B.; Wu, Y.; Gao, J.; Shi, F. Flower litters of alpine plants affect soil nitrogen and phosphorus rapidly in the eastern Tibetan Plateau. Biogeosciences 2016, 13, 5619–5631. [Google Scholar] [CrossRef]
- Córdova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biology and Biochemistry 2018, 125, 115–124. [Google Scholar] [CrossRef]
- López-Angulo, J.; de la Cruz, M.; Chacón-Labella, J.; Illuminati, A.; Matesanz, S.; Pescador, D.S.; Pías, B.; Sánchez, A.M.; Escudero, A. The role of root community attributes in predicting soil fungal and bacterial community patterns. New Phytologist 2020, 228, 1070–1082. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Liu, D.; Li, Y.; Liu, P. Soil net N mineralization and hydraulic properties of carbonate-derived laterite under different vegetation types in Karst forests of China. Science of The Total Environment 2023, 856, 159116. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Pei, X.; Peng, S.; Wang, G.; Smoak, J.M.; Duan, B. Litter inputs drive increases in topsoil organic carbon after scrub encroachment in an alpine grassland. Pedobiologia 2021, 85, 150731. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, environment, and floristic variation of western Amazonian forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yan, Y.; Xu, D.; Xu, X.; Wang, C.; Wang, X.; Chen, J.; Xin, X.; Eldridge, D.J. The fertile island effect collapses under extreme overgrazing: evidence from a shrub-encroached grassland. Plant and Soil 2020, 448, 201–212. [Google Scholar] [CrossRef]
- QU, W.-L.; YANG, X.-P.; ZHANG, C.-T.; Wei, B. Shrub-mediated “fertile island” effects in arid and semi-arid grassland. Acta Prataculturae Sinica 2015, 24, 201. [Google Scholar]
- Allington, G.R.; Valone, T.J. Islands of fertility: a byproduct of grazing? Ecosystems 2014, 17, 127–141. [Google Scholar] [CrossRef]
- Bai, Y.; She, W.; Zhang, Y.; Qiao, Y.; Fu, J.; Qin, S. N enrichment, increased precipitation, and the effect of shrubs collectively shape the plant community in a desert ecosystem in northern China. Science of the Total Environment 2020, 716, 135379. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustration of sampling location and design.
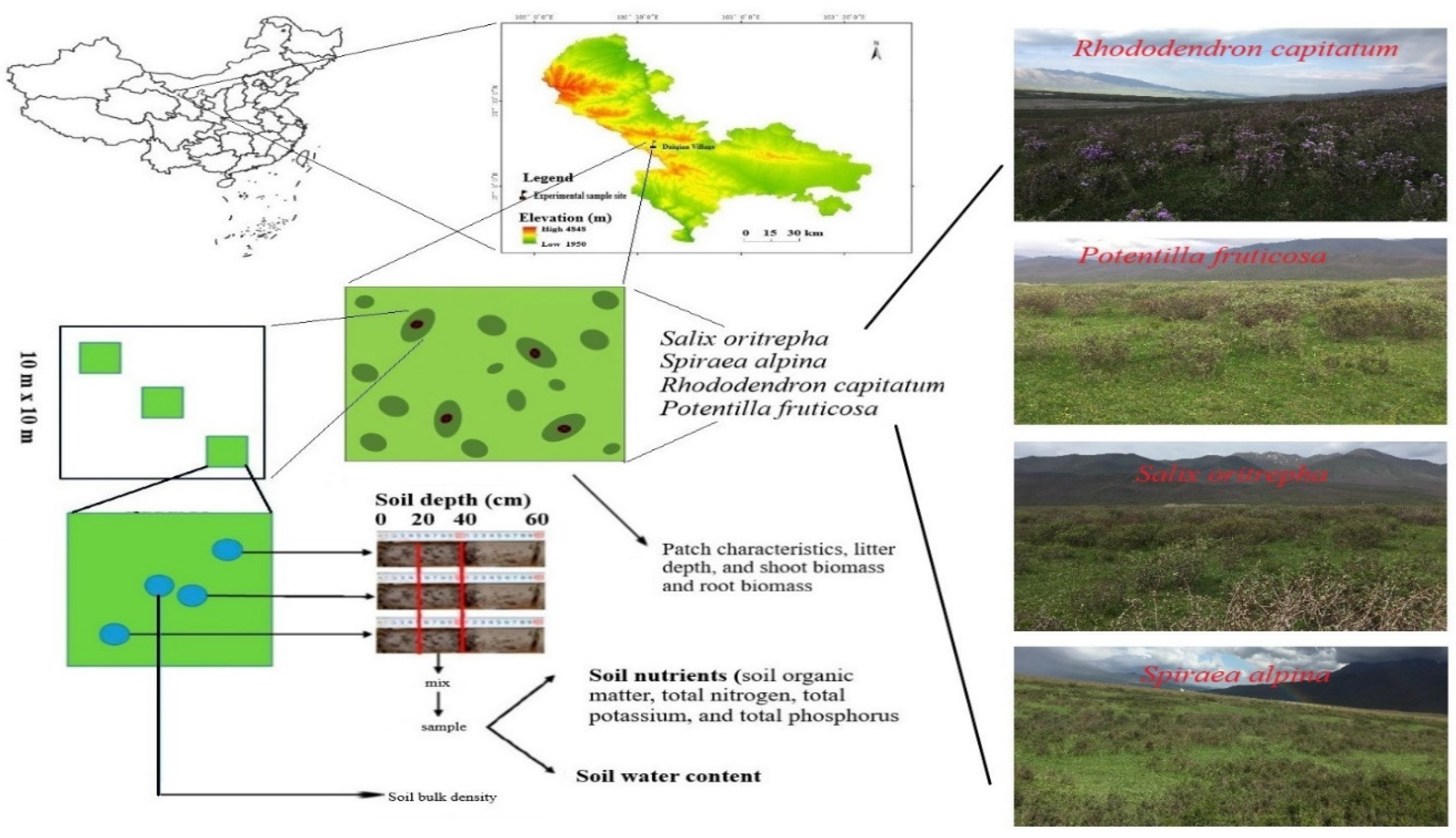
Figure 2.
Trends in SOM, TN, TK, and TP of the four shrub types.
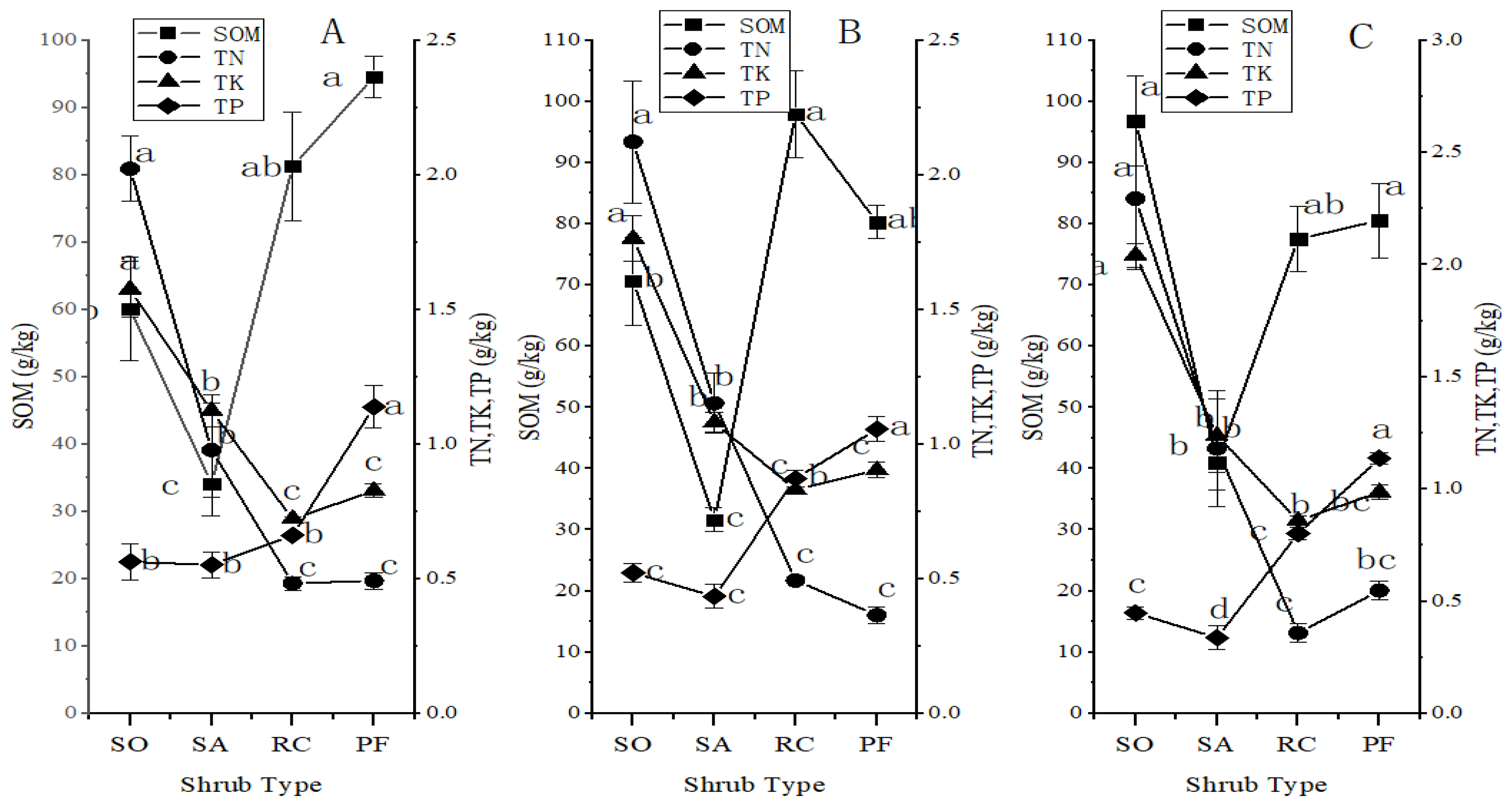
Figure 3.
The correlation between shrub patch characteristics and soil nutrients. (The darker the color, the higher the correlation between the two traits. The lighter the color, the weaker the correlation between the two traits).
Figure 3.
The correlation between shrub patch characteristics and soil nutrients. (The darker the color, the higher the correlation between the two traits. The lighter the color, the weaker the correlation between the two traits).
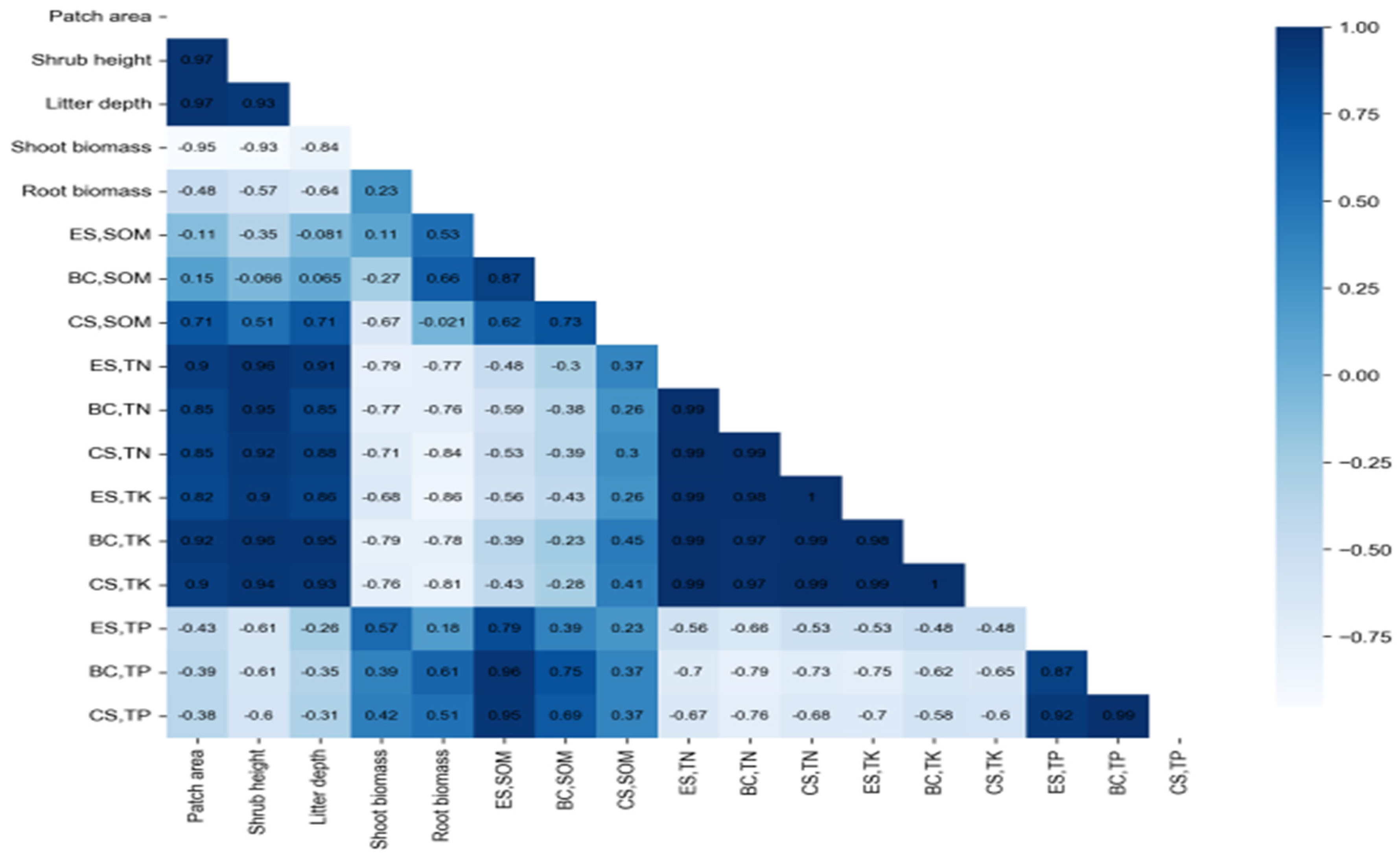
Table 1.
Characteristics of shrub patch. (SO, (Salix oritrepha); SA, (Spiraea alpine); RC, (Rhododendron capitatum) and PF, (Potentilla fruticosa) Mean (± SE).
Table 1.
Characteristics of shrub patch. (SO, (Salix oritrepha); SA, (Spiraea alpine); RC, (Rhododendron capitatum) and PF, (Potentilla fruticosa) Mean (± SE).
| Patch type | Patch area (m2) | Height (cm) | Litter depth (cm) | Shoot biomass (g/m2) | Root biomass (g/m2) |
| SO | 1.66±0.41a | 117.94±3.58a | 3.33±0.03a | 1098.53±64.52a | 2284.29±105.40b |
| SA | 0.08±0.01b | 61.32±3.04b | 0.70±0.04b | 1062.93±94.04a | 2581.55±186.08b |
| RC | 0.37±0.01b | 60.05±1.51b | 0.75±0.01b | 613.68±81.33b | 3983.36±490.31a |
| PF | 0.12±0.01b | 43.90±3.10c | 1.10±0.01b | 579.53±97.15b | 3291.81±264.77a |
Table 3.
The relative interaction intensity (RII) values for SOM, TN, TK, and TP of the soil depths for the four shrub species.
Table 3.
The relative interaction intensity (RII) values for SOM, TN, TK, and TP of the soil depths for the four shrub species.
| Shrub patch type | Soil depth (cm) | SOM (g/kg) | TN (g/kg) | TK (g/kg) | TP(g/kg) |
| SO | 0-20 | 0.205 | 0.177 | 0.078 | -0.175 |
| 20-40 | 0.296 | -0.057 | 0.164 | -0.087 | |
| 40-60 | 0.204 | 0.027 | 0.155 | -0.052 | |
| Mean Value | 0.235 | 0.049 | 0.132 | -0.105 | |
| SA | 0-20 | 0.082 | 0.153 | 0.155 | -0.189 |
| 20-40 | 0.027 | 0.000 | -0.020 | -0.219 | |
| 40-60 | 0.196 | 0.093 | -0.032 | -0.371 | |
| Mean Value | 0.102 | 0.082 | 0.034 | -0.260 | |
| RC | 0-20 | 0.112 | 0.026 | 0.130 | 0.122 |
| 20-40 | -0.188 | -0.273 | 0.053 | 0.041 | |
| 40-60 | -0.069 | -0.222 | 0.076 | 0.119 | |
| Mean Value | -0.048 | -0.156 | 0.087 | 0.094 | |
| PF | 0-20 | -0.027 | 0.037 | 0.073 | -0.080 |
| 20-40 | -0.131 | -0.022 | 0.121 | 0.125 | |
| 40-60 | -0.094 | 0.135 | 0.070 | -0.034 | |
| Mean Value | -0.084 | 0.050 | 0.088 | 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



