
Submitted:
13 December 2024
Posted:
16 December 2024
You are already at the latest version
Abstract
As of the end of 2024, despite the disruptions caused by the coronavirus disease 2019 (COVID-19) pandemic, the global human immunodeficiency virus (HIV) epidemic persists, with 39 million individuals living with the virus and 1.3 million new infections reported in that year. Opportunistic infections associated with acquired immunodeficiency syndrome (AIDS) claimed the lives of over 630,000 people in 2023, contributing to a cumulative death toll exceeding 40.4 million since the disease's emergence in 1981. Although advances in HIV diagnosis and antiretroviral therapy have transformed the disease into a manageable chronic condition, issues like drug resistance, low compliance, and high treatment costs persist. Until a vaccine becomes available, a comprehensive approach involving prevention and therapy is essential, particularly for high-risk populations. Research shows that prevention strategies are more cost-effective than treatment. The development of antiretroviral drugs has also spurred the creation of antivirals for other infections. The emergence of COVID-19 vaccines has opened new avenues for HIV vaccine development, as lessons from vaccine platform technologies like mRNA, viral vectors, and protein subunits can be applied to HIV. This review explores the most promising prophylactic and therapeutic HIV vaccine trials, emphasizing the need for a multi-pronged approach combining vaccines with non-vaccine strategies. Though significant progress is anticipated, the road to a viable HIV vaccine remains long, especially for high-risk populations such as medical workers, blood transfusion recipients, and drug users. The combinatorial approach, capitalizing on various aspects of immunity, may hold the key to defeating HIV in the future.
Keywords:
HIV
; preventive vaccines
; therapeutic vaccines
; AIDS
; vaccine targets
1. Introduction
According to UNAIDS, at the end of 2024, despite the changes brought about by the COVID-19 epidemic, the number of people living with HIV in the world was 39 million. In 2023, the number of new HIV infection cases was more than 1.3 million. More than 630,000 people died from opportunistic infections associated with AIDS in 2022. At the same time, by the end of 2023, more than 40.4 million AIDS patients have died worldwide since the first cases of infection were recorded in 1981 [1,2].
The problem of HIV infection is still far from being resolved. However, the increasingly available HIV serodiagnosis, combined with highly active antiretroviral therapy, has halted the spread of the epidemic and prolonged the lives of infected patients, making the highly lethal disease a chronic one. However, the ongoing treatment does not solve the problems of multiple drug resistance (MDR) formation, low compliance, and high cost [3,4]. More virulence and drug-resistant subtypes and circulating recombinant forms (CRF) of HIV are emerging [5,6]. Moreover, the problem of HIV-tuberculosis (TB) co-infection still remains relevant [7].
Until an effective vaccine is available, a comprehensive approach is needed in the fight against HIV infection and AIDS, using all possible means of therapy and prevention (involving all levels), including microbicides [8,9,10,11,12,13]. Moreover, studies clearly show that HIV prevention in high-risk groups is more cost-effective than therapy [13,14,15].
The development of antiretroviral drugs for the treatment of HIV has influenced the development of other antiviral drugs targeting different viruses. Some HIV drugs have served as a foundation for the development of drugs against other viral infections, e.g Based on HIV nucleoside reverse transcriptase, protease and integrase inhibitors other drugs for the treatment of Hepatitis C (HCV) infection were developed, such as sofosbuvir, telaprevir and bictegravir.
The development of a COVID-19 vaccine opens up opportunities for the development of an HIV vaccine, it can contribute to broader advancements in vaccine development and immunology. The development of COVID-19 vaccines has utilized various vaccine platform technologies, such as mRNA, viral vector, and protein subunit vaccines. These platforms can be adapted and applied to other viral diseases, including HIV. It is noteworthy that in 2023, Dr. Katalin Karikó and Dr. Drew Weissman were awarded the Nobel Prize in physiology or medicine due to their discoveries concerning nucleoside base modifications that eventually enabled the development of mRNA vaccines against COVID-19 [16]. They both hoped that mRNA vaccine technology will help create an effective anti-HIV vaccine. Lessons learned from the development of COVID-19 vaccines can help researchers explore and optimize similar platform technologies for HIV vaccine candidates.
The following comprehensive review covers some of the most exciting trials of prophylactic and therapeutic vaccines against HIV with a detailed description of the mechanism, entering a new milestone in their existence.
2. Structure, Protein Functions, and the Infectious Mechanisms of HIV
HIV, a single-stranded RNA-containing, enveloped, reverse transcribed virus of the Retroviridae family and genus Lentivirus, causes HIV infection, which in its terminal stage progresses to AIDS [17,18].
The subtypes of HIV include: HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 viruses can be further divided into groups M (major), N (new or non-M, non-O), O (outlier), and P. Most of the isolates belong to the M group, in which following subtypes are distinguished: A, B, C, D, F-1, F-2, G, H, J, K. While HIV-1 subtypes N, O, and P as well as HIV-2, have primarily been confined to West Africa, strains of HIV-1 group M have become ubiquitous [17,18,19,20,21,22,23].
The virion is spherical, 80-100 nm in size, covered on the outside with a shell formed during budding from the plasma membrane of the host cell. The envelope contains glycoprotein (gp) spikes consisting of surface gp120 and transmembrane gp41 (cleavage products of gp160 precursor protein). Beneath the shell are the p17 matrix protein, the p24 capsid protein, nucleoproteins (p7, and p9), as well as enzymes (reverse transcriptase - p51 / p66, integrase - p31-32 and protease - p10). The capsid contains two identical plus-strand copies of the RNA genome, which includes the structural genes gag, pol, env, etc. [24]. HIV-2 contains gp140, gp105, gp36, p26 proteins instead of HIV-1 proteins (gp120, gp41, gp160, p24). The structural elements of HIV should be considered as potential targets for vaccine candidates, which will be further discussed in this article (Figure 1).
2.1. HIV-1 Gene-Encoded Proteins and Their Functions
The HIV genome contains structural and regulatory genes. The structural genes include gag, pol, and env, while the regulatory genes can be divided into genes necessary for viral replication (rev, tat) and additional genes (nef, vif, vpr, vpu). A detailed description of the genes, the proteins they encode, and their respective functions is provided in Table 1 [17,25,26,27,28,29].
2.2. Mechanism of Infection
The virus binds via the glycoprotein gp120 to the CD4-T-helper receptor and cells of macrophage origin (macrophages, dendritic and microglial cells). The co-receptors for HIV are CCR5 and CXCR4 chemokine receptors on T-lymphocytes, monocytes, macrophages, Langerhans cells [43,44]. The majority of primary HIV-1 isolates in early disease use CCR5 as a coreceptor, whereas during disease progression with the emergence of syncytium-inducing viruses, CXCR4 is also used [45]. The gp120 comprises constant (C1-C4) and variable (V1-V5) regions, forming nine antigenic epitopes. The variable regions are prone to genetic mutations during the multi-stage replication process of the virus, which accounts for the high variability of HIV. HIV enters the cell by fusion of the envelope with the plasma membrane of the cell through the gp41. After deproteinization of the virion in the cytoplasm of the cell, complementary (proviral) DNA is synthesized on the RNA template using viral RT, which, as part of the preintegration complex, penetrates the nuclear membrane and integrates with the cell DNA using viral IN. The resulting DNA-provirus included in the chromosomal DNA can be latent or serve as the basis for the synthesis of viral genomic RNA and mRNA, which provide the synthesis and assembly of virions. Virions exit the cell by budding: the core of the virus "dresses" in the altered plasma membrane of the cell. If normally immune cells are activated, they begin to translate and transcribe the proteins necessary for the immune response, then in the presence of HIV in the body, infected immune cells facing any infection transcribe and translate new viruses that are cleaved from the cell membrane and infect more cells [17,18,19,20,21,22,46,47].
3. Vaccines Based on the Induction of Neutralizing Antibodies
Research into developing a vaccine against HIV began in 1986. The first trial used the assumption that activating the neutralizing antibodies would protect against a potential HIV infection. The first HIV vaccines were developed using recombinant DNA gene modification technology, which was similar to the hepatitis B vaccine licensed in 1986 [25,48,49]. From the period of 1988 to 2008, various HIV vaccine designs were developed around the viral envelope glycoproteins (mainly gp120 and gp160), which are responsible for binding the virus to the target cells and serve as the primary target for neutralizing antibodies. Among the first to begin testing was VaxSyn (1988-2003) a recombinant envelope glycoprotein (rgpl60) created in the baculovirus-insect cell system. Subsequently, numerous envelope proteins were assessed in additional 35 studies.
Collectively, these vaccine constructs induced binding and neutralizing antibodies, which were long-lasting, and also stimulated CD4+ T cell responses, but did not produce overt cytotoxic lymphocyte (CD8+ CTL) responses [50]. The induction of CTL responses, which was already perceived as a desirable characteristic of a HIV vaccine, could be achieved through the use of live vectors, especially poxvirus vectors [51]. Live vectors were found to be useful because they could simulate natural infection by presenting antigens to the immune system. Two different groups in 1986 reported the expression of the HIV envelope gene in vaccinia vectors [52,53].
One of them was HIVAC-1e (1988), a recombinant vaccinia virus designed to express HIV gp160. In the majority of subjects, the T-cell responses were transient and no antibodies to HIV were detected. However, the appearance of a sustained T-cell response in a small proportion of subjects led researchers to the idea that antibody responses would be enhanced by priming with a recombinant vaccinia vector expressing the HIV-1 envelope and then with an envelope booster protein [54]. VaxSyn + HIVAC-1e (1991) – combined approach. The prime-boosting approach enhanced both the humoral and cellular immune response and induced neutralizing antibodies [55,56]. At this stage, one of the features of the vector used became apparent: there was a marked decrease in the immunogenicity of the vector in individuals previously vaccinated against smallpox [49,54].
Of course, the further refusal to use the cowpox vector was largely influenced by the fact that the introduction of such a vaccine is contraindicated for people who already have immunosuppression, which means that its use is unsafe in groups at high risk of HIV infection [57,58]. It was decided to use non-replicating poxvirus vectors in the early 1990s, based on two models, namely, a highly attenuated strain of vaccinia virus (NYVAC) or an avian poxvirus, canarypox (ALVAC), that is not able to replicate in mammalian cells [51,59]. ALVAC-HIV vector vaccine, an HIV vaccine with the ALVAC vector, vCP125, expressing gp160 has been tested alone or as a combination primary booster with the gp160 subunit. The vaccine significantly stimulated the response of neutralizing antibodies to protein administration and induced CTL activity [60]. Other ALVAC vectors (vCP205, vCP300, vCP1433, vCP1452, and vCP1521) have been designed not only to express the HIV envelope, but also to express gag and other HIV genes to induce a broader cell-mediated immune response. It was vCP1521 that was then used in the most successful trial of RV144 [61,62]. Vax003-Vax004 ‒ (1994-) vaccines based on bivalent recombinant gp120 and alum provided protection to chimpanzees after HIV infection and were safe and immunogenic in phase 1/2 of human clinical trials [63,64]. As a result, high levels of both neutralizing antibodies (nAbs) binding (in VAX003 and VAX004) and antibody-dependent cellular inhibition of the virus (in VAX004) were observed, but both candidates were not effective in preventing HIV infection due to their narrow specificity[63,64].
3.1. Direct Cytotoxicity
Vaccines aimed at stimulating CTLs will cause pre-sensitized CTL to immediately recognize HIV-infected cells and destroy them by the mechanism of direct cytotoxicity, preventing further replication of the virus. The mechanism of the cytotoxic effect is described below and in Figure 2.
CTLs secrete perforin proteins, which are incorporated into the cell membrane and oligomerize therein to form a pore. Next, CTLs secrete granzyme B, which enters the cell through the previously created pore. The cell cytosol contains procaspase 7, which consists of prodomain, large subunit and small subunit. Granzyme B proteolytically detaches the prodomain procaspase from the large subunit and also detaches the large subunit from the small subunit. Two executioner procaspase 7 molecules are required to form an "active" caspase. As a result, the two large and two small procaspase subunits join together to form a heterotetramer, which is active executioner caspase 7. Executioner caspase 7 starts proteolytically destroying the cell proteins leading to cell death.
It is important to note that proapoptotic proteins such as Bax and Bak are constantly present in the cell. Moreover, Bak is built into the outer mitochondrial membrane and Bax is freely present in the cell cytoplasm. The Bax molecule oligomerizes with Bak on the outer mitochondrial membrane forming a large pore in it through which cytochrome c escapes from the intermembrane space into the cell cytosol and triggers apoptosis. However, in the normal cell cytoplasm there are anti apoptotic proteins such as Bcl2 and BCl-XL, which bind to Bax and Bak preventing their oligomerization. Granzyme B proteolytically destroys Bcl2 and BCl-XL proteins and thus creates conditions for cytochrome c release into the cell cytosol [65].
The cell cytosol contains Apoptotic protease activating factor 1 (APAF1) which has a caspase recruitment domain (CARD) that binds to the active center of APAF1 itself and blocks it. But cytochrome c, once in the cytosol of the cell, pushes CARD from the active center of APAF1 and binds itself to it instead of CARD. Further, about 7 APAF1 molecules with active centers bound by cytochrome c bind to each other using their own CARD domains, forming the apoptosome. Further, CARDs associated with the prodomain of seven initiator procaspase 9, interact with the CARDs that are part of the apoptosome. As a result of this interaction, the small subunit detaches from the large subunit of procaspase 9, and the large subunit also detaches from the prodomain. From there, a heterotetramer consisting of two large and two small subunits is assembled with the formation of the "active" initiator caspase 9, which itself will start proteolytically activating Caspase 3 and Caspase 6, which are the executioner caspases. Caspase 3 and Caspase 6 produce proteolysis of the cell proteins, which leads to cell death [66].
3.2. Chemokine-Mediated HIV Suppression
Chemokine based vaccines, especially CCL4 based vaccines, will lead to CCR5 desensitization. This would result in HIV having no entry gate for T-helper cells to be infected. Although this type of vaccines will not protect somatic cells from infection, they will prevent the development of AIDS. The mechanism of CCR5 desensitization is described below and in Figure 3.
Being a ligand for CCR5, CCL4 in high amounts can overstimulate CCR5, that will lead to the phosphorylation of CCR5 by CD28-induced G-protein-coupled receptor kinase 2 (GRK2). β-arrestin binds to the phosphorylated inorganic phosphate molecules. Then, β-arrestin changes its conformation and releases a site for binding to the Adaptor protein complex 2 (AP-2 complex). However, this is not enough for the AP-2 complex to join the β-arrestin. The AP-2 complex consists of four proteins called adaptins: two large adaptins (α and β), a medium adaptin (μ), and a small adaptin (σ). The α and β adaptins have so-called 'ears' that form the appendage domain. The remaining part of α and β adaptins, as well as μ and σ adaptins together form the core domain. In general, the AP-2 complex has one core domain and two appendage domains. The core domain binds to synaptotagmin ½, which is embedded in the cell membrane. It is important to know that this interaction is stable only if PIP2 is present in the cell membrane with which the AP-2 complex also binds. After the previously mentioned interactions, the AP-2 complex changes its conformation and is activated, which leads to the interaction of μ-adaptin with β-arrestin.
The cell cytosol contains clathrin protein, which consists of a heavy chain comprising three domains: proximal, distal, and N-terminal, and a light chain bound to the proximal domain of the heavy chain. Clathrin is not found as a monomer in the cell cytosol. The three clathrin molecules bind to each other using the proximal heavy chain domain to form a claritine triskelion, which is present in the cell cytosol. The claritine triskelion then binds to the appendage domains of the AP-2 complex, which will lead to polymerization of the triskelion, forming a closed three-dimensional network resembling a soccer ball. As a result of these processes, the plasma membrane is woven inward to form the Clatrin coated vesicle bud [67,68].
The cell requires Ca2+ for normal clathrin-dependent endocytosis. Ca2+ ions bind to calmodulin, forming the calmodulin/4Ca2+ complex. In turn, the calmodulin/4Ca2+ complex binds to calcineurin activating it. In the cytoplasm of the cell there is a protein dynamin 1 with which an inorganic phosphate molecule is bound and inhibiting it. Calcineurin dephosphorylates dynamin 1 and thereby activates it. Once activated, dynamin 1 binds to the "neck" of the Clatrin coated vesicle bud and oligomerizes, covering it completely. dynamin 1 has GTPase activity and having enveloped the "neck" of the clathrin coated vesicle bud it begins to hydrolyze GTP to GDP and inorganic phosphate, releasing energy which is used to enhance the torsion of oligomerized dynamin 1 around the "neck", narrowing and eventually cutting it, forming the clathrin coated endocytic vesicle. In the process of cutting the neck, endophilin also plays an important role, which is normally in the homodimerized state and also binds to the "neck" and then polymerizes around it in the form of a ring. This happens because endophilin has "+" charged amino acids, which are attracted to the "-" charged heads of the cell membrane phospholipids.
Phosphorylation of CCR5 by GRK2. β-arrestin binds with the inorganic phosphate molecules of CCR5. Then AP-2 complex binds with β-arrestin. To form a stable complex, AP-2 interacts with SYT and PIP2, embedded in the plasma membrane. Three clatrin molecules bind to each other and form clatrin triskelion. The clatrin triskelion then binds to the AP-2 complex. As a result of these processes, the plasma membrane is woven inward to form the Clatrin coated vesicle bud. Ca2+ ions bind to calmodulin, forming the calmodulin/4Ca2+ complex. The calmodulin/4Ca2+ complex binds to calcineurin and activates it. Calcineurin dephosphorylates dynamin 1 and thereby activates it. Once activated, dynamin 1 binds to the "neck" of the Clatrin coated vesicle bud and oligomerizes, covering it completely. Having enveloped the "neck" of the clathrin coated vesicle bud dynamin 1 begins to hydrolyze GTP to GDP and inorganic phosphate, releasing energy which is used to enhance the torsion of oligomerized dynamin 1 around the "neck", narrowing and eventually cutting it, forming the clathrin coated endocytic vesicle. Endophilin also plays an important role in forming the clathrin coated endocytic vesicle [69,70].
4. Stimulation of T-Cell Immune
Further study of the problem and analysis of the unsatisfactory results of previous years of research prompted scientists to use the T-cell immune response. In the corresponding period of time, the degree of knowledge of the virus also increased. The leading role of CD8+ T-cells in the containment of virus replication at one of the stages of its development in the body has been proven.
Subsequent studies have been conducted with increasing reliance specifically (and often, only) on the T-response [71,72]. Candidate model vaccine vectors used for T-cell vaccines were live recombinant viral vectors, mainly pox and adenovirus vectors (particularly replication-defective adenovirus 5 (Ad5)), as well as DNA vaccines [73].
STEP trial - HVTN 502 – HVTN 503 (Phambili) - recombinant replication-defective adenovirus type 5 (Ad5) vector vaccines: MRKAd5 HIV-1 gag/pol/nef clade B vaccine. Both trials were primarily aimed at reducing post-infection viral load, but were interrupted early because those who were vaccinated had an increased risk of infection [49,74,75].
HVTN 505 - DNA vaccine expressing clade B gag/pol/nef and env from branches A, B and C, which was used to prime rAd5 multiclade vaccines. The goal was to activate both arms of immunity. An increased risk of infection was not observed, but the vaccination schedule was not shown to be effective [76,77,78].
RV-144 - the study design consisted of a recombinant canarypox vector vaccine, ALVAC-HIV (vCP1521), expressing Env (branch E), group specific antigen (Gag) (branch B) and protease (Pro) (branch B), and AIDSVAX B vaccines /E with an alum adjuvant and a bivalent subunit vaccine based on HIV glycoprotein 120 (gp120). The vaccine showed significant effectiveness in protecting against HIV infection - 31.2% in 42 months after vaccination. Vaccine-induced reactions included binding of IgG antibodies to HIV Env variable loops 1 and 2 (V1V2) and antibody-dependent cellular cytotoxicity (ADCC) in low-IgA vaccine recipients. The study showed that the size and multifunctionality of Env-specific CD4+ T cells play a role in reducing the risk of HIV infection [79,80]. RV305- RV306- Evaluation of the immune response generated in recent booster vaccine recipients compared to RV144 vaccine recipients. The assumption was that late boosters will elicit a sustained immune response [80,81].
HVTN 097 - the schedule included 2 doses of ALVAC-HIV (vCP1521) HIV vaccine followed by 2 booster doses of AIDSVAX B/E. Tetanus immunization was included in the schedule to avoid cross-reaction to the vaccine [82]. The primary booster schedule induced mainly Env-specific CD4+ T cell responses at significantly higher levels compared to RV144 vaccine recipients (RV144 = 36.4%; HVTN 097 = 51.9%). IgG antibodies recognizing the V1V2 region and IgG3 binding antibody responses to gp120 and V1V2 antigens were also significantly higher among HVTN 097 vaccine recipients compared to RV144 recipients. ADCC antibody responses were also higher in HVTN 097 than in RV144 [82].
HVTN 100 is an ALVAC-HIV vector (vCP2438) expressing HIV gp120 Env (C branch of ZM96), gp41 Env, Gag and Pro (all B branches), and the bivalent protein vaccine with gp120 subtype C and MF59 adjuvant. The vaccine induced a greater frequency of IgG3 responses to Env gp120, significantly higher CD4+ T cell responses, and gp120 binding antibody responses compared to the RV144 schedule [83]. HVTN 100 exceeded levels that were modeled as necessary for protection.
HVTN 702 - evaluation of the efficacy, safety and tolerability of ALVAC-HIV (vCP2438) plus a bivalent gp120/MF59 subtype C vaccination regimen with primary booster. The vaccination schedule used in HVTN 702 has been modified to improve the efficiency and robustness of immune responses compared to RV144. The study was terminated due to the low efficacy of the vaccine [83,84].
5. Mosaic HIV Vaccines
Polyvalent HIV-1 Gag, Pol and Env mosaic immunogens expressed by replication-incompetent adenovirus serotype 26 (Ad26) vector and optimized to provide maximum coverage of potential T-cell epitopes. The mosaic HIV vaccine causes the formation of multifunctional antibodies - neutralizing, non-neutralizing and binding. This approach solves the problem of narrow specificity of the candidate vaccine and affects different circulating virus isolates [90]. However, large clinical studies of mosaic HIV vaccines have not shown their effectiveness, although some of these studies are currently underway (Table 3).
6. mRNA HIV Vaccine
mRNA platforms are designed to deliver a piece of genetic material that instructs the body to make a protein fragment of a target pathogen (such as HIV), which the immune system will hopefully recognize and mount a defense against [103].
mRNA technology could be important in accelerating the identification of appropriate target antigens for a protective response. However, it does not resolve other challenges in HIV vaccine development, such as determining the correct antigen. B-cells play an important role in the action of these vaccines, which implement their immune response through the BCR. The mechanism of mRNA HIV vaccine is discussed below and in Figure 4.
B cell antigen receptor (BCR) is embedded in the membrane of B cells. The BCR is composed of membrane immunoglobulin (mIg) molecules and associated Igα/Igβ (CD79a/CD79b) heterodimers (α/β). The mIg subunits bind antigen, resulting in receptor aggregation, while the α/β subunits transduce signals to the cell interior. BCR aggregation rapidly activates the Src family of Lyn kinases. The cytoplasmic part of CD79a/b molecules contains immunoreceptor tyrosine-based activation motifs (ITAMs). The Lyn protein, which phosphorylates CD79a/b by the OH groups of the two tyrosine residues that make up the ITAMs, is embedded in the membrane of B cells. The Syk protein binds to the phosphorylated tyrosine residues using SRC homology 2 (SH2) domain, after which it phosphorylates itself by the OH groups of tyrosine (autophosphorylation), leading to its own activation. It is important to note that the Lyn protein can also activate Syk by phosphorylation [104].
The adaptor protein BLNK (B-cell linker protein) binds to the phosphorylated tyrosine residues of CD79a/b using the SH2 domain. Next, Syk phosphorylates the BLNK molecule by the tyrosine OH-group, facilitating the binding of phospholipase Cγ2 (PLCγ2) to it. But this is not enough to activate PLCγ2. The fact is that the Bruton's tyrosine kinase (BTK) molecule binds to the phosphorylated tyrosine residues of BLNK protein using its SH2 domain, after which BTK binds to phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which is a phospholipid of the inner layer of the cell membrane using pleckstrin homology (PH) domain. The interaction of the PH domain with PIP3 leads to full activation of BTK protein. As a result, BTK phosphorylates and thereby activates PLCγ2. PLCγ2 begins to hydrolyze PIP2 (phosphotidylinositol diphosphate), which is a typical representative of cell membrane phospholipids, to inositol triphosphate (phospholipid head) and diacylglycerol (phospholipid tail). DAG remains in the cell membrane. IP3 binds to IP3 receptors located on the ER membrane and stimulates them. It is important to note that IP3 receptors are Ca2+ channels and that after interaction with IP3 they open, promoting the release of large amounts of Ca2+ ions from the ER into the cell cytosol along a concentration gradient. Ca2+ ions and DAG together activate PKCβ (Proteinkinase Cβ). In turn, Ca2+ binds to calmodulin, forming the calmodulin/4Ca2+ complex. In the quiescent state, the cell cytoplasm contains the molecule Nuclear factor of activated T-cells (NFAT), which is in a phosphorylated state and is a transcription factor. As long as NFAT is in the phosphorylated state it cannot pass into the nucleoplasm. The calmodulin/4Ca2+ complex binds to the protein calcineurine resulting in its activation. Calcineurine, a protein phosphatase, dephosphorylates NFAT and thereby changes its conformation, leading to the availability of nuclear localization signals (NLS) and transport of this transcription factor into the nucleoplasm [105].
The cell cytosol contains a caspase recruitment domain family member 11 (CARD11 or CARMA1), which is in the inactive, "closed" state. PKCβ phosphorylates CARMA1 by the serine/threonine OH-group and thereby promotes a change in the conformation of this molecule and its transition from the "closed" to the "open" state. There is a BCL10-MALT1 heterodimer (B cell CLL/lymphoma 10-MALT1 paracaspase heterodimer) in the cytoplasm. After CARMA1 is in the "open" state, it is bound to BCL10 molecule through caspase recruitment domain (CARD)-CARD domain interactions, which is a part of BCL10-MALT1 heterodimer, forming CBM signalosome complex (The caspase recruitment domain family member 11 (CARD11 or CARMA1)-B cell CLL/lymphoma 10 (BCL10)-MALT1 paracaspase (MALT1)). Once this complex is formed, the MALT1 molecule begins to oligomerize, leading to the recruitment of tumor necrosis factor receptor-associated factor 6 (TRAF6) protein, which is an E3 ubiquitin ligase. The cell cytoplasm contains the molecule IKK (IκB kinase), which consists of three subunits: IKK-α, IKK-β, and IKK-γ (NEMO-NF-kappa-B essential modulator). TRAF6 modifies IKKγ with Lys63-polyUb chains, recruiting the kinase TAK1 through its ubiquitin-binding cofactors TAB2 and TAB3. TAK1 then phosphorylates IKK-β, leading to activation of the IKK molecule. It is important to mention that the B-cell cytosol contains the NF-κb molecule, which consists of three subunits: IkBα (Inhibitor of Nuclear Factor (NF)-Κb α isoform), p50, and RelA. In the resting state, IkBα masks the NLS (nuclear localization sequence) of p50 and RelA proteins, preventing transport of these subunits as a heterodimer from the cell cytosol into the nucleoplasm. However, upon activation, IKK starts phosphorylating the IkBα subunit by the OH-groups of the serine/threonine residues. Further, the enzyme E3 ubiquitin ligase binds to the phosphorylated serine/threonine residues, promoting ubiquitination and further proteosomal degradation of IkBα. As a result, NLS is unmasked and the p50/RelA heterodimer is transferred from the cell cytoplasm into the nucleoplasm [106].
The B-cell membrane contains a CD19 protein, the intracellular part of which is phosphorylated by Lyn protein by the OH-group of tyrosine. The phosphorylated tyrosine residues of CD19 are bound to the BCAP (B cell adaptor molecule for PI3K) molecule, which recruits the p85 domain of PI3K protein to itself. This interaction leads to the activation of the p110 domain of PI3K, which with its catalytic activity phosphorylates phosphatidylinositol-4,5-bisphosphate (PIP2) to produce and accumulate phosphatidylinositol-3,4,5-trisphosphate (PIP3). On the one hand, PIP3 is important for binding the BTK molecule to the inner surface of the cell membrane, and on the other hand, it is important for the activation of the Akt/mTOR pathway.
BCR aggregation rapidly activates the Src family of Lyn kinases. The Lyn protein phosphorylates CD79a/b by the OH groups of the two tyrosine residues. The Syk protein binds to the phosphorylated tyrosine residues using SH2 domain, after which it phosphorylates itself by the OH groups of tyrosine (autophosphorylation), leading to its own activation. The adaptor protein BLNK binds to the phosphorylated tyrosine residues of CD79a/b using the SH2 domain. Syk phosphorylates the BLNK molecule, facilitating the binding of PLCγ2 to it. BTK molecule binds to the phosphorylated tyrosine residues of BLNK protein using its SH2 domain, after which BTK binds to PIP3 using the PH domain. As a result, BTK phosphorylates and thereby activates PLCγ2. PLCγ2 begins to hydrolyze PIP2 to IP3 and DAG. IP3 contributes to the output of Ca2+ from the ER. Ca2+ ions and DAG together activate PKCβ. Ca2+ binds to calmodulin, forming the calmodulin/4Ca2+ complex. The calmodulin/4Ca2+ complex binds to the protein calcineurin resulting in its activation. Calcineurin, a protein phosphatase, dephosphorylates NFAT and thereby leads to the transport of this transcription factor into the nucleoplasm. PKCβ phosphorylates CARMA1. CARMA1 binds to BCL-10 molecule, which is a part of BCL-10-MALT1 heterodimer, forming a CBM signalosome complex. Once this complex is formed, the MALT1 molecule begins to oligomerize, leading to the recruitment of TRAF6 protein. TRAF6 modifies IKKγ with Lys63-polyUb chains, recruiting the kinase TAK1 through its ubiquitin-binding cofactors TAB2 and TAB3. TAK1 then phosphorylates IKK-β, leading to activation of the IKK molecule. IKK starts phosphorylating the IkBα subunit of NF-κb. Further, the enzyme E3 ubiquitin ligase binds to the phosphate groups of IkBα, promoting ubiquitination and further proteosomal degradation of IkBα. As a result, NLS is unmasked and the p50/RelA heterodimer is transferred from the cell cytoplasm into the nucleoplasm. Intracellular part of CD19 protein is phosphorylated by Lyn. With that phosphate groups BCAP protein binds. BCAP recruits PI3K, which phosphorylates PIP2 to PIP3. On the one hand, PIP3 is important for binding the BTK molecule to the inner surface of the cell membrane, and on the other hand, it is important for the activation of the Akt/mTOR pathway [104,107].
The Table 4 below summarizes ongoing vaccine trials on the mRNA platform. The difference is in the antigens, the viral proteins that the immune system targets.
7. Dendritic Cell-Based HIV Vaccines
One of the problems of our immune system's fight against HIV is the reduced ability of professional antigen-presenting cells to present HIV antigens on their surface. This approach consists in using professional antigen presenting cells (dendritic) to activate T cells, which, in turn, are an important step in specific immune activity in both innate and adaptive pathways. To achieve this goal, antigens are targeted to dendritic cells using different strategies. The most serious trials of dendritic cell-based HIV vaccines are presented in Table 5. Most studies have been conducted on autologous DCs [111,112,113].
The great advantage of these vaccines is the activation of CTLs against HIV-infected cells, which fulfills a therapeutic role. The mechanism of CTLs activation by dendritic cells is described below and in Figure 5.
CD8+ T-cells activation plays an important role here. The CD8 glycoprotein is embedded in the Cytotoxic T-cells (CTL) membrane and its cytoplasmic part is reacted by lymphocyte-specific protein tyrosine kinase (Lck). Phosphoprotein associated with glycosphingolipid-enriched microdomains 1 (PAG1) is also embedded in the cell membrane. PAG1 is an adaptor protein whose cytoplasmic part contains inorganic phosphate molecules. C-terminal Src kinase (CSK) interacts with the previously mentioned inorganic phosphate molecules of PAG1. Thus, PAG1 brings CSK in close proximity to the Lck molecule. CSK aids to limit Lck activity via Tyr505 phosphorylation. However, it is important to note that the Protein tyrosine phosphatase, receptor type, C (PTPRC) molecule also known as CD45 is embedded in the cell membrane. After CD8 interacts with the MHC-1 molecule on the APC membrane, CD45 is activated and begins to dephosphorylate the cytoplasmic part of PAG1, resulting in the uncoupling of CSK from the PAG1 molecule. This mechanism results in the distancing of the CSK from the cell membrane and its inability to phosphorylate and inactivate the Lck. As a result, Lck is activated and phosphorylates the immunoreceptor tyrosine-based activation motifs (ITAMs) of the CD3 (cluster of differentiation 3) by the OH group of tyrosine. This creates the opportunity for the SH2 domain of the ZAP-70 protein (Zeta-chain-associated protein kinase 70) to bind to the phosphorylated tyrosine residues. Next, Lck phosphorylates ZAP-70 by the OH-group of tyrosine, which leads to a change in its conformation and activation of the kinase domain [118,119].
The linker for activation of T cells (LAT) protein is built into the membrane of CD8+ T- cells and is a target for ZAP-70 molecule, which phosphorylates LAT on multiple tyrosine residues. The inorganic phosphates on LAT protein serve as docking sites for SH2 domains of GRB2-related adaptor protein (GADS), growth-factor receptor-bound protein 2 (GRB2), and phospholipase Cγ1 (PLCγ1).
The SH3 domain of the GRB2 is bound to the proline-rich region of the Son of Sevenless (SOS) protein, which belongs to guanine nucleotide exchange factors (GEFs). In the quiescent state in the cell cytoplasm, there is RAS protein bound to GDP-inactive state. SOS detaches the GDP molecule from RAS and attaches GTP in its place-active state. Then RAS-GTP goes to the RAF-1 protein (proto-oncogene serine/threonine-protein kinase) and activates it. In turn, RAF-1 phosphorylates and thereby activates the enzyme MEK (MAPK/ERK kinase). MEK phosphorylates the mitogen-activated protein kinase (MAPK) by the OH group of serine/threonine (mitogen-activated protein kinase) and thereby activates it. Ultimately, MAPK phosphorylates and activates specific transcription factors that increase the expression of genes responsible for cytokine synthesis [120].
The SH3-domain of the GADS protein binds to the SH2-domain-containing leukocyte protein 76 kDa (SLP76), then ZAP-70 phosphorylates SLP-76 at critical tyrosine residues. This creates binding sites for the SH2 domain of VAV and ITK (interleukin-2-inducible T-cell kinase). In turn, VAV, being GEF, serves the same function as SOS-MAPK activation.
ZAP-70 phosphorylates ITK at tyrosine residues, which leads to its activation. As we mentioned above, the inorganic phosphate on the LAT protein serves as a docking site for the SH2 domain of PLCγ1. But this is not enough to fully activate PLCγ1. ITK phosphorylates PLCγ1 at tyrosine residues, which leads to its full activation. PLCγ1 activates the inositol triphosphate system, leading to DAG formation and increased Ca2+ concentration in the cell. Ca2+ by the already known mechanism activates the NFAT pathway. DAG activates and recruits the guanyl nucleotide-releasing protein RAS (RASGRP1) into the plasma membrane. RASGRP1, like SOS, is the GEF responsible for RAS activation in T cells. RAS, in turn, as we mentioned, activates the MAPK kinase pathway. DAG, together with Ca2+ ions, activates PKCθ, which acts in the same way as PKCβ in B cells. This will lead to the activation of the NF-κb pathway [118,119].
Cytoplasmic part of CD8 glycoprotein binds with Lck. CSK interacts with PAG1. Thus, PAG1 brings CSK in close proximity to the Lck molecule. CSK aids to limit Lck activity via Tyr505 phosphorylation. After CD8 interacts with the MHC-1 molecule on the APC membrane, CD45 is activated and begins to dephosphorylate the cytoplasmic part of PAG1, resulting in the uncoupling of CSK from the PAG1 molecule. As a result, Lck is activated and phosphorylates the ITAMs of the CD3. Next, Lck phosphorylates and activates ZAP-70. ZAP-70 molecule phosphorylates LAT on multiple tyrosine residues. LAT protein serves as a docking site for GADS, GRB2 and PLCγ1 molecules. The SH3 domain of the GRB2 is bound to SOS. SOS converts RAS-GDP to RAS-GTP. Then RAS-GTP activates RAF-1 protein. RAF-1 activates MEK and MEK activates MAPK. Ultimately, MAPK phosphorylates and activates specific transcription factors that increase the expression of genes responsible for cytokine synthesis. GADS protein binds to the SLP-76,then ZAP-70 phosphorylates SLP-76. This creates binding sites for VAV and ITK. In turn, VAV, being GEF, serves the same function as SOS-MAPK activation. ZAP-70 phosphorylates ITK. ITK phosphorylates PLCγ1 at tyrosine residues, which leads to its full activation. PLCγ1 activates the inositol triphosphate system. Ca2+ activates the NFAT pathway, DAG activates and recruits the RASGRP1 into the plasma membrane. RASGRP1, like SOS, is the GEF responsible for RAS activation in T cells. DAG, together with Ca2+ ions, activates PKCθ [118,119].
8. Peptide-Based HIV Vaccines
The approach is to use peptides to evoke a cellular immune response against the most immunogenic or conserved regions of the target protein. This response can be elicited by naturally occurring sub-immunodominant epitopes, by employing a multi-epitope approach that allows multiple strains and different stages of the virus life cycle to be targeted at once [121,122,123,124]. The most serious trials of peptide-based HIV vaccines are presented in Table 6.
9. DNA-Based HIV Vaccines
This approach allows both humoral and cellular immune responses to be elicited using small DNA fragments encoding the antigen (plasmids). Expression of the inserted gene of interest can be controlled using a strong mammalian promoter that can be found on a plasmid backbone of bacterial DNA. When target cells are transfected with DNA vaccines, the encoded proteins will be presented in the major histocompatibility complex (MHC) [130,131,132,133,134]. The most serious trials of DNA-based HIV vaccines are presented in Table 7.
The mechanism of DNA-based HIV vaccines is described in Figure 6. These vaccines are important because they enhance the CD4+T-cell response. The fact is that MHC-II receptors with the antigen epitope are built into the membrane of the dendritic cell. Dendritic cells are approached by naive T-helpers (T0-helpers) whose membrane contains TCR and CD4. CD4 binds to MHC-II receptors, and TCR is necessary to recognize the antigen that is expressed on MHC-II. Once the antigen is recognized, the TCR transmits the signal to the T0-helper (primary signal) via CD3. Furthermore, on the surface of the T0-helper there is CD28, with which CD80/CD86, located on the surface of the dendritic cell, bind and also activate the T0-helper (secondary signal-activation). In addition, the dendritic cell secretes IL-1, which binds to IL-1R, which is on the surface of the T0-helper (tertiary activation-coactivation). As a result of all these activations, the T0-helper secretes IL-2, which acts through an autocrine mechanism. In addition to the secretion of IL-2, the T0-helper starts synthesizing and integrating the alpha chain of IL-2R into its membrane (beta and gamma chains are permanently incorporated into the T0-helper membrane), triggering the biochemical cascade discussed below [138].
Janus kinase 1 (JAK1) is associated with the cytoplasmic part of the beta chain, and Janus kinase 3 (JAK3) is associated with the cytoplasmic part of the gamma chain. In the resting state, both are inactive. It is important to know that in the heterodimerized state, IL-2Rs have low affinity for IL-2. However, after attachment of alpha chain to beta and gamma chains the affinity of IL-2R to IL-2 increases dramatically. After the interaction of IL-2R with IL-2, JAK1/JAK3 change their conformation, which leads to transphosphorylation of JAK1 and JAK3 by the OH group of tyrosine residues located in a specific site called activation loop. This phosphorylation leads to a repeated change in the conformation of JAK1 and JAK3 so that they gain affinity to the cytoplasmic part of the beta chain and begin to phosphorylate it at the OH-group of tyrosine [138].
At rest, there is a Signal transducer and activator of transcription 5 (STAT5) protein in the cytosol of the cell that binds to the phosphorylated tyrosine residues of the beta chain using its SH2 domain. Next, JAK1 and JAK3 phosphorylate STAT5 by the OH-group of tyrosine residues and detaches back from the beta chain. In the cell cytoplasm, STAT5 homodimerizes (phosphorylated tyrosine residues of STAT5 proteins bind to each other's SH2 domains). Once STAT5 is homodimerized it undergoes conformational changes and NLS appears, which plays an important role in the transport of the STAT5 homodimer from the cytosol to the nucleoplasm. STAT5 is a transcription factor that increases the expression of genes responsible for T cell proliferation. This is important because when HIV infection occurs, the pool of CD4+ T-helper cells decreases significantly, which contributes negatively to the immune system's fight against HIV [138].
After the interaction of IL-2R with IL-2, JAK1/JAK3 transphosphorylate each other. This phosphorylation leads to a change in the conformation of JAK1 and JAK3 so that they gain affinity to the cytoplasmic part of the beta chain and begin to phosphorylate it. STAT5 binds to the phosphorylated tyrosine residues of the beta chain. JAK1 and JAK3 phosphorylate STAT5. STAT5 homodimerizes and enters the nucleoplasm.
10. Viral Vector-Based HIV Vaccines
The use of viral vectors makes it possible to induce a strong cytotoxic T-lymphocyte response through intracellular antigen expression, leading to the elimination of virus-infected cells.
Vaccinia virus and adenovirus are the most used vectors mostly due to their ability to induce an immune response, specifically including CTL, to the expressed foreign antigens. The most promising trials of viral vector-based HIV vaccines are presented in Table 8.
11. Broadly Neutralizing Antibodies Against HIV
For many years, passive immunization with protective antibodies has been used in the prevention and treatment of several bacterial and viral infections, subsequently influencing the current field of HIV vaccination. The mechanism of action of antibodies through their interaction with the innate and adaptive arms of the immune system, combined with their ability to bind and neutralize infectious agents, makes the antibody-based approach attractive to researchers.
HIV spike protein, or envelope (Env) protein, plays a role in infection of human (host) cells and has high levels of sequence variation between viral strains, but neutralizing antibodies exist against semi-conserved regions. Of greatest interest for the development of HIV-1 vaccines are broadly neutralizing antibodies (bnAbs), which are effective at targeting a wide range of viral strains. Most bnAbs target key regions of HIV’s Env protein that play a role in infection of human (host) cells.
Env is synthesized as a single gp160 molecule and is post-translationally cleaved by furin to a gp120 and gp41 that form trimeric fusion protein composed of three gp120 subunits and three gp41 subunits [144,145,146]. The Env complex is anchored to the viral membrane with gp41 transmembrane domain which binds with gp120 N-terminus domain.
The gp120 structure contains several variable and conserved regions that form loops, which are key targets for bNAbs. These loops include the V1/V2, V3, V4, and V5 regions [147]. The gp120 is heavily glycosylated, and many bNAbs are glycan-dependent, meaning they recognize both the peptide backbone and the attached glycans. The specific glycosylation patterns on the loops can create or obscure epitopes for bNAbs. Many bNAbs target epitopes that are formed by the three-dimensional structure of gp120, which includes specific arrangements of the loops. These conformational epitopes can span multiple loops or regions of gp120, highlighting the importance of maintaining the structural integrity of the protein for effective antibody binding. The balance between variability and conservation in the gp120 loops determines the accessibility and effectiveness of bNAbs. While variable regions allow for immune escape, the conserved regions that form functional sites are essential for viral entry and thus remain accessible targets for bNAbs.
The data from laboratory studies on bNAbs provide grounds to suggest that developing a vaccine using these antibodies will prevent HIV infection of cells. The gp120 subunit is responsible for binding to the CD4 receptor and a co-receptor (CCR5 or CXCR4) on the host cell surface, while gp41 facilitates the fusion of the viral and host cell membranes [148]. gp120 binding to CD4 leads to large conformational changes with rearrangement of the V1V2 and V3 loops, formation of a bridging sheet and the trimer “opening” [149]. The binding of CD4 and the corresponding conformational changes in gp120 allow the binding of the coreceptor [150]. Coreceptor binding leads to activation of the gp41 fusion peptide, which brings the viral and host cell membranes into contact to complete fusion [151,152].
The V1V2 site is located at the apex of the trimeric gp120 structure. The V1 and V2 loops form beta strands, stabilized by interstrand disulfide bridges and shielded by glycans [153]. V1V2 site is involved in shielding the coreceptor-binding site and is highly variable, which contributes to immune evasion [153,154,155]. Despite its variability, certain bNAbs target a glycan-dependent epitope in this region. These bNAbs recognize the conserved elements and glycosylation patterns of the V1V2 loop and stabilize the Env trimer thereby interfering with structural changes needed for coreceptor binding [156].
The V3 glycan supersite is centrally located in the gp120 structure and is often referred to as the "crown" of gp120. It plays a critical role in coreceptor binding (CXCR4 or CCR5) and is crucial for the conformational changes required for membrane fusion [157]. V3 glycan supersite contains a highly conserved 324GDIR327 motif that is part of the CCR5 coreceptor binding site [158]. It also contains various high-mannose glycans, most notably the N332 glycan, which is binded by all known bnAbs targeting this site [159,160,161,162]. The V3 loop is a major target for bNAbs, which recognize the conserved crown region and adjacent glycans. They neutralize this epitope by disrupting coreceptor binding and restricting conformational changes in the V3 loop to activate membrane fusion mediated by gp41 [157]. The CD4 binding site (CD4bs) is a conserved region of gp120 that interacts directly with the CD4, the primary receptor for viral entry, and is essential for the virus's ability to attach to the host cells. This interaction is the first step in HIV-1 entry, making the CD4bs a critical target for neutralization. This site overlaps with the V1, V2, and V3 loops and contains a glycan shield with multiple N-linked glycans [148]. Studies have identified two types of bNAbs that bind CD4bs: CD4 mimics, which contact the CD4bs with heavy chains encoded by either VH1-2 or VH1-46, and CDRH3-dependent bNAbs, interacting with the CD4bs by their CDRH3 loop.
Among CD4 mimics, VRC01 subclass is one of the most studied bNAbs targeting the CD4bs. VRC01 subclass characterized by a VH1-2-derived heavy chain, short CDRL1, and a 5-residue CDRL3 used to avoid clashes with the N276 glycan, present on most circulating strains [163,164].
VRC01 subclass bNAbs are generally broader and more potent than bnAbs with VH1-46. However, recently two CD4-mimic bNAbs with VH1-46 were identified. 1–18 and 1–55 have very high neutralization potency and breadth [165]. In addition to higher potency, 1–18 was resistant to VRC01 class escape mutations, indicating promise in potential combination therapies.
Unlike CD4 mimics, CDRH3-dependent bNAbs have different mode of binding with gp120. M1214_N1 is a newer CD4bs-targeting bNAb that binds to the CD4bs with a distinct orientation compared to CD4 mimics. M1214_N1 interacts differently with the region, utilizing its long CDRH3 loop to make deeper contacts with both the CD4bs and groove formed by the N197 and N386 glycosylation sites. The M1214_N1 epitope is centered on the CD4 binding loop and extends from V2 to V5, so called “V2V5 corridor” [166].
The gp120 "silent face" is a region on the outer domain of gp120 that is relatively inaccessible to immune recognition compared to other parts of the protein, such as the CD4bs. It is termed "silent" because it lacks the immunodominant features that typically attract a strong antibody response. The silent face is located on the opposite side of gp120 from the CD4 binding site and is covered by a dense glycan shield, which helps the virus evade immune recognition [167]. The epitope predominantly includes the N262, N295, and N448 glycans on a single gp120 monomer. By binding this epitope bNAbs can prevent conformational changes in gp120 or block interactions with host cell receptors, thereby neutralizing the virus [167].
The gp120-gp41 interface is a critical region where the two subunits of the HIV-1 Env trimer interact. This interface is responsible for maintaining the stability of the trimer and facilitating the conformational changes required for viral fusion with host cells. This region is targeted by the diverse bNAbs with different epitopes and dependency from glycans. Most of these bNAbs bind only trimeric Env protein. Their neutralization mechanisms are also different: some bnAbs, such as 8ANC195, can prevent the conformational changes required for membrane fusion [168], whereas others, such as 3BC176 and 3BC315 [169,170], destabilize the trimer by disrupting tryptophan clasps in gp41 [171].
The gp41 subunit of Env contains multiple epitopes, including the fusion peptide (FP) and membrane-proximal external region (MPER), which are both targets for bNAbs. The HIV-1 FP is one of the critical components of the virus-cell entry machinery [172]. It consists of 15-20 hydrophobic amino acid residues at the N terminus of the gp41 subunit [173]. Notably, some antibodies that interact with the gp120–gp41 interface, also bind the FP N-terminal residues, such as PGT151 [174]. The MPER is a highly conserved region of gp41 ectodomain located near the viral membrane and is also crucial for the fusion process. A feature of this epitope is that bNAbs against it recognize not only the MPER site, but also the lipid component of the viral membrane [175]. However, despite these difficulties, several antibodies binding this site and preventing the virus from merging with the host cell membrane have been identified.
Some of bNAbs targeting key regions of HIV Env protein, that play a role in infection of host cells are represented in Table 9.
To protect against HIV two or more bNAbs that target different parts of the virus are required. Combinations of bNAbs may lead to broader neutralization compared to giving bNAbs alone, and multispecific antibodies may have better efficiency than combinations. Many factors that determine effectiveness of antibody combinations exist. An alternative strategy to passive immunization with bNAbs is to use AAV-mediated gene transfer to enable in vivo production of desirable anti-HIV-1 antibodies [176,177]. The combinations of anti-HIV bNAbs under research are presented in Table 10.
The HIV-1 Env protein presents a variety of conserved epitopes that are targeted by bNAbs. The structure of Env, with its dense glycan shield and complex trimeric form, presents challenges for immune recognition but also provides multiple targets for bNAbs. These antibodies are essential for neutralizing the virus and are a central focus in efforts to develop an effective HIV vaccine.
12. Conclusions
Since none of the previously developed HIV vaccine concepts in isolation achieves a sufficient percentage of efficacy, and the combination of several types of vectors or antigens in some trials increases the possibility of achieving the goal, but does not lead to any stable result, the combinatorial approach should be applied in another direction. The key to defeating HIV may lie in the activation of several links of immunity at once, the combination of vaccine and non-vaccine approaches (pre-exposure prophylaxis, microbicides, multipurpose prevention technologies).
However, there is still a need for a vaccine - the percentage of infected medical workers, blood transfusion patients and drug users is high. Perhaps the COVID-19 pandemic and the subsequent surge in mRNA-based development will lead to significant results.
Author Contributions
Conceptualization, YVZ, VAZ; writing—original draft preparation, SOV, VOP, MYS, EA-k, KRP, NYuF; writing—review and editing, YVZ, SOV; visualization, SOV and MYS; supervision, YVZ; funding acquisition, YVZ. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Russian Science Foundation (grant no. 22-75-10140 for content analysis of HIV prevention, and grant no. 21-73-20202 for content analysis of HIV vaccine molecular mechanisms).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
Yury V. Zhernov was employed by the company Fomin Clinic. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- UNAIDS Available online: https://www.unaids.org/en/Homepage (accessed on 13 November 2024).
- AVAC | Global Health Advocacy, Access & Equity Available online: https://avac.org/ (accessed on 13 November 2024).
- Hodge, D.; Back, D.J.; Gibbons, S.; Khoo, S.H.; Marzolini, C. Pharmacokinetics and Drug–Drug Interactions of Long-Acting Intramuscular Cabotegravir and Rilpivirine. Clin. Pharmacokinet. 2021, 60, 835–853. [CrossRef]
- Temereanca, A.; Ruta, S. Strategies to Overcome HIV Drug Resistance-Current and Future Perspectives. Front. Microbiol. 2023, 14, 1133407. [CrossRef]
- Moskaleychik, F.F.; Laga, V.Y.; Delgado, E.; Vega, Y.; Fernandez-Garcia, A.; Perez-Alvarez, null; Kornilaeva, G.V.; Pronin, A.Y.; Zhernov, Y.V.; Thomson, M.M.; et al. [Rapid spread of the HIV-1 circular recombinant CRF02-AG in Russia and neighboring countries]. Vopr. Virusol. 2015, 60, 14–19.
- Karamov, E.; Epremyan, K.; Siniavin, A.; Zhernov, Y.; Cuevas, M.T.; Delgado, E.; Sánchez-Martínez, M.; Carrera, C.; Kornilaeva, G.; Turgiev, A.; et al. HIV-1 Genetic Diversity in Recently Diagnosed Infections in Moscow: Predominance of AFSU , Frequent Branching in Clusters, and Circulation of the Iberian Subtype G Variant. AIDS Res. Hum. Retroviruses 2018, 34, 629–634. [CrossRef]
- Frolova, O.P.; Butylchenko, O.V.; Gadzhieva, P.G.; Timofeeva, M.Yu.; Basangova, V.A.; Petrova, V.O.; Fadeeva, I.A.; Kashutina, M.I.; Zabroda, N.N.; Basov, A.A.; et al. Medical Care for Tuberculosis-HIV-Coinfected Patients in Russia with Respect to a Changeable Patients’ Structure. Trop. Med. Infect. Dis. 2022, 7, 86. [CrossRef]
- Zhernov, Y.V.; Kremb, S.; Helfer, M.; Schindler, M.; Harir, M.; Mueller, C.; Hertkorn, N.; Avvakumova, N.P.; Konstantinov, A.I.; Brack-Werner, R.; et al. Supramolecular Combinations of Humic Polyanions as Potent Microbicides with Polymodal Anti-HIV-Activities. New J. Chem. 2017, 41, 212–224. [CrossRef]
- Zhernov, Y. Natural Humic Substances Interfere with Multiple Stages of the Replication Cycle of Human Immunodeficiency Virus. J. Allergy Clin. Immunol. 2018, 141, AB233. [CrossRef]
- Zhernov, Yu.V.; Khaitov, M.R. Microbicides for Topical Immunoprevention of HIV Infection. Bull. Sib. Med. 2019, 18, 49–59. [CrossRef]
- Zhernov, Y.V.; Konstantinov, A.I.; Zherebker, A.; Nikolaev, E.; Orlov, A.; Savinykh, M.I.; Kornilaeva, G.V.; Karamov, E.V.; Perminova, I.V. Antiviral Activity of Natural Humic Substances and Shilajit Materials against HIV-1: Relation to Structure. Environ. Res. 2021, 193, 110312. [CrossRef]
- Zhernov, Y.V.; Petrova, V.O.; Simanduyev, M.Y.; Shcherbakov, D.V.; Polibin, R.V.; Mitrokhin, O.V.; Basov, A.A.; Zabroda, N.N.; Vysochanskaya, S.O.; Al-khaleefa, E.; et al. Microbicides for Topical HIV Immunoprophylaxis: Current Status and Future Prospects. Pharmaceuticals 2024, 17, 668. [CrossRef]
- Vassall, A.; Pickles, M.; Chandrashekar, S.; Boily, M.-C.; Shetty, G.; Guinness, L.; Lowndes, C.M.; Bradley, J.; Moses, S.; Alary, M.; et al. Cost-Effectiveness of HIV Prevention for High-Risk Groups at Scale: An Economic Evaluation of the Avahan Programme in South India. Lancet Glob. Health 2014, 2, e531–e540. [CrossRef]
- Wheatley, M.M.; Knowlton, G.S.; Butler, M.; Enns, E.A. Cost-Effectiveness of HIV Retention and Re-Engagement Interventions in High-Income Countries: A Systematic Literature Review. AIDS Behav. 2022, 26, 2159–2168. [CrossRef]
- Bozzani, F.M.; Terris-Prestholt, F.; Quaife, M.; Gafos, M.; Indravudh, P.P.; Giddings, R.; Medley, G.F.; Malhotra, S.; Torres-Rueda, S. Costs and Cost-Effectiveness of Biomedical, Non-Surgical HIV Prevention Interventions: A Systematic Literature Review. PharmacoEconomics 2023, 41, 467–480. [CrossRef]
- The Nobel Prize in Physiology or Medicine 2023 Available online: https://www.nobelprize.org/prizes/medicine/2023/summary/ (accessed on 13 November 2024).
- Fanales-Belasio, E.; Raimondo, M.; Suligoi, B.; Buttò, S. HIV Virology and Pathogenetic Mechanisms of Infection: A Brief Overview. Ann. Ist. Super. Sanita 2010, 46, 5–14. [CrossRef]
- Khalid, K.; Padda, J.; Khedr, A.; Ismail, D.; Zubair, U.; Al-Ewaidat, O.A.; Padda, S.; Cooper, A.C.; Jean-Charles, G. HIV and Messenger RNA Vaccine. Cureus 2021. [CrossRef]
- Nyamweya, S.; Hegedus, A.; Jaye, A.; Rowland-Jones, S.; Flanagan, K.L.; Macallan, D.C. Comparing HIV-1 and HIV-2 Infection: Lessons for Viral Immunopathogenesis. Rev. Med. Virol. 2013, 23, 221–240. [CrossRef]
- Azevedo-Pereira, J.M.; Santos-Costa, Q. HIV Interaction With Human Host: HIV-2 As a Model of a Less Virulent Infection. AIDS Rev. 2016, 18, 44–53.
- Travers, K.; Mboup, S.; Marlink, R.; Guèye-Nidaye, A.; Siby, T.; Thior, L.; Traore, I.; Dieng-Sarr, A.; Sankalé, J.-L.; Mullins, C.; et al. Natural Protection Against HIV-1 Infection Provided by HIV-2. Science 1995, 268, 1612–1615. [CrossRef]
- Rowland-Jones, S. Protective Immunity Against HIV Infection: Lessons from HIV-2 Infection. Future Microbiol. 2006, 1, 427–433. [CrossRef]
- Aberg, J.A.; Kaplan, J.E.; Libman, H.; Emmanuel, P.; Anderson, J.R.; Stone, V.E.; Oleske, J.M.; Currier, J.S.; Gallant, J.E. Primary Care Guidelines for the Management of Persons Infected with Human Immunodeficiency Virus: 2009 Update by the HIV Medicine Association of the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 49, 651–681. [CrossRef]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 Envelope Glycoprotein Biosynthesis, Trafficking, and Incorporation. J. Mol. Biol. 2011, 410, 582–608. [CrossRef]
- Ng’uni, T.; Chasara, C.; Ndhlovu, Z.M. Major Scientific Hurdles in HIV Vaccine Development: Historical Perspective and Future Directions. Front. Immunol. 2020, 11, 590780. [CrossRef]
- Bour, S.; Geleziunas, R.; Wainberg, M.A. The Human Immunodeficiency Virus Type 1 (HIV-1) CD4 Receptor and Its Central Role in Promotion of HIV-1 Infection. Microbiol. Rev. 1995, 59, 63–93. [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV Gp120 Envelope Glycoprotein in Complex with the CD4 Receptor and a Neutralizing Human Antibody. Nature 1998, 393, 648–659. [CrossRef]
- Ding, C.; Patel, D.; Ma, Y.; Mann, J.F.S.; Wu, J.; Gao, Y. Employing Broadly Neutralizing Antibodies as a Human Immunodeficiency Virus Prophylactic & Therapeutic Application. Front. Immunol. 2021, 12, 697683. [CrossRef]
- Chen, X.; Wang, X. The HIV-1 Gag P6: A Promising Target for Therapeutic Intervention. Retrovirology 2024, 21, 1. [CrossRef]
- Marie, V.; Gordon, M.L. The HIV-1 Gag Protein Displays Extensive Functional and Structural Roles in Virus Replication and Infectivity. Int. J. Mol. Sci. 2022, 23, 7569. [CrossRef]
- Mailler, E.; Bernacchi, S.; Marquet, R.; Paillart, J.-C.; Vivet-Boudou, V.; Smyth, R.P. The Life-Cycle of the HIV-1 Gag–RNA Complex. Viruses 2016, 8, 248. [CrossRef]
- Hill, M.; Tachedjian, G.; Mak, J. The Packaging and Maturation of the HIV-1 Pol Proteins. Curr. HIV Res. 3, 73–85. [CrossRef]
- Cafaro, A.; Schietroma, I.; Sernicola, L.; Belli, R.; Campagna, M.; Mancini, F.; Farcomeni, S.; Pavone-Cossut, M.R.; Borsetti, A.; Monini, P.; et al. Role of HIV-1 Tat Protein Interactions with Host Receptors in HIV Infection and Pathogenesis. Int. J. Mol. Sci. 2024, 25, 1704. [CrossRef]
- Brigati, C.; Giacca, M.; Noonan, D.M.; Albini, A. HIV Tat, Its TARgets and the Control of Viral Gene Expression. FEMS Microbiol. Lett. 2003, 220, 57–65. [CrossRef]
- Kula, A.; Marcello, A. Dynamic Post-Transcriptional Regulation of HIV-1 Gene Expression. Biology 2012, 1, 116–133. [CrossRef]
- Miller, R.H.; Sarver, N. HIV Accessory Proteins: Emerging Therapeutic Targets. Mol. Med. 1995, 1, 479–485. [CrossRef]
- Sudderuddin, H.; Kinloch, N.N.; Jin, S.W.; Miller, R.L.; Jones, B.R.; Brumme, C.J.; Joy, J.B.; Brockman, M.A.; Brumme, Z.L. Longitudinal Within-Host Evolution of HIV Nef-Mediated CD4, HLA and SERINC5 Downregulation Activity: A Case Study. Retrovirology 2020, 17, 3. [CrossRef]
- Andrew, A.; Strebel, K. HIV-1 Accessory Proteins: Vpu and Vif. In Human Retroviruses; Vicenzi, E., Poli, G., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, 2014; Vol. 1087, pp. 135–158 ISBN 978-1-62703-669-6.
- Rashid, F.; Zaongo, S.D.; Iqbal, H.; Harypursat, V.; Song, F.; Chen, Y. Interactions between HIV Proteins and Host Restriction Factors: Implications for Potential Therapeutic Intervention in HIV Infection. Front. Immunol. 2024, 15. [CrossRef]
- Dubé, M.; Bego, M.G.; Paquay, C.; Cohen, É.A. Modulation of HIV-1-Host Interaction: Role of the Vpu Accessory Protein. Retrovirology 2010, 7, 114. [CrossRef]
- Kogan, M.; Rappaport, J. HIV-1 Accessory Protein Vpr: Relevance in the Pathogenesis of HIV and Potential for Therapeutic Intervention. Retrovirology 2011, 8, 25. [CrossRef]
- Vanegas-Torres, C.A.; Schindler, M. HIV-1 Vpr Functions in Primary CD4+ T Cells. Viruses 2024, 16, 420. [CrossRef]
- Bleul, C.C.; Wu, L.; Hoxie, J.A.; Springer, T.A.; Mackay, C.R. The HIV Coreceptors CXCR4 and CCR5 Are Differentially Expressed and Regulated on Human T Lymphocytes. Proc. Natl. Acad. Sci. 1997, 94, 1925–1930. [CrossRef]
- Zaitseva, M.; Blauvelt, A.; Lee, S.; Lapham, C.K.; Kiaus-Kovrun, V.; Mostowski, H.; Manischewitz, J.; Golding, H. Expression and Function of CCR5 and CXCR4 on Human Langerhans Cells and Macrophages: Implications for HIV Primary Infection. Nat. Med. 1997, 3, 1369–1375. [CrossRef]
- Ostrowski, M.A.; Justement, S.J.; Catanzaro, A.; Hallahan, C.A.; Ehler, L.A.; Mizell, S.B.; Kumar, P.N.; Mican, J.A.; Chun, T.-W.; Fauci, A.S. Expression of Chemokine Receptors CXCR4 and CCR5 in HIV-1-Infected and Uninfected Individuals. J. Immunol. 1998, 161, 3195–3201. [CrossRef]
- Woodham, A.W.; Skeate, J.G.; Sanna, A.M.; Taylor, J.R.; Da Silva, D.M.; Cannon, P.M.; Kast, W.M. Human Immunodeficiency Virus Immune Cell Receptors, Coreceptors, and Cofactors: Implications for Prevention and Treatment. AIDS Patient Care STDs 2016, 30, 291–306. [CrossRef]
- Barmania, F.; Pepper, M.S. C-C Chemokine Receptor Type Five (CCR5): An Emerging Target for the Control of HIV Infection. Appl. Transl. Genomics 2013, 2, 3–16. [CrossRef]
- McAleer, W.J.; Buynak, E.B.; Maigetter, R.Z.; Wampler, D.E.; Miller, W.J.; Hilleman, M.R. Human Hepatitis B Vaccine from Recombinant Yeast. Nature 1984, 307, 178–180. [CrossRef]
- Esparza, J. What Has 30 Years of HIV Vaccine Research Taught Us? Vaccines 2013, 1, 513–526. [CrossRef]
- Dolin, R.; Graham, B.S.; Greenberg, S.B.; Tacket, C.O.; Belshe, R.B.; Midthun, K.; Clements, M.L.; Gorse, G.J.; Horgan, B.W.; Atmar, R.L.; et al. The Safety and Immunogenicity of a Human Immunodeficiency Virus Type 1 (HIV-1) Recombinant Gp160 Candidate Vaccine in Humans. Ann. Intern. Med. 1991, 114, 119–127. [CrossRef]
- Perkus, M.E.; Tartaglia, J.; Paoletti, E. Poxvirus-Based Vaccine Candidates for Cancer, AIDS, and Other Infectious Diseases. J. Leukoc. Biol. 1995, 58, 1–13. [CrossRef]
- Chakrabarti, S.; Robert-Guroff, M.; Wong-Staal, F.; Gallo, R.C.; Moss, B. Expression of the HTLV-III Envelope Gene by a Recombinant Vaccinia Virus. Nature 1986, 320, 535–537. [CrossRef]
- Hu, S.-L.; Kosowski, S.G.; Dalrymple, J.M. Expression of AIDS Virus Envelope Gene in Recombinant Vaccinia Viruses. Nature 1986, 320, 537–540. [CrossRef]
- Cooney, E. Safety of and Immunological Response to a Recombinant Vaccinia Virus Vaccine Expressing HIV Envelope Glycoprotein. The Lancet 1991, 337, 567–572. [CrossRef]
- Cooney, E.L.; McElrath, M.J.; Corey, L.; Hu, S.L.; Collier, A.C.; Arditti, D.; Hoffman, M.; Coombs, R.W.; Smith, G.E.; Greenberg, P.D. Enhanced Immunity to Human Immunodeficiency Virus (HIV) Envelope Elicited by a Combined Vaccine Regimen Consisting of Priming with a Vaccinia Recombinant Expressing HIV Envelope and Boosting with Gp160 Protein. Proc. Natl. Acad. Sci. 1993, 90, 1882–1886. [CrossRef]
- Graham, B.S.; Matthews, T.J.; Belshe, R.B.; Clements, M.L.; Dolin, R.; Wright, P.F.; Gorse, G.J.; Schwartz, D.H.; Keefer, M.C.; Bolognesi, D.P.; et al. Augmentation of Human Immunodeficiency Virus Type 1 Neutralizing Antibody by Priming with Gp160 Recombinant Vaccinia and Boosting with Gp160 in Vaccinia-Naive Adults. J. Infect. Dis. 1993, 167, 533–537. [CrossRef]
- Redfield, R.R.; Wright, D.C.; James, W.D.; Jones, T.S.; Brown, C.; Burke, D.S. Disseminated Vaccinia in a Military Recruit with Human Immunodeficiency Virus (HIV) Disease. N. Engl. J. Med. 1987, 316, 673–676. [CrossRef]
- Zagury, D.; Bernard, J.; Cheynier, R.; Desportes, I.; Leonard, R.; Fouchard, M.; Reveil, B.; Ittele, D.; Lurhuma, Z.; Mbayo, K.; et al. A Group Specific Anamnestic Immune Reaction against HIV-1 Induced by a Candidate Vaccine against AIDS. Nature 1988, 332, 728–731. [CrossRef]
- Cox, W.I.; Tartaglia, J.; Paoletti, E. Induction of Cytotoxic T Lymphocytes by Recombinant Canarypox (ALVAC) and Attenuated Vaccinia (NYVAC) Viruses Expressing the HIV-1 Envelope Glycoprotein. Virology 1993, 195, 845–850. [CrossRef]
- Pialoux, G.; Excler, J.-L.; Rivière, Y.; Gonzalez-Canali, G.; Feuillie, V.; Coulaud, P.; Gluckman, J.-C.; Matthews, T.J.; Meignier, B.; Kieny, M.-P.; et al. A Prime-Boost Approach to HIV Preventive Vaccine Using a Recombinant Canarypox Virus Expressing Glycoprotein 160 (MN) Followed by a Recombinant Glycoprotein 160 (MN/LAI). AIDS Res. Hum. Retroviruses 1995, 11, 373–381. [CrossRef]
- Marovich, M.A. ALVAC-HIV Vaccines: Clinical Trial Experience Focusing on Progress in Vaccine Development. Expert Rev. Vaccines 2004, 3, S99–S104. [CrossRef]
- Thongcharoen, P.; Suriyanon, V.; Paris, R.M.; Khamboonruang, C.; De Souza, M.S.; Ratto-Kim, S.; Karnasuta, C.; Polonis, V.R.; Baglyos, L.; Habib, R.E.; et al. A Phase 1/2 Comparative Vaccine Trial of the Safety and Immunogenicity of a CRF01_AE (Subtype E) Candidate Vaccine: ALVAC-HIV (vCP1521) Prime With Oligomeric Gp160 (92TH023/LAI-DID) or Bivalent Gp120 (CM235/SF2) Boost. JAIDS J. Acquir. Immune Defic. Syndr. 2007, 46, 48–55. [CrossRef]
- The rgp120 HIV Vaccine Study Group Placebo-Controlled Phase 3 Trial of a Recombinant Glycoprotein 120 Vaccine to Prevent HIV-1 Infection. J. Infect. Dis. 2005, 191, 654–665. [CrossRef]
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W.; Choopanya, K.; Bangkok Vaccine Evaluation Group Randomized, Double-Blind, Placebo-Controlled Efficacy Trial of a Bivalent Recombinant Glycoprotein 120 HIV-1 Vaccine among Injection Drug Users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671. [CrossRef]
- Loreto, C.; La Rocca, G.; Anzalone, R.; Caltabiano, R.; Vespasiani, G.; Castorina, S.; Ralph, D.J.; Cellek, S.; Musumeci, G.; Giunta, S.; et al. The Role of Intrinsic Pathway in Apoptosis Activation and Progression in Peyronie’s Disease. BioMed Res. Int. 2014, 2014, 1–10. [CrossRef]
- Shakeri, R.; Kheirollahi, A.; Davoodi, J. Apaf-1: Regulation and Function in Cell Death. Biochimie 2017, 135, 111–125. [CrossRef]
- Cheng, J.; Lucas, P.C.; McAllister-Lucas, L.M. Canonical and Non-Canonical Roles of GRK2 in Lymphocytes. Cells 2021, 10, 307. [CrossRef]
- Laporte, S.A.; Miller, W.E.; Kim, K.-M.; Caron, M.G. β-Arrestin/AP-2 Interaction in G Protein-Coupled Receptor Internalization. J. Biol. Chem. 2002, 277, 9247–9254. [CrossRef]
- Sundborger, A.; Soderblom, C.; Vorontsova, O.; Evergren, E.; Hinshaw, J.E.; Shupliakov, O. An Endophilin–Dynamin Complex Promotes Budding of Clathrin-Coated Vesicles during Synaptic Vesicle Recycling. J. Cell Sci. 2011, 124, 133–143. [CrossRef]
- Royle, S.J. The Cellular Functions of Clathrin. Cell. Mol. Life Sci. 2006, 63, 1823–1832. [CrossRef]
- McMichael, A.; Hanke, T. The Quest for an AIDS Vaccine: Is the CD8+ T-Cell Approach Feasible? Nat. Rev. Immunol. 2002, 2, 283–291. [CrossRef]
- Mudd, P.A.; Martins, M.A.; Ericsen, A.J.; Tully, D.C.; Power, K.A.; Bean, A.T.; Piaskowski, S.M.; Duan, L.; Seese, A.; Gladden, A.D.; et al. Vaccine-Induced CD8+ T Cells Control AIDS Virus Replication. Nature 2012, 491, 129–133. [CrossRef]
- Schoenly, K.A.; Weiner, D.B. Human Immunodeficiency Virus Type 1 Vaccine Development: Recent Advances in the Cytotoxic T-Lymphocyte Platform “Spotty Business.” J. Virol. 2008, 82, 3166–3180. [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; Del Rio, C.; et al. Efficacy Assessment of a Cell-Mediated Immunity HIV-1 Vaccine (the Step Study): A Double-Blind, Randomised, Placebo-Controlled, Test-of-Concept Trial. The Lancet 2008, 372, 1881–1893. [CrossRef]
- Duerr, A.; Huang, Y.; Buchbinder, S.; Coombs, R.W.; Sanchez, J.; Del Rio, C.; Casapia, M.; Santiago, S.; Gilbert, P.; Corey, L.; et al. Extended Follow-up Confirms Early Vaccine-Enhanced Risk of HIV Acquisition and Demonstrates Waning Effect Over Time Among Participants in a Randomized Trial of Recombinant Adenovirus HIV Vaccine (Step Study). J. Infect. Dis. 2012, 206, 258–266. [CrossRef]
- Catanzaro, A.T.; Roederer, M.; Koup, R.A.; Bailer, R.T.; Enama, M.E.; Nason, M.C.; Martin, J.E.; Rucker, S.; Andrews, C.A.; Gomez, P.L.; et al. Phase I Clinical Evaluation of a Six-Plasmid Multiclade HIV-1 DNA Candidate Vaccine. Vaccine 2007, 25, 4085–4092. [CrossRef]
- McEnery, R. HVTN 505 Trial Expanded to See If Vaccine Candidates Can Block HIV Acquisition. IAVI Rep. Newsl. Int. AIDS Vaccine Res. 2011, 15, 17.
- Catanzaro, A.T.; Koup, R.A.; Roederer, M.; Bailer, R.T.; Enama, M.E.; Moodie, Z.; Gu, L.; Martin, J.E.; Novik, L.; Chakrabarti, B.K.; et al. Phase 1 Safety and Immunogenicity Evaluation of a Multiclade HIV-1 Candidate Vaccine Delivered by a Replication-Defective Recombinant Adenovirus Vector. J. Infect. Dis. 2006, 194, 1638–1649. [CrossRef]
- Fischinger, S.; Shin, S.; Boudreau, C.M.; Ackerman, M.; Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kim, J.H.; Robb, M.L.; Michael, N.L.; et al. Protein-Based, but Not Viral Vector Alone, HIV Vaccine Boosting Drives an IgG1-Biased Polyfunctional Humoral Immune Response. JCI Insight 2020, 5, e135057. [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Excler, J.-L.; Nitayaphan, S.; Kaewkungwal, J.; Premsri, N.; Kunasol, P.; Karasavvas, N.; Schuetz, A.; Ngauy, V.; et al. Randomized, Double-Blind Evaluation of Late Boost Strategies for HIV-Uninfected Vaccine Recipients in the RV144 HIV Vaccine Efficacy Trial. J. Infect. Dis. 2017, 215, 1255–1263. [CrossRef]
- Pitisuttithum, P.; Nitayaphan, S.; Chariyalertsak, S.; Kaewkungwal, J.; Dawson, P.; Dhitavat, J.; Phonrat, B.; Akapirat, S.; Karasavvas, N.; Wieczorek, L.; et al. Late Boosting of the RV144 Regimen with AIDSVAX B/E and ALVAC-HIV in HIV-Uninfected Thai Volunteers: A Double-Blind, Randomised Controlled Trial. Lancet HIV 2020, 7, e238–e248. [CrossRef]
- Gray, G.E.; Huang, Y.; Grunenberg, N.; Laher, F.; Roux, S.; Andersen-Nissen, E.; De Rosa, S.C.; Flach, B.; Randhawa, A.K.; Jensen, R.; et al. Immune Correlates of the Thai RV144 HIV Vaccine Regimen in South Africa. Sci. Transl. Med. 2019, 11, eaax1880. [CrossRef]
- Bekker, L.-G.; Moodie, Z.; Grunenberg, N.; Laher, F.; Tomaras, G.D.; Cohen, K.W.; Allen, M.; Malahleha, M.; Mngadi, K.; Daniels, B.; et al. Subtype C ALVAC-HIV and Bivalent Subtype C Gp120/MF59 HIV-1 Vaccine in Low-Risk, HIV-Uninfected, South African Adults: A Phase 1/2 Trial. Lancet HIV 2018, 5, e366–e378. [CrossRef]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-Correlates Analysis of an HIV-1 Vaccine Efficacy Trial. N. Engl. J. Med. 2012, 366, 1275–1286. [CrossRef]
- Zolla-Pazner, S.; Gilbert, P.B. Revisiting the Correlate of Reduced HIV Infection Risk in the Rv144 Vaccine Trial. J. Virol. 2019, 93, e00629-19. [CrossRef]
- National Institute of Allergy and Infectious Diseases (NIAID) A Phase-1 Open-Label Clinical Trial to Evaluate the Safety and Immunogenicity of Synthetic DNAs Encoding NP-GT8 and IL-12, With or Without a TLR-Agonist- Adjuvanted HIV Env Trimer 4571 Boost, in Adults Without HIV; clinicaltrials.gov, 2024;
- Pitisuttithum, P.; Nitayaphan, S.; Chariyalertsak, S.; Kaewkungwal, J.; Dawson, P.; Dhitavat, J.; Phonrat, B.; Akapirat, S.; Karasavvas, N.; Wieczorek, L.; et al. Late Boosting of the RV144 Regimen with AIDSVAX B/E and ALVAC-HIV in HIV-Uninfected Thai Volunteers, a Randomised Controlled Trial. Lancet HIV 2020, 7, e238–e248. [CrossRef]
- HIV Vaccine Trials Network Phase 1b Randomized Double Blind Placebo Controlled Clinical Trial to Evaluate the Safety and Immunogenicity of the Vaccine Regimen ALVAC-HIV (vCP1521) Followed by AIDSVAX® B/E in Healthy, HIV-1 Uninfected Adult Participants in South Africa; clinicaltrials.gov, 2019;
- National Institute of Allergy and Infectious Diseases (NIAID) A Phase 1-2 Randomized, Double-Blind, Placebo-Controlled Clinical Trial of Clade C ALVAC-HIV (vCP2438) and Bivalent Subtype C gp120/MF59® in HIV-Uninfected Adults at Low Risk of HIV Infection; clinicaltrials.gov, 2021;
- Trovato, M.; D’Apice, L.; Prisco, A.; De Berardinis, P. HIV Vaccination: A Roadmap among Advancements and Concerns. Int. J. Mol. Sci. 2018, 19, 1241. [CrossRef]
- Tsoukas, C.M.; Raboud, J.; Bernard, N.F.; Montaner, J.S.; Gill, M.J.; Rachlis, A.; Fong, I.W.; Schlech, W.; Djurdjev, O.; Freedman, J.; et al. Active Immunization of Patients with HIV Infection: A Study of the Effect of VaxSyn, a Recombinant HIV Envelope Subunit Vaccine, on Progression of Immunodeficiency. AIDS Res. Hum. Retroviruses 1998, 14, 483–490. [CrossRef]
- Cooney, E.L.; Collier, A.C.; Greenberg, P.D.; Coombs, R.W.; Zarling, J.; Arditti, D.E.; Hoffman, M.C.; Hu, S.L.; Corey, L. Safety of and Immunological Response to a Recombinant Vaccinia Virus Vaccine Expressing HIV Envelope Glycoprotein. Lancet Lond. Engl. 1991, 337, 567–572. [CrossRef]
- VaxGen A Phase III Trial to Determine the Efficacy of AIDSVAX B/E Vaccine in Intravenous Drug Users in Bangkok, Thailand; clinicaltrials.gov, 2005;
- VaxGen A Phase III Trial to Determine the Efficacy of Bivalent AIDSVAX B/B Vaccine in Adults at Risk of Sexually Transmitted HIV-1 Infection in North America; clinicaltrials.gov, 2005;
- Gray, G.; Buchbinder, S.; Duerr, A. Overview of STEP and Phambili Trial Results: Two Phase IIb Test of Concept Studies Investigating the Efficacy of MRK Ad5 Gag/Pol/Nef Sub-Type B HIV Vaccine. Curr. Opin. HIV AIDS 2010, 5, 357–361. [CrossRef]
- National Institute of Allergy and Infectious Diseases (NIAID) A Multicenter Double-Blind Randomized Placebo-Controlled Phase IIB Test-of-Concept Study to Evaluate the Safety and Efficacy of a Three-Dose Regimen of the Clade B-Based Merck Adenovirus Serotype 5 HIV-1 Gag/Pol/Nef Vaccine in HIV-1 Uninfected Adults in South Africa; clinicaltrials.gov, 2021;
- U.S. Army Medical Research and Development Command A Phase III Trial of Aventis Pasteur Live Recombinant ALVAC-HIV (vCP1521) Priming With VaxGen Gp120 B/E (AIDSVAX B/E) Boosting in HIV-Uninfected Thai Adults; clinicaltrials.gov, 2019;
- National Institute of Allergy and Infectious Diseases (NIAID) Phase 2b, Randomized, Placebo-Controlled Test-of-Concept Trial to Evaluate the Safety and Efficacy of a Multiclade HIV-1 DNA Plasmid Vaccine Followed by a Multiclade HIV-1 Recombinant Adenoviral Vector Vaccine in HIV-Uninfected, Adenovirus Type 5 Neutralizing Antibody Negative, Circumcised Men and Male-to-Female (MTF) Transgender Persons, Who Have Sex With Men; clinicaltrials.gov, 2021;
- National Institute of Allergy and Infectious Diseases (NIAID) A Pivotal Phase 2b/3 Multisite, Randomized, Double-Blind, Placebo-Controlled Clinical Trial to Evaluate the Safety and Efficacy of ALVAC-HIV (vCP2438) and Bivalent Subtype C Gp120/MF59 in Preventing HIV-1 Infection in Adults in South Africa; clinicaltrials.gov, 2024;
- Janssen Vaccines & Prevention B.V. A Multicenter, Randomized, Double-Blind, Placebo-Controlled Phase 2b Efficacy Study of a Heterologous Prime/Boost Vaccine Regimen of Ad26.Mos4.HIV and Aluminum Phosphate-Adjuvanted Clade C Gp140 in Preventing HIV-1 Infection in Adult Women in Sub-Saharan Africa; clinicaltrials.gov, 2023;
- Janssen Vaccines & Prevention B.V. A Multi-Center, Randomized, Double-Blind, Placebo-Controlled Phase 3 Efficacy Study of a Heterologous Vaccine Regimen of Ad26.Mos4.HIV and Adjuvanted Clade C Gp140 and Mosaic Gp140 to Prevent HIV-1 Infection Among Cis-Gender Men and Transgender Individuals Who Have Sex With Cis-Gender Men and/or Transgender Individuals; clinicaltrials.gov, 2024;
- MRC/UVRI and LSHTM Uganda Research Unit A Phase IIb Three-Arm, Two-Stage HIV Prophylactic Vaccine Trial With a Second Randomisation to Compare TAF/FTC to TDF/FTC as Pre-Exposure Prophylaxis; clinicaltrials.gov, 2023;
- Mu, Z.; Haynes, B.F.; Cain, D.W. HIV mRNA Vaccines—Progress and Future Paths. Vaccines 2021, 9, 134. [CrossRef]
- Hobeika, E.; Nielsen, P.J.; Medgyesi, D. Signaling Mechanisms Regulating B-Lymphocyte Activation and Tolerance. J. Mol. Med. 2015, 93, 143–158. [CrossRef]
- Debant, M.; Hemon, P.; Brigaudeau, C.; Renaudineau, Y.; Mignen, O. Calcium Signaling and Cell Fate: How Can Ca2+ Signals Contribute to Wrong Decisions for Chronic Lymphocytic Leukemic B Lymphocyte Outcome? Int. J. Dev. Biol. 2015, 59, 379–389. [CrossRef]
- Grondona, P.; Bucher, P.; Schulze-Osthoff, K.; Hailfinger, S.; Schmitt, A. NF-κB Activation in Lymphoid Malignancies: Genetics, Signaling, and Targeted Therapy. Biomedicines 2018, 6, 38. [CrossRef]
- Patterson, S.D.; Huang, X.; Jørgensen, H.G.; Michie, A.M. Transcriptional Regulation by the NFAT Family in Acute Myeloid Leukaemia. Hemato 2021, 2, 556–571. [CrossRef]
- International AIDS Vaccine Initiative A Phase 1, Randomized, First-in-Human, Open-Label Study to Evaluate the Safety and Immunogenicity of eOD-GT8 60mer mRNA Vaccine (mRNA-1644) and Core-G28v2 60mer mRNA Vaccine (mRNA-1644v2-Core) in HIV-1 Uninfected Adults in Good General Health; clinicaltrials.gov, 2024;
- International AIDS Vaccine Initiative A Phase 1 Study to Evaluate the Safety and Immunogenicity of eOD-GT8 60mer mRNA Vaccine (mRNA-1644) in HIV-1 Uninfected Adults in Good General Health; clinicaltrials.gov, 2023;
- National Institute of Allergy and Infectious Diseases (NIAID) A Phase 1, Randomized, Open-Label Clinical Trial to Evaluate the Safety and Immunogenicity of BG505 MD39.3, BG505 MD39.3 Gp151, and BG505 MD39.3 Gp151 CD4KO HIV Trimer mRNA Vaccines in Healthy, HIV-Uninfected Adult Participants; clinicaltrials.gov, 2024;
- Saxena, M.; Bhardwaj, N. Re-Emergence of Dendritic Cell Vaccines for Cancer Treatment. Trends Cancer 2018, 4, 119–137. [CrossRef]
- Sabado, R.L.; Bhardwaj, N. Cancer Immunotherapy: Dendritic-Cell Vaccines on the Move. Nature 2015, 519, 300–301. [CrossRef]
- Espinar-Buitrago, M.; Muñoz-Fernández, M.A. New Approaches to Dendritic Cell-Based Therapeutic Vaccines Against HIV-1 Infection. Front. Immunol. 2022, 12, 719664. [CrossRef]
- Jacobson, J.M.; Routy, J.-P.; Welles, S.; DeBenedette, M.; Tcherepanova, I.; Angel, J.B.; Asmuth, D.M.; Stein, D.K.; Baril, J.-G.; McKellar, M.; et al. Dendritic Cell Immunotherapy for HIV-1 Infection Using Autologous HIV-1 RNA: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. JAIDS J. Acquir. Immune Defic. Syndr. 2016, 72, 31–38. [CrossRef]
- Macatangay, B.J.C.; Riddler, S.A.; Wheeler, N.D.; Spindler, J.; Lawani, M.; Hong, F.; Buffo, M.J.; Whiteside, T.L.; Kearney, M.F.; Mellors, J.W.; et al. Therapeutic Vaccination With Dendritic Cells Loaded With Autologous HIV Type 1–Infected Apoptotic Cells. J. Infect. Dis. 2016, 213, 1400–1409. [CrossRef]
- García, F.; Plana, M.; Climent, N.; León, A.; Gatell, J.M.; Gallart, T. Dendritic Cell Based Vaccines for HIV Infection: The Way Ahead. Hum. Vaccines Immunother. 2013, 9, 2445–2452. [CrossRef]
- García, F.; Climent, N.; Guardo, A.C.; Gil, C.; León, A.; Autran, B.; Lifson, J.D.; Martínez-Picado, J.; Dalmau, J.; Clotet, B.; et al. A Dendritic Cell–Based Vaccine Elicits T Cell Responses Associated with Control of HIV-1 Replication. Sci. Transl. Med. 2013, 5. [CrossRef]
- Muro, R.; Takayanagi, H.; Nitta, T. T Cell Receptor Signaling for γδT Cell Development. Inflamm. Regen. 2019, 39, 6. [CrossRef]
- Park, I.; Yun, Y. Transmembrane Adaptor Proteins Positively Regulating the Activation of Lymphocytes. Immune Netw. 2009, 9, 53. [CrossRef]
- Thatcher, J.D. The Ras-MAPK Signal Transduction Pathway. Sci. Signal. 2010, 3. [CrossRef]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [CrossRef]
- Larijani, M.S.; Sadat, S.M.; Bolhassani, A.; Pouriayevali, M.H.; Bahramali, G.; Ramezani, A. In Silico Design and Immunologic Evaluation of HIV-1 P24-Nef Fusion Protein to Approach a Therapeutic Vaccine Candidate. Curr. HIV Res. 2019, 16, 322–337. [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-Based Synthetic Vaccines. Chem. Sci. 2016, 7, 842–854. [CrossRef]
- Rockstroh, J.K.; Asmuth, D.; Pantaleo, G.; Clotet, B.; Podzamczer, D.; van Lunzen, J.; Arastéh, K.; Mitsuyasu, R.; Peters, B.; Silvia, N.; et al. Re-Boost Immunizations with the Peptide-Based Therapeutic HIV Vaccine, Vacc-4x, Restores Geometric Mean Viral Load Set-Point during Treatment Interruption. PLoS ONE 2019, 14, e0210965. [CrossRef]
- Pollard, R.B.; Rockstroh, J.K.; Pantaleo, G.; Asmuth, D.M.; Peters, B.; Lazzarin, A.; Garcia, F.; Ellefsen, K.; Podzamczer, D.; van Lunzen, J.; et al. Safety and Efficacy of the Peptide-Based Therapeutic Vaccine for HIV-1, Vacc-4x: A Phase 2 Randomised, Double-Blind, Placebo-Controlled Trial. Lancet Infect. Dis. 2014, 14, 291–300. [CrossRef]
- Gharakhanian, S.; Katlama, C.; Launay, O.; Bodilis, H.; Calin, R.; Ho, R.; Fang, T.; Marcu, M.; Autran, B.; Vieillard, V.; et al. VAC-3S, an Immunoprotective HIV Vaccine Directed to the 3S Motif of Gp41, in Patients Receiving ART: Safety, Dose & Immunization Schedule Assessment. Phase I Study Results: Immunologic Endpoints & Safety I ● Introduction and Background Phase I Study: Methods Phase I Study Results: Immunologic Endpoints & Safety.; October 2013.
- Brekke, K.; Sommerfelt, M.; Ökvist, M.; Dyrhol-Riise, A.M.; Kvale, D. The Therapeutic HIV Env C5/Gp41 Vaccine Candidate Vacc-C5 Induces Specific T Cell Regulation in a Phase I/II Clinical Study. BMC Infect. Dis. 2017, 17, 228. [CrossRef]
- Jensen, K.J.; Gómez Román, V.R.; Jensen, S.S.; Leo-Hansen, C.; Karlsson, I.; Katzenstein, T.L.; Rodrigues, C.M.; Jespersen, S.; Janitzek, C.M.; Té, D. da S.; et al. Clade A HIV-1 Gag-Specific T Cell Responses Are Frequent but Do Not Correlate with Viral Loads in a Cohort of Treatment-Naive HIV-Infected Individuals Living in Guinea-Bissau. Clin. Vaccine Immunol. CVI 2012, 19, 1999–2001. [CrossRef]
- Boffito, M.; Fox, J.; Bowman, C.; Fisher, M.; Orkin, C.; Wilkins, E.; Jackson, A.; Pleguezuelos, O.; Robinson, S.; Stoloff, G.A.; et al. Safety, Immunogenicity and Efficacy Assessment of HIV Immunotherapy in a Multi-Centre, Double-Blind, Randomised, Placebo-Controlled Phase Ib Human Trial. Vaccine 2013, 31, 5680–5686. [CrossRef]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical Applications of DNA Vaccines: Current Progress. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 53, 296–302. [CrossRef]
- van Diepen, M.T.; Chapman, R.; Douglass, N.; Galant, S.; Moore, P.L.; Margolin, E.; Ximba, P.; Morris, L.; Rybicki, E.P.; Williamson, A.-L. Prime-Boost Immunizations with DNA, Modified Vaccinia Virus Ankara, and Protein-Based Vaccines Elicit Robust HIV-1 Tier 2 Neutralizing Antibodies against the CAP256 Superinfecting Virus. J. Virol. 2019, 93, e02155-18. [CrossRef]
- Lisziewicz, J.; Calarota, S.A.; Lori, F. The Potential of Topical DNA Vaccines Adjuvanted by Cytokines. Expert Opin. Biol. Ther. 2007, 7, 1563–1574. [CrossRef]
- Felber, B.K.; Valentin, A.; Rosati, M.; Bergamaschi, C.; Pavlakis, G.N. HIV DNA Vaccine: Stepwise Improvements Make a Difference. Vaccines 2014, 2, 354–379. [CrossRef]
- Munson, P.; Liu, Y.; Bratt, D.; Fuller, J.T.; Hu, X.; Pavlakis, G.N.; Felber, B.K.; Mullins, J.I.; Fuller, D.H. Therapeutic Conserved Elements (CE) DNA Vaccine Induces Strong T-Cell Responses against Highly Conserved Viral Sequences during Simian-Human Immunodeficiency Virus Infection. Hum. Vaccines Immunother. 2018, 14, 1820–1831. [CrossRef]
- Colby, D.J.; Sarnecki, M.; Barouch, D.H.; Tipsuk, S.; Stieh, D.J.; Kroon, E.; Schuetz, A.; Intasan, J.; Sacdalan, C.; Pinyakorn, S.; et al. Safety and Immunogenicity of Ad26 and MVA Vaccines in Acutely Treated HIV and Effect on Viral Rebound after Antiretroviral Therapy Interruption. Nat. Med. 2020, 26, 498–501. [CrossRef]
- Jacobson, J.M.; Zheng, L.; Wilson, C.C.; Tebas, P.; Matining, R.M.; Egan, M.A.; Eldridge, J.; Landay, A.L.; Clifford, D.B.; Luetkemeyer, A.F.; et al. The Safety and Immunogenicity of an Interleukin-12–Enhanced Multiantigen DNA Vaccine Delivered by Electroporation for the Treatment of HIV-1 Infection. JAIDS J. Acquir. Immune Defic. Syndr. 2016, 71, 163–171. [CrossRef]
- Tebas, P.; Ramirez, L.; Morrow, M.; Yan, J.; Shah, D.; Lee, J.; Weiner, D.; Boyer, J.; Bagarazzi, M.; Sardesai, N. Potent Cellular Immune Responses after Therapeutic Immunization of HIV-Positive Patients with the PENNVAX®-B DNA Vaccine in a Phase I Trial. Retrovirology 2012, 9, P276, 1742-4690-9-S2-P276. [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [CrossRef]
- Riddler, S. A Phase I Study to Evaluate the Safety, Tolerability and Immunogenicity of a Therapeutic HIV Vaccine Composed of Autologous Dendritic Cells Loaded with Autologous Inactivated Whole Virus or Conserved Peptides in ART-Treated HIV-Infected Adults; clinicaltrials.gov, 2024;
- GeneCure Biotechnologies A Phase I Dose-Escalation Clinical Trial to Evaluate the Safety and Immunogenicity of a Replication-Defective HIV-1 Vaccine (HIVAXTM) in HIV-1 Infected Subjects Receiving Highly Active Antiretroviral Therapy; clinicaltrials.gov, 2020;
- ANRS, Emerging Infectious Diseases A Phase I/II Randomised Therapeutic HIV Vaccine Trial in Individuals Who Started Antiretrovirals During Primary or Chronic Infection; clinicaltrials.gov, 2019;
- Theravectys S.A. A Multi-Center, Randomized, Double-Blind, Placebo-Controlled Phase I/II Trial to Compare the Safety, Tolerability and Immunogenicity of the Therapeutic THV01 Vaccination at 5.10E+6 TU (Transducing Unit) , 5.10E+7 TU (Transducing Unit) or 5.10E+8 TU (Transducing Unit) Doses to Placebo in HIV-1 Clade B Infected Patients Under Highly Active Antiretroviral Therapy (HAART); clinicaltrials.gov, 2019;
- Janssen Vaccines & Prevention B.V. A Safety, Tolerability and Immunogenicity Study of 2 Different Regimens of Tetravalent Ad26.Mos4.HIV Prime Followed by Boost With MVA-Mosaic OR Ad26.Mos4.HIV Plus a Combination of Mosaic and Clade C Gp140 Protein in HIV-1 Infected Adults on Suppressive ART; clinicaltrials.gov, 2022;
- McCune, J.M.; Rabin, L.B.; Feinberg, M.B.; Lieberman, M.; Kosek, J.C.; Reyes, G.R.; Weissman, I.L. Endoproteolytic Cleavage of Gp160 Is Required for the Activation of Human Immunodeficiency Virus. Cell 1988, 53, 55–67. [CrossRef]
- Zhu, P.; Chertova, E.; Bess, J.; Lifson, J.D.; Arthur, L.O.; Liu, J.; Taylor, K.A.; Roux, K.H. Electron Tomography Analysis of Envelope Glycoprotein Trimers on HIV and Simian Immunodeficiency Virus Virions. Proc. Natl. Acad. Sci. 2003, 100, 15812–15817. [CrossRef]
- Caillat, C.; Guilligay, D.; Torralba, J.; Friedrich, N.; Nieva, J.L.; Trkola, A.; Chipot, C.J.; Dehez, F.L.; Weissenhorn, W. Structure of HIV-1 Gp41 with Its Membrane Anchors Targeted by Neutralizing Antibodies. eLife 2021, 10, e65005. [CrossRef]
- Parker Miller, E.; Finkelstein, M.T.; Erdman, M.C.; Seth, P.C.; Fera, D. A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target. Viruses 2021, 13, 1774. [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV Gp120 Envelope Glycoprotein in Complex with the CD4 Receptor and a Neutralizing Human Antibody. Nature 1998, 393, 648–659. [CrossRef]
- Da, L.-T.; Quan, J.-M.; Wu, Y.-D. Understanding of the Bridging Sheet Formation of HIV-1 Glycoprotein Gp120. J. Phys. Chem. B 2009, 113, 14536–14543. [CrossRef]
- Chen, B.; Vogan, E.M.; Gong, H.; Skehel, J.J.; Wiley, D.C.; Harrison, S.C. Structure of an Unliganded Simian Immunodeficiency Virus Gp120 Core. Nature 2005, 433, 834–841. [CrossRef]
- Bartesaghi, A.; Merk, A.; Borgnia, M.J.; Milne, J.L.S.; Subramaniam, S. Prefusion Structure of Trimeric HIV-1 Envelope Glycoprotein Determined by Cryo-Electron Microscopy. Nat. Struct. Mol. Biol. 2013, 20, 1352–1357. [CrossRef]
- Tamm, L.K.; Lee, J.; Liang, B. Capturing Glimpses of an Elusive HIV Gp41 Prehairpin Fusion Intermediate. Structure 2014, 22, 1225–1226. [CrossRef]
- Pan, R.; Gorny, M.K.; Zolla-Pazner, S.; Kong, X.-P. The V1V2 Region of HIV-1 Gp120 Forms a Five-Stranded Beta Barrel. J. Virol. 2015, 89, 8003–8010. [CrossRef]
- Gorman, J.; Soto, C.; Yang, M.M.; Davenport, T.M.; Guttman, M.; Bailer, R.T.; Chambers, M.; Chuang, G.-Y.; DeKosky, B.J.; Doria-Rose, N.A.; et al. Structures of HIV-1 Env V1V2 with Broadly Neutralizing Antibodies Reveal Commonalities That Enable Vaccine Design. Nat. Struct. Mol. Biol. 2016, 23, 81–90. [CrossRef]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.-P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 Gp120 V1/V2 Domain with Broadly Neutralizing Antibody PG9. Nature 2011, 480, 336–343. [CrossRef]
- Löving, R.; Sjöberg, M.; Wu, S.-R.; Binley, J.M.; Garoff, H. Inhibition of the HIV-1 Spike by Single-PG9/16-Antibody Binding Suggests a Coordinated-Activation Model for Its Three Protomeric Units. J. Virol. 2013, 87, 7000–7007. [CrossRef]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell Binding and Entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866–a006866. [CrossRef]
- Sok, D.; Pauthner, M.; Briney, B.; Lee, J.H.; Saye-Francisco, K.L.; Hsueh, J.; Ramos, A.; Le, K.M.; Jones, M.; Jardine, J.G.; et al. A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity 2016, 45, 31–45. [CrossRef]
- Wang, Z.; Barnes, C.O.; Gautam, R.; Cetrulo Lorenzi, J.C.; Mayer, C.T.; Oliveira, T.Y.; Ramos, V.; Cipolla, M.; Gordon, K.M.; Gristick, H.B.; et al. A Broadly Neutralizing Macaque Monoclonal Antibody against the HIV-1 V3-Glycan Patch. eLife 2020, 9, e61991. [CrossRef]
- Bonsignori, M.; Kreider, E.F.; Fera, D.; Meyerhoff, R.R.; Bradley, T.; Wiehe, K.; Alam, S.M.; Aussedat, B.; Walkowicz, W.E.; Hwang, K.-K.; et al. Staged Induction of HIV-1 Glycan–Dependent Broadly Neutralizing Antibodies. Sci. Transl. Med. 2017, 9, eaai7514. [CrossRef]
- Kumar, S.; Ju, B.; Shapero, B.; Lin, X.; Ren, L.; Zhang, L.; Li, D.; Zhou, Z.; Feng, Y.; Sou, C.; et al. A VH 1-69 Antibody Lineage from an Infected Chinese Donor Potently Neutralizes HIV-1 by Targeting the V3 Glycan Supersite. Sci. Adv. 2020, 6, eabb1328. [CrossRef]
- Simonich, C.A.; Williams, K.L.; Verkerke, H.P.; Williams, J.A.; Nduati, R.; Lee, K.K.; Overbaugh, J. HIV-1 Neutralizing Antibodies with Limited Hypermutation from an Infant. Cell 2016, 166, 77–87. [CrossRef]
- Travers, S.A. Conservation, Compensation, and Evolution of N-Linked Glycans in the HIV-1 Group M Subtypes and Circulating Recombinant Forms. ISRN AIDS 2012, 2012, 1–9. [CrossRef]
- Kong, L.; Ju, B.; Chen, Y.; He, L.; Ren, L.; Liu, J.; Hong, K.; Su, B.; Wang, Z.; Ozorowski, G.; et al. Key Gp120 Glycans Pose Roadblocks to the Rapid Development of VRC01-Class Antibodies in an HIV-1-Infected Chinese Donor. Immunity 2016, 44, 939–950. [CrossRef]
- Schommers, P.; Gruell, H.; Abernathy, M.E.; Tran, M.-K.; Dingens, A.S.; Gristick, H.B.; Barnes, C.O.; Schoofs, T.; Schlotz, M.; Vanshylla, K.; et al. Restriction of HIV-1 Escape by a Highly Broad and Potent Neutralizing Antibody. Cell 2020, 180, 471-489.e22. [CrossRef]
- Jia, M.; Liberatore, R.A.; Guo, Y.; Chan, K.-W.; Pan, R.; Lu, H.; Waltari, E.; Mittler, E.; Chandran, K.; Finzi, A.; et al. VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA. Cell Host Microbe 2020, 27, 963-975.e5. [CrossRef]
- Zhou, T.; Zheng, A.; Baxa, U.; Chuang, G.-Y.; Georgiev, I.S.; Kong, R.; O’Dell, S.; Shahzad-ul-Hussan, S.; Shen, C.-H.; Tsybovsky, Y.; et al. A Neutralizing Antibody Recognizing Primarily N-Linked Glycan Targets the Silent Face of the HIV Envelope. Immunity 2018, 48, 500-513.e6. [CrossRef]
- Scharf, L.; Scheid, J.F.; Lee, J.H.; West, A.P.; Chen, C.; Gao, H.; Gnanapragasam, P.N.P.; Mares, R.; Seaman, M.S.; Ward, A.B.; et al. Antibody 8ANC195 Reveals a Site of Broad Vulnerability on the HIV-1 Envelope Spike. Cell Rep. 2014, 7, 785–795. [CrossRef]
- Dubrovskaya, V.; Tran, K.; Ozorowski, G.; Guenaga, J.; Wilson, R.; Bale, S.; Cottrell, C.A.; Turner, H.L.; Seabright, G.; O’Dell, S.; et al. Vaccination with Glycan-Modified HIV NFL Envelope Trimer-Liposomes Elicits Broadly Neutralizing Antibodies to Multiple Sites of Vulnerability. Immunity 2019, 51, 915-929.e7. [CrossRef]
- Lee, J.H.; Leaman, D.P.; Kim, A.S.; Torrents De La Peña, A.; Sliepen, K.; Yasmeen, A.; Derking, R.; Ramos, A.; De Taeye, S.W.; Ozorowski, G.; et al. Antibodies to a Conformational Epitope on Gp41 Neutralize HIV-1 by Destabilizing the Env Spike. Nat. Commun. 2015, 6, 8167. [CrossRef]
- Pancera, M.; Zhou, T.; Druz, A.; Georgiev, I.S.; Soto, C.; Gorman, J.; Huang, J.; Acharya, P.; Chuang, G.-Y.; Ofek, G.; et al. Structure and Immune Recognition of Trimeric Pre-Fusion HIV-1 Env. Nature 2014, 514, 455–461. [CrossRef]
- Harrison, S.C. Viral Membrane Fusion. Nat. Struct. Mol. Biol. 2008, 15, 690–698. [CrossRef]
- Kong, R.; Xu, K.; Zhou, T.; Acharya, P.; Lemmin, T.; Liu, K.; Ozorowski, G.; Soto, C.; Taft, J.D.; Bailer, R.T.; et al. Fusion Peptide of HIV-1 as a Site of Vulnerability to Neutralizing Antibody. Science 2016, 352, 828–833. [CrossRef]
- Torrents De La Peña, A.; Rantalainen, K.; Cottrell, C.A.; Allen, J.D.; Van Gils, M.J.; Torres, J.L.; Crispin, M.; Sanders, R.W.; Ward, A.B. Similarities and Differences between Native HIV-1 Envelope Glycoprotein Trimers and Stabilized Soluble Trimer Mimetics. PLOS Pathog. 2019, 15, e1007920. [CrossRef]
- Irimia, A.; Sarkar, A.; Stanfield, R.L.; Wilson, I.A. Crystallographic Identification of Lipid as an Integral Component of the Epitope of HIV Broadly Neutralizing Antibody 4E10. Immunity 2016, 44, 21–31. [CrossRef]
- Shipulin, G.A.; Glazkova, D.V.; Urusov, F.A.; Belugin, B.V.; Dontsova, V.; Panova, A.V.; Borisova, A.A.; Tsyganova, G.M.; Bogoslovskaya, E.V. Triple Combinations of AAV9-Vectors Encoding Anti-HIV bNAbs Provide Long-Term In Vivo Expression of Human IgG Effectively Neutralizing Pseudoviruses from HIV-1 Global Panel. Viruses 2024, 16, 1296. [CrossRef]
- Leontyev D.S., Urusov F.A., Glazkova D.V., Belugin B.V., Orlova O.V., Mintaev R.R., Tsyganova G.M., Bogoslovskaya E.V., Shipulin G.A. Study of the protective efficacy of CombiMab-2 against human immunodeficiency virus type 1 in mice humanised with CD4+ T-lymphocytes. Biological Products. Prevention, Diagnosis, Treatment. 2024;24(3):312-321. (In Russ.). [CrossRef]
Figure 1.
The structure of HIV with vaccine targets. Created with Biorender.com.
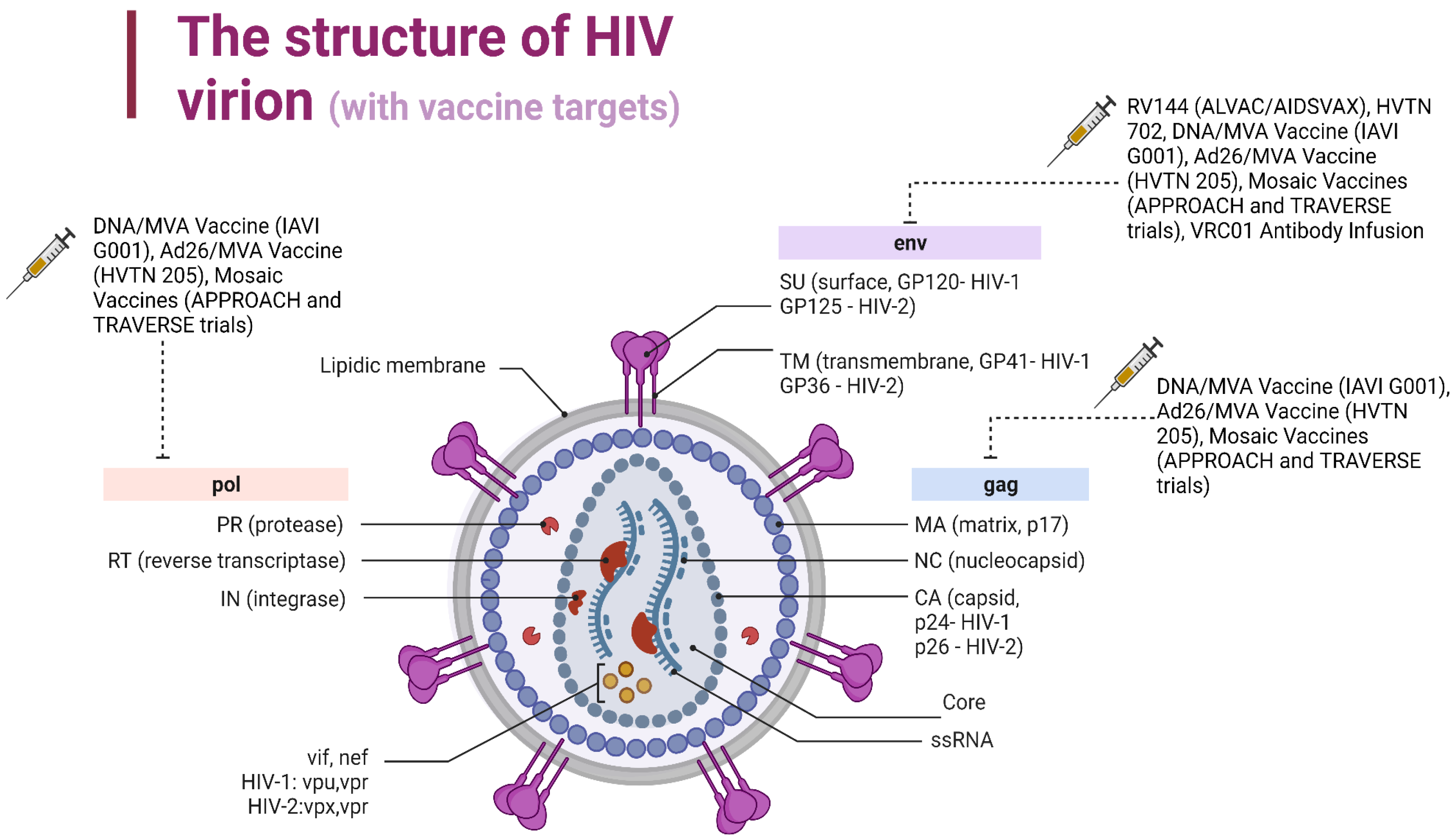
Figure 2.
The mechanism of HIV vaccines aimed at stimulating CTLs. Perforins form a pore in the plasma membrane. Through this pore Granzyme B (GrB) enters the cell. Granzyme B activates procaspase 7. Caspase 7 starts proteolytically destroying the cell proteins leading to cell death. Antiapoptotic proteins B-cell lymphoma 2 (Bcl2) and BCl-XL bind to Bax and Bcl-2 homologous antagonist killer (Bak), preventing their oligomerization. Granzyme B proteolytically destroys Bcl2 and BCl-XL proteins and thus creates conditions for cytochrome c release into the cell cytosol. Cytochrome c binds to the Apoptotic protease activating factor 1 (APAF1) molecule. Further, about 7 APAF1 molecules with active centers bound by cytochrome c bind to each other using their own caspase recruitment domain (CARD domains), forming the apoptosome. Apoptosome activates initiator caspase 9, which in turn activates executioner caspases 3 and 6. Caspase 3 and Caspase 6 produce proteolysis of the cell proteins, which leads to cell death.
Figure 2.
The mechanism of HIV vaccines aimed at stimulating CTLs. Perforins form a pore in the plasma membrane. Through this pore Granzyme B (GrB) enters the cell. Granzyme B activates procaspase 7. Caspase 7 starts proteolytically destroying the cell proteins leading to cell death. Antiapoptotic proteins B-cell lymphoma 2 (Bcl2) and BCl-XL bind to Bax and Bcl-2 homologous antagonist killer (Bak), preventing their oligomerization. Granzyme B proteolytically destroys Bcl2 and BCl-XL proteins and thus creates conditions for cytochrome c release into the cell cytosol. Cytochrome c binds to the Apoptotic protease activating factor 1 (APAF1) molecule. Further, about 7 APAF1 molecules with active centers bound by cytochrome c bind to each other using their own caspase recruitment domain (CARD domains), forming the apoptosome. Apoptosome activates initiator caspase 9, which in turn activates executioner caspases 3 and 6. Caspase 3 and Caspase 6 produce proteolysis of the cell proteins, which leads to cell death.
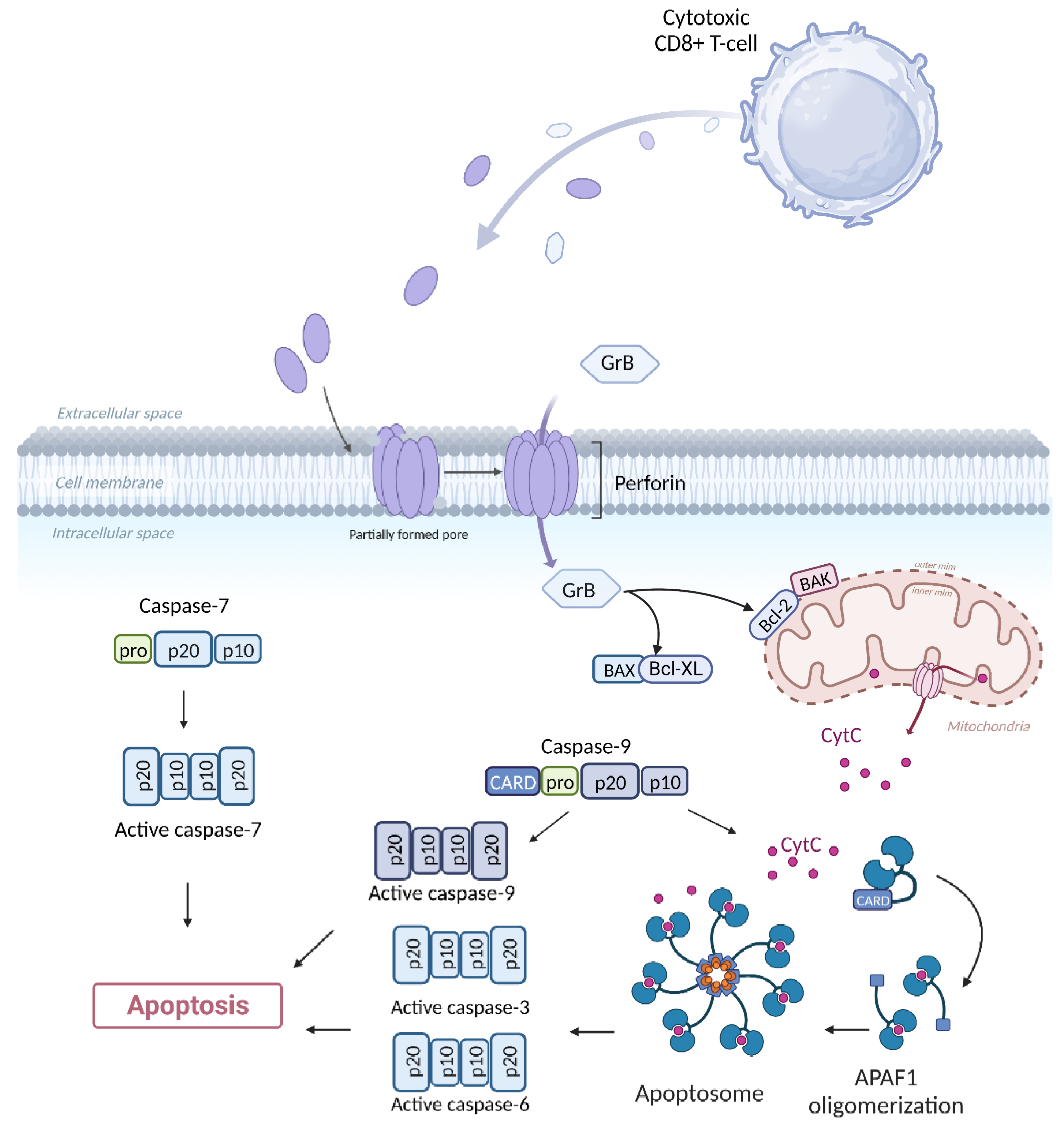
Figure 3.
The mechanism of HIV vaccines aimed at chemokine-mediated suppression. CCL4 – chemokine (C-C motif) ligands 4; CCR5 – C-C chemokine receptor type 5; GRK2 - G-protein-coupled receptor kinase 2; AP-2 - Adaptor protein complex; GTP – Guanosine triphosphate; GDP – Guanosine diphosphate. Created with Biorender.com.
Figure 3.
The mechanism of HIV vaccines aimed at chemokine-mediated suppression. CCL4 – chemokine (C-C motif) ligands 4; CCR5 – C-C chemokine receptor type 5; GRK2 - G-protein-coupled receptor kinase 2; AP-2 - Adaptor protein complex; GTP – Guanosine triphosphate; GDP – Guanosine diphosphate. Created with Biorender.com.
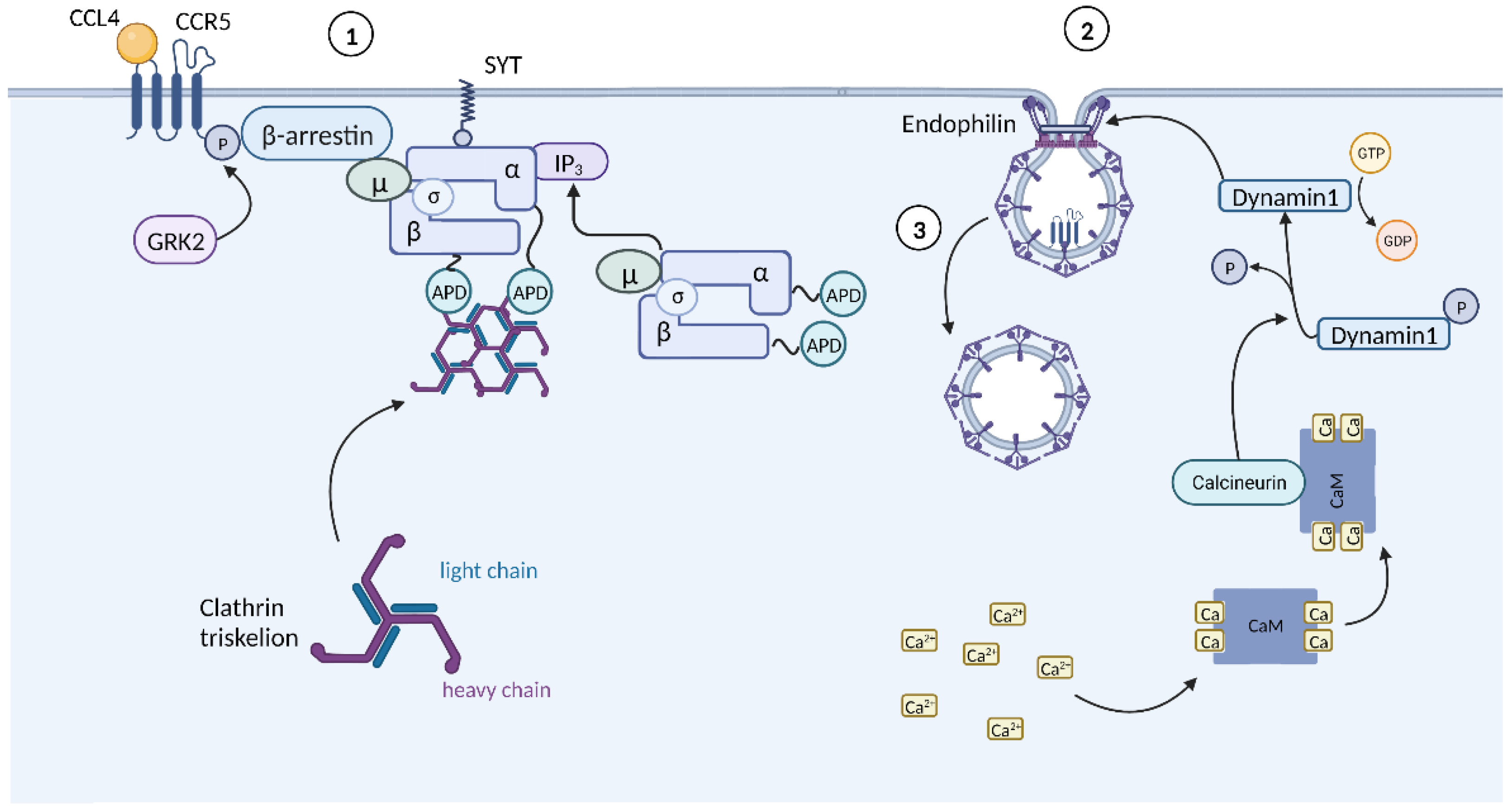
Figure 4.
The mechanism of mRNA HIV vaccine. BCR - B cell antigen receptor; SH2 - SRC homology 2; BLNK - B-cell linker protein; BTK - Bruton's tyrosine kinase; PIP3 - phosphatidylinositol (3,4,5)-trisphosphate; PH - pleckstrin homology; IP3 - inositol triphosphate; DAG – diacylglycerol; PLCγ2 - phospholipase Cγ2; PIP-2 - phosphotidylinositol diphosphate; PKCβ - Proteinkinase Cβ; NFAT - Nuclear factor of activated T-cells; CARMA11 - caspase recruitment domain family member 11; MALT1 – Mucosa-associated lymphoid tissue lymphoma translocation protein 1; BCL-10 - B cell CLL/lymphoma 10; TRAF6 - tumor necrosis factor receptor-associated factor 6; IKK - IκB kinase; NF-κb - Nuclear Factor kappa b; BCAP - B cell adaptor molecule for PI3K. Created with Biorender.com.
Figure 4.
The mechanism of mRNA HIV vaccine. BCR - B cell antigen receptor; SH2 - SRC homology 2; BLNK - B-cell linker protein; BTK - Bruton's tyrosine kinase; PIP3 - phosphatidylinositol (3,4,5)-trisphosphate; PH - pleckstrin homology; IP3 - inositol triphosphate; DAG – diacylglycerol; PLCγ2 - phospholipase Cγ2; PIP-2 - phosphotidylinositol diphosphate; PKCβ - Proteinkinase Cβ; NFAT - Nuclear factor of activated T-cells; CARMA11 - caspase recruitment domain family member 11; MALT1 – Mucosa-associated lymphoid tissue lymphoma translocation protein 1; BCL-10 - B cell CLL/lymphoma 10; TRAF6 - tumor necrosis factor receptor-associated factor 6; IKK - IκB kinase; NF-κb - Nuclear Factor kappa b; BCAP - B cell adaptor molecule for PI3K. Created with Biorender.com.
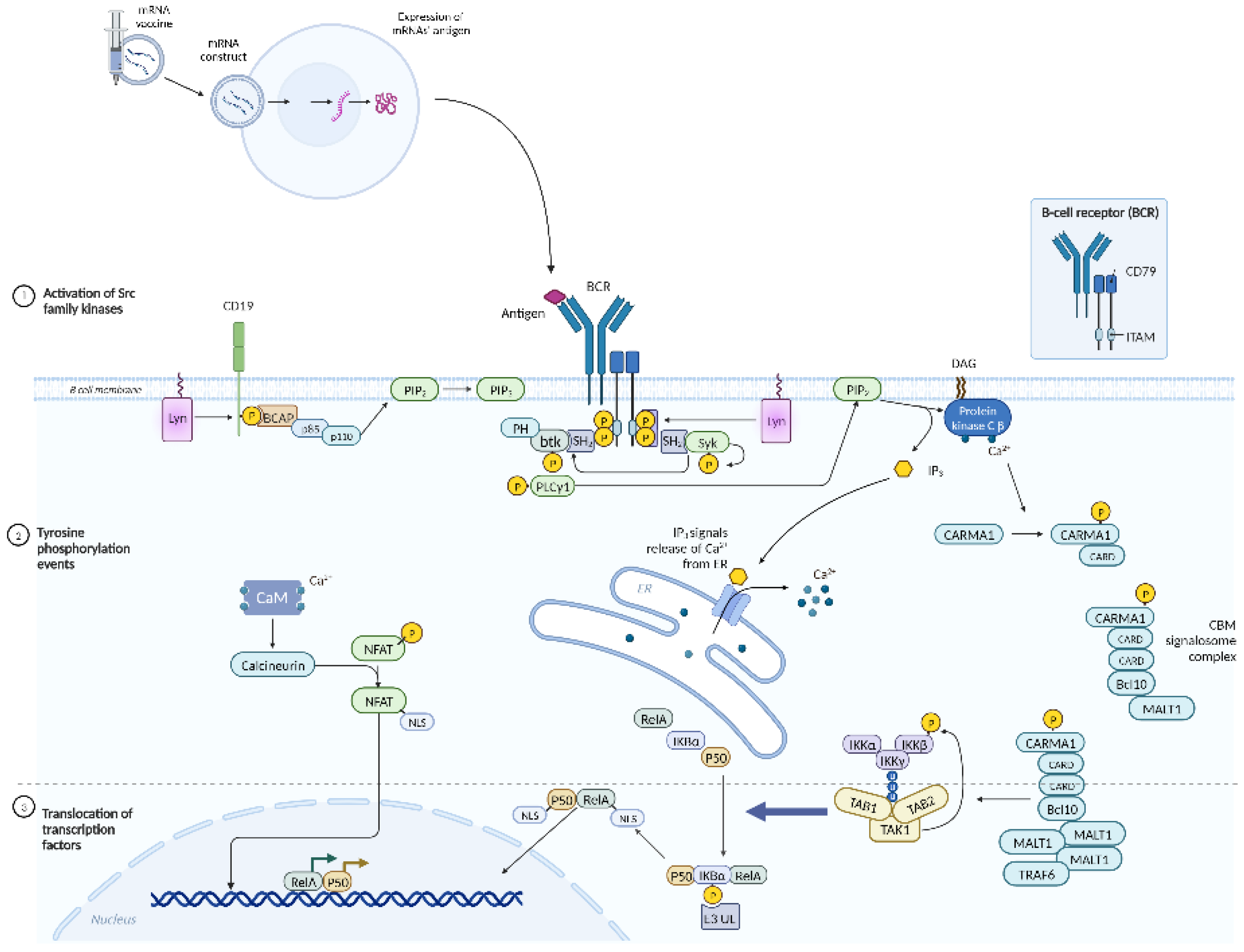
Figure 5.
The mechanism of dendritic cell-based HIV vaccine. Lck - lymphocyte-specific protein tyrosine kinase; CSK - C-terminal Src kinase; PAG1 - Phosphoprotein associated with glycosphingolipid-enriched microdomains 1; ITAMs - immunoreceptor tyrosine-based activation motifs; ZAP-70 binds to the phosphorylated tyrosine residues of CD3; LAT - linker for activation of T cells; GADS - GRB2-related adaptor protein of Shc; GRB2 - growth-factor receptor-bound protein 2; PLCγ1 - phospholipase Cγ1; SOS - Son of Sevenless; RAF-1 - proto-oncogene serine/threonine-protein kinase; GDP – Guanosine diphosphate; GTP – Guanosine triphosphate; MEK - MAPK/ERK kinase; MAPK - mitogen-activated protein kinase; ITK - interleukin-2-inducible T-cell kinase; SLP-76 - SH2-domain-containing leukocyte protein 76 kDa; RASGRP1 - guanyl nucleotide-releasing protein RAS; DAG - diacylglycerol; NFAT - Nuclear factor of activated T-cells. Created with Biorender.com.
Figure 5.
The mechanism of dendritic cell-based HIV vaccine. Lck - lymphocyte-specific protein tyrosine kinase; CSK - C-terminal Src kinase; PAG1 - Phosphoprotein associated with glycosphingolipid-enriched microdomains 1; ITAMs - immunoreceptor tyrosine-based activation motifs; ZAP-70 binds to the phosphorylated tyrosine residues of CD3; LAT - linker for activation of T cells; GADS - GRB2-related adaptor protein of Shc; GRB2 - growth-factor receptor-bound protein 2; PLCγ1 - phospholipase Cγ1; SOS - Son of Sevenless; RAF-1 - proto-oncogene serine/threonine-protein kinase; GDP – Guanosine diphosphate; GTP – Guanosine triphosphate; MEK - MAPK/ERK kinase; MAPK - mitogen-activated protein kinase; ITK - interleukin-2-inducible T-cell kinase; SLP-76 - SH2-domain-containing leukocyte protein 76 kDa; RASGRP1 - guanyl nucleotide-releasing protein RAS; DAG - diacylglycerol; NFAT - Nuclear factor of activated T-cells. Created with Biorender.com.
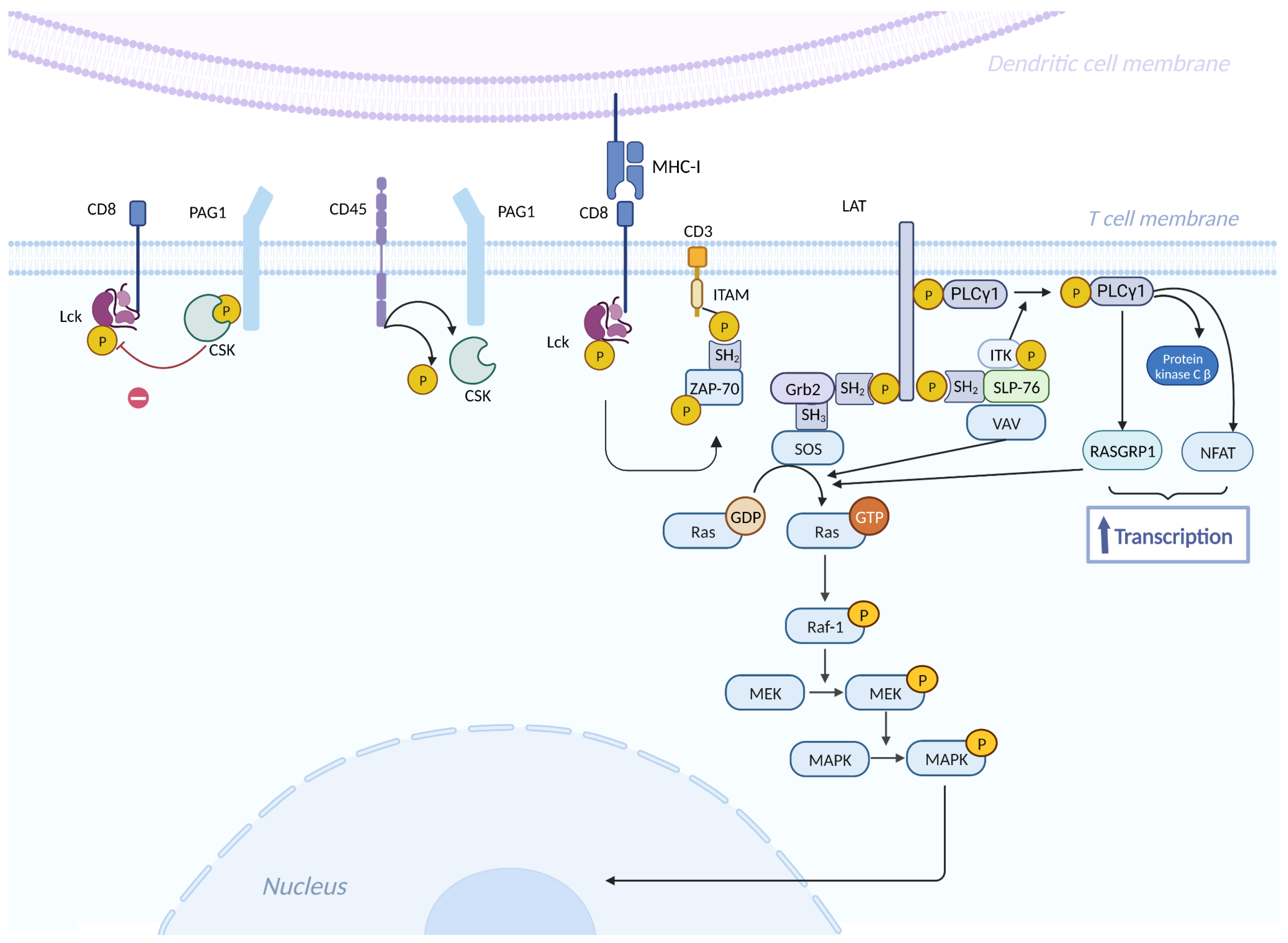
Figure 6.
The mechanism of DNA-based HIV vaccines. JAK-1 - Janus kinase 1; JAK-3 - Janus kinase 3; STAT5 - Signal transducer and activator of transcription 5. Created with Biorender.com.
Figure 6.
The mechanism of DNA-based HIV vaccines. JAK-1 - Janus kinase 1; JAK-3 - Janus kinase 3; STAT5 - Signal transducer and activator of transcription 5. Created with Biorender.com.
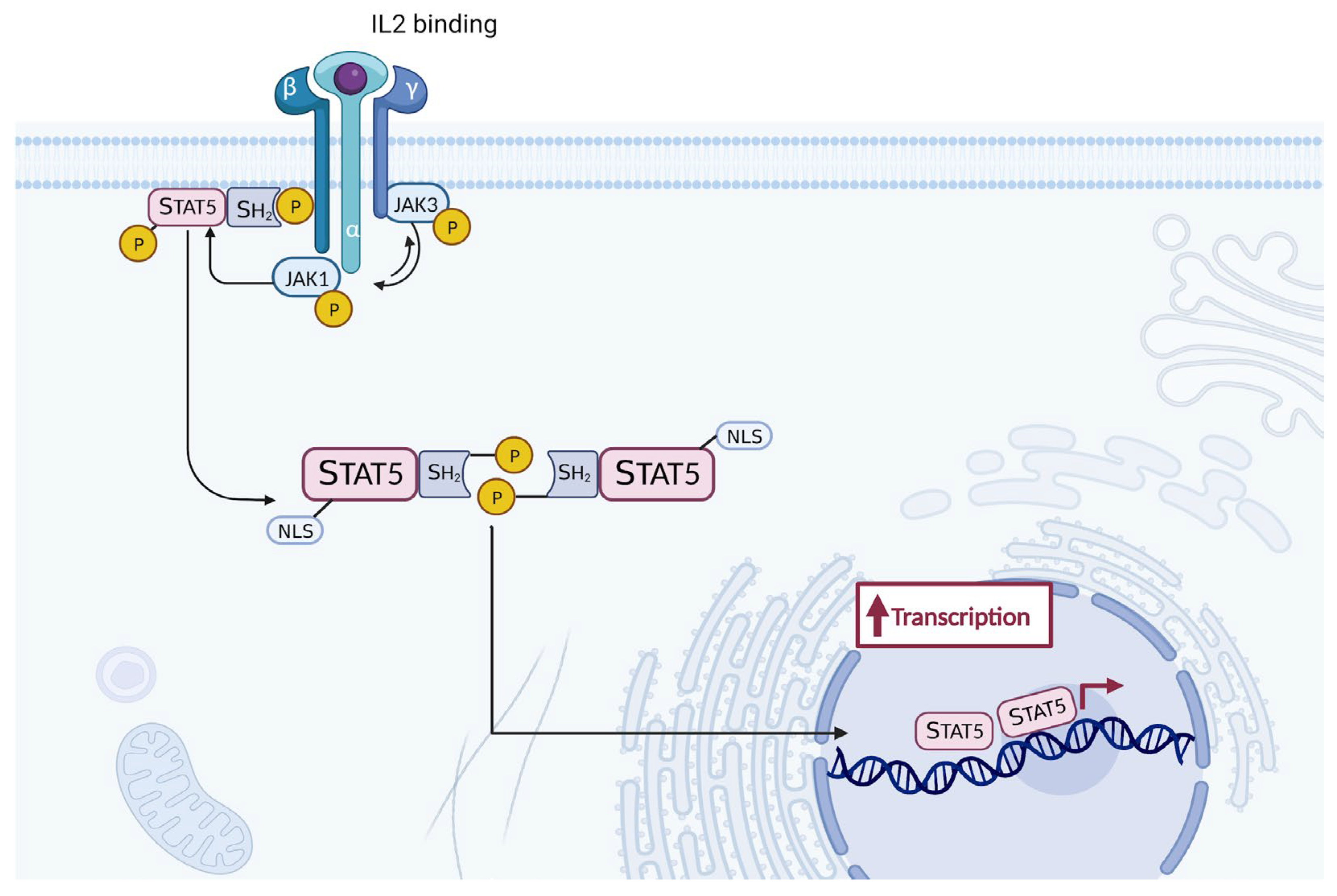

Table 1.
HIV genes and their functions.
| Gene | Encoded protein | Function | References |
| 1. Structural genes | |||
| gag (group antigen) | capsid protein (p24) matrix protein (p17) nucleoprotein (p7) p6 |
forms conical capsid covers the conical capsid binds the RNA and forms nucleoprotein/RNA complex facilitates the budding process of virions |
[29,30,31] |
| pol (polyprotein) | reverse transcriptase (RT, p51) ribonuclease H (RNase H, p15), RNase plus RT (p66) protease (PR, p10) integrase (IN, p32) |
transforms viral RNA into DNA degrades the viral RNA processes Gag and Gag-Pol protein precursors into mature viral proteins incorporates viral DNA into the host genome |
[32] |
| env (envelope) | surface protein (SU, gp120) transmembrane protein (TM, gp41) |
contains a domain responsible for binding the virus to the CD4 receptors and co-receptors facilitates the fusion of the viral envelope with the cellular membrane through anchoring |
[24] |
| 2. Regulatory genes | |||
| tat (transactivator of transcription) | transactivator protein (p14) | activates the viral transcription | [33,34] |
| rev (regulator of expression of virion proteins) | RNA splicing regulator (p19) | regulates the nuclear export of unspliced and partially spliced mRNAs from nucleus to the cytoplasm | [35] |
| 3. Auxiliary genes | |||
| nef (negative factor) | negative regulating factor (p27) | modulates cellular signaling, increases the downregulation of the CD4 receptors and MHC I, allows viral replication | [36,37] |
| vif (virion infectivity factor) | viral infectivity protein (p23) | promotes the viral infectivity by preventing restriction factor APOBEC3G from acting | [38,39] |
| vpu | virus protein unique (p16) | causes the degradation of CD4, viral particle release and the arrest of the cell cycle | [38,40] |
| vpr | virus protein r (p15) | facilitates viral infectivity and affects the cell cycle | [41,42] |
Table 2.
Clinical studies based on RV-144 results.
| Study | Vaccine (Immunogen) | Location (site) | Target group | Date | Efficacy | References |
|---|---|---|---|---|---|---|
| HVTN 305 (NCT05781542) | ALVAC-HIV and AIDSVAX B/E | Thailand (Clade B) | 162 women and men | 2012-2017 | No | [86] |
| HVTN 306 | ALVAC-HIV and AIDSVAX B/E | Thailand (Clade B) | 360 men and women aged 20–40 years |
2012-2017 | No | [87] |
| HVTN 097 (NCT02109354) | ALVAC-HIV (vCP1521) and AIDSVAX B/E | South Africa (Clade B/E) | 100 black Africans (men and women) aged 18–40 years | 2012-2013 | No | [88] |
| HVTN 100 (NCT02404311) | ALVAC-HIV (vCP2438) and bivalent subtype C gp120/MF59 | South Africa (Clade C) | 252 men and women | 2015-2018 | No | [89] |
Table 3.
Largest clinical study of mosaic HIV vaccines.
| Study | Vaccine (Immunogen) | Location (site) | Target group | Date | Efficacy | References |
|---|---|---|---|---|---|---|
| VaxSyn | Recombinant envelope glycoprotein subunit (rgp160) of HIV | Canada (Clade B) | 72 adults | 1987 | No | [91] |
| HIVAC-1e | Recombinant vaccinia virus designed to express HIV gp160 | USA (Clade B) | 35 male adults | 1988 | No | [92] |
| VAX003 (VaxGen) (NCT00006327) | AIDSVAX B/E (subtype B - MN; subtype AE - A244 rgp120) | Thailand (Clade B/E) | 2,545 men and women IDUs |
1998-2002 | No | [93] |
| VAX004 (VaxGen) (NCT00002441) | AIDSVAX B/B (subtype B - MN and GNE8 rgp120) | North America (Clade B) | 5,417 MSM and 300 women |
1999-2003 | No | [94] |
| STEP HVTN502 | Ad5 expressing subtype B Gag (CAM-1), Pol (IIIB), Nef (JR-FL) | North America the Caribbean South America, and Australia (Clade B) | 3,000 MSM and heterosexual men and women |
2004-2007 | No | [95] |
| Phambili HVTN 503 (NCT00413725) | Ad5 expressing subtype B Gag (CAM-1), Pol (IIIB), Nef (JR-FL) | South Africa (Clade C) | 801 adults | 2003-2007 | No | [96] |
| RV 144 (NCT00223080) | ALVAC-HIV (vCP1521) expressing Gag and Pro (subtype B LAI), CRF01_AE gp120 (92TH023) linked to transmembrane anchoring portion of gp41 (LAI) AIDSVAX B/E Aluminium hydroxide |
Thailand (Clade B) | 16,402 community-risk men and women |
2003-2009 | Yes 31% | [97] |
| HVTN 505 (NCT00865566) | 6 DNA plasmids - subtype B Gag, Pol, Nef and subtypes A, B and C Env 4 rAd5 vectors - subtype B Gag/Pol and subtypes A, B and C Env |
United States (Clade B) | 2,504 men or transgender women who have sex with men | 2009-2017 | No | [98] |
| Uhambo HVTN 702 (NCT02968849) | ALVAC-HIV (vCP2438) expressing Gag and Pro (subtype B LAI), subtype C gp120 (ZM96.C) linked to transmembrane anchoring portion of gp41 (LAI) | South Africa (Clade C) | 5,400 men and women | 2016-2021 | No | [99] |
| IMBOKODO HVTN 705 (NCT03060629) | Ad26.Mos4.HIV Subtype C gp140 |
Sub-Saharan Africa (Clade C) | 2600 women | 2017-2022 | No data | [100] |
| MOSAICO HVTN 706 (NCT03964415) | Ad26.Mos4.HIV Subtype C gp140 or bivalent gp140 (subtype C/Mosaic) |
Europe North America and South America (Clade C) | 3800 MSM and transgender persons | 2019-2024 | No data | [101] |
| PrEPVacc (NCT04066881) | DNA-HIV-PT123 plasmid and AIDSVAX B/E or DNA-HIV-PT123 plasmid with trimeric CN54gp140; MVA-CMDR (Chang Mai double recombinant) and trimeric CN54gp140 Concurrent PrEP administration of either TAF/FTC or TDF/FTC |
Uganda, Tanzania, Mozambique, Republic of South Africa (Clade C) | 1668 Adults Men and Women | 2020-2023 | No data | [102] |
Table 4.
Ongoing trials of preventive mRNA HIV vaccines.
| Trial | Name | Hypothesis | Year | Target group | Site | Vaccine Candidates | Immunogene design | Vaccine Manufacturer | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| IAVI G002 NCT05001373 |
A Phase 1 Study to Evaluate the Safety A Phase I Trial to and Immunogenicity of eOD-GT8 and Immunogenic 60mer mRNA Vaccine (mRNA-1644) delivered by an m and Core-g28v2 60mer mRNA Vaccine negative adults (mRNA-1644v2-Core |
Sequential vaccination by a germline-targeting prime followed by directional boost immunogens can induce specific classes of B-cell responses and guide their early maturation toward broadly neutralizing antibody (bnAb) development through an mRNA platform |
2021- 2023 | 56 adults ages 18 to 50 | 4 sites in the US (Atlanta; San Antonio; Seattle; Washington, DC) | Two experimental HIV vaccines based on messenger RNA (mRNA) platform: 1. eOD-GT8 60mer mRNA Vaccine (mRNA-1644) 2. Core-g28v2 60mer mRNA Vaccine (mRNA-1644v2-Core) |
IAVI Neutralizing Antibody Center (NAC) at Scripps Research |
Moderna | [108] |
| IAVI G003 NCT05414786 |
A Phase I Trial to Evaluate the Safety and Immunogenicity of eOD-GT8 60mer delivered by an mRNA platform in HIV negative adults | eOD-GT8 60mer delivered by an mRNA platform in HIV negative adults will induce immune responses in African populations as was seen in IAVI G001, which demonstrated this recombinant protein (eOD-GT8 60mer) safely induced immune responses in 97% of recipients, who were healthy U.S. adults | 2022- 2023 | 18 healthy, HIV- negative adults | 2 sites: Kigali, Rwanda, and Tembisa, South Africa | One experimental HIV vaccine based on messenger RNA (mRNA) platform: 1. eOD-GT8 60mer delivered by an mRNA Vaccine platform (mRNA- 1644) |
IAVI Neutralizing Antibody Center (NAC) at Scripps Research |
Moderna | [109] |
| HVTN 302 NCT05217641 |
A Clinical Trial to Evaluate the Safety and Immunogenicity of BG505 MD39.3, BG505 MD39.3 gp151, and BG505 MD39.3 gp151 CD4KO HIV Trimer mRNA Vaccines in Healthy, HIV-uninfected Adult Participants | The BG505 MD39.3 soluble and membrane-bound trimer mRNA vaccines will be safe and well-tolerated among HIV-uninfected individuals and will elicit autologous neutralizing antibodies | 2022- 2023 | 108 adults ages 18 to 55 years | 11 sites in the US (Birmingham; Boston; Los Angeles; New York City; Philadelphia; Pittsburgh; Rochester; Seattle) |
Three experimental HIV vaccines based on messenger RNA (mRNA) platform: 1. BG505 MD39.3 mRNA 2. BG505 MD39.3 gp151 mRNA 3. BG505 MD39.3 gp151 CD4K0 mRNA |
Scripps Consortium for HIV/AIDS Vaccine Development (CHAVD) and IAVI Neutralizing Antibody Center (NAC) at Scripps Research |
Moderna | [110] |
Table 5.
Ongoing trials of dendritic cell-based HIV vaccines.
| Trial | Phase | Registry Identifier | Result | Status | Last Update | Ref. |
|---|---|---|---|---|---|---|
| AGS-004 (personalized therapeutic vaccine utilizing patient-derived dendritic cells and HIV antigens) | IIb | NCT00672191 | Induction of CD4 and CD8, reduction of VL, One severe adverse event | Completed | 2013 | [114] |
| Autologous HIV-1 ApB DC Vaccine | I/II | NCT00510497 | Safe and immunogenic Reduction of HIV blood reservoir | Completed | 2016 | [115] |
| Dendritic cell vaccine (DCV-2) | I/II | NCT00402142 | Safely induced marginal immune responses, whereas markedly increased Vacc-C5-induced regulatory T cell | Completed | 2014 | [116] |
| Dendritic cells loaded with HIV-1 lipopeptides | I | NCT00796770 | Safe and showed few CD8+ T cell responses | Completed | 2017 | [117] |
Table 6.
Ongoing trials of peptide-based HIV vaccines.
| Trial | Phase | Registry Identifier | Result | Status | Last Update | Ref. |
|---|---|---|---|---|---|---|
| Vacc-4x | II | NCT00659789 | Induction of CD4 and CD8, reduction of VL, One severe adverse event | Completed | 2017 | [125] |
| VAC-3S | I/II | NCT01549119 | Safe and immunogenic Reduction of HIV blood reservoir | Completed | 2015 | [126] |
| Vacc-C5 | I/II | NCT01627678 | Safely induced marginal immune responses, whereas markedly increased Vacc-C5-induced regulatory T cell | Completed | 2014 | [127] |
| AFO-18 | I | NCT01141205 | Safe and showed few CD8 T cell responses | Completed | 2013 | [128] |
| HIV-v | I | NCT01071031 | Safe and can elicit T- and B-cell responses that significantly reduce viral load. | Completed | 2012 | [129] |
Table 7.
Ongoing trials of DNA-based HIV vaccines.
| Trial | Phase | Registry Identifier | Result | Status | Last Update | Ref. |
|---|---|---|---|---|---|---|
| Ad26.Mos.HIV + MVA-Mosaic | II | NCT02919306 | Generated robust immune responses, but did not lead to viremic control after treatment interruption. | Completed | 2022 | [135] |
| MAG pDNA vaccine +/- IL-12 | I | NCT01266616 | Elicited CD4+ but not CD8+ T-cell responses to multiple HIV-1 antigens. | Completed | 2015 | [136] |
| PENNVAX-B (Gag, Pol, Env) + electroporation | I | NCT01082692 | Strong induction of CD8 T cell responses | Completed | 2012 | [137] |
Table 8.
Ongoing trials of viral vector-based HIV vaccines.
| Trial | Phase | Registry Identifier | Result | Status | Last Update | Ref. |
|---|---|---|---|---|---|---|
| DC-HIV04 Comparison of Dendritic Cell-Based Therapeutic Vaccine Strategies for HIV Functional Cure |
I | NCT03758625 | Not reported | Active | 2024 | [139] |
| GCHT01 | I | NCT01428596 | Not reported | Unknown | 2019 | [140] |
| GTU-MultiHIV B-clade + MVA HIV-B (DNA + viral vector vaccines) | II | NCT02972450 | Not reported | Terminated | 2019 | [141] |
| THV01 (lentiviral vector-based therapeutic vaccine) | I/II | NCT02054286 | Not reported | Completed | 2019 | [142] |
| Ad26.Mos4.HIV + MVA-Mosaic or clade C gp140 + mosaic gp140 |
I | NCT03307915 | Not reported | Completed | 2022 | [143] |
Table 9.
bNAbs targeting key regions of HIV Env protein.
| Neutralizing activities of different bNAbs | ||
| Targets of bNAbs | Functions of target | bNAbs |
| V3-glycan | Involved in the initial interaction and resulting fusion between the Env protein and a coreceptor in the host cell membrane. | 10-1074 |
| 10-1074-LS | ||
| 10-1074-LS-J | ||
| PGT121 | ||
| PGT121LS | ||
| PGT121.414.LS | ||
| DH270 | ||
| PGT128 | ||
| PGT135 | ||
| CD4 binding site | Binds to the CD4 receptor on the surface of the host cell, causing structural changes that allow the gp120 protein to bind to the host cell membrane. | 3BNC117 |
| 3BNC117-LS | ||
| 3BNC117-LS-J | ||
| VRC01 | ||
| VRC01-LS | ||
| VRC07-523LS | ||
| 12A12 | ||
| BANC131 | ||
| CH103 | ||
| CH235.12 | ||
| CH31 | ||
| N49P7 | ||
| NIH 45i | ||
| PG04 | ||
| VRC07 | ||
| VRC13 | ||
| V1/V2-glycan | Involved in a structural change to the Env protein enabling HIV to fuse and infect the host cell. | CAP256V2LS |
| PGDM1400 | ||
| CH01-04 | ||
| PG16 | ||
| PGT141-145 | ||
| gp41 MPER | Helps disrupt viral membrane during fusion of HIV with host cell. | 10E8.4 |
| 2F5 | ||
| 4E10 | ||
| 10E8VLS | ||
| Fusion peptide (FP) | Inserts into and disrupts the host cell membrane, allowing HIV to release genetic material into the cell. | ACS202 |
| VRC34.01 | ||
| gp120-41 interface | Where gp120 and gp41 meet; involved in structural changes to the Env protein during entry into the host cell. | 35022 |
| LN02 | ||
| SANC 195 | ||
| 8ANC195 | ||
| PGT151 | ||
| 3BC176 | ||
| 3BC315 | ||
| gp120 Silent face | A region of the Env protein that is heavily sugar-coated and has little functional role in virus entry. | SF12 |
| VRC-PG05 | ||
Table 10.
Ongoing trials of bNAbs against HIV.
| bNAbs combinations: Two or more antibodies in a regimen | |||||||
| Regimen | Status | Route | Research Institution | Trial Name | |||
| 10-1074 + 3BNC117 | Phase 1, Completed | IV | Rockefeller University | YCO-0899 | |||
| 10-1074-LS + 3BNC117-LS | Phase 1, Ongoing | IV, SC | Rockefeller University | YCO-0971 | |||
| 10-1074-LS-J + 3BNC117-LS-J | Phase 1/2, Ongoing | IV, SC | IAVI, Rockefeller University, University of Washington | IAVI C100 | |||
| PGT121 + VRC07-523LS + PGDM1400 | Phase 1, Completed Phase 1/2a, Ongoing | IV | BIDMC, IAVI, NIAID | IAVI T002 IAVI T003 | |||
| PGT121 + PGDM1400 + 10-1074 + VRC07-523LS | Phase I, Ongoing | IV | NIAID | HVTN 130/ HPTN 089 | |||
| PGT121.414.LS + VRC07-523LS | Phase I, Ongoing | IV, SC | NIAID | HVTN 136/ HPTN 092 | |||
| VRC07-523LS + CAP256V2LS | Phase I, Ongoing | SC | NIAID | HVTN 138/ HPTN 098 | |||
| CAP256V2LS + VRC07-523LS + PGT121 | Phase I, Ongoing | IV, SC | CAPRISA, NIAID | CAPRISA 012B | |||
| CombiMab1 (AAV9-vectors encoding bNAbs: N6 + 10E8 + 10-1074) | Preclinical | IM | CSP FMBA | - | |||
| CombiMab2 (AAV9-vectors encoding bNAbs: VRC07-523 + PGDM1400 + 10-1074) | Preclinical | IM | CSP FMBA | - | |||
| Multispecific: Parts of two or more antibodies on a single antibody | |||||||
| Regimen | Status | Route | Research Institution | Trial Name | |||
| SAR441236 (CD4bs specificity of VRC01-LS V1/V2 glycan-directed binding of PGDM1400 gp41 MPER binding of 10E8v4) |
Phase I, Planned | IV | Sanofi, NIAID | HVTN 129/ HPTN 088 | |||
| 10E8.4/iMab | Phase I, Ongoing | IV, SC | ADARC | AAAS1239 | |||
ADARC: Aaron Diamond AIDS Research Center; BIDMC: Beth Israel Deaconess Medical Center; CAPRISA: Centre for the AIDS Programme of Research in South Africa; IAVI: International AIDS Vaccine Initiative; NIAID: National Institute of Allergy and Infectious Diseases; VRC: Vaccine Research Center of NIAID; CSP FMBA: Centre for Strategic Planning and Management of Biomedical Health Risks, Federal Medical Biological Agency; IV: Intravenous; SC: Subcutaneous; IM: Intramuscular.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



