
Submitted:
07 September 2023
Posted:
11 September 2023
You are already at the latest version
Abstract
Anthocyanins (ACNs) have attracted considerable attention for their potential to modulate the immune system. Research has revealed their antioxidant and anti-inflammatory properties, which play a crucial role in immune regulation by influencing key immune cells such as lymphocytes, macrophages, and dendritic cells. Moreover, ACNs contribute to maintaining a balance between proinflammatory and anti-inflammatory cytokines, thus promoting immune health. Beyond their direct effects on immune cells, ACNs significantly impact gut health and the microbiota, essential factors in immune regulation. Emerging evidence suggests that they positively influence the composition of the gut microbiome that enhance the immunomodulatory effects of ACNs. Furthermore, these compounds synergize with other bioactive substances, such as vitamins and minerals, further enhancing their potential as immune-supporting dietary supplements. However, detailed clinical studies must fully validate these findings and determine safe dosages for different populations. Incorporating these natural compounds into functional foods or supplements can revolutionize the management of immune-related conditions. Personalized nutrition and healthcare strategies can be developed to enhance overall well-being and immune resilience by fully understanding the mechanisms underlying their actions. Recent advancements in delivery methods have focused on improving the bioavailability and effectiveness of ACNs, providing promising avenues for future applications.
Keywords:
Anthocyanins
; immune cells
; gut microbiota
; bioavailability
; efficacy
; nano-delivery methods.
1. Introduction
The immune system is vital in safeguarding the body against diseases and maintaining its physiological functions. Any disturbance in the immune system can result in various health problems, such as autoimmune diseases, inflammatory disorders, cancer, etc. A debilitated immune system can cause infections and tumors [1], while an overactive one may lead to autoimmune conditions like type I diabetes, systemic lupus erythematosus, and rheumatoid arthritis [2,3]. The immune response can be categorized into two types: innate and adaptive. Innate immunity comprises pre-existing responses triggered without prior exposure to an antigen and inherent in individuals from birth [4]. Conversely, adaptive immunity includes humoral and cell-mediated immunity, activated only after exposure to a specific antigen [5]. Immune cells, such as lymphocytes, dendritic cells, monocytes/macrophages, natural killer cells, CD4+and CD8+ T-cells, and myeloid-derived suppressor cells, play crucial roles in regulating innate and adaptive immunity through their distinct structures and functions [6]. In addition to these immune cells, various inflammatory cytokines and chemokines, including interleukins (ILs), tumor necrosis factor-alpha (TNF-α), transforming growth factor beta (TGF-β), and interferon-gamma (IFN-γ), are triggered and regulated [7,8,9]. Signaling pathways such as nuclear factor kappa B (NF-κB), programmed cell death protein 1 (PD-1), cytotoxic T-lymphocyte–associated antigen 4 (CTLA-4), mitogen-activated protein kinases (MAPKs), extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), Janus kinase/signal transducer and activator of transcription (JAK/STAT) and nucleotide-binding, oligomerization domain (NOD)-like receptors (NLRs) also participate in immunomodulation [10,11,12]. Modifying the immune system means enhancing, inhibiting, activating, amplifying, or expressing various elements or processes of the immune response. This modulation ultimately leads to an improvement in the immune response.
Amidst the global COVID-19 or coronavirus pandemic, it is evident that markets worldwide have become inundated with a plethora of products claiming to be "immunity boosters." These offerings emerge alongside various speculative cures, treatments, and preventive strategies. However, it is crucial to recognize that the notion of an "immunity booster" is scientifically misleading and frequently exploited to promote unverified products and therapies [13,14]. The "immunity boosters" market comprises vitamins, minerals, antioxidants, probiotics, functional foods, nutraceuticals, and other complementary and alternative medicines. In a study conducted using data from the US National Health and Nutrition Examination Survey, more than 50% of the US population acknowledged using supplements [15]. This widespread usage has significant economic implications, with the global dietary supplements market estimated to be approximately 133.1 billion USD and projected to accelerate at a CAGR of 9.6% from 2016 to 2024 [16]. It is crucial to underscore that promoting "immunity boosters" can mislead consumers and give rise to false hopes. The scientific community consistently emphasizes the importance of evidence-based practices and rigorous research to substantiate the claims of any product or therapy. Unfortunately, many offerings flooding the market lack the necessary scientific backing to validate their effectiveness.
Natural products have been extensively studied for their potential immunomodulatory properties [17]. These natural products can modulate the immune system by enhancing or suppressing the immune response. They can also promote the production of cytokines and other immune system molecules, leading to an overall improvement in immune function. One of their key functions is to stimulate non-specific innate immune responses, which involve immune system mediators like innate leukocytes (natural killer cells, eosinophils, basophils, and mast cells) and phagocytic cells (neutrophils, macrophages, and dendritic cells) to defend against pathogens [18]. By enhancing innate immunity, these products facilitate effector innate immune responses such as immune cell infiltration, phagocytosis, and cytotoxic mechanisms like natural killer cell-mediated cytotoxicity, ultimately destroying tumor cells [19]. Certain natural compounds also act as immunomodulatory agents, activating the adaptive immune system. In adaptive immunity, T- and B-lymphocytes recognize tumor antigens through cell-surface antigen-specific receptors, leading to an augmented humoral response and the elimination of tumor cells through various mechanisms, including T-cell mediated cancer cell death [20].
Insufficient nutrition, malnutrition or deficiencies in specific nutrients can disrupt the functioning of the immune system, leaving the body vulnerable to diseases. To maintain a robust immune system, consuming a well-balanced diet that provides all the necessary nutrients in appropriate quantities [21]. Functional foods and nutraceuticals offer an alternative approach to boosting immune function and supporting the management of diverse diseases. Several studies have explored the potential of plant-based nutraceuticals as immunomodulating agents, owing to their wide range of effects that can positively impact the immune system [22,23,24]. These substances are often better tolerated than conventional pharmaceutical treatments, making them an attractive supplement for enhancing immune system function [21]. Among them, ACN, naturally occurring water-soluble flavonoids, has been shown to stimulate immunomodulatory and antioxidant effects. This, in turn, helps to reduce the harmful cooperative and synergistic effects of oxidative stress and proinflammatory cytokines. Consequently, ACN may protect against chronic diseases [25,26]. This review provides a comprehensive update on the current understanding of the scientific foundation behind immunomodulatory properties and the clinical significance of ACNs, thereby highlighting their potential as dietary supplements/ nutraceuticals in the rapidly expanding global food industry.
2. Chemistry and Natural Sources of ACNs
ACNs are glucosides of anthocyanidins and are predominantly found as 3-glucosides. The primary configuration of their parent nucleus consists of a strongly conjugated 2-phenyl benzopyran cation. Three carbon atoms connect the benzene rings to form a C6–C3–C6 skeleton, the ACN motif [27,28]. These ACNs can be classified into sugar-free anthocyanidin aglycones and glycosides. ACNs include over 700 different derivatives of 27 aglycons [29]. These compounds are formed via the phenylpropanoid pathway (Figure 1) from anthocyanidins, which serve as their precursors [30].
The chemical structures of flavylium cation and ACNs are given in Figure 2. The aglycon form of anthocyanidins is rarely observed due to their inherent instability and reactivity, primarily caused by the electron-deficient flavylium cation. Conversely, ACNs are commonly observed in a glycosylated state, as this modification enhances their stability and solubility [31]. They are typically composed of one of six anthocyanidin bases, which differ in their molecular structure at the B-ring and are attached to a sugar moiety at the third position of the C-ring. Approximately 90% of all known anthocyanidins found in plants consist of six bases, namely pelargonidin, cyanidin, peonidin, delphinidin, petunidin, and malvidin [32,33], with the glycosides of the nonmethylated anthocyanidins (cyanidin, delphinidin, and pelargonidin) being the most abundant natural ACNs, accounting for up to 60% of the total content [34].
The color of ACNs becomes bluer as the number of hydroxyl groups in the B-ring increases—conversely, methylation results in a red shift in the color of ACNs. Moreover, the color is also pH dependent i.e., blue in basic condition while red in acidic condition (where ACNs are positively charged). Methylation of the B-ring offers improved resistance to oxidation and helps stabilize the ACNs. Methyl-modified flavonoids are commonly present on the surfaces of leaves and flowers [35]. On the other hand, glycosylation of ACNs causes a hypsochromic shift in the absorption maxima of the spectra and enhances their stability during storage in vacuoles [36,37]. The sugar molecules, such as glucose, galactose, rhamnose, or arabinose, are primarily attached to the aglycone to form derivatives of 3-glycosides or 3,5-diglycosides [25]. The sugar moieties of acylated ACNs, usually attached to the hydroxyl group in C-3 and C-5 of the aglycone, have a covalent ester linkage to one or more aliphatic (acetic, malonic, oxalic, and succinic) or aromatic (caffeic, coumaric, ferulic, hydroxybenzoic, and sinapic) acids [38,39]. The acylation of glycosyl moieties in ACNs alters their chemical properties, enhancing stability. Aliphatic acylation does not affect color, while aromatic acylation causes a blue shift. As a result, acylated ACNs are more suitable for use as natural colorants and bioactive components in innovative functional foods and nutraceuticals. This modification improves stability and expands their potential applications [37,39,40,41].
These fruits are rich in ACNs, which are pigments responsible for their vibrant colors. Berries like blackcurrants, blackberries, blueberries, cranberries, and vegetables like black carrots, red cabbage, and purple potato are well-known sources of ACNs [42,43]. For instance, pistachios contain significant amounts of cyanidin 3-O-galactoside, while blackcurrants contain delphinidin 3-O-rutinoside and cyanidin 3-O-rutinoside. On the other hand, red wine, elderberries, and pomegranate juice are known to contain malvidin 3-O-glucoside, cyanidin 3-O-glucoside, and cyanidin 3,5-O-diglucoside, respectively [44,45]. These compounds exhibit strong antioxidant properties and have the potential to function as preventive bioactive molecules against various illnesses. Additionally, various flowers, particularly those with red, purple, and blue shades, are used in traditional medicine and consumed as food. These flowers, such as red clover, red rose, red hibiscus, red pineapple sage, pink blossom, blue rosemary, blue chicory, cornflower, purple passion flower, purple mint, common violet, purple sage, and lavender, are also rich in ACNs [34]. Acylated ACNs with varying structures are also present in fruits, berries, vegetables, and tubers. Some rich sources of these compounds are purple sweet potato, red radish, purple carrot, and red cabbage [46]. Within the pigmented members of the Solanaceae family, such as potatoes, peppers, tomatoes, and eggplants, acylated ACNs are identified with the structure of anthocyanidin-3-hydroxycinnamoyl-rutinoside-5-glucoside, with delphinidin being the primary anthocyanidin, excluding pigmented potatoes [39,47].
3. Immunomodulatory Potential of ACNs
Over the years, researchers have explored various avenues to improve immune function and address immune-related disorders. Recently, there has been growing interest in the potential of natural compounds to influence the immune response. ACNs, documented in both in vitro and in vivo studies, have been found to possess diverse health-promoting properties. In addition to their notable antioxidant property, ACNs play a crucial role in modulating the immune system. They protect the immune system through the following cellular processes:
- Inflammation disrupts immune homeostasis, leading to diverse diseases or disease conditions. However, ACNs have been proven to play a crucial role in regulating inflammatory pathways by inhibiting the production of proinflammatory cytokines (IL-1β, IL-6, and TNF-α) and proinflammatory mediators (COX, LOX, MPO, and PGE2), thereby offering protection against the development of various inflammatory conditions [29,32,50].
- ACNs influence immune cell activation and proliferation by modulating various pathways. Moreover, they have a significant impact on gene expression within immune cells, leading to heightened expression of genes responsible for antioxidant defense, anti-inflammatory pathways, and immune cell activation [29,51,52].
- Recent research indicates that ACNs can influence gut microbiota composition and functionality. Given the pivotal role of the gut microbiota in regulating the immune system, this interaction has the potential to contribute significantly to the immunomodulatory effects exhibited by ACNs [53,54,55,56,57].
ACNs, along with other polyphenols such as flavones, flavone-3-ols, and flavanones have shown potential in promoting a balanced T helper cell type 1 (Th1)/Th2 response and reducing the production of allergen-specific immunoglobulin (Ig) E antibodies [58]. Additionally, ACNs can activate gamma-delta (γδ) T cells, a type of immune cell involved in both acquired and innate immunity, by mimicking pathogen-associated molecular patterns [59]. This interaction with γδ T cells enhances the activity of essential immune components, such as natural killer cells, cytokines, and lymphocytes, which are crucial in defending against pathogens that can enter the body through the digestive and respiratory systems. Cyanidin 3-O-glucoside inhibits osteoclast differentiation and formation in a dose-dependent manner by downregulating osteoclast differentiation marker genes and suppressing the activation of specific kinases. It specifically targets osteoclasts without affecting osteoblasts and promotes osteoblast differentiation and matrix formation [60]. Protocatechuic acid (PCA), a major metabolite of ACNs, dose-dependently inhibited the differentiation of osteoclasts and suppressed their bone-resorbing activity by targeting the JNK signaling pathway and down-regulating osteoclastogenesis-related genes. Moreover, PCA administration effectively restored bone loss in mice induced by lipopolysaccharide, suggesting its potential therapeutic application for inflammatory bone disorders [61].
Delphinidin increased cytosolic-free Ca2+ concentration by releasing Ca2+ from intracellular stores and enhancing Ca2+ entry through the putative Ca2+ release-activated Ca2+ (CRAC) channel. This led to the activation of the nuclear factor of activated T cells (NFAT) and subsequent stimulation of cytokine production, particularly IL-2 and IFN-γ, indicating its immunostimulatory effects on T cells through the CRAC channel and NFAT pathway [62]. The ortho-dihydroxy phenyl structure of anthocyanidins on the B-ring is linked to their inhibitory actions. Anthocyanidins with an ortho-dihydroxy phenyl structure on the B-ring exhibited inhibitory effects on cell transformation and activator protein-1 transactivation induced by 12-O-tetradecanoylphorbol-13-acetate (TPA). Delphinidin, but not peonidin, specifically blocked the phosphorylation of protein kinases in the ERK and JNK pathways, while p38 kinase was unaffected. Moreover, specific inhibitors of JNK and ERK, but not p38, could block cell transformation, indicating that anthocyanidins inhibit tumorigenesis by targeting the MAPK pathway [63]. Delphinidin, the most potent inhibitor, suppressed cyclooxygenase (COX)-2 expression by blocking MAPK-mediated pathways, including NF-κB, activator protein-1 (AP-1), and CCAAT/enhancer binding protein-delta (C/EBPδ) activation. This provides molecular evidence for the anti-inflammatory properties of anthocyanidins with an ortho-dihydroxyphenyl structure by inhibiting MAPK-mediated COX-2 expression [64].
In a recent study, researchers investigated the immune-modulatory effects of Sambucus ebulus L. fruit infusion, which contains significant amounts of cyanidin 3-O-galactoside (Cy3Gal, 48.15 mg/g dry weight) and cyanidin 3-sambubioside (43.41 mg/g dry weight). The study involved healthy volunteers, with fifty-three participants who underwent a 4-week intervention. The intervention results were promising, as a noteworthy reduction in various markers of inflammation and complement activity was observed. Specifically, levels of total protein decreased by 2.82%, IL-6 by 20.15%, TNFα by 5.38%, IL-8 by 5.50%, C3 by 4.16%, and C4 by 14.29%. Interestingly, hemoglobin and hematocrit levels decreased in the entire group, particularly among women. This indicates that consuming S. ebulus fruits could enhance the functioning of the immune system [65]. Another study investigated the anti-inflammatory properties of a specific fraction of black rice, rich in cyanidin 3-O-glucoside and peonidin 3-O-glucoside. The researchers found that this fraction effectively suppressed the expression of inflammatory genes (NLRP3, IL-1β, and IL-18) and the secretion of cytokines (IL-6, IL-1β, and IL-18) induced by the spike glycoprotein S1 subunit of SARS-CoV-2 in A549 and THP-1 cells. Moreover, the black rice fraction inhibited the activation of NF-κB and downregulated inflammasome-dependent inflammatory pathway proteins (NLRP3, ASC, and caspase-1). These results indicate that ACNs from black rice could be utilized in preventive strategies against the long-term effects of COVID-19 infection [66].
Cyanidin 3-O-glucoside (C3G) was delivered using enteric sodium alginate to assess its anti-food allergy effects in vivo. The targeted rectal and colonic delivery of C3G proved highly effective in reducing allergic symptoms, diarrhea, and serological markers. Additionally, C3G improved the intestinal barrier, increased secretory IgA and β-defensin, and regulated the Th1/Th2 immune balance. Beneficial bacteria like Lactobacillus and Odoribacter thrived, while pathogenic bacteria like Helicobacter and Turicibacter decreased [67]. A study investigated ACN-rich roselle extract (ARRE) from Hibiscus sabdariffa in broiler chickens—supplemented diets with ARRE increased antioxidant capacity, lysozyme levels, antimicrobial enzymes, and anti-inflammatory cytokine IL-10. The ARRE at 200 and 400 mg/kg body weight doses showed higher levels of serum complement 3, indicating enhanced immune response [68]. Furthermore, consuming C3G provided lasting cardioprotection against ischemia/reperfusion injury, independent of its anti-inflammatory properties. Immune-deficient mice experienced similar protection, suggesting other mechanisms at play. The C3G-enriched diet altered the gut microbiome, and transferring fecal microbiota from C3G-fed mice replicated the cardioprotective effects [69].
A research study explored the potential effects of C3G on Alzheimer's disease (AD) and other conditions by analyzing gene expression patterns in the spleen. The investigation involved wild-type mice (C57BL/6J Jms), mice with an Alzheimer's disease model (APPswe/PS1dE9 mice), and mice with the Alzheimer's model treated with C3G. By comparing the gene expression profiles, the study identified specific genes related to antioxidants, immune responses, and AD pathways that were differentially expressed in the C3G-treated group. Notably, six crucial antioxidant genes (S100a8, S100a9, Prdx2, Hp, Mpst, and Prxl2a) and various immune-related genes were upregulated in the treated mice. These findings support the potential of C3G to have immunomodulatory effects [70]. Another investigation focused on the effects of a boysenberry and apple juice concentrate rich in cyanidin glycosides (including cyanidin 3-O-sophoroside, cyanidin 3-O-glucoside, and cyanidin 3-O-[2-glucosylrutinoside]) on acute lung inflammation and M2 macrophage-associated cytokines in a male C57BL/6J mouse model of allergic airways disease. The mice received oral administration of the concentrate (equivalent to 0.2 mg/kg in humans; 2.5 mg/kg total ACN content) prior to an intranasal ovalbumin challenge. Consumption of the concentrate led to significant reductions in eosinophil, neutrophil, and T-cell infiltration in the lung and mucous production. Furthermore, gene expression analysis revealed increased expression of anti-inflammatory macrophage markers (Arg1) and specific cytokines (CXCL10 and CCL4) associated with an anti-inflammatory response [71]. In SARS-CoV-2 viral replication, the papain-like (PLpro) and main proteases (Mpro) are crucial in processing viral replicase polypeptides. The PLpro protease also possesses deubiquitinating activity, impacting important signaling pathways, inhibiting the interferon response, and antagonism innate immune responses. Interestingly, C3G demonstrated concentration-dependent inhibition of the SARS-CoV-2 PLpro in an in vitro enzymatic inhibition assay within a micromolar range [72,73].
Effects of C3G and hydrochlorothiazide (HCT) on T-cell function in spontaneously hypertensive rats (SHR) were investigated. Male SHR and Wistar-Kyoto (WKY) rats were given water, C3G (10 mg/kg per day), HCT (10 mg/kg per day), or a combination of both for 15 weeks. SHR showed lower proportions of specific helper T-cells (CD62Llo, CD62L-, and CD25+) than WKY. C3G increased specific T-cell proportions (CD62Llo and CD62L-) in SHR, while HCT had mixed effects (higher CD62Lhi and CD62Llo and lower CD62L-). C3G increased TNF-α and IFN-γ concentrations, while HCT decreased them. This suggests that C3G positively affects T-cell function, while HCT further suppresses it in SHR [74]. In rheumatoid arthritis (RA), fibroblast-like synoviocytes (FLS) contribute to synovitis, chronic inflammation, and joint damage. Cyanidin effectively inhibited IL-17A-induced migration and proliferation of FLS cells in rats with adjuvant-induced arthritis (AA). Cyanidin also reduced the overexpression of IL-17 receptor A (IL-17RA) and downregulated IL-17A-dependent factors. Additionally, cyanidin modulated JAK/STAT-3 signaling and activated the PIAS3 protein, suppressing STAT-3-specific transcriptional activation. In AA rats, cyanidin treatment alleviated clinical symptoms, synovial pannus growth, immune cell infiltration, and bone erosion. It also reduced serum levels of IL-23 and GM-CSF and decreased the level of p-STAT-3 protein in the synovial tissue [52]. In a study on bovine type II collagen-induced arthritis (CIA) rats, C3G was administered via tail vein injections. C3G reduced CD38+ cell proportion, suppressed rheumatoid arthritis synovial fibroblast (RASF) proliferation, and decreased proinflammatory cytokine secretion. C3G also increased regulatory T (Treg) cell proportion and upregulated Sirtuin 6 (SIRT6) expression, suppressing natural killer group 2D (NKG2D) expression in CD38+ NK cells. This elevated TNF-α secretion and decreased IFN-γ secretion in CD38+ NK cells, promoting mononuclear cells differentiation into Treg cells. Inhibition of Treg cell differentiation by CD38+ NK cells may contribute to the immune imbalance observed in RA and CIA [75].
Delphinidin 3-O-glucoside (D3G) and delphinidin (DC) were found to inhibit human colorectal cancer cells (HCT-116 and HT-29) at concentrations ranging from 100 to 600 µg/mL. The IC50 values for D3G were 329 µg/mL (HCT-116) and >600 µg/mL (HT-29), while for DC, they were 242 µg/mL (HCT-116) and >600 µg/mL (HT-29). C3G, D3G, DC, and D3G-rich extracts also reduced PD-L1 protein expression in HCT-116 cells. C3G specifically decreased PD-L1 fluorescence intensity by 39%. Moreover, C3G reduced PD-1 expression in peripheral blood mononuclear cells by 41% in monoculture and by 39% and 26% in co-culture with HCT-116 and HT-29 cells, respectively. D3G also reduced PD-1 expression by 50% and 51% in co-culture with HCT-116 and HT-29 cells, respectively [76]. CAN supplementation, specifically glycosides of delphinidin and cyanidin, alleviated the negative effects of a high-fat diet (HFD) in rats. The HFD caused intestinal permeabilization, endotoxemia, reduced tight junction protein expression, increased nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and nitric oxide synthase (NOS)-2 expression, oxidative stress, and activation of redox-sensitive signals. ACN supplementation increased glucagon-like peptide (GLP)-2 levels, preventing permeabilization and oxidative stress and activating signaling pathways in cell cultures. In HFD-induced obese mice, ACN restored gut microbiota composition and MUC2 levels, protective mucin in the intestinal barrier, and immune response [77]. An ACN-rich Lonicera caerulea L. fruit extract containing C3G (387.60 mg/g), cyanidin 3-rutinoside (23.62 mg/g), peonidin 3-glucoside (22.20 mg/g), and cyanidin 3,5-diglucoside (8.16 mg/g) showed significant inhibition of SMMC-7721 cell growth at concentrations of 0.1-0.8 mg/mL. The extract arrested the cell cycle in the G2/M phase, causing DNA damage and apoptosis. In H22 tumor-bearing mice, the extract at doses of 50, 100, and 200 mg/kg bw/day for 15 days enhanced antioxidant activity, reduced lipid peroxidation levels, and regulated immune cytokines, including IL-2, IFN-γ, and TNF-α. These findings demonstrate the potent anti-tumor effect of the ACN-rich L. caerulea fruit extract by maintaining redox balance and enhancing immunoregulatory activity [78].
The protective effects of C3G against TNF-α-stimulated intestinal cell-induced endothelial cell activation were investigated in an in vitro co-culture system. TNF-α induced NF-κB translocation and increased TNF-α and IL-8 gene expression in Caco-2 cells, which were reduced by C3G pre-treatment. TNF-α-stimulated Caco-2 cells activated endothelial cells, increasing E-selectin and vascular cell adhesion molecule (VCAM)-1 mRNA levels, leukocyte adhesion, and NF-κB levels in HUVECs. C3G inhibited these effects by selectively inhibiting the NF-κB pathway in epithelial cells. These findings suggest that ACNs, like C3G, may hold promise for managing chronic gut inflammatory diseases [79]. The ethanol extract (25-400 µg/mL) of phenolic-rich elderberry (Sambucus nigra L. subsp. canadensis) and specific ACNs, such as cyanidin chloride and C3G (6.25-25 μM), showed potent dose-dependent inhibition of lipopolysaccharide or IFNγ-induced reactive oxygen species (ROS) production, indicating strong anti-inflammatory and antioxidant properties [80]. C3G (1.25-5 µg/mL) suppressed the production of Th2 cytokines IL-4 and IL-13 in phorbol myristic acetate/ionomycin-activated EL-4 T cells, potentially inhibiting allergic reactions. This suppression occurred at the transcriptional level by downregulating the GATA-3 transcription factor, suggesting that C3G could be a promising anti-allergic agent targeting Th2 activation [81].
The ethanol extract from black raspberries (100 and 200 µg/mL) and its metabolites, cyanidin 3-rutinoside (10-200 μM) and quercetin-3-rutinoside (10-200 μM) exhibited immunomodulatory effects by inhibiting T-cell proliferation, limiting myeloid-derived suppressor cell expansion, and attenuating IL-6-mediated STAT3 signaling. These findings suggest that black raspberries and their components could be developed as targeted immunotherapies or drugs to inhibit specific STAT-regulated signaling pathways [82]. Novel strategies are sought to prevent relapse in adult patients with Philadelphia chromosome-positive acute lymphoblastic leukemia (Ph+ ALL). ACN-rich berry extract and imatinib effectively inhibited leukemia cell proliferation more than 6-mercaptopurine. However, the most promising results were observed with DNA vaccination and the berry extract, leading to higher survival rates and reduced relapse rates. This suggests its potential to enhance leukemia-specific immunity and be a maintenance therapy for other malignancies [83]. Two polyphenol-rich juices were investigated in a randomized crossover study involving healthy men on a low-polyphenol diet. These juices contained cyanidin glycosides (236 mg) and epigallocatechin gallate (226 mg) and showed positive effects, including improved antioxidant status, reduced oxidative DNA damage, and enhanced immune cell functions. Interestingly, there was a time delay between juice intake and the observed reductions in oxidative DNA damage and increases in interleukin-2 secretion [84].
Proanthocyanidins in lingonberry enhance anti-inflammatory macrophage (M2) activation and inhibit proinflammatory M1 activation in mouse and human macrophages. They also promote STAT6 phosphorylation, indicating their potential for anti-inflammatory effects, making them a beneficial addition to a healthy diet [85]. D3G has been found to stimulate mesenchymal stem cell (MSC) proliferation and enhance the production of anti-inflammatory cytokines, such as IL-10 and TGF-β, while reducing NF-κB expression. Conditioned media from MSCs treated with D3G inhibits macrophage metabolism and reduces the production of proinflammatory cytokines (IL-1β, IL-12, and TNF-α) while increasing IL-10, demonstrating its potential to modify immunoregulatory properties and promote IL-10 production by macrophages [86]. In a study using a C57BL/6 mice model treated with azoxymethane (AOM) and dextran sodium sulfate (DSS) for 11 weeks, researchers compared the effects of black lentil (BL) water with delphinidin 3-O-(2-O-β-d-glucopyranosyl-α-l-arabinopyranoside) (D3G)-rich lentil extract (41 mg/kg body weight) on tumor development, inflammation, and immune response. The AOM/DSS + BL group displayed a lower disease activity index and fewer neoplasms than the AOM/DSS control group. Additionally, proinflammatory cytokines like IL-1β and IL-6 were downregulated in the colon mucosa of the AOM/DSS + BL group. Both BL and D3G-rich extracts exhibited anti-inflammatory and pro-immune effects, while BL also prevented neoplasia growth [87].
ACN supplementation (glycosides of cyanidin and delphinidin) was found to combat obesity, dyslipidemia, insulin resistance, and steatosis in mice on a high-fat diet (HFD) for 14 weeks. The HFD caused intestinal permeabilization, endotoxemia, reduced expression of tight junction proteins (occludin, ZO-1, and claudin-1) in the ileum, increased NADPH oxidase (NOX1 and NOX4), NOS2, oxidative stress, and activation of redox-sensitive signals (NF-κB and ERK1/2). ACN supplementation reversed these effects, raised GLP-2 levels, prevented permeabilization in vitro, and restored gut microbiota composition and MUC2 levels (protective mucin). In vitro experiments showed that ACNs prevented Caco-2 monolayer permeabilization induced by TNF-α. ACNs also inhibited upregulation of NOX1/4, oxidative stress, and activation of NF-κB and ERK in inflammatory processes [76]. Delphinidin chloride and its degradation product, gallic acid, promoted the differentiation of Tregs and enhanced immune response control. Induced Tregs secreted more immunosuppressive proteins and demonstrated increased functionality. In an allograft model, treatment with these compounds reduced activated T cells by promoting Treg differentiation. Delphinidin and gallic acid are promising candidates for treating immune cell activation-related diseases, including autoimmune conditions, and could be relevant for immune system-associated food products [88]. Delphinidin inhibited the proliferation and differentiation of specific T-lymphocytes (Th1, Th17, and Treg) without affecting Th2 subsets. The inhibition of T-cell activity was attributed to its ability to inhibit calcium signaling, histone deacetylase (HDAC), NFAT activations, and ERK1/2 activation through the estrogen receptor (ER)-α pathway [89].
Delphinidin has shown promising results in both in vitro and in vivo experiments, effectively inhibiting proliferation, activation of pathway components, and secretion of proinflammatory cytokines/chemokines, ultimately improving skin conditions in a mouse model of psoriasis. These findings highlight the potential of delphinidin as a modulator of the phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway for psoriasis treatment [90]. In prostate cancer, the combination of delphinidin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) has been found to activate caspase pathways, relying on the activation of death receptor 5 (DR5) and cleavage of HDAC3. This exciting finding suggests that delphinidin could be used for prostate cancer chemoprevention by enhancing TRAIL-induced apoptosis [91]. Delphinidin effectively suppresses inflammatory signaling by blocking the acetylation of p65, leading to the accumulation of p65 in the cytosol and the nuclear localization of IKBα. This inhibitory effect prevents the expression of genes regulated by NF-κB and the production of proinflammatory cytokines, making delphinidin a potential therapeutic candidate for preventing inflammatory arthritis [92].
Resveratrol and peonidin 3-O-glucoside (Peo3G)-rich red grape vine leaf extract exhibit preventive properties against DNA entry into macrophages, leading to the inhibition of apoptosis-associated speck-like protein containing a caspase activation and recruitment domain (CARD) (ASC) oligomerization, caspase-1 activation, and the secretion of IL-1β and IL-18. In a mouse model of psoriasis induced by imiquimod, the extract reduces psoriasis-related inflammation, including caspase-1 activation, IL-1β maturation, IL-17 production, and overall disease severity [93]. Another ACN, malvidin 3-O-glucoside (M3G), has been investigated for its therapeutic potential in reducing inflammasome-induced inflammation. The research shows that M3G targets various types of inflammasomes, such as nucleotide-binding domain, leucine-rich–containing family, pyrin domain–containing-3 (NLRP3), NLR family CARD domain containing 4 (NLRC4), and absent in melanoma 2 (AIM2), resulting in decreased levels of caspase-1 and IL-1β proteins in microglia and the brain. Additionally, M3G exhibits beneficial effects in alleviating anxiety and depression symptoms and mitigating bacterial-mediated inflammation and stress-induced inflammasome-mediated innate responses [94]. Furthermore, M3G promotes resilience by modulating brain synaptic plasticity and peripheral inflammation. In a mouse model of increased systemic inflammation caused by transplanting stress-susceptible hematopoietic progenitor cells, M3G, in combination with deep hypothermic circulatory arrest (DHCA), reduces depression-like symptoms. This reduction is achieved by enhancing histone acetylation of the regulatory sequences of the Rac1 gene, thereby modulating synaptic plasticity [95].
4. Gut Microbiota in Immunity and Metabolism of ACNs
Gut health has become a global concern due to its significant impact on human well-being, often called “the second brain” [96]. The human body houses a substantial number of bacteria, estimated to be 3.9×1013 in the entire body of a 70 kg “reference man” [97]. The gut microbiota includes Bacteroides, Clostridium, Prevotella, Eubacterium, Ruminococcus, Fusobacterium, Peptococcus, and Bifidobacterium and a smaller presence of Escherichia and Lactobacillus [98,99]. These microbiota plays a vital role in the innate and adaptive immune system, while the immune system ensures a symbiotic relationship with the microbiota [100]. The gut microbiota plays a significant role in metabolizing ACNs into phenolic acids and aldehyde metabolites, primarily through specific bacteria in the large intestine. These metabolites can influence various physiological processes [101,102,103]. Conversely, ACNs contribute to the enhancement of gut health by modulating the function of the gut barrier and promoting the colonization of beneficial bacterial species. This dynamic interplay leads to heightened protection against pathogens, optimized nutrient metabolism, and an overall reinforcement of the immune response [57].
A significant portion of dietary ACNs remain unabsorbed and reaches the colon without absorption during digestion, interacting with gut microbiota and undergoing degradation, potentially increasing their bioavailability through bacterial or chemical processes. Notably, certain bacteria in the colon, such as Bacteroides spp., Enterococcus casseliflavus, Eubacterium spp., and Clostridium spp., play a key role in this process by producing enzymes like β-glucosidase, α-galactosidase, and α-mannosidase that break down ACNs [104,105]. In the colon, unabsorbed ACNs are hydrolyzed by microbial enzymes capable of cleaving sugar linkages and releasing ACN aglycones [106,107], which then further metabolized into phloroglucinol aldehyde (e.g., 2,4,6-trohydroxybenzaldehyde, 3,4-dihydroxy benzaldehyde) and phenolic acids (PAs) derived from phloroglucinol (A ring) and benzoic acids (B ring) [106]. Research findings have elucidated distinct metabolic pathways associated with the fermentation of various anthocyanidins (aglycones), leading to diverse phenolic compounds. For example, the fermentation of cyanidin-3-O-rutinoside and C3G yields PCA, p-coumaric acid, and phloroglucinol aldehyde [108]. Similarly, delphinidin-3-O-rutinoside undergoes transformations resulting in gallic acid, syringic acid, and phloroglucinol aldehyde [108]. In the case of M3G, it is converted to syringic acid [109], while pelargonidin-3-O-glucoside (Pel3G) is metabolized into 4-hydroxybenzoic acid [110]. The initial conversion of petunidin yields 3-O-methyl gallic acid, and subsequent O-demethylation leads to the production of gallic acid [110]. Furthermore, peonidin undergoes sequential transformations, first into vanillic acid (3-methoxy-4-hydroxybenzoic acid) and then into PCA [109]. In red radish, acylated ACNs, such as pelargonidin-3-sophorosid-5-glucoside acylated with ferulic acid and malonic acid, undergo degradation, resulting in the formation of 4-hydroxybenzoic acid, p-coumaric acid, ferulic acid, and caffeic acid [111].
A high-fat, high-sucrose diet-induced metabolic and inflammatory changes in mice, resulting in decreased levels of beneficial gut bacteria (Bacteroidetes and Muribaculaceae) and propionate. However, supplementation with Saskatoon berry powder, C3G, or PCA mitigated these effects and increased fecal Muribaculaceae and propionate [112]. Another study using an ApcMin/+ mouse model compared the effectiveness of PCA and black raspberries (BRB) dietary administration in preventing colorectal cancer. Both 5% BRBs and 500 ppm PCA-supplemented diets significantly reduced adenoma development in the small intestine and colon. They promoted a shift to anti-inflammatory bacterial profiles, suggesting potential benefits for colorectal cancer patients. Interestingly, 500 ppm PCA increased IFN-γ and SMAD4 (mothers against decapentaplegic homolog 4) in primary cultured human natural killer cells. In comparison, 1,000 ppm PCA did not show the same effect [113]. Furthermore, a study investigated the long-lasting cardioprotective effect of dietary ACNs by examining gene expression, histology, and resistance to ischemia/reperfusion in mice hearts after a month of ceasing the C3G-enriched diet. The results indicated that the cardioprotective benefits persisted, independent of immune responses, and were attributed to changes in the gut microbiota induced by the C3G-enriched diet [69].
C3G also alleviated polystyrene (PS)-induced toxicities in Caco-2 cells and Caenorhabditis elegans by promoting autophagy and discharge. In C57BL/6 mice, C3G supplementation reduced tissue accumulation, promoted fecal PS discharge, alleviated oxidative stress and inflammation caused by PS, and modulated PS-associated gut microbiome perturbations, indicating its potential protective role against PS-induced toxicity and gut dysbiosis [114]. In a study investigating the potential of purple sweet potato ACN extract in modulating the gut microbiota in a dextran sulfate sodium (DSS)-induced chronic colitis mouse model, extract treatment prevented the loss of beneficial bacteria (Bifidobacterium and Lactobacillus), inhibited the increase of harmful bacteria (Gammaproteobacteria and Helicobacter), and maintained colonic tight junction protein expression and architecture, resulting in attenuated intestinal inflammation. These findings suggest the extract could be an effective and safe treatment for chronic colitis [115]. Combining bilberry ACN with chitosan and low molecular citrus pectin was proposed to enhance digestive stability and modulate the microbiome for programmed cell death-ligand 1 (PD-L1) blockade treatment. This combo enriches subdominant species, increases butyrate concentration, enhances intra-tumoral CD8+ T cell infiltrations, and restores gut microbiome diversity, improving tumor growth control. These findings indicate potential opportunities for probiotic combos to enhance the therapeutic efficiency of immune checkpoint inhibitors through gut microbiome manipulation [116].
Bilberry ACN supplementation attenuated western diet-induced serum aspartate aminotransferase, alanine transaminase, low-density lipoprotein cholesterol levels, liver fat content, thiobarbituric acid reactive substances, and alpha-smooth muscle actin. Additionally, it modified the gut microbiota by reducing the Firmicutes/Bacteroidetes ratio and increasing the relative abundance of Akkermansia spp. and Parabacteroides spp., suggesting its potential to ameliorate nonalcoholic fatty liver disease by addressing dyslipidemia and gut microbiome dysbiosis [117]. M3G ingestion improved histopathological scores, enhanced IL-10 expression, promoted beneficial microbial interactions, and reduced pathogenic bacteria abundance and inflammatory mediators, demonstrating its unique mechanism of action on the gut microbiome compared to whole blueberry in DSS-induced colitis mouse model [118]. C3G has shown the potential to support gut integrity and overall health by producing phenolic compounds that combat gut inflammation and oxidative stress. These metabolites activate nuclear factor erythroid 2-related factor 2 (Nrf2) and influence TGFβ-activated kinase 1 (TAK1)-mediated mitogen-activated protein kinase (MAKP), and sphingosine kinase (SphK)/sphingosine-1-phosphate(S1P)-mediated NF-κB pathways, collectively reducing inflammation and creating an optimal environment for gut metabolism [119].
In a study investigating the effects of six berries, each containing different ACN profiles, on metabolic risk factors in a mouse model of polygenic obesity, the biological activities varied based on their delphinidin/malvidin-versus cyanidin-type ACNs, which were influenced by their structure and metabolism in the gut. Berry consumption shifted gastrointestinal bacterial communities towards obligate anaerobes, reducing luminal oxygen and oxidative stress [120]. A study investigated the distribution of black currant ACNs and their metabolites in lean and diet-induced obese mice with intact or disrupted gut microbiomes. Daily consumption of a black currant extract-supplemented diet improved glucose metabolism and reduced weight gain in mice with an intact gut microbiome but not those with a disrupted one. The results highlight the role of the gut microbiome and ACN aglycone type in modulating the protective effects of ACNs against obesity and insulin resistance [121]. Another study examined the metabolism of M3G, gallic acid, and a mixture of ACNs by gut bacteria. Most ACNs disappeared after 5 hours of incubation, while gallic acid degraded almost completely after 24 hours. M3G resulted in the formation of syringic acid, whereas the mixture of ACNs led to the formation of gallic, syringic, and p-coumaric acids. Furthermore, the tested ACNs promoted the growth of Bifidobacterium spp. and Lactobacillus–Enterococcus spp. [122]. The findings from all the studies above collectively support the potential of ACNs as promising natural interventions to enhance gut health and contribute to preventing and alleviating diseases.
5. Nutraceutical Potential of ACNs
ACNs have gained considerable attention in the nutraceutical industry, primarily for their role as natural colorants and antioxidants (Figure 3). ACN chalcones and quinoidal bases have been reported as efficient antioxidants, and the glycosylated B-ring structure of ACNs enhances its antioxidant activity, especially with ortho-hydroxylation and methoxylation [33,48,123]. However, anthocyanidin, the aglycone form of ACNs, exhibits higher ORAC values due to its greater stability and reactivity [124]. Acylation of ACNs with phenolic acids significantly increases their antioxidant activity, while diacylation enhances the activity further, and 5-glycosylation reduces the activity [124,125]. Studies have shown that various ACNs extracted from plants possess antioxidative properties. C3G, D3G, and Peo3G from black bean seed coats exhibit strong antioxidative activity and reduce malondialdehyde formation by ultraviolet B (UV-B) irradiation [126]. Delphinidin and D3G show the highest inhibitory effect on lipid peroxidation and O2•− scavenging activity. On the other hand, cyanidin and C3G demonstrate the highest inhibitory effect on copper (II)-induced low-density lipoprotein (LDL) oxidation, while delphinidin has intermediate efficacy [127]. Moreover, ACNs activate Nrf2 and antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), enhancing their enzymatic activity [128,129]. In addition to their antioxidant effects, ACNs demonstrate potent activity against inflammation and associated disease conditions [50]. The imbalance between proinflammatory molecules and anti-inflammatory mediators leads to chronic inflammation, linked to various chronic diseases such as neurodegenerative diseases, arthritis, allergies, liver diseases, diabetes, obesity, cardiovascular diseases, and cancer [130]. Several studies showed that the ACNs suppress the activation of NF-κB [131] and MAPK signaling cascades [64], inhibiting the production of proinflammatory cytokines and enzymes like COX-2 [132]. ACNs directly inhibit COX-1 and COX-2 enzymes, thus reducing the production of prostaglandin E2 (PGE2) [132]. Moreover, ACNs also suppress NLRP3 inflammasomes, a complex in inflammatory responses [133,134].
Further, ACNs exhibit promising potential as oral hepatoprotective agents against chemically-induced liver damage. They effectively combat liver lipid accumulation, inhibit lipogenesis, promote lipolysis, and counteract oxidative stress. They decrease liver enzyme levels, enhance antioxidant enzymes, and suppress the expression of COX-2 and iNOS, suggesting their potential therapeutic use in treating liver oxidative stress and associated diseases [135]. C3G–lauric acid ester displayed enhanced antioxidant activity compared to its precursor C3G, safeguarding cells through oxidative stress regulation and PI3K/Akt-mediated Nrf2–heme oxygenase 1 (HO-1)/ nicotinamide adenine dinucleotide (phosphate) reduced: quinone oxidoreductase (NQO1) pathway activation, highlighting the potential of acylation for improved antioxidative effects and stability [136]. Myrciaria jaboticaba (Vell.) O.Berg and Syzygium cumini (L.) Skeels peel powders exhibited protective effects against liver fibrosis and hepatocarcinogenesis, while Syzygium malaccense (L.) Merr. & L.M.Perry showed antioxidative benefits, emphasizing the anthocyanin profile effect [137]. Lonicera caerulea L. anthocyanin extract demonstrated hepatoprotective potential against alcoholic steatohepatitis through AMP-activated protein kinase (AMPK)-mediated anti-inflammatory and lipid-reducing pathways [138]. Bilberry fruit anthocyanin extract showed hepatoprotective effects against acute liver failure, reducing damage markers, oxidative stress, inflammation, and necrosis, attributed to their antioxidative and anti-inflammatory properties [139].
Due to high lipid content and energy demand, the central nervous system (CNS), particularly the brain, is highly vulnerable to excessive ROS [140]. ROS can be generated internally during cellular respiration and externally through environmental factors such as pollution, smoking, UV-B radiation, and infections [141]. An excessive inflammatory response in the CNS can contribute to neuronal apoptosis and the progression of neurodegenerative diseases [142]. In this context, ACNs found in various fruits and vegetables have shown neuroprotective effects in preclinical models of neurodegenerative diseases and cerebral ischemia [143]. ACNs exert their neuroprotective effects through multiple mechanisms. They act as direct scavengers of ROS [144], enhance antioxidant response pathways [145], and inhibit neuroinflammation by modulating specific signaling pathways [146,147,148]. Studies in animal models have demonstrated that ACNs can prevent cognitive decline, delay the onset of neurodegenerative diseases like Alzheimer's [149] and Parkinson's [150], and reduce brain damage in cerebral ischemia. In a randomized controlled trial involving twelve participants with mild cognitive impairment, older adults demonstrated enhanced learning and memory capacity after 12 weeks of grape juice supplementation compared to the placebo group [151]. Similarly, in another study, a 12-week supplementation of blueberry juice in the same group improved memory function [152,153]. These findings suggest that ACNs could be a potential co-adjuvant therapy to pharmacological treatments or a preventive strategy to reduce drug dosage and adverse effects in managing neurodegenerative diseases, cerebral ischemia, and cognition.
Dietary ACNs have demonstrated beneficial effects in protecting against cardiovascular diseases (CVDs) in animal and human studies [153]. Animal studies have shown that ACNs can prevent hypertriglyceridemia, hypercholesterolemia, and platelet hyperactivity induced by high-fat or high-fructose diets, while also protecting cardiac tissue against oxidative stress in ischemia/reperfusion conditions [154]. Human intervention studies have revealed that berry ACN intake can increase high-density lipoprotein (HDL)-cholesterol and reduce LDL-cholesterol, triglycerides, blood pressure, and inflammatory markers [155,156]. Furthermore, higher ACN intake has been associated with a reduced risk of myocardial infarction and decreased incidence of coronary heart disease and CVD-related mortality [157,158]. The cardioprotective mechanisms of ACNs include increasing plasma antioxidant capacity and nitric oxide levels, reducing LDL oxidation and platelet aggregation, and enhancing glutathione and omega-3 fatty acid synthesis. Cardiovascular diseases, including myocardial infarction, ischemic heart disease, and stroke, are major causes of mortality globally. Atherosclerosis, driven by oxidized low-density lipoproteins (ox-LDL), plays a key role in these diseases. ACNs have been found to protect against atherosclerosis in animal models by reducing the formation of atherosclerotic plaques and improving dyslipidemia. ACNs also prevent LDL-cholesterol oxidation [159], reduce inflammation [160], and lower the expression of adhesion molecules in the aorta [160,161], leading to decreased leukocyte infiltration and proinflammatory cytokines. Additionally, ACNs have shown cardioprotective effects in animal models of myocardial infarction and chemotherapeutic drug-induced cardiotoxicity [162].
Obesity is a significant global health issue associated with numerous metabolic disorders. Studies have shown that dietary ACNs can have beneficial effects in combating obesity. One mechanism through which ACNs act as anti-obesity agents is by increasing energy expenditure [163]. Berries rich in ACNs, such as petunidin and malvidin, were found to effectively reduce metabolic damage caused by a high-fat diet by boosting energy expenditure and reducing mitochondrial dysfunction in adipose tissue [164]. Moreover, ACNs have been found to modulate the AMPK, a key regulator of energy balance [165,166]. By activating AMPK, ACNs can promote mitochondrial biogenesis, reduce lipid metabolism, increase fatty acid oxidation, and improve glucose metabolism. This AMPK activation was associated with improved glucose uptake in skeletal muscle, decreased glucose production in the liver, and decreased liver lipid content and serum lipoproteins. Furthermore, dietary ACNs have been shown to affect lipid metabolism by downregulating key molecules such as fatty acid synthase and sterol-regulatory element-binding proteins. ACNs from different sources were found to reduce hyperglycemia and inhibit hepatic lipogenesis, potentially suppressing inflammation in obese individuals [167,168].
In addition to their effects on energy expenditure and lipid metabolism, ACNs have been linked to the modulation of gut microbiota [169]. Gut microbiota changes in obese individuals can contribute to metabolic disorders, and ACNs have been studied for their potential role in regulating gut microbiota to combat obesity-related conditions. Moreover, ACNs have demonstrated promising results in protecting pancreatic β cells, improving insulin resistance, and reducing cholesterol levels in diabetic subjects [170]. ACNs-rich beverages have also positively affected postprandial glycemic response and insulin excretion [171]. Numerous in vitro and in vivo studies have consistently supported the potential anti-obesity effects of ACNs, demonstrating their ability to inhibit fat accumulation [172], improve lipid profiles [168], and reduce adipocyte hypertrophy [173]. Furthermore, ACNs have shown anti-inflammatory properties by suppressing NF-κB signaling and promoting an anti-inflammatory phenotype in adipose tissue macrophages [174]. Thus, ACNs present a promising natural approach to combat obesity and related metabolic disorders. The mechanisms through which ACNs exert their anti-obesity effects involve increased energy expenditure, modulation of AMPK, lipid metabolism, gut microbiota, and anti-inflammatory properties.
ACNs have emerged as potential antitumoral agents, attracting significant interest in recent decades [175,176]. Cancer is characterized by uncontrolled cell proliferation, resistance to apoptosis, and cell migration, often involving dysregulation of pathways like Notch, Wnt/β-catenin, NF-κB, and MAPK [177]. Dietary ACNs from berries have been found to modulate the levels of proteins involved in these pathways, leading to cell growth inhibition. Notably, ACN mixtures have shown enhanced effects compared to single purified ACNs, indicating overlapping pathways in cell growth inhibition [178]. ACNs exhibit diverse cancer prevention and inhibition mechanisms, targeting pathways related to cell survival, proliferation, apoptosis, inflammation, and angiogenesis. They have been shown to suppress NF-κB [179], PI3K/Akt [180,181], and Akt/mTOR [182] signalling pathways. ACNs can also inhibit tumor growth, promote apoptosis and autophagy in cancer cells, and inhibit angiogenesis and metastatic migration [183]. In vitro and in vivo studies have demonstrated the anticancer potential of ACNs from various sources, including berry extracts [78,184] and pure ACNs like delphinidin [185] and cyanidin 3-rutinoside [181]. These findings suggest that ACNs hold promise with potential anticancer properties, making them a subject of considerable research and exploration in cancer therapy and prevention.
ACN pigments found in various berries are beneficial for maintaining good vision and have been associated with improved night vision [48,186]. Berries rich in ACNs have long been known for their positive effects on eye health. For instance, administering bilberry extract, containing about 39% ACNs, to mice has been shown to protect photoreceptor cell function during retinal inflammation [187]. In a study with 132 patients having normal-tension glaucoma, daily supplementation with two capsules containing 60.0 mg ACNs from bilberry resulted in improved visual function [188]. Additionally, other berries like blackcurrant [189] and purple corn seed [190] have shown a protective effect on eyesight, with blackcurrant ACNs supplementation increasing ocular blood flow in patients with open-angle glaucoma, and purple corn seed extract reducing lens opacity and malonaldehyde levels. Furthermore, ACNs from black soybean seed coat has effectively prevented retinal degeneration and suppressed human lens epithelial cell death under oxidative stress [191].
ACNs exhibit potent antimicrobial properties against a wide range of microorganisms, particularly in inhibiting the growth of food-borne pathogens. Their antimicrobial effects are attributed to various mechanisms, including cell wall, membrane, and intercellular matrix damage [175]. For instance, maqui berry extracts have shown antibacterial activity, being highly effective against Aeromonas hydrophilia and Listeria innocua [192], commonly associated with refrigerated foods as indicators of pathogenic or spoilage microorganisms [193]. Cranberry extract also demonstrates antibacterial activity against certain resistant strains of Enterococcus faecium, Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli, with its effect not solely due to low pH but rather to specific bioactive components like ACNs and flavonols [194]. ACN-rich extracts from various fruits like blueberry, raspberry, blackcurrant, and strawberry display inhibitory effects on Gram-negative bacteria but not on Gram-positive bacteria [195], likely due to differences in cell wall structures. These antimicrobial actions are likely a result of synergistic effects from various phytochemicals in the extracts, including ACNs, weak organic acids, phenolic acids, and other chemical forms [196]. Hence, the complex nature of these compounds necessitates further comprehensive analysis of their antimicrobial potential.
From a nutraceutical point of view, ACNs are highly valued in the food and supplement industry for their diverse health benefits. As natural colorants, they add vibrant hues to foods and beverages. Besides the immunomodulatory potential ACNs possess potent antioxidant properties, neutralizing free radicals and protecting cells from oxidative damage. Moreover, they exhibit anti-inflammatory actions by inhibiting pathways involved in inflammation and activating antioxidant enzymes. ACNs have shown neuroprotective effects, making them potential candidates for supporting brain health. They also protect against cardiovascular diseases by improving lipid profiles and reducing blood pressure. In obesity, ACNs are linked to increased energy expenditure and improved lipid metabolism. They have potential as antitumoral agents by targeting pathways involved in cancer cell growth. Additionally, ACNs contribute to maintaining good vision and display antimicrobial properties against food-borne pathogens. Thus, these compounds hold great promise as natural compounds with multifaceted health benefits, making them valuable in nutraceutical products for promoting overall well-being and preventing chronic diseases. As a nutraceutical, the bioavailability of ACN is the key factor for biological action. The low bioavailability of ACNs causes low absorption of these compounds into the blood circulating system and a high excretion rate of ACNs in urine and feces, thus reducing the efficacy of ACNs bioactivity.
A Considerable body of research has been devoted to the assessment of biological effects of ACNs including their immune modulation potential. Array of signaling pathways and cellular process is involved in immune modulation activities of ACNs, and may provide potential therapeutic targets and strategies for the improvement of immune mediated disorders in future. However, much remains to be elucidated before their application in clinical settings. The metabolites of ACNs need in depth assessment for their potential to modulate immune processes, as ACNs are transported in blood and urine primarily as metabolites. In addition, the nutraceutical potential of ACNs is mostly demonstrated using in vitro and in vivo experimental models; however, their low bioavailability is one of the key obstacles in achieving the desired biological effects. Novel approaches are desired to developed, intended to enhance the bioavailability of ACNs in human subjects.
6. Factors Influencing Pharmacokinetics of ACNs
The pharmacokinetics of ACNs are influenced by various factors, including the site of administration, chemical structure, pH, temperature, food matrix, and gut microbiota [107,197]. Interactions between ACNs and salivary proteins within the oral cavity can potentially lead to degradation [198]. Both dietary intake and binding with salivary proteins can reduce ACN levels in the mouth [199,200]. ACN stability is also impacted by the composition of the food matrix and the molecular structure, with higher B-ring hydroxylation leading to decreased stability [201,202,203]. ACNs exhibit sensitivity to oral pH levels within the range of 6.0 to 7.0 and temperature variations. Some conversions occur under neutral to basic pH conditions [202,204]. For instance, approximately 30% of C3G can convert into the chalcone form under these conditions [202].
Upon oral consumption, ACNs enter the stomach, where the acidic pH (ranging from 1.5 to 5.0) impacts their stability [205]. In this environment, ACNs can either be absorbed or progress to the small intestine for further processing before entering the bloodstream. Studies on rats have demonstrated the presence of ACNs in both systemic and portal plasma shortly after ingestion, suggesting stomach absorption. A 14-week experiment involving 32 rats fed diets containing chokeberry, bilberry, and grape extracts revealed plasma ACN levels below 2 μmol/L, with higher presumed metabolites. Urine analysis showed varying concentrations of intact ACNs and methylated derivatives (e.g., 17.4 nmol/L for bilberry and 52.6 nmol/L for chokeberry) [206]. Approximately 75-97% of total ACNs were recovered from the gastrointestinal tracts of rats within 30-120 minutes after oral intake, as demonstrated in research [207]. In vitro digestion studies indicated a 75-88% ACN recovery rate under gastric conditions, underscoring the limited degradation of ACNs by gastric enzymes [101,208]. Evidence suggests that ACNs likely traverse cell membranes through active transport mechanisms owing to their intricate structure [25,209]. Membrane carriers, such as organic anion carriers like bilitranslocase and glucose transporters (GLUT), participate in this process [110,210,211]. ACN absorption involves GLUT1 and GLUT3, along with other transporters [209]. At gastric pH levels ranging from 1.5 to 4, ACNs maintain their glycoside form [212,213].
ACNs experience significant degradation within the intestinal tract due to elevated pH levels (ranging from pH 5.6 to 7.9), the influence of gut microbiota, and intestinal enzyme activity. The pH within the small intestine spans from 5.0 to 7.0, while the large intestine maintains a pH of 7.0 to 8.0 [214]. Enzymatic conversion of ACNs into anthocyanidins, facilitated by β-glucosidase and lactase-phlorizin hydrolase, occurs in this environment [211]. The gastrointestinal epithelial tissues absorb several bioactive ACN derivatives, including protocatechuic acid, vanillic acid, gallic acid, and phloroglucinol aldehyde [215]. The absorption of ACNs from the small intestine occurs through active transport mechanisms involving transporters such as the sodium-dependent glucose cotransporter (SGLT1), GLUT2, and bilitranslocase [216,217,218]. The hydrolysis of ACNs into anthocyanidins enables their passive diffusion, a process facilitated by enzymes like β-glucosidase [211,216,218,219].
In a laboratory experiment, treating a Caco-2 monolayer with a 200 mg/L ACN extract for four days led to an increase in the expression of the glucose transporter 2 (GLUT2) and the rate of ACN transport [220]. However, prolonged exposure (16 hours) of Caco-2 cells to berry extracts (0.125%, w/v) resulted in reduced expression of sodium-dependent glucose transporter 1 (SGLT1) and GLUT2 mRNA [221]. ACNs are predominantly absorbed via the SGLT1 and GLUT2 pathways, with their phase II metabolites (glucuronide and sulfate) also being actively absorbed through ATP-binding cassette (ABC) proteins [222]. Unabsorbed ACNs within the colon undergo enzymatic breakdown through the activity of colonic bacteria, which includes enzymes like α-galactosidase, β-D-glucuronidase, β-D-glucosidase, and α-rhamnosidase. These enzymatic processes result in the cleavage of glycosidic bonds within 20 minutes to 2 hours. This cleavage yields smaller phenolic compounds, including benzaldehydes or hydroxytyrosol, as well as phenol aldehydes and phenolic acids like hydroxybenzoic, homovanillic, phenyl propionic, protocatechuic, syringic, gallic, and vanillic acids [33,223].
In a 48-hour human study, 44% of the 13C label from C3G intake was excreted in urine (5.4%), breath (6.9%), and faeces (32.1%), indicating complete catabolism. Over 50% of the label remained in the body, attributed to slow clearance due to colonic metabolism and other factors [224]. In mice, after 3 hours, almost 90% of the 14C label from oral 14C C3G intake was in adipose and gastrointestinal tissues, decreasing to 50% in faeces and 3.3% in urine after 24 hours [225]. Human studies indicated a slower anthocyanin clearance than in mice, with variations in recovery percentages and plasma concentrations [224]. Methylation and glucuronidation were major in vivo C3G conjugation routes, as shown by various intervention models [224,226,227]. Liver metabolism was evident, with bile presence of C3G and Peo3G after oral administration to rats [228]. An intravenous (IV) administration revealed rapid fluctuations of methylated C3G in plasma, indicating absorption or decomposition [229]. Anthocyanidin stability and fate depended on conjugation possibilities [224,226,227].
In swine studies, M3G reached peak concentrations in brain tissue (4.43 pmol/g) after bilberry extract intake and malvidin glycosides dominated in various tissues of blueberry-fed pigs [230,231,232]. Mice also showed malvidin predominance in plasma and tissues after bilberry anthocyanin consumption [233]. In humans, malvidin glycosides ranked among the top urinary anthocyanin metabolites during blueberry juice consumption, alongside other C6-C3-C6 forms [228]. These findings suggest malvidin's resistance to metabolism due to its structural attributes, supported by evidence of enzymatic methylation from petunidin to M3G [234,235]. The observed pelargonidin-based metabolite recovery is attributed to functional group removal, yielding pelargonidin glycoside and pelargonidin [236,237,238]. Urinary pelargonidin glucuronide resulted from cyanidin dehydroxylation in a human study with various cyanidin glycosides [226]. Pel3G formation was observed upon IV administration of C3G [229]. Notably, blueberries lack pelargonidin glycosides, yet pelargonidin-based metabolites were abundant in urine after long-term juice intake [228], potentially making them a flavonoid-based anthocyanin intake biomarker due to pelargonidin's high membrane solubility [239].
In an investigation, four healthy older women were administered 720 mg of anthocyanins, leading to the identification of anthocyanin glycosides in both plasma and urine. Notably, the peak plasma concentration averaged 97.4 nmol/L after 71.3 minutes, displaying elimination patterns adhering to first-order kinetics with a half-life of 132.6 minutes. Most anthocyanins were excreted within the initial 4 hours, characterized by an initial excretion rate of 77 microg/h followed by 13 microg/h subsequently [240]. Further investigations explored plasma anthocyanins in overweight adults (BMI: 26 ± 2 kg/m²) consuming strawberry beverages at varying meal timings. Optimal concentrations and bioavailability of pelargonidin-based anthocyanins were observed when the strawberry drink was consumed two hours prior to the meal, in contrast to during or after the meal, highlighting the mealtime-dependent impact on anthocyanin pharmacokinetics [241]. A separate study investigated the pharmacokinetics of bilberry anthocyanins and their interaction with glucose transporters (sGLT1 and GLUT2) during absorption. Rats were administered a standardized bilberry extract containing 15 distinct anthocyanins (Mirtoselect®), with liquid chromatography-electrospray ionization tandem mass spectrometry (LC/ESI-MS/MS) utilized to measure absorption. Fasting significantly augmented anthocyanin bioavailability sevenfold compared to fed rats, and glucose co-administration exhibited no interference with uptake. Both aglycone and sugar constituents influenced the variance in bioavailability among anthocyanins. Computational analysis revealed a correlation between anthocyanin absorption and recognition by glucose transporters sGLT1 and GLUT2, offering valuable insights into this complex interplay [242].
The absorption and clearance kinetics of anthocyanins, with reported maximum plasma concentrations ranging from 1.4 to 592 nmol/l, occurring within 0.5-4 hours post-consumption (doses: 68-1300 mg) [243]. Urinary excretion averages 0.03-4% of the dose, with elimination half-lives of 1.5-3 hours [244]. Although anthocyanin metabolism is not fully understood, recent evidence indicates that they are primarily absorbed and transported as metabolites in human serum and urine, constituting 68-80% of urinary anthocyanin content [245,246]. These findings significantly advance our understanding of the intricate processes governing ACN absorption and cellular responses to their metabolites. Due to uncertainties surrounding ACN bioavailability, numerous researchers are actively exploring innovative strategies to enhance the stability and bioavailability of these natural compounds.
7. Strategies for Enhanced ACN Bioavailability
The daily consumption of ACNs can vary depending on individual factors such as age, health status, and dietary preferences. Though anthocyanin-rich foods offer potential health benefits, there is no specific recommended daily intake (RDI) for ACNs, as they are not considered essential nutrients like vitamins and minerals. Despite the numerous health benefits associated with anthocyanin, its bioavailability in the plasma is limited, with less than 6% of the initial dose being absorbed after consuming a meal rich in anthocyanin. This restricted absorption is attributed to the rapid degradation of ACN under various external conditions, such as temperature, pH, light, oxygen, water activity, and enzymatic activity. In addition, interaction of ACNs with other food matrix (such as carbohydrates, proteins, and dietary fibers) in processed foodstuffs may also affect the release of ACNs from food matrix, thus affecting their bioaccessibility and bioavailability. These conditions lead to its high reactivity, resulting in reduced availability for the body [247]. In the United States, the reported daily consumption of ACN pigments is around 12.5 mg per person, but the bioavailability of these compounds in diets is relatively poor, typically averaging between 1-2% [248]. ACN stability refers to the ability of these compounds to resist degradation and maintain their chemical structure, color, and bioactivity over time and under various environmental conditions. Improving the stability and bioavailability of ACNs could maximize their health benefits and promote their inclusion in daily diets.
Using ACNs to form a polymeric matrix of polysaccharides, proteins, or liposomes nano-delivery vehicles represents a promising approach to enhance stability and bioavailability [249]. Polysaccharides offer several advantages, including their abundant availability, cost-effectiveness, ease of chemical modification, and biodegradability, making them favorable compared to other matrices. Proteins possess a complex bioactive structure that renders them sensitive to environmental factors such as pH, ionic strength, and temperature changes. This sensitivity may affect their stability during delivery. Liposomes exhibit reduced stability and are prone to aggregation and phospholipid oxidation, especially when stored over extended periods [250]. These polymeric matrix serves as a protective barrier for ACNs, shielding them from factors that might otherwise degrade or inactivate them, such as heat, light, oxygen, and changes in pH. Moreover, incorporating ACNs into a polymeric matrix can facilitate their controlled release, enabling a gradual and sustained release of these bioactive compounds in the body [48]. Starch [251], chitosan [55,252], cyclodextrins [253], cellulose [254], and sodium alginate [255] are frequently employed as encapsulation materials because of their excellent biocompatibility and biodegradability. Additionally, emulsion cross-linking (e.g., ionic cross-linking, covalent cross-linking, etc.), self-assembly (e.g., layer-by-layer, self-assembled micelles, etc.), and spray drying techniques are commonly utilized to prepare polysaccharide-ACN nano-systems [250].
A study found that the incorporation of β-cyclodextrin (β-CD) slowed the degradation of C3G and reduced its deterioration during simulated in vitro digestion. This finding suggests the potential of β-CD to enhance the stability and delay the degradation of blackberry ACNs under conditions similar to the gastrointestinal tract [253]. To enhance the stability of saffron ACNs under harsh gastric conditions, ACNs were encapsulated using spray drying with β-glucan and β-CD. The resulting microcapsules successfully protected the ACNs against stomach conditions and displayed positive attributes such as controlled release and antioxidant behavior [256]. Through spray drying, ethanolic pomace extract was microencapsulated with whey protein isolate. In vitro, testing showed that while the ACN concentration decreased after intestinal digestion for microcapsules, it remained consistent for the non-encapsulated system. This indicates the potential of microencapsulation to influence the release of ACNs during digestion [257]. A complex consisting of Bryophyllum pinnatum (Lam.) Oken extract and β-CD was created and integrated into an oil-in-water emulsion for topical anti-inflammatory evaluation. The incorporation of β-CD enhanced the aggregate properties and antioxidant activity of the emulsion, suggesting its potential in topical applications [258]. Microencapsulation of anthocyanin-rich extracts from Leum Pua black rice bran was achieved through spray drying with various wall materials, including maltodextrin, gum arabic, and whey protein isolate. Optimal retention of ACNs was observed with maltodextrin and maltodextrin-whey protein combinations, while whey protein exhibited high phenolic content and antioxidant activity. During in vitro digestion, microcapsules with pure whey protein showed the highest release of phenolic content and antioxidant activity in intestinal fluids [259].
Sour cherry skin ACNs were successfully encapsulated in whey protein isolate microcapsules, demonstrating substantial polyphenol content and facilitating controlled release in the intestine. Additionally, the encapsulated ACNs promoted the growth of Lactobacillus casei 431®, a beneficial probiotic bacterium [260]. A study investigated the impact of whey protein and citrus pectin encapsulation on ACN bioavailability and intestinal accessibility in humans. The results indicated that whey protein encapsulation influenced short-term bioavailability. In contrast, citrus pectin encapsulation enhanced intestinal accessibility, affecting the formation of degradation products in human plasma during passage through the small intestine [261]. The bioavailability of ACNs from blackberry pomace microcapsules and microcapsule-supplemented yogurt after in vitro digestion was evaluated. Three microencapsulation methods (spray-drying, freeze-drying, and ionic gelation) were compared, with spray-drying and ionic gelation producing stable microcapsules that improved C3G bioavailability in yogurt formulations [262]. Another study aimed to enhance the stability and bioavailability of ACNs through cyclodextrin encapsulation. Encapsulation slowed down ACN degradation and led to the formation of phenolic acids. The study also highlighted the potential of encapsulation to enhance ACN release in the colon, potentially conferring health benefits [263]. A novel approach utilized chitosan/β-lactoglobulin nano complexes to encapsulate ACNs for enhanced sustained release, stability, and bioavailability. The optimized nano complexes exhibited improved stability and bioavailability under simulated gastrointestinal conditions, showing promise for sustained release applications [264]. Encapsulation of anthocyanin-rich bilberry extract (BE) aimed to improve its stability. The study revealed that encapsulated BE exhibited slower release and higher availability of ACNs, indicating the role of encapsulation in inhibiting early degradation in the intestinal system [265].
Researchers developed a thermally induced matrix system using strongly acidic whey protein isolate gels to encapsulate anthocyanin-rich bilberry extract. The encapsulation process influenced release kinetics, stabilization, and release characteristics, with pH-dependent interactions impacting overall stability [266]. Alginate/chitosan beads were fabricated through spray drying and external gelation methods to encapsulate ACNs, enhancing their stability and bioavailability. The study found that bead degradation and electrostatic interactions influenced ACN release, with certain beads identified as optimal microcarriers [267]. For Iranian borage ACNs, maltodextrin/modified maize starch was identified as the optimal carrier to target drug delivery to the intestines. Modifying starch improved its solubility and swelling behavior, suggesting its potential as an effective drug carrier [251]. Porous microgels derived from oxidized starch hydrolysate were created to enhance ACN delivery. These microgels exhibited efficient adsorption and release, ultimately improving ACN stability and release [268]. Chitosan nanoparticles were employed to encapsulate ACNs from black carrots, leading to pH-dependent release profiles that protected against rapid degradation within the digestive environment. This technique augmented antioxidant effectiveness, stability, and overall ACN presence [269]. A composite co-precipitation technique using chitosan and pectin was utilized to fabricate nanocarriers for ACNs, offering protection against degradation and delayed release upon exposure to intestinal fluid [270]. Cellulose nanocrystals were integrated into chitosan-based microencapsulation for blueberry ACN extracts, resulting in durable microcapsules with enhanced stability and controlled release properties [271].
Lipid-based nanocarriers encompass diverse systems that utilize lipids, such as fats and oils, providing versatility in solubilizing bioactive ingredients. They can dissolve or entrap these ingredients within the oil phase for hydrophobic components or the aqueous phase for hydrophilic ones. One notable category within this group is nanoemulsions, comprising oil, water, and surfactants/biopolymers in various forms, including single O/W or W/O nanoemulsions, double nanoemulsions (O/W/O or W/O/W), Pickering nanoemulsions stabilized by biopolymer nanoparticles, and structural nanoemulsions stabilized by single or double layers of biopolymer coatings [272,273,274]. Another significant group of lipid-based nanocarriers is nanoliposomes, which consist of phospholipids, oils, and different solvents. Nanoliposomes feature a central aqueous cavity suitable for entrapping hydrophilic ingredients. At the same time, hydrophobic components can be encapsulated within the nanoliposome membrane, composed of either a single bilayer or a double layer of phospholipids [275]. One study aimed to enhance the durability of ACNs from red cabbage within the gastrointestinal system by incorporating red cabbage extract (RCE) into solid lipid nanoparticles (SLNs) using a water-in-oil microemulsion. The resilience of RCE-SLNs was found to be stronger at pH 3.0 (gastric fluid) compared to pH 5.0 (intestinal fluid), and storing them at 25 °C preserved ACNs, ensuring increased absorption and potential health benefits [276]. Similarly, a stable mangosteen peel extract (MPE) nanoemulsion was achieved through high-speed homogenization with specific ratios of virgin coconut oil, Tween 80, and Span 80. This resulting MPE-NE remained physically stable for 28 days and exhibited a shelf life of up to one year, suggesting its potential for topical applications [277]. Furthermore, nanoliposomes incorporating pistachio green hull extract (PHE-NL) and soy lecithin (1-3%) demonstrated improved preservation of ACNs and lipid oxidation stability compared to PHE or empty nanoliposomes, attributed to the presence of polyphenols [278].
8. Conclusions and Perspectives
Exploring ACNs as immunomodulatory dietary supplements presents a promising avenue for harnessing the potential of natural compounds to support and enhance immune system function. The extensive research discussed in this review highlights the multifaceted mechanisms through which ACNs exert their immunomodulatory effects, ranging from antioxidant and anti-inflammatory activities to direct interactions with immune cells. The diverse sources of ACNs, including various fruits, vegetables, and plants, offer various options for individuals seeking to incorporate these compounds into their diets. However, the full realization of the immunomodulatory benefits of ACNs hinges upon overcoming the challenges associated with their limited bioavailability. Nano strategies have emerged as innovative solutions for enhancing the absorption and bioavailability of ACNs, thereby maximizing their therapeutic potential. Using nanocarriers and encapsulation techniques holds the promise of optimizing the delivery of ACNs to target tissues and cells, ultimately amplifying their immunomodulatory effects.
Future research should focus on a comprehensive understanding of the molecular mechanisms underlying anthocyanin-immune cell interactions, unraveling intricate signaling pathways, and deciphering the precise impact on immune system regulation. Additionally, continued efforts in developing advanced nano-formulations are paramount to ensure the effective delivery and bioavailability enhancement of ACNs. As personalized nutrition gains prominence, tailoring anthocyanin-based interventions to specific populations and immune-related conditions could provide valuable insights into their clinical applications. Collaborative endeavors between nutritionists, immunologists, pharmacologists, chemists, and nanotechnologists are essential to translate the potential demonstrated in preclinical and early clinical studies into tangible health benefits for individuals of all ages. Continued interdisciplinary research and translational efforts will pave the way for a future where ACNs contribute significantly to optimizing immune system function and promoting overall health.
Author Contributions
Conceptualization, T.P.I., H.U. and M.D.; methodology, T.P.I.; formal analysis, T.P.I. and L.F.D.L.; investigation, T.P.I., L.F.D.L., A.B. and H.U.; resources, T.P.I., L.F.D.L., D.G.B., M.R.P.G. and A.B.; writing—original draft preparation, T.P.I. and L.F.D.L.; writing—review and editing, T.P.I., L.F.D.L., M.R.P.G., S.S., P.S., H.U. and D.G.B.; visualization, T.P.I. and P.S.; supervision, M.R.P.G., H.U. and M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mei, H.; Dong, X.; Wang, Y.; Tang, L.; Hu, Y. Managing Patients with Cancer during the COVID-19 Pandemic: Frontline Experience from Wuhan. Lancet Oncol. 2020, 21, 634–636. [Google Scholar] [CrossRef]
- Mastrandrea, L.D. An Overview of Organ-Specific Autoimmune Diseases Including Immunotherapy. Immunol. Invest. 2015, 44, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jia, S.; Xu, H. Potential Therapeutic Applications of Exosomes in Different Autoimmune Diseases. Clin. Immunol. 2019, 205, 116–124. [Google Scholar] [CrossRef]
- Rusek, P.; Wala, M.; Druszczyńska, M.; Fol, M. Infectious Agents as Stimuli of Trained Innate Immunity. Int. J. Mol. Sci. 2018, 19, 456. [Google Scholar] [CrossRef]
- Mrak, D.; Tobudic, S.; Koblischke, M.; Graninger, M.; Radner, H.; Sieghart, D.; Hofer, P.; Perkmann, T.; Haslacher, H.; Thalhammer, R.; et al. SARS-CoV-2 Vaccination in Rituximab-Treated Patients: B Cells Promote Humoral Immune Responses in the Presence of T-Cell-Mediated Immunity. Ann. Rheum. Dis. 2021, 80, 1345–1350. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, A.; Bi, Y.; Wang, Y.; Yang, Q.; Cao, Y.; Li, Y.; Liu, G. Targeting Myeloid-Derived Suppressor Cells in Cancer Immunotherapy. Cancers (Basel) 2020, 12, 2626. [Google Scholar] [CrossRef]
- Ivashkiv, L.B. IFNγ: Signalling, Epigenetics and Roles in Immunity, Metabolism, Disease and Cancer Immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef]
- Galon, J.; Bruni, D. Tumor Immunology and Tumor Evolution: Intertwined Histories. Immunity 2020, 52, 55–81. [Google Scholar] [CrossRef]
- Bilanges, B.; Posor, Y.; Vanhaesebroeck, B. PI3K Isoforms in Cell Signalling and Vesicle Trafficking. Nat. Rev. Mol. Cell Biol. 2019, 20, 515–534. [Google Scholar] [CrossRef]
- Peluso, I.; Yarla, N.S.; Ambra, R.; Pastore, G.; Perry, G. MAPK Signalling Pathway in Cancers: Olive Products as Cancer Preventive and Therapeutic Agents. Semin. Cancer Biol. 2019, 56, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting Signalling Pathways and the Immune Microenvironment of Cancer Stem Cells - a Clinical Update. Nat. Rev. Clin. Oncol. 2020, 17, 204–232. [Google Scholar] [PubMed]
- Macedo, A.C; de Faria, A.O.V.; Ghezzi, P. Boosting the Immune System, from Science to Myth: Analysis the Infosphere with Google. Front. Med. (Lausanne) 2019, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.N.; Marcon, A.R.; Caulfield, T. “Immune Boosting” in the Time of COVID: Selling Immunity on Instagram. Allergy Asthma Clin. Immunol. 2020, 16. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.C.; Black, L.I.; Stussman, B.J.; Barnes, P.M.; Nahin, R.L. Trends in the Use of Complementary Health Approaches among Adults: United States, 2002-2012. Natl. Health Stat. Report. 2015, 1–16. [Google Scholar]
- Grand View Research, Dietary Supplements Market Size, Share & Trend Analysis Report by Ingredient (Botanicals, Vitamins, Minerals, Amino Acids, Enzymes), By Product, By Application, By End-use, And Segment Forecasts, 2018 - 2024. (2018). Archived at: https://web.archive.org/web/20190115112358/https://www.grandviewresearch.com/industry-analysis/dietary-supplements-market (accessed June 2, 2023).
- Wainwright, C.L.; Teixeira, M.M.; Adelson, D.L.; Buenz, E.J.; David, B.; Glaser, K.B.; Harata-Lee, Y.; Howes, M.-J.R.; Izzo, A.A.; Maffia, P.; et al. Future Directions for the Discovery of Natural Product-Derived Immunomodulating Drugs: An IUPHAR Positional Review. Pharmacol. Res. 2022, 177, 106076. [Google Scholar] [CrossRef]
- Alhazmi, H.A.; Najmi, A.; Javed, S.A.; Sultana, S.; Al Bratty, M.; Makeen, H.A.; Meraya, A.M.; Ahsan, W.; Mohan, S.; Taha, M.M.E.; et al. Medicinal Plants and Isolated Molecules Demonstrating Immunomodulation Activity as Potential Alternative Therapies for Viral Diseases Including COVID-19. Front. Immunol. 2021, 12, 637553. [Google Scholar] [CrossRef]
- Demaria, O.; Cornen, S.; Daëron, M.; Morel, Y.; Medzhitov, R.; Vivier, E. Publisher Correction: Harnessing Innate Immunity in Cancer Therapy. Nature 2019, 576, E3. [Google Scholar] [CrossRef]
- Di Sotto, A.; Vitalone, A.; Di Giacomo, S. Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines (Basel) 2020, 8, 468. [Google Scholar] [CrossRef]
- Maheshwari, S.; Kumar, V.; Bhadauria, G.; Mishra, A. Immunomodulatory Potential of Phytochemicals and Other Bioactive Compounds of Fruits: A Review. Food Front. 2022, 3, 221–238. [Google Scholar] [CrossRef]
- Jantan, I.; Ahmad, W.; Bukhari, S.N.A. Corrigendum: Plant-Derived Immunomodulators: An Insight on Their Preclinical Evaluation and Clinical Trials. Front. Plant Sci. 2018, 9, 1178. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Singh, V.R.; Verma, S.; Meshram, N.; Dhruw, L.; Sharma, R.; Ghosh, K.K.; Gupta, R.C. Plant and Food Derived Immunomodulators as Nutraceuticals for Performance Enhancing Activities. In Nutraceuticals in Veterinary Medicine; Springer International Publishing: Cham, 2019; pp. 593–601. [Google Scholar]
- Kim, J.H.; Kim, D.H.; Jo, S.; Cho, M.J.; Cho, Y.R.; Lee, Y.J.; Byun, S. Immunomodulatory Functional Foods and Their Molecular Mechanisms. Exp. Mol. Med. 2022, 54, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Chemistry, Pharmacology and Health Benefits of Anthocyanins: Anthocyanins and Human Health. Phytother. Res. 2016, 30, 1265–1286. [Google Scholar] [CrossRef]
- do Rosario, V.A.; Chang, C.; Spencer, J.; Alahakone, T.; Roodenrys, S.; Francois, M.; Weston-Green, K.; Hölzel, N.; Nichols, D.S.; Kent, K.; et al. Anthocyanins Attenuate Vascular and Inflammatory Responses to a High Fat High Energy Meal Challenge in Overweight Older Adults: A Cross-over, Randomized, Double-Blind Clinical Trial. Clin. Nutr. 2021, 40, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Amaya, D.B. Natural Food Pigments and Colorants. In Reference Series in Phytochemistry; Springer International Publishing: Cham, 2018; pp. 1–35. ISBN 9783319264783. [Google Scholar]
- Qi, Q.; Chu, M.; Yu, X.; Xie, Y.; Li, Y.; Du, Y.; Liu, X.; Zhang, Z.; Shi, J.; Yan, N. Anthocyanins and Proanthocyanidins: Chemical Structures, Food Sources, Bioactivities, and Product Development. Food Rev. Int. 2022, 1–29. [Google Scholar] [CrossRef]
- Kozłowska, A.; Dzierżanowski, T. Targeting Inflammation by Anthocyanins as the Novel Therapeutic Potential for Chronic Diseases: An Update. Molecules 2021, 26, 4380. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Anthocyanins. Adv. Nutr. 2015, 6, 620–622. [Google Scholar] [CrossRef]
- Riaz, M.; Zia-Ul-Haq, M.; Saad, B. Biosynthesis and Stability of Anthocyanins. In Anthocyanins and Human Health: Biomolecular and Therapeutic Aspects; Springer International Publishing: Cham, 2016; pp. 71–86. [Google Scholar]
- Lee, Y.-M.; Yoon, Y.; Yoon, H.; Park, H.-M.; Song, S.; Yeum, K.-J. Dietary Anthocyanins against Obesity and Inflammation. Nutrients 2017, 9, 1089. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the Art of Anthocyanins: Antioxidant Activity, Sources, Bioavailability, and Therapeutic Effect in Human Health. Antioxidants (Basel) 2020, 9, 451. [Google Scholar] [CrossRef]
- Calderaro, A.; Barreca, D.; Bellocco, E.; Smeriglio, A.; Trombetta, D.; Laganà, G. Colored Phytonutrients: Role and Applications in the Functional Foods of Anthocyanins. In Phytonutrients in Food: From Traditional to Rational Usage; Nabavi, S.M., Suntar, I., Barreca, D., Khan, H., Eds.; Elsevier: Woodhead Publishing, 2020; pp. 177–195. [Google Scholar]
- Onyilagha, J. C.; Grotewold, E. The Biology and Structural Distribution of Surface Flavonoids. Recent Res. Dev. Plant Sci 2004, 2, 53–71. [Google Scholar]
- Harbone, J.B. The Flavonoids: Recent Advances. In Plant Pigments; Goodwin, T.W., Ed.; Academic Press: San Diego, CA, USA, 1988; pp. 299–343. [Google Scholar]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of Plant Pigments: Anthocyanins, Betalains and Carotenoids. Plant J. 2008, 54 (4), 733–749. [Google Scholar] [CrossRef]
- Sasaki, N.; Nishizaki, Y.; Ozeki, Y.; Miyahara, T. The Role of Acyl-Glucose in Anthocyanin Modifications. Molecules 2014, 19, 18747–18766. [Google Scholar] [CrossRef]
- Jokioja, J.; Yang, B.; Linderborg, K.M. Acylated Anthocyanins: A Review on Their Bioavailability and Effects on Postprandial Carbohydrate Metabolism and Inflammation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5570–5615. [Google Scholar] [CrossRef]
- Zhao, C.-L.; Yu, Y.-Q.; Chen, Z.-J.; Wen, G.-S.; Wei, F.-G.; Zheng, Q.; Wang, C.-D.; Xiao, X.-L. Stability-Increasing Effects of Anthocyanin Glycosyl Acylation. Food Chem. 2017, 214, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Alappat, B.; Alappat, J. Anthocyanin Pigments: Beyond Aesthetics. Molecules 2020, 25, 5500. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible Berries: Bioactive Components and Their Effect on Human Health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Virgous, C.; Si, H. Synergistic Anti-Inflammatory Effects and Mechanisms of Combined Phytochemicals. J. Nutr. Biochem. 2019, 69, 19–30. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Luján-Barroso, L.; Slimani, N.; Romieu, I.; Touillaud, M.; Kaaks, R.; Teucher, B.; Mattiello, A.; Grioni, S.; et al. Estimation of the Intake of Anthocyanidins and Their Food Sources in the European Prospective Investigation into Cancer and Nutrition (EPIC) Study. Br. J. Nutr. 2011, 106, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Bellocco, E.; Barreca, D.; Laganà, G.; Calderaro, A.; El Lekhlifi, Z.; Chebaibi, S.; Smeriglio, A.; Trombetta, D. Cyanidin-3- O -Galactoside in Ripe Pistachio (Pistachia vera L. Variety Bronte) Hulls: Identification and Evaluation of Its Antioxidant and Cytoprotective Activities. J. Funct. Foods 2016, 27, 376–385. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Acylated Anthocyanins from Edible Sources and Their Applications in Food Systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Sobhani, M.; Farzaei, M.H.; Kiani, S.; Khodarahmi, R. Immunomodulatory; Anti-Inflammatory/Antioxidant Effects of Polyphenols: A Comparative Review on the Parental Compounds and Their Metabolites. Food Rev. Int. 2021, 37, 759–811. [Google Scholar] [CrossRef]
- Ma, Z.; Du, B.; Li, J.; Yang, Y.; Zhu, F. An Insight into Anti-Inflammatory Activities and Inflammation Related Diseases of Anthocyanins: A Review of Both in Vivo and in Vitro Investigations. Int. J. Mol. Sci. 2021, 22, 11076. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A.A.; Sarmast, E.; Fatehi, P.; Jafari, T. Impact of Dietary Anthocyanins on Systemic and Vascular Inflammation: Systematic Review and Meta-Analysis on Randomised Clinical Trials. Food Chem. Toxicol. 2020, 135, 110922. [Google Scholar] [CrossRef]
- Samarpita, S.; Ganesan, R.; Rasool, M. Cyanidin Prevents the Hyperproliferative Potential of Fibroblast-like Synoviocytes and Disease Progression via Targeting IL-17A Cytokine Signalling in Rheumatoid Arthritis. Toxicol. Appl. Pharmacol. 2020, 391, 114917. [Google Scholar] [CrossRef] [PubMed]
- Hair, R.; Sakaki, J.R.; Chun, O.K. Anthocyanins, Microbiome and Health Benefits in Aging. Molecules 2021, 26, 537. [Google Scholar] [CrossRef] [PubMed]
- Verediano, T.A.; Stampini Duarte Martino, H.; Dias Paes, M.C.; Tako, E. Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients 2021, 13, 1331. [Google Scholar] [CrossRef]
- Chen, K.; Kortesniemi, M.K.; Linderborg, K.M.; Yang, B. Anthocyanins as Promising Molecules Affecting Energy Homeostasis, Inflammation, and Gut Microbiota in Type 2 Diabetes with Special Reference to Impact of Acylation. J. Agric. Food Chem. 2023, 71, 1002–1017. [Google Scholar] [CrossRef]
- Kapoor, P.; Tiwari, A.; Sharma, S.; Tiwari, V.; Sheoran, B.; Ali, U.; Garg, M. Effect of Anthocyanins on Gut Health Markers, Firmicutes-Bacteroidetes Ratio and Short-Chain Fatty Acids: A Systematic Review via Meta-Analysis. Sci. Rep. 2023, 13, 1729. [Google Scholar] [CrossRef]
- Liang, A.; Leonard, W.; Beasley, J.T.; Fang, Z.; Zhang, P.; Ranadheera, C.S. Anthocyanins-Gut Microbiota-Health Axis: A Review. Crit. Rev. Food Sci. Nutr. 2023, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, Y.; Takimoto, H.; Matsumoto, T.; Kawaguchi, K. Potential Use of Dietary Natural Products, Especially Polyphenols, for Improving Type-1 Allergic Symptoms. Curr. Pharm. Des. 2014, 20, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.S. Grape Consumption Supports Immunity in Animals and Humans. J. Nutr. 2009, 139, 1801S–5S. [Google Scholar] [CrossRef]
- Park, K.H.; Gu, D.R.; So, H.S.; Kim, K.J.; Lee, S.H. Dual Role of Cyanidin-3-Glucoside on the Differentiation of Bone Cells. J. Dent. Res. 2015, 94, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Kim, J.-Y.; Cheon, Y.-H.; Baek, J.M.; Ahn, S.-J.; Yoon, K.-H.; Lee, M.S.; Oh, J. Protocatechuic Acid Attenuates Osteoclastogenesis by Downregulating JNK/c-Fos/NFATc1 Signaling and Prevents Inflammatory Bone Loss in Mice: PCA Attenuates Osteoclastogenesis. Phytother. Res. 2016, 30, 604–612. [Google Scholar] [CrossRef]
- Jara, E.; Hidalgo, M.A.; Hancke, J.L.; Hidalgo, A.I.; Brauchi, S.; Nuñez, L.; Villalobos, C.; Burgos, R.A. Delphinidin Activates NFAT and Induces IL-2 Production through SOCE in T Cells. Cell Biochem. Biophys. 2014, 68, 497–509. [Google Scholar] [CrossRef]
- Hou, D.-X.; Kai, K.; Li, J.-J.; Lin, S.; Terahara, N.; Wakamatsu, M.; Fujii, M.; Young, M.R.; Colburn, N. Anthocyanidins Inhibit Activator Protein 1 Activity and Cell Transformation: Structure-Activity Relationship and Molecular Mechanisms. Carcinogenesis 2003, 25, 29–36. [Google Scholar] [CrossRef]
- Hou, D.-X.; Yanagita, T.; Uto, T.; Masuzaki, S.; Fujii, M. Anthocyanidins Inhibit Cyclooxygenase-2 Expression in LPS-Evoked Macrophages: Structure–Activity Relationship and Molecular Mechanisms Involved. Biochem. Pharmacol. 2005, 70, 417–425. [Google Scholar] [CrossRef]
- Kiselova-Kaneva, Y.; Nashar, M.; Roussev, B.; Salim, A.; Hristova, M.; Olczyk, P.; Komosinska-Vassev, K.; Dincheva, I.; Badjakov, I.; Galunska, B.; et al. Sambucus Ebulus (Elderberry) Fruits Modulate Inflammation and Complement System Activity in Humans. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Semmarath, W.; Mapoung, S.; Umsumarng, S.; Arjsri, P.; Srisawad, K.; Thippraphan, P.; Yodkeeree, S.; Dejkriengkraikul, P. Cyanidin-3-O-Glucoside and Peonidin-3-O-Glucoside-Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS-CoV-2 Spike Glycoprotein S1-Induction in Vitro in A549 Lung Cells and THP-1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway. Nutrients 2022, 14, 2738. [Google Scholar]
- Li, J.; Zou, C.; Liu, Y. Amelioration of Ovalbumin-Induced Food Allergy in Mice by Targeted Rectal and Colonic Delivery of Cyanidin-3-O-Glucoside. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.A.; Al-Khalaifah, H.S.; Gouda, A.; Osman, A.; Goda, N.I.A.; Mohammed, H.A.; Darwish, M.I.M.; Hassan, A.M.; Mohamed, S.K.A. Potential Effects of Anthocyanin-Rich Roselle (Hibiscus Sabdariffa L.) Extract on the Growth, Intestinal Histomorphology, Blood Biochemical Parameters, and the Immune Status of Broiler Chickens. Antioxidants (Basel) 2022, 11, 544. [Google Scholar] [CrossRef] [PubMed]
- Trinei, M.; Carpi, A.; Menabo’, R.; Storto, M.; Fornari, M.; Marinelli, A.; Minardi, S.; Riboni, M.; Casciaro, F.; DiLisa, F.; et al. Dietary Intake of Cyanidin-3-Glucoside Induces a Long-Lasting Cardioprotection from Ischemia/Reperfusion Injury by Altering the Microbiota. J. Nutr. Biochem. 2022, 101, 108921. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, V.; Park, M.; Lee, H.-J. Comparative Transcriptome Analysis of the Expression of Antioxidant and Immunity Genes in the Spleen of a Cyanidin 3-O-Glucoside-Treated Alzheimer’s Mouse Model. Antioxidants (Basel) 2021, 10, 1435. [Google Scholar] [CrossRef]
- Shaw, O.M.; Hurst, R.D.; Cooney, J.; Sawyer, G.M.; Dinnan, H.; Martell, S. Boysenberry and Apple Juice Concentrate Reduced Acute Lung Inflammation and Increased M2 Macrophage-Associated Cytokines in an Acute Mouse Model of Allergic Airways Disease. Food Sci. Nutr. 2021, 9, 1491–1503. [Google Scholar] [CrossRef]
- Pitsillou, E.; Liang, J.; Ververis, K.; Lim, K.W.; Hung, A.; Karagiannis, T.C. Identification of Small Molecule Inhibitors of the Deubiquitinating Activity of the SARS-CoV-2 Papain-like Protease: In Silico Molecular Docking Studies and in Vitro Enzymatic Activity Assay. Front. Chem. 2020, 8, 623971. [Google Scholar] [CrossRef]
- Pitsillou, E.; Liang, J.; Ververis, K.; Hung, A.; Karagiannis, T.C. Interaction of Small Molecules with the SARS-CoV-2 Papain-like Protease: In Silico Studies and in Vitro Validation of Protease Activity Inhibition Using an Enzymatic Inhibition Assay. J. Mol. Graph. Model. 2021, 104, 107851. [Google Scholar] [CrossRef]
- Aloud, B.M.; Petkau, J.C.; Yu, L.; McCallum, J.; Kirby, C.; Netticadan, T.; Blewett, H. Effects of Cyanidin 3-O-Glucoside and Hydrochlorothiazide on T-Cell Phenotypes and Function in Spontaneously Hypertensive Rats. Food Funct. 2020, 11, 8560–8572. [Google Scholar] [CrossRef]
- Wang, H.; Li, S.; Zhang, G.; Wu, H.; Chang, X. Potential Therapeutic Effects of Cyanidin-3-O-Glucoside on Rheumatoid Arthritis by Relieving Inhibition of CD38+ NK Cells on Treg Cell Differentiation. Arthritis Res. Ther. 2019, 21, 220. [Google Scholar] [CrossRef]
- Mazewski, C.; Kim, M.S.; Gonzalez de Mejia, E. Anthocyanins, Delphinidin-3-O-Glucoside and Cyanidin-3-O-Glucoside, Inhibit Immune Checkpoints in Human Colorectal Cancer Cells in Vitro and in Silico. Sci. Rep. 2019, 9, 11560. [Google Scholar] [CrossRef]
- Cremonini, E.; Daveri, E.; Mastaloudis, A.; Adamo, A.M.; Mills, D.; Kalanetra, K.; Hester, S.N.; Wood, S.M.; Fraga, C.G.; Oteiza, P.I. Anthocyanins Protect the Gastrointestinal Tract from High Fat Diet-Induced Alterations in Redox Signaling, Barrier Integrity and Dysbiosis. Redox Biol. 2019, 26, 101269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, H.; Yi, J.; Yang, B.; Li, M.; He, D.; Yang, W.; Zhang, Y.; Ni, H. Anti-Tumor Properties of Anthocyanins from Lonicera caerulea ‘Beilei’ Fruit on Human Hepatocellular Carcinoma: In Vitro and in Vivo Study. Biomed. Pharmacother. 2018, 104, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Cimino, F.; Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Busà, R.; Saija, A.; Speciale, A. Cyanidin-3-O-Glucoside Modulates the in Vitro Inflammatory Crosstalk between Intestinal Epithelial and Endothelial Cells. Mediators Inflamm. 2017, 3454023. [Google Scholar] [CrossRef] [PubMed]
- Simonyi, A.; Chen, Z.; Jiang, J.; Zong, Y.; Chuang, D.Y.; Gu, Z.; Lu, C.-H.; Fritsche, K.L.; Greenlief, C.M.; Rottinghaus, G.E.; et al. Inhibition of Microglial Activation by Elderberry Extracts and Its Phenolic Components. Life Sci. 2015, 128, 30–38. [Google Scholar] [CrossRef]
- Pyo, M.Y.; Yoon, S.J.; Yu, Y.; Park, S.; Jin, M. Cyanidin-3-Glucoside Suppresses Th2 Cytokines and GATA-3 Transcription Factor in EL-4 T Cells. Biosci. Biotechnol. Biochem. 2014, 78, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Mace, T.A.; King, S.A.; Ameen, Z.; Elnaggar, O.; Young, G.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Knobloch, T.J.; Weghorst, C.M.; et al. Bioactive Compounds or Metabolites from Black Raspberries Modulate T Lymphocyte Proliferation, Myeloid Cell Differentiation and Jak/STAT Signaling. Cancer Immunol. Immunother. 2014, 63, 889–900. [Google Scholar] [CrossRef]
- Köchling, J.; Schmidt, M.; Rott, Y.; Sagner, M.; Ungefroren, H.; Wittig, B.; Henze, G. Can Anthocyanins Improve Maintenance Therapy of Ph(+) Acute Lymphoblastic Leukaemia? Eur. J. Haematol. 2013, 90, 291–300. [Google Scholar] [CrossRef]
- Bub, A.; Watzl, B.; Blockhaus, M.; Briviba, K.; Liegibel, U.; Müller, H.; Pool-Zobel, B.L.; Rechkemmer, G. Fruit Juice Consumption Modulates Antioxidative Status, Immune Status and DNA Damage. J. Nutr. Biochem. 2003, 14, 90–98. [Google Scholar] [CrossRef]
- Ryyti, R.; Hämäläinen, M.; Leppänen, T.; Peltola, R.; Moilanen, E. Phenolic Compounds Known to Be Present in Lingonberry (Vaccinium Vitis-Idaea L.) Enhance Macrophage Polarization towards the Anti-Inflammatory M2 Phenotype. Biomedicines 2022, 10, 3045. [Google Scholar] [CrossRef]
- Neves, B.R.O.; de Freitas, S.; Borelli, P.; Rogero, M.M.; Fock, R.A. Delphinidin-3-O-Glucoside in Vitro Suppresses NF-ΚB and Changes the Secretome of Mesenchymal Stem Cells Affecting Macrophage Activation. Nutrition 2023, 105, 111853. [Google Scholar] [CrossRef]
- Mazewski, C.; Luna-Vital, D.; Berhow, M.; Gonzalez de Mejia, E. Reduction of Colitis-Associated Colon Carcinogenesis by a Black Lentil Water Extract through Inhibition of Inflammatory and Immunomodulatory Cytokines. Carcinogenesis 2020, 41, 790–803. [Google Scholar] [CrossRef]
- Hyun, K.H.; Gil, K.C.; Kim, S.G.; Park, S.-Y.; Hwang, K.W. Delphinidin Chloride and Its Hydrolytic Metabolite Gallic Acid Promote Differentiation of Regulatory T Cells and Have an Anti-Inflammatory Effect on the Allograft Model. J. Food Sci. 2019, 84, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Dayoub, O.; Le Lay, S.; Soleti, R.; Clere, N.; Hilairet, G.; Dubois, S.; Gagnadoux, F.; Boursier, J.; Martínez, M.C.; Andriantsitohaina, R. Estrogen Receptor α/HDAC/NFAT Axis for Delphinidin Effects on Proliferation and Differentiation of T Lymphocytes from Patients with Cardiovascular Risks. Sci. Rep. 2017, 7, 9378. [Google Scholar] [CrossRef] [PubMed]
- Chamcheu, J.C.; Adhami, V.M.; Esnault, S.; Sechi, M.; Siddiqui, I.A.; Satyshur, K.A.; Syed, D.N.; Dodwad, S.-J.M.; Chaves-Rodriquez, M.-I.; Longley, B.J.; et al. Dual Inhibition of PI3K/Akt and MTOR by the Dietary Antioxidant, Delphinidin, Ameliorates Psoriatic Features in Vitro and in an Imiquimod-Induced Psoriasis-like Disease in Mice. Antioxid. Redox Signal. 2017, 26, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.; Jeong, M.-H.; Jeon, H.; Sung, G.-J.; So, Y.; Kim, I.; Son, J.; Lee, S.-W.; Yoon, H.-G.; Choi, K.-C. Delphinidin Sensitizes Prostate Cancer Cells to TRAIL-Induced Apoptosis, by Inducing DR5 and Causing Caspase-Mediated HDAC3 Cleavage. Oncotarget 2015, 6, 9970–9984. [Google Scholar] [CrossRef] [PubMed]
- Seong, A.-R.; Yoo, J.-Y.; Choi, K.; Lee, M.-H.; Lee, Y.-H.; Lee, J.; Jun, W.; Kim, S.; Yoon, H.-G. Delphinidin, a Specific Inhibitor of Histone Acetyltransferase, Suppresses Inflammatory Signaling via Prevention of NF-ΚB Acetylation in Fibroblast-like Synoviocyte MH7A Cells. Biochem. Biophys. Res. Commun. 2011, 410, 581–586. [Google Scholar] [CrossRef]
- Chung, I.-C.; Yuan, S.-N.; OuYang, C.-N.; Hu, S.-I.; Lin, H.-C.; Huang, K.-Y.; Lin, W.-N.; Chuang, Y.-T.; Chen, Y.-J.; Ojcius, D.M.; et al. EFLA 945 Restricts AIM2 Inflammasome Activation by Preventing DNA Entry for Psoriasis Treatment. Cytokine 2020, 127, 154951. [Google Scholar] [CrossRef]
- Iban-Arias, R.; Sebastian-Valverde, M.; Wu, H.; Lyu, W.; Wu, Q.; Simon, J.; Pasinetti, G.M. Role of Polyphenol-Derived Phenolic Acid in Mitigation of Inflammasome-Mediated Anxiety and Depression. Biomedicines 2022, 10, 1264. [Google Scholar] [CrossRef]
- Wang, J.; Hodes, G.E.; Zhang, H.; Zhang, S.; Zhao, W.; Golden, S.A.; Bi, W.; Menard, C.; Kana, V.; Leboeuf, M.; et al. Epigenetic Modulation of Inflammation and Synaptic Plasticity Promotes Resilience against Stress in Mice. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Eisenstein, M. Microbiome: Bacterial Broadband. Nature 2016, 533, S104–6. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Williamson, G.; Clifford, M.N. Colonic Metabolites of Berry Polyphenols: The Missing Link to Biological Activity? Br. J. Nutr. 2010, 104 Suppl 3, S48–66. [Google Scholar] [CrossRef]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of Polyphenols and Polyphenol-Rich Dietary Sources on F. Han et al. Trends in Food. J. Agric. Food Chem. 2013, 83, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Hanske, L.; Engst, W.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Contribution of Gut Bacteria to the Metabolism of Cyanidin 3-Glucoside in Human Microbiota-Associated Rats. Br. J. Nutr. 2013, 109, 1433–1441. [Google Scholar] [CrossRef]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.-O.; Carbonero, F. Impact of Tart Cherries Polyphenols on the Human Gut Microbiota and Phenolic Metabolites in Vitro and in Vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, L.; Angelino, D.; Vivas, E.I.; Kerby, R.L.; García-Viguera, C.; Del Rio, D.; Rey, F.E.; Mena, P. Differential Catabolism of an Anthocyanin-Rich Elderberry Extract by Three Gut Microbiota Bacterial Species. J. Agric. Food Chem. 2020, 68, 1837–1843. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut Microbiota Functions: Metabolism of Nutrients and Other Food Components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Rimac Brnčić, S.; El, S.N.; Karakaya, S.; Simsek, S.; Manach, C.; Wiczkowski, W.; de Pascual-Teresa, S. A Review of Factors Affecting Anthocyanin Bioavailability: Possible Implications for the Inter-Individual Variability. Foods 2019, 9, 2. [Google Scholar] [CrossRef]
- Han, F.; Yang, P.; Wang, H.; Fernandes, I.; Mateus, N.; Liu, Y. Digestion and Absorption of Red Grape and Wine Anthocyanins through the Gastrointestinal Tract. Trends Food Sci. Technol. 2019, 83, 211–224. [Google Scholar] [CrossRef]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of Anthocyanins and Consequent Effects on the Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Q.; Zhao, T.; Zhang, Z.; Mao, G.; Feng, W.; Wu, X.; Yang, L. Biotransformation and Metabolism of Three Mulberry Anthocyanin Monomers by Rat Gut Microflora. Food Chem. 2017, 237, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants (Basel) 2021, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of Anthocyanins and Derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Fleschhut, J.; Kratzer, F.; Rechkemmer, G.; Kulling, S.E. Stability and Biotransformation of Various Dietary Anthocyanins in Vitro. Eur. J. Nutr. 2006, 45, 7–18. [Google Scholar] [CrossRef]
- Zhao, R.; Shen, G.X. Impact of Anthocyanin Component and Metabolite of Saskatoon Berry on Gut Microbiome and Relationship with Fecal Short Chain Fatty Acids in Diet-Induced Insulin Resistant Mice. J. Nutr. Biochem. 2023, 111, 109201. [Google Scholar] [CrossRef]
- Dong, A.; Lin, C.-W.; Echeveste, C.E.; Huang, Y.-W.; Oshima, K.; Yearsley, M.; Chen, X.; Yu, J.; Wang, L.-S. Protocatechuic Acid, a Gut Bacterial Metabolite of Black Raspberries, Inhibits Adenoma Development and Alters Gut Microbiome Profiles in Apcmin/+ Mice. J. Cancer Prev. 2022, 27, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhu, R.; Ye, X.; Sun, Y.; Tang, Q.; Liu, Y.; Yan, F.; Yu, T.; Zheng, X.; Tu, P. Food-Derived Cyanidin-3-O-Glucoside Reverses Microplastic Toxicity via Promoting Discharge and Modulating the Gut Microbiota in Mice. Food Funct. 2022, 13, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Xu, J.; Wang, L.; Chen, C.; Chen, P. Anti-Inflammatory Effects of Purple Sweet Potato Anthocyanin Extract in DSS-Induced Colitis: Modulation of Commensal Bacteria and Attenuated Bacterial Intestinal Infection. Food Funct. 2021, 12, 11503–11514. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Jing, N.; Jiang, G.; Liu, Z. Biostimulating Gut Microbiome with Bilberry Anthocyanin Combo to Enhance Anti-PD-L1 Efficiency against Murine Colon Cancer. Microorganisms 2020, 8, 175. [Google Scholar] [CrossRef]
- Nakano, H.; Wu, S.; Sakao, K.; Hara, T.; He, J.; Garcia, S.; Shetty, K.; Hou, D.-X. Bilberry Anthocyanins Ameliorate NAFLD by Improving Dyslipidemia and Gut Microbiome Dysbiosis. Nutrients 2020, 12, 3252. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, T.T.Y.; Tang, Q.; Xue, C.; Li, R.W.; Wu, V.C.H. Malvidin 3-glucoside Modulated Gut Microbial Dysbiosis and Global Metabolome Disrupted in a Murine Colitis Model Induced by Dextran Sulfate Sodium. Mol. Nutr. Food Res. 2019, 63, 1900455. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.-X.; Wu, S. The Effects and Mechanisms of Cyanidin-3-Glucoside and Its Phenolic Metabolites in Maintaining Intestinal Integrity. Antioxidants (Basel) 2019, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Overall, J.; Bonney, S.; Wilson, M.; Beermann, A., III; Grace, M.; Esposito, D.; Lila, M.; Komarnytsky, S. Metabolic Effects of Berries with Structurally Diverse Anthocyanins. Int. J. Mol. Sci. 2017, 18, 422. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.E.; Gibson, G.R.; de Pascual-Teresa, S. Metabolism of Anthocyanins by Human Gut Microflora and Their Influence on Gut Bacterial Growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Wang, H.; Cao, G.; Prior, R.L. Oxygen Radical Absorbing Capacity of Anthocyanins. J. Agric. Food Chem. 1997, 45, 304–309. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Stintzing, A.S.; Carle, R.; Frei, B.; Wrolstad, R.E. Colour and Antioxidant Properties of Cyanidin-Based Anthocyanin Pigments. J. Agric. Food Chem. 2002, 50, 6172–6181. [Google Scholar] [CrossRef]
- Tamura, H.; Yamagami, A. Antioxidative Activity of Monoacylated Anthocyanins Isolated from Muscat Bailey A Grape. J. Agric. Food Chem. 1994, 42, 1612–1615. [Google Scholar] [CrossRef]
- Tsuda, T.; Shiga, K.; Ohshima, K.; Kawakishi, S.; Osawa, T. Inhibition of Lipid Peroxidation and the Active Oxygen Radical Scavenging Effect of Anthocyanin Pigments Isolated from Phaseolus Vulgaris L. Biochem. Pharmacol. 1996, 52, 1033–1039. [Google Scholar] [CrossRef]
- Brown, J.E.; Kelly, M.F. Inhibition of Lipid Peroxidation by Anthocyanins, Anthocyanidins and Their Phenolic Degradation Products. Eur. J. Lipid Sci. Technol. 2007, 109, 66–71. [Google Scholar] [CrossRef]
- Speciale, A.; Chirafisi, J.; Saija, A.; Cimino, F. Nutritional Antioxidants and Adaptive Cell Responses: An Update. Curr. Mol. Med. 2011, 11, 770–789. [Google Scholar] [CrossRef] [PubMed]
- Speciale, A.; Anwar, S.; Canali, R.; Chirafisi, J.; Saija, A.; Virgili, F.; Cimino, F. Cyanidin-3-O -Glucoside Counters the Response to TNF-Alpha of Endothelial Cells by Activating Nrf2 Pathway. Mol. Nutr. Food Res. 2013, 57, 1979–1987. [Google Scholar] [CrossRef]
- Vendrame, S.; Klimis-Zacas, D. Anti-Inflammatory Effect of Anthocyanins via Modulation of Nuclear Factor- B and Mitogen-Activated Protein Kinase Signaling Cascades. Nutr. Rev. 2015, 73, 348–358. [Google Scholar] [CrossRef]
- Poulose, S.M.; Fisher, D.R.; Larson, J.; Bielinski, D.F.; Rimando, A.M.; Carey, A.N.; Schauss, A.G.; Shukitt-Hale, B. Anthocyanin-Rich Açai (Euterpe Oleracea Mart.) Fruit Pulp Fractions Attenuate Inflammatory Stress Signaling in Mouse Brain BV-2 Microglial Cells. J. Agric. Food Chem. 2012, 60, 1084–1093. [Google Scholar] [CrossRef]
- Hassimotto, N.M.A.; Moreira, V.; Nascimento, N.G. do; Souto, P.C.M. de C.; Teixeira, C.; Lajolo, F.M. Inhibition of Carrageenan-Induced Acute Inflammation in Mice by Oral Administration of Anthocyanin Mixture from Wild Mulberry and Cyanidin-3-Glucoside. Biomed Res. Int. 2013, 1–10. [Google Scholar]
- Cui, H.-X.; Chen, J.-H.; Li, J.-W.; Cheng, F.-R.; Yuan, K. Protection of Anthocyanin from Myrica Rubra against Cerebral Ischemia-Reperfusion Injury via Modulation of the TLR4/NF-ΚB and NLRP3 Pathways. Molecules 2018, 23, 1788. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, Y.; He, Q.; Li, L.; Xie, H.; Zhao, Y.; Zhao, J. Nrf2 Inhibits NLRP3 Inflammasome Activation through Regulating Trx1/TXNIP Complex in Cerebral Ischemia Reperfusion Injury. Behav. Brain Res. 2018, 336, 32–39. [Google Scholar] [CrossRef]
- Putta, S.; Yarla, N.S.; Peluso, I.; Tiwari, D.K.; Reddy, G.V.; Giri, P.V.; Kumar, N.; Malla, R.; Rachel, V.; Bramhachari, P.V.; et al. Anthocyanins: Multi-Target Agents for Prevention and Therapy of Chronic Diseases. Curr. Pharm. Des. 2018, 23, 6321–6346. [Google Scholar] [CrossRef]
- Zhou, P.; Pan, Y.; Yang, W.; Yang, B.; Ou, S.; Liu, P.; Zheng, J. Hepatoprotective Effect of Cyanidin-3-O-Glucoside–Lauric Acid Ester against H2O2-Induced Oxidative Damage in LO2 Cells. J. Funct. Foods 2023, 107, 105642. [Google Scholar] [CrossRef]
- Romualdo, G.R.; de Souza, I.P.; de Souza, L.V.; Prata, G.B.; Fraga-Silva, T.F. de C.; Sartori, A.; Borguini, R.G.; Santiago, M.C.P. de A.; Fernandes, A.A.H.; Cogliati, B.; et al. Beneficial Effects of Anthocyanin-Rich Peels of Myrtaceae Fruits on Chemically-Induced Liver Fibrosis and Carcinogenesis in Mice. Food Res. Int. 2021, 139, 109964. [Google Scholar] [CrossRef]
- Zuo, A.; Wang, S.; Liu, L.; Yao, Y.; Guo, J. Understanding the Effect of Anthocyanin Extracted from Lonicera Caerulea L. on Alcoholic Hepatosteatosis. Biomed. Pharmacother. 2019, 117, 109087. [Google Scholar] [CrossRef] [PubMed]
- Popović, D.; Kocić, G.; Katić, V.; Zarubica, A.; Veličković, L.J.; Ničković, V.P.; Jović, A.; Veljković, A.; Petrović, V.; Rakić, V.; et al. Anthocyanins Protect Hepatocytes against CCl4-Induced Acute Liver Injury in Rats by Inhibiting pro-Inflammatory Mediators, Polyamine Catabolism, Lipocalin-2, and Excessive Proliferation of Kupffer Cells. Antioxidants (Basel) 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, L.; Hogea, L.M.; Călina, D.; Andreescu, N.; Grădinaru, R.; Ştefănescu, R. Modern Treatment Approaches in Psychoses. Pharmacogenetic, Neuroimagistic and Clinical Implications. Farmacia 2017, 65, 75–81. [Google Scholar]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Russo, M.V.; McGavern, D.B. Immune Surveillance of the CNS Following Infection and Injury. Trends Immunol. 2015, 36, 637–650. [Google Scholar] [CrossRef]
- Min, J.; Yu, S.-W.; Baek, S.-H.; Nair, K.M.; Bae, O.-N.; Bhatt, A.; Kassab, M.; Nair, M.G.; Majid, A. Neuroprotective Effect of Cyanidin-3-O-Glucoside Anthocyanin in Mice with Focal Cerebral Ischemia. Neurosci. Lett. 2011, 500, 157–161. [Google Scholar] [CrossRef]
- Cásedas, G.; González-Burgos, E.; Smith, C.; López, V.; Gómez-Serranillos, M.P. Regulation of Redox Status in Neuronal SH-SY5Y Cells by Blueberry (Vaccinium Myrtillus L.) Juice, Cranberry (Vaccinium Macrocarpon A.) Juice and Cyanidin. Food Chem. Toxicol. 2018, 118, 572–580. [Google Scholar] [CrossRef]
- Pacheco, S.M.; Soares, M.S.P.; Gutierres, J.M.; Gerzson, M.F.B.; Carvalho, F.B.; Azambuja, J.H.; Schetinger, M.R.C.; Stefanello, F.M.; Spanevello, R.M. Anthocyanins as a Potential Pharmacological Agent to Manage Memory Deficit, Oxidative Stress and Alterations in Ion Pump Activity Induced by Experimental Sporadic Dementia of Alzheimer’s Type. J. Nutr. Biochem. 2018, 56, 193–204. [Google Scholar] [CrossRef]
- Amin, F.U.; Shah, S.A.; Badshah, H.; Khan, M.; Kim, M.O. Anthocyanins Encapsulated by PLGA@PEG Nanoparticles Potentially Improved Its Free Radical Scavenging Capabilities via P38/JNK Pathway against Aβ1–42-Induced Oxidative Stress. J. Nanobiotechnology 2017, 15. [Google Scholar] [CrossRef]
- Kim, M.J.; Rehman, S.U.; Amin, F.U.; Kim, M.O. Enhanced Neuroprotection of Anthocyanin-Loaded PEG-Gold Nanoparticles against Aβ1-42-Induced Neuroinflammation and Neurodegeneration via the NF-KB /JNK/GSK3β Signaling Pathway. Nanomedicine 2017, 13, 2533–2544. [Google Scholar] [CrossRef]
- Mulabagal, V.; Lang, G.A.; DeWitt, D.L.; Dalavoy, S.S.; Nair, M.G. Anthocyanin Content, Lipid Peroxidation and Cyclooxygenase Enzyme Inhibitory Activities of Sweet and Sour Cherries. J. Agric. Food Chem. 2009, 57, 1239–1246. [Google Scholar] [CrossRef]
- Joseph, J.A.; Arendash, G.; Gordon, M.; Diamond, D.; Shukitt-Hale, B.; Morgan, D.; Denisova, N.A. Blueberry Supplementation Enhances Signaling and Prevents Behavioral Deficits in an Alzheimer Disease Model. Nutr. Neurosci. 2003, 6, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cassidy, A.; Schwarzschild, M.A.; Rimm, E.B.; Ascherio, A. Habitual Intake of Dietary Flavonoids and Risk of Parkinson Disease. Neurology 2012, 78, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Krikorian, R.; Nash, T.A.; Shidler, M.D.; Shukitt-Hale, B.; Joseph, J.A. Concord Grape Juice Supplementation Improves Memory Function in Older Adults with Mild Cognitive Impairment. Br. J. Nutr. 2010, 103, 730–734. [Google Scholar] [CrossRef]
- Krikorian, R.; Shidler, M.D.; Nash, T.A.; Kalt, W.; Vinqvist-Tymchuk, M.R.; Shukitt-Hale, B.; Joseph, J.A. Blueberry Supplementation Improves Memory in Older Adults. J. Agric. Food Chem. 2010, 58, 3996–4000. [Google Scholar] [CrossRef] [PubMed]
- Ockermann, P.; Headley, L.; Lizio, R.; Hansmann, J. A Review of the Properties of Anthocyanins and Their Influence on Factors Affecting Cardiometabolic and Cognitive Health. Nutrients 2021, 13, 2831. [Google Scholar] [CrossRef]
- Yang, Y.; Andrews, M.C.; Hu, Y.; Wang, D.; Qin, Y.; Zhu, Y.; Ni, H.; Ling, W. Anthocyanin Extract from Black Rice Significantly Ameliorates Platelet Hyperactivity and Hypertriglyceridemia in Dyslipidemic Rats Induced by High Fat Diets. J. Agric. Food Chem. 2011, 59, 6759–6764. [Google Scholar] [CrossRef] [PubMed]
- García-Conesa, M.-T.; Chambers, K.; Combet, E.; Pinto, P.; Garcia-Aloy, M.; Andrés-Lacueva, C.; de Pascual-Teresa, S.; Mena, P.; Konic Ristic, A.; Hollands, W.; et al. Meta-Analysis of the Effects of Foods and Derived Products Containing Ellagitannins and Anthocyanins on Cardiometabolic Biomarkers: Analysis of Factors Influencing Variability of the Individual Responses. Int. J. Mol. Sci. 2018, 19, 694. [Google Scholar] [CrossRef]
- Luís, Â.; Domingues, F.; Pereira, L. Association between Berries Intake and Cardiovascular Diseases Risk Factors: A Systematic Review with Meta-Analysis and Trial Sequential Analysis of Randomized Controlled Trials. Food Funct. 2018, 9, 740–757. [Google Scholar] [CrossRef]
- Cassidy, A.; Mukamal, K.J.; Liu, L.; Franz, M.; Eliassen, A.H.; Rimm, E.B. High Anthocyanin Intake Is Associated with a Reduced Risk of Myocardial Infarction in Young and Middle-Aged Women. Circulation 2013, 127, 188–196. [Google Scholar] [CrossRef]
- Cassidy, A.; Bertoia, M.; Chiuve, S.; Flint, A.; Forman, J.; Rimm, E.B. Habitual Intake of Anthocyanins and Flavanones and Risk of Cardiovascular Disease in Men. Am. J. Clin. Nutr. 2016, 104, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Choi, S.; Lee, Y.; Lee, E.; Park, M.; Park, K.; Kim, C.-S.; Lim, Y.; Park, J.-T.; Jeon, B. Anthocyanin-Rich Extract from Red Chinese Cabbage Alleviates Vascular Inflammation in Endothelial Cells and APO E−/− Mice. Int. J. Mol. Sci. 2018, 19, 816. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wei, X.; Yan, X.; Jin, T.; Ling, W. Protocatechuic Acid, a Metabolite of Anthocyanins, Inhibits Monocyte Adhesion and Reduces Atherosclerosis in Apolipoprotein E-Deficient Mice. J. Agric. Food Chem. 2010, 58, 12722–12728. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Kang, J.; Xie, C.; Burris, R.; Ferguson, M.E.; Badger, T.M.; Nagarajan, S. Dietary Blueberries Attenuate Atherosclerosis in Apolipoprotein E-Deficient Mice by Upregulating Antioxidant Enzyme Expression. J. Nutr. 2010, 140, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Trinei, M.; Fornari, M.; Calvenzani, V.; Marinelli, A.; Micheli, L.A.; Pilu, R.; Matros, A.; Mock, H.-P.; Tonelli, C.; et al. Dietary Cyanidin 3-Glucoside from Purple Corn Ameliorates Doxorubicin-Induced Cardiotoxicity in Mice. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 462–469. [Google Scholar] [CrossRef]
- Skates, E.; Overall, J.; DeZego, K.; Wilson, M.; Esposito, D.; Lila, M.A.; Komarnytsky, S. Berries Containing Anthocyanins with Enhanced Methylation Profiles Are More Effective at Ameliorating High Fat Diet-Induced Metabolic Damage. Food Chem. Toxicol. 2018, 111, 445–453. [Google Scholar] [CrossRef]
- Bognar, E.; Sarszegi, Z.; Szabo, A.; Debreceni, B.; Kalman, N.; Tucsek, Z.; Sumegi, B.; Gallyas, F. Antioxidant and Anti-Inflammatory Effects in RAW264.7 Macrophages of Malvidin, a Major Red Wine Polyphenol. PLoS One 2013, 8, e65355. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ashford, M.L.J. AMPK: Regulating Energy Balance at the Cellular and Whole Body Levels. Physiology (Bethesda) 2014, 29, 99–107. [Google Scholar] [CrossRef]
- López, M. Hypothalamic AMPK and Energy Balance. Eur. J. Clin. Invest. 2018, 48, e12996. [Google Scholar] [CrossRef]
- Park, S.; Kang, S.; Jeong, D.-Y.; Jeong, S.-Y.; Park, J.J.; Yun, H.S. Cyanidin and Malvidin in Aqueous Extracts of Black Carrots Fermented with Aspergillus Oryzae Prevent the Impairment of Energy, Lipid and Glucose Metabolism in Estrogen-Deficient Rats by AMPK Activation. Genes Nutr. 2015, 10. [Google Scholar] [CrossRef]
- Wu, T.; Jiang, Z.; Yin, J.; Long, H.; Zheng, X. Anti-Obesity Effects of Artificial Planting Blueberry (Vaccinium Ashei) Anthocyanin in High-Fat Diet-Treated Mice. Int. J. Food Sci. Nutr. 2016, 67, 257–264. [Google Scholar] [CrossRef]
- Jamar, G.; Estadella, D.; Pisani, L.P. Contribution of Anthocyanin-Rich Foods in Obesity Control through Gut Microbiota Interactions: Anthocyanin-Rich Foods in Obesity Control. Biofactors 2017, 43, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Kang, M.; Xie, Q.; Xu, B.; Sun, C.; Chen, K.; Wu, Y. Anthocyanins from Chinese Bayberry Extract Protect β Cells from Oxidative Stress-Mediated Injury via HO-1 Upregulation. J. Agric. Food Chem. 2011, 59, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Fu, D.X.; Wilkinson, M.; Simmons, B.; Wu, M.; Betts, N.M.; Du, M.; Lyons, T.J. Strawberries Decrease Atherosclerotic Markers in Subjects with Metabolic Syndrome. Nutr. Res. 2010, 30, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Gu, L.; Hager, T.J.; Hager, A.; Howard, L.R. Whole Berries versus Berry Anthocyanins: Interactions with Dietary Fat Levels in the C57BL/6J Mouse Model of Obesity. J. Agric. Food Chem 2008, 56, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Daugherty, A.; Kristo, A.S.; Riso, P.; Klimis-Zacas, D. Wild Blueberry (Vaccinium Angustifolium) Consumption Improves Inflammatory Status in the Obese Zucker Rat Model of the Metabolic Syndrome. J. Nutr. Biochem. 2013, 24, 1508–1512. [Google Scholar] [CrossRef]
- Tomay, F.; Marinelli, A.; Leoni, V.; Caccia, C.; Matros, A.; Mock, H.-P.; Tonelli, C.; Petroni, K. Purple Corn Extract Induces Long-Lasting Reprogramming and M2 Phenotypic Switch of Adipose Tissue Macrophages in Obese Mice. J. Transl. Med. 2019, 17. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The Case for Anthocyanin Consumption to Promote Human Health: A Review: Anthocyanins and Human Health. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Lin, B.-W.; Gong, C.-C.; Song, H.-F.; Cui, Y.-Y. Effects of Anthocyanins on the Prevention and Treatment of Cancer: Anthocyanins and Prevention & Treatment of Cancer. Br. J. Pharmacol. 2017, 174, 1226–1243. [Google Scholar]
- Dreesen, O.; Brivanlou, A.H. Signaling Pathways in Cancer and Embryonic Stem Cells. Stem Cell Rev. 2007, 3, 7–17. [Google Scholar] [CrossRef]
- Kausar, H.; Jeyabalan, J.; Aqil, F.; Chabba, D.; Sidana, J.; Singh, I.P.; Gupta, R.C. Berry Anthocyanidins Synergistically Suppress Growth and Invasive Potential of Human Non-Small-Cell Lung Cancer Cells. Cancer Lett. 2012, 325, 54–62. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Son, Y.-O.; Wang, X.; Divya, S.P.; Joseph, B.; Hitron, J.A.; Wang, L.; Kim, D.; Yin, Y.; Roy, R.V.; et al. Cyanidin-3-Glucoside Inhibits UVB-Induced Oxidative Damage and Inflammation by Regulating MAP Kinase and NF-ΚB Signaling Pathways in SKH-1 Hairless Mice Skin. Toxicol. Appl. Pharmacol. 2014, 280, 127–137. [Google Scholar] [CrossRef]
- Choi, Y.J.; Choi, Y.J.; Kim, N.; Nam, R.H.; Lee, S.; Lee, H.S.; Lee, H.-N.; Surh, Y.-J.; Lee, D.H. Açaí Berries Inhibit Colon Tumorigenesis in Azoxymethane/Dextran Sulfate Sodium-Treated Mice. Gut Liver 2017, 11, 243–252. [Google Scholar] [CrossRef]
- Fragoso, M.F.; Romualdo, G.R.; Vanderveer, L.A.; Franco-Barraza, J.; Cukierman, E.; Clapper, M.L.; Carvalho, R.F.; Barbisan, L.F. Lyophilized Açaí Pulp (Euterpe Oleracea Mart) Attenuates Colitis-Associated Colon Carcinogenesis While Its Main Anthocyanin Has the Potential to Affect the Motility of Colon Cancer Cells. Food Chem. Toxicol. 2018, 121, 237–245. [Google Scholar] [CrossRef]
- Long, H.-L.; Zhang, F.-F.; Wang, H.-L.; Yang, W.-S.; Hou, H.-T.; Yu, J.-K.; Liu, B. Mulberry Anthocyanins Improves Thyroid Cancer Progression Mainly by Inducing Apoptosis and Autophagy Cell Death. Kaohsiung J. Med. Sci. 2018, 34, 255–262. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The Therapeutic Potential of Anthocyanins: Current Approaches Based on Their Molecular Mechanism of Action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef]
- Lippert, E.; Ruemmele, P.; Obermeier, F.; Goelder, S.; Kunst, C.; Rogler, G.; Dunger, N.; Messmann, H.; Hartmann, A.; Endlicher, E. Anthocyanins Prevent Colorectal Cancer Development in a Mouse Model. Digestion 2017, 95, 275–280. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, Y.; Zhang, W.; Peng, X.; Zhou, J.; Li, F.; Han, B.; Liu, X.; Ou, Y.; Yu, X. Delphinidin Induced Protective Autophagy via MTOR Pathway Suppression and AMPK Pathway Activation in HER-2 Positive Breast Cancer Cells. BMC Cancer 2018, 18. [Google Scholar] [CrossRef]
- Nomi, Y; Iwasaki-Kurashige, K; Matsumoto, H. Therapeutic Effects of Anthocyanins for Vision and Eye Health. Molecules 2019, 24, 3311. [Google Scholar] [CrossRef]
- Miyake, S.; Takahashi, N.; Sasaki, M.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Vision Preservation during Retinal Inflammation by Anthocyanin-Rich Bilberry Extract: Cellular and Molecular Mechanism. Lab. Invest. 2012, 92, 102–109. [Google Scholar] [CrossRef]
- Shim, S.H.; Kim, J.M.; Choi, C.Y.; Kim, C.Y.; Park, K.H. Ginkgo Biloba Extract and Bilberry Anthocyanins Improve Visual Function in Patients with Normal Tension Glaucoma. J. Med. Food 2012, 15, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Ohguro, H.; Ohguro, I.; Katai, M.; Tanaka, S. Two-Year Randomized, Placebo-Controlled Study of Black Currant Anthocyanins on Visual Field in Glaucoma. Ophthalmologica 2012, 228, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Thiraphatthanavong, P.; Wattanathorn, J.; Muchimapura, S.; Wipawee, T.-M.; Wannanon, P.; Terdthai, T.-U.; Suriharn, B.; Lertrat, K. Preventive Effect of Zea Mays L. (Purple Waxy Corn) on Experimental Diabetic Cataract. Biomed Res. Int. 2014, 507435. [Google Scholar]
- Paik, S.-S.; Jeong, E.; Jung, S.W.; Ha, T.J.; Kang, S.; Sim, S.; Jeon, J.H.; Chun, M.-H.; Kim, I.-B. Anthocyanins from the Seed Coat of Black Soybean Reduce Retinal Degeneration Induced by N-Methyl-N-Nitrosourea. Exp. Eye Res. 2012, 97, 55–62. [Google Scholar] [CrossRef]
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of Polyphenolic Profile, Antioxidant Activity and Antibacterial Properties of Maqui [Aristotelia Chilensis (Molina) Stuntz] a Chilean Blackberry: Antioxidant and Antibacterial Properties of Maqui. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, L.; Olabarrieta, I.; de Marañón, I.M. Antimicrobial Assays of Natural Extracts and Their Inhibitory Effect against Listeria Innocua and Fish Spoilage Bacteria, after Incorporation into Biopolymer Edible Films. Int. J. Food Microbiol. 2012, 158, 58–64. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Dussault, D.; Sylvain, J.-F.; Lacroix, M. Antimicrobial Effect of Cranberry Juice and Extracts. Food Control 2011, 22, 1413–1418. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Kähkönen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.M. Antimicrobial Properties of Phenolic Compounds from Berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as Antimicrobial Agents of Natural Plant Origin. Nat. Prod. Commun. 2011, 6, 149–156. [Google Scholar] [CrossRef]
- Manolescu, B.N.; Oprea, E.; Mititelu, M.; Ruta, L.L.; Farcasanu, I.C. Dietary Anthocyanins and Stroke: A Review of Pharmacokinetic and Pharmacodynamic Studies. Nutrients 2019, 11, 1479. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Soares, S.; Mateus, N.; Rivas-Gonzalo, J.; Escribano-Bailón, M.T.; Freitas, V. de New Anthocyanin–Human Salivary Protein Complexes. Langmuir 2015, 31, 8392–8401. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Staticin VitroDigestion Method Suitable for Food – an International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Sandhu, A.; Huang, Y.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. The Effect of Dietary Factors on Strawberry Anthocyanins Oral Bioavailability. Food Funct. 2017, 8, 3970–3979. [Google Scholar] [CrossRef]
- Kamonpatana, K.; Giusti, M.M.; Chitchumroonchokchai, C.; MorenoCruz, M.; Riedl, K.M.; Kumar, P.; Failla, M.L. Susceptibility of Anthocyanins to Ex Vivo Degradation in Human Saliva. Food Chem. 2012, 135, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Kamonpatana, K.; Failla, M.L.; Kumar, P.S.; Giusti, M.M. Anthocyanin Structure Determines Susceptibility to Microbial Degradation and Bioavailability to the Buccal Mucosa. J. Agric. Food Chem. 2014, 62, 6903–6910. [Google Scholar] [CrossRef] [PubMed]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling Anthocyanin Bioavailability for Human Health. Annu. Rev. Food Sci. Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef]
- Pedersen, A.M.; Bardow, A.; Jensen, S.B.; Nauntofte, B. Saliva and Gastrointestinal Functions of Taste, Mastication, Swallowing and Digestion: Saliva and Gastrointestinal Functions. Oral Dis. 2002, 8, 117–129. [Google Scholar] [CrossRef]
- Henriques, J.F.; Serra, D.; Dinis, T.C.P.; Almeida, L.M. The Anti-Neuroinflammatory Role of Anthocyanins and Their Metabolites for the Prevention and Treatment of Brain Disorders. Int. J. Mol. Sci. 2020, 21, 8653. [Google Scholar] [CrossRef]
- He, J.; Magnuson, B.A.; Lala, G.; Tian, Q.; Schwartz, S.J.; Giusti, M.M. Intact Anthocyanins and Metabolites in Rat Urine and Plasma after 3 Months of Anthocyanin Supplementation. Nutr. Cancer 2006, 54, 3–12. [Google Scholar] [CrossRef]
- Borges, G.; Roowi, S.; Rouanet, J.-M.; Duthie, G.G.; Lean, M.E.J.; Crozier, A. The Bioavailability of Raspberry Anthocyanins and Ellagitannins in Rats. Mol. Nutr. Food Res. 2007, 51, 714–725. [Google Scholar] [CrossRef]
- Stalmach, A.; Edwards, C.A.; Wightman, J.D.; Crozier, A. Gastrointestinal Stability and Bioavailability of (Poly)Phenolic Compounds Following Ingestion of Concord Grape Juice by Humans. Mol. Nutr. Food Res. 2012, 56, 497–509. [Google Scholar] [CrossRef]
- Oliveira, H.; Roma-Rodrigues, C.; Santos, A.; Veigas, B.; Brás, N.; Faria, A.; Calhau, C.; de Freitas, V.; Baptista, P.V.; Mateus, N.; et al. GLUT1 and GLUT3 Involvement in Anthocyanin Gastric Transport- Nanobased Targeted Approach. Sci. Rep. 2019, 9, 789. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Mattivi, F. The Interaction of Anthocyanins with Bilitranslocase. Biochem. Biophys. Res. Commun. 2002, 296, 631–636. [Google Scholar] [CrossRef]
- Krga, I.; Milenkovic, D. Anthocyanins: From Sources and Bioavailability to Cardiovascular-Health Benefits and Molecular Mechanisms of Action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Lamaison, J.-L.; Rémésy, C. Anthocyanins Are Efficiently Absorbed from the Stomach in Anesthetized Rats. J. Nutr. 2003, 133, 4178–4182. [Google Scholar] [CrossRef]
- Shahidi, F.; Peng, H. Bioaccessibility and Bioavailability of Phenolic Compounds. J. Food Bioact. 2018, 4. [Google Scholar] [CrossRef]
- Hribar, U.; Ulrih, N.P. The Metabolism of Anthocyanins. Curr. Drug Metab. 2014, 15, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.-M. The Role of Metabolism (and the Microbiome) in Defining the Clinical Efficacy of Dietary Flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, S.; Terdoslavich, M.; Franca, R.; Vanzo, A.; Tramer, F.; Braidot, E.; Petrussa, E.; Vianello, A. Bioavailability of Flavonoids: A Review of Their Membrane Transport and the Function of Bilitranslocase in Animal and Plant Organisms. Curr. Drug Metab. 2009, 10, 369–394. [Google Scholar] [CrossRef]
- Zou, T.-B.; Feng, D.; Song, G.; Li, H.-W.; Tang, H.-W.; Ling, W.-H. The Role of Sodium-Dependent Glucose Transporter 1 and Glucose Transporter 2 in the Absorption of Cyanidin-3-o-β-Glucoside in Caco-2 Cells. Nutrients 2014, 6, 4165–4177. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Capanoglu, E.; Grootaert, C.; Van Camp, J. Anthocyanin Absorption and Metabolism by Human Intestinal Caco-2 Cells—A Review. Int. J. Mol. Sci. 2015, 16, 21555–21574. [Google Scholar] [CrossRef]
- Figueira, I.; Menezes, R.; Macedo, D.; Costa, I.; Dos Santos, C.N. Polyphenols beyond Barriers: A Glimpse into the Brain. Curr. Neuropharmacol. 2017, 15, 562–594. [Google Scholar] [CrossRef]
- Faria, A.; Pestana, D.; Azevedo, J.; Martel, F.; de Freitas, V.; Azevedo, I.; Mateus, N.; Calhau, C. Absorption of Anthocyanins through Intestinal Epithelial Cells - Putative Involvement of GLUT2. Mol. Nutr. Food Res. 2009, 53, 1430–1437. [Google Scholar] [CrossRef]
- Alzaid, F.; Cheung, H.-M.; Preedy, V.R.; Sharp, P.A. Regulation of Glucose Transporter Expression in Human Intestinal Caco-2 Cells Following Exposure to an Anthocyanin-Rich Berry Extract. PLoS One 2013, 8, e78932. [Google Scholar] [CrossRef]
- Lewandowska, U.; Szewczyk, K.; Hrabec, E.; Janecka, A.; Gorlach, S. Overview of Metabolism and Bioavailability Enhancement of Polyphenols. J. Agric. Food Chem. 2013, 61, 12183–12199. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human Metabolism and Elimination of the Anthocyanin, Cyanidin-3-Glucoside: A 13C-Tracer Study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef]
- Felgines, C.; Krisa, S.; Mauray, A.; Besson, C.; Lamaison, J.-L.; Scalbert, A.; Mérillon, J.-M.; Texier, O. Radiolabelled Cyanidin 3-O-Glucoside Is Poorly Absorbed in the Mouse. Br. J. Nutr. 2010, 103, 1738–1745. [Google Scholar] [CrossRef]
- Kalt, W.; McDonald, J.E.; Vinqvist-Tymchuk, M.R.; Liu, Y.; Fillmore, S.A.E. Human Anthocyanin Bioavailability: Effect of Intake Duration and Dosing. Food Funct. 2017, 8, 4563–4569. [Google Scholar] [CrossRef]
- Cooney, J.M.; Jensen, D.J.; McGhie, T.K. LC-MS Identification of Anthocyanins in Boysenberry Extract and Anthocyanin Metabolites in Human Urine Following Dosing. J. Sci. Food Agric. 2004, 84, 237–245. [Google Scholar] [CrossRef]
- Felgines, C.; Talavera, S.; Texier, O.; Gil-Izquierdo, A.; Lamaison, J.-L.; Remesy, C. Blackberry Anthocyanins Are Mainly Recovered from Urine as Methylated and Glucuronidated Conjugates in Humans. J. Agric. Food Chem. 2005, 53, 7721–7727. [Google Scholar] [CrossRef]
- Kalt, W.; McDonald, J.E.; Liu, Y.; Fillmore, S.A.E. Flavonoid Metabolites in Human Urine during Blueberry Anthocyanin Intake. J. Agric. Food Chem. 2017, 65, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Fornasaro, S.; Ziberna, L.; Gasperotti, M.; Tramer, F.; Vrhovšek, U.; Mattivi, F.; Passamonti, S. Determination of Cyanidin 3-Glucoside in Rat Brain, Liver and Kidneys by UPLC/MS-MS and Its Application to a Short-Term Pharmacokinetic Study. Sci. Rep. 2016, 6, 22815. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Blumberg, J.B.; McDonald, J.E.; Vinqvist-Tymchuk, M.R.; Fillmore, S.A.E.; Graf, B.A.; O’Leary, J.M.; Milbury, P.E. Identification of Anthocyanins in the Liver, Eye, and Brain of Blueberry-Fed Pigs. J. Agric. Food Chem. 2008, 56, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Milbury, P.E.; Kalt, W. Xenobiotic Metabolism and Berry Flavonoid Transport across the Blood-Brain Barrier. J. Agric. Food Chem. 2010, 58, 3950–3956. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Kritchevsky, J.; Hargett, K.; Feller, K.; Klobusnik, R.; Song, B.J.; Cooper, B.; Jouni, Z.; Ferruzzi, M.G.; Janle, E.M. Plasma Bioavailability and Regional Brain Distribution of Polyphenols from Apple/Grape Seed and Bilberry Extracts in a Young Swine Model. Mol. Nutr. Food Res. 2015, 59, 2432–2447. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Ogawa, T.; Koyanagi, A.; Kobayashi, S.; Goda, T.; Kumazawa, S.; Kobayashi, H.; Shimoi, K. Correction to Distribution and Excretion of Bilberry Anthocyanins in Mice. J. Agric. Food Chem. 2009, 57, 9856–9856. [Google Scholar] [CrossRef]
- Zimman, A.; Waterhouse, A.L. Enzymatic Synthesis of [3‘-O-Methyl-3H]Malvidin-3-Glucoside from Petunidin-3-Glucoside. J. Agric. Food Chem. 2002, 50, 2429–2431. [Google Scholar] [CrossRef]
- Wen, X.; Walle, T. Methylated Flavonoids Have Greatly Improved Intestinal Absorption and Metabolic Stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef]
- El Mohsen, M.A.; Marks, J.; Kuhnle, G.; Moore, K.; Debnam, E.; Kaila Srai, S.; Rice-Evans, C.; Spencer, J.P.E. Absorption, Tissue Distribution and Excretion of Pelargonidin and Its Metabolites Following Oral Administration to Rats. Br. J. Nutr. 2006, 95, 51–58. [Google Scholar] [CrossRef]
- Wu, X.; Pittman, H.E., 3rd; Prior, R.L. Fate of Anthocyanins and Antioxidant Capacity in Contents of the Gastrointestinal Tract of Weanling Pigs Following Black Raspberry Consumption. J. Agric. Food Chem. 2006, 54, 583–589. [Google Scholar] [CrossRef]
- Carkeet, C.; Clevidence, B.A.; Novotny, J.A. Anthocyanin Excretion by Humans Increases Linearly with Increasing Strawberry Dose. J. Nutr. 2008, 138, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Bednár, P.; Barták, P.; Lemr, K.; Sevcík, J. Estimation of Partition Coefficients by MEKC Part 2: Anthocyanins. J. Sep. Sci. 2005, 28, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Muccitelli, H.U.; Sánchez-Moreno, C.; Prior, R.L. Anthocyanins Are Absorbed in Glycated Forms in Elderly Women: A Pharmacokinetic Study. Am. J. Clin. Nutr. 2001, 73, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, A.K.; Huang, Y.; Xiao, D.; Park, E.; Edirisinghe, I.; Burton-Freeman, B. Pharmacokinetic Characterization and Bioavailability of Strawberry Anthocyanins Relative to Meal Intake. J. Agric. Food Chem. 2016, 64, 4891–4899. [Google Scholar] [CrossRef] [PubMed]
- Baron, G.; Altomare, A.; Regazzoni, L.; Redaelli, V.; Grandi, S.; Riva, A.; Morazzoni, P.; Mazzolari, A.; Carini, M.; Vistoli, G.; et al. Pharmacokinetic Profile of Bilberry Anthocyanins in Rats and the Role of Glucose Transporters: LC–MS/MS and Computational Studies. J. Pharm. Biomed. Anal. 2017, 144, 112–121. [Google Scholar] [CrossRef]
- Kay, C.D.; Mazza, G.; Holub, B.J.; Wang, J. Anthocyanin Metabolites in Human Urine and Serum. Br. J. Nutr. 2004, 91, 933–942. [Google Scholar] [CrossRef]
- Kay, C.D. Aspects of Anthocyanin Absorption, Metabolism and Pharmacokinetics in Humans. Nutr. Res. Rev. 2006, 19, 137–146. [Google Scholar] [CrossRef]
- Felgines, C.; Talavéra, S.; Texier, O.; Lamaison, J.-L.; Gonthier, M.-P.; Scalbert, A.; Rémésy, C. Strawberry Anthocyanins Are Recovered in Urine as Glucuro- and Sulfoconjugates in Humans. J. Nutr. 2003, 133, 1296–1301. [Google Scholar] [CrossRef]
- Kay, C.D.; Mazza, G. (joe); Holub, B.J. Anthocyanins Exist in the Circulation Primarily as Metabolites in Adult Men. J. Nutr. 2005, 135, 2582–2588. [Google Scholar] [CrossRef]
- Xie, L.; Lee, S.G.; Vance, T.M.; Wang, Y.; Kim, B.; Lee, J.-Y.; Chun, O.K.; Bolling, B.W. Bioavailability of Anthocyanins and Colonic Polyphenol Metabolites Following Consumption of Aronia Berry Extract. Food Chem. 2016, 211, 860–868. [Google Scholar] [CrossRef]
- Li, A.; Xiao, R.; He, S.; An, X.; He, Y.; Wang, C.; Yin, S.; Wang, B.; Shi, X.; He, J. Research Advances of Purple Sweet Potato Anthocyanins: Extraction, Identification, Stability, Bioactivity, Application, and Biotransformation. Molecules 2019, 24, 3816. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, Y. Biopolymer-Based Encapsulation of Anthocyanins as Reinforced Natural Colorants for Food Applications. J. Agric. Food Res. 2023, 11, 100488. [Google Scholar] [CrossRef]
- Yuan, Y.; Fan, Q.; Xu, X.; Wang, O.; Zhao, L.; Zhao, L. Nanocarriers Based on Polysaccharides for Improving the Stability and Bioavailability of Anthocyanins: A Review. Carbohydr. Polym. Technol. Appl. 2023, 100346. [Google Scholar] [CrossRef]
- Mehran, M.; Masoum, S.; Memarzadeh, M. Improvement of Thermal Stability and Antioxidant Activity of Anthocyanins of Echium Amoenum Petal Using Maltodextrin/Modified Starch Combination as Wall Material. Int. J. Biol. Macromol. 2020, 148, 768–776. [Google Scholar] [CrossRef]
- Wang, M.; Li, L.; Wan, M.; Lin, Y.; Tong, Y.; Cui, Y.; Deng, H.; Tan, C.; Kong, Y.; Meng, X. Preparing, Optimising, and Evaluating Chitosan Nanocapsules to Improve the Stability of Anthocyanins from Aronia Melanocarpa. RSC Adv. 2021, 11, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, Z.; Wang, C.; Liu, S.; Wang, Y.; Zhang, M.; Tian, Y.; Lv, J.; Xu, H.; Xia, G. Stability and Antioxidant Activity of Chitosan/β-Lactoglobulin on Anthocyanins from Aronia Melanocarpa. Lebenson. Wiss. Technol. 2023, 173, 114335. [Google Scholar] [CrossRef]
- Fernandes, A.; Rocha, M.A.A.; Santos, L.M.N.B.F.; Brás, J.; Oliveira, J.; Mateus, N.; de Freitas, V. Blackberry Anthocyanins: β-Cyclodextrin Fortification for Thermal and Gastrointestinal Stabilization. Food Chem. 2018, 245, 426–431. [Google Scholar] [CrossRef]
- Vukoja, J.; Buljeta, I.; Pichler, A.; Šimunović, J.; Kopjar, M. Formulation and Stability of Cellulose-Based Delivery Systems of Raspberry Phenolics. Processes (Basel) 2021, 9, 90. [Google Scholar] [CrossRef]
- Kanokpanont, S.; Yamdech, R.; Aramwit, P. Stability Enhancement of Mulberry-Extracted Anthocyanin Using Alginate/Chitosan Microencapsulation for Food Supplement Application. Artif. Cells Nanomed. Biotechnol. 2018, 46, 773–782. [Google Scholar] [CrossRef]
- Ahmad, M.; Ashraf, B.; Gani, A.; Gani, A. Microencapsulation of Saffron Anthocyanins Using β Glucan and β Cyclodextrin: Microcapsule Characterization, Release Behaviour & Antioxidant Potential during in-Vitro Digestion. Int. J. Biol. Macromol. 2018, 109, 435–442. [Google Scholar]
- Flores, F.P.; Singh, R.K.; Kerr, W.L.; Phillips, D.R.; Kong, F. In Vitro Release Properties of Encapsulated Blueberry (Vaccinium Ashei) Extracts. Food Chem. 2015, 168, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Lanna, E.G.; Bittencourt, V.C.E.; Moreira, A.M.S.; Da Silva, J.G.; Sousa, O.V.; Denadai, Â.M.L. Physicochemical Characterization and Biological Activities of the Ethanol Extract of Bryophyllum Pinnatum (Lam.) Oken Incorporated in β-Cyclodextrin. J. Incl. Phenom. Macrocycl. Chem. 2016, 85, 247–259. [Google Scholar] [CrossRef]
- Norkaew, O.; Thitisut, P.; Mahatheeranont, S.; Pawin, B.; Sookwong, P.; Yodpitak, S.; Lungkaphin, A. Effect of Wall Materials on Some Physicochemical Properties and Release Characteristics of Encapsulated Black Rice Anthocyanin Microcapsules. Food Chem. 2019, 294, 493–502. [Google Scholar] [CrossRef]
- Oancea, A.-M.; Hasan, M.; Vasile, A.M.; Barbu, V.; Enachi, E.; Bahrim, G.; Râpeanu, G.; Silvi, S.; Stănciuc, N. Functional Evaluation of Microencapsulated Anthocyanins from Sour Cherries Skins Extract in Whey Proteins Isolate. Lebenson. Wiss. Technol. 2018, 95, 129–134. [Google Scholar] [CrossRef]
- Mueller, D.; Jung, K.; Winter, M.; Rogoll, D.; Melcher, R.; Kulozik, U.; Schwarz, K.; Richling, E. Encapsulation of Anthocyanins from Bilberries – Effects on Bioavailability and Intestinal Accessibility in Humans. Food Chem. 2018, 248, 217–224. [Google Scholar] [CrossRef]
- Santos, S.S. dos; Paraíso, C.M.; Romanini, E.B.; Correa, V.G.; Peralta, R.M.; Costa, S.C. da; Santos Junior, O. de O.; Visentainer, J.V.; Reis, M.H.M.; Madrona, G.S. Bioavailability of Blackberry Pomace Microcapsules by Using Different Techniques: An Approach for Yogurt Application. Innov. Food Sci. Emerg. Technol. 2022, 81, 103111. [Google Scholar] [CrossRef]
- Flores, G.; Ruiz del Castillo, M.L.; Costabile, A.; Klee, A.; Bigetti Guergoletto, K.; Gibson, G.R. In Vitro Fermentation of Anthocyanins Encapsulated with Cyclodextrins: Release, Metabolism and Influence on Gut Microbiota Growth. J. Funct. Foods 2015, 16, 50–57. [Google Scholar] [CrossRef]
- Ge, J.; Yue, X.; Wang, S.; Chi, J.; Liang, J.; Sun, Y.; Gao, X.; Yue, P. Nanocomplexes Composed of Chitosan Derivatives and β-Lactoglobulin as a Carrier for Anthocyanins: Preparation, Stability and Bioavailability in Vitro. Food Res. Int. 2019, 116, 336–345. [Google Scholar] [CrossRef]
- Oidtmann, J.; Schantz, M.; Mäder, K.; Baum, M.; Berg, S.; Betz, M.; Kulozik, U.; Leick, S.; Rehage, H.; Schwarz, K.; et al. Preparation and Comparative Release Characteristics of Three Anthocyanin Encapsulation Systems. J. Agric. Food Chem. 2012, 60, 844–851. [Google Scholar] [CrossRef]
- Betz, M.; Kulozik, U. Whey Protein Gels for the Entrapment of Bioactive Anthocyanins from Bilberry Extract. Int. Dairy J. 2011, 21, 703–710. [Google Scholar] [CrossRef]
- Ji, Y. Synthesis of Porous Starch Microgels for the Encapsulation, Delivery and Stabilization of Anthocyanins. J. Food Eng. 2021, 302, 110552. [Google Scholar] [CrossRef]
- Chatterjee, N.S.; Dara, P.K.; Perumcherry Raman, S.; Vijayan, D.K.; Sadasivam, J.; Mathew, S.; Ravishankar, C.N.; Anandan, R. Nanoencapsulation in Low-molecular-weight Chitosan Improves in Vivo Antioxidant Potential of Black Carrot Anthocyanin. J. Sci. Food Agric. 2021, 101, 5264–5271. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, X.; Tie, S.; Hou, S.; Wang, H.; Song, Y.; Rai, R.; Tan, M. Facile Synthesis of Nano-Nanocarriers from Chitosan and Pectin with Improved Stability and Biocompatibility for Anthocyanins Delivery: An in Vitro and in Vivo Study. Food Hydrocoll. 2020, 109, 106114. [Google Scholar] [CrossRef]
- Wang, W.; Jung, J.; Zhao, Y. Chitosan-Cellulose Nanocrystal Microencapsulation to Improve Encapsulation Efficiency and Stability of Entrapped Fruit Anthocyanins. Carbohydr. Polym. 2017, 157, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.M.; Ghalegi Ghalenoei, M.; Dehnad, D. Influence of Spray Drying on Water Solubility Index, Apparent Density, and Anthocyanin Content of Pomegranate Juice Powder. Powder Technol. 2017, 311, 59–65. [Google Scholar] [CrossRef]
- Akhavan, S.; Assadpour, E.; Katouzian, I.; Jafari, S.M. Lipid Nano Scale Cargos for the Protection and Delivery of Food Bioactive Ingredients and Nutraceuticals. Trends Food Sci. Technol. 2018, 74, 132–146. [Google Scholar] [CrossRef]
- Assadpour, E.; Jafari, S.M. A Systematic Review on Nanoencapsulation of Food Bioactive Ingredients and Nutraceuticals by Various Nanocarriers. Crit. Rev. Food Sci. Nutr. 2019, 59, 3129–3151. [Google Scholar] [CrossRef]
- Demirci, M.; Caglar, M.Y.; Cakir, B.; Gülseren, İ. Encapsulation by Nanoliposomes. In Nanoencapsulation Technologies for the Food and Nutraceutical Industries; Jafari, S.M., Ed.; Elsevier: Academic Press, 2017; pp. 74–113. [Google Scholar]
- Ravanfar, R.; Tamaddon, A.M.; Niakousari, M.; Moein, M.R. Preservation of Anthocyanins in Solid Lipid Nanoparticles: Optimization of a Microemulsion Dilution Method Using the Placket–Burman and Box–Behnken Designs. Food Chem. 2016, 199, 573–580. [Google Scholar] [CrossRef]
- Mulia, K.; Putri, G.A.; Krisanti, E. Encapsulation of Mangosteen Extract in Virgin Coconut Oil Based Nanoemulsions: Preparation and Characterization for Topical Formulation. Mater. Sci. For. 2018, 929, 234–242. [Google Scholar] [CrossRef]
- Rafiee, Z.; Barzegar, M.; Sahari, M.A.; Maherani, B. Nanoliposomal Carriers for Improvement the Bioavailability of High – Valued Phenolic Compounds of Pistachio Green Hull Extract. Food Chem. 2017, 220, 115–122. [Google Scholar] [CrossRef]
Figure 1.
Overview of phenylpropanoid pathway. PAL, phenylalanine ammonia-lyase; C4H, cinnamic acid 4-hydroxylase; 4CL, 4-coumarate:CoA ligase ; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone3-hydroxylase; F3’H, flavonoid 3′-hydroxylase; F3’5’H, flavonoid 3′5′-hydroxylase; DFR, dihydroflavonol 4-reductase; ANS, anthocyanidin synthase; OMT, ortho-methyltransferase; UFGT, UDP flavonoid glycosyltransferase.
Figure 1.
Overview of phenylpropanoid pathway. PAL, phenylalanine ammonia-lyase; C4H, cinnamic acid 4-hydroxylase; 4CL, 4-coumarate:CoA ligase ; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone3-hydroxylase; F3’H, flavonoid 3′-hydroxylase; F3’5’H, flavonoid 3′5′-hydroxylase; DFR, dihydroflavonol 4-reductase; ANS, anthocyanidin synthase; OMT, ortho-methyltransferase; UFGT, UDP flavonoid glycosyltransferase.
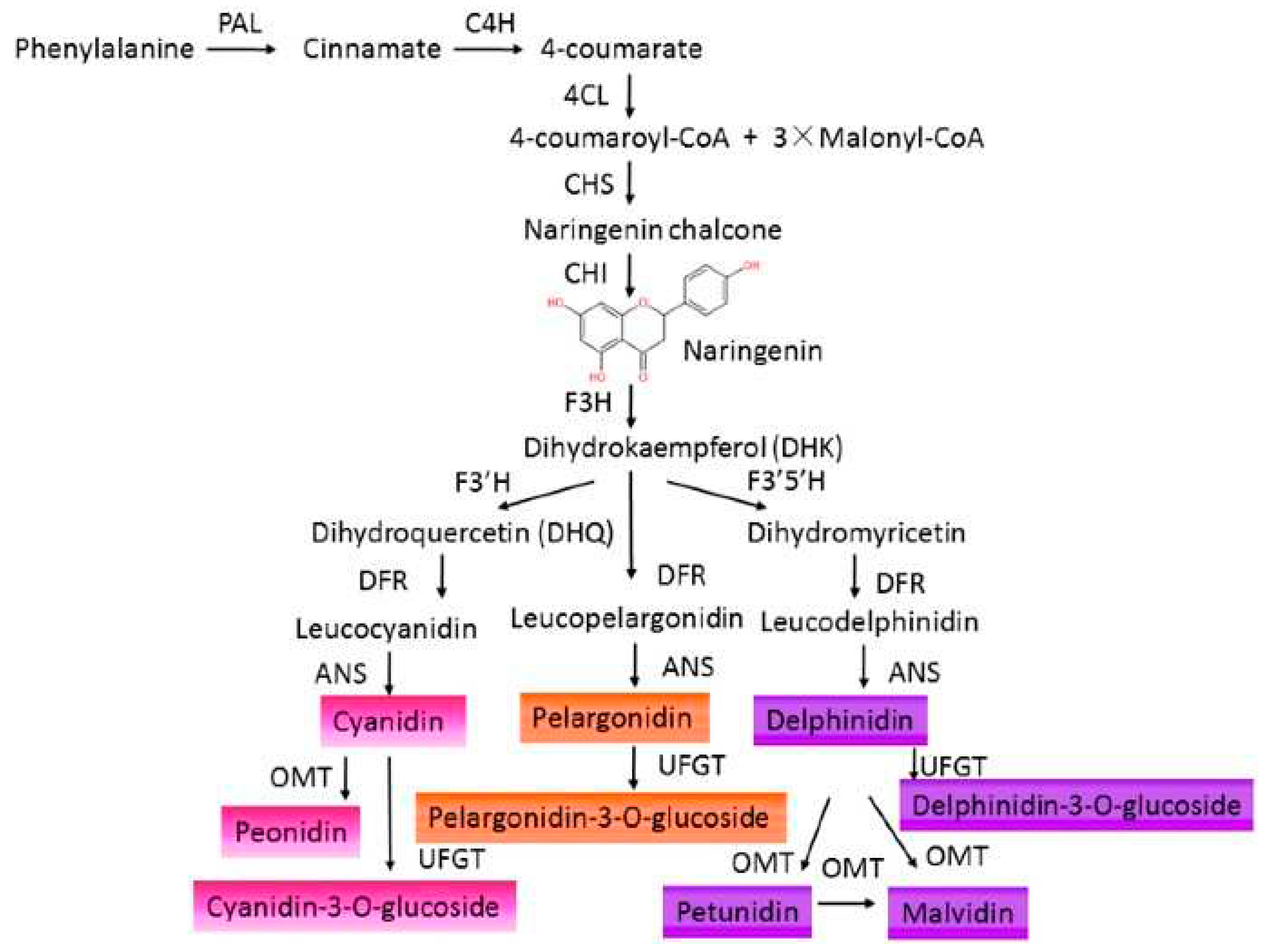
Figure 2.
Chemical skeleton of anthocyanin. (a) flavylium cation, (b) most common anthocyanidins, (c) basic structure of anthocyanin.
Figure 2.
Chemical skeleton of anthocyanin. (a) flavylium cation, (b) most common anthocyanidins, (c) basic structure of anthocyanin.
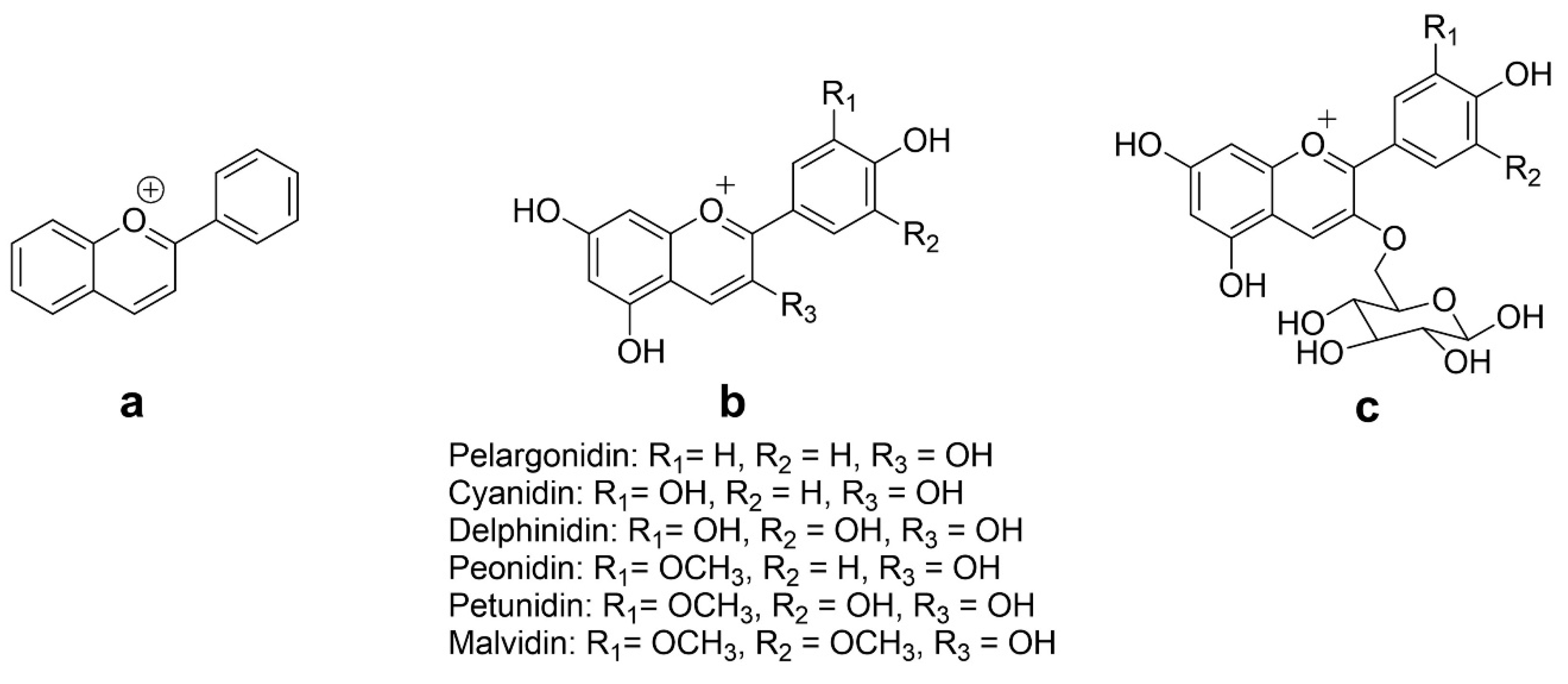
Figure 3.
Nutraceutical potential of anthocyanins based on reported studies.
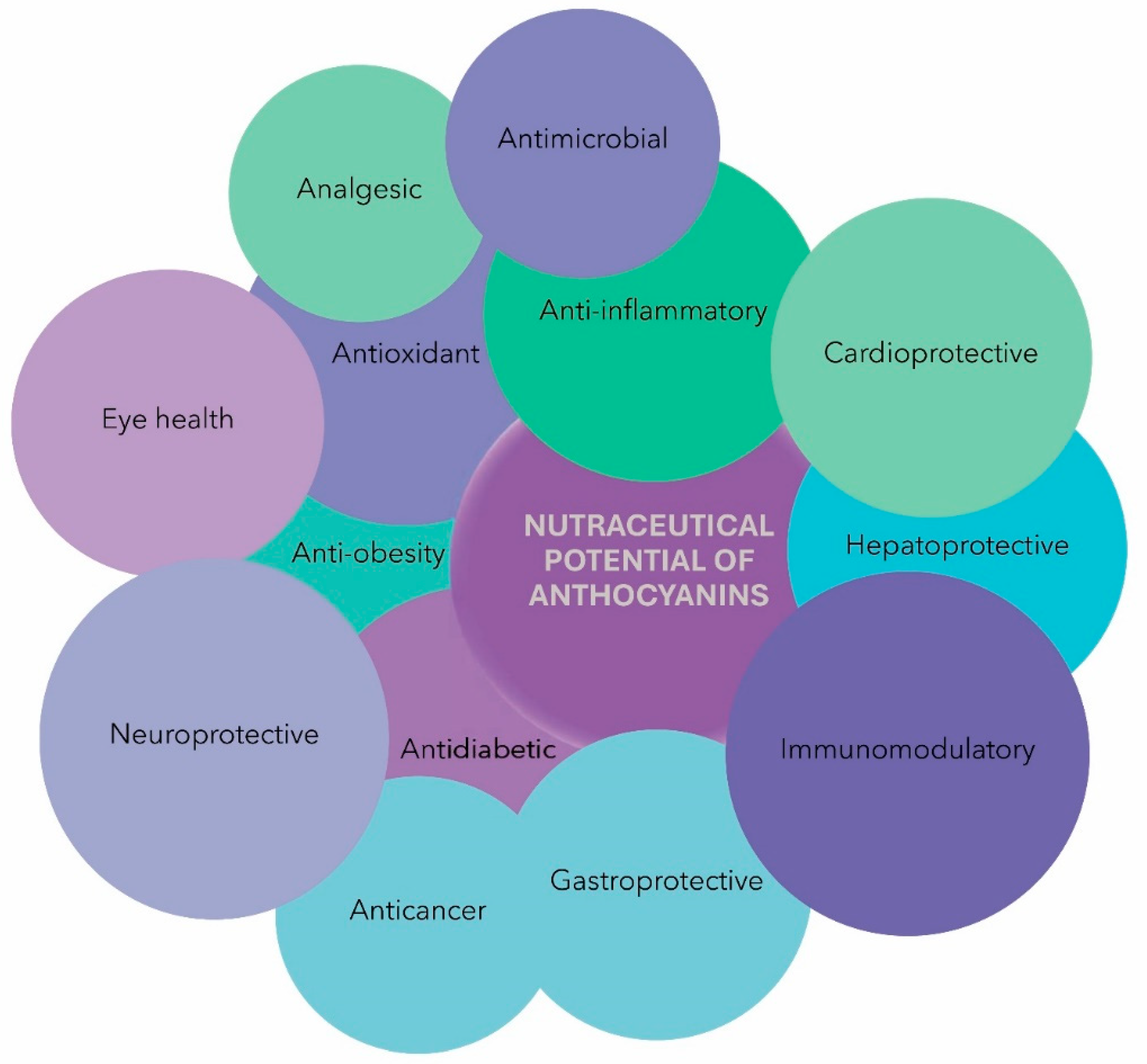
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



