Submitted:
24 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Mycobacterium tuberculosis is the agent of tuberculosis, one of the most important infectious diseases in the world. This microorganism stands out from other bacteria, not only for its extremely high infection capacity, but also for its cellular characteristics that include an extremely resistant and hydrophobic cellular parade the passage of antibiotics. An incredible ability to adapt to adverse conditions inside the host as well as its vast arsenal of virulence factors that allow its survival within the inhospitable environment within the macrophage can be highlighted. This review aims to discuss several aspects of MTB microbiology, genetics, and physiology. We will address in this review details of the metabolism of MTB that allows it to replicate in the active phase and remain viable during latency, as well as the characteristics of its cell wall that contribute to the blockade of the immune response and its resistance to antibiotics.
Keywords:
tuberculosis
; Mycobacterium
; physiology
; mycolic acids
; cell wall
1. Introduction
The World Health Organization (WHO) estimates that one-third of the world's population is infected with Mycobacterium tuberculosis (MTB), the agent of tuberculosis (TB), and that about 5 to 10% of those infected will develop active disease at some stage of your life. MTB was first described in 1882 by Robert Koch, thereafter, being commonly referred to as Koch's bacilli in honor of its discoverer. Since its discovery, the bacillus has been the target of several studies and campaigns aimed at its eradication [1,2,3].
MTB is transmitted through the inhalation of aerosols emitted by patients in the active phase of the disease. Upon inhaling MTB is phagocytosed by alveolar macrophages, yet other cells such as dendritic cells and neutrophils also can phagocyte MTB, triggering an immune response that recruits more immune cells to the site. Finally, with the development of the progressive immune response, a local lesion called granuloma appears. This lesion is a hallmark of TB and although it is not pathognomonic helped in the diagnosis of active disease [4,5,6,7,8].
MTB infects only human hosts, and the infecting dose is extremely low, only 5-10 bacilli can establish the initial infection. In the interior of the macrophage, the MTB is inside of modified phagosomes, to establish the infection and to avoid the immune system the MTB is used of diverse strategies like the inhibition of phagosome maturation, inhibition of the presentation of antigens, among other [4,9].
It is important to note that several people will contact the MTB and will be able to control the spread of the disease due to the action of the immune system. However, these individuals have great potential to have residual bacilli and, therefore, to become ill phagosome throughout their lives. In infected people, the risk of developing active disease increases over time, but in patients with immune deficiencies such as HIV, this risk is even greater, Figure 1 [10,11,12,13].
2. Mycobacteria and Mycobacterium tuberculosis: An Interesting Genus
Mycobacteria are bacilli, phylogenetically related to actinobacteria, with large amounts of C-G in their DNA. They present an extremely complex cellular envelope, which is characterized by the rich presence of mycolic acids. The Mycobacterium genus comprises more than 150 species including Mycobacterium tuberculosis Complex (MTBC), M. avium Complex (MAC), M. terrae Complex, M. abcessus-chelonae Complex, M. simiae Complex, M. fortuitum-smegmatis Group, M. celatum Group, M. xenopi Group and others non-tuberculous mycobacteria (NTM). Mycobacteria are identified by Ziehl-Neelsen staining. They are acid-fast bacteria, which during staining with fuchsin retain this dye even after washing with alcohol-acid due to the characteristics of its cell wall. This cell wall is composed of a layer of peptidoglycan, and a micomembrane composed of mycolic acids, arabinose, and other sugars, which make this membrane highly hydrophobic and provide great protection to the bacillus against external agents such as antibiotics [15,16,17,18,19,20,21].
Potentially pathogenic can cause lung disease or even disseminated disease in immunocompromised patients. The MTBC is formed by the species M. tuberculosis, M. canettii, M. africanum, M. microti and M. bovis, recently was reported more two new species M. mungi and M. orygis. In addition to two subspecies M. caprae and M. pinnipedii. The bacteria belonging to the MTBC present about 99.95% of genetic identity and although there is clearly a great similarity in genome level, the members present different preferences regarding the host, pathogenicity, and phenotype [11,22,23].
M. bovis can cause disease in humans whose disease cannot be distinguished from that caused by MTB. Although its incidence is low nowadays thanks to the advent of pasteurization, which has virtually eliminated the main route of transmission, transmission through the ingestion of contaminated milk produced by contaminated cattle, M. bovis disease has been increasing in immunocompromised patients, especially those infected with HIV. M. africanum is an important agent of TB, accounting for half of the cases in West Africa [24,25,26].
The M. Avium Complex is formed by M. avium subsp. avium (MAA), M. avium subsp. hominissuis (MAH), M. avium subsp. paratuberculosis (MAP), M. avium subsp. silvaticum (MAS), M. avium subsp. lepraemurinum, M. arosiense, M. bouchedurhonense, M. colombiense and M. intracellulare, which may cause lung disease and cavitary lung disease in immunocompromised patients, but in immunocompetent patients, the disease is rare [27,28,29,30,31,32].
Other important NTM is the M. ulcerans, agent responsible for Buruli ulcer. This mycobacterium presents the production of an important cytotoxin, mycolactone, responsible for cellular necrosis and immunomodulation. In addition, the other important pathogenic slow growers are M. marinum, M. haemophilum, M. genavese, M. kansasii, M. malmoense, M. simiae, M. xenopi and M. szulgai [20,33,34,35,36,37].
Among the species, we must also give relevance to Mycobacterium leprae, which is the agent of leprosy. Although the disease is one of the oldest, still today the culture of this bacterium cannot be performed in laboratory media. NTMs are in the environment, mainly soil and water and are classified according to the growth rate in culture. Currently many NTMs have been causing illness in patients who have undergone invasive procedures, such as ophthalmic, aesthetic, and cardiac surgeries, or constant use of injectable drugs. The most frequent NMTs in these cases are the fast-growing M. fortuitum, M. abscessus and M. chelonae. However, the isolation of NTMs requires great care since it may be the result of simple contamination or temporary colonization. Table 1 shows the main groups and complexes of the Mycobacteriaceae family and their representatives [33,38,39].
The history of TB is closely linked to the history of humankind, with signs of TB being found in European and Middle Eastern skeletons of 4000 years until the description of tuberculosis by Hippocrates in ancient Greece. In Europe in the 16th and 17th centuries, TB was one of the main infectious diseases of the time, with thousands of associated deaths41. Now is widely accepted that MTB originated in Africa more than 7000 years ago from environmental mycobacteria. With the aid of the establishment of agriculture, livestock, establishment of the civilization and increase of the population density it was possible to select virulent strains and the populations transmitted that[40,41,42,43,44].
It is interesting to note that MTB in the Americas has become a major problem for indigenous communities during colonization by the European peoples. MTB was believed to be a microorganism evolved from M. bovis that causes the same human pathology in cattle; yet new studies have shown that the only difference between the members of the M. tuberculosis Complex corresponds to the polymorphisms in certain genes, suggesting, therefore, that both bacilli evolved in parallel [45].
3. Mycobacteria and Mycobacterium tuberculosis: An Interesting Genus
MTB is a straight or slightly curved bacillus, immobile and non-sporulating. It has a lipid-rich cell wall, is resistant to alcohol-acid discoloration, and is called alcohol-acid resistant bacilli. MTB exhibits an extremely slow generation time of 20 to 22 hours, nonchromogenic colonies, rugose-like suede appearance, growing between 14 to 28 days at 37 °C in solid medium. In addition, samples from non-sterile sites may affect the bacillus growth. The cell wall of the MTB has as main constituents the mycolic acid, peptidoglycan and arabinogalactans, being covered by polysaccharides and proteins that are not covalently bound. This wall is responsible for an important degree of microorganism resistance to the microbicidal mechanisms present inside the macrophage [46,47,48,49].
Recent studies of cryomicroscopy allied to tomography suggest that the mycomembrane of MTB, is organized like the external wall of Gram negative and that presents proteins with function of porins. It shows that the cell wall of this important pathogen would be much more complex than it has been described today [50,51].
It is estimated that the MTB genome shelters about 140 proteins in the mycomembrane, none of which have been identified and characterized. This fact is even more striking considering that in M. smegmatis; the MspA protein was structurally identified and characterized. MspA consists of the major pore of M. smegmatis, and its deletion showed decrease in permeability to glucose, phosphate, metal ions and amino acids. This interesting porin consists of eight monomers, showed the presence of a channel formed by β-sheets, besides a large external protuberance also formed by β-sheets. The core presents 16 asthmatic acid residues that generate a highly negative network, which explains the pore preference for cations. However, no similar protein was identified in MTB [52,53,54,55,56,57,58].
The MTB peptidoglycan consists of repeats of the disaccharides N-acetylglucosamine and N-acetylmuramic acid (NAG-NAM), which are produced in the cytoplasm from lipid precursors. The precursors receive the addition of amino acids in their chains, through the Mur proteins, which constitute in ATP-dependent ligases. The MTB peptidoglycan is highly interlaced, with cross-links 3-3 instead of the traditional 4-3, in addition, they have different alterations such as NAM glycosylation, D-Glu amidation and mesodiaminopimelic acid (mDAP) in lateral chains. It is interesting to note that imidation may inhibit the recognition of MTB by the immune system; however, the presence of glycosylation induces the production of inflammatory cytokines by the macrophage. In addition, there is the action of hydrolases that will act on the anterior peptidoglycan, opening space for the insertion of the new peptide, whereas penicillin-binding proteins (PBPs), such as PonA1 and PonA2, incorporate the units in the structure. Finally, PBPA, PBPB, LdtA and LdtB are transpeptidases that promote crosslinks between the nascent structures [59,60,61,62].
MTB shows the MurA, MurB, MurC, MurD, MurE and MurF proteins and the product of these proteins are exported with the action of the transmembrane protein MviN. The proteins MurI, DapF and DdlA are responsible to produce the substrates for the Mur proteins Figure 2 [62,63,64,65,66].
The arabinogalactan present in the cell wall of the MTB is composed of a galactan formed by the repetition of the disaccharide of 6-d-Galfβ1-5-d-Galfβ, being formed by the action of galactofuranosil transferases GlfT1 and GlfT2. Like NAM, galactan is modified by the addition of long arabinan polymers which are produced by DprE1, DprE2, AftA, AftB, EmbA and EmbB and their ends are branched by AftC, AftD and Rv1459. Arabinans may also receive modifications as addition of succinyl or non-N-acetylated galactosamines (GalN), which is found in many pathogenic mycobacteria and appears to be important for an efficient infection. Finally, the arabinans are bound to mycolic acids, which are large hydroxylated fatty acids of branched chains, being covalently attached to the cell wall, forming the mycomembrane, Figure 3 [67,68,69,70].
The FASI (FASI enzyme) and FAS2 enzyme complexes (composed of MabA, HadABC, InhA, KasAB and an isomerase) that promote the union and branching of two fatty acids form myasolic acids. Subsequently undergo maturation promoted by a series of enzymes which results in three different molecules at the end: α-meroacids, methoxy-meroacids and ketomeroacids, all three being important for virulence. Other lipids that are components of the cell wall of mycobacteria are trehalose dimycolate (TDM) that promotes the cord factor (the cord factor consists of serpentine MTB clusters observed in the lamina stained by Ziehl Neelsen), glycerol monomicolate, glucose monomicolate (GMM), triacyl-trehalose (TAT), diacyl trehalose (DAT), sulfolipid (SLs), pentacyl trehalose (PAT), mannosyl-β-1-phosphomicocetos, phenolic glycolipids (PGLs) and phthiocerol dimycocesate [71,72,73,74].
Most genes involved in lipid synthesis are essential for MTB survival. Mycobacterial Membrane Proteins Large (MmpLs) transports the mycolic acids produced. Mmpl are members of the superfamily Resistance Nodulation Division (RND). MTB has fifteen genes coding for RND-type transporters, 13 of which are from Mmpl proteins. In addition to the transport of lipids, these transporters have also been involved in the transportation of drugs, the acquisition of iron and the export of siderophores [75,76,77,78,79,80,81,82].
The formation of the MTB cell wall is an extremely complex process and requires several post-translational transformations, which include phosphorylation, pupilation which is a process of addition of ubiquitin-like proteins, and which direct the proteins for proteolysis through proteosomes. Complex regulation of cell wall formation allows MTB to regulate its growth under different conditions within the host, even being a likely mechanism of persistent infection. Therefore, the MTB cell wall has several molecules that induce the immune response [83,84,85,86,87,88].
MTB presents for cell division the FtsZ protein, a GTPase that forms the central ring of the division and recruits the other proteins involved in the process; FtsE, a protein like ABC type transporters, but without known function. In additional, FtsK, a translocase DNA responsible for chromosomal segregation and septal formation; FtsW, responsible to produce peptide glycol in the septum and translocation of the peptide. A finally FtsI, acts together with FtsW promoting peptidoglycan transpeptidation in the septum; AmiC, an amidase responsible for septal degradation for cell separation and EnvC, also involved in cell separation through septal degradation. In relation to the division, the MTB divides through the elongation and subsequent division of the mother cell into two daughter cells, this division occurring asymmetrically and generating daughter cells of different sizes [89,90,91,92,93,94,95,96].
4. A Complex Metabolism for Survive and Infect
MTB is a prototrophic bacterium that can synthesize amino acids, vitamins, and cofactors. It presents a complete set of genes involved in the metabolism of fatty acids, tricarboxylic acid pathway and pentose phosphate pathway. In addition, MTB have enzymes that bind the cycle of tricarboxylic acids and the glycolytic pathway. Studies show that MTB can grow rapidly in the first three weeks of infection, reaching its plateau after 2 to 3 weeks after establishment of a Th1-type immune response and increased interferon-γ levels. However, auxotrophic mutants for proline, leucine, lysine, tryptophan, purines, or magnesium transport deficiency related to the mgt gene, present growth deficits in the early stages. Some genes that code for enzymes involved in lipid metabolism, such as isocitrate lyase, phospholipase C and pcaA, when disrupted and mutated, cause defects in MTB persistence in the late stages of growth, with most of them showing no sign of active disease [97,98,99,100,101].
Cholesterol is an important source of energy for MTB, in addition to being used for the synthesis of lipids linked to virulence. During latent infection, mainly, cholesterol provides many of the products needed for survival for long periods. The metabolism of cholesterol requires the activity of many oxygenates and is probably an explanation of why MTB would infect the lungs, where large concentrations of oxygen are found. The mce 4 operon has been shown to be responsible for encoding a cholesterol transport system and host membrane lipid uptake, in addition to participating in energy pathways from these lipids, which allows the infection and permanence of the bacilli by long periods of time [102,103,104,105,106,107,108,109,110,111,112].
The MTB to survive inside the host needs carbon and nitrogen sources and for this, it explores several metabolic pathways for these nutrients to be obtained. Carbon sources inside the macrophage are reduced when compared to other environments of the organism, inside the macrophage the MTB increases the expression of isocitrate lyase, which acts on the transformation of acetyl-CoA into carbohydrates through gluconeogenesis. In the macrophage, glucose availability is also one of the critical points for the establishment of MTB, for this we note an increase in the production of enzymes involved with β-oxidation and gluconeogenesis that allows the use of abundant fatty acids in the cellular interior. Phosphate is important for processes of obtaining energy, producing membrane phospholipids, nucleic acids, and many other cellular reactions [98,112,113,114,115,116].
MTB can use ammonia as a source of nitrogen, which is incorporated into glutamate, and glutamine, this process is catalyzed by glutamine synthetase and oxaglutarate synthetase. The bacillus can still grow with NO3- as the only source of carbon, or even use nitrates that are transported by importers during hypoxia to serve as final electron acceptor. Nitrites generated in the cell interior can be exported out of the cell through exporters. It is interesting to note that in MTB there are several ways to capture and use organic sources of nitrogen, mainly amino acids, the most important being aspartate, asparagine, glutamate, and glutamine that will undergo deamination processes by specific enzymes. Finally, urea can be used as a source of nitrogen, albeit with less efficiency than other organic and inorganic sources. Sulfur in turn is important for the initial process of protein synthesis and for maintaining a redox environment. Therefore, these metabolic systems in MTB present several proteins related to the detection of critical concentrations, capture and transformation of organic and inorganic sources in the molecules of interest [114,117,118,119,120,121,122,123,124].
Among other nutritional requirements, MTB needs the presence of vitamins and other co-factors for survival. For example, many MTB enzymes involved in the synthesis of fatty acids and other lipids are biotin-dependent, so MTB has several genes related to the de novo production of biotin. Another essential cofactor for MTB is nicotinamide adenine dinucleotide (NAD), which is important for various redox reactions, reactions linked to energetic metabolism processes, cobalamin synthesis, and DNA ligase activity. In this way, the MTB can carry out the synthesis of new or recycle from external sources [125,126].
The uptake of metals is an important aspect for the survival of any microorganism, in MTB in addition to the transporters there are the production of siderophores, which are carboxymycobactins produced and exported to the extracellular medium. In the extracellular medium, these molecules bind to non-heme iron and other metals, are internalized by specific transporters that allow the use of these metals, while the less polar mycobactins are associated with the cell and sequester iron. Carboxymycobactins can sequester the iron contained in transferrin and ferritin from the host and IrtAB ABC transporter is responsible for the translocation of these siderophores through the inner membrane. Other essential metals for MTB are zinc, manganese, and cobalt, which are used as cofactors to produce various molecules such as vitamin B12 or even act in response to oxidative stress. MTB presents sensors capable of identifying the levels of these metals in the cellular interior, in addition to the presence of transporters that capture these important metals. It is interesting to note that the host organism often responds by decreasing the concentration of these metals through the action of chelating proteins such as calprotetin [127,128].
Finally, in terms of metabolism, one important point is the oxidative phosphorylation, which allows MTB to produce huge amounts of ATP required for several essential cellular functions. The complex responsible to produce ATP is the F1Fo ATPase synthase, a big complex formed by 10 subunits, and produces ATP utilizing a proton motive force. This complex is closely linked to electron transport chain, responsible for generating a gradient of the protons in the periplasm and recycle donors and acceptors of electrons. Due to its importance for the physiology of MTB, ATPase synthase is an important target for drug development. The new option for the treatment of TB consisted of the inhibitor of ATPase synthase bedaquiline [50,129,130,131,132].

Table 2.
Nutritional Source Table and Metabolism of MTB.
| Nutrient/ Metabolism |
Gene | Compound | Keeg Code |
|---|---|---|---|
| Ammonium/ Nitrogen |
Rv1737c, Rrv2329c, Rv0261c, Rv0267, Rv1161, Rv1736c, Rv1162, Rv1164, Rv0252, Rv0253, Rv0021c, Rv2781c, Rv2476c, Rv2220, Rv2222c, Rv2860c, Rv1878, Rv3859c, Rv3858c, Rv3588c, Rv1284 and Rv3273 |
C00011, C00014, C00025, C00058, C00064, C00088, C00169, C00192, C00244, C00288, C00488, C00533, C00697, C00726, C00887, C01417, C01563, C05361 and C06058 | Mtu00910 |
| Glucose Carbohydrate/ Carbon and energy |
Rv0650, Rv0946c, Rv2029c, Rv3010c, Rv1099c, Rv0363c, Rv1438, Rv1436, Rv1437, Rv0489, Rv1023, Rv1617, Rv1127c, Rv2241, Rv2215, Rv0462, Rv2455c, Rv2454c, Rv0761c, Rv1862, Rv0162c, Rv1530, Rv3045, Rv0768, Rv0147, Rv0223c, Rv0458, Rv3667, Rv3068c, Rv2702 and Rv0211 |
C00022, C00024, C00031, C00033, C00036, C00068, C00074, C00084, C00103, C00111, C00118, C00186, C00197, C00221, C00236, C00267, C00469, C00631, C00668, C01159, C01172, C01451, C05125, C05345, C05378, C06186, C06187, C06188, C15972, C15973 and C16255 | Mtu00010 |
| Within the macrophage MTB increases the expression of isocitrate lyase, which acts in the transformation of acetyl-CoA into carbohydrates through gluconeogenesis |
Rv0889c, Rv0896, Rv1131, Rv1475c, Rv0066c, Rv3339c, Rv1248c, Rv0462, Rv2455c, Rv2454c, Rv0952, Rv0951, Rv3318, Rv0248c, Rv3319, Rv0247c, Rv3316, Rv3317, Rv1552, Rv1553, Rv1554, Rv1555, Rv1098c, Rv1240, Rv2852c, Rv2967c, Rv0211, Rv2241 and Rv2215 | C00022, C00024, C00026, C00036, C00042, C00068, C00074, C00091, C00122, C00149, C00158, C00311, C00417, C05125, C05379, C05381, C15972, C15973, C16254 and C16255 | Mtu00020 |
| Phosphate/ Energy source or phospholipid biosynthesis |
Rv0946c, Rv1121, Rv1447c, Rv1445c, Rv1122, Rv1844c, Rv1408, Rv1449c, Rv1448c, Rv2465c, Rv0478, Rv2436, Rv3068c, Rv1017c, Rv0363c, Rv1099c, Rv2029c, and Rv3010c |
C00022, C00031, C00117, C00118, C00119, C00121, C00197, C00198, C00199, C00204, C00221, C00231, C00257, C00258, C00279, C00345, C00577, C00620, C00631, C00668, 00672, C00673, C01151, C01172, C01182, C01218, C01236, C01801, C03752, C04442, C05345, C05378, C05382, C06019, C06473 and C20589. | Mtu00030 |
| Sulfur/ It is important for the initial process of protein synthesis and for maintaining a redox environment |
Rv2400c, Rv2399c, Rv2398c, Rv2397c, Rv1286, Rv1285, Rv2131c, Rv2837c, Rv2392, Rv2391, Rv0331, Rv0815c, Rv2291, Rv3117, Rv3283, Rv2335, Rv2334, Rv3684, Rv3341, Rv1079, Rv0391 and Rv3238c |
C00033, C00042, C00053, C00054, C00059, C00065, C00084, C00087, C00094, C00097, C00155, C00224, C00245, C00263, C00283, C00320, C00409, C00580, C00979, C01118, C01861, C02084, C03920, C04022, C08276, C11142, C11143, C11145, C15521, C17267, C19692, C20870 and C20955 |
Mtu00920 |
| Biotin/ Metabolism of cofactors and vitamins/ Necessary for lipid biosynthesis and gluconeogenesis |
Rv1350, Rv0242c, Rv3559c, Rv0769, Rv0032, Rv1569, Rv1568, Rv1570, Rv1589, Rv3279c and Rv1442 |
C01209, C01894, C01909, C02656, C05552, C05921, C06250, C19673, C19845, C19846, C20372, C20373, C20374, C20375, C20376, C20377, C20378, C20384, C20385, C20386, C20387, C20683 and C22458 | Mtu00780 |
| Colesterol/ Lipid Synthesis of virulence- related lipids. Providing latent infection survival products |
Rv0764c |
C00187, C00448, C00751, C01054, C01164, C01189, C01561, C01673, C01694, C01724, C01753, C01789, C01802, C01902, C01943, C02141, C02530, C03428, C03845, C04525, C05103, C05107, C05108, C05109, C05437, C05439, C05440, C05441, C05442, C05443, C07712, C08813, C08821, C08830, C11455, C11508, C11522, C11523, C15776, C15777, C15780, C15781, C15782, C15783, C15808, C15816, C15915, C18231, C21106, C22112, C22116, C22119, C22120, C22121, C22122, C22123 and C22136. | Mtu00100 |
The electron transport chain is formed for different complex of protein, and it is interesting to note that the MTB is extremely adapted to survive in different conditions of availability of energy substrates and oxygen concentration. The MTB can control your metabolism and utilized different sources of nutrients depends on your environment. In the case of aerobic conditions, like in exponential phase in vitro, the MTB uses a classical electron transport chain pathway. The classical electron transporter chain of MTB is formed by the complex Cox, Nuo, Sdh1, Hyd1, Qcr, Cta and participating of this pathway alanine dehydrogenase, proline dehydrogenase, glycerol3-phosphate dehydrogenase and Hyd2. In energy-limit conditions the complex Cox, Nuo, Sdh1, Hyd1 and Hyd2 are upregulated [50,129,132,133,134].
In case of limited concentration of oxygen, MTB uses an alternative electron transporter chain to made oxidative phosphorylation. This response for limitation in oxygen availability utilized alternate donors and acceptors of electrons, in this electron transporter chain the complexes are Mqo, Ndh, Sdh2, Hyd1, Nar, Cyd, Frd, Hyd3 and Hyd2. In this alternative electron transporter chain, the complexes upregulated are Mqo, Ndh, Sdh2, Hyd1, Hyd2, Hyd3, Cyd and Frd, Figure 4 [133,135].
5. Transport Substrates Across the Membrane: The Set Transporters from MTB
The MTB presents membrane proteins important for the transport of several substrates, these transport systems correspond to the four main superfamilies: ATP Binding Cassette (ABC) that are considered primary transporters, since they use ATP as energy source for the system; Small Multidrug Resistance (SMR), Resistance Nodulation Division (RND), Major Facilitator Super-family (MFS), which are considered secondary transporters because they use the proton motor force (H+ or Na+) as energy. The Multidrug and Toxic-compound Extrusion (MATE) family is another important superfamily of transporters but not reported in MTB [103,136,137,138,139,140,141].
MTB has the ability to capture various carbon and nitrogen sources, organic and inorganic sources of phosphate and sulfur, many of which are transported by ABC transporters. In addition, probable MTB has the presence of porins in the mycomembrane without which the uptake and essential nutrients would not be possible. However, in MTB many of these transporters still need to be characterized. About 2.5% of the MTB genome encodes for components of importers ABC transporters with at least 37 complete transporters. Importers of carbohidrates such as the UgpABCE and SugABC; anions such as the PhoT, PstACBS, CysATW and ModABC; and peptides, DppABCD and OppABCD. In the case of exporters, MTB has exporters of polysaccharides, lipids, virulence factors and antimicrobial peptides (AMPs) [142,143,144].
Figure 5.
MTB transporters. MTB presents different types of transporters that act as importers and exporters. The main MTB transporters are ABC, RND, MATE and MFS type. Also noteworthy is the presence of 5 type VII secretion systems that act on MTB virulence.
Figure 5.
MTB transporters. MTB presents different types of transporters that act as importers and exporters. The main MTB transporters are ABC, RND, MATE and MFS type. Also noteworthy is the presence of 5 type VII secretion systems that act on MTB virulence.
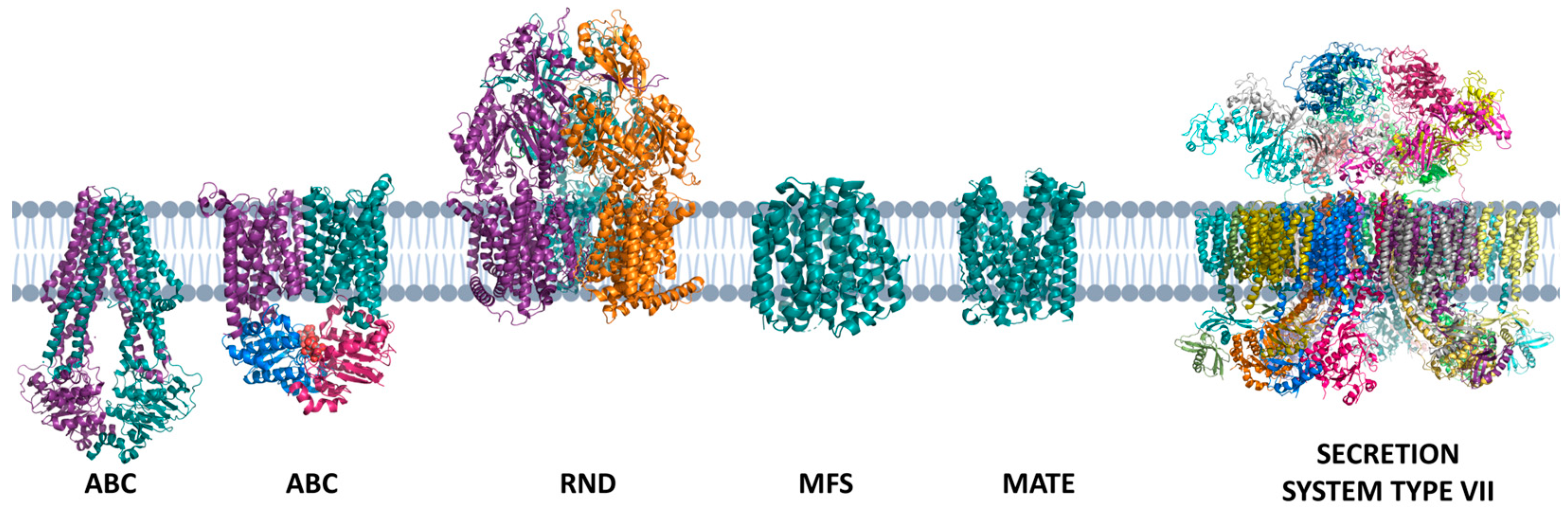
Most important are multidrug exporters ABC transporters that cause resistance to antibiotics used in the treatment of tuberculosis. Currently, four drugs are used: isoniazid (INH), pyrazinamide (PZR), ethambutol (ETH) and rifampicin (RIF) during the first two months, and the combination of rifampicin and isoniazid for another four months. The increased incidence and spread of MDR (Multi-Drug Resistance) and extensive drug resistance (XDR) has become a serious public health problem. MDR strains are resistant to isoniazid and rifampicin, first-line drugs, and XDR strains are resistant to these and to fluoroquinolones and injectable aminoglycosides [140,145,146,147].
Some ABC transporters already have a direct relationship with resistance, such as Rv1217c/1218c (RIF and INH), Rv2686c/2687c/2688c [fluoroquinolones (FLQ), such as ciprofloxacin (CPX)], and Rv1456c/1457c/1458c [RIF, INH, ETH and streptomycin (STM)]. The drug exporter best characterized in MTB and responsible for a significant increase in resistance is the DrrA/DrrB, where DrrA is the ATPase domain and the DrrB the transmembrane domain. The expression of this drug caused an increase in MIC (Minimum Inhibitory Concentration) for daunorubicin (DRB), doxorubicin (DOX), puromycin (PUR), norfloxacin (NOR), chloramphenicol (CLOR), tetracycline (TET), anthracyclines (ANT), and ETH and STM. Finally, the Rv0194 transporter, consisting of a single gene coding for both transmembrane domains and ATPAse, is a multidrug efflux pump, which promotes the increase of MIC for ampicillin (AMP), vancomycin (VAN), erythromycin (ERY), novobiocin (NOV), STM, TET and CLOR [148,149,150,151].
In addition, metals like copper need to be exported out or metabolized by MTB because are toxic for the bacteria. The MTB encodes a copper transporter, in addition to a copper-binding metallothionein, which demonstrate the presence of a detoxifying metabolism for that metal [152].
The MSF transporters are a large family of transporters that includes uniporters, simporters and antiporters. MFS has 12 or 14 transmembrane helix and transport many types of substrates, like anions, cations, amino acids, simple sugars, and drugs. The MTB has 16 open reading frames identified as MSF type efflux pumps; the Rv1634 as associated to fluoroquinolone resistance to NOR and CPX; Rv1258 confer resistance to TET, and aminoglycosides like STM; and EfpA increase the resistance to INH and other antimicrobial compounds [153].
SMR transporters are formed by homodimers or heterodimers and have four transmembranes. The protomers are antiparallel, three transmembranes are responsible for forming the pore, and the fourth transmembrane is responsible for dimerization. Only Rv3065 was described in MTB and is responsible for erythromycin (ERY), acriflavine and ethidium bromide [154,155].
MTB also has members of the RND family; these transporters are presented as homotrimers with 12 transmembranes and a large periplasmic region. The MTB has 15 putative RND transporters known as Mmpl (mycobacterial membrane protein, large), these proteins are apparently similar in structure and sequence. Four of these proteins has in our operons mmpS (mycobacterial membrane protein, small), and would have a role of adapters for the formation of a channel to the micomembrane. MmpL proteins are linked to the transport of lipids, whereas Mmpl6 is part of a detoxification system of INH and other compounds. The bacillus also presents mechanosensitive channels that are activated by the increase of tension in the bilipid layer. These channels during the severe osmotic downshock can create large pores in the membrane in a transient way that collaborate for the relief of the tension and guarantee the maintenance of cellular integrit [75,79,156,157,158,159,160]y.
The Type VII secretion system (T7SS), also known as ESX (ESAT-6 secretion) system, is a specialized protein secretion system found in mycobacteria and other Actinobacteria. It is involved in the secretion of effector proteins across the bacterial cell envelope, playing crucial roles in virulence, pathogenesis, and bacterial survival. The T7SS is composed of a complex machinery of proteins that work together to facilitate the secretion process. The core components of the T7SS include membrane-associated ATPases (EccB and EccC), membrane proteins (EccD and EccE), and a substrate recognition complex (EccB1-EccC1) [3]. These components form a transmembrane channel that spans the mycobacterial cell envelope and allows for the secretion of effector proteins [161,162,163].
The T7SS is classified into five paralogous loci, known as esx-1 to esx-5, which are present in different combinations in various mycobacterial strains. Each locus encodes a distinct set of proteins that contribute to the overall function of the T7SS. For example, the esx-1 locus is essential for the virulence of Mycobacterium tuberculosis, while the esx-3 locus is involved in iron acquisition and nutrient uptake. The secretion of effector proteins through the T7SS is mediated by specific secretion signals present in the substrates. These signals include conserved motifs such as YXXXD/e and WXG, which are recognized by the substrate recognition complex and facilitate the translocation of the effector proteins across the cell envelope [164,165,166,167,168].
The T7SS has been implicated in various functions that contribute to bacterial fitness and survival. It is involved in the modulation of host immune responses, allowing mycobacteria to evade the immune system and establish persistent infections. The T7SS also plays a role in nutrient acquisition, including iron uptake, which is crucial for mycobacterial growth and survival. Additionally, the T7SS has been implicated in interbacterial competition, where it can secrete toxins that target competitor bacteria, providing a selective advantage for the secreting strain [161,162,167,168,169].
6. Who Are MTB? The Genes Answer the Question
The MTB lineage H37Rv (most common and first to be fully sequenced) has a genome consisting of approximately 4000 genes, 225 genes linked to lipid metabolism, 227 information pathways, 517 cell wall and cellular processes, 50 stable RNAs which include rRNA and tRNA, 137 insertion elements and bacteriophages, 167 of PE and PPE proteins that are acidic and rich in N-terminal proline and glutamate, 877 genes concerning intermediate and respiration metabolism, 190 regulatory protein genes, 91 genes linked to virulence, detoxification and adaptation, 911 hypothetical conserved function genes and 605 genes of unknown function [170,171].
About 190 proteins encoded by the MTB genome are regulatory proteins, of which 11 correspond to two-component systems, five unpaired response regulators, two unpaired kinases, 11 protein kinases and another 140 regulatory transcripts. They are involved in situations of hypoxia, thermal shock, lack of nutrients, lack of metals, and responses to oxidative stress and reactive nitrogen species. MTB as well as other bacteria exhibits DNA replication from an OriC origin to the terminal ter region, in a process that occurs in two directions of the circular genome. The ribosomes have the same fractions seen in E. coli: 16s, 23s and 5s and MTB rRNAs are encoded by the rrn operon, which in MTB corresponds to a single operon [172,173,174].
MTB complex can be divide in human-adapted pathogens, which 5 lineages (lineage 1: the Philippines and Indian Ocean; lineage 2: East Asia; lineage 3: India and East Africa; lineage 4: Europe and Americas; lineage 7: Ethiopia), the Mycobacterium africanum with 2 lineages (lineage 5: M. africanum 1 in West Africa; lineage 6: M. africanum 2 in West Africa) and the animal-related species like M. bovis, M. pinnipedii, M. microti and M. orygis; and the M. canetti in East Africa. Except by M. canetti, the members of MTB complex have high genomic similarity, and the differences are found in single nucleotides polymorphisms (SNPs) and deletions in regions called regions of difference (RDs), besides phenotypic differences in virulence in host adaptability, Figure 6 [175,176,177,178,179].
The studies help to characterize 14 RDs (RD1-14) presents in M. tuberculosis H37Rv, while the regions (RvD1-5) are absent in the strain H37Rv in addition to a specific deletion region known as TbD1. M canetti has all RDs including RvD and TbD1, while M. africanum from West Africa lacks RD9 and the isolates from East Africa lacks RD3. The M. microti lacks RD7, RD8, RD9 and RD10, besides a specific region called RDMIC, M. bovis shows deletions of RD4, RD5, RD6, RD7, RD8, RD9, RD10, RD12 and RD13; while The BCG strain show deletions of the RD1, RD2 and RD14 that occurs in the attenuation process. Finally, the M. caprae is related to M. bovis except to a several nucleotides substitutions in gyrB gene. For example, the RD1 region in MTB is 9.1 kilobases in size, it consists of 9 genes, among them the gene coding for the protein 10-kDa CFP-10 and ESAT-6. The genes surrounding these two proteins encode specialized secretion systems. In addition, MTB has four more locus of secretion homologous to that found in the RD1 region [179,180,181,182].
The MTB presents two families of genes that are unique in the genus Mycobacterium, families PE and PPE that encode proteins very similar to ESAT-6 and CFP-10 proteins. In the pathogenic mycobacteria, these two families of genes are present in great quantity when compared with other gene families [183,184,185].
In Mycobacterium tuberculosis, several types of non-coding RNA have been identified, including antisense transcripts, riboswitches, and small intergenic RNAs (sRNAs). Antisense transcripts are short RNA molecules that base-pair with the 5' regions of mRNAs. They have been found to regulate gene expression in M. tuberculosis, including genes involved in the transposition of foreign genetic elements. These antisense transcripts can range in size from less than 50 nucleotides to several kilobases and can map to sequences throughout the length of cognate mRNAs [186,187].
Riboswitches are RNA elements that can bind specific ligands and regulate gene expression. In M. tuberculosis, riboswitches have been identified, such as the cobalamin riboswitch, which represses gene expression in the presence of specific ligands. Small intergenic RNAs (sRNAs) are short transcripts that are encoded between open reading frames. They can regulate gene expression by base-pairing with target mRNAs, thereby modulating translation efficiency and mRNA stability. These sRNAs have been shown to be differentially expressed in response to changing environments and may play important roles in the regulation of gene expression and growth in M. tuberculosis [188,189].
The presence of these non-coding RNAs in M. tuberculosis suggests that post-transcriptional regulation plays a significant role in the pathogenesis of the disease. They have the potential to regulate the expression of genes involved in virulence and adaptation, contributing to the ability of the bacterium to cause disease and persist in the host [186,187].
A regulatory component of MTB gene transcription consists of the DosR-DosS/DosT system, which controls other regulons known to activate the bacillus dormancy stages under microaerophilic conditions. This regulon also controls genes linked to the production and activation of respiration using alternative electron acceptors. Two sensors that are histidine kinases proteins, DosS and DosT, phosphorylate the Dos regulon. Once activated this regulon controls about fifty genes directly, many of which still have unknown function. It is known that besides the hypoxia condition, situations such as nitric oxide concentration also activate this regulon. The redox stress that is inside the macrophage is also another factor that can activate the Dos regulon. Although this regulon is activated in the early stages of hypoxia, studies have shown that it is not essential for the survival of bacillus in the state of dormancy. It is also noted that in mutants with deletion in genes coding for heat shock proteins, the ability to remain inside macrophages is lost [135,190,191,192,193,194,195].
MTB presents three distinct systems for DNA repair, such as homologous recombination, non-homologous junction and single-strand annealing. Each of these repair systems is activated according to the type of lesion that has occurred and its extension, in addition, are extremely coordinated processes by different sets of genes and proteins. MDR strains have shown mutations in genes linked to DNA repair, which may be the cause of hypermutations in the genes that code for the targets of the drugs and that, therefore, would promote a rapid adaptation of the strain to the treatment and action of the immune system [196,197,198,199,200,201,202].
In MTB the genes that are regulated by the sigH, rpoV and whiB3 genes are related to controls of host immune response that facilitate the spread of MTB to new individuals. Furthermore, MTB have a set of genes designated to lipid and fatty acid metabolism, including the catabolism of cholesterol; inhibitors of antimicrobial effectors of macrophage, like inhibitors of apoptosis, proteins to oxidative and nitrosative stress and phagosome arresting; metal transporters, importers and exporters; proteases, including metalloproteases and β-lactamases; cell wall proteins, like lipoproteins and secretion systems; gene expression regulators, including sigma factors, two component systems and transcriptional regulators; and others destinates to survive and virulence of MTB [203,204,205,206].
MTB presents 13 sigma factors currently described, all belonging to the σ70 family, which recognizes regions -35 and -10 of the promoter. Interestingly, MTB does not show any sigma factor of the σ54 family, which recognizes regions -12 and -24 of the promoter. The σ70 MTB, present representatives of the 4 groups of this family: Group 1, represented by σA; group 2, represented by σB; group 3, represented by σF; and finally, another 10 sigmas representing group 4. The factors σA and σB are present in all mycobacteria, as well as σE, the sigmas σD, σF, σG, σH and σj are found in all mycobacteria except M. leprae. However, σC is present only in pathogenic mycobacteria, Table 4 [207,208,209,210,211,212,213,214].
Another regulation in gene transcription can be performed through anti-sigmas. These proteins interact with sigma factors, preventing them from binding to the promoter or promoting the disengagement of the factor from promoter. In MTB the anti-sigmas RsmA, σM anti-sigma were described; RseA, σE anti-sigma; RshA, σH anti-sigma; RslA; σL anti-sigma; RskA; σK anti-sigma; RsdA; σD anti-sigma; RsfA, σF anti-sigma. Anti- sigma Rv0093c and Rv0181c, which are sigma anti-sigma σC and σG, have been identified putatively. The anti-sigmas σE, σF e σH were identified as proteins present in the cytoplasm, whereas the other anti-sigmas would be proteins located in the plasma membrane.
In addition, anti-anti-sigma factors, whose function is to block anti-sigmas, are Rv1635c, Rv3687c, Rv0516c, Rv1364c, Rv1904 and Rv2638 blockers of σ F anti- sigma. Interestingly, some sigma factors self-regulate their levels, binding in their own promoters, is the case of factors σA, σB, σD, σF, σE, σH, σK, σL and σM. While sigmas also regulate other sigmas, as is the case of σH regulates the sigmas σB and σE; the σE, regulates the σB; which is also regulated by sigma σL and σF. The σF also regulates the σC. The σG also regulates the sigmas σh and σF. Finally, σJ regulates the factor σI. Regarding the MTB promoters, research indicates that the -10 region of the promoters are similarity to the E. coli promoters, however, the -35 region appears to be totally different from the regions described for E. coli. In addition, there is variation in the distance between these two regions when compared to these two bacteria, as in E. coli an AT-rich region at position -15 is observed, due to the high content of CG present in the genome, is absent in MTB[154,204,212,213,214,215,216].
In MTB the virulence and pathogenesis are directly linked to its ability to survive within the host. The disease is a consequence of the survival and replication of MTB in the host and its relation to the immune system. It is interesting to note that MTB is extremely adapted to survive under the extreme conditions that lie within the macrophages and other cells of the immune system. In addition to possessing an incredible capacity to modulate not only its metabolism and cell division under these conditions but also, manipulate the immune responses of the host [16,118,180,217,218,219,220].
A curious fact of MTB is the absence of classical virulence factors such as toxins. However, the presence of several genes whose functions allow the survival, adaptation, persistence, control and dissemination of MTB in the host are observed. The main virulence factors of MTB are discussed below, Table 5 [128,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235].
Table 4.
Sigma factor of MTB.
| Gene | Function | Category |
|---|---|---|
| Rv2940 | Promotes the synthesis of long chain fatty acids | Cell Wall |
| Rv2930 | Acyl-coenzyme A, promotes the degradation of fatty acids | |
| Rv2941 | Fatty-acid-CoA synthetase that acts in the lipidic pathway |
|
| Rv2942 | Promotes the transport of lipids and synthesis of the mycomembrane | |
| Rv3804c | Promotes the transfer of mycolic acids to trehalose |
|
| Rv0642c and Rv0410 | A methyltransferase bound to the synthesis of mycolic acids | Cell Wall |
| Rv0899 | It is a protein expressed at low pH | |
| Rv0475 | It is a heparin-binding hemagglutinin protein | |
|
RD1 region |
Inhibition of phagosome maturation and apoptosis. Responsible for encoding a network of secretion systems |
|
|
Rv2246 |
It is a culture filtrate protein whose function is a chaperonin linked to latency and persistence | |
| Rv3763 | It is a culture filtration protein that promotes the regulation of IL-12 | |
|
Rv1811 |
Acts on magnesium uptake |
|
|
Rv3083 to Rv3089, and Rv2869c |
MTB related to mycolic acid synthesis is mymA operon |
|
|
Rv2946C, Rv1660, Rv2048c, Rv2941, Rv2938, 1527c, Rv1661, Rv3823c, Rv1345 and Rv1916 |
Linked to complex lipid synthesis | |
| Rv1411c, Rv1410c, Rv0934, Rv1235 and Rv1857 | Lipoproteins that constitute virulence factors |
|
| Rv3682 | Transglucosylases and transpeptidase | |
| Rv2136 | Involved in the synthesis of peptidoglycan |
|
| Rv0198c and Rv2869c | Metallo-proteases | |
| Rv2097c and Rv2115c | Proteasome associated proteins |
|
| Rv2382c, Rv1348, Rv1349, Rv2711 and Rv1811 | Related to metal transporter | Cell Wall |
| Rv3270 | Zinc efflux | |
| Rv0969 | Cupper efflux | |
|
Rv3367, Rv1818c and Rv2136c |
Rv3367, Rv1818c and Rv2136c |
|
|
MT18B_4990, Rv1411c, Rv1270c and Rv0934 |
Lipoproteins that are Toll Like Receptor-2 (TLR2) agonists and their Myeloid Differentiation Primary-Response protein 88 (MYD88) |
|
|
Rv0350, Rv1860, MT18B_4990 and Rv1436 |
Allow the bacillus to bind to cells, either phagocytosed and continue to replicate |
Intermediary metabolism and respiration |
|
Rv2220 |
A glutamine synthetase, also constitutes a culture filtration protein, acts on the metabolism of nitrogen |
|
| Rv0467 | Isocytrase lyase, converts isocitrate to succinate and allows bacterial growth under fatty acids and acetate | |
| Rv3487 | It is a lipase esterase that acts on lipid degradation |
|
| Rv1345 | Acts on β- oxidation of fatty acids | |
|
Rv2351c, Rv2350c, Rv2349c and Rv1755c |
Phospholipases involved in the cycles of obtaining energy |
|
|
Rv3602c and Rv3601 |
Consist of pantothenate synthase proteins, this molecule that acts on the degradation of lipids and other cell signaling |
|
| Rv2987, Rv2192, Rv0500 and Rv0780 | Act on the biosynthesis of leucine, tryptophan, proline and purines respectively | |
|
Rv1161 |
Involved with respiration under anaerobic conditions and the conversion of nitrate to nitrite |
Intermediary metabolism and respiration |
|
Rv0475, Rv0930, Rv0820, Rv2224c, Rv3236c, (Rv3666c to Rv3663c) and Rv2200c |
Linked to cell wall | |
| Rv3883c | Proteases involved with virulence | |
| Rv0983 and Rv3671c | Serine proteases | |
| Rv3810 | Act with multiplication and intracellular growth | |
| Rv3671c | Encodes a membrane protein responsible for MTB resistance to the acidic environment of IFN-γ-activated phagosomes | |
|
Rv0195, Rv0386, Rv0491, Rv0890c, Rv0894, Rv3416, Rv3133c, Rv1013, Rv2946c, Rv2488c and Rv3133c |
Formation of biofilms. This structure is involved in bacterial persistence and protects it from chemical and physical agents |
Regulator Proteins |
|
Rv2711 |
Binds to regions of genes involved in iron uptake and nitrate reductase |
|
| Rv0757 | Controls the expression of virulence genes by magnesium deficiency | |
| Rv0903c | Regulates macrophage virulence genes | |
| Rv0981 | Two-component system that regulates macrophage virulence genes | |
|
Rv3416 |
Cytoplasmic redox sensor, linked to pH resistance |
|
| Rv0931c and Rv0410c | Proteins kinases related to virulence | Regulator Proteins |
| Rv2745c | ATP-dependent protease | |
|
Rv1908c |
Catalase: peroxidase that degrades peroxides and other organic peroxides |
Virulence, detoxification, adaptation |
| Rv2428 | Protein whose function is to detoxify hydroperoxides |
|
| Rv3846 and Rv0342 | Act in the detoxification of superoxide | |
| Rv0432 | Superoxide dismutase | |
| Rv1936 to Rv1941 and Rv1908c | Catalase-peroxidase protein |
|
| Rv1932 | Thiol peroxidase | |
| Rv0353 | gene repressor of proteins of heat shock |
|
| Rv0251c | Possibly a molecular chaperone | |
| Rv3409c, Rv3568, Rv34545c, Rv3544c, Rv3543c, Rv3542c, Rv3541c and Rv3540c | Linked to catabolism of cholesterol | Lipid metabolism |
| Rv2383c | Linked to production mycobactin, an important siderophore in MTB |
|
| Rv2246 | protein involved in lipid and fatty acid metabolism |
|
| Rv3151, Rv1743, Rv3654c and Rv3655c | Involved with the inhibition of apoptosis |
Conserved hypotheticals |
| Rv2027c, Rv0490, Rv0981, Rv0982, Rv2395A and Rv2395B | Regulatory proteins | |
| Rv2032, Rv0211, Rv0153c and Rv0990c | Virulence factors are found the region of difference RD2 | Conserved hypotheticals |
| Rv2445c, Rv2234 and Rv1651c | Involved in phagosome arresting | PE/PPE |
5. Conclusions
The ability of MTB to survive within the macrophage and to spread during active disease is well known. MTB is one of the most adapted bacteria to survive against the human immune system and demonstrates great metabolic and regulatory versatility that allowed it to evolve with its host for thousands of years.
The MTB cell wall is extremely complex and protects against various external agents. There is still much debate about its organization; could mycolic acids be organized in a similar way to the Gram-negative outer membrane? More studies are needed to better understand their characteristics and components.
The pathogenic capacity of the MTB is due mainly to its unique characteristics, whether genetic, proteomic or metabolic. Understanding the biology of these microorganisms is extremely important, as it is one of the world's major infectious diseases with few treatment options.
In addition, a better understanding of the physiology and molecular biology of MTB will allow the development of new diagnostic and treatment techniques, which are currently scarce due to the emergence of resistant strains.
Author Contributions
The authors also contributed to the conception, research and realization of this work.
Funding
The authors would like the National Council for Scientific and Technological Development, CNPq - Brazil (process number 163258/2022-8) for all support during the experiment.
Acknowledgments
The authors would like to thank the Universidade Cidade de São Paulo - Unicid, for supporting the work developed and the coordination of the Biomedicine course for all the support during the execution of the project.
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- World Health Organization, (WHO) WHO Report on TB 2020; 2020; Vol. 1; ISBN 9783642253874.
- World Health Organization WHO Consolidated Guidelines on Tuberculosis.; 2022; ISBN 978-92-4-155051-2.
- Organización Mundial de la Salud Global Tuberculosis Report 2019 OMS - WHO. World Health Organization 2019.
- Zhang, Q. ao; Ma, S.; Li, P.; Xie, J. The Dynamics of Mycobacterium Tuberculosis Phagosome and the Fate of Infection. Cell Signal 2023, 108. [Google Scholar] [CrossRef]
- Huang, Y.; Ai, L.; Wang, X.; Sun, Z.; Wang, F. Review and Updates on the Diagnosis of Tuberculosis. J Clin Med 2022, 11. [Google Scholar] [CrossRef]
- Natarajan, A.; Beena, P.M.; Devnikar, A. V.; Mali, S. A Systemic Review on Tuberculosis. Indian Journal of Tuberculosis 2020, 67. [Google Scholar] [CrossRef] [PubMed]
- Belhaouane, I.; Pochet, A.; Chatagnon, J.; Hoffmann, E.; Queval, C.J.; Deboosère, N.; Boidin-Wichlacz, C.; Majlessi, L.; Sencio, V.; Heumel, S.; et al. Tirap Controls Mycobacterium Tuberculosis Phagosomal Acidification. PLoS Pathog 2023, 19. [Google Scholar] [CrossRef] [PubMed]
- Echeverría-Valencia, G. Phagocytosis of Mycobacterium Tuberculosis : A Narrative of the Uptaking and Survival. IntechOpen 2023. [Google Scholar] [CrossRef]
- Pethe, K.; Swenson, D.L.; Alonso, S.; Anderson, J.; Wang, C.; Russell, D.G. Isolation of Mycobacterium Tuberculosis Mutants Defective in the Arrest of Phagosome Maturation. Proc Natl Acad Sci U S A 2004, 101, 13642–13647. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat Rev Dis Primers 2016, 2. [Google Scholar] [CrossRef]
- Park, C.K.; Kwon, Y.S. Respiratory Review of 2014: Tuberculosis and Nontuberculous Mycobacterial Pulmonary Disease. Tuberc Respir Dis (Seoul) 2014. [Google Scholar] [CrossRef]
- Hartman-Adams, H.; Clark, K.; Juckett, G. Update on Latent Tuberculosis Infection. Am Fam Physician 2014. [Google Scholar]
- Kroon, E.E.; Kinnear, C.; Orlova, M.; Fischinger, S.; Shin, S.; Boolay, S.; Walzl, G.; Jacobs, A.; Wilkinson, R.; Alter, G.; et al. A Case-Control Study Identifying Highly Tuberculosis-Exposed, HIV-1-Infected but Persistently TB, Tuberculin and IGRA Negative Persons with M. Tuberculosis Specific Antibodies in Cape Town, South Africa. medRxiv 2020, null. [Google Scholar] [CrossRef]
- WHO The End TB Strategy. J Chem Inf Model 2013, 53, 1689–1699.
- Van Soolingen, D.; Hoogenboezem, T.; De Haas, P.E.W.; Hermans, P.W.M.; Koedam, M.A.; Teppema, K.S.; Brennan, P.J.; Besra, G.S.; Portaels, F.; Top, J.; et al. A Novel Pathogenic Taxon of the Mycobacterium Tuberculosis Complex, Canetti: Characterization of an Exceptional Isolate from Africa. Int J Syst Bacteriol 1997, 47, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Orgeur, M.; Brosch, R. Evolution of Virulence in the Mycobacterium Tuberculosis Complex. Curr Opin Microbiol 2018, 41, 68–75. [Google Scholar] [CrossRef]
- Cole, S.T. Comparative and Functional Genomics of the Mycobacterium Tuberculosis Complex. Microbiology (N Y) 2015, 1851, 11–2919. [Google Scholar]
- Žmak, L.; Janković, M.; Obrovac, M.; Katalinić-Janković, V. Non-Tuberculous Mycobacteria. Infektoloski Glasnik 2013, 33. [Google Scholar] [CrossRef]
- George, M. Cutaneous Non-Tuberculous Mycobacterial Infections: An Update. Journal of Skin and Sexually Transmitted Diseases 2023, 0. [Google Scholar] [CrossRef]
- Katoch, V.M. Infections Due to Non-Tuberculous Mycobacteria (NTM). Indian Journal of Medical Research 2004, 120. [Google Scholar]
- Bhalla, G.S.; Sarao, M.S.; Kalra, D.; Bandyopadhyay, K.; John, A.R. Methods of Phenotypic Identification of Non-Tuberculous Mycobacteria. Pract Lab Med 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- de Lima, C.A.M.; Gomes, H.M.; Oelemann, M.A.C.; Ramos, J.P.; Caldas, P.C.; Campos, C.E.D.; Pereira, M.A. da S.; Montes, F.F.O.; de Oliveira, M. do S.C.; Suffys, P.N.; et al. Nontuberculous Mycobacteria in Respiratory Samples from Patients with Pulmonary Tuberculosis in the State of Rondônia, Brazil. Mem Inst Oswaldo Cruz 2013, 108, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Refaya, A.K.; Kumar, N.; Raj, D.; Veerasamy, M.; Balaji, S.; Shanmugam, S.; Rajendran, A.; Tripathy, S.P.; Swaminathan, S.; Peacock, S.J.; et al. Whole-Genome Sequencing of a Mycobacterium Orygis Strain Isolated from Cattle in Chennai, India. Microbiol Resour Announc 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Zimpel, C.K.; Brandão, P.E.; de Souza Filho, A.F.; de Souza, R.F.; Ikuta, C.Y.; Neto, J.S.F.; Soler Camargo, N.C.; Heinemann, M.B.; Guimarães, A.M.S. Complete Genome Sequencing of Mycobacterium Bovis SP38 and Comparative Genomics of Mycobacterium Bovis and M. Tuberculosis Strains. Front Microbiol 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Onipede, A.; Gagneux, S.; Niemann, S.; Kremer, K.; Desmond, E.; Kato-Maeda, M.; Behr, M. Genomic Analysis Distinguishes Mycobacterium Africanum. J Clin Microbiol 2004, 42, 3594–3599. [Google Scholar] [CrossRef] [PubMed]
- Brudey, K.; Gutierrez, M.; Vincent, V.; Parsons, L.; Salfinger, M.; Rastogi, N.; Sola, C. Mycobacterium Africanum Genotyping UsingNovel Spacer Oligonucleotides in the Direct RepeatLocus. J Clin Microbiol 2004, 42, 5053–5057. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bannantine, J.P.; Zhang, Q.; Amonsin, A.; May, B.J.; Alt, D.; Banerji, N.; Kanjilal, S.; Kapur, V. The Complete Genome Sequence of Mycobacterium Avium Subspecies Paratuberculosis. Proc Natl Acad Sci U S A 2005, 102, 12344–12349. [Google Scholar] [CrossRef]
- Wu, C.-W.; Schmoller, S.K.; Bannantine, J.P.; Eckstein, T.M.; Inamine, J.M.; Livesey, M.; Albrecht, R.; Talaat, A.M. A Novel Cell Wall Lipopeptide Is Important for Biofilm Formation and Pathogenicity of Mycobacterium Avium Subspecies Paratuberculosis. Microb Pathog 2009, 46, 222–230. [Google Scholar] [CrossRef]
- Sugino, K.; Ono, H.; Ando, M.; Tsuboi, E. Pleuroparenchymal Fibroelastosis in Mycobacterium Avium Complex Lung Disease. Respirol Case Rep 2023, 11. [Google Scholar] [CrossRef]
- Urabe, N.; Sakamoto, S.; Masuoka, M.; Kato, C.; Yamaguchi, A.; Tokita, N.; Homma, S.; Kishi, K. Efficacy of Three Sputum Specimens for the Diagnosis of Mycobacterium Avium Complex Pulmonary Disease. BMC Pulm Med 2023, 23. [Google Scholar] [CrossRef]
- Hendrix, C.; McCrary, M.; Hou, R.; Abate, G. Diagnosis and Management of Pulmonary NTM with a Focus on Mycobacterium Avium Complex and Mycobacterium Abscessus: Challenges and Prospects. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Comincini, S.; Barbarini, D.; Telecco, S.; Bono, L.; Marone, P. Rapid Identification of Mycobacterium Tuberculosis and Mycobacterium Avium by Polymerase Chain Reaction and Restriction Enzyme Analysis within Sigma Factor Regions. New Microbiologica 1998, 21. [Google Scholar]
- Nqwata, L.; Ouédrago, A.R. Non-Tuberculous Mycobacteria Pulmonary Disease: A Review of Trends, Risk Factors, Diagnosis and Management. African Journal of Thoracic and Critical Care Medicine 2022, 28. [Google Scholar]
- Ayerakwa, E.A.; Abban, M.K.; Isawumi, A.; Mosi, L. Profiling Mycobacterium Ulcerans: Sporulation, Survival Strategy and Response to Environmental Factors. Future Sci OA 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Tchan, B.G.O.; Ngazoa-Kakou, S.; Aka, N.; Apia, N.K.B.; Hammoudi, N.; Drancourt, M.; Saad, J. PPE Barcoding Identifies Biclonal Mycobacterium Ulcerans Buruli Ulcer, Côte d’Ivoire. Microbiol Spectr 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, T.; Hagiya, H.; Iio, K.; Yamasaki, O.; Miyamoto, Y.; Hoshino, Y.; Kakehi, A.; Okura, M.; Minabe, H.; Yokoyama, Y.; et al. Buruli Ulcer Caused by Mycobacterium Ulcerans Subsp. Shinshuense: A Case Report. Journal of Infection and Chemotherapy 2023, 29. [Google Scholar] [CrossRef]
- Tweedale, B.; Collier, F.; Waidyatillake, N.T.; Athan, E.; O’Brien, D.P. Mycobacterium Ulcerans Culture Results According to Duration of Prior Antibiotic Treatment: A Cohort Study. PLoS One 2023, 18. [Google Scholar] [CrossRef]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-Tuberculous Mycobacteria and the Rise of Mycobacterium Abscessus. Nat Rev Microbiol 2020, 18, 392–407. [Google Scholar] [CrossRef]
- Stout, J.E.; Koh, W.J.; Yew, W.W. Update on Pulmonary Disease Due to Non-Tuberculous Mycobacteria. International Journal of Infectious Diseases 2016, 45, 123–134. [Google Scholar] [CrossRef]
- Zammarchi, L.; Tortoli, E.; Borroni, E.; Bartalesi, F.; Strohmeyer, M.; Baretti, S.; Simonetti, M.T.; Liendo, C.; Santini, M.G.; Rossolini, G.M.; et al. High Prevalence of Clustered Tuberculosis Cases in Peruvian Migrants in Florence, Italy. Infect Dis Rep 2014. [Google Scholar] [CrossRef]
- Váradi, O.A.; Rakk, D.; Spekker, O.; Terhes, G.; Urbán, E.; Berthon, W.; Pap, I.; Szikossy, I.; Maixner, F.; Zink, A.; et al. Verification of Tuberculosis Infection among Vác Mummies (18th Century CE, Hungary) Based on Lipid Biomarker Profiling with a New HPLC-HESI-MS Approach. Tuberculosis 2021, 126. [Google Scholar] [CrossRef]
- Lalremruata, A.; Ball, M.; Bianucci, R.; Welte, B.; Nerlich, A.G.; Kun, J.F.J.; Pusch, C.M. Molecular Identification of Falciparum Malaria and Human Tuberculosis Co-Infections in Mummies from the Fayum Depression (Lower Egypt). PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Zmak, L.; Obrovac, M.; Makek, M.J.; Perko, G.; Trkanjec, J.T. From Peruvian Mummies to Living Humans: First Case of Pulmonary Tuberculosis Caused by Mycobacterium Pinnipedii. International Journal of Tuberculosis and Lung Disease 2019, 23. [Google Scholar] [CrossRef] [PubMed]
- Zink, A.R.; Sola, C.; Reischl, U.; Grabner, W.; Rastogi, N.; Wolf, H.; Nerlich, A.G. Characterization of Mycobacterium Tuberculosis Complex DNAs from Egyptian Mummies by Spoligotyping. J Clin Microbiol 2003, 41. [Google Scholar] [CrossRef] [PubMed]
- Basta, P.C.; Camacho, L.A.B. Tuberculin Skin Test to Estimate the Prevalence of Mycobacterium Tuberculosis Infection in Indigenous Populations in the Americas: A Literature Review. Cad Saude Publica 2006, 22, 245–254. [Google Scholar] [CrossRef]
- VanderVen, B.; Huang, L.; Rohde, K.; Russell, D. The Minimal Unit of Infection: Mycobacterium Tuberculosis in the Macrophage. Microbiol Spectr 2016, 4 6, null. [Google Scholar] [CrossRef]
- Xu, G.; Wang, J.; Gao, G.F.; Liu, C.H. Insights into Battles between Mycobacterium Tuberculosis and Macrophages. Protein Cell 2014, 5, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Jackson, M.; Ma, Y.; McNeil, M. Cell Wall Core Galactofuran Synthesis Is Essential for Growth of Mycobacteria. J Bacteriol 2001, 183. [Google Scholar] [CrossRef]
- Jacobo-Delgado, Y.M.; Rodríguez-Carlos, A.; Serrano, C.J.; Rivas-Santiago, B. Mycobacterium Tuberculosis Cell-Wall and Antimicrobial Peptides: A Mission Impossible? Front Immunol 2023, 14. [Google Scholar] [CrossRef]
- Hoffmann, C.; Leis, A.; Niederweis, M.; Plitzko, J.M.; Engelhardt, H. Disclosure of the Mycobacterial Outer Membrane: Cryo-Electron Tomography and Vitreous Sections Reveal the Lipid Bilayer Structure. Proc Natl Acad Sci U S A 2008, 105, 3963–3967. [Google Scholar] [CrossRef]
- Huang, K.C.; Mukhopadhyay, R.; Wen, B.; Gitai, Z.; Wingreen, N.S. Cell Shape and Cell-Wall Organization in Gram-Negative Bacteria. Proc Natl Acad Sci U S A 2008, 105. [Google Scholar] [CrossRef]
- Heinz, C.; Engelhardt, H.; Niederweis, M. The Core of the Tetrameric Mycobacterial Porin MspA Is an Extremely Stable β-Sheet Domain. Journal of Biological Chemistry 2003, 278. [Google Scholar] [CrossRef]
- Mahfoud, M.; Sukumaran, S.; Hülsmann, P.; Grieger, K.; Niederweis, M. Topology of the Porin MspA in the Outer Membrane of Mycobacterium Smegmatis. Journal of Biological Chemistry 2006, 281. [Google Scholar] [CrossRef]
- Stahl, C.; Kubetzko, S.; Kaps, I.; Seeber, S.; Engelhardt, H.; Niederweis, M. MspA Provides the Main Hydrophilic Pathway through the Cell Wall of Mycobacterium Smegmatis. Mol Microbiol 2001, 40. [Google Scholar] [CrossRef]
- Pavlenok, M.; Niederweis, M. Hetero-Oligomeric MspA Pores in Mycobacterium Smegmatis. FEMS Microbiol Lett 2016, 363. [Google Scholar] [CrossRef]
- Hillmann, D.; Eschenbacher, I.; Thiel, A.; Niederweis, M. Expression of the Major Porin Gene MspA Is Regulated in Mycobacterium Smegmatis. J Bacteriol 2007, 189. [Google Scholar] [CrossRef] [PubMed]
- Sharbati-Tehrani, S.; Meister, B.; Appel, B.; Lewin, A. The Porin MspA from Mycobacterium Smegmatis Improves Growth of Mycobacterium Bovis BCG. International Journal of Medical Microbiology 2004, 294. [Google Scholar] [CrossRef] [PubMed]
- Huff, J.; Pavlenok, M.; Sukumaran, S.; Niederweis, M. Functions of the Periplasmic Loop of the Porin MspA from Mycobacterium Smegmatis. Journal of Biological Chemistry 2009, 284. [Google Scholar] [CrossRef] [PubMed]
- Maitra, A.; Munshi, T.; Healy, J.; Martin, L.T.; Vollmer, W.; Keep, N.H.; Bhakta, S. Cell Wall Peptidoglycan in Mycobacterium Tuberculosis: An Achilles’ Heel for the TB-Causing Pathogen. FEMS Microbiol Rev 2019, 43, 548–575. [Google Scholar] [CrossRef]
- Wang, Q.; Matsuo, Y.; Pradipta, A.R.; Inohara, N.; Fujimoto, Y.; Fukase, K. Synthesis of Characteristic Mycobacterium Peptidoglycan (PGN) Fragments Utilizing with Chemoenzymatic Preparation of Meso-Diaminopimelic Acid (DAP), and Their Modulation of Innate Immune Responses. Org Biomol Chem 2016, 14, 1013–1023. [Google Scholar] [CrossRef]
- Block, A.M.; Namugenyi, S.B.; Palani, N.P.; Brokaw, A.M.; Zhang, L.; Beckman, K.B.; Tischler, A.D. Mycobacterium Tuberculosis Requires the Outer Membrane Lipid Phthiocerol Dimycocerosate for Starvation-Induced Antibiotic Tolerance. mSystems 2023, 8. [Google Scholar] [CrossRef]
- Babajan, B.; Chaitanya, M.; Rajsekhar, C.; Gowsia, D.; Madhusudhana, P.; Naveen, M.; Chitta, S.K.; Anuradha, C.M. Comprehensive Structural and Functional Characterization of Mycobacterium Tuberculosis UDP-NAG Enolpyruvyl Transferase (Mtb-MurA) and Prediction of Its Accurate Binding Affinities with Inhibitors. Interdiscip Sci 2011, 3. [Google Scholar] [CrossRef]
- Shinde, Y.; Ahmad, I.; Surana, S.; Patel, H. The Mur Enzymes Chink in the Armour of Mycobacterium Tuberculosis Cell Wall. Eur J Med Chem 2021, 222. [Google Scholar] [CrossRef]
- Verma, A.; Kumar, V.; Naik, B.; Masood Khan, J.; Singh, P.; Erik Joakim Saris, P.; Gupta, S. Screening and Molecular Dynamics Simulation of Compounds Inhibiting MurB Enzyme of Drug-Resistant Mycobacterium Tuberculosis: An in-Silico Approach. Saudi J Biol Sci 2023, 30. [Google Scholar] [CrossRef] [PubMed]
- Eniyan, K.; Rani, J.; Ramachandran, S.; Bhat, R.; Khan, I.A.; Bajpai, U. Screening of Antitubercular Compound Library Identifies Inhibitors of Mur Enzymes in Mycobacterium Tuberculosis. SLAS Discovery 2020, 25. [Google Scholar] [CrossRef]
- Rossini, N. de O.; Silva, C.; Dias, M.V.B. The Crystal Structure of Mycobacterium Thermoresistibile MurE Ligase Reveals the Binding Mode of the Substrate M-Diaminopimelate. J Struct Biol 2023, 215. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Singh, N.; Kumar, R.; Kushwaha, N.K.; Prajapati, V.M.; Singh, S.K. GlfT1 Down-Regulation Affects Mycobacterium Tuberculosis Biofilm Formation and Its in-Vitro and in-Vivo Survival. Tuberculosis 2023, 141. [Google Scholar] [CrossRef] [PubMed]
- Alderwick, L.J.; Dover, L.G.; Veerapen, N.; Gurcha, S.S.; Kremer, L.; Roper, D.L.; Pathak, A.K.; Reynolds, R.C.; Besra, G.S. Expression, Purification and Characterisation of Soluble GlfT and the Identification of a Novel Galactofuranosyltransferase Rv3782 Involved in Priming GlfT-Mediated Galactan Polymerisation in Mycobacterium Tuberculosis. Protein Expr Purif 2008, 58. [Google Scholar] [CrossRef] [PubMed]
- Babu Sait, M.R.; Koliwer-Brandl, H.; Stewart, J.A.; Swarts, B.M.; Jacobsen, M.; Ioerger, T.R.; Kalscheuer, R. PPE51 Mediates Uptake of Trehalose across the Mycomembrane of Mycobacterium Tuberculosis. Sci Rep 2022, 12. [Google Scholar] [CrossRef]
- Chiaradia, L.; Lefebvre, C.; Parra, J.; Marcoux, J.; Burlet-Schiltz, O.; Etienne, G.; Tropis, M.; Daffé, M. Dissecting the Mycobacterial Cell Envelope and Defining the Composition of the Native Mycomembrane. Sci Rep 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Grzegorzewicz, A.E.; Eynard, N.; Quémard, A.; North, E.J.; Margolis, A.; Lindenberger, J.J.; Jones, V.; Korduláková, J.; Brennan, P.J.; Lee, R.E.; et al. Covalent Modification of the Mycobacterium Tuberculosis FAS-II Dehydratase by Isoxyl and Thiacetazone. ACS Infect Dis 2016, 1. [Google Scholar] [CrossRef]
- Grzegorzewicz, A.E.; Lelièvre, J.; Esquivias, J.; Angala, B.; Liu, J.; Lee, R.E.; McNeil, M.R.; Jackson, M. Lack of Specificity of Phenotypic Screens for Inhibitors of the Mycobacterium Tuberculosis FAS-II System. Antimicrob Agents Chemother 2021, 65. [Google Scholar] [CrossRef]
- Cantaloube, S.; Veyron-Churlet, R.; Haddache, N.; Daffé, M.; Zerbib, D. The Mycobacterium Tuberculosis FAS-II Dehydratases and Methyltransferases Define the Specificity of the Mycolic Acid Elongation Complexes. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.J.; Dechow, S.J.; Abramovitch, R. Acid Fasting: Modulation of Mycobacterium Tuberculosis Metabolism at Acidic PH. Trends Microbiol 2019, null, null. [Google Scholar] [CrossRef] [PubMed]
- Moolla, N.; Bailo, R.; Marshall, R.; Bavro, V.N.; Bhatt, A. Structure-Function Analysis of MmpL7-Mediated Lipid Transport in Mycobacteria. The Cell Surface 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Nakata, N.; Mukai, T.; Kawagishi, I.; Ato, M. Coexpression of MmpS5 and MmpL5 Contributes to Both Efflux Transporter MmpL5 Trimerization and Drug Resistance in Mycobacterium Tuberculosis. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Ma, S.; Huang, Y.; Xie, F.; Gong, Z.; Zhang, Y.; Stojkoska, A.; Xie, J. Transport Mechanism of Mycobacterium Tuberculosis MmpL/S Family Proteins and Implications in Pharmaceutical Targeting. Biol Chem 2020, 401. [Google Scholar] [CrossRef]
- Williams, J.T.; Abramovitch, R.B. Molecular Mechanisms of MmpL3 Function and Inhibition. Microbial Drug Resistance 2023, 29. [Google Scholar] [CrossRef]
- Pérez, J.; Garcia, R.; Bach, H.; de Waard, J.H.; Jacobs, W.R.; Av-Gay, Y.; Bubis, J.; Takiff, H.E. Mycobacterium Tuberculosis Transporter MmpL7 Is a Potential Substrate for Kinase PknD. Biochem Biophys Res Commun 2006, 348. [Google Scholar] [CrossRef]
- Adams, O.; Deme, J.C.; Parker, J.L.; Fowler, P.W.; Lea, S.M.; Newstead, S. Cryo-EM Structure and Resistance Landscape of M. Tuberculosis MmpL3: An Emergent Therapeutic Target. Structure 2021, 29. [Google Scholar] [CrossRef]
- Li, W.; Obregón-Henao, A.; Wallach, J.B.; North, E.J.; Lee, R.E.; Gonzalez-Juarrero, M.; Schnappinger, D.; Jackson, M. Therapeutic Potential of the Mycobacterium Tuberculosis Mycolic Acid Transporter, MmpL3. Antimicrob Agents Chemother 2016, 60. [Google Scholar] [CrossRef]
- Chim, N.; Torres, R.; Liu, Y.; Capri, J.; Batot, G.; Whitelegge, J.P.; Goulding, C.W. The Structure and Interactions of Periplasmic Domains of Crucial MmpL Membrane Proteins from Mycobacterium Tuberculosis. Chem Biol 2015, 22. [Google Scholar] [CrossRef]
- Bode, N.J.; Heran Darwin, K. The Pup-Proteasome System of Mycobacteria. Microbiology spectrum 2014, 2. [Google Scholar] [CrossRef]
- Samanovic, M.I.; Li, H.; Darwin, K.H. The Pup-Proteasome System of Mycobacterium Tuberculosis. Subcell Biochem 2013. [Google Scholar] [CrossRef]
- Becker, S.H.; Jastrab, J.B.; Dhabaria, A.; Chaton, C.T.; Rush, J.S.; Korotkov, K. V.; Ueberheide, B.; Heran Darwin, K. The Mycobacterium Tuberculosis Pup-Proteasome System Regulates Nitrate Metabolism through an Essential Protein Quality Control Pathway. Proc Natl Acad Sci U S A 2019, 116. [Google Scholar] [CrossRef] [PubMed]
- Barandun, J.; Delley, C.L.; Weber-Ban, E. The Pupylation Pathway and Its Role in Mycobacteria. BMC Biol 2012. [Google Scholar] [CrossRef] [PubMed]
- Striebel, F.; Imkamp, F.; Özcelik, D.; Weber-Ban, E. Pupylation as a Signal for Proteasomal Degradation in Bacteria. Biochim Biophys Acta Mol Cell Res 2014, 1843. [Google Scholar] [CrossRef]
- Janssen, G. V.; Zhang, S.; Merkx, R.; Schiesswohl, C.; Chatterjee, C.; Darwin, K.H.; Geurink, P.P.; van der Heden van Noort, G.J.; Ovaa, H. Development of Tyrphostin Analogues to Study Inhibition of the Mycobacterium Tuberculosis Pup Proteasome System**. ChemBioChem 2021, 22. [Google Scholar] [CrossRef]
- Mir, M.A.; Rajeswari, H.S.; Veeraraghavan, U.; Ajitkumar, P. Molecular Characterisation of ABC Transporter Type FtsE and FtsX Proteins of Mycobacterium Tuberculosis. Arch Microbiol 2006, 185, 147–158. [Google Scholar] [CrossRef]
- Mir, M.A.; Arumugam, M.; Mondal, S.; Rajeswari, H.S.; Ramakumar, S.; Ajitkumar, P. Mycobacterium Tuberculosis Cell Division Protein, FtsE, Is an ATPase in Dimeric Form. Protein Journal 2015, 34, 35–47. [Google Scholar] [CrossRef]
- White, E.L.; Suling, W.J.; Ross, L.J.; Seitz, L.E.; Reynolds, R.C. 2-Alkoxycarbonylaminopyridines: Inhibitors of Mycobacterium Tuberculosis FtsZ. Journal of Antimicrobial Chemotherapy 2002, 50. [Google Scholar] [CrossRef]
- Roy, S.; Vijay, S.; Arumugam, M.; Anand, D.; Mir, M.; Ajitkumar, P. Mycobacterium Tuberculosis Expresses FtsE Gene through Multiple Transcripts. Curr Microbiol 2011. [Google Scholar] [CrossRef]
- Mavrici, D.; Marakalala, M.J.; Holton, J.M.; Prigozhin, D.M.; Gee, C.L.; Zhang, Y.J.; Rubin, E.J.; Alber, T. Mycobacterium Tuberculosis FtsX Extracellular Domain Activates the Peptidoglycan Hydrolase, RipC. Proc Natl Acad Sci U S A 2014, 111, 8037–8042. [Google Scholar] [CrossRef] [PubMed]
- Mavrici, D.; Marakalala, M.J.; Holton, J.M.; Prigozhin, D.M.; Gee, C.L.; Zhang, Y.J.; Rubin, E.J.; Alber, T. Mycobacterium Tuberculosis FtsX Extracellular Domain Activates the Peptidoglycan Hydrolase, RipC. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 8037–8042. [Google Scholar] [CrossRef] [PubMed]
- Plocinska, R.; Martinez, L.; Gorla, P.; Pandeeti, E.; Sarva, K.; Blaszczyk, E.; Dziadek, J.; Madiraju, M. V.; Rajagopalan, M. Mycobacterium Tuberculosis MtrB Sensor Kinase Interactions with FtsI and Wag31 Proteins Reveal a Role for MtrB Distinct from That Regulating MtrA Activities. J Bacteriol 2014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Y.; Zhang, Y.; Qiao, L.; Chi, X.; Han, Y.; Lin, Y.; Si, S.; Jiang, J. Identification of Anti-Mycobacterium Tuberculosis Agents Targeting the Interaction of Bacterial Division Proteins FtsZ and SepFe. Acta Pharm Sin B 2023, 13. [Google Scholar] [CrossRef]
- Paritala, H.; Carroll, K.S. New Targets and Inhibitors of Mycobacterial Sulfur Metabolism. Infectious disorders drug targets 2013, 13, 85–115. [Google Scholar] [CrossRef]
- Rhee, K.; de Carvalho, L.P.S.; Bryk, R.; Ehrt, S.; Marrero, J.; Park, S.W.; Schnappinger, D.; Venugopal, A.; Nathan, C. Central Carbon Metabolism in Mycobacterium Tuberculosis: An Unexpected Frontier. Trends Microbiol 2011, 19 7, 307–314. [Google Scholar] [CrossRef]
- Ehrt, S.; Rhee, K. Mycobacterium Tuberculosis Metabolism and Host Interaction: Mysteries and Paradoxes. Curr Top Microbiol Immunol 2013, 374, 163–188. [Google Scholar] [CrossRef]
- Rhee, Kyu Y. ; Carvalho, L.; Bryk, Ruslana; Ehrt, Sabine; Nathan, C. Central Carbon Metabolism in Mycobacterium Tuberculosis: An Unexpected Frontiers. Trends Microbiol 2011, 23, 1–12. [Google Scholar]
- Morita, Y.S.; Fukuda, T.; Sena, C.B.C.; Yamaryo-Botte, Y.; McConville, M.J.; Kinoshita, T. Inositol Lipid Metabolism in Mycobacteria: Biosynthesis and Regulatory Mechanisms. Biochim Biophys Acta Gen Subj 2011, 1810, 630–641. [Google Scholar] [CrossRef]
- Rodríguez, D.C.; Ocampo, M.; Varela, Y.; Curtidor, H.; Patarroyo, M.A.; Patarroyo, M.E. Mce4F Mycobacterium Tuberculosis Protein Peptides Can Inhibit Invasion of Human Cell Lines. Pathog Dis 2015, 73, 1–12. [Google Scholar] [CrossRef]
- Fieweger, R.A.; Wilburn, K.M.; Montague, C.R.; Roszkowski, E.K.; Kelly, C.M.; Southard, T.L.; Sondermann, H.; Nazarova, E. V.; VanderVen, B.C. MceG Stabilizes the Mce1 and Mce4 Transporters in Mycobacterium Tuberculosis. Journal of Biological Chemistry 2023, 299. [Google Scholar] [CrossRef]
- Pawełczyk, J.; Brzostek, A.; Minias, A.; Płociński, P.; Rumijowska-Galewicz, A.; Strapagiel, D.; Zakrzewska-Czerwińska, J.; Dziadek, J. Cholesterol-Dependent Transcriptome Remodeling Reveals New Insight into the Contribution of Cholesterol to Mycobacterium Tuberculosis Pathogenesis. Sci Rep 2021, 11. [Google Scholar] [CrossRef]
- Larrouy-Maumus, G. Cholesterol Acquisition by Mycobacterium Tuberculosis. Virulence 2015, 6, 412–413. [Google Scholar] [CrossRef]
- Miner, M.D.; Chang, J.C.; Pandey, A.K.; Sassetti, C.M.; Sherman, D.R. Role of Cholesterol in Mycobacterium Tuberculosis Infection. Indian J Exp Biol 2009, 47, 407–411. [Google Scholar]
- Pandey, A.K.; Sassetti, C.M. Mycobacterial Persistence Requires the Utilization of Host Cholesterol. Proc Natl Acad Sci U S A 2008, 105, 4376–4380. [Google Scholar] [CrossRef]
- Nazarova, E.; Montague, C.R.; La, T.; Wilburn, K.M.; Sukumar, N.; Lee, W.; Caldwell, S.; Russell, D.; VanderVen, B. Rv3723/LucA Coordinates Fatty Acid and Cholesterol Uptake in Mycobacterium Tuberculosis. Elife 2017, 6, null. [Google Scholar] [CrossRef]
- Wilburn, K.M.; Fieweger, R.; VanderVen, B. Cholesterol and Fatty Acids Grease the Wheels of Mycobacterium Tuberculosis Pathogenesis. Pathog Dis 2018, 76 2, null. [Google Scholar] [CrossRef]
- Szulc-Kielbik, I.; Kielbik, M.; Przygodzka, P.; Brzostek, A.; Dziadek, J.; Klink, M. Mycobacterium Tuberculosis Requires Cholesterol Oxidase to Disrupt TLR2 Signalling in Human Macrophages. Mediators Inflamm 2019, 2019. [Google Scholar] [CrossRef]
- Crowe, A.M.; Workman, S.D.; Watanabe, N.; Worrall, L.J.; Strynadka, N.C.J.; Eltis, L.D. IpdAB, a Virulence Factor in Mycobacterium Tuberculosis, Is a Cholesterol Ring-Cleaving Hydrolase. Proc Natl Acad Sci U S A 2018, 115. [Google Scholar] [CrossRef]
- Marques, M.A.M.; Berr??do-Pinho, M.; Rosa, T.L.S.A.; Pujari, V.; Lemes, R.M.R.; Lery, L.M.S.; Silva, C.A.M.; Guimar??es, A.C.R.; Atella, G.C.; Wheat, W.H.; et al. The Essential Role of Cholesterol Metabolism in the Intracellular Survival of Mycobacterium Leprae Is Not Coupled to Central Carbon Metabolism and Energy Production. J Bacteriol 2015, 197, 3698–3707. [Google Scholar] [CrossRef]
- Pethe, K.; Sequeira, P.C.; Agarwalla, S.; Rhee, K.; Kuhen, K.; Phong, W.Y.; Patel, V.; Beer, D.; Walker, J.R.; Duraiswamy, J.; et al. A Chemical Genetic Screen in Mycobacterium Tuberculosis Identifies Carbon-Source-Dependent Growth Inhibitors Devoid of in Vivo Efficacy. Nat Commun 2010. [Google Scholar] [CrossRef]
- Xu, Y.; Pooja; Borah, K. Mycobacterium Tuberculosis Carbon and Nitrogen Metabolic Fluxes. Biosci Rep 2022, 42, null. [Google Scholar] [CrossRef]
- Shi, L.; Sohaskey, C.D.; Pfeiffer, C.; Datta, P.; Parks, M.; McFadden, J.; North, R.J.; Gennaro, M.L. Carbon Flux Rerouting during Mycobacterium Tuberculosis Growth Arrest. Mol Microbiol 2010, 78. [Google Scholar] [CrossRef]
- Baughn, A.D.; Rhee, K.Y. Metabolomics of Central Carbon Metabolism in Mycobacterium Tuberculosis. Microbiol Spectr 2014, 2. [Google Scholar] [CrossRef]
- Gouzy, A.; Poquet, Y.; Neyrolles, O. Amino Acid Capture and Utilization within the Mycobacterium Tuberculosis Phagosome. Future Microbiol 2014, 9 5, 631–637. [Google Scholar] [CrossRef]
- Gouzy, A.; Poquet, Y.; Neyrolles, O. Nitrogen Metabolism in Mycobacterium Tuberculosis Physiology and Virulence. Nat Rev Microbiol 2014, 12, 729–737. [Google Scholar] [CrossRef]
- Gouzy, A.; Larrouy-Maumus, G.; Wu, T. Di; Peixoto, A.; Levillain, F.; Lugo-Villarino, G.; Gerquin-Kern, J.L.; De Carvalho, L.P.S.; Poquet, Y.; Neyrolles, O. Mycobacterium Tuberculosis Nitrogen Assimilation and Host Colonization Require Aspartate. Nat Chem Biol 2013, 9. [Google Scholar] [CrossRef]
- Williams, K.J.; Jenkins, V.A.; Barton, G.R.; Bryant, W.A.; Krishnan, N.; Robertson, B.D. Deciphering the Metabolic Response of Mycobacterium Tuberculosis to Nitrogen Stress. Mol Microbiol 2015, 97. [Google Scholar] [CrossRef]
- Xu, Y.; Ma, S.; Huang, Z.; Wang, L.; Raza, S.H.A.; Wang, Z. Nitrogen Metabolism in Mycobacteria: The Key Genes and Targeted Antimicrobials. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Larsen, R.; Kloosterman, T.G.; Kok, J.; Kuipers, O.P. GlnR-Mediated Regulation of Nitrogen Metabolism in Lactococcus Lactis. J Bacteriol 2006. [Google Scholar] [CrossRef]
- He, J.; Kang, X.; Wu, J.; Shao, Z.; Zhang, Z.; Wu, Y.; Yuan, H.; Zhao, G.; Wang, J. Transcriptional Self-Regulation of the Master Nitrogen Regulator GlnR in Mycobacteria. J Bacteriol 2023, 205. [Google Scholar] [CrossRef]
- Borah, K.; Beyß, M.; Theorell, A.; Wu, H.; Basu, P.; Mendum, T.A.; Nӧh, K.; Beste, D.J.V.; McFadden, J. Intracellular Mycobacterium Tuberculosis Exploits Multiple Host Nitrogen Sources during Growth in Human Macrophages. Cell Rep 2019, 29. [Google Scholar] [CrossRef]
- Sharma, R.; Hartman, T.E.; Beites, T.; Kim, J.-H.; Eoh, H.; Engelhart, C.A.; Zhu, L.; Wilson, D.J.; Aldrich, C.C.; Ehrt, S.; et al. Metabolically Distinct Roles of NAD Synthetase and NAD Kinase Define the Essentiality of NAD and NADP in Mycobacterium Tuberculosis. mBio 2023. [Google Scholar] [CrossRef]
- Mori, S.; Yamasaki, M.; Maruyama, Y.; Momma, K.; Kawai, S.; Hashimoto, W.; Mikami, B.; Murata, K. NAD-Binding Mode and the Significance of Intersubunit Contact Revealed by the Crystal Structure of Mycobacterium Tuberculosis NAD Kinase-NAD Complex. Biochem Biophys Res Commun 2005, 327. [Google Scholar] [CrossRef]
- Neyrolles, O.; Wolschendorf, F.; Mitra, A.; Niederweis, M. Mycobacteria, Metals, and the Macrophage. Immunol Rev 2015, 264, 249–263. [Google Scholar] [CrossRef]
- Reddy, P.V.; Puri, R.V.; Chauhan, P.; Kar, R.; Rohilla, A.; Khera, A.; Tyagi, A.K. Disruption of Mycobactin Biosynthesis Leads to Attenuation of Mycobacterium Tuberculosis for Growth and Virulence. Journal of Infectious Diseases 2013. [Google Scholar] [CrossRef]
- Liang, Y.; Rubinstein, J.L. Structural Analysis of Mycobacterial Electron Transport Chain Complexes by CryoEM. Biochem Soc Trans 2023, 51, 183–193. [Google Scholar]
- Anand, P.; Akhter, Y. A Review on Enzyme Complexes of Electron Transport Chain from Mycobacterium Tuberculosis as Promising Drug Targets. Int J Biol Macromol 2022, 212, 474–494. [Google Scholar] [CrossRef]
- Zeng, S.; Zhang, J.; Sun, M.; Zhang, X.; Cook, G.M.; Zhang, T. Nitric Oxide-Dependent Electron Transport Chain Inhibition by the Cytochrome Bc1inhibitor and Pretomanid Combination Kills Mycobacterium Tuberculosis. Antimicrob Agents Chemother 2021, 65. [Google Scholar] [CrossRef]
- Mathiyazakan, V.; Wong, C.-F.; Harikishore, A.; Pethe, K.; Grüber, G. Cryo-Electron Microscopy Structure of the Mycobacterium Tuberculosi s Cytochrome Bcc : Aa 3 Supercomplex and a Novel Inhibitor Targeting Subunit Cytochrome c I. Antimicrob Agents Chemother 2023, 67. [Google Scholar] [CrossRef]
- Ofori-Anyinam, B.; Riley, A.J.; Jobarteh, T.; Gitteh, E.; Sarr, B.; Faal-Jawara, T.I.; Rigouts, L.; Senghore, M.; Kehinde, A.; Onyejepu, N.; et al. Comparative Genomics Shows Differences in the Electron Transport and Carbon Metabolic Pathways of Mycobacterium Africanum Relative to Mycobacterium Tuberculosis and Suggests an Adaptation to Low Oxygen Tension. Tuberculosis 2020, 120. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.M.; Denny, W.A. Inhibitors of Enzymes in the Electron Transport Chain of Mycobacterium Tuberculosis. In Annual Reports in Medicinal Chemistry 2019, 52, 97–130. [Google Scholar] [CrossRef]
- Sousa, E.H.S.; Tuckerman, J.R.; Gonzalez, G.; Gilles-Gonzalez, M.-A. DosT and DevS Are Oxygen-Switched Kinases in Mycobacterium Tuberculosis. Protein Science 2007, 16. [Google Scholar] [CrossRef]
- Slotboom, D.J.; Ettema, T.W.; Nijland, M.; Thangaratnarajah, C. Bacterial Multi-Solute Transporters. FEBS Lett 2020, 1–10. [Google Scholar] [CrossRef]
- Putman, M.; van Veen, H.W.; Konings, W.N. Molecular Properties of Bacterial Multidrug Transporters. Microbiol Mol Biol Rev 2000. [Google Scholar] [CrossRef]
- Khan, M.T.; Khan, T.A.; Ahmad, I.; Muhammad, S.; Wei, D.Q. Diversity and Novel Mutations in Membrane Transporters of Mycobacterium Tuberculosis. Brief Funct Genomics 2023, 22, 168–179. [Google Scholar] [CrossRef]
- De Rossi, E.; Arrigo, P.; Bellinzoni, M.; Silva, P.A.E.; Martín, C.; Aínsa, J.A.; Guglierame, P.; Riccardi, G. The Multidrug Transporters Belonging to Major Facilitator Superfamily in Mycobacterium Tuberculosis. Mol Med 2002. [Google Scholar] [CrossRef]
- Rossi, E.D.; Aínsa, J.A.; Riccardi, G. Role of Mycobacterial Efflux Transporters in Drug Resistance: An Unresolved Question. FEMS Microbiol Rev 2006, 30, 36–52. [Google Scholar] [CrossRef]
- De Rossi, E.; Arrigo, P.; Bellinzoni, M.; Silva, P.E.A.; Martín, C.; Aínsa, J.A.; Guglierame, P.; Riccardi, G. The Multidrug Transporters Belonging to Major Facilitator Superfamily (MFS) in Mycobacterium Tuberculosis. Molecular Medicine 2002, 8. [Google Scholar] [CrossRef]
- Fenn, J.S.; Nepravishta, R.; Guy, C.S.; Harrison, J.; Angulo, J.; Cameron, A.D.; Fullam, E. Structural Basis of Glycerophosphodiester Recognition by the Mycobacterium Tuberculosis Substrate-Binding Protein UgpB. ACS Chem Biol 2019, 14, 1879–1887. [Google Scholar] [CrossRef]
- Jiang, D.; Zhang, Q.; Zheng, Q.; Zhou, H.; Jin, J.; Zhou, W.; Bartlam, M.; Rao, Z. Structural Analysis of Mycobacterium Tuberculosis ATP-Binding Cassette Transporter Subunit UgpB Reveals Specificity for Glycerophosphocholine. FEBS Journal 2014, 281, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhang, Q.; Zheng, Q.; Zhou, H.; Jin, J.; Zhou, W.; Bartlam, M.; Rao, Z. Structural Analysis of Mycobacterium Tuberculosis ATP-Binding Cassette Transporter Subunit UgpB Reveals Specificity for Glycerophosphocholine. FEBS Journal 2014. [Google Scholar] [CrossRef] [PubMed]
- Boritsch, E.C.; Brosch, R. Evolution of Mycobacterium Tuberculosis: New Insights into Pathogenicity and Drug Resistance. Microbiol Spectr 2016, 4 5, null. [Google Scholar] [CrossRef]
- Rostamian, M.; Kooti, S.; Abiri, R.; Khazayel, S.; Kadivarian, S.; Borji, S.; Alvandi, A. Prevalence of Mycobacterium Tuberculosis Mutations Associated with Isoniazid and Rifampicin Resistance: A Systematic Review and Meta-Analysis. J Clin Tuberc Other Mycobact Dis 2023, 32. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Dwivedi, S.P.; Gaharwar, U.S.; Meena, R.; Rajamani, P.; Prasad, T. Recent Updates on Drug Resistance in Mycobacterium Tuberculosis. J Appl Microbiol 2020, 128, 1547–1567. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, B.S.; Bhakta, S.; Barik, R.; Basu, J.; Kundu, M.; Chakrabarti, P. Overexpression and Functional Characterization of an ABC (ATP-Binding Cassette) Transporter Encoded by the Genes DrrA and DrrB of Mycobacterium Tuberculosis. Biochem J 2002. [Google Scholar] [CrossRef]
- Khosravi, A.D.; Sirous, M.; Absalan, Z.; Tabandeh, M.R.; Savari, M. Comparison of DrrA and DrrB Efflux Pump Genes Expression in Drug-Susceptible and -Resistant Mycobacterium Tuberculosis Strains Isolated from Tuberculosis Patients in Iran. Infect Drug Resist 2019, 12, 3437–3444. [Google Scholar] [CrossRef]
- Koch, A.; Mizrahi, V.; Warner, D.F. The Impact of Drug Resistance on Mycobacterium Tuberculosis Physiology: What Can We Learn from Rifampicin? 2014, 17. [CrossRef]
- Drobniewski, F.; Balabanova, Y.; Ruddy, M.; Weldon, L.; Jeltkova, K.; Brown, T.; Malomanova, N.; Elizarova, E.; Melentyey, A.; Mutovkin, E.; et al. Rifampin- and Multidrug-Resistant Tuberculosis in Russian Civilians and Prison Inmates: Dominance of the Beijing Strain Family. Emerg Infect Dis 2002, 8, 1320–1326. [Google Scholar] [CrossRef]
- Du, D.; van Veen, H.W.; Murakami, S.; Pos, K.M.; Luisi, B.F. Structure, Mechanism and Cooperation of Bacterial Multidrug Transporters. Curr Opin Struct Biol 2015, 33, 76–91. [Google Scholar] [CrossRef]
- Singh, G.; Akhter, Y. Molecular Insights into the Differential Efflux Mechanism of Rv1634 Protein, a Multidrug Transporter of Major Facilitator Superfamily in Mycobacterium Tuberculosis. Proteins: Structure, Function and Bioinformatics 2022, 90. [Google Scholar] [CrossRef]
- Dutta, N.K.; Mehra, S.; Kaushal, D. A Mycobacterium Tuberculosis Sigma Factor Network Responds to Cell-Envelope Damage by the Promising Anti-Mycobacterial Thioridazine. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Dutta, N.K.; Mehra, S.; Didier, P.J.; Roy, C.J.; Doyle, L.A.; Alvarez, X.; Ratterree, M.; Be, N.A.; Lamichhane, G.; Jain, S.K.; et al. Genetic Requirements for the Survival of Tubercle Bacilli in Primates. Journal of Infectious Diseases 2010, 201, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, P.; Akhter, Y. The Drug Binding Sites and Transport Mechanism of the RND Pumps from Mycobacterium Tuberculosis: Insights from Molecular Dynamics Simulations. Arch Biochem Biophys 2016, 592. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.M.; Babii, S.O.; Pandya, A.N.; Li, W.; Li, Y.; Mehla, J.; Scott, R.; Hegde, P.; Prathipati, P.K.; Acharya, A.; et al. Proton Transfer Activity of the Reconstituted Mycobacterium Tuberculosis MmpL3 Is Modulated by Substrate Mimics and Inhibitors. Proc Natl Acad Sci U S A 2022, 119. [Google Scholar] [CrossRef] [PubMed]
- Thouvenel, L.; Rech, J.; Guilhot, C.; Bouet, J.Y.; Chalut, C. In Vivo Imaging of MmpL Transporters Reveals Distinct Subcellular Locations for Export of Mycolic Acids and Non-Essential Trehalose Polyphleates in the Mycobacterial Outer Membrane. Sci Rep 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Duan, H.; Yan, H.; Liu, X.; Peng, L.; Hu, Y.; Liu, W.; Liang, L.; Shi, H.; Zhao, G.; et al. Specifically Targeting Mtb Cell-Wall and TMM Transporter: The Development of MmpL3 Inhibitors. Curr Protein Pept Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.C.; Hsu, F.F.; Arnett, E.; Dunaj, J.L.; Davidson, P.M.; Pacheco, S.A.; Harriff, M.J.; Lewinsohn, D.M.; Schlesinger, L.S.; Purdy, G.E. The Mycobacterium Tuberculosis MmpL11 Cell Wall Lipid Transporter Is Important for Biofilm Formation, Intracellular Growth, and Nonreplicating Persistence. Infect Immun 2017, 85. [Google Scholar] [CrossRef]
- Famelis, N.; Geibel, S.; Van Tol, D. Mycobacterial Type VII Secretion Systems. Biol Chem 2023, 404, 691–702. [Google Scholar] [CrossRef]
- Abdallah, AM.; Pittius, G.; Nicolaas, C.; DiGiuseppe Champion, P.; Cox, J.; Luirink, J.; Vandenbroucke-Grauls, CMJE. ; Appelmelk, BJ.; Bitter, W. Type VII Secretion - Mycobacteria Show the Way. Nat Rev Microbiol 2007, 5, 883–891. [Google Scholar] [CrossRef]
- Abdallah, A.M.; Gey van Pittius, N.C.; Champion, P.A.D.; Cox, J.; Luirink, J.; Vandenbroucke-Grauls, C.M.J.E.; Appelmelk, B.J.; Bitter, W. Type VII Secretion--Mycobacteria Show the Way. Nat Rev Microbiol 2007. [Google Scholar] [CrossRef]
- Rivera-Calzada, A.; Famelis, N.; Llorca, O.; Geibel, S. Type VII Secretion Systems: Structure, Functions and Transport Models. Nat Rev Microbiol 2021, 19, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Zahra, A.; Thompson, V.; Sirisaengtaksin, N.; Wells, A.; Porto, M.; Köster, S.; Penberthy, K.; Kubota, Y.; Dricot, A.; et al. Mycobacterium Tuberculosis Type VII Secreted Effector EsxH Targets Host ESCRT to Impair Trafficking. PLoS Pathog 2013. [Google Scholar] [CrossRef] [PubMed]
- Crosskey, T.D.; Beckham, K.S.H.; Wilmanns, M. The ATPases of the Mycobacterial Type VII Secretion System: Structural and Mechanistic Insights into Secretion. Prog Biophys Mol Biol 2020, 152, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, Y.; Lin, C.; Zhang, J.; Mai, J.; Jiang, J.; Gao, X.; Li, Y.; Zhao, G.; Zhang, L.; et al. Crosstalk between the Ancestral Type VII Secretion System ESX-4 and Other T7SS in Mycobacterium Marinum. iScience 2022, 25. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, N.; Padmanaban, S.; Dutta, S. Cryo-EM Reveals the Membrane-Binding Phenomenon of EspB, a Virulence Factor of the Mycobacterial Type VII Secretion System. Journal of Biological Chemistry 2023, 299. [Google Scholar] [CrossRef]
- Gijsbers, A.; Eymery, M.; Gao, Y.; Menart, I.; Vinciauskaite, V.; Siliqi, D.; Peters, P.J.; McCarthy, A.; Ravelli, R.B.G. The Crystal Structure of the EspB-EspK Virulence Factor-Chaperone Complex Suggests an Additional Type VII Secretion Mechanism in Mycobacterium Tuberculosis. Journal of Biological Chemistry 2023, 299. [Google Scholar] [CrossRef]
- Rengarajan, J.; Bloom, B.R.; Rubin, E.J. Genome-Wide Requirements for Mycobacterium Tuberculosis Adaptation and Survival in Macrophages. Proc Natl Acad Sci U S A 2005, 102, 8327–8332. [Google Scholar] [CrossRef]
- Turkarslan, S.; Peterson, E.J.R.; Rustad, T.R.; Minch, K.J.; Reiss, D.J.; Morrison, R.; Ma, S.; Price, N.D.; Sherman, D.R.; Baliga, N.S. A Comprehensive Map of Genome-Wide Gene Regulation in Mycobacterium Tuberculosis. Sci Data 2015. [Google Scholar] [CrossRef]
- Namouchi, A.; Didelot, X.; Schöck, U.; Gicquel, B.; Rocha, E.P.C. After the Bottleneck: Genome-Wide Diversification of the Mycobacterium Tuberculosis Complex by Mutation, Recombination, and Natural Selection. Genome Res 2012. [Google Scholar] [CrossRef]
- Sobkowiak, B.; Romanowski, K.; Sekirov, I.; Gardy, J.L.; Johnston, J.C. Comparing Mycobacterium Tuberculosis Transmission Reconstruction Models from Whole Genome Sequence Data. Epidemiol Infect 2023. [Google Scholar] [CrossRef]
- Cole, S. T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S. V.; Eiglmeier, K.; Gas, S.; Barry, C. E.; et al. Deciphering the Biology of Mycobacterium Tuberculosis from the Complete Genome Sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Shabbeer, A.; Cowan, L.; Ozcaglar, C.; Rastogi, N.; Vandenberg, S.; Yener, B.; Bennett, K.P. TB-Lineage: An Online Tool for Classification and Analysis of Strains of Mycobacterium Tuberculosis Complex. Infect Genet Evol 2012, 12 4, 789–797. [Google Scholar] [CrossRef]
- Padane, A.; Harouna Hamidou, Z.; Drancourt, M.; Saad, J. CRISPR-Based Detection, Identification and Typing of Mycobacterium Tuberculosis Complex Lineages. Microbiol Spectr 2023, 11. [Google Scholar] [CrossRef]
- He, C.; Cheng, X.; Kaisaier, A.; Wan, J.; Luo, S.; Ren, J.; Sha, Y.; Peng, H.; Zhen, Y.; Liu, W.; et al. Effects of Mycobacterium Tuberculosis Lineages and Regions of Difference (RD) Virulence Gene Variation on Tuberculosis Recurrence. Ann Transl Med 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Sahal, M.R.; Senelle, G.; La, K.; Molina-Moya, B.; Dominguez, J.; Panda, T.; Cambau, E.; Refregier, G.; Sola, C.; Guyeux, C. An Updated Evolutionary History and Taxonomy of Mycobacterium Tuberculosis Lineage 5, Also Called M. Africanum. bioRxiv 2022. [Google Scholar] [CrossRef]
- Krishnan, N.; Malaga, W.; Constant, P.; Caws, M.; Thi Hoang Chau, T.; Salmons, J.; Thi Ngoc Lan, N.; Bang, N.D.; Daffé, M.; Young, D.B.; et al. Mycobacterium Tuberculosis Lineage Influences Innate Immune Response and Virulence and Is Associated with Distinct Cell Envelope Lipid Profiles. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Mehrotra, J.; Bishai, W.R. Regulation of Virulence Genes in Mycobacterium Tuberculosis. Int J Med Microbiol 2001, 291. [Google Scholar] [CrossRef]
- Rindi, L.; Medici, C.; Bimbi, N.; Buzzigoli, A.; Lari, N.; Garzelli, C. Genomic Variability of Mycobacterium Tuberculosis Strains of the Euro-American Lineage Based on Large Sequence Deletions and 15-Locus MIRU-VNTR Polymorphism. PLoS One 2014, 9, null. [Google Scholar] [CrossRef]
- Malinga, L.A.; Abeel, T.; Desjardins, C.A.; Dlamini, T.C.; Cassell, G.; Chapman, S.B.; Birren, B.W.; Earl, A.M.; van der Walt, M. Draft Genome Sequences of Two Extensively Drug-Resistant Strains of Mycobacterium Tuberculosis Belonging to the Euro-American S Lineage. Genome Announc 2016, 4, 2–3. [Google Scholar] [CrossRef]
- De Leon, J.; Jiang, G.; Ma, Y.; Rubin, E.; Fortune, S.; Sun, J. Mycobacterium Tuberculosis ESAT-6 Exhibits a Unique Membrane-Interacting Activity That Is Not Found in Its Ortholog from Non-Pathogenic Mycobacterium Smegmatis. Journal of Biological Chemistry 2012, 287. [Google Scholar] [CrossRef]
- Gey Van Pittius, N.C.; Sampson, S.L.; Lee, H.; Kim, Y.; Van Helden, P.D.; Warren, R.M. Evolution and Expansion of the Mycobacterium Tuberculosis PE and PPE Multigene Families and Their Association with the Duplication of the ESAT-6 (Esx) Gene Cluster Regions. BMC Evol Biol 2006, 6. [Google Scholar] [CrossRef] [PubMed]
- García-Pelayo, M.; Caimi, K.; Inwald, J.; Hinds, J.; Bigi, F.; Romano, M.; van Soolingen, D.; Hewinson, R.G.; Cataldi, A.; Gordon, S. Microarray Analysis of Mycobacterium Microti Reveals Deletion of Genes Encoding PE-PPE Proteins and ESAT-6 Family Antigens. Tuberculosis, 2004; 84, 159–166. [Google Scholar] [CrossRef]
- Arnvig, K.B.; Young, D.B. Non-Coding RNA and Its Potential Role in Mycobacterium Tuberculosis Pathogenesis. RNA Biol 2012, 9, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Arnvig, K.B.; Comas, I.; Thomson, N.R.; Houghton, J.; Boshoff, H.I.; Croucher, N.J.; Rose, G.; Perkins, T.T.; Parkhill, J.; Dougan, G.; et al. Sequence-Based Analysis Uncovers an Abundance of Non-Coding RNA in the Total Transcriptome of Mycobacterium Tuberculosis. PLoS Pathog 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Mujwar, S.; Pardasani, K.R. Prediction of Riboswitch as a Potential Drug Target and Design of Its Optimal Inhibitors for Mycobacterium Tuberculosis. Int J Comput Biol Drug Des 2015, 8. [Google Scholar] [CrossRef]
- Bahoua, B.; Sevdalis, S.E.; Soto, A.M. Effect of Sequence on the Interactions of Divalent Cations with M-Box Riboswitches from Mycobacterium Tuberculosis and Bacillus Subtilis. Biochemistry 2021, 60. [Google Scholar] [CrossRef]
- Sivaramakrishnan, S.; De Montellano, P.R.O. The DosS-DosT/DosR Mycobacterial Sensor System. Biosensors (Basel) 2013, 3, 259–282. [Google Scholar] [CrossRef]
- Honaker, R.W.; Leistikow, R.L.; Bartek, I.L.; Voskui, M.I. Unique Roles of DosT and DosS in DosR Regulon Induction and Mycobacterium Tuberculosis Dormancy. Infect Immun 2009, 77. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, K.J.; Ko, I.J.; Kim, Y.M.; Oh, J. Il Different Roles of DosS and DosT in the Hypoxic Adaptation of Mycobacteria. J Bacteriol 2010, 192. [Google Scholar] [CrossRef]
- Kumar, A.; Toledo, J.C.; Patel, R.P.; Lancaster, J.R.; Steyn, A.J.C. Mycobacterium Tuberculosis DosS Is a Redox Sensor and DosT Is a Hypoxia Sensor. Proc Natl Acad Sci U S A 2007, 104. [Google Scholar] [CrossRef]
- Sevalkar, R.R.; Glasgow, J.N.; Pettinati, M.; Marti, M.A.; Reddy, V.P.; Basu, S.; Alipour, E.; Kim-Shapiro, D.B.; Estrin, D.A.; Lancaster, J.R.; et al. Mycobacterium Tuberculosis DosS Binds H2S through Its Fe3+ Heme Iron to Regulate the DosR Dormancy Regulon. Redox Biol 2022, 52. [Google Scholar] [CrossRef]
- Zheng, H.; Colvin, C.J.; Johnson, B.K.; Kirchhoff, P.D.; Wilson, M.; Jorgensen-Muga, K.; Larsen, S.D.; Abramovitch, R.B. Inhibitors of Mycobacterium Tuberculosis DosRST Signaling and Persistence. Nat Chem Biol 2017, 13. [Google Scholar] [CrossRef]
- Chadda, A.; Jensen, D.; Tomko, E.J.; Manzano, A.R.; Nguyen, B.; Lohman, T.M.; Galburt, E.A. Mycobacterium Tuberculosis DNA Repair Helicase UvrD1 Is Activated by Redox-Dependent Dimerization via a 2B Domain Cysteine. Proc Natl Acad Sci U S A 2022, 119. [Google Scholar] [CrossRef]
- Dos Vultos, T.; Mestre, O.; Tonjum, T.; Gicquel, B. DNA Repair in Mycobacterium Tuberculosis Revisited. In Proceedings of the FEMS Microbiology Reviews 2009, 33, 471–487. [Google Scholar] [CrossRef]
- O’Sullivan, D.M.; Hinds, J.; Butcher, P.D.; Gillespie, S.H.; McHugh, T.D. Mycobacterium Tuberculosis DNA Repair in Response to Subinhibitory Concentrations of Ciprofloxacin. Journal of Antimicrobial Chemotherapy 2008, 62. [Google Scholar] [CrossRef]
- Mizrahi, V.; Andersen, S.J. DNA Repair in Mycobacterium Tuberculosis. What Have We Learnt from the Genome Sequence? Mol Microbiol 1998, 29. [Google Scholar] [CrossRef]
- Abeldenov, S.; Saparbayev, M.; Khassenov, B. Biochemical Characterization of Mycobacterium Tuberculosis DNA Repair Enzymes – Nfo, XthA and Nei2. Cent Asian J Glob Health 2014, 2. [Google Scholar] [CrossRef]
- Miggiano, R.; Morrone, C.; Rossi, F.; Rizzi, M. Targeting Genome Integrity in Mycobacterium Tuberculosis: From Nucleotide Synthesis to DNA Replication and Repair. Molecules 2020, 25, 25. [Google Scholar] [CrossRef]
- Singh, A. Guardians of the Mycobacterial Genome: A Review on DNA Repair Systems in Mycobacterium Tuberculosis. Microbiology (United Kingdom) 2017, 163, 1740–1758. [Google Scholar] [CrossRef] [PubMed]
- Sharp, J.D.; Singh, A.K.; Park, S.T.; Lyubetskaya, A.; Peterson, M.W.; Gomes, A.L.C.; Potluri, L.P.; Raman, S.; Galagan, J.E.; Husson, R.N. Comprehensive Definition of the SigH Regulon of Mycobacterium Tuberculosis Reveals Transcriptional Control of Diverse Stress Responses. PLoS One 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.; Song, T.; Puyang, X.; Bardarov, S.; Jacobs, J.; Husson, R.N. The Alternative Sigma Factor Sigh Regulates Major Components of Oxidative and Heat Stress Responses in Mycobacterium Tuberculosis. J Bacteriol 2001, 183. [Google Scholar] [CrossRef] [PubMed]
- Cioetto-Mazzabò, L.; Boldrin, F.; Beauvineau, C.; Speth, M.; Marina, A.; Namouchi, A.; Segafreddo, G.; Cimino, M.; Favre-Rochex, S.; Balasingham, S.; et al. SigH Stress Response Mediates Killing of Mycobacterium Tuberculosis by Activating Nitronaphthofuran Prodrugs via Induction of Mrx2 Expression. Nucleic Acids Res 2023, 51. [Google Scholar] [CrossRef] [PubMed]
- Steyn, A.J.C.; Collins, D.M.; Hondalus, M.K.; Jacobs, W.R.; Pamela Kawakami, R.; Bloom, B.R. Mycobacterium Tuberculosis WhiB3 Interacts with RpoV to Affect Host Survival but Is Dispensable for in Vivo Growth. Proc Natl Acad Sci U S A 2002, 99. [Google Scholar] [CrossRef] [PubMed]
- Heimann, J.D. The Extracytoplasmic Function (ECF) Sigma Factors. Adv Microb Physiol 2002, 46, 47–110. [Google Scholar] [CrossRef]
- Manganelli, R.; Dubnau, E.; Tyagi, S.; Kramer, F.R.; Smith, I. Differential Expression of 10 Sigma Factor Genes in Mycobacterium Tuberculosis. Mol Microbiol 1999, 31. [Google Scholar] [CrossRef] [PubMed]
- Grosse-Siestrup, B.T.; Gupta, T.; Helms, S.; Tucker, S.L.; Voskuil, M.I.; Quinn, F.D.; Karls, R.K. A Role for Mycobacterium Tuberculosis Sigma Factor c in Copper Nutritional Immunity. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Chauhan, R.; Ravi, J.; Datta, P.; Chen, T.; Schnappinger, D.; Bassler, K.E.; Balázsi, G.; Gennaro, M.L. Reconstruction and Topological Characterization of the Sigma Factor Regulatory Network of Mycobacterium Tuberculosis. Nat Commun 2016, 7. [Google Scholar] [CrossRef]
- Mustyala, K.K.; Malkhed, V.; Chittireddy, V.R.; Vuruputuri, U. Virtual Screening Studies to Identify Novel Inhibitors for Sigma F Protein of Mycobacterium Tuberculosis. Int J Mycobacteriol 2015, 4. [Google Scholar] [CrossRef]
- Greenstein, A.E.; MacGurn, J.A.; Baer, C.E.; Falick, A.M.; Cox, J.S.; Alber, T. M. Tuberculosis Ser/Thr Protein Kinase D Phosphorylates an Anti-Anti-Sigma Factor Homolog. PLoS Pathog 2007, 3. [Google Scholar] [CrossRef]
- Baruzzo, G.; Serafini, A.; Finotello, F.; Sanavia, T.; Cioetto-Mazzabò, L.; Boldrin, F.; Lavezzo, E.; Barzon, L.; Toppo, S.; Provvedi, R.; et al. Role of the Extracytoplasmic Function Sigma Factor SigE in the Stringent Response of Mycobacterium Tuberculosis. Microbiol Spectr 2023, 11. [Google Scholar] [CrossRef]
- Burian, J.; Yim, G.; Hsing, M.; Axerio-Cilies, P.; Cherkasov, A.; Spiegelman, G.B.; Thompson, C.J. The Mycobacterial Antibiotic Resistance Determinant WhiB7 Acts as a Transcriptional Activator by Binding the Primary Sigma Factor SigA (RpoV). Nucleic Acids Res 2013, 41. [Google Scholar] [CrossRef]
- Singha, B.; Behera, D.; Khan, M.Z.; Singh, N.K.; Sowpati, D.T.; Gopal, B.; Nandicoori, V.K. The Unique N-Terminal Region of Mycobacterium Tuberculosis Sigma Factor A Plays a Dominant Role in the Essential Function of This Protein. Journal of Biological Chemistry 2023, 299. [Google Scholar] [CrossRef] [PubMed]
- Barik, S.; Sureka, K.; Mukherjee, P.; Basu, J.; Kundu, M. RseA, the SigE Specific Anti-Sigma Factor of Mycobacterium Tuberculosis, Is Inactivated by Phosphorylation-Dependent ClpC1P2 Proteolysis. Mol Microbiol 2010, 75. [Google Scholar] [CrossRef] [PubMed]
- Rahlwes, K.C.; Dias, B.R.S.; Campos, P.C.; Alvarez-Arguedas, S.; Shiloh, M.U. Pathogenicity and Virulence of Mycobacterium Tuberculosis. Virulence 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Forrellad, M.A.; Klepp, L.I.; Gioffré, A.; García, J.S.; Morbidoni, H.R.; de la Paz Santangelo, M.; Cataldi, A.A.; Bigi, F. Virulence Factors of the Mycobacterium Tuberculosis Complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef]
- Orgeur, M.; Brosch, R. Evolution of Virulence in the Mycobacterium Tuberculosis Complex. Curr Opin Microbiol 2018, 41, 68–75. [Google Scholar] [CrossRef]
- Ramon-Luing, L.A.; Palacios, Y.; Ruiz, A.; Téllez-Navarrete, N.A.; Chavez-Galan, L. Virulence Factors of Mycobacterium Tuberculosis as Modulators of Cell Death Mechanisms. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Mir, M.; Prisic, S.; Kang, C.-M.; Lun, S.; Guo, H.; Murry, J.P.; Rubin, E.J.; Husson, R.N. Mycobacterial Gene CuvA Is Required for Optimal Nutrient Utilization and Virulence. Infect Immun 2014, 82, 4104–4117. [Google Scholar] [CrossRef]
- Ghazaei, C. Mycobacterium Tuberculosis and Lipids: Insights into Molecular Mechanisms from Persistence to Virulence. Journal of Research in Medical Sciences 2018, 63, 23. [Google Scholar] [CrossRef]
- Modlin, S.J.; Elghraoui, A.; Gunasekaran, D.; Zlotnicki, A.M.; Dillon, N.A.; Dhillon, N.; Kuo, N.; Robinhold, C.; Chan, C.K.; Baughn, A.D.; et al. Structure-Aware Mycobacterium Tuberculosis Functional Annotation Uncloaks Resistance, Metabolic, and Virulence Genes. mSystems 2021, 6. [Google Scholar] [CrossRef]
- Aguilar-López, B.A.; Correa, F.; Moreno- Altamirano, M.M.B.; Espitia, C.; Hernández-Longoria, R.; Oliva-Ramírez, J.; Padierna-Olivos, J.; Sánchez-García, F.J. LprG and PE_PGRS33 Mycobacterium Tuberculosis Virulence Factors Induce Differential Mitochondrial Dynamics in Macrophages. Scand J Immunol 2019, 89. [Google Scholar] [CrossRef]
- Dong, W.; Nie, X.; Zhu, H.; Liu, Q.; Shi, K.-X.; You, L.; Zhang, Y.; Fan, H.; Yan, B.; Niu, C.; et al. Mycobacterial Fatty Acid Catabolism Is Repressed by FdmR to Sustain Lipogenesis and Virulence. Proceedings of the National Academy of Sciences 2021, 118, null. [Google Scholar] [CrossRef]
- Rieck, B.; Degiacomi, G.; Zimmermann, M.; Cascioferro, A.; Boldrin, F.; Lazar-Adler, N.R.; Bottrill, A.R.; le Chevalier, F.; Frigui, W.; Bellinzoni, M.; et al. PknG Senses Amino Acid Availability to Control Metabolism and Virulence of Mycobacterium Tuberculosis; 2017; Vol. 13; ISBN 1111111111.
- Parthasarathy, G.; Lun, S.; Guo, H.; Ammerman, N.C.; Geiman, D.E.; Bishai, W.R. Rv2190c, an NlpC/P60 Family Protein, Is Required for Full Virulence of Mycobacterium Tuberculosis. PLoS One 2012. [Google Scholar] [CrossRef]
- Velmurugan, K.; Chen, B.; Miller, J.L.; Azogue, S.; Gurses, S.; Hsu, T.; Glickman, M.; Jacobs, W.R.; Porcelli, S.A.; Briken, V. Mycobacterium Tuberculosis NuoG Is a Virulence Gene That Inhibits Apoptosis of Infected Host Cells. PLoS Pathog 2007, 3. [Google Scholar] [CrossRef]
- Rodriguez, G.M.; Smith, I. Identification of an ABC Transporter Required for Iron Acquisition and Virulence in Mycobacterium Tuberculosis. J Bacteriol 2006, 188, 424–430. [Google Scholar] [CrossRef]
- Beisiegel, M.; Mollenkopf, H.J.; Hahnke, K.; Koch, M.; Dietrich, I.; Reece, S.T.; Kaufmann, S.H.E. Combination of Host Susceptibility and Mycobacterium Tuberculosis Virulence Define Gene Expression Profile in the Host. Eur J Immunol 2009, 39. [Google Scholar] [CrossRef]
- Camacho, L.R.; Ensergueix, D.; Perez, E.; Gicquel, B.; Guilhot, C. Identification of a Virulence Gene Cluster of Mycobacterium Tuberculosis by Signature-Tagged Transposon Mutagenesis. Mol Microbiol 1999, 34, 257–267. [Google Scholar] [CrossRef]
- Brodin, P.; Majlessi, L.; Marsollier, L.; De Jonge, M.I.; Bottai, D.; Demangel, C.; Hinds, J.; Neyrolles, O.; Butcher, P.D.; Leclerc, C.; et al. Dissection of ESAT-6 System 1 of Mycobacterium Tuberculosis and Impact on Immunogenicity and Virulence. Infect Immun 2006, 74. [Google Scholar] [CrossRef]
- Walters, S.B.; Dubnau, E.; Kolesnikova, I.; Laval, F.; Daffe, M.; Smith, I. The Mycobacterium Tuberculosis PhoPR Two-Component System Regulates Genes Essential for Virulence and Complex Lipid Biosynthesis. Mol Microbiol 2006, 60. [Google Scholar] [CrossRef]
- Perkowski, E.F.; Miller, B.K.; Mccann, J.R.; Sullivan, J.T.; Malik, S.; Allen, I.C.; Godfrey, V.; Hayden, J.D.; Braunstein, M. An Orphaned Mce-Associated Membrane Protein of Mycobacterium Tuberculosis Is a Virulence Factor That Stabilizes Mce Transporters. Mol Microbiol 2016, 100, 90–107. [Google Scholar] [CrossRef]
- Garg, R.; Tripathi, D.; Kant, S.; Chandra, H.; Bhatnagar, R.; Banerjee, N. The Conserved Hypothetical Protein Rv0574c Is Required for Cell Wall Integrity, Stress Tolerance, and Virulence of Mycobacterium Tuberculosis. Infect Immun 2015. [Google Scholar] [CrossRef]
Figure 1.
Tuberculosis pathophysiology. The pathophysiology of tuberculosis includes phagocytosis of the bacillus by lung macrophages, followed by escape into the cell cytoplasm after preventing lysosomal fusion to the phagosome. Escape can induce cell necrosis, which induces an increase in cellular immune response that culminates in granuloma formation. The granuloma may contain the infection or function as a site for replication of the bacilli in the latent disease.
Figure 1.
Tuberculosis pathophysiology. The pathophysiology of tuberculosis includes phagocytosis of the bacillus by lung macrophages, followed by escape into the cell cytoplasm after preventing lysosomal fusion to the phagosome. Escape can induce cell necrosis, which induces an increase in cellular immune response that culminates in granuloma formation. The granuloma may contain the infection or function as a site for replication of the bacilli in the latent disease.
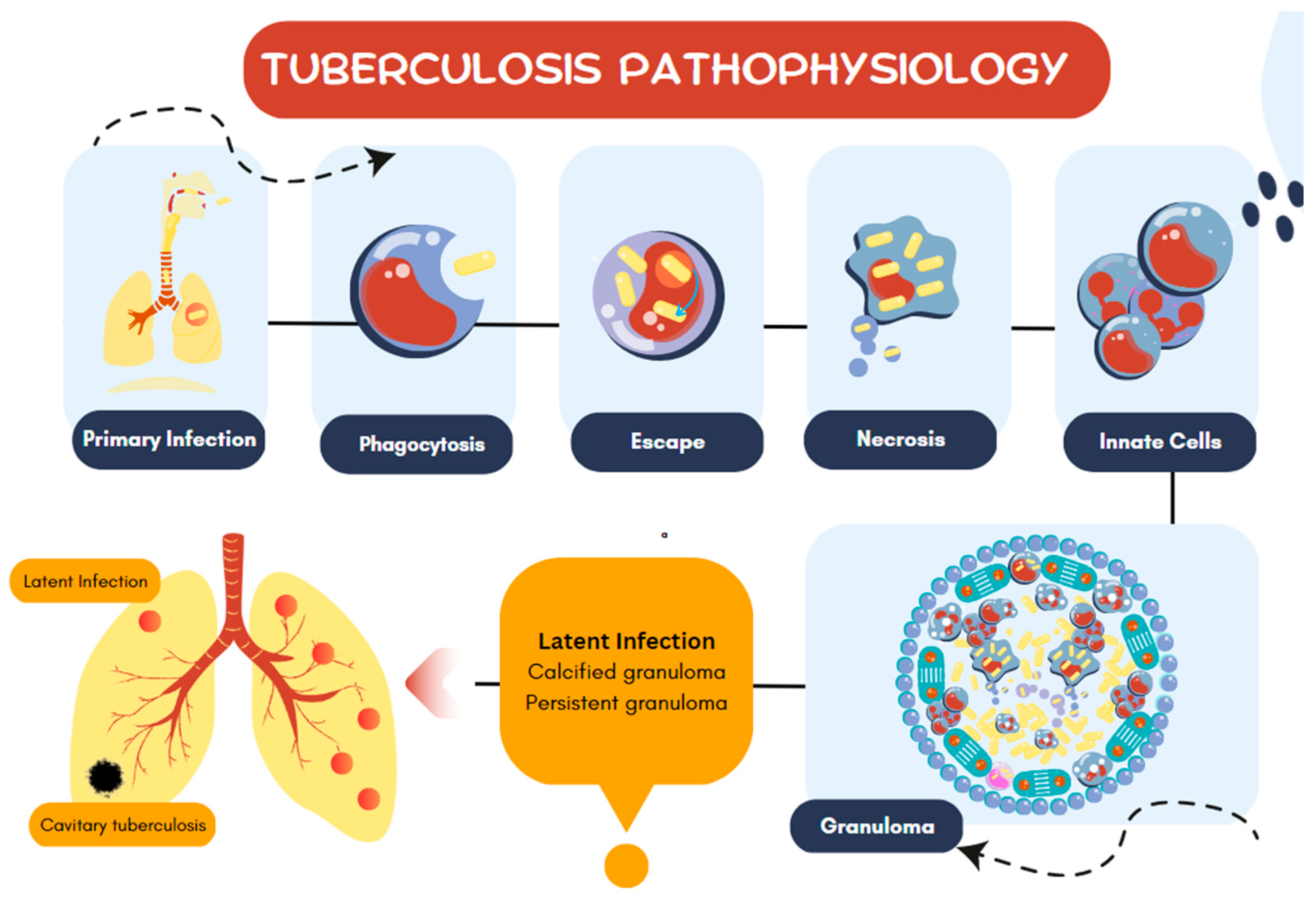
Figure 2.
MTB peptidoglycan synthesis. MTB peptidoglycan is synthesized from precursor sugars present in the bacillus cytoplasm. Mur enzymes sequentially add five amino acids to the NAM, then the NAM is inserted into the inner leaf of the cytoplasmic membrane. The MurG protein links NAM to NAG and MurJ transports the block to the outer layer of the plasma membrane. In the periplasm transglycosylase proteins and transpeptidases such as PBPs and Pon will link NAG-NAM building blocks to form peptidoglycan layers.
Figure 2.
MTB peptidoglycan synthesis. MTB peptidoglycan is synthesized from precursor sugars present in the bacillus cytoplasm. Mur enzymes sequentially add five amino acids to the NAM, then the NAM is inserted into the inner leaf of the cytoplasmic membrane. The MurG protein links NAM to NAG and MurJ transports the block to the outer layer of the plasma membrane. In the periplasm transglycosylase proteins and transpeptidases such as PBPs and Pon will link NAG-NAM building blocks to form peptidoglycan layers.
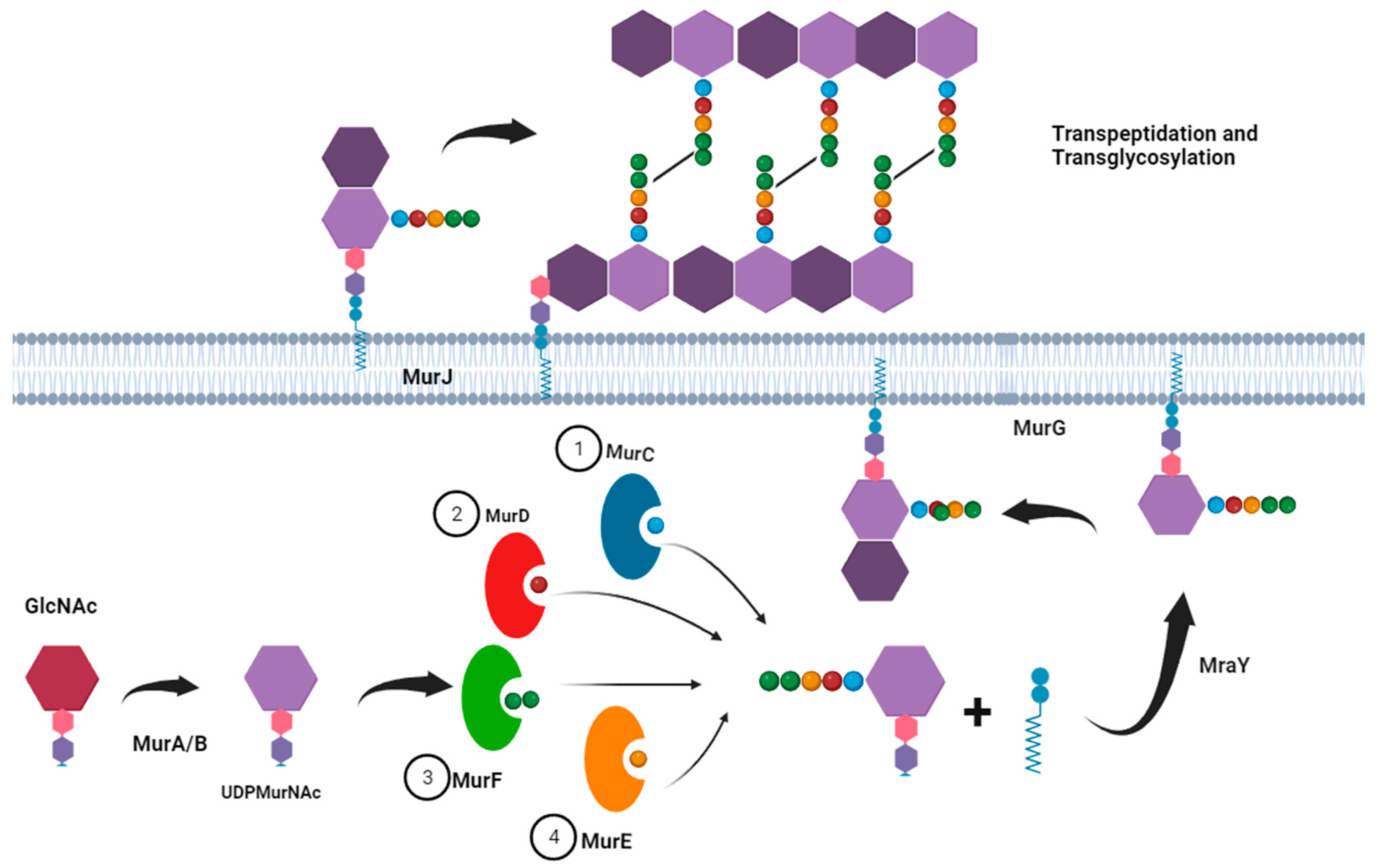
Figure 3.
MTB mycomembrane. The MTB mycomembrane is a complex association of several molecules. The first layer is composed of the phospholipid bilayer of the cytoplasmic membrane, followed by a small periplasm. Also in the periplasm is a thin layer of peptidoglycan. Associated with the plasma membrane are several liporabinogalactans that branch with arabinogalactans. Mycolic acids and surface lipids are also linked to arabinogalactans, in addition to the presence of proteins such as the 85B antigen.
Figure 3.
MTB mycomembrane. The MTB mycomembrane is a complex association of several molecules. The first layer is composed of the phospholipid bilayer of the cytoplasmic membrane, followed by a small periplasm. Also in the periplasm is a thin layer of peptidoglycan. Associated with the plasma membrane are several liporabinogalactans that branch with arabinogalactans. Mycolic acids and surface lipids are also linked to arabinogalactans, in addition to the presence of proteins such as the 85B antigen.
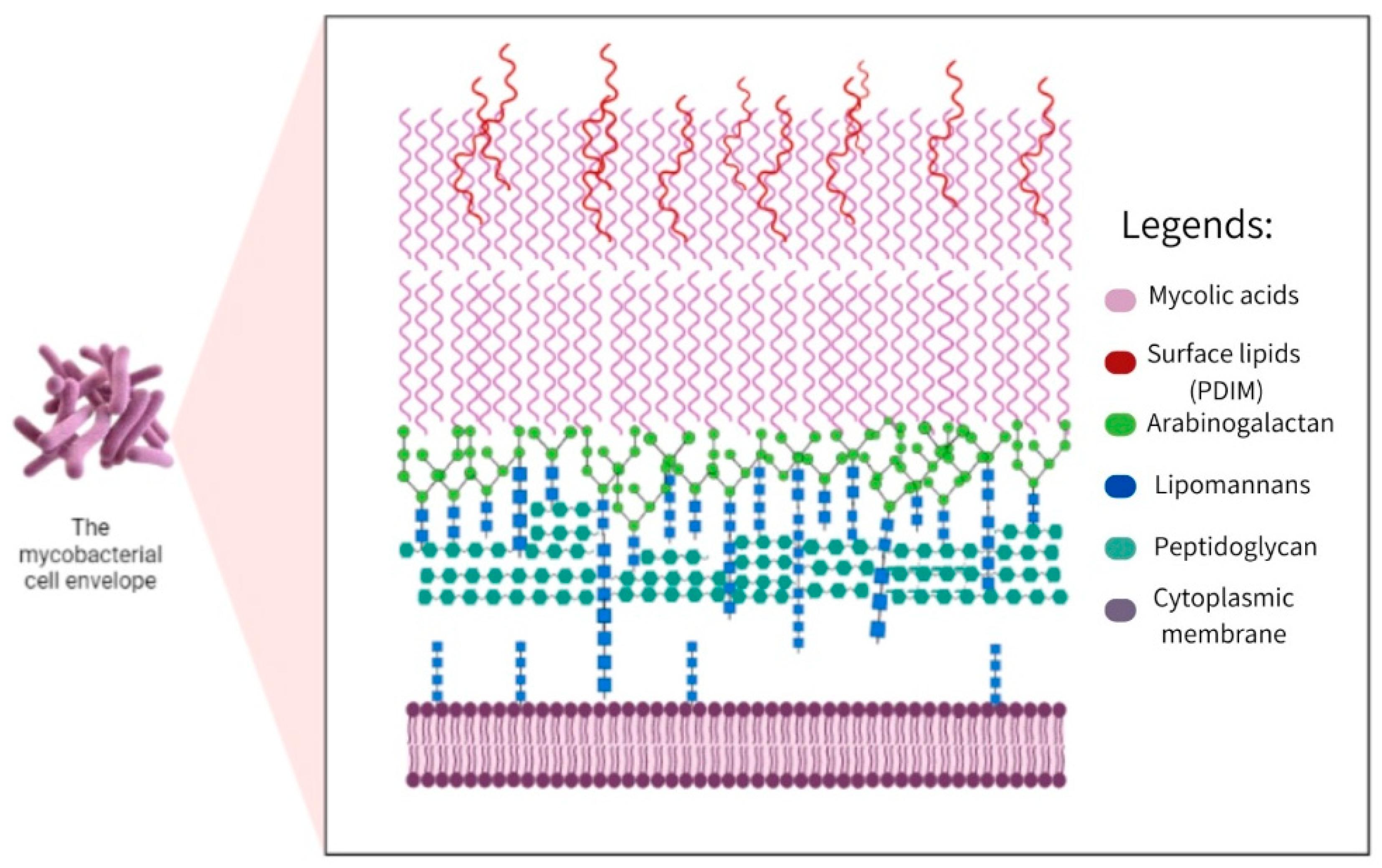
Figure 4.
Comparison of electron transport chains. Point A represents the classic electron transport chain used in aerobic conditions, for example, in the in vitro phase, formed by the complex Cox, Nuo, Sdh1, Hyd1, Qcr, Cta participates in this pathway alanine dehydrogenase, proline dehydrogenase, glycerol3-phosphate dehydrogenase and Hyd2. Under energy boundary conditions, the Cox complex, Nuo, Sdh1, Hyd1 and Hyd2 are upregulated. Point B represents the alternative electron transport chain, used in case of limited oxygen concentration, to carry out oxidative phosphorylation, in which it makes use of alternative electron donors and acceptors, the complexes that make up the chain are Mqo, Ndh, Sdh2, Hyd1, Nar, Cyd, Frd, Hyd3 and Hyd2, and the upregulated complexes are Mqo, Ndh, Sdh2, Hyd1, Hyd2, Hyd3, Cyd and Frd.
Figure 4.
Comparison of electron transport chains. Point A represents the classic electron transport chain used in aerobic conditions, for example, in the in vitro phase, formed by the complex Cox, Nuo, Sdh1, Hyd1, Qcr, Cta participates in this pathway alanine dehydrogenase, proline dehydrogenase, glycerol3-phosphate dehydrogenase and Hyd2. Under energy boundary conditions, the Cox complex, Nuo, Sdh1, Hyd1 and Hyd2 are upregulated. Point B represents the alternative electron transport chain, used in case of limited oxygen concentration, to carry out oxidative phosphorylation, in which it makes use of alternative electron donors and acceptors, the complexes that make up the chain are Mqo, Ndh, Sdh2, Hyd1, Nar, Cyd, Frd, Hyd3 and Hyd2, and the upregulated complexes are Mqo, Ndh, Sdh2, Hyd1, Hyd2, Hyd3, Cyd and Frd.
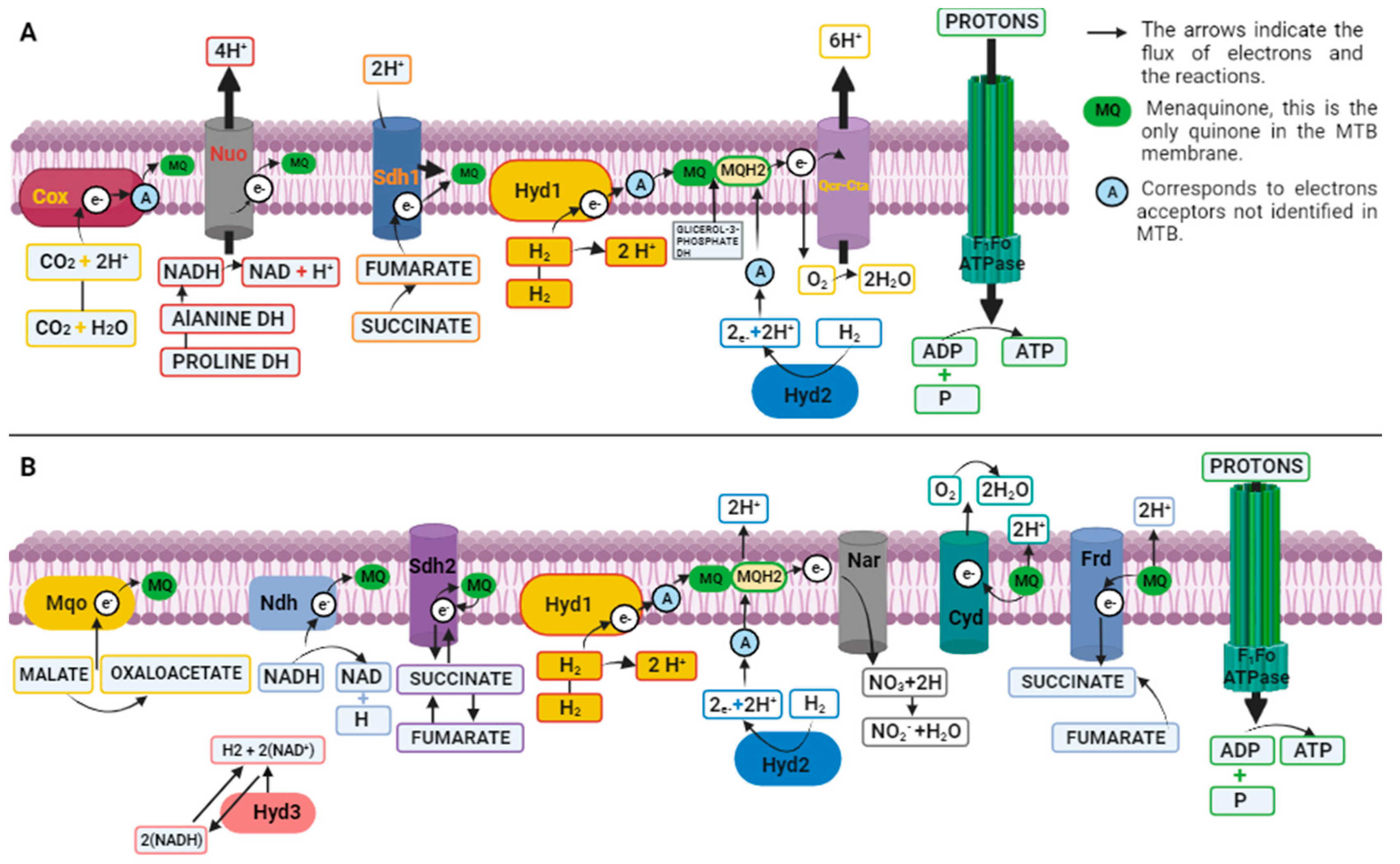
Figure 6.
MTB lineages adapted to man. Lineage 1: Philippines and Indian Ocean; Lineage 2: East Asia; lineage 3: India and East Africa; Lineage 4: Europe and the Americas; Lineage 5: M. africanum 1 in West Africa; Lineage 6 M. africanum 2 in West Africa; Lineage 7: Ethiopia; And species related to animals such as: M. bovis, M. pinnipedii, M. microti, M. orygis; and M. canetti. Also, which deletions from RDs 1 to 14 do some strains have; strain 5 RD9, strain 6 with RD3, M. microti with RD7, RD8, RD9, RD10 and RDmic and M.bovis with RD4, RD5, RD6, RD7, RD8, RD9 and RD10; And M. canetti has no deletion, harboring all RDs including RvD and TBD1.
Figure 6.
MTB lineages adapted to man. Lineage 1: Philippines and Indian Ocean; Lineage 2: East Asia; lineage 3: India and East Africa; Lineage 4: Europe and the Americas; Lineage 5: M. africanum 1 in West Africa; Lineage 6 M. africanum 2 in West Africa; Lineage 7: Ethiopia; And species related to animals such as: M. bovis, M. pinnipedii, M. microti, M. orygis; and M. canetti. Also, which deletions from RDs 1 to 14 do some strains have; strain 5 RD9, strain 6 with RD3, M. microti with RD7, RD8, RD9, RD10 and RDmic and M.bovis with RD4, RD5, RD6, RD7, RD8, RD9 and RD10; And M. canetti has no deletion, harboring all RDs including RvD and TBD1.
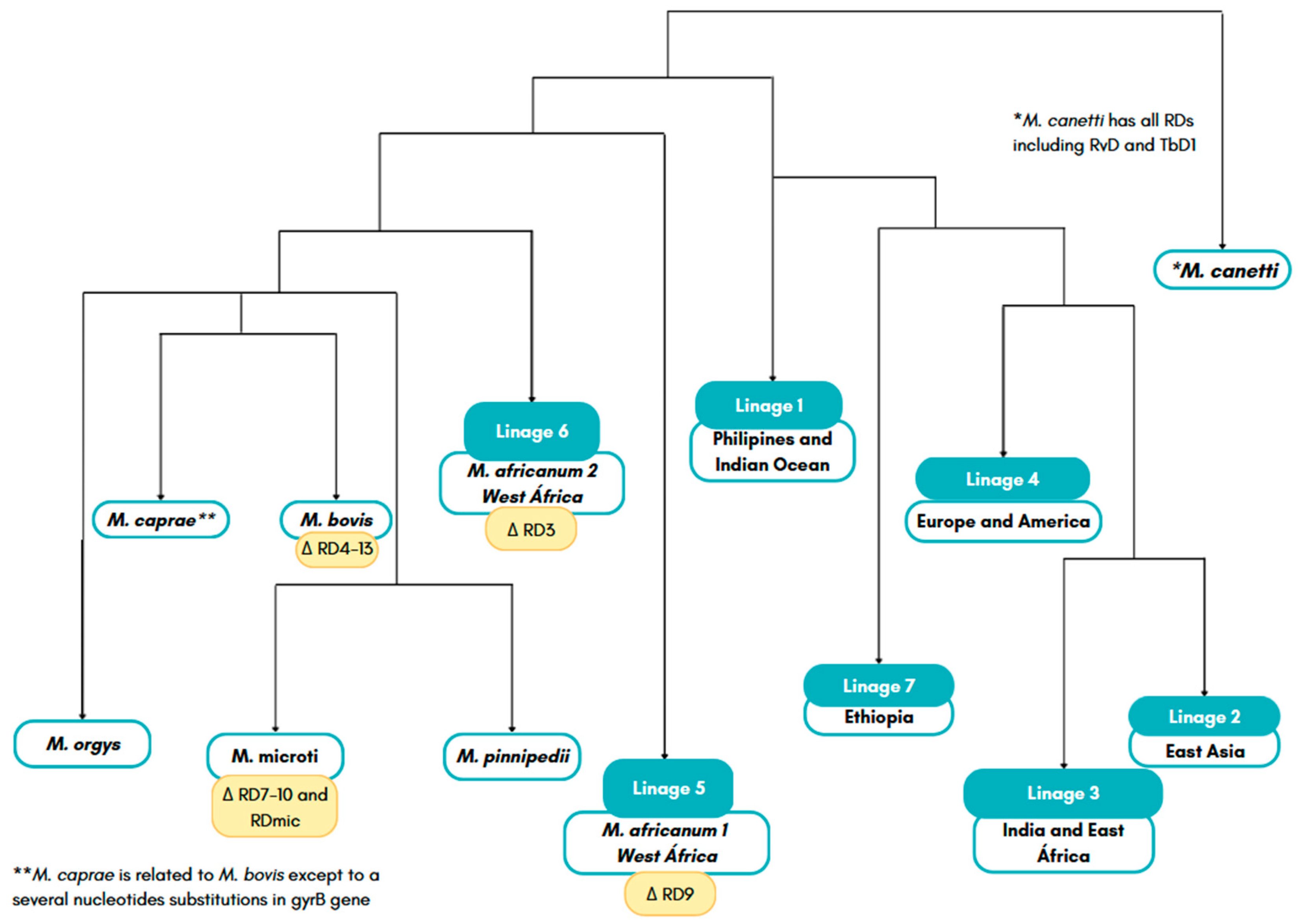
Table 1.
Mycobacteriacea groups and complex.
| Group or Complex | Principal Species | Pathogenicity and Disease1 |
|---|---|---|
|
M. tuberculosis Complex (MTBC) |
tuberculosis kansasii gastri bovis africanum |
Pathogenic/ tuberculosis |
| M. kansasii Group |
kansasii gastri |
Pathogenic/ pulmonary and soft tissue |
| M. avium Complex |
avium colombiense intracellulare |
Pathogenic/pulmonary in immunocompromised |
| M. simiae Complex |
kubicae florentinum |
Generally non-pathogenic |
| M. celatum Group |
xenopi celatum |
Pathogenic/pulmonary in immunocompromised |
| M. terrae Complex |
terrae hibernae |
Generally non-pathogenic |
| M. smegmatis Group |
smegmatis thermoresistible |
Generally non-pathogenic |
| M. fortuitum Group |
fortuitum peregrinum |
Pathogenic/soft tissue |
| M. abscessus-chelonae Complex |
abscessus chelonae |
Pathogenic/pulmonary in immunocompromised |
1 Pathogenicity is considered as the main characteristic, some mycobacteria only cause disease in immunosuppressed patients and therefore were not considered pathogenic.
Table 4.
Sigma factor of MTB.
| Sigma | Presence | Response | Observation |
|---|---|---|---|
| σA | All mycobacteria | Nutritional deficiency and the general response to the stress |
During the infection it was observed that its expression increases in some strains but not for the H37Rv strain |
| σB | All mycobacteria | Nutritional deficiency and the general response to the stress |
It is not essential for the survival of the bacillus |
| σC | All pathogenic mycobacteria | All pathogenic mycobacteria |
Act on the Immunopathological phenotype of tuberculosis |
| σD | All mycobacteria except M. leprae | It regulates genes linked to starving response during nutritional deficiency |
Act on the Immunopathological phenotype of tuberculosis |
| σE | All mycobacteria | Action during stress on the bacillary surface and during the response to thermal shock |
Influence on the immunopathological phenotype |
| σF | All mycobacteria except M. leprae | Involved in the biosynthesis of the mycobacterial envelope |
Influencing the immunopathological phenotype |
| σG | All mycobacteria except M. leprae |
Acts on the SOS response, a global response to DNA damage, and acts on the survival of the bacillus during macrophage infection |
Present as characteristics the presence of over 120 amino acid residues in the C-terminal |
| σH |
All mycobacteria except M. leprae |
Includes participation during oxidative stress, heat stress |
Influence on the immunopathogenic phenotype |
| σI |
Only in pathogenic mycobacteria |
It is induced during thermal shock processes |
Present as characteristics the presence of over 120 amino acid residues in the C-terminal |
| σJ |
MTB | Acting during oxidative stress |
Present as characteristics the presence of over 120 amino acid residues in the C-terminal |
| σL | MTB | Is bound to the virulence and biosynthesis of phthiocerol dimycocerosate |
An essential lipid for the virulence of MTB |
| σM |
MTB |
Is responsible for the long-term adaptation of the bacillus in vivo |
- |
| σK | MTB |
Yet its performance is unknown |
- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



