
Submitted:
15 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
In Pakistan, bats are one of the dominant mammals that play an important role in the ecosystem in terms of pollination, seed dispersal, and control of pest insects. Bats have also played an im-portant role in the emergence and transmission of zoonotic pathogens; however, most current studies focus on viral pathogens, not potential bacterial pathogens. This study was designed to estimate the prevalence and antibiotic profiling of Staphylococcus (S.) aureus in oral and rectal samples from bats captured in northern Pakistan and to determine the factors associated with in-fection. Two hundred individual bats of five species: Pipistrellus javanicus (n = 17), Pipistrellus pipistrellus (n = 10), Rhinopoma microphyllum (n = 48), Rousettus leschenaultii (n = 124), and Scotophilus kuhlii (n = 1) were captured for non-lethal collection of oral and rectal samples to iso-late S. aureus. Bats were sampled from three sites: a natural cave, a man-made castle, and an an-imal shed, in Khyber Pakhtunkhwa and Punjab provinces. Oral (n = 200) and rectal (n = 200) swabs were collected from each individual bat using sterile cotton swabs specifically for use in bacteriological studies. Each isolate of bacteria was identified by using phenotypic tests and con-firmed as S. aureus based on PCR assay. Out of a cumulative four hundred samples, 80 swabs were positive for S. aureus including 47 rectal and 33 oral swabs. Prevalence of S. aureus infection varied significantly among species, with Rousettus leschenaultii exhibiting the highest prevalence (n = 77; 37.90%). In addition to bat species, prevalence varied significantly among habitats but not between sex, age class, or reproductive status. This study confirmed the occurrence of S. aureus in oral and rectal microbiota of bats in Pakistan. Importantly, S. aureus isolates showed resistance to tetracycline, gentamicin, and erythromycin and carried resistant genes such as TetK, TetM, Erma, and aacA-D. In this regard, efforts should be taken to educate the local communities on how to minimize exposure to an antibiotic-resistant bacterial pathogen through contact with bats while simultaneously increasing the awareness of protecting bats as a vital component of our ecosystem.
Keywords:
antibiotic resistance
; Chiroptera
; Staphylococcus aureus
; microbiota
; molecular identification
; risk factors
; bacteria
1. Introduction
Pakistan has a high diversity of bat species with 50 recognized species belonging to 26 genera and 8 families [1,2]. There are numerous ecological and epidemiological traits of bats that make them suitable as reservoirs of pathogens [3,4]. Some bat species can be highly mobile which could facilitate the movement of pathogens between populations, such as Australian flying-fox individuals have been observed flying up to 50 km in a night [5,6]. Different bat species have varied social structures, some are solitary roosters and other species roost in colonies of hundreds to thousands of individuals and form mixed species roosts, a potential factor of transmitting pathogens to co-habituating bats. Furthermore, bats occupy a wide range of habitats from caves, woodlands, and forests to house openings, roof trusses, and crevices in urban areas. During the early migration season fruit bats such as Eidolon helvum often roost by the thousands in trees located in dense urban areas which increases human exposure to the bats’ excreta [7,8,9].
Bats are reservoir hosts to zoonotic and non-zoonotic pathogens such as bacteria, fungi, and viruses [9,10]. However, most research has focused on viruses associated with bats but relatively little research has been conducted on bacterial associations [13]. Previous studies have shown that the nasopharynx and digestive system of fruit bats are probably colonized by the S. aureus complex which showed resistance against tetracycline and erythromycin [14]. Members of the S. aureus complex include S. aureus, S. argenteus, and S. schweitzeri [15]. In humans, S. argenteus can cause skin and soft tissue infections, as well as bacteremia [16,17]. Staphylococcus schweitzeri, on the other hand, mostly colonizes non-human primates and bats [14,18]. However, three cases of S. schweitzeri infection in humans have been documented, with a suspected zoonotic source [19]. Apart from Staphylococcus spp., one study identified 34 bacterial species isolated from fecal swabs of bats kept in captivity. Most of these bacteria are not known to cause sickness in animals and humans except Leptospira sp., Bartonella, and Barrelia, of which some may be pathogenic to humans [20]. Some enteric bacteria such as Shigella sp., Yersinia sp., and Salmonella sp., which are causative agents of human and animal diseases that were also isolated from fecal and tissue samples (i.e., kidney, heart, spleen, liver, lung) of different species of bats [19,20,21].
Some bat species can be a source of multidrug resistance bacteria which pose a risk human and animal health [24,25,26,27]. Antimicrobial resistance genes of bacterial species have been isolated from bat guano samples of Myotis blythii and M. myotis [23], and antibiotic resistance genes were detected in fecal samples of wild and captive grey-headed flying foxes [28]. Antibiotic resistant genes have also been detected in other volant wildlife species [29]. For example, antibiotic resistant genes such as tet(M), erm(A), blaZ, erm(T), fexA, str, eta, and tst were detected in tracheal samples from white storks captured in residential areas [30]. Tetracycline is the drug of choice for the control of certain infections in dairy cattle and poultry, however, excessive use of such widely used antibiotics can result in the development of resistance. The prevalence of antimicrobial resistance (AMR) genes in wildlife species, including bats, could contribute to the epidemiology of what is a major threat to human populations all over the globe. In this circumstance, it is important to study AMR from a ‘One Health’ perspective that includes the diversity of free-ranging wildlife [29,30].
There are few studies on the occurrence of antibiotic resistant bacteria associated with oral and rectal microbiota in bats, however, such studies are lacking in Pakistan. The present study was conducted using molecular and bacteriological diagnostics to identify the presence and antibiotic resistance profile of bacteria in oral and rectal samples collected from bat species captured in KPK and Punjab regions of Pakistan. We also assess the environmental and demographic factors associated with S. aureus prevalence in sampled bat populations.
2. Materials and Methods
2.1. Study area
The present study was conducted at three sites in two different provinces of Pakistan specifically Khyber Pakhtunkhwa and Punjab (Figure 1). Bats were captured and sampled from Malakand caves and a nearby livestock shed in August 2018 and Sheikhupura fort in October 2019. Additional details on the three sampling sites is as follows:
Site I: A cave in district Malakand of Khyber Pakhtunkhwa province (34.4897° N, 71.7978° E), previously used for the mining of chromite but now has become a roosting site for bats after the government banned mining. The cave consisted of five passages which were interconnected mainly by a narrow passage internally. Forty-eight greater mouse-tailed bats (Rhinopoma microphyllum) and 42 fulvous fruit bats (Rousettus leschenaultii) were captured from this site.
Site II: Bats were captured from the cracks of a dilapidated brick shed traditionally used to house livestock. The shed was in Malakand district of Khyber Pakhtunkhwa province (34.07757 N° E71.83435° E), located near to Site I. We captured 10 common pipistrelles (Pipistrellus pipistrellus).
Site III: Sheikhupura fort dates back to 1607 AD. Sheikhupura fort is a very old multi-story building, and due to its dilapidated condition bats occupied crevices in the brick buildings as their roosting site (31.70813° N, 73.99078° E). We captured 17 Java pipistrellus (Pipistrellus javanicus), 82 fulvous fruit bats (Rousettus leschenaultii), and 1 Asiatic lesser yellow house bat (Scotophilus kuhlii).
2.2. Collection of samples and data
Bats were captured (n = 200) by using different types of nets (hand and mist net) and were kept in hygienic cotton bags to prevent cross-contamination. Oral (n = 200) and rectal (n = 200) swabs were collected from each bat using a sterile cotton swab and transferred into an Eppendorf tube (1.5 mL) containing phosphate buffer solution and transported in a dry shipper to the laboratory for bacteriological studies. Bat species, age class, reproductive status, and sex were recorded in addition to the habitat type (natural vs. manmade). All bats were released after non-lethal sample collection.
2.3. Isolation and phenotypic identification of bacteria
Samples were enriched in nutrient broth and incubated at 37oC for 24 hours. After enrichment, samples were streaked on mannitol salt agar and kept at 37°C for 24 hours. Suspected colonies were subculture on tryptic soya agar plates for purification. After 24 hours of incubation, individual S. aureus colonies were identified colony morphology, shape, and color. Each isolate was identified using Gram staining, oxidase, catalase, indole, voges Proskauer, and urease tests according to standard procedure [33]. Finally, bacterial isolates were preserved in 50% glycerol till further analysis.
2.4. Molecular confirmation of bacteria
Nucleic acid was extracted from isolated bacteria using the FavorPerp TM DNA extraction mini kit (Favorgen Biotech Crop, Taiwan). Molecular confirmation of S. aureus using 16S rRNA gene of 484 base pairs was done [34]. For PCR initial denaturation was done at 94 ºC for 3 min, followed by 35 cycles of amplification with 94 °C for 30 sec, annealing at 55 °C for 30 sec, and extension at 72 °C for 30 sec. Five microliters of the amplified product were subjected to 1.5% agarose gel having ethidium bromide stain. Gel documentation system was used for the confirmation of PCR products (Gel Doc EZ imager). PCR products were sequenced in one direction only using primer 16S rRNA (1ST BASE, Malaysia) and sequences were checked using BioEdit (Version 7.2.5) and EMBOSS Seqret. New sequences were submitted to GenBank (accession numbers: OP001633- OP001637).
2.5. Phenotypic and genotypic antibiotic resistance profiling
After confirmation, Staphylococcus aureus isolates were further tested for antibiotic sensitivity against commonly used antimicrobials agents i.e. Tetracycline (30 µg), erythromycin (15 µg), and gentamicin (10 µg) by disc diffusion method [35]. Moreover, antibiotic-resistant genes of tetracycline such as Tetk and TetM were detected with modification as described previously [36]. PCR conditions for the detection of Tetk resistance gene were initial denaturation at 94°C for 3 min, followed by denaturation at 94°C for 30 sec, annealing 55°C for 30 sec, elongation at 72°C for 30 sec, and final elongation at 72°C for 4 min for 30 cycles [36]. In the case of TetM gene PCR conditions were initial denaturation at 96°C for 3 min, denaturation at 96°C for 30 sec, annealing at 54°C for 30 sec, elongation at 72°C for 30 sec, and final elongation at 72°C for 4 min for 30 cycles using a thermocycler (T100TM Thermal cycler, BioRad, Singapore). The detection of erythromycin resistance gene (ermA) was done as described by [37] with certain modifications; PCR conditions were initial denaturation at 94°C for 3 min, denaturation at 94°C for 30 sec, annealing at 55°C for 30 sec, elongation 72°C for 30 sec, and final elongation at 72°C for 4 min for 30 cycles. Gentamicin resistance gene (aacA-D) was detected with modifications as described by[38]. PCR conditions were initial denaturation at 94°C for 3 min, denaturation at 94°C for 30 sec, annealing at 52°C for 30 sec, elongation at 72°C for 30 sec, and final elongation 72°C for 4 min for 30 cycles.
2.6. Statistical analysis
The role of biological (bat species, age class, sex, and reproductive condition) and environmental (i.e., habitat type - natural, manmade) factors on the prevalence of S. aureus in oral and rectal swabs were analyzed using Chi-square tests. Post hoc pairwise Wilcox tests were performed if a factor had a significant influence on prevalence. A p-value less than 0.05 was considered significant. All statistical analyses were conducted using R ver. 4.2.2. with base functions [39]
3. Results
In our study, four hundred (200 oral and 200 rectal) swabs from 5 bat species were collected for isolation of Staphylococcus aureus. Overall, fifty-eight (14.5%) swabs were positive for Staphylococcus aureus and were found positive for voges proskauer, oxidase, and urease while negative for Gram stain, catalase, and indole test. We sequenced PCR products of the 16S rRNA gene for 5 samples, confirming these isolates were S. aureus (Genbank Accession numbers: OP001633- OP001637). All five sequences were identical across the 484 base pairs. The highest prevalence of Staphylococcus aureus was isolated from Scotophilus kuhlii (n = 1, 100%), followed by Rousettus leschenaultii (n = 47, 37.90%), and Pipistrellus javanicus (n = 4, 23.52%). Isolates of Staphylococcus sp. were recovered more frequently from rectal swabs (n=31/200, 15.50%) vs. oral swabs (n=19/200, 9.50%) of bats. Isolates were recovered from both rectal and oral swabs collected from 8 individual bats (4% of animals). For oral samples, the highest prevalence of Staphylococcus aureus was recorded in Rousettus leschenaultii (n = 16, 12.90%), followed by Pipistrellus javanicus (n = 2, 11.76%) and Rhinopoma microphyllum (n = 1, 2.08%), respectively. However, rectal samples positive for Staphylococcus aureus were highest in Scotophilus kuhlii (n = 1, 100%), followed by Rousettus leschenaultii (n = 25, 20.16%) and Pipistrellus javanicus (n = 2, 11.76%), respectively. Only two bat species had both swab types positive for Staphylococcus aureus, specifically Rousettus leschenaultii (n = 6, 4.84%) and Rhinopoma microphyllum (n = 2, 4.17%).

Table 1.
Prevalence of Staphylococcus aureus isolated from oral and rectal swabs from bat species sampled at sites across northern Pakistan.
Table 1.
Prevalence of Staphylococcus aureus isolated from oral and rectal swabs from bat species sampled at sites across northern Pakistan.
| Bat species | Swabs collected | S. aureusPrevalence (%) | ||
| Oral/Rectal (no. each) |
Oral (95% CI) |
Rectal (95% CI) |
Total (95% CI) |
|
| Pipistrellus javanicus | 17/17 | 11.76 (1.46 – 36.44) |
11.76 (1.46 – 36.44) |
23.52 (6.81 – 49.89) |
| Pipistrellus pipistrellus | 10/10 | 0 | 0 | 0 |
| Rhinopoma microphyllum | 48/48 | 2.08 (1.31 – 17.20) |
10.42 (3.47 – 22.58) |
12.5 (4.73 – 25.25) |
| Rousettus leschenaultii | 124/124 | 12.90 (11.47 – 25.62) |
20.16 (17.66 – 33.57) |
37.90 (29.35 – 47.05) |
| Scotophilus kuhlii | 1/1 | 0 | 100 (2.50 – 100) |
100 (2.50 – 100) |
| Total | 200/200 | 13.50 (9.10 – 19.03) |
19.50 (14.25 – 25.68) |
29.00 (22.82 – 35.82) |
The association between prevalence of S. aureus and factors such as sampling site, bat species, age, reproductive status, and sex is given in Table 2. The prevalence of S. aureus was significantly different (p < 0.05) amongst sampling sites and bat species. Sampling sites were also a contributing factor for the occurrence of S. aureus in oral and rectal samples which were collected from bats at roosting sites in caves (23.3%) and castle (37 %) while all bats collected from animal sheds were negative. We observed significant variation in prevalence by bat species: Rousettus leschenaultii (35.90 %), followed by Pipistrellus javanicus (23.52 %), Pipistrellus pipistrellus (0 %), and Rhinopoma microphyllum (12.5 %). While only one sample of Scotophilus kuhlii was captured and tested positive (Table 2). Male individuals (36.8 %) tended to be positive for S. aureus more often than female bats (24.1%), but the trend was not significant. There were no significant differences in infection status based on age class or reproductive status.
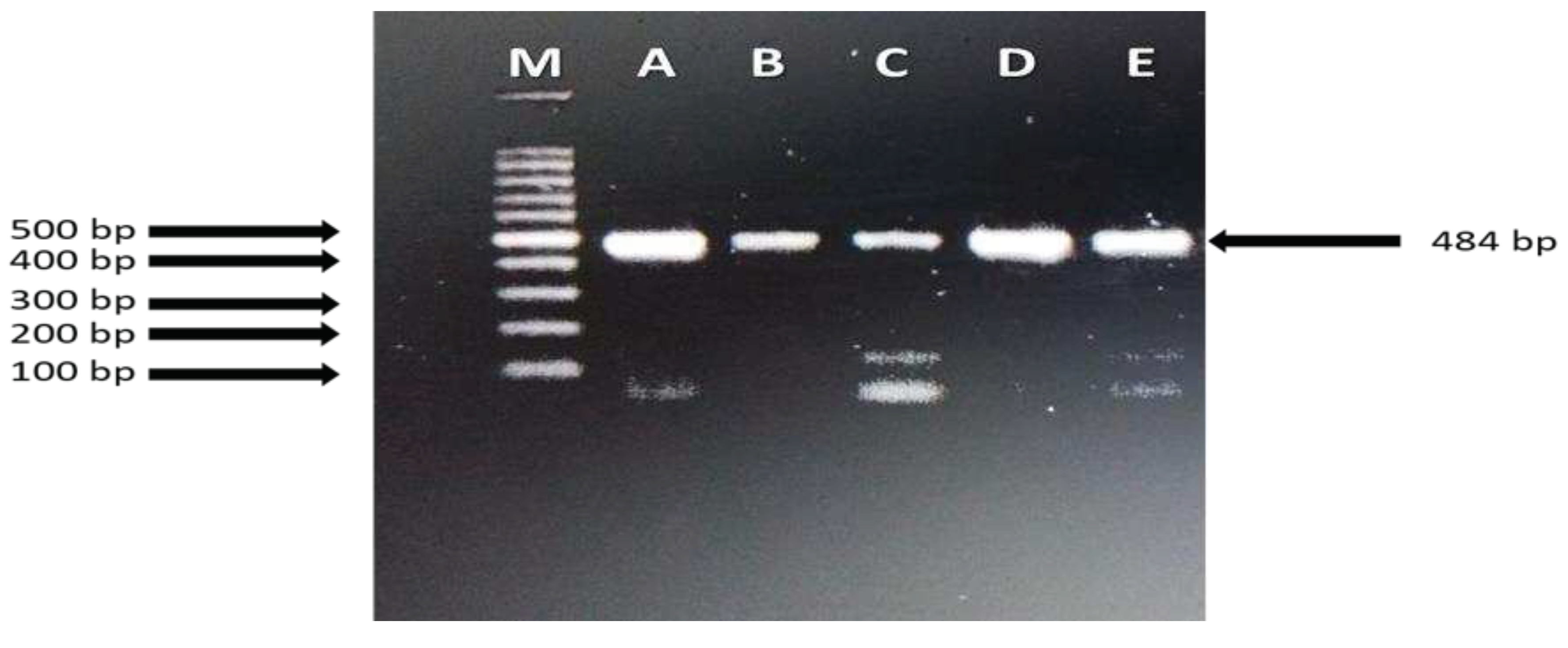
Figure 1.
PCR-based confirmation using primers for 16S rRNA of Staphylococcus isolated from oral and rectal swabs from five bat species. Sequences of these PCR products confirmed all samples were S. aureus. Lane M: (left to right) 100 bp ladder, Lane A to E: Field isolates of S. aureus from bat oral and rectal swab samples.
Figure 1.
PCR-based confirmation using primers for 16S rRNA of Staphylococcus isolated from oral and rectal swabs from five bat species. Sequences of these PCR products confirmed all samples were S. aureus. Lane M: (left to right) 100 bp ladder, Lane A to E: Field isolates of S. aureus from bat oral and rectal swab samples.
Based on the disc diffusion method, 58 isolates of S. aureus were subjected to antimicrobial susceptibility tests against three antibiotics: tetracycline, erythromycin, and gentamicin. S. aureus isolates were found to be highly resistant to tetracycline (43.10 %) and gentamicin (43.10 %). While 27.58 % isolates were also resistant to erythromycin (Table 3).
Further, all 58 isolates of S. aureus were analyzed for the presence of resistance genes using PCR assays. Among 58 isolates, 24 oral and 34 rectal isolates were subjected to tetracycline, erythromycin and gentamicin resistance genes detection. A comparatively higher number of oral S. aureus isolates were resistant against TetM (n = 18, 75 %), followed by TetK (n = 17, 70.08 %), aacA-D (n = 15, 62.5 %) and ermA (n = 8, 33.33 %) genes, respectively. Similarly in rectal isolates TetM and TetK genes were detected in 14 (41.17 %) and 12 (35.29 %) isolates while 13 (38.23 %) isolates of S. aureus carried aacA-D genes. In the case of erythromycin resistance gene; ermA was detected only in 6 (17.64 %) isolates (Table 4).
4. Discussion
Here we found that several bat species in Pakistan can harbor S. aureus in oral and fecal swabs, including strains with important antibiotic-resistant genes. S. aureus can cause serious infections both in animals and humans due to their zoonotic potential. In both developed and developing countries livestock and wildlife are the potential source for methicillin resistant S. aureus (MRSA) which can cause serious infections in humans and animals [40]. In this study we captured insectivorous and frugivorous bats and the highest prevalence of S. aureus (37.90% to 23.52%) was observed in fruit bats, although our sampling was not even and only included a few representative species from each category.
Previous studies have also identified S. aureus in diverse bat species globally. In Nigeria, a comparatively higher prevalence (78.6%) of S. aureus was found in fecal samples of straw-colored fruit bats (Eidolon helvum). Cross-species transmission of these S. aureus was inferred by multiple sequence typing (MLST) which revealed their close association with isolates of monkeys and other bats species in central and west Africa [41]. Another study conducted by Held et al. [14] in Gabon which sampled 55 R. aegyptiacus individuals found a much lower rate of S. aureus at 6.0 %. Held et al. [14] collected samples from bat species native to Gabon, i.e. Hipposideros ruber, Rhinolophus sp., Miniopterus minor, Pipistrellus annulus, and Chaerephon pumila and screened for S. aureus and S. schweitzeri. A low prevalence (4-6%) of S. schweitzeri and S. aureus was reported in Micropteropus pusillus and Rousettus aegyptiacus. Staphylococcus species were also prevalent in gut samples including feces, small intestine, and large intestine of Myotis altarium and Rhinolophus sinicus in China [42]. Additionally, some other studies have also reported low prevalence of S. aureus from African monkeys and bats [24,43]. In our analysis of 200 oral and 200 rectal swabs from four insectivorous and frugivorous bat species, we observed significant differences in prevalence of S. aureus among bat species. Variation has been seen in prevalence of bacteria with respect to host species of bats in Trinidad. Salmonella spp. was dominant bacteria isolated from two insectivorous (i.e. Molossus major and M. ater) and one fish-eating bat species (i.e. Noctilio leporinus), while E. coli was isolated from fruigivorous (i.e. Artebius sp. and Carollia perspicillata), sanguivorous (i.e. Desmodus rotundus, and Diaemus youngi), nectivorous (i.e. Glossophaga sp.), insectivorous (i.e. Molossus major, Mormoops sp., and Pteronotus parnelli), omnivorous (i.e. Phyllostomus discolor, P. hastatus, and Pteronotus parnelli) bats [22]. The interspecific factors that drive differences in S. aureus prevalence is not known, nor the mechanisms that lead to difference in prevelance even among co-roosting bat species. A study was conducted in Romania, in which bat guano, cave air and swabs of bat fur were collected from caves to determine of aerosolized microbes [44]. Myotis capaccinii, M. daubentonii, M. myotis, Miniopterus schreibersii, and Rhinolophus euryale were the main guano contributing bats species in the cave. Additional bacteria, including E. coli, Klebsiella pneumoniae, Chryseomonas luteola, Salmonella spp., Micrococcus spp., and Streptococcus; Staphylococcus were also detected in samples from Romania. Based on these findings it was concluded that guano of bats in caves had a higher concentration of airborne pathogens which could be a potential source of biohazards for other animals, including other bat species, as well as human beings [44].
We investigated the association of prevalence of S. aureus with respect to habitat type/location, bat species, age class, reproductive status, and sex. We only found significant differences between locations and species in our study. We observed S. aureus in oral and rectal swabs collected from 5 bat species from natural caves but also man-made sites including an abandoned castle and livestock shed. Little is known about the potential for S. aureus cross-species transmission from bats to people or other species. Some previous studies have sugested cross-species transmission of S. aureus from bat, particuarlaly in areas of high human habitation. Five out of eight Egyptian fruit bats kept in Copenhagen Zoo with other animals were found positive for S. aureus. Moreover, it was also observed that some isolates prevalent in these zoo animals were comparable to those observed in the human population suggesting their zoonotic and reverse-zoonotic potential [48]. Similarly, bat guano samples collected from Myotis blythii and M. myotis roosts located inside a Church building in Slovakia were positive for Staphylococcus sp. It was suggested that the presence of bats and accumulation of bat guano inside human settlements may be a potential risk for human health [25]. Presence of Staphylococcus in bats living in diverse habitats, e.g. forests, university campus, and caves, have been reported from previous studies from Brazil, Nigeria, and Romania, respectively [24,44,45]. Sampling site specific prevalence of Staphylococcus species in the gut microbiome of lesser horseshoe bats were revealed in bats captured from caves in India using metagenomics and culture dependent methods [49]. Similar reports of isolation of Staphylococcus species in guano layers and caves have been reported in the United States, Serbia, Slovenia [47,50,51]. However, a study from Italy revealed that bats species such as Tadarida teniotis, Miniopterus schreibersi, Myotis capaccinii, Myotis daubentoni, Pipistrellus kuhli and Myotis myotis had a diverse microflora but no detection of Staphylococcus was noted. Citrobacter freundii, Streptococcus faecalis, Pseudomonas putida, Proteus mirabilis, Kluyvera ascorbata, Klebsiella oxytoca, Escherichia coli, Escherichia blattae, Enterobacter cloacae and Hafnia alvei were dominant bacterial species in enteric samples of these bats [52].
Species level differences in bacterial diversity and prevelance have been previously noted in other bat studies. Bats captured from the Atlantic forest of Brazil identified Serratia marcescens, Pseudomonas sp., Klebsiella oxytoca and Escherichia coli and found that baterial richness was greater in frugivorous bats as compared to nectarivores, insectivores, carnivores, and sanguivores. Staphylococcus sp. was also detected from oral and rectal samples [45]. These bats have the habit of seasonal migration, and it was also observed that they abandon their colonies during the rainy season. These factors might be responsible for seasonal variation in bacterial richness of bats [46]. In another interesting study, fecal samples of twelve bat species had varying levels of prevalence of Staphylococcus. These bats were captured from eight parts of Slovenia during their autumn migration across central Europe [47].
It is well documented that antimicrobial resistance is an emerging issue for veterinary, wildlife, and human health. We found that S. aureus isolates were resistant against tetracycline, gentamicin and erythromycin based on disc diffusion methods. These findings were supported by previous investigations outside of Pakistan, but with differeing results. Previous studies of guano samples of Myotis blythii and M. myotis from Slovakia using similar methods and found S. aureus isolates resistant to tetracycline and erythromycin while sensitive for gentamicin [25]. In contrast to our study, one study from forest areas of Gabon found that all isolates of S. aureus derived from bat pharyngeal swabs were susceptible to tetracycline [14]. In another study, the antibiotic resistance pattern was determined from S. aureus isolates from bat guano collected from a cave in Algeria. Seven isolates were sensitive to all antibiotics while remaining four isolates were shown resistant to fusidic acid, cefoxitin and penicillin G [27]. Characterization of S. aureus isolated from wild and zoo animals in Germany few isolates were resistant against tetracycline [26]. Samples collected from straw colored fruit bats (Eidolon helvum) in Nigeria found that all isolates were susceptible to tetracycline and gentamicin and only 7.4% isolates showed resistance against erythromycin [24]. This previously observed variation in antimicrobial resistance might be due to difference in bat species, roosting sites, and geographical regions, or perhaps due to variation in exposure to antibiotics, or resistance genes in the environment based on location, foraging patterns, and other factors.
In this study, we also investigated the prevalence and provided evidence of antibiotic resistance genes such as TetA in S. aureus isolated from oral and rectal samples of Pakistani bats. Such type of resistance genes detection pattern was also reported from other wild animals. In Spain tetracycline resistance genes such as tet(K) and tet(L) were detected in S. aureus isolated from wild birds [29]. Another study from Spain reported detection of tet(M), erm(A), blaZ, erm(T), fexA, str, eta, tst, antibiotic resistant genes in tracheal samples of nestlings of white stork captured from human residential areas [30]. Apart from these, aacA-D genes associated with gentamicin resistance was reported from companion and wild animals in European countries which is verification of the presence of such genes in S. aureus isolates of bats [53,53]. Tetracycline is the drug of choice for the control of certain infections in dairy cattle and poultry birds. However, unregulated use of this antibiotic can be results in development of resistance, as observed in a study conducted by Liu et al. ([56] in China in S. aureus isolates of four different animals (cows, swine, chickens, ducks) and tetracycline resistance genes such tetK (22.38%) and tetM (52.45%) were detected.
5. Conclusions
In conclusion, the present study confirmed that multidrug resistance S. aureus is prevalent in the oral and rectal microbiota of bats in Pakistan. Sampling site (an environmental factor) and bat species (a biological factor) contributed significantly to differences in prevalence of S. aureus. We provide important baseline data and a first study for Pakistan and the locations and species we investigated. Together with other studies, our finding provide evidence that bats across the globe can harbor multidrug resistance bacteria which could pose human and animal health risks, and if associated with bat morbitity or mortality could be a threat to bat populations themselves. A deeper understanding of these bacterial associations and the animal and human interactions with bats are needed to understand and minimize the chance of cross-species transmission of such pathogens.
Author Contributions
Data curation, Attaullah, Shahzad Ali; Formal analysis, Attaullah and Kendra Phelps; Funding acquisition, Shahzad Ali and Kevin Olival; Investigation, Attaullah and Shahzad Ali; Methodology, Shahzad Ali, Kendra Phelps and Kevin Olival; Resources, Shahzad Ali, Kendra Phelps and Kevin Olival; Supervision, Shahzad Ali; Validation, Shahzad Ali, Attaullah; Writing – original draft, Attaullah, Shahzad Ali, Kendra Phelps and Kevin Olival; Writing review & editing, Attaullah, Shahzad Ali, Kendra Phelps and Kevin Olival.
Funding
This study was partially funded (sample collection, data analysis, and writing) by the US Department of the Defense, Defense Threat Reduction Agency (DTRA), under the award HDTRA1-17-0064 (‘Understanding the Risk of Bat-Borne Zoonotic Disease Emergence in Western Asia’). The content of the information does not necessarily reflect the position or the policy of the US federal government, and no official endorsement should be inferred.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Acknowledgments
We are thankful to provisional wildlife departments for their permission to conduct this study. We thank Mr. Luke Hamel from EcoHealth Alliance for his support of this work and assistance in the field.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mahmood ul Hassan, M.; Nameer, P.O. Diversity role and threats to the survival of bats in Pakistan. J. Anim. Plant. Sci. 2006, 16, 38–42. [Google Scholar]
- Mahmood-ul-Hassan, M.; Jones, G.; DietzI, C. Bats of Pakistan. The least known mammals. Verlag Dr. Muller, Saarbrucken, Germany. 2009, pp.168.
- Turmelle, A.S.; Olival, K.J. Correlates of viral richness in bats (order Chiroptera). Eco Health. 2009, 6, 522–539. [Google Scholar] [PubMed]
- Letko, M.; Seifert, S. N.; Olival, K. J.; Plowright, R. K.; Munster, V. J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [PubMed]
- Strahan, R. (ed.). Mammals of Australia.1995. Reed.
- Hall, L.; Richards, R. Flying foxes and fruit and blossom bats of Australia. Australian Natural History Series. 2000. UNSW Press.
- Wilkinson, G.S.; South, J.M. Life history, ecology and longevity in bats. Aging Cell. 2002, 1, 124–131. [Google Scholar] [CrossRef]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: important reservoir hosts of emerging viruses. Clin Microbiol. 2006, 19, 531–545. [Google Scholar]
- Plowright, R.K.; Foley, P.; Field, H.E.; Dobson, A.P.; Foley, J.E.; Eby, P.; Daszak, P. Urban habituation, ecological connectivity and epidemic dampening: the emergence of Hendra virus from flying foxes (Pteropus spp.). Proc Biol Sci. 2011, 278, 3703–3712. [Google Scholar] [CrossRef]
- Wibbelt, G.; Kurth, A.; Yasmum, N.; Bannert, M.; Nagel, S.; Nitsche, A.; Ehlers, B. Discovery of herpesviruses in bats. J. Gen. Virol. 2007, 88, 2651–2655. [Google Scholar] [CrossRef]
- Julg, B.; Elias, J.; Zahn, A.; Koppen, S.; Becker-Gaab, C.; Bogner, J.R. Bat-associated histoplasmosis can be transmitted at entrances of bat caves and not only inside the caves. J. Travel. Med. 2008, 15, 133–136. [Google Scholar] [CrossRef]
- Whitaker, Jr.; JO, Ritzi.; C.M.; Dick, C.W. Collecting and preserving bat ectoparasites for ecological study. Ecological and behavioral methods for the study of bats. JHU Press. 2009, 2, 806–827.
- Muhldorfer, K.; Speck, S.; Wibbelt, G. Diseases in free-ranging bats from Germany. BMC Vet Res. 2011, 7, 1–11. [Google Scholar] [CrossRef]
- Held, J.; Gmeiner, M.; Mordmüller, B.; Matsiegui, P.B.; Schaer, J.; Eckerle, I.; Schaumburg, F. Bats are rare reservoirs of Staphylococcus aureus complex in Gabon. Infect Genet Evol. 2016, 47, 118–120. [Google Scholar] [PubMed]
- Tong, S.Y.; Schaumburg, F.; Ellington, M. J.; Corander, J.; Pichon, B.; Leendertz, F.; Giffard, P. M. 2015. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: the non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J.Syst. Evol. Microbiol. 2015, 65, 15. [Google Scholar]
- Dupieux, C.; Blondé, R.; Bouchiat, C.; Meugnier, H.; Bes, M.; Laurent, S.; Tristan, A. Community-acquired infections due to Staphylococcus argenteus lineage isolates harbouring the Panton-Valentine leucocidin, France, 2014. Eurosurveillance. 2015, 20, 21154. [Google Scholar] [PubMed]
- Chantratita, N.; Wikraiphat, C.; Tandhavanant, S.; Wongsuvan, G.; Ariyaprasert, P.; Suntornsut, P.; Peacock, S.J. Comparison of community-onset Staphylococcus argenteus and Staphylococcus aureus sepsis in Thailand: a prospective multicentre observational study. Clin. Microbiol. Ifect. 2016, 22, 458–11. [Google Scholar]
- Schaumburg, F.; Pauly, M.; Anoh, E.; Mossoun, A.; Wiersma, L.; Schubert, G.; Peters, G. Staphylococcus aureus complex from animals and humans in three remote African regions. Clin. Microbiol. Infect. 2015, 21, 345–e1. [Google Scholar]
- Okuda, S. , Sherman, D. J.; Silhavy, T. J.; Ruiz, N.; Kahne, D. Lipopolysaccharide transport and assembly at the outer membrane: the PEZ model. Nat Rev Microbiol. 2016, 14, 337–345. [Google Scholar]
- Tulsiani, S.M.; Cobbold, R.N.; Graham, G.C.; Dohnt, M.F.; Burns, M.A.; Leung, L.P.; Craig, S.B. The role of fruit bats in the transmission of pathogenic leptospires in Australia. Ann Trop Med Parasite. 2011, 105, 71–84. [Google Scholar] [CrossRef]
- Brygoo, E.R.; Simond, J.P.; Mayoux, A.M. The pathogenic enterobacteria of Pteropus rufus (Megachiroptera) in Madagascar. Societé de biologie de Madagascar. Bactériologie. 1971, 165, 1793–1795. [Google Scholar]
- Adesiyun, A.A.; Stewart-Johnson, A.; Thompson, N.N. Isolation of enteric pathogens from bats in Trinidad. J. Wild. Dis. 2009, 45, 952–961. [Google Scholar] [CrossRef]
- Reyes A., W. B.; Rovira, H.G.; Masangkay, J.S.; Ramirez, T.J.; Yoshikawa, Y.; Baticados, W.N. Polymerase chain reaction assay and conventional isolation of Salmonella spp. from Philippine bats. Acta. Sci. Vet. 2011, 39, 1–7. [Google Scholar]
- Akobi, B.; Aboderin, O.; Sasaki. T.; Shittu, A. Characterization of Staphylococcus aureus isolates from faecal samples of the straw-colored fruit bat (Eidolon helvum) in Obafemi Awolowo University (OAU), Nigeria. BMC Microbiol. 2012, 12, 1–8.
- Vandžurová, A.; Bačkor, P.; Javorský, P.; Pristaš, P. Staphylococcus nepalensis in the guano of bats (Mammalia). Vet. Microbiol. 2013, 164, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Febler, A.T, Thomas P.; Mühldorfer, K.; Grobbel, M.; Brombach, J.; Eichhorn. I.; Monecke, S.; Ehricht, R.; Schwarz, S. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from zoo and wild animals. Vet. Microbiol. 2018, 218, 98–103.
- Mairi, A.; Touati, A.; Pantel, A.; Yahiaoui Martinez, A.; Ahmim, M.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. First Report of CC5-MRSA-IV-SCCfus "Maltese Clone" in Bat Guano. Mcrooganisms. 2021, 9, 2264. [Google Scholar] [CrossRef] [PubMed]
- McDougall, F.; Boardman, W.; Power, M. High Prevalence of Beta-Lactam-Resistant Escherichia coli in South Australian Grey-Headed Flying Fox Pups (Pteropus poliocephalus). Microorganisms. 2022, 10, 1589. [Google Scholar]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C. A.; Camacho, M.C.; de la Puente, J.; Fernández-Fernández, R.; Torres, C. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu (A) in Magpies and Cinereous Vultures in Spain. Microbial Ecology. 2019, 78, 409–415. [Google Scholar]
- Gómez, P.; Lozano, C.; Camacho, M.C.; Lima-Barbero, J.F.; Hernández, J.M.; Zarazaga, M.; Torres, C. Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 2016, 71, 53–57. [Google Scholar]
- Loh, E.H.; Zambrana-Torrelio, C.; Olival, K.J.; Bogich, T.L.; Johnson, C.K.; Mazet, J.; A.k, Karesh, W.;Daszak. Targeting Transmission Pathways for Emerging Zoonotic Disease Surveillance and Control. Vec-Bor. Zoono Dis 2015, 432–437.
- White, A.; Hughes, J. M. Critical importance of a one health approach to antimicrobial resistance. EcoHealth. 2019, 16, 404–409. [Google Scholar]
- Holt JG. Bergey's manual of systematic bacteriology. vol. 1, Williams and Wilkins. 1986. Baltimore.
- Atyah, M.A.S.; Zamri-Saad, M.; Siti-Zahrah, A. First report of methicillin-resistant Staphylococcus aureus from cage-cultured tilapia (Oreochromis niloticus). Vet. Microbiol. 2010, 144, 502–504. [Google Scholar]
- Bauer, A.W.; Perry, D.M.; Kirby.; W.M. Drug usage and antibiotic susceptibility of staphylococci. J. A.Med. Ass. 1960, 173, 475–480. [CrossRef] [PubMed]
- Momtaz, H.; Dehkordi, F.S.; Rahimi, E.; Asgarifar, A.; Momeni, M. Virulence genes and antimicrobial resistance profiles of Staphylococcus aureus isolated from chicken meat in Isfahan province, Iran. J. Appl. Poult. Res. 2013, 22, 913–921. [Google Scholar]
- Anisimova, E.; Yarullina, D. Characterization of erythromycin and tetracycline resistance in Lactobacillus fermentum strains. Intl. J. Microbiol. 2018, 1–9. [Google Scholar]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R. A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. 2022.https://www.R-project.org/.
- Graveland, H.; Duim, B.; van Duijkeren, E.; Heederik, D.; Wagenaar, J.A. Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int.J. Med Microbiol. 2011, 301, 630–634. [Google Scholar]
- Olatimehin, A.; Shittu, A.O.; Onwugamba, F.C.; Mellmann, A.; Becker, K.; Schaumburg, F. Staphylococcus aureus complex in the straw-colored fruit bat (Eidolon helvum) in Nigeria. Front. Microbiol. 2018, 9, 162. [Google Scholar]
- Wu, H.; Xing, Y.; Sun, H.; Mao, X. Gut microbial diversity in two insectivorous bats: Insights into the effect of different sampling sources. Microbiology. Open. 2019, 8, 00670. [Google Scholar]
- Schaumburg, F.; Alabi, A.S.; Köck, R.; Mellmann, A.; Kremsner, P.G.; Boesch, C.; Peters, G. Highly divergent Staphylococcus aureus isolates from African non-human primates. Environ. Microbiol. Rep. 2012, 4, 141–146. [Google Scholar] [CrossRef]
- Borda, D.R.; Nastase-Bucur, R.M.; Spinu, M.; Uricariu, R.; Mulec, J. Aerosolized microbes from organic rich materials: case study of bat guano from Caves in Romania. J. Cave. Karst. Stud. 2014, 76, 114–126. [Google Scholar] [CrossRef]
- Cláudio, V.C.; Gonzalez, I.; Barbosa, G.; Rocha, V.; Moratelli, R.; Rassy, F. Bacteria richness and antibiotic-resistance in bats from a protected area in the Atlantic Forest of Southeastern Brazil. PloS. One. 2018, 13, 0203411. [Google Scholar] [CrossRef]
- Okon, E. Fruit bats at Ife: their roosting and food preferences (Ife fruit bat project no. 2). Nierian Field. 1974, 39, 33–40. [Google Scholar]
- Vengust, M.; Knapic, T.; Weese, J.S. The fecal bacterial microbiota of bats; Slovenia. PloS. One. 2018, 13, 0196728. [Google Scholar]
- Espinosa-Gongora, C.; Chrobak, D.; Moodley, A.; Bertelsen, M.F.; Guardabassi, L. Occurrence and distribution of Staphylococcus aureus lineages among zoo animals. Vet Microbiol. 2012, 2, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Selvin, J.; Lanong, S.; Syiem, D.; De Mandal, S.; Kayang, H.; Kumar, N.S.; Kiran, G.S. Culture-dependent and metagenomic analysis of lesser horseshoe bats’ gut microbiome revealing unique bacterial diversity and signatures of potential human pathogens. Microb. Pathog. 2019, 137, 103675. [Google Scholar] [PubMed]
- Newman, M.M.; Kloepper, L.N.; Duncan, M.; McInroy, J.A; Kloepper, J.W. Variation in bat guano bacterial community composition with depth. Front. Microbiol. 2018, 9, 914. [Google Scholar]
- Dimkić, I.; Stanković, S.; Kabić, J.; Stupar, M.; Nenadić, M.; Ljaljević-Grbić,M.; Žikić, V.; Vujisić, P.; Tešević, V.; Vesović, N.; Pantelić, D.; Savić-Šević, S.; Vukojević, J.; Ćurčić, S. Bat guano-dwelling microbes and antimicrobial properties of the pygidial gland secretion of a troglophilic ground beetle against them. Appl. Microbiol. Biotechnol. 2020, 104, 4109–4126.
- Di-bella, C.; C. Piraino, S.; Caracappa, L.; Fornasari, C.; Violani, B. Zava. Enteric Microflora in Italian Chiroptera. J.Mt Ecol. 2003, 7, 221–224.
- Strommenger, B.; Kehrenberg, C.; Kettlitz, C.; Cuny, C.; Verspohl, J.; Witte, W. Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and their relationship to human isolates. J. Antimicrob. Chemother. 2006, 57, 461–465. [Google Scholar]
- Luzzago, C.; Locatelli, C.; Franco, A.; Scaccabarozzi, L.; Gualdi, V.; Viganò, R. Clonal diversity, virulence-associated genes and antimicrobial resistance profile of Staphylococcus aureus isolates from nasal cavities and soft tissue infections in wild ruminants in Italian Alps. Veterinary Microbiol. 2014, 170, 157–161. [Google Scholar]
- Loncaric, I.; Kunzel, F.; Licka, T.; Simhofer, H.; Spergser, J.; Rosengarten, R. 2014 Identification and characterization of methicillin-resistant Staphylococcus aureus (MRSA) from Austrian companion animals and horses. Vet. Microbiol. 2014, 168, 381–387. [Google Scholar]
- Liu, B.; Sun, H.; Pan, Y.; Zhai, Y.; Cai, T.; Yuan, X.; Hu, G. Prevalence, resistance pattern, and molecular characterization of Staphylococcus aureus isolates from healthy animals and sick populations in Henan Province, China. Gut. Pathog. 2018, 10, 1–13. [Google Scholar]
Figure 1.
Study area map showing sampling sites in districts Malakand and Sheikhupura in Pakistan.
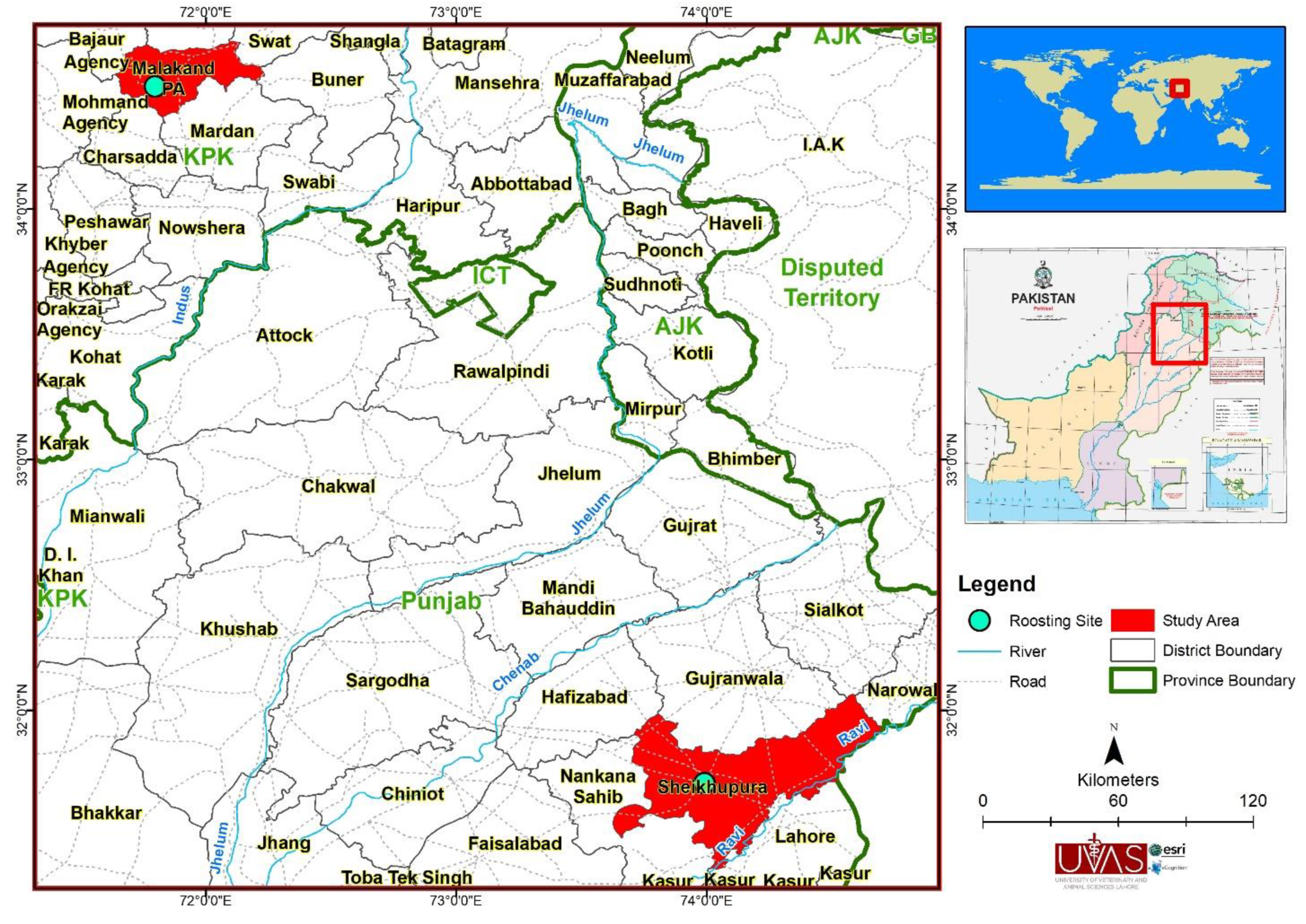
Table 2.
Identification of biological and environmental factors associated with the prevalence of Staphylococcus aureus.
Table 2.
Identification of biological and environmental factors associated with the prevalence of Staphylococcus aureus.
| Factors | Variables | No. samples collected |
Prevalence (%) (95% CI) |
Chi-square χ2 |
p-value |
|---|---|---|---|---|---|
| Site | Castle | 100 | 37.00 (27.57 – 47.24) |
8.60 | 0.01 |
| Cave | 90 | 23.33 (15.06 – 33.43) |
|||
| Animal Shed | 10 | 0 | |||
| Species | Pipistrellus javanicus | 17 | 23.52 (6.81 – 49.89) |
17.90 | < 0.01 |
| Pipistrellus pipistrellus | 10 | 0 | |||
| Rhinopoma microphyllum | 48 | 12.5 (4.73 – 25.25) |
|||
| Rousettus leschenaultii | 124 | 37.90 (29.35 – 47.05) |
|||
| Scotophilus kuhlii | 1 | 100 (2.50 – 100) |
|||
| Age | Adult | 149 | 29.53 (22.34 – 37.55) |
0.01 | 0.91 |
| Juvenile | 51 | 27.45 (15.89 – 41.74) |
|||
| Reproductive Status | Lactating | 2 | 50.00 (1.26 – 98.75) |
5.11 | 0.28 |
| Non-breeding | 90 | 22.22 (14.13 – 32.31) |
|||
| Non-scrotal | 1 | 0 | |||
| Post-lactating | 34 | 29.41 (15.10 – 47.79) |
|||
| Scrotal | 73 | 36.99 (25.97 – 49.09) |
|||
| Sex | Female | 124 | 24.19 (16.95 – 32.70) |
3.07 | 0.08 |
| Male | 76 | 36.84 (26.06 – 48.69) |
Table 3.
Breakpoint values of each antimicrobial agent and phenotypic antimicrobial susceptibility profiles of 58 Staphylococcus aureus isolate detected in this study according to Clinical & Laboratory Standards Institute (CLSS).
Table 3.
Breakpoint values of each antimicrobial agent and phenotypic antimicrobial susceptibility profiles of 58 Staphylococcus aureus isolate detected in this study according to Clinical & Laboratory Standards Institute (CLSS).
|
Antimicrobial class |
Antimicrobial agents |
Conc. (µg) |
Staphylococcus aureus | ||
| S | I | R | |||
| Tetracycline | Tetracycline | 30 µg | 15 (25.86) | 18 (31.03) | 25 (43.10) |
| Erythromycin | Macrolides | 15 µg | 20 (34.48) | 22 (37.93) | 16 (27.58) |
| Gentamicin | Aminoglycosides | 10 µg | 15 (25.86) | 18 (31.03) | 25 (43.10) |
Table 4.
PCR assays-based detection of antimicrobial resistance genes in Staphylococcus aurues isolated from oral and rectal samples from bat species in Pakistan.
Table 4.
PCR assays-based detection of antimicrobial resistance genes in Staphylococcus aurues isolated from oral and rectal samples from bat species in Pakistan.
| Total isolates tested | Source of isolation | No. isolates | Antibiotic resistance genes in Staphylococcus aureus | |||
| TetK (%) | TetM (%) | ermA (%) | aacA-D (%) | |||
| 58 | Oral | 24 | 17 (70.08) | 18 (75.00) | 8 (33.33) | 15( 62.5) |
| Rectal | 34 | 12 (35.29) | 14 (41.17) | 6 (17.64) | 13 (38.23) | |
| Total | 58 | 29 (50) | 32 (55.17) | 14 (24.13) | 28 (48.27) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



