Submitted:
07 October 2023
Posted:
08 October 2023
You are already at the latest version
Abstract
This paper explores the ethical considerations surrounding the research on echinoderms, a group of invertebrates that has recently garnered attention in the scientific community. We emphasize the importance of responsible animal handling and the need for an ethical framework that encompasses echinoderms. The 3R's principle, advocating for the replacement of conscious living vertebrates with non-sentient material in research, is discussed as a guiding tool in current animal research practices. As invertebrates are generally classified as non-sentient animals, the Replacement dimension tends to favor them as prevalent models in experimental research. While we currently lack the means to assess the mental states of invertebrates, there is undeniable evidence of social behavior in many species, suggesting that a lack of interactions with these organisms could potentially adversely affect their well-being. In the last few years, considerable progress has been made in developing an ethical framework that takes invertebrates into account, particularly cephalopods, crustaceans, and echinoderms. In this context, we discuss the development of a broader conceptual framework of 5R’s that includes Responsibility and Respect, which may guide practices ensuring welfare in echinoderms, even in the absence of any particular normative.
Keywords:
echinoderms welfare
; 5 R’s principle
; respect
; responsibility
; Invertebrates
; Responsible echinoderms use
; Echinoderms sentience
1. Introduction
Animal use and interaction have played a crucial role in human endeavors throughout recorded human history. Food, transportation, research (primarily in the field of medical investigation), clothing, and companionship are among the main applications. Notably, this engagement includes invertebrates, indicating their natural involvement in various aspects of human existence. While some aspects of these relationships are unequivocally positive, such as providing sustenance, serving as research models, or providing companionship, others are negative, lacking purpose, or even inflicting harm. This dichotomy may stem from the classification of certain invertebrates as pests or vectors of human diseases. Such adverse interactions have elicited widespread aversion or apprehension towards numerous invertebrate species [1] (Kellert, 1993). As a result, ethical concerns about these creatures must be addressed to ensure their viability as experimental modeling alternatives to vertebrates.
Invertebrates account for more than 90% of the total biodiversity on Earth [1] (Kellert, 1993). This vast biological realm includes 36 phyla of invertebrates, eight of which share the most common associations with humans: Porifera, Cnidaria, Platyhelminthes, Nematoda, Annelida, Arthropoda (the largest phylum in the animal kingdom), Mollusca (the second largest phylum in the animal kingdom), and Echinodermata [1] (Kellert, 1993). Although these phyla are all classified as invertebrates, their diversity is nothing short of astounding. Morphological, nervous system, and behavioral characteristics are unique to each phylum and can even vary within one [2,3,4] (Schmidt-Rhaese & Schwartz, 2016; Pollo & Vitale, 2019; Crespi-Abril & Rubilar, 2018).
Recently, there has been a shift in the ethical consideration of invertebrates, prompting a number of studies to lay the philosophical groundwork for incorporating invertebrates into ethical discourse [5,6,7,8] (Hovarth et al., 2013; Mather, 2016; Carrere & Mather, 2019; Crespi-Abril & Rubilar, 2021). The intricate behaviors of specific invertebrates, particularly octopuses (Cephalopods), have primarily prompted this transformation. Close interaction with octopuses in aquarium settings has aided in the identification of individuality and observable behaviors (personality traits) in these creatures [7,9] (Mather, 2012; Mather & Carrere, 2019). Despite their distinct nervous system configuration from vertebrates, these studies have revealed octopuses’ remarkable intelligence and sentience [10] (Schmidt-Rhaesa et al., 2015). Furthermore, in 2013, cephalopods were included in European Union legislation regarding the protection of animals used for scientific purposes, putting them on par with vertebrates (EU, 2010, Directive 2010/63/EU). This pivotal advancement not only adds to the ethical consideration for invertebrates in general, but it also represents a pivotal advancement in the field [3,8,11] (Pollo & Vitale, 2019; Mikhalevich & Powell, 2020; Crespi-Abril & Rubilar, 2021). It is paramount to recognize that a lack of understanding of invertebrate behaviors does not preclude their capacity for sentience or their ability to respond to negative experiences in a non-anthropocentric manner that could cause suffering [12] (Elwood, 2019).
Concurrent with this growing ethical awareness, significant efforts have been made in experimental research to improve invertebrate welfare [5,7] (Hovarth et al., 2013; Carrere & Mather, 2019). However, relevant information about the implications for invertebrate welfare is dispersed, limited, and sometimes contradictory. As a result, the current study conducts a comprehensive review of the current landscape of animal ethics and echinoderm welfare, with the goal of contributing to the development of a comprehensive framework for invertebrate welfare.
2. Ethical concerns and the 5R principle
In general, ethics is concerned with situations involving various types of conflicts. While there are many different types of conflicts in science, not all of them require ethical consideration. The most widely accepted criterion for determining the relevance of ethical reflection is whether or not it has an impact on humans [13] (Guariglia & Videla, 2011). This viewpoint clearly states that all human beings are morally concerned (an anthropocentric doctrine). However, the same cannot be said for non-anthropocentric living beings. As a result, determining which living beings should be regarded as ethical considerations becomes difficult.
One approach is to use humans as a reference point to determine whether these living beings meet criteria for sentience or consciousness, which include behavioral, evolutionary, and physiological considerations [14] (Edelman & Seth, 2009). Scientific evidence supports arguments about which living beings should be considered ethically [6,15,16,17,18] (Chandroo, Duncan, & Moccia, 2004; Griffin & Speck, 2004; Ellwood, 2011; Lewbart & Mosley, 2012; Mather, 2016). With our improved understanding of animal suffering, the scope of moral consideration has expanded to include all vertebrates, as well as octopuses as the sole group of invertebrates.
Nonetheless, the ethical discourse is increasingly shifting toward considering animal species regardless of scientific knowledge about their capacity for suffering [19] (Birch, 2017). Significant efforts have recently been directed toward developing an ethical framework that takes invertebrates into account. Significant progress has been made in this regard by two distinct groups, namely crustaceans [20] (Birch et al., 2021) and echinoderms [8,21] (Rubilar & Crespi-Abril, 2017; Crespi-Abril & Rubilar, 2021).
The 3R’s principle, proposed by Russell and Burch (1959) [22], is the globally accepted tool that currently guides animal research practices. Replacement is one aspect of this principle that advocates for the use of scientific methods involving non-sentient material to replace those involving conscious living vertebrates [22] (Russell & Burch 1959). Replacement should ideally encourage the use of lower levels of organization, such as cell culture, and even artificial models, such as computational simulations. However, because invertebrates are generally considered non-sentient animals, the Replacement dimension tends to favor them as common models for experimental research.
In this regard, the principle falls short in considering the inclusion of those invertebrates whose capacity for sentience is still debated. The 3R’s principle is built on a strong assumption: that non-human animals lower on the zoological scale lack sentience [23] (Tomasik, 2014). This assumption presents a significant limitation, especially considering the vast number and diversity of invertebrates used in scientific research. Despite the widespread acceptance of the 3R’s as a policy tool aimed at alleviating animal suffering and reducing their use in research, its effectiveness in achieving these goals has been notably criticized [24] (Blattner, 2019). In fact, the number of animals used for research in the European Union (EU) is now similar to the 1980s [25] (Taylor & Rego, 2016). The Three Rs Principle was originally intended to establish ethical guidelines for animal experimentation, albeit with species-specific limitations. However, it did not address the epistemological issues raised by animal manipulation adequately [26] (Johnson & Degeling, 2012). This principle was later expanded to the 5R’s principle (Figure 1) to include two additional concepts emphasizing personal commitment: Respect (establishing a respectful relationship with any living being regardless of its complexity or the knowledge we have of that living being) and Responsibility (personal commitment of researchers to apply ethics concepts conscientiously) [8] (Crespi-Abril & Rubilar, 2021).
3. Ethical approach in Echinoderms
3.1. Phylum Echinodermata
Echinoderms are a phylum of marine invertebrates. They are often key, long-lived species that shape and maintain the status of many marine ecosystems, inhabiting a wide range of ecological niches from the abyssal depths of the oceans to the intertidal zone. Echinoderms, distinguished by their pentamerous radial body arrangement, form a monophyletic group with hemichordates, also known as acorn worms. These organisms exhibit significant diversity and widespread distribution across a variety of marine habitats, playing critical ecological roles in each setting [27] (Brusca, Moore, Schuster, 2016). The fundamental characteristics of this Phylum have exhibited remarkable consistency since the Ordovician epoch (approximately 495-440 million years ago) [28] (Telford et al., 2014), with an approximate enumeration of 7000 species [29] (Smith, 2005).
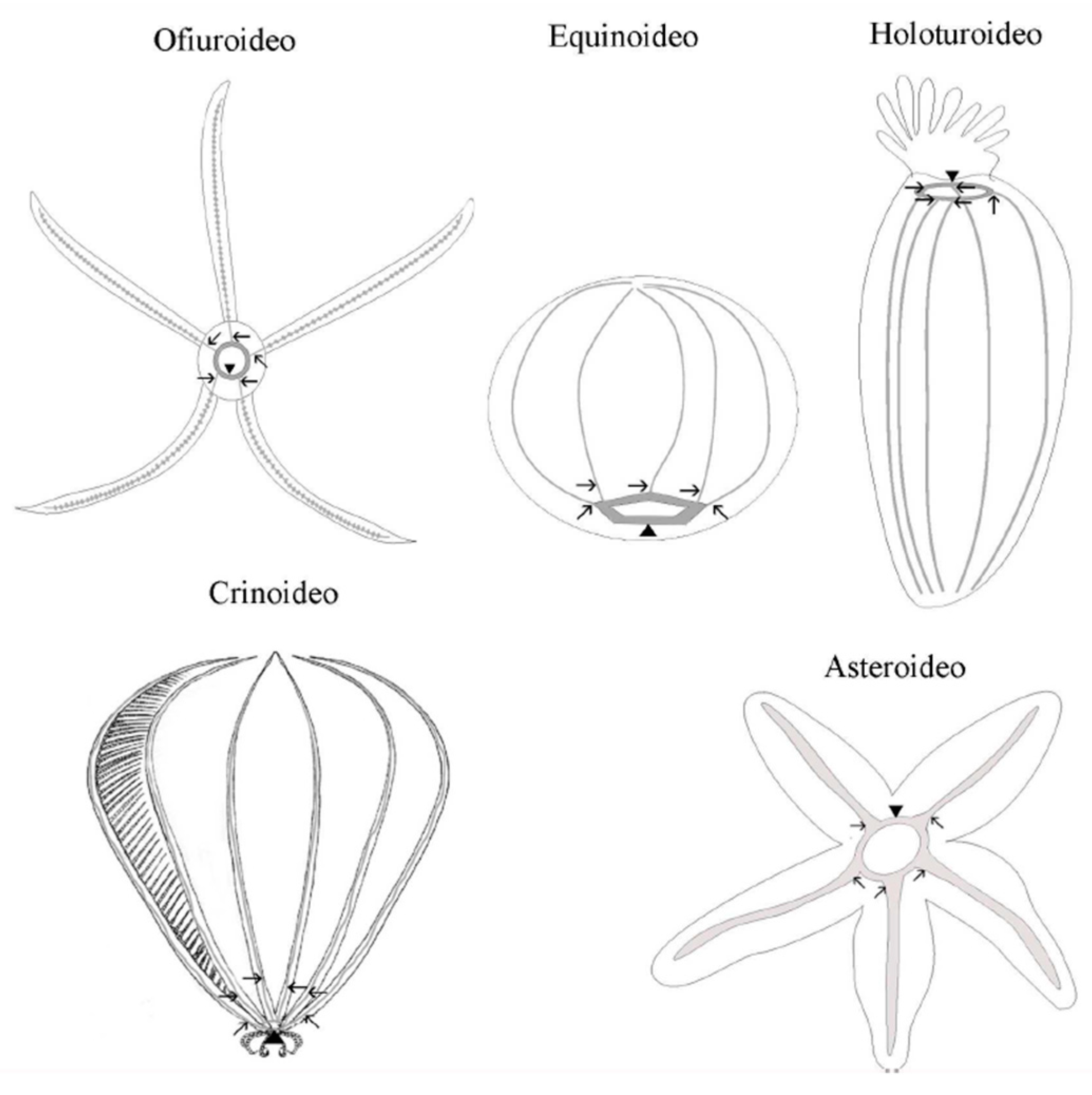
The extant echinoderm assemblage includes approximately 7000 species divided into five distinct taxonomic groups: Asteroidea, which includes starfishes; Echinoidea, which includes sea urchins, sand dollars, and sea biscuits; Crinoidea, which includes sea lilies and feather stars; Ophiuroidea, which includes basket stars and brittle stars and Holothuroidea, which includes sea cucumbers. This collective lineage has an exceptionally notable fossil record dating back to the Cambrian period, providing a solid foundation for comparative molecular studies spanning a wide range of meticulously documented divergence intervals [28,29] (Smith, 2005; Telford et al., 2014) (Figure 2).
Echinoderms have a number of distinguishing characteristics that set them apart from their zoological counterparts. Notably, among these characteristics is a hydraulic water vascular system that is intricately linked with a distinct arrangement of calcium carbonate endoskeleton known as stereom. Most echinoderms developmental trajectories begin with larval stages, where intricate metamorphic processes culminate in the eventual attainment of the adult form. These organisms exhibit a diverse range of life history traits, most notably sexual reproduction, though asexual reproduction is also observed. Echinoderm larvae are primitive, free-living, and planktonic in nature, with a wide range of morphology and functional characteristics that occasionally resemble those observed in hemichordate larvae. Following the metamorphic stage, the majority of mature echinoderms adopt a benthic lifestyle with radial symmetry and a typical pentameric structural composition. Organisms internal architecture is intricately organized, with a reliance on calcium carbonate ossicles reinforced by a complex network of collagenous ligaments. Notably, any skeletal structures found in larvae adopt intricate rod-like configurations with distinct origins in Ophiuroidea and Echinoidea, whereas such features are absent in the larvae of the remaining three echinoderm classes Echinoderms have distinct pentaradial symmetry in their adult forms. Nonetheless, their developmental origins can be traced back to bilaterally symmetrical larvae, a shared feature of the deuterostome clade (shown in Figure 1A). These distinguishing characteristics place echinoderms within an intriguing evolutionary framework, identifying them as invertebrate deuterostomes inextricably linked to vertebrate organisms [30] (Arnone et al., 2015).
Echinoderm species exhibit a variety of developmental strategies, ranging from direct development from a fertilized egg to an adult to indirect development, in which adults emerge from the metamorphosis of a larva with no relation to the adult. The cell structure and count of indirect developing species’ long-lived, feeding, bilaterally symmetrical larvae are very simple. There are numerous intermediate developmental stages, including facultative larval feeding and non-feeding larvae. Indirect development is primitive in echinoderms, and all five surviving classes, as well as the sister phylum Hemichordata, have dipleurula-type larvae [31] (Amemiya et al., 2005).
Crinoids (Crinoidea) are sessile or free-living benthic animals with microphagous filter-feeding habits. Some comatulids feed during the day, while others feed by extending the tips of their arms or moving only at night to avoid predators. Some forms prefer and actively seek out areas with flowing water for feeding (rheophilic), while others do not (rheophobic). They can be found in a variety of habitats, including attachment to substrates via their arms or cirri, within caves or small crevices, beneath rocks, or attached via cirri to other invertebrates such as corals (epizoic). Crinoids are important in developmental biology because they are the only echinoderms that have a primitive tripartite coelom (Mooi and David, 1998). They are significant in paleontology because they evolved during the Cambrian period and were a dominant and diverse component of Paleozoic benthic fauna [32] (Hess and Messing, 2011).
Sea cucumbers (Holothuroidea) are mostly found in benthic marine habitats ranging from the littoral zone to the abyss. They are found in all oceans, with a particular abundance in coral reefs. Sea cucumbers are primarily detritivores, feeding on both suspended particles and organic matter bound in sediment. The distribution of sea cucumbers is influenced by the type of substrate, as well as other environmental variables such as dominant currents, temperature, water salinity, and depth. Sea cucumbers’ ecological importance is linked to their bioturbation activities, which involve the movement of organic material within sediments as well as the transfer of energy and materials at the water-sediment interface. The commercialization of sea cucumbers’ body walls, also known as beche de mer or trepang, is economically significant. Out of the over a thousand known species, the fishing industry primarily focuses on around thirty. This practice has a long history in Chinese and Japanese culinary and medical traditions. Larger species are especially valuable. Trepang curing is applied to the body wall, resulting in a product with high nutritional value due to its high protein and low fat content. Furthermore, sea cucumbers contain biologically active compounds that are used to treat a variety of medical conditions, including HIV, cancer, and osteoarthritis [33,34] (Chen, 2003; Slater and Chen, 2015).
Echinoids (Echinoidea) are found in a wide variety of geographical and bathymetric environments, from intertidal zones to depths of 5000 meters. Regular sea urchins live on a variety of substrates, with the majority of them living on rocky or mobile substrates. Sand dollars and heart urchins, for example, can only be found on soft bottoms and frequently bury themselves. Many morphological differences between regular and irregular echinoids are caused by differences in lifestyle and feeding habits. Some populations of regular sea urchins exposed to wave action have developed digging behaviors to bury themselves slightly, whereas species not exposed to such conditions typically exhibit cryptic behaviors [35] (Lawrence, 2013). The sea urchins, as well as the sea cucumbers are valuable species, the commercialization of sea urchins’ gonads, also known as roe, is economically significant around the world [35,36] (Lawrence, 2013; Sun and Chiang, 2015).
Ophiuroids (Ophiuroidea) have a wide geographic distribution and a benthic lifestyle, having adapted to live in a variety of environments. They have been discovered in submarine hydrothermal vents and on bottoms with cold methane seeps, as well as in intertidal zones and abyssal regions from the tropics to the poles. The majority of species are usually found on soft substrates. They can be carnivorous, scavengers, sediment-consuming, or filter feeders. The majority use multiple feeding methods, though one usually takes precedence over the others. The majority are carnivorous, eating polychaetes, mollusks, and small crustaceans. Because of their diverse feeding strategies, they play an important role in trophic chains. They are not commercially important, unlike some other echinoderm groups. Nonetheless, due to their abundance, they can be used as environmental indicators [37] (Pérez et al., 2014).
Sea stars (Asteroidea) have a wide geographical range and a benthic habitat, inhabiting a variety of marine substrates where they can be abundant and visible. The vast majority are scavengers or opportunistic predators. Many species have generalist feeding habits and play an important role in the structure and functioning of marine communities as apex predators. They play an ecologically important role across latitudes by occupying different levels of the trophic chain, particularly as apex predators in rocky and coral reef ecosystems. This group’s ecological success can be attributed to a variety of morphological and life history characteristics, including indeterminate growth, extraoral and intraoral digestion (providing access to a diverse diet), rapid prey detection and response, and the ability to anchor themselves to substrates using ambulacral tube feet [37,38] (Salguero and Bonilla, 2010; Pérez et al., 2014).
3.2. Historical use of echinoderms
Because sea urchins have been consumed by humans throughout history, human understanding of this Phylum dates back to prehistoric times [39] (Lawrence, 2007). They are depicted in “frescos cretenses,” which date back 4000 years. For a long time, Eastern cultures have consumed and used sea cucumbers for medicinal purposes [40] (Brown & Eddy, 2015). The earliest known of these animals dates from Aristotle, who described the first known echinoderm in 350 B.C.E., over 2000 years ago. He described the feeding apparatus of sea urchins in his work “Historia Animalium,” which is now known as Aristotle’s Lantern. It is worth noting that Aristotle classified echinoderms as ostracoderms. The scholars of the time rekindled their interest in nature and began studying these creatures again. Klein coined the term “Echinodermata” in his 1734 work “Naturalis dispositio echinodermatum.” However, he only used it to refer to sea urchins, not all the classes that are now known. Linnaeus classified the genera Asterias, Echinus, and Holothuria as Mollusca in the 10th Edition of Systema Naturae (1758). The term “Echinodermata” resurfaced in 1792, when it was recognized that these animals were a distinct group of invertebrates, though sea cucumbers were not included. Later, Lamarck (1809) [41] grouped Echinodermata with the true Coelenterata in the Radiata group of invertebrates. It took nearly four decades for De Tornos (1839) [42] and Salacroux (1840) [43] to coin the term “Echinodermata.” Due to its more advanced structural characteristics, it was argued in 1854 that Echinodermata did not belong with Coelelenterata. Echinodermata have since been recognized as a distinct clade of invertebrates.
Since the nineteenth century, the description of Echinodermata species has been a dominant focus in literature. Initially the focus was on species found along Europe’s coasts, as evidenced by Frey and Leuckart’s work in 1847 [44]. As a result of numerous oceanographic expeditions, this trend has expanded to include species from all over the world. While it is impractical to list every expedition and paper that resulted from it, some of the pioneering ones are worth mentioning. The HMS Challenger and the Albatross laid the groundwork for hundreds of subsequent expeditions around the world. These expeditions have recently expanded to include deep-sea species as well as those found in intertidal and shallow waters [45,46,47,48,49] (Agassiz, 1881; Théel, 1882; Agassiz, 1890; Clark, 1902; De Morgan, 1913). Given that echinoderms have been collected worldwide for over 300 years to describe species, determine distributions, and populate museum collections, it is clear that hundreds of thousands of echinoderms were collected and preserved without much ethical thought.
3.3. Echinoderms as models
Since the widespread use of the microscope in scientific research, echinoderms have served as experimental models. Due to the ease with which gametes could be obtained and the optical transparency of sea urchin embryos, they became valuable animal models in the mid-nineteenth century. Dufossé (1847) [50] and Derbès (1847) [51] provided early insights into fertilization and sea urchin embryo development via metamorphosis. Despite the fact that their work was overlooked in the scientific literature [52] (Briggs & Wessel, 2006), sea urchin embryos remained useful as model organisms. Hertwig (1876) [53] made a pivotal discovery by demonstrating how sperm entered the female gamete, leading to embryo formation in the sea urchin Toxopneustes lividus. This discovery established sea urchin embryos as the gold standard for embryonic research, advancing our understanding of fertilization mechanisms, egg activation, cleavage, gastrulation, and early embryonic differentiation. They have also been useful in research on nervous system development, evolutionary development, and regeneration [54,55,56,57,58] (Boveri, 1901; Dolmatov, 1988; Briggs & Wessel, 2006; McClay, 2011; Garner et al., 2016; http://www.echinobase.org).
Sea stars were also recognized as important model organisms in the early twentieth century. Metchnikoff (1893) [59] discovered a cellular immune response in bipinnaria larvae of sea stars, observing the rapid migration of mesenchymal cells to injury sites. He also discovered amoeboid cells’ phagocytic activities [59] (Metchnikoff, 1893). Despite these groundbreaking discoveries, echinoderm larvae were not widely used as model organisms in immunology until the twenty-first century (Figure 2A) [60,61,62] (Pinsino, Thorndyke, & Matranga, 2007; Furukawa, Funabashi, Matsumoto, & Kaneko, 2012; Ho, 2015).
One of the most remarkable characteristics of echinoderms is their ability to regenerate lost structures, such as the arms of sea stars. In recent years, there has been a surge in research into the regenerative capacity of the echinoderm nervous system (NS). Studies have shown that holothurians can regenerate their radial nerve cord (RNC) after it has been transected, with the regenerated cords displaying similar structure and function to the originals and without scar tissue formation—a common issue in vertebrate CNS regenerative responses. Follow-up studies have revealed that radial glia play an important role in the regenerative process, both in forming the bridge that connects the severed ends of the RNC and in generating new neurons and glia in the regenerated structure [63,64] (Mashanov, Zueva, & Garcia-Arraras, 2013; Mashanov and Zueva, 2019). Several laboratories are currently working to identify the genes required for NS regeneration [65,66,67,68,69] (Ben Khadra et al., 2014; Diaz-Balzac & García-Arrarás, 2018; Dolmatov, 2021; Medina et al., 2021; Mashanov et al., 2023).
The capacity for evisceration and subsequent regeneration in sea cucumbers, as well as brittle stars and sea stars due to their ability to regrow arms, has positioned them as invaluable models in regeneration research [70,71,72,73,74,75,76,77,78,79,80,81,82,83,84] (Candia-Carnevali, Bonasoro, Lucca, Thorndyke, 1995; Dolmatov & Ginanova, 2001; Candelaria, Murray, File, García-Arrarás, 2006; San Miguel-Ruiz & García-Arrarás, 2007; Mashanov, Zueva, Heinzeiller, 2008; Carnevali & Burighel, 2010; Mashanov et al., 2014; Mashanov et al., 2015; Mashanov et al., 2017; Ferrario et al., 2018; Dolmatov, 2020; Byrne, 2020; Dolmatov et al., 2020; Medina et al., 2021; Magalhães et al., 2023). With recent technological advancements and the availability of new tools, echinoderms have become outstanding model organisms for both scientific research and educational purposes and in most cases individuals are handled alive [85,86] (Eisen, 1995; Burke et al., 2006; www.Echinobase.org).
3.3. Echinoderms nervous system
In echinoderms, the nervous system is organized in accordance with the general pentameric pattern of the body plan. Each radius has its own radial nerve cord, which runs the length of the proximo-distal axis and terminates at the distal tip of the arm (in stellate echinoderms) or near the aboral pole (in globose forms). A circumoral nerve ring joins all five individual radial nerve cords at the body’s oral pole to form a single anatomical entity. The radial nerve cords and the nerve ring comprise the echinoderm’s central nervous system (CNS) (Figure 3). This CNS is an anatomically and histologically distinct agglomeration of neurons and glial cells associated with an extensive neuropil (densely interwoven neuronal processes) found nowhere else in the body and is in charge of the initiation and coordination of various body-wide responses [87] (Mashanov et al., 2016).
Adult echinoderm neuroanatomy is distinguished by the presence of distinct superimposed domains or layers of nervous tissue located at different levels relative to the oral-aboral axis. These domains are known as the ectoneural and hyponeural systems [88,89,90] (Cobb 1987, Heinzeller and Welsch 2001, Hyman 1955). The ectoneural system is located around the mouth, either within or directly beneath the oral epidermis. It is always present both in the nerve ring and radial nerve cords of all echinoderms, shows the most consistent organization across the phylum, and is the predominant part of the nervous system in all classes except crinoids. The hyponeural system may or may not be a part of the nerve ring or radial nerve cords. Its organization differs between classes and is generally related to the degree of development of large muscles. When hyponeural tissue is present, it acts as a second (usually thinner) layer of nervous tissue that directly overlies the aboral surface of the respective ectoneural cords [90,91] (Heinzeller and Welsch 1994, Hyman, 1955).
This simplified classification of the echinoderm nervous system, however, does not fully account for recent reports from the last decade, revealing unexpected nervous system elements that do not neatly align with the three aforementioned divisions [66,87] (Mashanov et al., 2016; Diaz-Balzac & García-Arrarás, 2018). The nervous system (NS) of echinoderms is one of the most fascinating aspects. Given their close evolutionary relationship with chordates, as well as their radial symmetry and lack of prominent ganglia or centralized nervous structures associated with cephalization found in most other animals, the echinoderm nervous system has been regarded as pivotal to understanding the evolution of the chordate nervous system [87,92] (Mashanov et al., 2016; Arnone et al., 2021). In fact, it has been suggested that the centralized nervous system is a plesiomorphic (ancestral) condition in echinoderms and may also be a plesiomorphic trait at the level of the Deuterostomia [87] (Mashanov et al., 2016).
The nervous system, including the ectoneural and hyponeural subsystems, tissue is organized as a neuroepithelium made up of two major cell types: radial glial cells and neurons. The cell types have a similar relative abundance in the radial nerve cord’s ectoneural and hyponeural bands, with radial glial cells accounting for 60-70% of the total cell population. On the other hand, neuronal cells are more abundant in the circumoral nerve ring, accounting for only 45% of the cell composition formed by glia [93] (Mashanov et al. 2010). Echinoderm glial cells share significant morphological similarities with the radial glia of chordates, including the orthogonal orientation of the cell’s main axis to the plane of the neuroepithelium, the presence of long thick bundles of intermediate filaments, and the presence of short protrusions branching off at a right angle from the main processes and penetrating into the surrounding neural parenchyma. The cell bodies of the majority of radial glial cells in echinoderms are located at the apical surface of the neuroepithelium. Some of them, however, are bipolar, with the apical and basal processes extending from opposite poles of cell bodies located at different depths within the neural parenchyma. Radial glial cells are the most common type of glial cell in echinoderms (though they are unlikely to be perfectly homogeneous), and they perform a variety of functions. In addition to the radial glia of the CNS, other glial cell types associated with the peripheral nervous system may exist [87] (Mashanov et al., 2016). Radial glia has more morphological similarities than chordates because they are the main proliferative population in nervous tissue and thus capable of giving rise to new neurons, both in the non-injured and regenerating CNS. There are, however, differences between radial glia in echinoderms and chordates. Another significant difference is that radial glia is the only major glial cell type in the adult CNS of echinoderms, whereas in higher vertebrates, radial glia predominates in embryogenesis but then mostly disappear from the mature nervous system by giving rise to a plethora of more specialized cell types [94,95] (Kriegstein and Alvarez-Buylla 2009, Malatesta et al. 2008). Radial glia is common in the adult CNS of lower vertebrates, but they frequently co-exist with other abundant specialized glia, such as oligodendrocytes [96,97] (Tanaka and Ferretti 2009, Zamora 1978).
The neural parenchyma of the CNS is made up of neurons with somata and neurites. The most common neuronal morphology ranges from unipolar to bipolar to multipolar. In echinoderms, the neurons can be classified quite easily by size: The normal neurons, which are dominant in most Classes, and the giant neurons of ophiuroids. The first class has a small somata (about 5 m in diameter) that produces very thin give processes (0.1-1 m in cross-section) with numerous local swellings (varicosities) along their length. Because they are typically found near or embedded within calcareous structures, they are less appealing to neurobiologists, particularly electrophysiologists who study their electrical properties. As a result, while other animal groups became popular neurobiology research subjects, echinoderms were largely overlooked for a long time. Even today, there are surprisingly few electrophysiological studies on echinoderms [98,99] (Binyon & Hasler, 1970; Millott & Okumura, 1968).
Communication between neurons in traditional chemical synapses was previously thought to be absent in echinoderms [88,100] (Cobb 1987, 1995). However, this long-held belief may be due to a lack of adequate tools for dealing with the difficulties imposed by the endoskeleton. Since the optimized sample preparation protocols, there has been evidence demonstrating the presence of typical chemical synapses that occur on a regular basis in the CNS of Echinodermata [93,102] (Mashanov et al. 2006, 2010). These findings are consistent with those found in the sea urchin genome, where the genes required for synapse formation were discovered [101] (Burke et al. 2006). There are also different types of synapses as: unsheathing synapses (with the pre-synaptic terminal wrapped around the post-synaptic process), passant synapses between parallel nerve fibers, and complex synapses with a pre-synaptic terminal forming two or more synapses on different post-synaptic processes, or, conversely, a single post-synaptic neuron receiving synaptic input from multiple pre-synaptic axons [102] Mashanov et al., 2006). Echinoderm CNS have regional differences in cellular composition as well as a complex internal spatial segregation of different cell types. The radial nerve cord is made up of repetitive units [88,103,104] (de Bremaeker et al. 1997, Cobb 1987, Cobb and Stubbs 1981).
The CNS of echinoderms generates complex, coordinated, and directional behavioral responses to various sensory stimuli. Although the molecular and cellular mechanisms underlying these behaviors remain unknown, it is known that an echinoderm’s CNS contains a large number of neurotransmitters from all major groups, including acetylcholine, aminoacids, monoamines, neuropeptides, and gases. Acetylcholine appears to mediate muscle contraction due to its function as a major excitatory neurotransmitter [105,106,107] (Devlin 2001, Florey and Cahill 1980, Heinzeller and Welsch 1994). Post-synaptic nicotinic and muscarinic receptors have also been identified, and acetylcholinesterase (the enzyme required to hydrolyze acetylcholine at synapses) activity has been detected in both the ectoneural and hyponeural systems [88,108] (Cobb 1987, Ryberg 1974). GABAergic neurons proliferate throughout the CNS, including the radial nerve cord, nerve ring, and podial nerves, as well as the nerves and visceral plexi [87,109] (Newman and Thorndyke, 1994; Mashanov et al., 2016). GABA is involved in both echinoderm muscle contraction and relaxation, depending on the post-synaptic receptor (GABA A or GABA B) present in neuromuscular junctions [105] (Devlin, 2001). L-glutamate is and excitatory neurotransmitter in the ectoneural subsystem of echinoderms [110,111] (Wilkie et al. 2010, 2013). L-glutamate is also a neurotransmitter capable of eliciting arm autotomy response, whereas acetylcholine acts as an antagonist of L-glutamate [110] (Wilkie et al. 2010). The research on serotonine as a neurotransmitter in the adult CNS is limited. There have been reports of its presence in muscles and basiepithelial plexi [112,113,114] (Candia Carnevali et al. 2001, Inoue et al. 2002, Sugni et al. 2004). Furthermore, neuropharmacological studies have revealed that serotonin regulates muscular contraction by inhibiting the excitatory effect of acetylcholine [113] (Inoue et al. 2002) and may also be involved in the regulation of post-traumatic regeneration [114] (Sugni et al. 2004). Catecholamines such as dopamine and noradrenaline have been found in the ectoneural only and appear to be involved in the movement of the tube feet [115,116] (Cottrell and Pentreath 1970, Díaz-Balzac et al. 2010) and to be fundamental for the righting response of the sea urchin [117] (Howell et al., 2023). Histamine data is extremely limited; it has only been studied in one species of cucumber, and it appears to be involved in sensory systems, as it was found in tentacles and body wall papillae and projected their axons directly to the nerve ring and the radial nerve cord [118] (Hoekstra et al. 2012).
The enzyme nitric oxide synthase, which produces NO, was discovered in both the ectoneural and hyponeural parts of the radial nerve cord of adult sea stars, as well as in some radial glial cells [119,120] (Kotsiuba and Kotsiuba 2004, Martinez et al. 1994). Apparently NO is involved in the relaxation of the viscera and tube feet [121,122] (Elphick and Melarange 1998; 2001).
In addition to these phylogenically widespread neurotransmitters, echinocherm contains specific neuropeptides from the SALMFamide family [103,107,122,123,124,128] (Beer et al. 2001, de Bremaeker et al. 1997, Elphick and Melarange 2001, Elphick et al. 2013, Heinzeller and Welsch 1994). These neuropeptides relax the visceral musculature as well as the muscles of the body wall [122,125] (Elphick and Melarange 2001, Elphick et al. 1995).
All of the information above suggests that the Echinoderm CNS is more complex than previously thought and that, despite the lack of a centralized brain, it is possible to elicit complex individual and social behavior. Recent advances in knowledge of this group have provided new insights suggesting that echinoderms are sentient animals capable of suffering pain.
3.3. Pain and echinoderms
Even though the debate over pain perception in invertebrates is still ongoing, it is critical to recognize that the absence of evidence of painful sensations should not be interpreted as conclusive proof of pain absence in this group. As scientists, we are responsible for treating the organisms we study with dignity and ensuring their well-being. To accomplish this, we must first understand the concepts of analgesia, sedation, and anesthesia. It is also critical to understand how to correctly administer anesthesia, which response variables to consider, and which substances to use.
Various agents have been used to anesthetize echinoderms [126] (Kaplan, 1969). Iso-osmotic solutions such as MgCl2, MgSO4, or Ca2+-free seawater are common echinoderm anesthetics. The mechanism of action of these agents is to destabilize membrane potential, preventing pain signals from propagating. Additionally, local anesthetics that block neural stimulation of the muscle have been employed when necessary. The local anesthetics “MS222” [127,128,129,130,131] (Wilkie, 1978; O’Neill, 1989, 1990, 1994; Motokawa and Wainwright, 1991) and “propylene phenoxetol” [127,132,133,134,135,136,137] (Wilkie, 1978, 1983, 1992; Byrne, 1985, 1986; Wilkie, et al., 1984; Wilkie, et al., 1990) have both demonstrated effectiveness in studies involving echinoderm connective tissue.
MS222 (IUPAC name 3-amino benzoic acid, ethyl ester, methanesulfonate salt, also known as ethyl m-aminobenzoate or tricaine methanesulfonate) is a local anesthetic of the ester type. Its structure is similar to that of other local anesthetics, such as benzocaine, implying that it likely functions similarly by impeding axonal conduction via interference with membrane depolarization [138] (MacDonald and Wann, 1978). Originally developed as a fish anesthetic [139] (Jolly et al., 1972), it has since been widely used on a variety of invertebrates (National Research Council, 1981).
In ophiuroids, “propylene phenoxetol” is thought to act as a local anesthetic that inhibits axonal conduction [137] (Wilkie et al., 1990). Because this volatile liquid is not water soluble, maintaining known concentrations of the compound in the medium is difficult. Furthermore, there have been some doubts about the precise identity of this compound.
While certain compounds possessing anesthetic effects in echinoderms have been studied, the practice of using them during individual manipulations is not yet standardized because there is neither awareness nor normative mandate for it.
4. Echinoderms welfare
Animal welfare is now recognized as a scientific discipline that includes ethology, physiology, pathology, biochemistry, genetics, immunology, nutrition, cognitive-neural studies, veterinary care, and ethics [140,141,142,143,144,145,146] (Fraser, 2008a, Fraser, 2008b; Fraser et al., 1997; Lassen et al., 2006; Mason & Mendl, 1993; Mellor et al., 2009; Sande & Simonsen, 1992). While the assessment of animal welfare has traditionally focused on vertebrates [147] (Hemsworth et al., 2015), the vast diversity of invertebrates presents a significant challenge.
Firstly, the fundamental indicators (such as cortisol levels, longevity, feeding rate, and behavior) utilized in welfare assessments follow a reductionist approach. Secondly, this assessment only encompasses two of the “Three Conceptions” (Basic health and function, and Natural living) and four of the “Five Domains” (Nutrition, Environment, Health, and Behavior) that should ideally be evaluated. By definition, this basic assessment remains incomplete. Moreover, in invertebrates, except for cephalopods, evaluating the Mental State Domain and the Conception of Affective State is currently extremely challenging. Although we lack the tools to assess invertebrate mental states, there is undeniable evidence of social behavior in many species, implying that a lack of interactions with these organisms may have a negative impact on their mental well-being [148] (Bovenkerk & Verweij, 2016). Even if a comprehensive welfare assessment for invertebrates is not currently feasible, we must ensure that the best welfare conditions are met.
A comprehensive assessment of animal welfare, according to Botreau et al. (2007) [149], requires a well-defined set of criteria. These guidelines should be followed when developing these criteria:
- Each and every significant aspect must be addressed in order for the assessment to be comprehensive;
- The criteria must not be redundant or irrelevant;
- Each criterion must be independent of the others.
- All stakeholders must agree on the criteria, and they must have a practical basis
- The criteria, as well as their application, should be transparent and simple to understand.
- The number of criteria should be limited to 12 at most.
Given the diversity of invertebrates, it is critical to recognize the importance of developing a specific set of criteria for assessing the welfare of each Phylum or even each Order within this group.
There are several indicators that provide information about an animal’s well-being in response to its experiences. These indicators are based on a thorough understanding of how individuals respond physiologically and behaviorally to various conditions. The interpretation of these indicators is dependent on one’s attitude toward animal welfare. Exploration, hunting or foraging, socialization, parental care, play, and sexual activity are all expected to develop as inherent behavioral elements [150,151] (Mellor, 2015a,b). As a result, when an animal is kept in captivity, it is critical to understand the specific fundamental characteristics of each species that exist in their natural environment in order to recreate them. This meets the need to “explore, solve problems, and overcome challenges” [152] (p.623) (Lahvis 2017, p.623).
In order to achieve the application of echinoderm well-being, the following non-invasive echinoderm assessments have recently been developed as indicators for use in the aquaculture industry and research, providing valuable insights into individual well-being:
- Tube feet adhesion: Echinoderms move by using their tube feet, which are frequently adhesive to the bottom or walls of the aquarium in captivity. A detached or loosely attached echinoderm indicates that the individual is not in good health.
- Echinoderms have a distinct righting behavior because they are most vulnerable with their oral face above [153,154] (Percy, 1973; Himmelman, 1984). The speed at which this behavior is carried out can be used to gauge its physiological activity, health, and overall condition. This indicator must first be established for the species, and size and sex must be evaluated before establishing a normal time of righting for each species. After you have set a time, you can use it as a stress indicator.
- Echinoderms use their spines / Pedicelaria / Arms for a variety of functions (feeding, moving, defense, eating), so their response to stimuli is a good indicator of their health. An echinoderm that is nonresponsive or responds slowly indicates that the individual is not in good health.
- Feeding behavior: like any other animal’s, is an indicator of health. Echinoderms feeding and defecating indicate a healthy individual.
- Epidermis appearance: A healthy echinoderm has a shiny, non-interrumped epidermis. In contrast, the presence of reddish or blackish coloration, as well as inflammation and mucus, indicates the presence of an infection.
5. Conclusions
The majority of ethical and welfare approaches in animal research have primarily focused on vertebrates. Addressing invertebrate welfare presents unique challenges, and researchers will require time to fully integrate these concepts. However, significant progress in this area has been made [7,8,20] (Carrere and Mather, 2019, Birch et al., 2021; Crespi-Abril and Rubilar, 2021). With rising public awareness and concern, there is a chance that ethical concepts will be adopted more quickly. In the near future, this could lead to the establishment of guidelines, norms, and laws in this domain.
In recent years, it has become clear that advanced invertebrates have qualities such as self-awareness and sentience, as well as the ability to experience pain, though precisely defining and understanding it in their context is difficult. Animal welfare legislation in various countries supports this viewpoint. While many invertebrates have learning and memory abilities, there are significant structural and physiological differences between animal groups. Some researchers believe that these differences indicate that, despite their self-protective behaviors, advanced invertebrates are incapable of feeling pain. Regardless of the validity of this argument, it is critical for humanity, particularly scientists, to take invertebrate welfare seriously and to treat them with care, both in captivity and in their natural habitats. The scientific community has responded by advocating for invertebrate welfare to be considered in breeding or holding facilities, laboratories, and under natural conditions whenever possible. In addition, if advanced invertebrate experimentation is deemed necessary, appropriate anesthesia methods should be considered.
Author Contributions
Both Authors, Crespi-Abril and Rubilar, contributed to conceptualization, writing and editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONICET, grant number PIP 11220210100013CO”.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kellert, S. R. Values and perceptions of invertebrates. Conservation Biology 1993, 7, 845–855. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, & schwartz 2016. Nematomorpha from the Philippines, with description of two new species. Zootaxa, 4158(2), 246-260.
- Pollo, S., & Vitale, A. (2019). Invertebrates and humans: science, ethics, and policy. In C. Carere & J. Mather (Eds.), The Welfare of Invertebrate Animals (pp. 7–22). Springer.
- Crespi-Abril, A. C., & Rubilar, T. Ética e invertebrados: análisis de los casos de los cefalópodos y equinodermos. Revista Latinoamericana de Estudios Críticos Animales 2018, 8, 210–232.
- Horvath, K., Angeletti, D., Nascetti, G., Carere, C. Invertebrate welfare: an overlooked issue. Annali dell’Istituto superiore di sanità 2013, 49, 9–17.
- Mather, J. A. An invertebrate perspective on pain. Animal Sentience 2016, 1, 12. [Google Scholar] [CrossRef]
- Carere, C., & Mather, J. (2019). The Welfare of Invertebrate Animals. Springer. [CrossRef]
- Crespi-Abril, A. C., & Rubilar, T. Moving forward in the ethical consideration of invertebrates in experimentation: Beyond the Three R’s Principle. Revista de Biología Tropical 2021, 69(Suppl. 1), 346–357. [CrossRef]
- Mather, J.A. Why (and how) personalities in invertebrates? Current Zoology 2012, 58, 566. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, A., Harzsch, S., & Purschke, G. (2015). Structure and Evolution of Invertebrate Nervous Systems. Oxford University Press.
- Mikhalevich, I., & Powell, R. Minds without spines: evolutionarily inclusive animal ethics. Animal Sentience 2020, 29, 1–25.
- Ellwood, R.W. Pain and suffering in invertebrates? Institute for Laboratory Animal Research 2011, 52, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Guariglia, O., & Vidiella, G. (2011). Breviario de ética. Buenos Aires: Edhasa.
- Edelman, D. B., & Seth, A. K. Animal Consciousness: A Synthetic Approach. Trends in Neurosciences 2009, 32, 476–484.
- Chandroo, K. P., Duncan, I. J. H., & Moccia, R. D. Can fish suffer?: Perspectives on Sentience, Pain, Fear, and Stress. Applied Animal Behavior Science 2004, 86, 225–250.
- Griffin, D. R., & Speck, G. B. New Evidence of Animal Consciousness. Animal Cognition 2004, 7, 5–18.
- Ellwood, R. W. Pain and suffering in invertebrates? Institute for Laboratory Animal Research 2011, 52, 175–184. [Google Scholar] [CrossRef]
- Lewbart, G. A. , & Mosley, C. Clinical Anesthesia and Analgesia in Invertebrates. Journal of Exotic Pet Medicine 2012, 21, 59–70. [Google Scholar]
- Birch, J (2017) Animal Sentience and the Precautionary Principle. Animal Sentience, 16(1). [CrossRef]
- Birch, J., Burn, C., Schnell, A., Browning, H., y Crump, A. (2021). Review of the Evidence of Sentience in Cephalopod Molluscs and Decapod Crustaceans. London: London School of Economics and Politics.
- Rubilar, C. T., & Crespi-Abril, A. C. Does Echinoderm research deserve an ethical consideration? International Journal of Tropical Biology 2017, 65, S11–S22.
- Russell, W. M. S., & Burch, R. L. (1959). The Principles of Humane Experimental Technique. London: Methuen.
- Tomasik, B. (2014). Suffering in Animals vs. humans, essays on reducing suffering. Retrieved from http://reducing-suffering.org/suffering-in-animals-vs-humans/.
- Blattner, C. E. (2019). Rethinking the 3Rs: From Whitewashing to Rights. In K. Hermann y K. Jayne (Eds.), Animal Experimentation: Working Towards a Paradigm Change (pp. 168-193). Amsterdam: Brill.
- Taylor, K., & Rego, L. EU statistics on animal experiments for 2014. ALTEX 2016, 33, 465–468.
- Johnson, J., & Degeling, C. Animals-as-Patients: Improving the Practice of Animal Experimentation. Between the Species 2012, 15, 43–58.
- Brusca, R. C., Moore, W., & Schuster, M. (2016). Invertebrates. Massachusetts: Sinauer Associated, Inc, Publishers.
- Telford, M. J., Lowe, C. J., Cameron, C. B., Ortega-Martinez, O., Aronowicz, J., Oliveri, P., & Copley, R. R. Phylogenomic analysis of echinoderm class relationships supports Asterozoa. Proceedings of the Royal Society of London B: Biological Sciences 2014, 281, 20140479.
- Smith, A. B. Echinoderms (Other Than Echinoids). In R. Cocks & I. Plimer (Eds.), Encyclopedia of Geology (pp. 334–341). Elsevier, Oxford.
- Arnone, M.I., Byrne, M., Martinez, P. (2015). Echinodermata. In: Wanninger, A. (eds) Evolutionary Developmental Biology of Invertebrates 6. Springer, Vienna. [CrossRef]
- Amemiya, C. T., Miyake, T., & Rast, J. P. Echinoderms. Current biology 2005, 15, R944–R946.
- Hess, H., Messing, C., & Ausich, W. I. (2011). Treatise on Invertebrate Paleontology. University of Kansas Paleontological Institute.
- Chen, J. Overview of sea cucumber farming and sea ranching practices in China. SPC beche-de-mer Information Bulletin 2003, 18, 18–23. [Google Scholar]
- Slater, M., & Chen, J. Sea cucumber biology and ecology. Echinoderm Aquaculture 2015, 47–55.
- Lawrence, J. M. (Ed.). (2013). Sea Urchins: Biology and Ecology. Academic Press.
- Sun, J., & Chiang, F. S. Use and exploitation of sea urchins. Echinoderm aquaculture 2015, 25–45.
- López-Pérez, Andrés, Rebeca Granja-Fernández, Cuauhtémoc Aparicio-Cid, Ronald C. Zepeta-Vilchis, Ana M. Torres-Huerta, Francisco Benítez-Villalobos, Daniel A. López-López, Carlos Cruz-Antonio, and Omar Valencia-Méndez. “Corales pétreos, equinodermos y peces asociados a comunidades y arrecifes coralinos del Parque Nacional Huatulco, Pacífico sur mexicano.” Revista mexicana de biodiversidad 85, no. 4 (2014): 1145-1159.
- Luna Salguero, B and Bonilla, H. “Estructura comunitaria y trófica de las estrellas de mar (Echinodermata: Asteroidea) en arrecifes rocosos de Loreto, Golfo de California, México.” Hidrobiológica 20, no. 2 (2010): 127-134.
- Lawrence, J. M. (2007). Sea urchin roe cuisine. In Developments in Aquaculture and Fisheries Science (Vol. 37, pp. 521–523). Elsevier.
- Brown, N., & Eddy, S. (2015). Echinoderm Aquaculture. New Jersey: Wiley-Blackwell.
- Lamarck, J. B. (1809). Philosophie zoologique, ou, Exposition des considérations relative à l’histoire naturelle des animaux. Pzaris.
- De Tornos, L. (1839). Compendio de Historia Natural Dividido en los Tres Ramos de Mineralogía, Botánica y Zoología. Madrid, Imprenta Salvador Albert.
- Salacroux, A. P. G. (1840). Nuevos Elementos de Historia Natural. Madrid, Imprenta de Verges.
- Frey, H., & Luckart, R. (1847). Zootomie. Voss, Germany: Leipzig. Agassiz, A. (1881). Report on the Echinoidea dredged by H.M.S. Challenger during the Years 1873-1876. ZOOLOGY Part IX Challenger Reports.
- Agassiz, A. Report on the scientific results of the voyage of the H.M.S. Challenger during the years 1873–76, report on the Echinoidea. Challenger Rep 1881, 3, 1–321. [Google Scholar]
- Théel, H. (1882). Report on the Holothurioidea, dredged by H.M.S. Challenger during the Years 1873-1876. ZOOLOGY Part IX Challenger Reports.
- Agassiz, A. Notice of Calamocrinus diomedeae, a new stalked Crinoid from the Galapagos, dredged by the U. S. Fish Commission Steamer ‘Albatross’. Bulletin of the Museum of Comparative Zoology 1890, 20, 165–167. [Google Scholar]
- Clark, H. L. (1902). Papers from the Hopkins Stanford Galapagos Expedition. Proceeding of the Washington Academy of Science, IV, 521-531.
- De Morgan, W. The Echinoderms collected by the “Huxley” from the North Side of the Bay of Biscay in August, 1906. Journal of the Marine Biological Association of the United Kingdom 1913, 9, 530–541. [Google Scholar] [CrossRef]
- Dufossé, A. (1847). Observations sur le developpement des oursins. In Annales des Sciences Naturelles (Vol. 7, pp. 44–52).
- Derbès, A. A. Observations sur le Méchanisme et les Phenomènes qui Accompagnent la Formation de l’Embryon chez l’Oursin Comestible. Ann Sci Nat Zool 1847, 8, 80–98. [Google Scholar]
- Briggs, E., & Wessel, G. M. In the beginning… animal fertilization and sea urchin development. Developmental biology 2006, 300, 15–26.
- Hertwig, R. (1876). Zur Histologie der Radiolarien: Untersuchungen über den Bau und die Entwicklung der Sphaerozoiden und Thalassicolliden. W. Englemann.
- Boveri, T. Die Polarität von Oocyte, Ei und Larve des Strongylocentrotus lividus. Zoologische Jahrbücher. Abteilung für Anatomie und Ontogenie der Tiere 1901, 14, 630–653. [Google Scholar]
- Dolmatov, I. Y. (1988). Structure of the aquapharyngeal complex in the holothurian Eupentacta fraudatrix under the normal conditions and during regeneration. Canadian Science Dissertation. Vladivostok: Institute of Marine Biology, Far East Division of the USSR Academy of Sciences.
- Briggs, E., & Wessel, G. M. In the beginning… animal fertilization and sea urchin development. Developmental biology 2006, 300, 15–26.
- McClay, D. R. Evolutionary crossroads in developmental biology: sea urchins. Development 2011, 138, 2639–2648. [Google Scholar] [CrossRef]
- Garner, S., Zysk, I., Byrne, G., Kramer, M., Moller, M. Taylor, V., & Burke, R. D. Neurogenesis in sea urchin embryos and the diversity of deuterostome neurogenic mechanisms. Development 2016, 143, 286–297.
- Metchnikoff, I. (1893). Lectures on the Comparative Pathology of Inflammation, Delivered at the Pasteur Institute in 1891. London.
- Pinsino, A., Thorndyke, M. C., & Matranga, V. Coelomocytes and post-traumatic response in the common sea Star Asterias rubens. Cell Stress & Chaperones 2007, 12, 331–341.
- Furukawa, R. , Funabashi, H., Matsumoto, M., & Kaneko, H. Starfish ApDOCK protein essentially functions in larval defense system operated by mesenchyme cells. Immunology and Cell Biology 2012, 90, 955–965. [Google Scholar]
- Ho, E. C. H. (2016). A simple animal model for characterizing gene regulatory control of an immune response. University of Toronto (Canada).
- Mashanov, V. S., Zueva, O. R., & Garcia-Arraras, J. E. Radial glial cells play a key role in echinoderm neural regeneration. BMC Biology 2013, 11, 49.
- Mashanov, V., & Zueva, O. Radial glia in echinoderms. Developmental neurobiology 2019, 79, 396–405.
- Ben Khadra, Y., Said, K., Thorndyke, M., & Martinez, P. Homeobox genes expressed during echinoderm arm regeneration. Biochemical genetics 2014, 52, 166–180.
- Díaz-Balzac, C. A., & García-Arrarás, J. E. (2018). Echinoderm nervous system. In Oxford Research Encyclopedia of Neuroscience.
- Dolmatov, I. Y. Molecular aspects of regeneration mechanisms in holothurians. Genes 2021, 12, 250. [Google Scholar] [CrossRef]
- Medina-Feliciano, J. G., & García-Arrarás, J. E. Regeneration in echinoderms: Molecular advancements. Frontiers in Cell and Developmental Biology 2021, 9, 768641.
- Mashanov, V. S., Ademiluyi, S., Machado, D. J., Reid, R., & Janies, D. (2023) Echinoderm Radial Glia in Adult Cell Renewal, Indeterminate Growth, and Regeneration. Frontiers in Neural Circuits, 17, 1258370. [CrossRef]
- Candia-Carnevali, M. D., Bonasoro, F., Lucca, E., & Thorndyke, M. C. Pattern of cell proliferation in the early stages of arm regeneration in the feather star Antedon mediterranea. The Journal of Experimental Zoology 1995, 272, 464–474.
- Dolmatov, I. Y., & Ginanova, T. T. Muscle Regeneration in Holothurians. Microscopy Research and Technique 2001, 55, 452–463.
- Candelaria, A. G., Murray, M., File, S. K., & García-Arrarás, J. E. Contribution of mesenterial muscle dedifferentiation to intestine regeneration in the sea cucumber Holothuria glaberrima. Cell and Tissue Research 2006, 325, 55–65.
- San Miguel-Ruiz, J. E., & García-Arrarás, J. E. Common cellular events occur during wound healing and organ regeneration in the sea cucumber Holothuria glaberrima. BioMed Central Developmental Biology 2007, 7, 115–124.
- Mashanov, V. S., Zueva, O. R., & Heinzeller, T. Regeneration of the radial nerve cord in a holothrian: A promising new model system for studying post-traumatic recovery in the adult nervous system. Tissue & Cell 2008, 40, 351–372.
- Carnevali, M. D. C., & Burighel, P. (2010). Regeneration in echinoderms and ascidians. eLS.
- Mashanov, V. S., Zueva, O. R., & García-Arrarás, J. E. Transcriptomic changes during regeneration of the central nervous system in an echinoderm. BMC genomics 2014, 15, 1–21.
- Mashanov, V. S., Zueva, O. R., & García-Arrarás, J. E. Expression of pluripotency factors in echinoderm regeneration. Cell and tissue research 2015, 359, 521–536.
- Mashanov, V. S., Zueva, O. R., & García-Arrarás, J. E. Inhibition of cell proliferation does not slow down echinoderm neural regeneration. Frontiers in zoology 2017, 14, 1–9.
- Ferrario, C., Khadra, Y. B., Czarkwiani, A., Zakrzewski, A., Martinez, P., Colombo, G.,... & Sugni, M. Fundamental aspects of arm repair phase in two echinoderm models. Developmental biology 2018, 433, 297–309.
- Dolmatov, I. Y. Variability of regeneration mechanisms in echinoderms. Russian Journal of Marine Biology 2020, 46, 391–404. [Google Scholar] [CrossRef]
- Byrne, M. The Link between Autotomy and CNS Regeneration: Echinoderms as Non-Model Species for Regenerative Biology. BioEssays 2020, 42, 1900219. [Google Scholar] [CrossRef]
- Dolmatov, I. Y., Kalacheva, N. V., Mekhova, E. S., & Frolova, L. T. Autotomy and regeneration of the visceral mass in feather stars. Zoomorphology 2020, 139, 171–187.
- Medina-Feliciano, J. G., & García-Arrarás, J. E. Regeneration in echinoderms: Molecular advancements. Frontiers in Cell and Developmental Biology 2021, 9, 768641.
- Magalhães, F., Andrade, C., Simões, B., Brigham, F., Valente, R., Martinez, P., ... & Coelho, A. V. (2023). Regeneration of Starfish Radial Nerve Cord restores animal mobility and unveils a new coelomocyte population. Cell and Tissue Research, 1-16.
- Eisen, A. (1995). A Holistic Approach to Teaching a Laboratory Using Sea Urchin. Development as an Example System. In C. A. Goldman (Ed.), Tested Studies for Laboratory Teaching (pp. 25-32). Atlanta, GA: Proceedings of the 16th Conference of the Association for Biology Laboratory Education.
- Burke, R. D., et al. A genomic view of the sea urchin nervous system. Developmental biology 2006, 300, 434–460. [CrossRef]
- Mashanov, V. Mashanov, V., Zueva, O., Rubilar, T., Epherra, L., & García-Arrarás, J. E. (2016). Echinodermata. In A. Schmidt-Rhaesa, S. Harzsch, & G. Purschke (Eds.), Structure and Evolution of Invertebrate Nervous Systems (pp. 665–688). Oxford: Oxford University Press.
- Cobb, J. L. S. (1987). Neurobiology of the Echinodermata. In Nervous Systems in Invertebrates (pp. 483-525). Boston, MA: Springer US.
- Heinzeller, T. and Welsch, U. (2001). The echinoderm nervous system and its phylogenetic interpretation. In G. Roth and M. Wullimann, eds. Brain Evolution and Cognition, pp. 41–75. Wiley-Spektrum, New York.
- Hyman, L. (1955). The Invertebrates. IV. Echinodermata. The Coelomate Bilateria. McGraw-Hill Book Co., New York.
- Heinzeller, T. and Welsch, U. (1994). Crinoidea. In F. Harrison and F.-S. Chia, eds. Microscopic Anatomy of Invertebrates. Vol. 14. Echinodermata. pp. 1–148. Wiley-Liss, New York.
- Arnone, M.I. , Byrne, M., Martinez, P. (2015). Echinodermata. In: Wanninger, A. (eds) Evolutionary Developmental Biology of Invertebrates 6. Springer, Vienna. [CrossRef]
- Mashanov, V. S., Zueva, O. R., & Garcia-Arraras, J. E. Organization of glial cells in the adult sea cucumber central nervous system. Glia 2010, 58, 1581–1593.
- Kriegstein, A. and Alvarez-Buylla, A. The glial nature of embryonic and adult neural stem cells. Annual Reviews in Neurosciences 2009, 32, 149–184. [Google Scholar] [CrossRef]
- Malatesta, P., Appolloni, I., and Calzolari, F. Radial glia and neural stem cells. Cell & Tissue Research 2008, 331, 165–178.
- Tanaka, E.M. and Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nature Reviews in Neuroscience 2009, 10, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Zamora, A.J. The ependymal and glial configuration in the spinal cord of urodeles. Anatomy and Embryology (Berlin) 1978, 154, 67–82. [Google Scholar] [CrossRef]
- Binyon, J., & Hasler, B. Electrophysiology of the starfish radial nerve cord. Comparative Biochemistry and Physiology 1970, 32, 747–753.
- Millott, N. , & Okumura, H. The electrical activity of the radial nerve in Diadema antillarum Philippi and certain other echinoids. Journal of Experimental Biology 1968, 48, 279–287. [Google Scholar]
- Cobb, J. (1995). The nervous systems of Echinodermata: recent results and new approaches. In O. Breidbach and W. Kutsch, eds. The Nervous Systems of Invertebrates: An Evolutionary and Comparative Approach, pp. 407–424. Birkhhäuser Verlag, Basel.
- Burke, R.D., Angerer, L.M., Elphick, M.R., et al. A genomic view of the sea urchin nervous system. Developmental Biology 2006, 300, 434–460. [CrossRef]
- Mashanov, V. , Zueva, O., Heinzeller, T., and Dolmatov, I. Ultrastructure of the circumoral nerve ring and the radial nerve cords in holothurians (Echinodermata). Zoomorphology 2006, 125, 27–38. [Google Scholar] [CrossRef]
- De Bremaeker, N., Deheyn, D., Thorndyke, M.C., Baguet, F., and Mallefet, J. Localization of S1- and S2-like immunoreactivity in the nervous system of the brittle star Amphipholis squamata (Delle Chiaje 1828). Proceedings of the Royal Society of London B Biological Sciences 1997, 264, 667–674. [CrossRef] [PubMed]
- Cobb, J.L. and Stubbs, T.R. The giant neurone system in Ophiuroids. I. The general morphology of the radial nerve cords and circumoral nerve ring. Cell & Tissue Research 1981, 219, 197–207. [Google Scholar]
- Devlin, C.L. The pharmacology of gamma-aminobutyric acid and acetylcholine receptors at the echinoderm neuromuscular junction. Journal of Experimental Biology 2001, 204, 887–896. [Google Scholar] [CrossRef]
- Florey, E. and Cahill, M.A. Cholinergic motor control of sea urchin tube feet: evidence for chemical transmission without synapses. Journal of Experimental Biology 1980, 88, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Heinzeller, t. and Welsch, U. (1994). crinoidea. in f. Harrison and f.-S. chia, eds. Microscopic Anatomy of Invertebrates. vol. 14. Echinodermata. pp. 1–148. Wiley-liss, new york.
- Ryberg, E. The localization of cholinesterases and non-specific esterases in the Echinopluteus. Zoologica Scripta 1974, 2, 163–170. [Google Scholar] [CrossRef]
- Newman, S.J. and Thorndyke, M.C. (1994). Localisation of gamma aminobutyric acid (GABA)-like immunoreactivity in the echinoderm Asterias rubens. Cell & Tissue Research.
- Wilkie, I.C., Barbaglio, A., Maclaren, W.M., and Carnevali, M.D.C. Physiological and immunocytochemical evidence that glutamatergic neurotransmission is involved in the activation of arm autotomy in the featherstar Antedon mediterranea (Echinodermata: Crinoidea). Journal of Experimental Biology 2010, 213, 2104–2115.
- Wilkie, I.C., Barbaglio, A., and Carnevali, M.D.C. The elusive role of L-glutamate as an echinoderm neurotransmitter: evidence for its involvement in the control of crinoid arm muscles. Zoology 2013, 116, 1–8. [CrossRef]
- Candia Carnevali, M., Bonasoro, F., Thorndyke, M.C., and Patruno, M. (2001). Role of the nervous system in echinoderm regeneration. In M. Barker, ed. Echinoderms 2000, pp. 5–20. Balkema, Rotterdam.
- Inoue, M., Tamori, M., and Motokawa, T. Innervation of holothurian body wall muscle: inhibitory effects and localization of 5-HT. Zoological Science 2002, 19, 1217–1222. [CrossRef]
- Sugni, M., Ferreri, F., Bonasoro, F., Candia Carnevali, M., and Wilkie, I.C. (2004). New evidence for serotonergic control of regenerative processes in crinoids. In T. Heinzeller and J. Nebelsick, eds. Echinoderms: München, pp. 141–146. Taylor & Francis Group, London.
- Cottrell, G.A. and Pentreath, V.W. Localization of catecholamines in the nervous system of a starfish, Asterias rubens, and of a brittlestar, Ophiothrix fragilis. Comparative and General Pharmacology 1970, 1, 73–81. [Google Scholar] [CrossRef]
- Díaz-Balzac, C.A., Mejías, W., Jiménez, L.B., and García-Arrarás, J.E. The catecholaminergic nerve plexus of Holothuroidea. Zoomorphology 2010, 129, 99–109.
- Howell, E., Lancaster, A., Besh, J., Richardson, B., Gomez, E., Harnew-Spradley, S., & Shelley, C. The dopamine receptor antagonist haloperidol disrupts behavioral responses of sea urchins and sea stars. Journal of Experimental Biology 2023, 226.
- Hoekstra, L.A., Moroz, L.L., and Heyland, A. Novel insights into the echinoderm nervous system from histaminergic and FMRFaminergic-like cells in the sea cucumber Leptosynapta clarki. PloS One 2012, 7, e44220.
- Kotsiuba, E.P. and Kotsiuba, A.E. NADPH-diaphorase localization in the radial nerve cords of the starfish Patiria pectinifera. Tsitologiia 2004, 46, 346–351. [Google Scholar] [PubMed]
- Martinez, A., Riveros-Moreno, V., Polak, J.M., Moncada, S., and Sesma, P. Nitric oxide (NO) synthase immunoreactivity in the starfish Marthasterias glacialis. Cell & Tissue Research 1994, 275, 599–603.
- Elphick, M.R., and Melarange, R. Nitric oxide function in an echinoderm. Biological Bulletin 1998, 194, 260–266. [CrossRef] [PubMed]
- Elphick, M.R. and Melarange, R. Neural control of muscle relaxation in echinoderms. Journal of Experimental Biology 2001, 204, 875–885. [Google Scholar] [CrossRef]
- Beer, A. J., Moss, C., & Thorndyke, M. Development of serotonin-like and SALMFamide-like immunoreactivity in the nervous system of the sea urchin Psammechinus miliaris. Biological Bulletin 2001, 200, 268–280.
- Elphick, M. R., Achhala, S., & Martynyuk, N. The evolution and diversity of SALMFamide neuropeptides. PLoS One 2013, 8, e59076.
- Elphick, M. R., Kemenes, G., Staras, K., & O’Shea, M. Behavioral role for nitric oxide in chemosensory activation of feeding in a mollusc. Journal of Neuroscience 1995, 15, 7653–7664.
- Kaplan , H.M. 1969 . Anesthesia in invertebrates . Fed. Proc. , 28 : 1557 – 1569.
- Wilkie, I. C. Nervously mediated change in the mechanical properties of a brittlestar ligament. Marine Behaviour and Physiology 1978, 5, 289–306. [Google Scholar] [CrossRef]
- O’Neill, P. L. Structure and mechanics of starfish body wall. J. Exp. Biol. 1989, 147, 53–89. [Google Scholar] [CrossRef]
- O’Neill, P. L. Torsion in the asteroid ray. J. Morph. 1990, 203, 141–149. [Google Scholar] [CrossRef]
- O’Neill, P. L. The effect of anaesthesia on spontaneous contraction of the body wall musculature in the asteroid Coscinasterias calamaria. Marine Behaviour and Physiology 1994, 24, 137–150. [Google Scholar] [CrossRef]
- Motokawa, T., & Wainwright, S. A. Stiffness of starfish arm and involvement of catch connective tissue in the stiffness change. Comp. Biochem. Physiol. 1991, 100A, 393–397.
- Wilkie, I. C. Nervously mediated change in the mechanical properties of the cirral ligaments of a crinoid. Marine Behaviour and Physiology 1983, 9, 229–248. [Google Scholar] [CrossRef]
- Wilkie, I. C. Variable tensility of the oral arm plate ligaments of the brittlestar Ophiura ophiura (Echinodermata: Ophiuroidea). Journal of Zoology 1992, 228, 5–26. [Google Scholar] [CrossRef]
- Byrne, M. The mechanical properties of the autotomy tissues of the holothurian Eupentacta quinquesemita and the effects of certain physico-chemical agents. J. exp. Biol. 1985, 117, 69–86. [Google Scholar] [CrossRef]
- Byrne, M. Induction of evisceration in the holothurian Eupentacta quinquesemita and the evidence for the existence of an endogenous evisceration factor. J. exp. Biol. 1986, 120, 25–40. [Google Scholar] [CrossRef]
- Wilkie, I. C., Emson, R. H., & Mladenov, P. V. Morphological and mechanical aspects of fission in Ophiocomella ophiactoides (Echinodermata, Ophiuroidea). Zoomorphology 1984, 104, 310–332.
- Wilkie, I. C., Griffiths, G. V. R., & Glennie, S. F. (1990). Morphological and physiological aspects of the autotomy plane in the aboral integument of Asterias rubens L. (Echinodermata). In: Echinoderm Research. C. De Ridder, P. Dubois, M-C. Lahaye and M. Jangoux (Eds.), A.A. Balkema, Rotterdam, pp. 301–313.
- MacDonald, A. G., & Wann, K. T. (1978). Physiological Aspects of Anaesthetics and Inert Gases. Academic Press.
- Jolly, D. W., Mawdsley-Thomas, L. E., & Bucke, D. Anaesthesia of fish. Vet. Record 1972, 91, 424–426.
- Fraser, D. (2008a). Understanding animal welfare. Acta Veterinaria Scandinavica, 50(1), 1–7. [CrossRef]
- Fraser, D. (2008b). Understanding Animal Welfare: The Science in its Cultural Context. Wiley-Blackwell Publishing.
- Fraser, D., Weary, D. M., Pajor, E. A., & Milligan, B. N. A scientific conception of animal welfare that reflects ethical concerns. Animal Welfare 1997, 6, 187–205.
- Lassen, J., Sandøe, P., & Forkman, B. Happy pigs are dirty!–conflicting perspectives on animal welfare. Livestock Science 2006, 103, 221–230. [CrossRef]
- Mason, G. J., & Mendl, M. (1993). Why is there no simple way of measuring animal welfare? Animal Welfare, 2, 301–319.
- Mellor, D., Patterson-Kane, E., & Stafford, K. J. (2009). The Sciences of Animal Welfare. John Wiley & Sons.
- Sandøe, P., & Simonsen, H. B. (1992). Assessing animal welfare: where does science end and philosophy begin? Animal Welfare, 1(4), 257–267.
- Hemsworth, P. H., Mellor, D. J., Cronin, G. M., & Tilbrook, A. J. Scientific assessment of animal welfare. New Zealand Veterinary Journal 2015, 63, 24–30. [CrossRef]
- Bovenkerk, B., & Verweij, M. (2016). Between individualistic animal ethics and holistic environmental ethics blurring the boundaries. In B. Bovenkerk & J. Keulartz (Eds.), Animal Ethics in the Age of Humans (pp. 369–385). Springer. [CrossRef]
- Botreau, R., Veissier, I., Butterworth, A., Bracke, M. B., & Keeling, L. J. Definition of criteria for overall assessment of animal welfare. Animal Welfare 2007, 16, 225–228.
- Mellor, D. (2015a). Enhancing Animal Welfare by Creating Opportunities for Positive Affective Engagement. New Zealand Veterinary Journal, 63, 3–8.
- Mellor, D. (2015b). Positive Animal Welfare States and Encouraging Environment-Focused and Animal-to-Animal Interactive Behaviours. New Zealand Veterinary Journal, 63, 9–16.
- Lahvis, G. Animal Welfare: Make Animal Models More Meaningful. Nature 2017, 543, 623. [Google Scholar] [CrossRef] [PubMed]
- Percy, J. A. (1973). Thermal Adaptation in the Boreo-Arctic Echinoid Strongylocentrotus droebachiensis (O. F. Müller, 1776). II. Seasonal Acclimatization and Urchin Activity. Physiology and Zoology, 46, 129–138.
- Himmelman, J. H., Guderley, H., Vignault, G., Drouin, G., & Wells, P. G. Response of the sea urchin, Strongylocentrotus droebachiensis, to reduced salinities: importance of size, acclimation, and interpopulation differences. Canadian journal of zoology 1984, 62, 1015–1021.
Figure 1.
Five R’s Principle which balances the level of importance of the ethical and epistemological approaches [8].
Figure 1.
Five R’s Principle which balances the level of importance of the ethical and epistemological approaches [8].
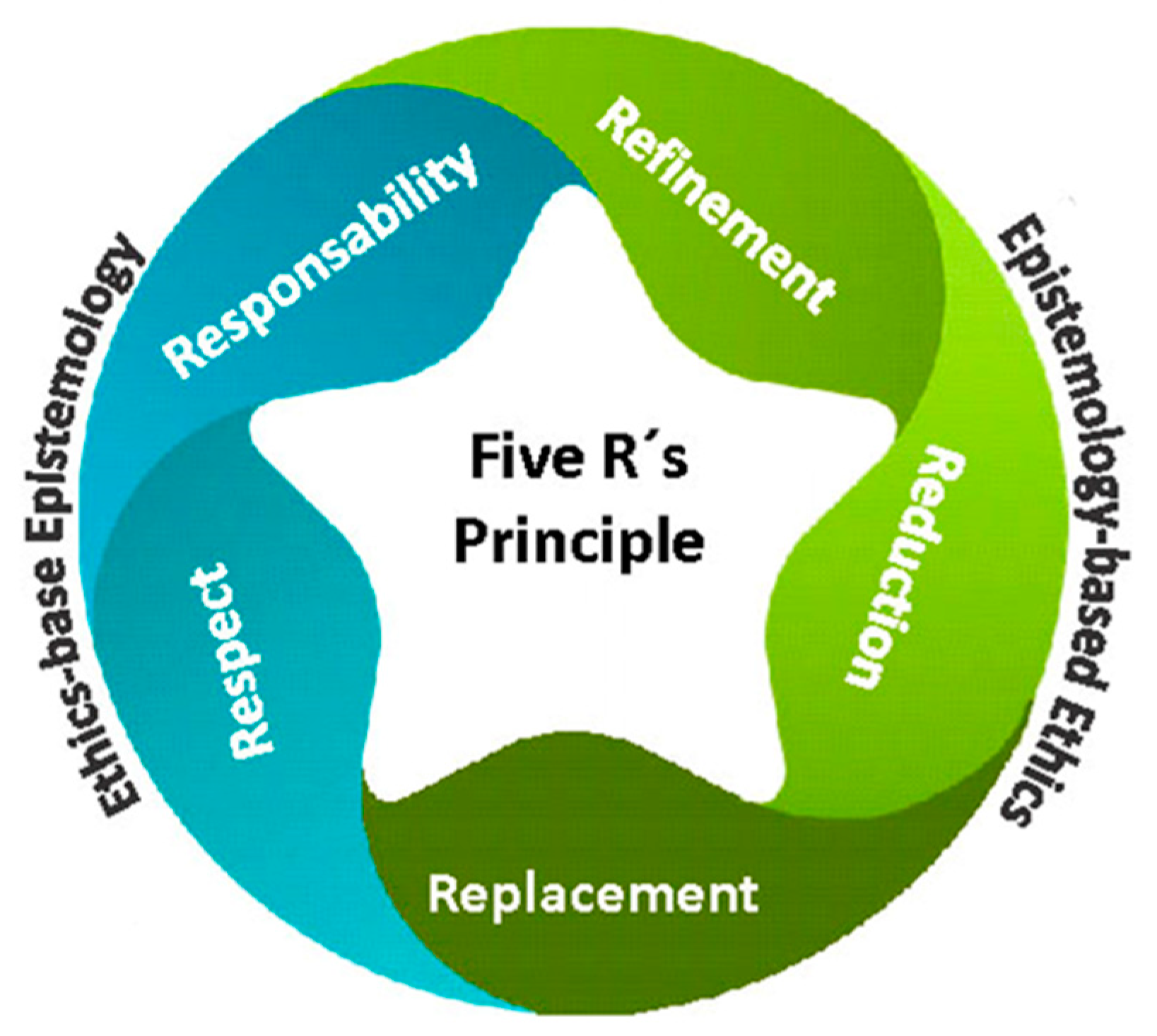
Figure 2.
Classes of Echinoderm A. Asteroidea B. Echinoidea C. Crinoidea D. Ophirouidea E. Holothuroidea.
Figure 2.
Classes of Echinoderm A. Asteroidea B. Echinoidea C. Crinoidea D. Ophirouidea E. Holothuroidea.

Figure 3.
Schematic representation of the central nervous system of the phylum Echinodermata.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



