
Submitted:
09 September 2023
Posted:
12 September 2023
You are already at the latest version
Abstract
Neurogenesis in the adult brain plays a major role in the cognitive ability of animals including learning and memory. Poria cocos (Schw.) Wolf (P. cocos) has been known as a traditional Chinese edible medicinal herb with the potential effect to improve learning and memory. However, the effect mechanisms are still elusive. In this study, significant neuronal differentiation-promoting activities of the ethanol extracts of P. cocos (EEPC) were tested in Neuro-2a cells and mouse cortical neural stem/progenitor cells (NSPCs). Subsequently, 97 compounds of EEPC were identified using UHPLC-Q-Exactive-MS/MS. After investigation of neuronal differentiation-promoting activities of 4 major compounds (Ethyl palmitoleate, L-(-)-Arabitol, Choline and Adenosine), only Choline has significant neuronal differentiation-promoting activity. In summary, we investigated the neuronal differentiation-promoting activity and chemical compositions of EEPC, the study of the neuronal differentiation-promoting effect of ether extract of P. cocos indicated P. cocos’s positive effect on learning and memory functions. This is the first investigation of the neuronal differentiation-promoting effect of ether extract of P. cocos based on similar research references.
Keywords:
Poria cocos (Schw.) Wolf
; Edible medicinal herb
; Neural progenitor cells
; Neurogenesis
; Neuronal Differentiation
1. Introduction
Neurogenesis and subsequent neurite outgrowth is a critical process for the formation of functional neural networks during brain development[1]. Neurodegenerative diseases, such as Alzheimer’s disease (AD) or Parkinson’s disease (PD), extensive neuronal loss and neurite atrophy are the major characteristics during the pathogenesis of disease[2]. Furthermore, Neurite loss is one of the typical features of neuronal injury. Neural stem/progenitor cells (NSPCs) are multi-potent cells that are able to self-renew and differentiate into neurons[3]. Therefore, promoting neuronal differentiation is an effective strategy for stem cell therapy and neural regeneration, which contributes to the reconstruction of neural circuits after neurodegeneration and brain injury[4, 5]. So far, a variety of natural products and their derivatives have been found to have neuronal differentiation-promoting activity and have been studied for neural regeneration.
Poria cocos (Schw.) Wolf (P. cocos) is a fungus used as a traditional Chinese edible medicinal herb[6]. According to the Chinese Pharmacopoeia, P. cocos has the effect of promoting diuresis, eliminating dampness, strengthening the spleen, and calming the heart[7]. Modern pharmacological research indicates that the chemical groups in P. cocos have been widely used in clinical treatments due to their various effects, such as diuretic, anti-oxidant, anti-tumor, anti-inflammatory, anti-bacterial, and anti-aging effects, among others[8-11]. Current studies have found that some traditional Chinese medicine prescriptions with P. cocos as the main raw material have potential effects on the treatment of neurodegenerative diseases. Bu-wang-san (BWS) is a classical traditional Chinese medicinal formula for enhancing the memory. It has been reported that BWS could improve the learning and memory functions through its effect on synaptic plasticity in ovariectomised rats[12]. Kai-xin-san, significantly alleviated the depressive symptoms by up-regulates the expressions of synaptic proteins on hippocampus of chronic mild stress induced rats[13]. Liu-wei-di-huang decoction has demonstrated neuroprotective effect on cognition deficits of diabetic encephalopathy in streptozotocin-induced diabetic rat[14]. Shen-hui soup enhanced neurite growth in PC12 cells which may be related to its effects in up-regulating the expression of GAP-43[15]. Moreover, our previous study has revealed that the ethanol extract of P. cocos induces differentiation in Neuro-2a cells. However, despite the potential benefits of P. cocos for the prevention and treatment of neurodegenerative disease, it has not been reported whether P. cocos can promote the differentiation of NSPCs. As a continuation of our previous study[16], in this study, we have further examined the effects of P. cocos on NSPCs differentiation. In addition, the UHPLC-MS method was used to explore the chemical basis of the promoting activity of P. cocos on neuronal differentiation. To our knowledge, this is the first report which shows that P. cocos has neuronal differentiation-promoting activity in NSPCs.
2. Materials and methods
2.1. Reagents
P. cocos (Figure 1a) was purchased from Hunan Province, China; FR180204, SP600125, Primary antibodies against anti-ERK1/2, anti-p-ERK1/2, anti-JNK1/2/3, anti-p-JNK1/2/3, anti-β-actin, HRP-conjugated anti-mouse and anti-rabbit were purchased from Beyotime Biotechnology (Shanghai, China); anti-β-tubulin III, DMSO and MTT were purchased from Sigma (St. Louis, MO, USA). Minimum Eagle’s Medium (MEM), foetal bovine serum (FBS), penicillin, and streptomycin were obtained from Hyclone (Logan, UT, USA). Choline, Adenosine, L-(-)-Arabitol, and Ethyl palmitoleate from Yuanye Biotechnology (Shanghai, China); all other chemicals and reagents are of analytical grade.
2.2. Preparation of Ethanol Extracts of P. cocos (EEPC)
EEPC was prepared as previously described. The dried sclerotium of P. cocos (Figure 1b) was grounded into powder and extracted twice with 10 volumes of 80% ethanol at 85-90˚C in a reflux condenser for 3 h. After being filtered through a 0.2 μm filter, the extract was concentrated and lyophilized by vacuum evaporation. The solid form of the extract (Figure 1c) was dissolved in DMSO prior to the experiment.
2.3. Identification and analysis of chemical constituents in EEPC using UHPLC-Q-Exactive-MS/MS
Chromatographic analysis was performed using a Thermo Vanquis UHPLC system (Thermo Fisher Scientific, USA). Chromatographic separation was carried on Agilent Zorbax Eclipse C18 (1.8 μm×2.1×100 mm) column by gradient elution with the optimal mobile phase of 0.1% formic acid aqueous solution (solvent A) and acetonitrile (solvent B); the column temperature was maintained at 35°C and the temperature of the sample chamber was set to 8°C. The gradient elution was set as follows: 0−5 min, 2% B; 5–20 min, 2%-98% B; 20–25 min, 98%–2% B; 25–30 min, 2% B. The flow rate was 0.30 mL·min-1 and the injection volume was 2 µL.
The mass spectrometer was operated with the heated electrospray ionization source in both positive and negative ion modes. The key parameters were as follows: spray voltage, +3.8 kV/-3.2 kV; sheath gas flow rate, 45 arbitrary units (Arbs); auxiliary gas flow rate, 15 Arbs; sweep gas flow rate, 1 Arbs; capillary temperature, 350 °C; auxiliary gas heater temperature, 300 °C. Scan modes, full MS with a resolution of 70,000 FWHM and data-dependent MS/MS with a resolution of 17,500 FWHM; stepped normalized collision energy, 20, 40 and 60 eV and scan range, m/z 75-1,050. Data acquisition and processing were carried out with Compound Discoverer 3.0, Thermo mzCloud and Thermo mzValut software (Thermo Scientific), respectively.
2.4. Cell Culture
Mouse neuroblastoma Neuro-2a cells were grown in MEM medium (Hyclone) supplemented with 10% heat-inactivated fetal bovine serum and 1% penicillin/streptomycin, and maintained at 37℃ in 5% CO2 humidified atmosphere. Cells were passaged every 3-4 days.
Primary NSPCs were isolated from E13.5 ICR mouse brain and cultured as previously described[17]. For NSPCs differentiation, single cells dissociated from neurospheres were seeded on poly-D-lysine (100 ng/mL) and laminin (20 μg/mL) coated coverslips at 2 × 104 cells/mL. Cells were incubated in DMEM/F12 medium supplemented with 10% FBS and 1% penicillin–streptomycin for 5 days to allow differentiation into multiple linages in the population.
2.5. Cytotoxicity Test
Cell viability was assessed by MTT (3,4,5-dimethylthiazol-2-yl)-2-5-diphenyltetrazolium bromide) reduction assay. For assay, cells (5×103) were plated in 96-well microtiter plates and grown for 24 h. Afterwards cells were treated with different concentrations of EEPC (5, 10, 20, 50 and 100 µg/mL). After 24 h incubation, the media containing EEPC were carefully removed and 100 µL of MTT solution (0.5 mg/mL in MEM) was added to each well and further incubated for four hours. 200 µL DMSO was added to each well to dissovled the formazon crystals, and the absorbance was measured by a microplate reader at 570 nm. Cell viability was shown relative to the control in a graph.
2.6. Western Blotting
Neuro-2a cells were seeded in 35 mm dish (6 × 104 /dish) overnight and then incubated with EEPC for different times (0−240 min) at 37 °C. Cells were lysed in RIPA buffer containing protease and phosphatase inhibitors and whole cell lysates were quantified using a BCA protein assay kit according to the manufacturer’s instructions. Those cell lysates were subsequently separated by SDS-PAGE and transferred to PVDF membranes. The membranes were probed with primary antibodies and then subsequently with secondary antibody, followed by electrochemiluminescence (ECL) detection.
2.7. Differentiation Assay
For neurite outgrowth assay, Neuro-2a cells were seeded into 24-well plates at a density of 1×104 cells /mL and grown for 24 h. After 24 h incubation, the culture medium was replaced with differentiation medium (MEM supplied with 0.5% FBS and 1% penicillin/streptomycin) containing different test compounds for 24 h. Neuro-2a cells were captured and counted under a phase contrast microscope. Neurites were defined as a protrusion with lengths longer than one diameter of the cell body. The neurite length of each cell was measured by Image J software.
2.8. Immunostaining
For immunostaining, cells were fixed in freshly prepared 4% PFA for 20 min and then permeabilized in PBS with 0.4% Triton X-100. Following blocked in PBS with 5% goat serum and 1% bovine serum albumin (BSA) for 20 min. The cells were incubated at 4 ℃ overnight with primary antibody, followed by incubation with Alexa Fluor-546 goat anti-mouse IgG or Alexa Fluor-488 goat anti-rabbit IgG as secondary antibodies for 1 h at room temperature. DAPI was added to visualize the nuclei. Images were taken using a fluorescence microscope (Olympus IX71, Japan).
2.9. Statistical Analysis
The results are expressed as the mean ±standard error of the mean (SEM). Data were subjected to student’s t test or one way analysis of variance (ANOVA) followed by Tukey’s test to assess the differences between the relevant control and each experimental group. A value of P < 0.05 was considered statistically significant.
3. Results
3.1. EEPC induces neuronal differentiation and neurite outgrowth in Neuro-2a cells
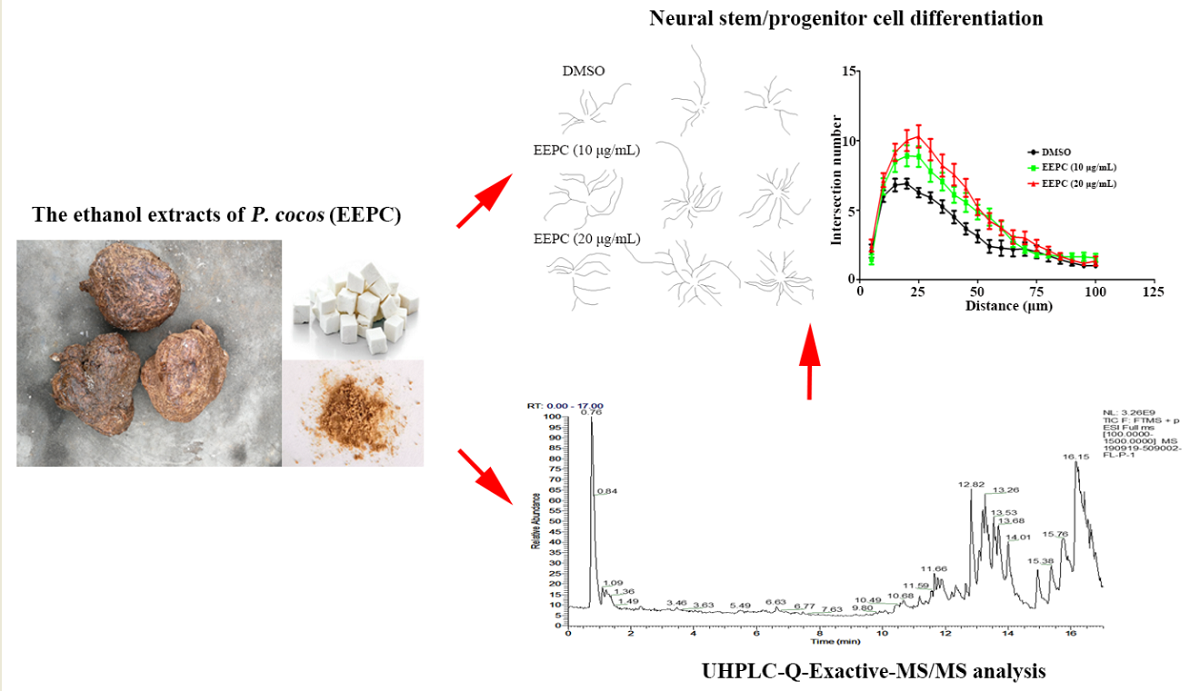
Our previous findings showed that the ability of EEPC in induced apoptosis and differentiation in Neuro-2a neuroblastoma cells[16]. To investigate the influences of EEPC on neuronal differentiation, Neuro-2a cells were treated with different concentrations of EEPC (10 and 20 μg/mL). Retinoic acid (RA, 10 μM) has been used as positive control. After 48h of differentiation, cells were labeled by immunostaining with an antibody against β-tubulin III to show the presence of neurite. As shown in Figure 2A, the untreated cells (DMSO) have a round shape with few neurites and the RA-treated cells apparently display long neurites. Here, we compared the effect of the differentiation of Neuro-2a cells induced by EEPC. Notably, all aspects we examined, including differentiation rate (Figure 2B) and the longest neurite length (Figure 2C); EEPC exhibited stronger activities as RA did. Moreover, EEPC promoted neuronal differentiation and neurite growth of Neuro-2a cells in a concentration-dependent manner.
3.2. EEPC activates JNK1/2/3 during EEPC-induced neuronal differentiation
Previously studies have demonstrated that the ERK1/2 and JNK1/2/3 signaling pathways are involved in neurite outgrowth[18]. To investigate whether ERK1/2 and JNK1/2/3 signaling pathways underlie EEPC-induced neuronal differentiation, a western blot assay was performed in a time-dependent manner using protein extracts of EEPC (20 μg/mL)-treated cells. As shown in Figure 3A-C, phosphorylation of ERK1/2 and JNK1/2/3 were significantly increased by EEPC treatment for 30-240 min. These results suggested that EEPC treatment promotes ERK1/2 and JNK1/2/3 phosphorylation in Neuro-2a cells and this may be the mechanism of EEPC-induced promotion of neurite outgrowth. To further determine whether activation of ERK1/2 and JNK1/2/3 signaling are required for EEPC promoted Neuro-2a cell differentiation and neurite outgrowth, we conducted a blocking study with ERK1/2 inhibitor (FR18024, 10 mM) and JNK 1/2/3 inhibitor (SP600125, 10 mM). As shown in Figure 2D-F, SP600125 but not FR18024 completely blocks the effect of EEPC on promoting Neuro-2a cell differentiation and neurite outgrowth. These results suggest that EEPC promotes neuronal differentiation in Neruo-2a cells through activation of JNK1/2/3 signaling pathway.
3.3. EEPC Promotes Neuronal Differentiation and Maturation of NSPCs
To reveal whether EEPC has abilities to promote neurogenesis in NSPCs, NSPCs isolated from E13.5 ICR mouse brain are multi-potent and can differentiate into neurons and astrocytes under differentiation conditions. EEPC treatment for 5 days enhanced neural induction, as we observed that the ratio of β-tubulin III positive cells (Neurons) was significantly increased from 71.6 ± 1.8% (DMSO) to 75.9 ± 1.3% (EEPC, 10 μg/mL) and 83.9 ± 1.3% (EEPC, 20 μg/mL), respectively (Figure 4A, B). This result suggested that EEPC could promote NSPC differentiation into neurons. Neuronal differentiation is a sequential multi-step process in which cells change from progenitor cells to premature neurons and finally into mature neurons. During neuron development, cell morphology changes dramatically, the neurites become more extensive, and the branch number of each neuron increases. To gain insights into the effects of EEPC during neuronal maturation, the total multiple-neurite neurons were measured for β-tubulin III positive cells. With EEPC treatment, the percentage of multiple neurite neurons (more than two branches) was significantly increased from 10.68 ± 1.0 % (DMSO) to 14.2 ± 1.2 % (EEPC, 10 μg/mL) and 14.7 ± 1.0 % (EEPC, 20 μg/mL), respectively (Figure 4C). Mature-like neurons had long and extensive dendrites. Furthermore, the percentage of mature-like neurons were counted. EEPC significantly increased the percentage of mature-like neurons from 23.6 ± 4.5 % (DMSO) to 40.2 ± 4.7 % (EEPC, 10 μg/mL) and 46.5 ± 5.1 % (EEPC, 20 μg/mL), respectively (Figure 4D). Finally, the effect of EEPC on dentritic complexity was assessed by Sholl analysis (Figure 4E). These results indicate that EEPC could help newborn neurons to form a more complicated neurite structure. Above all, we conclude that EEPC treatment promotes neuronal differentiation of NSPCs and developed greater morphological maturity.
3.4. Identification of the chemical constituents of EEPC extract
In order to find out the active substances of EEPC, the UHPLC-Q-Exactive-MS/MS technique was chosen for our study to determine chemical constituents of EEPC. The TIC of EEPC in positive and negative ion modes is portrayed in Figure 5A. A total of 97 compounds were identified from EEPC (Table S1), and these compounds were classified into 33 classes, including Carboxylic acids and derivatives (18.6%), Steroids and steroid derivatives (10.3%), Prenol lipids (7.2%), Organonitrogen compounds (6.2%), Organooxygen compounds (6.2%), Pyridines and derivatives (5.2%), Pyridines and derivatives (5.2%) and Benzene and substituted derivatives (5.2%) (Figure 5B). Among the compounds, the relative content of Choline (17.6 %) was the maximum, and that of (3β, 5ξ, 9ξ)-3,6,19-Trihydroxyurs-12-en-28-oic acid (9.7 %), Adenosine (7.2 %), L-(-)-Arabitol (6.5 %) and Ethyl palmitoleate (5.4 %), respectively (Table 1).
3.5. Choline is identified as the major effective component of EEPC
To determine which component was the active one, the neuronal differentiation-promoting activities of the 4 compounds were firstly investigated using Neuro-2a cells. The results showed Adenosine, L-(-)-Arabitol and Ethyl palmitoleate had weak effects on Neuro-2a cells differentiation; only Choline had dramatic neuronal differentiation-promoting effects in Neuro-2a cells (Figure 6A-C). Additionally, Choline also showed obvious neuronal differentiation-promoting effects in NPCs (Figure 6D-G). Therefore, our results showed that Choline was identified as the major compound with neuronal differentiation-promoting activity in EEPC.
4. Discussion
The discovery of adult brain stem cells and the development of adult neurogenesis research have brought hope for the ultimate treatment of neurodegenerative diseases[19-21]. From this perspective, finding drugs that can induce neurogenesis of adult NSPCs has become an effective therapeutic strategy for neurodegenerative diseases[22-24]. Recently, many herbal plant extracts and natural compounds were screened for promoting neurogenesis, with some of them being promising drug candidates[25-27]. In China, a number of herb ingredients known from Traditional Chinese Medicine (TCM) have a long history of use for improved learning and memory[28-30]. Therefore, these medicinal herbs provide efficient resources for drug discovery to promote neurogenesis. P. cocos is a well-known traditional medicinal fungus and dietary supplement. Previous studies demonstrated that P. cocos exhibited a series of neuropharmacological activities, including memory improvement[31], antidepressive[32] and neuroprotective[33]. However, the bioactive components and neuropharmacological mechanisms of P. cocos remain unclear. In this work, we explored the influence of the P. cocos on neurogenesis and unraveled the material basis for its action.
The Neuro-2a cell line has neuron-like properties that are commonly used as a model for the study of neuronal differentiation[34]. Here we demonstrated that the ethanol extracts of the P. cocos have neuronal differentiation-promoting activity in cultured Neuro-2a cell, leading to marked neurite outgrowth. Previous studies on the extracellular signal-regulated kinase (ERK) and c-jun N-terminal (JNK) are involved in the regulation of neurite outgrowth[18]. Interestingly, although EEPC markedly activated ERK1/2 at 30 min and thereafter, inhibition of ERK activity cannot offset the promotion of EEPC on neurite outgrowth. However, inhibition of JNK activity can offset the promotion of EEPC on neurite outgrowth. These results were consistent with a previous study which showed that JNK activation is required for neurite outgrowth in Neuro-2a cells. To confirm whether the effects of EEPC on neurogenesis are true, we examined the extract in primary cortical NSPCs cultures. Indeed, EEPC can not only induce the differentiation of NSPCs into neuronal cell types, but also promote the differentiation of newborn neurons into more mature type of cell morphology.
In order to obtain a better insight into the chemical constituents that could be contributing to the activity, a total of 97 metabolites were identified by UHPLC-Q-Exactive-MS/MS in EEPC (Table S1). The analysis of the relative proportion showed that there were 20 components (Table 1) accounting for 78.6% of the total amount. In addition, the neuronal differentiation-promoting activities of Choline (17.6%), Adenosine (7.2%), L-(-)-Arabitol (6.5%) and Ethyl palmitoleate (5.4%) were further confirmed both in Neuro-2a cells and NSPCs. Choline, which accounts for the largest proportion of EEPC, was proved to have significant neuronal differentiation promoting activity. For the possible active component, choline is an essential nutrient involved in one-carbon metabolism and is the precursor for many important compounds, including phospholipids, acetylcholine, and the methyl donor betaine[35]. Recent evidence suggests that choline is important during the perinatal period, especially for neural tube closure and brain development[36, 37]. It is interesting to note that dietary intake of choline by the pregnant rat and later by the infant directly affects the structure and function of hippocampal pyramida cells, including larger soma and increased numbers of primary and secondary basal dendritic branches[38]. Notably, these structural changes are often associated with memory function. In rodents, perinatal supplementation of choline enhances memory and learning functions, and these changes persist throughout life[39]. Conversely, choline deficiency during these sensitive periods leads to memory and cognitive deficits that also persist[40]. Maternal choline supplementation improves spatial learning and adult hippocampal neurogenesis in the Ts65Dn mouse model of Down syndrome[41]. Although the cellular mechanisms by which choline exerts these effects are unclear, numerous studies have implicated choline involves DNA methylation, altered gene expression, and associated changes in stem cell proliferation and differentiation[42, 43]. Therefore, choline was considered to be the main active component of P. cocos that promotes the neuronal differentiation of NSPCs.
In summary, we showed that EEPC treatment promotes neurogenesis in NSPCs in vitro. Based on these results, it is suggest that P.cocos has a positive effect on learning and memory functions and this might be due to its neurogenesis effect. However, in-depth mechanisms are required for more concrete information on the pharmacological features of P.cocos. To the best of our knowledge, this is the first investigation of the neurogenesis effect of P.cocos and the main biologically active substance is choline. Our findings provide an experimental basis for the application of P.cocos in the treatment of neurodegenerative diseases.
Acknowledgments
This work was financially supported by the Basic and Applied-Basic Research Project of Huaihua Science and Technology Bureau (2021R3132); The Scientific Research Foundation of Hunan Provincial Education Department (22A0549); The Hunan Provincial Natural Science Foundation (2020JJ5450); Hunan Innovation and Entrepreneurship Training project for College Students (201810548007), The Foundation of Hunan Double First-rate Discipline Construction Projects of Bioengineering (YYZW2018-5, YYZW2019-15, YYZW2019-32).
Conflicts of interest
The authors declare that they have no conflict interests.
References
- Gotz, M.; Huttner, W. B. , The cell biology of neurogenesis. Nature reviews. Molecular cell biology 2005, 6, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Mehler, M. F.; Gokhan, S. , Developmental mechanisms in the pathogenesis of neurodegenerative diseases. Progress in neurobiology 2001, 63, 337–363. [Google Scholar] [CrossRef] [PubMed]
- Farkas, L. M.; Huttner, W. B. , The cell biology of neural stem and progenitor cells and its significance for their proliferation versus differentiation during mammalian brain development. Current opinion in cell biology 2008, 20, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D. W.; Goodman, R. R.; Fraser, R. A.; Nedergaard, M.; Goldman, S. A. , Neural stem and progenitor cells: a strategy for gene therapy and brain repair. Neurosurgery 1998, 42, 858–867, discussion 867–868. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, J. W.; Shi, H. Y.; Ma, Y. M. , Neural stem cell therapy for brain disease. World journal of stem cells 2021, 13, 1278–1292. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, X. L.; Jiang, Z. M.; Li, X. F.; Qi, Y.; Yu, J.; Yang, X. X.; Zhang, M. , Quantification of Chemical Groups and Quantitative HPLC Fingerprint of Poria cocos (Schw.) Wolf. Molecules 2022, 27, 6383. [Google Scholar] [CrossRef]
- Song, Z.; Bi, K.; Luo, X.; Chan, K. , The isolation, identification and determination of dehydrotumulosic acid in Poria cocos. Analytical sciences : the international journal of the Japan Society for Analytical Chemistry 2002, 18, 529–531. [Google Scholar] [CrossRef]
- Rios, J. L.; Andujar, I.; Recio, M. C.; Giner, R. M. , Lanostanoids from fungi: a group of potential anticancer compounds. Journal of natural products 2012, 75, 2016–2044. [Google Scholar] [CrossRef]
- Rios, J. L. , Chemical constituents and pharmacological properties of Poria cocos. Planta medica 2011, 77, 681–691. [Google Scholar] [CrossRef]
- Jiang, Y. H.; Wang, L.; Chen, W. D.; Duan, Y. T.; Sun, M. J.; Huang, J. J.; Peng, D. Y.; Yu, N. J.; Wang, Y. Y.; Zhang, Y. , Poria cocos polysaccharide prevents alcohol-induced hepatic injury and inflammation by repressing oxidative stress and gut leakiness. Frontiers in nutrition 2022, 9, 963598. [Google Scholar] [CrossRef]
- Fang, C. L.; Paul, C. R.; Day, C. H.; Chang, R. L.; Kuo, C. H.; Ho, T. J.; Hsieh, D. J.; Viswanadha, V. P.; Kuo, W. W.; Huang, C. Y. , Poria cocos (Fuling) targets TGFbeta/Smad7 associated collagen accumulation and enhances Nrf2-antioxidant mechanism to exert anti-skin aging effects in human dermal fibroblasts. Environmental toxicology 2021, 36, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, S. L.; Gong, L.; Wang, J. L.; Li, Y. Z.; Wu, Z. H. , The effects of an herbal medicine Bu-Wang-San on learning and memory of ovariectomized female rat. Journal of ethnopharmacology 2008, 117, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Duan, X.; Cheng, X.; Cheng, X.; Li, X.; Zhang, L.; Liu, P.; Su, S.; Duan, J. A.; Dong, T. T.; Tsim, K. W.; Huang, F. , Kai-Xin-San, a standardized traditional Chinese medicine formula, up-regulates the expressions of synaptic proteins on hippocampus of chronic mild stress induced depressive rats and primary cultured rat hippocampal neuron. Journal of ethnopharmacology 2016, 193, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. P.; Feng, L.; Zhang, M. H.; Ma, D. Y.; Wang, S. Y.; Gu, J.; Fu, Q.; Qu, R.; Ma, S. P. , Neuroprotective effect of Liuwei Dihuang decoction on cognition deficits of diabetic encephalopathy in streptozotocin-induced diabetic rat. Journal of ethnopharmacology 2013, 150, 371–381. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Z. J.; Wang, X. H.; Ma, J.; Song, Y. H.; Liang, M.; Lin, S. X.; Zhao, J.; Zhang, A. Z.; Li, F.; Hua, Q. , The prescriptions from Shenghui soup enhanced neurite growth and GAP-43 expression level in PC12 cells. BMC complementary and alternative medicine 2016, 16, 369. [Google Scholar] [CrossRef]
- Xia Jiang, Jie Xie, Zhaotun Hu, Xiaoliang Xiang, Ethanol extract of Poria cocos induces apoptosis and differentiation in Neuro-2a neuroblastoma cells. Bangladesh Journal of Pharmacology 2022, 17, 9. [CrossRef]
- Xiang, X.; Zhuang, X.; Li, S.; Shi, L. , Arhgef1 is expressed in cortical neural progenitor cells and regulates neurite outgrowth of newly differentiated neurons. Neuroscience letters 2017, 638, 27–34. [Google Scholar] [CrossRef]
- Waetzig, V.; Herdegen, T. , The concerted signaling of ERK1/2 and JNKs is essential for PC12 cell neuritogenesis and converges at the level of target proteins. Molecular and cellular neurosciences 2003, 24, 238–249. [Google Scholar] [CrossRef]
- Stepien, T. , Neurogenesis in neurodegenerative diseases in the adult human brain. Postepy psychiatrii neurologii 2021, 30, 287–292. [Google Scholar]
- Peretto, P.; Bonfanti, L. , Adult neurogenesis 20 years later: physiological function vs. brain repair. Frontiers in neuroscience 2015, 9, 71. [Google Scholar] [CrossRef]
- Conti, L.; Cataudella, T.; Cattaneo, E. , Neural stem cells: a pharmacological tool for brain diseases? Pharmacological research 2003, 47, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Chang, K. A.; Kim, J. A.; Kim, S.; Joo, Y.; Shin, K. Y.; Kim, S.; Kim, H. S.; Suh, Y. H. , Therapeutic potentials of neural stem cells treated with fluoxetine in Alzheimer's disease. Neurochemistry international 2012, 61, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Grochowski, C.; Radzikowska, E.; Maciejewski, R. , Neural stem cell therapy-Brief review. Clinical neurology and neurosurgery 2018, 173, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J. D.; Ming, G. L.; Song, H. , Adult neurogenesis as a potential therapy for neurodegenerative diseases. Discovery medicine 2006, 6, 144–147. [Google Scholar]
- Deb, S.; Phukan, B. C.; Dutta, A.; Paul, R.; Bhattacharya, P.; Manivasagam, T.; Thenmozhi, A. J.; Babu, C. S.; Essa, M. M.; Borah, A. , Natural Products and Their Therapeutic Effect on Autism Spectrum Disorder. Advances in neurobiology 2020, 24, 601–614. [Google Scholar]
- Liu, J. W.; Tian, S. J.; de Barry, J.; Luu, B. , Panaxadiol glycosides that induce neuronal differentiation in neurosphere stem cells. Journal of natural products 2007, 70, 1329–1334. [Google Scholar] [CrossRef]
- Park, H. R.; Kim, J. Y.; Lee, Y.; Chun, H. J.; Choi, Y. W.; Shin, H. K.; Choi, B. T.; Kim, C. M.; Lee, J. , PMC-12, a traditional herbal medicine, enhances learning memory and hippocampal neurogenesis in mice. Neuroscience letters 2016, 617, 254–263. [Google Scholar] [CrossRef]
- Ren, H.; Gao, S.; Wang, S.; Wang, J.; Cheng, Y.; Wang, Y.; Wang, Y. , Effects of Dangshen Yuanzhi Powder on learning ability and gut microflora in rats with memory disorder. Journal of ethnopharmacology 2022, 296, 115410. [Google Scholar] [CrossRef]
- Sun, C.; Liu, J.; Li, N.; Liu, M.; Luo, Z.; Li, H. , Traditional Chinese Medicine Shenmayizhi Decoction Ameliorates Memory and Cognitive Impairment Induced by Multiple Cerebral Infarctions. Evidence-based complementary and alternative medicine : eCAM 2021, 2021, 6648455. [Google Scholar] [CrossRef]
- Sun, H.; Hu, Y.; Zhang, J. M.; Li, S. Y.; He, W. , [Effects of one Chinese herbs on improving cognitive function and memory of Alzheimer's disease mouse models]. Zhongguo Zhong yao za zhi = Zhongguo zhongyao zazhi = China journal of Chinese materia medica 2003, 28, 751–754. [Google Scholar]
- Wu, K. J.; Chen, Y. F.; Tsai, H. Y.; Wu, C. R.; Wood, W. G. , Guizhi-Fuling-Wan, a Traditional Chinese Herbal Medicine, Ameliorates Memory Deficits and Neuronal Apoptosis in the Streptozotocin-Induced Hyperglycemic Rodents via the Decrease of Bax/Bcl2 Ratio and Caspase-3 Expression. Evidence-based complementary and alternative medicine : eCAM 2012, 2012, 656150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, L.; Li, P.; Zhao, J.; Duan, J. , Antidepressant and immunosuppressive activities of two polysaccharides from Poria cocos (Schw.) Wolf. International journal of biological macromolecules 2018, 120 Pt B, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Di, X.; Bian, B.; Li, K.; Guo, J. , Neuroprotective Effects of Poria cocos (Agaricomycetes) Essential Oil on Abeta1-40-Induced Learning and Memory Deficit in Rats. International journal of medicinal mushrooms 2022, 24, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Tang, G.; Yang, J.; Ding, J.; Lin, H.; Xiang, X. , Synthesis of some new acylhydrazone compounds containing the 1,2,4-triazole structure and their neuritogenic activities in Neuro-2a cells. RSC advances 2020, 10, 18927–18935. [Google Scholar] [CrossRef]
- Hollenbeck, C. B. , An introduction to the nutrition and metabolism of choline. Central nervous system agents in medicinal chemistry 2012, 12, 100–113. [Google Scholar] [CrossRef]
- Derbyshire, E.; Obeid, R. , Choline, Neurological Development and Brain Function: A Systematic Review Focusing on the First 1000 Days. Nutrients 2020, 12, 1731. [Google Scholar] [CrossRef]
- Caudill, M. A. , Pre- and postnatal health: evidence of increased choline needs. Journal of the American Dietetic Association 2010, 110, 1198–1206. [Google Scholar] [CrossRef]
- Bastian, T. W.; von Hohenberg, W. C.; Kaus, O. R.; Lanier, L. M.; Georgieff, M. K. , Choline Supplementation Partially Restores Dendrite Structural Complexity in Developing Iron-Deficient Mouse Hippocampal Neurons. The Journal of nutrition 2022, 152, 747–757. [Google Scholar] [CrossRef]
- Tees, R. C. , The influences of rearing environment and neonatal choline dietary supplementation on spatial learning and memory in adult rats. Behavioural brain research 1999, 105, 173–188. [Google Scholar] [CrossRef]
- Mudd, A. T.; Getty, C. M.; Sutton, B. P.; Dilger, R. N. , Perinatal choline deficiency delays brain development and alters metabolite concentrations in the young pig. Nutritional neuroscience 2016, 19, 425–433. [Google Scholar] [CrossRef]
- Velazquez, R.; Ash, J. A.; Powers, B. E.; Kelley, C. M.; Strawderman, M.; Luscher, Z. I.; Ginsberg, S. D.; Mufson, E. J.; Strupp, B. J. , Maternal choline supplementation improves spatial learning and adult hippocampal neurogenesis in the Ts65Dn mouse model of Down syndrome. Neurobiology of disease 2013, 58, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Blusztajn, J. K.; Mellott, T. J. , Choline nutrition programs brain development via DNA and histone methylation. Central nervous system agents in medicinal chemistry 2012, 12, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Nagakura, T.; Uchino, H.; Inazu, M.; Yamanaka, T. , Functional Expression of Choline Transporters in Human Neural Stem Cells and Its Link to Cell Proliferation, Cell Viability, and Neurite Outgrowth. Cells 2021, 10, 453. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Poria cocos (Schw.) Wolf (a). The dried sclerotia of Poria cocos (Schw.) Wolf (b). The ethanol extract of Poria cocos (Schw.) Wolf (c).
Figure 1.
Poria cocos (Schw.) Wolf (a). The dried sclerotia of Poria cocos (Schw.) Wolf (b). The ethanol extract of Poria cocos (Schw.) Wolf (c).

Figure 2.
Effect of EEPC on the differentiation of Neuro-2a cells. (A) Neuro-2a cells were treated with EEPC at different concentrations from 1 to 20 μg/mL for 48 h. RA, retinoic acid as a positive control. Scale bar, 50 mm. The differentiation rate (B) and the longest neurite length of each differentiated cell (C) were analyzed. At least 300 cells/group were analyzed in each experiment, One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). ***P < 0.001.
Figure 2.
Effect of EEPC on the differentiation of Neuro-2a cells. (A) Neuro-2a cells were treated with EEPC at different concentrations from 1 to 20 μg/mL for 48 h. RA, retinoic acid as a positive control. Scale bar, 50 mm. The differentiation rate (B) and the longest neurite length of each differentiated cell (C) were analyzed. At least 300 cells/group were analyzed in each experiment, One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). ***P < 0.001.
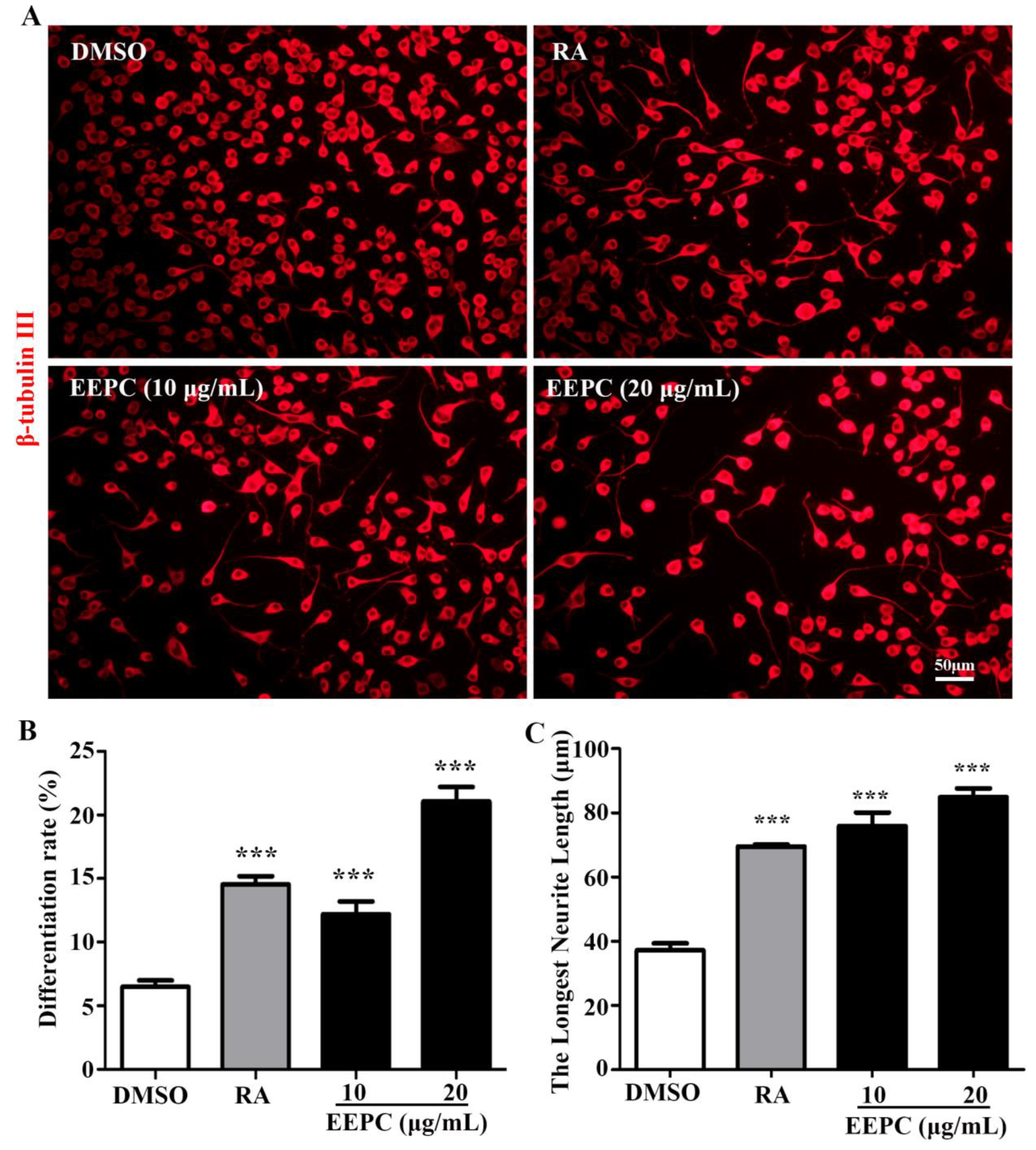
Figure 3.
Activation of JNK1/2/3 signaling pathway is required for EEPC induced Neuro-2a cells differentiation. EEPC (20 μg/mL) was added in Neuro-2a cells for indicated time points (0–240 min). Cells lysates were subjected to western blot analysis for the phosphorylated (p-ERK1/2 or p-JNK1/2/3) and total (ERK1/2 or JNK1/2/3) forms of different signaling proteins (A-C). Neuro-2a cells were pretreated with different inhibitors, including ERK inhibitor (FR18024, 10 μM) or JNK inhibitor (SP600125, 10 μM) for 1 h, followed by EEPC treatment (20 μg/mL) for 48 h. Neurites were visualized by inverted phase contrast microscope (D). Cell differentiation rate (E) and the longest length of neurites per differentiated cell (F) were quantified. At least 300 cells/group were analyzed in each experiment (n= 3), One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). **P < 0.01, ***P < 0.001, DMSO or inhibitor and EEPC cotreatment vs. EEPC single treatment.
Figure 3.
Activation of JNK1/2/3 signaling pathway is required for EEPC induced Neuro-2a cells differentiation. EEPC (20 μg/mL) was added in Neuro-2a cells for indicated time points (0–240 min). Cells lysates were subjected to western blot analysis for the phosphorylated (p-ERK1/2 or p-JNK1/2/3) and total (ERK1/2 or JNK1/2/3) forms of different signaling proteins (A-C). Neuro-2a cells were pretreated with different inhibitors, including ERK inhibitor (FR18024, 10 μM) or JNK inhibitor (SP600125, 10 μM) for 1 h, followed by EEPC treatment (20 μg/mL) for 48 h. Neurites were visualized by inverted phase contrast microscope (D). Cell differentiation rate (E) and the longest length of neurites per differentiated cell (F) were quantified. At least 300 cells/group were analyzed in each experiment (n= 3), One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). **P < 0.01, ***P < 0.001, DMSO or inhibitor and EEPC cotreatment vs. EEPC single treatment.
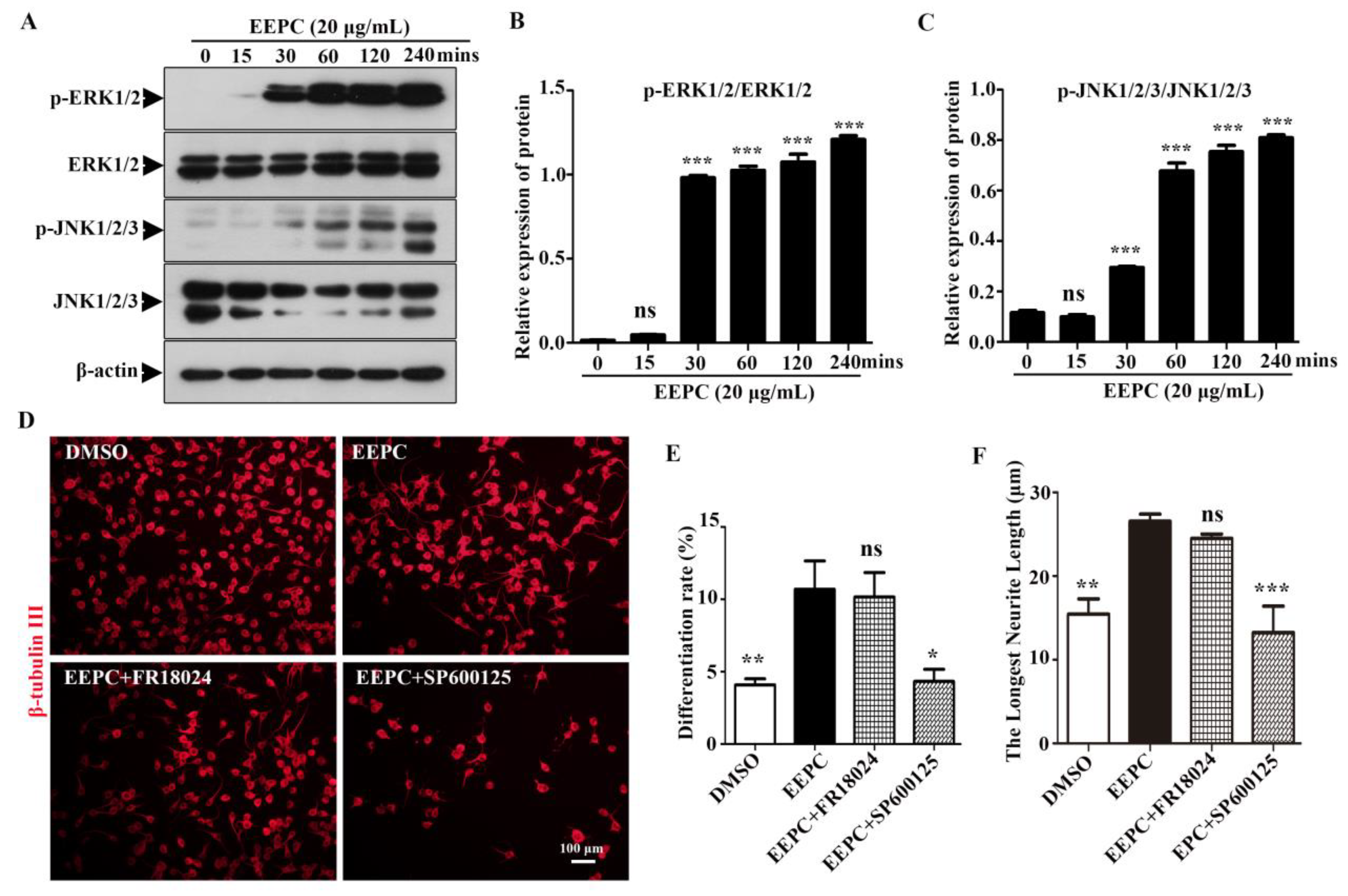
Figure 4.
Newborn neurons derived from neural stem cells (NSCs) in the presence of EEPC developed more mature morphology. (A) Newborn neurons were visualized by immunostanining for β-tubulin III. Nuclei were stained with DAPI (blue). Scale bars, 50 μm. The percentage of neurons with multiple neurites (B), average neurite length (C) and mature-like neurons (D) were measured. **P < 0.05, **P < 0.01, ***p <0.001. One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). (E) EEPC influenced the morphology of mature-like neurons. (F) Numbers of dendritic intersections at 0–200 μm from the cell bodies were accessed by Sholl analysis.
Figure 4.
Newborn neurons derived from neural stem cells (NSCs) in the presence of EEPC developed more mature morphology. (A) Newborn neurons were visualized by immunostanining for β-tubulin III. Nuclei were stained with DAPI (blue). Scale bars, 50 μm. The percentage of neurons with multiple neurites (B), average neurite length (C) and mature-like neurons (D) were measured. **P < 0.05, **P < 0.01, ***p <0.001. One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3). (E) EEPC influenced the morphology of mature-like neurons. (F) Numbers of dendritic intersections at 0–200 μm from the cell bodies were accessed by Sholl analysis.
Figure 5.
UHPLC-Q-Exactive-MS/MS analysis of the chemical constituents in the EEPC. (A) Positive ion mode (a) and Negative ion mode (b). (B) Classification of chemical constituents.
Figure 5.
UHPLC-Q-Exactive-MS/MS analysis of the chemical constituents in the EEPC. (A) Positive ion mode (a) and Negative ion mode (b). (B) Classification of chemical constituents.
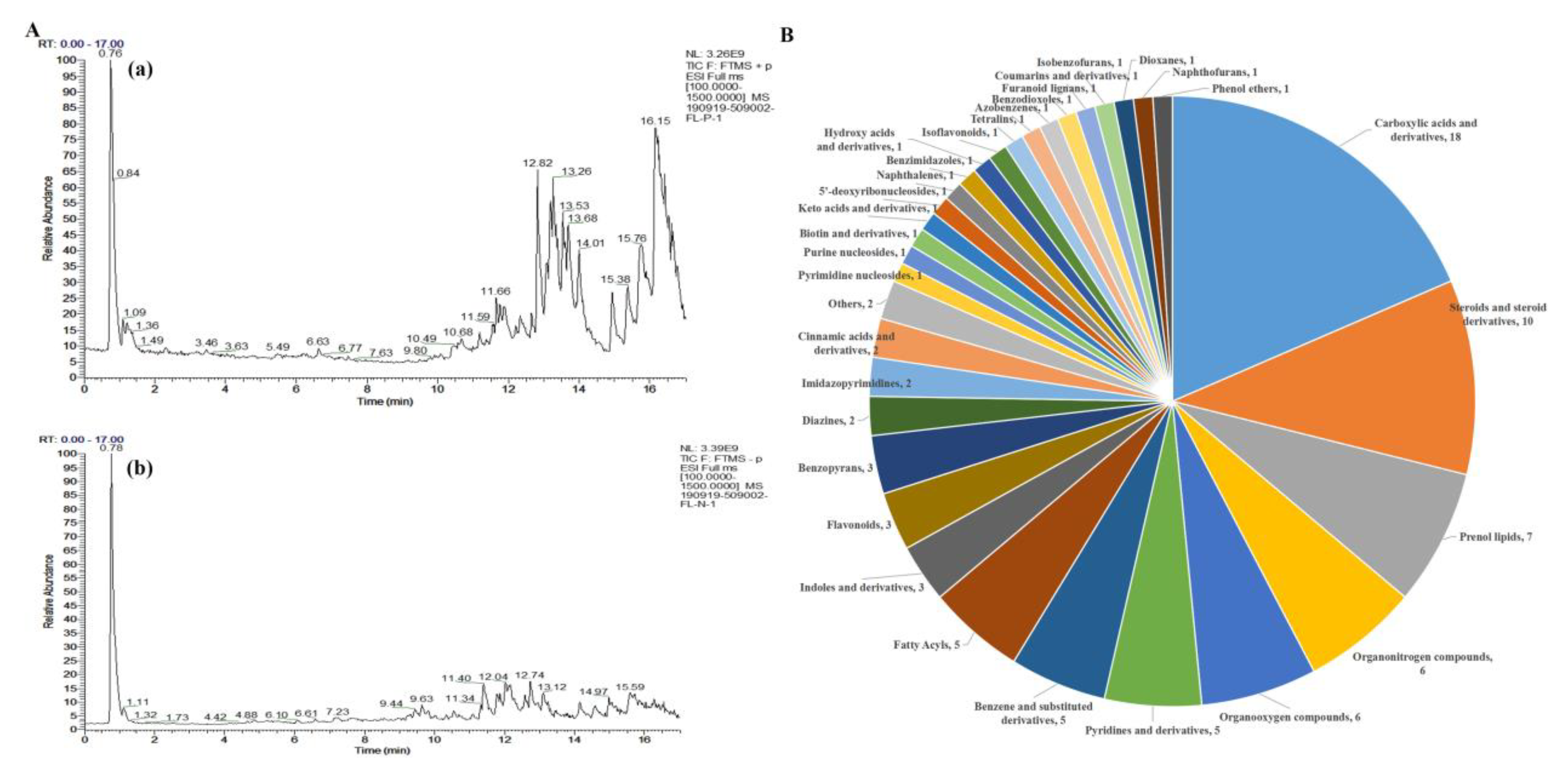
Figure 6.
Choline was identified as the main active component of EEPC. (A) Neuro-2a cells were treated with Ethyl palmitoleate (3.8 μM), L-(-)-Arabitol (8.6 μM), Choline (34.4 μM) and Adenosine (5.4 μM) for 48 h. Scale bar, 50 mm. The differentiation rate (B) and the longest neurite length of each differentiated cell (C) were analyzed. (D) NPCs were treated with Choline (34.4 μM), neurons were visualized by immunostanining for β-tubulin III. Nuclei were stained with DAPI (blue). Scale bars, 50 μm. The percentage of neurons with multiple neurites (E), average neurite length (F) and mature-like neurons (G) were measured. **P < 0.05, **P < 0.01, ***p <0.001. One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3).
Figure 6.
Choline was identified as the main active component of EEPC. (A) Neuro-2a cells were treated with Ethyl palmitoleate (3.8 μM), L-(-)-Arabitol (8.6 μM), Choline (34.4 μM) and Adenosine (5.4 μM) for 48 h. Scale bar, 50 mm. The differentiation rate (B) and the longest neurite length of each differentiated cell (C) were analyzed. (D) NPCs were treated with Choline (34.4 μM), neurons were visualized by immunostanining for β-tubulin III. Nuclei were stained with DAPI (blue). Scale bars, 50 μm. The percentage of neurons with multiple neurites (E), average neurite length (F) and mature-like neurons (G) were measured. **P < 0.05, **P < 0.01, ***p <0.001. One-way ANOVA followed by Dunnett’s test. Error bars represent SEM (n = 3).
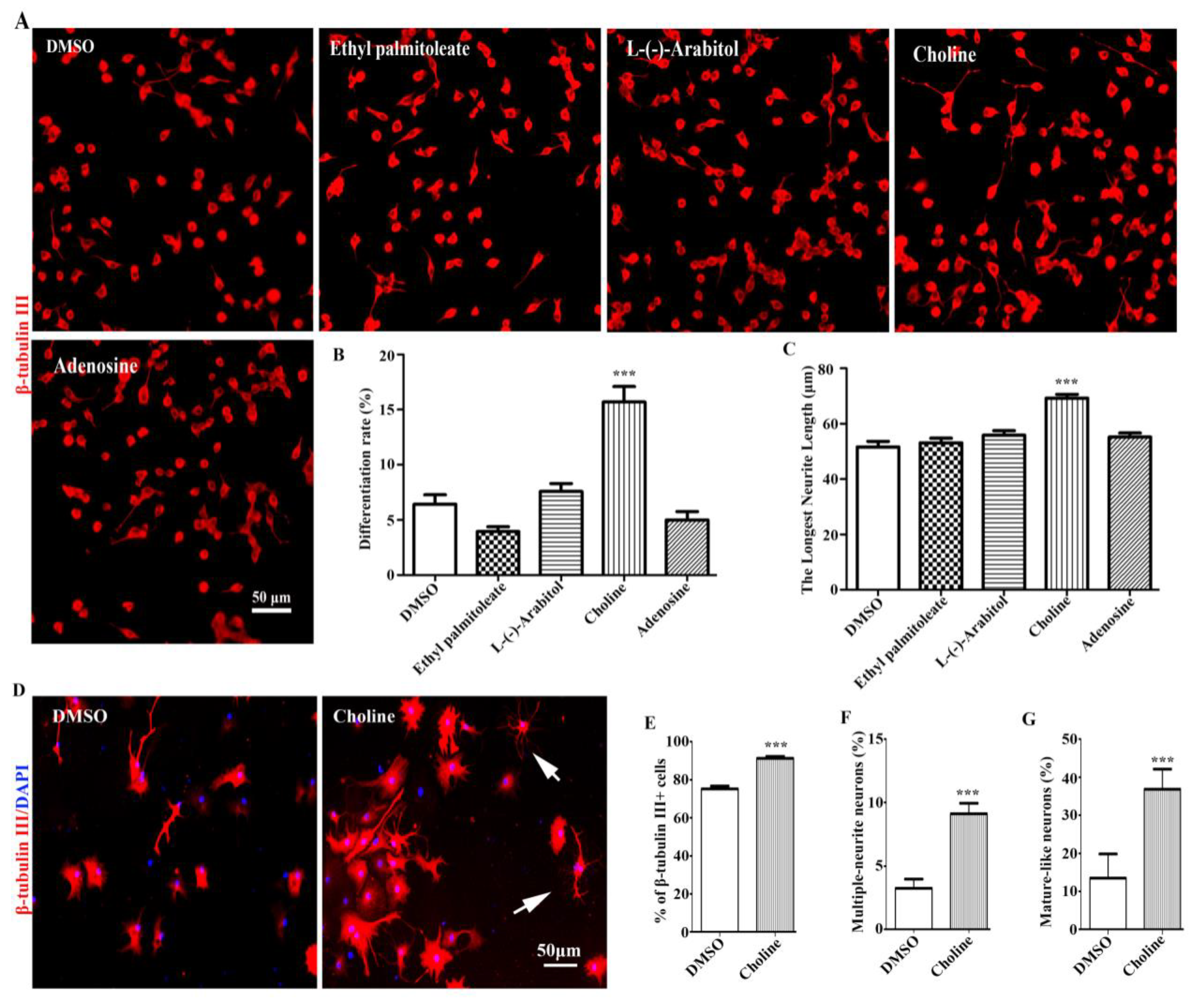

Table 1.
Identification of chemical constituents of EEPC by UHPLC-MS/MS (Relative proportion≥ 1%.).
| NO. | Name | Formula | Class | M.W. | RT [min] | Relative proportion (1%) |
|---|---|---|---|---|---|---|
| 1 | Choline | C5 H13NO | Organonitrogen compounds | 103.09982 | 0.769 | 17.75 |
| 2 | L-Glutamic acid | C5H9NO4 | Carboxylic acids and derivatives | 147.05274 | 0.79 | 1.14 |
| 3 | N,N-Diethylethanolamine | C6H15NO | Organonitrogen compounds | 117.11529 | 0.797 | 2.20 |
| 4 | Betaine | C5H11NO2 | Carboxylic acids and derivatives | 117.0789 | 0.809 | 1.82 |
| 5 | Trigonelline | C7H7NO2 | Pyridines and derivatives | 137.04733 | 0.813 | 2.46 |
| 6 | L-(-)-Arabitol | C5H12O5 | Organooxygen compounds | 152.06812 | 0.82 | 6.51 |
| 7 | Cytosine | C4H5N3O | Diazines | 111.0433 | 0.837 | 1.81 |
| 8 | Cytidine | C9H13N3O5 | Pyrimidine nucleosides | 243.0849 | 0.84 | 1.01 |
| 9 | Adenosine | C10H13 N5O4 | Purine nucleosides | 267.09604 | 1.243 | 7.23 |
| 10 | Acetophenone | C8H8O | Organooxygen compounds | 120.05739 | 1.321 | 1.14 |
| 11 | Leucine | C6H13NO2 | Carboxylic acids and derivatives | 131.0944 | 1.36 | 2.53 |
| 12 | L-Phenylalanine | C9H11NO2 | Carboxylic acids and derivatives | 165.07866 | 2.292 | 1.50 |
| 13 | Phenylacetylene | C8H6 | Benzene and substituted derivatives | 102.0469 | 2.292 | 1.46 |
| 14 | Picolinic acid | C6H5NO2 | Pyridines and derivatives | 123.03191 | 3.231 | 1.02 |
| 15 | Triphenylphosphine oxide | C18H15OP | Benzene and substituted derivatives | 278.0851 | 10.705 | 3.17 |
| 16 | 2-Amino-1,3,4-octadecanetriol | C18H39NO3 | Organonitrogen compounds | 317.29191 | 11.185 | 3.17 |
| 17 | (3β,5ξ,9ξ)-3,6,19-Trihydroxyurs-12-en-28-oic acid | C30H48O5 | Prenol lipids | 488.34864 | 11.697 | 9.67 |
| 18 | Bis(4-ethylbenzylidene)sorbitol | C24H30O6 | Dioxanes | 414.20291 | 11.918 | 2.50 |
| 19 | 18-β-Glycyrrhetinic acid | C30H46O4 | Prenol lipids | 470.33821 | 13.044 | 1.05 |
| 20 | Ethyl palmitoleate | C18H34O2 | Fatty Acyls | 282.25502 | 14.152 | 5.42 |
| 21 | 3-Acetyl-11-keto-β-boswellic acid | C32H48O5 | Prenol lipids | 512.34893 | 14.93 | 2.90 |
| 22 | 4-Methoxycinnamic acid | C10H10O3 | Cinnamic acids and derivatives | 178.06257 | 16.726 | 1.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



