
Submitted:
30 August 2023
Posted:
31 August 2023
You are already at the latest version
Abstract
Recent Socio-economic changes resulted to mass abandonment of agricultural lands in Central-Eastern Europe. This leads to landscapes homogenisation, especially the disappearance of semi-open habitats of the farmland-woodland interface. The information on potentials of transforming feral, abandoned landscapes to wood-pastures is limited. We studied the ecological features and silvopastoral benefits of a landscape subjected to intensive cattle grazing after ~20 years of abandonment, i.e. the mosaic of habitats, cattle’s preference for habitats and for arboreal forages. The nutritional characteristics of arboreal forage and herbaceous forage were compared. After the initial increase in the landscape’s woodland share, cattle grazing halted further woody succession and stabilised the landscape structure, with treeless grassland occupying 49% of the landscape (most preferred by cattle with regard to the time spent), followed by treed grassland (19%), pioneering birch/alder groves (13%), riparian vegetation (12%, least preferred by cattle), and close-canopy woods (6%). The consumption intensity of arboreal forages was on about 4.5 folds higher than that of herbaceous forage. Our study proved that the grazing herd of arobust cattle breed can turn a feral, post-agricultural land into a working wood pasture, consisting of interconnected open grasslands and various facets of woodland, which seldom occur in contemporary high forests. We advocate for the transition management of abandoned farmlands towards integrated silvopastoral landscapes for sustainable provision of multiple ecosystem services that cannot be provided by segregated agriculture and forestry.
Keywords:
Carpathian foothills
; ecosystem services
; grazing ecology
; land abandonment
; wood-pasture
; tree hay
1. Introduction
The European Union (EU) Forest strategy for 2030 [1] acknowledges the contribution of spontaneous forest regrowth through the abandonment of agriculture and rural areas, which corresponds with the rewilding approach that is advocated as the most efficient form of land-use from the biological-conservation perspective [1,2]. However, a growing number of studies reveals the adverse effects of abandonment and succession of woody vegetation on various aspects of biodiversity [3,4,5]. According to Lomba et al. [6], high nature value farmlands (HNVfs), supporting a wide range of ecosystem services, including the conservation of biodiversity, provide a more socio-economically viable option than rewilding. The low-intensity HNV farming systems, depending on socio-economic embedment in local landscapes’ ecologies, are essential for the long-lasting resilience of bio-cultural systems [7,8]. Social and economic decoupling from immediate ecological context, in particular landscapes [9], leads to progressing human alienation [10], generational amnesia [11], irreversible erosion of traditional ecological knowledge [12], and threatens food security [13,14].
Considering that the Potential Natural Vegetation (sensu [15]) of CE is almost entirely dominated by various types of deciduous and mixed forest communities [16,17], the end point of abandoned landscapes’ succession are dense forests. The evidence of this is particularly obvious in the eastern part of the Carpathian region, shared by Poland, Ukraine, and Slovakia, where, as a consequence of World War II (WWII), many communities were displaced and their villages deserted. Seventy years later, scores of former pastures, arable land, fruit orchards, and semi-open grazed woods have turned to high forests – either due to ecological succession or to afforestation [18,19]. Although this may seem beneficial from the viewpoint of the land’s carbon storage capacity, considering the tree lifespan-related inertia of forests adaptability to environmental change, vast tracts of dense woodlands may not turn a desired panacea [20].
Among the species-rich and best-adapted ecosystems are extensively grazed treed grasslands and semi-open woods [21,22,23]. The resilience of these highly diverse communities could be linked to a high-level of plants’ dissimilar responses to ecological stress and disturbances [24,25]. With diversity-friendly management, variegated treed landscapes can exhibit higher resistance to environmental hazards than vast tracks of structurally simplified dense forests and act as supplementary carbon sinks and biodiversity hubs [26,27,28]. The latter particularly refer to the role of semi-open woods and veteran, sun-exposed, trees [29,30].
Instead of farmlands transformation into dense forests, silvopastoralism could be (re-)introduced for simultaneous provisioning of multiple ecosystem services. Along with the provision of food and fibre, grazing may contribute to multiple environmental benefits, such as sustaining species-rich, semi-open habitats, facilitating the survival of shade-intolerant trees, or securing zoochoric plant dispersal across landscapes [21,31,32]. Moreover, silvopastoralism benefits the conservation of old livestock breeds, an important part of biodiversity and cultural legacy [13,33]. Among the most important factors, responsible for the loss of the ecological patterns related to the historical use of woodlands, was almost a universal ban on forest grazing imposed in Central Europe along with the progress of the modern, timber-oriented, forestry. In Poland, such a legal prohibition, enacted in the early 1970s, has radically changed the ecological character of the woodland-farmland interface, leading to ecological isolation of woodlands from their agricultural context [23,34].
In the present study, conducted in the rural neighbourhood of city of Rzeszów, SE Poland, we demonstrate the vegetation structure and the cattle grazing pattern in the wood pasture (Borówki village), which has developed from the former mosaic farming landscape, consisting of arable fields, meadow and pasture as well as small patches of groves. That working landscape had been abandoned in the late 1990s and became subject to progressing succession of pioneering trees, in particular, silver birch (Betula pendula) and black alder (Alnus glutinosa). First herd of Galloway cattle was introduced there in 2008, with no special site preparation, becoming the new key-factor affecting habitats and shaping the landscape mosaic. We investigated the relationship between the vegetation structure of the wood-pasture and the pattern of free-range cattle grazing. Experimental exposure of the harvested tree leafage to the livestock was also done to assess the attractiveness of the arboreal forage, unavailable in conventional CE Europe’s pastures, to cattle. We compared the local leafage to herbaceous forage with respect to selected chemical and nutritional traits, considering such tree use benefits for landscape quality and animal welfare.
We hypothesised that, unlike in landscapes, where grazing-prevented woods have developed at their edges an efficient ecological barrier of a dense wedge of shrubs [23,34], in working wood-pasture, livestock, wandering and crossing habitats’ borders, sustains ecological connectivity between woods and open habitats (H1). We expected that well-developed ecotone zones, combining features of different categories, are of particular appreciation to grazers (H2). We anticipate that experimentally provided arboreal forage will reveal, depending on tree species, various levels of forage attractiveness to cattle (H3). Finally, considering the past development of the studied landscape and its alternative future transformations, we reflected on the changing structure of ecosystem services resulting from particular scenarios.
We suggest that the present study could be used as a reference for transformation of abandoned and neglected farmlands into working silvopastoral landscapes as an ecologically and economically viable alternative to feralization of cultural rural landscapes.
2. Materials and Methods
2.1. Study area
The study area is located in the Carpathian foothills, ca. 10 km south-east from the centre of Rzeszów (200,000 inhabitants), south-east Poland (Figure 1A). When major socio-economic changes of the 1990s and early 2000s (opening Poland to global economy and accession to EU) led thousands of multiple-use (semi-)subsistence family farms to decline, scores of ploughfields, meadows, and pastures, including those in the study area, were abandoned [11].
In early 2000s, the local agriculture and craft association acquired ca. 30 ha farmland consisting of abandoned ploughfields, patches of woods as well as grassland with scattered wild cherry trees (Cerasus avium), common oaks (Quercus robur), sycamore maples (Acer pseudoplatanus), and black alders (Alnus glutinosa) in the hilly area of Dynowskie foothills. By then, parts of farrow fields were already covered by pioneering young birch (Betula pendula) and alder thickets, accompanied by young oaks, sycamore maples, and hornbeams (Carpinus betulus). The study was focused on the half of that area (16.3 ha) fenced, and subjected to grazing by the Galloway cattle, first introduced in 2008 (Figure 1). The herd, substantially exceeding the pasture grazing capacity (80 adult individuals and ca. 40 calves) receives supplementary fodder – locally acquired hay and haylage. Being a part of a circular farming system, animals are also fed with vegetable left-overs from the local fruit-vegetable operation. The herd spends most of the daytime of the grazing season (late April to early November) in the wood-pasture, gathering under large shelter after the dark or during major weather breakdowns.
2.2. Vegetation analysis of the cattle-grazed wood-pasture
We used Yuneec H520 with E90 camera (23 mm F/2.8) to acquire the high-resolution ortophotomap (20 MP, ca. 1.2 cm/px), acquired in the optimal autumn trees phenological stage (late October, 2020). The ortophotomap and the terrain model was used for the delineation of the apparent vegetation categories: open, treeless grassland (OGR), pioneering groves dominated by birch or alder (PIG), treed grassland (SOW), close-canopy woods (CCW), and riparian woods (RIW). In early July 2021, in 32 arbitrarily chosen locations, representing typical aspects of each vegetation category, 10 × 10 m phytosociological relevés were made. Using six-interval relevés’ coverage (“+” < 1%, “1” 1–5%, “2” 6–25%, “3” 26–50%, “4” 51–75%, “5” > 75%), abundances of vascular plant species were assessed [35] as a basis for the zones’ phytosociological classification. For the nomenclature of phytosociological syntaxonomic categories and for the species’ phytosociological affiliation we referred to [16] (supplementary material 1, SM1). In detailed phytosociological analyses, neither tree nor shrub seedlings were considered, due to their small diagnostic value. In addition, the ortophotomap was used to digitise the stems of the stand’s canopy layer (Figure 1 and Figure 2). With the “Kernel Density Estimation” QGIS plug-in (radius 25 m, ca. 2000 m2), the point values of stand density were estimated. Based on the resulting “heat map”, we generated isolines (QGIS, contour cutting = 5; Figure 1) and tree stem density zones, corresponding with five distinguished vegetation categories (Figure 2).
In order to assess the possible effect of livestock grazing on plant movement across the landscape mosaic, we categorized the herbaceous species affiliations to phytosociological classes as either grassland species, or forest species, or ruderal species, or species belonging to mantle and fringe shrubland vegetation. As a proxy measure of the plant species migration (facilitated by cattle) between various types of habitat, we compared the ratios of species categories with the 2-sample test for equality of proportions with continuity correction.
2.3. Spatial pattern and dynamics of the wood-pasture use by cattle
2.3.1. Cattle itineraries on the wood-pasture
Involving four, randomly selected adult herd’s cows, the round-the-clock movement pattern of animals through the mosaic of five habitat categories was explored. They were tagged with COBAN TK108B 10000 mAh track-loggers placed in canvas bags, fixed to the neck collars on June 24th 2020 and the cows’ position data logging, with 10-s interval, started on midnight 25/26 June and lasted till midnight 28/29 June 2020 of four cows. Based on the spatial density of the impulses (i.e., cows’ geo-impulses per 100 m2) recorded by the track-loggers, we estimated the relative exploitation intensities of particular habitat types as well as inter-habitats’ 10-m-wide transition zones (ecotones). The ecotones were determined in QGIS as the borderlines’ each side 5-m-wide buffers. Analysing the movements between neighbouring habitats, we assessed the spatial-dynamic pattern of cattle’s use of the variegated landscape.
2.3.2. Temporary cattle groupings and livestock temporal impact on the wood-pasture
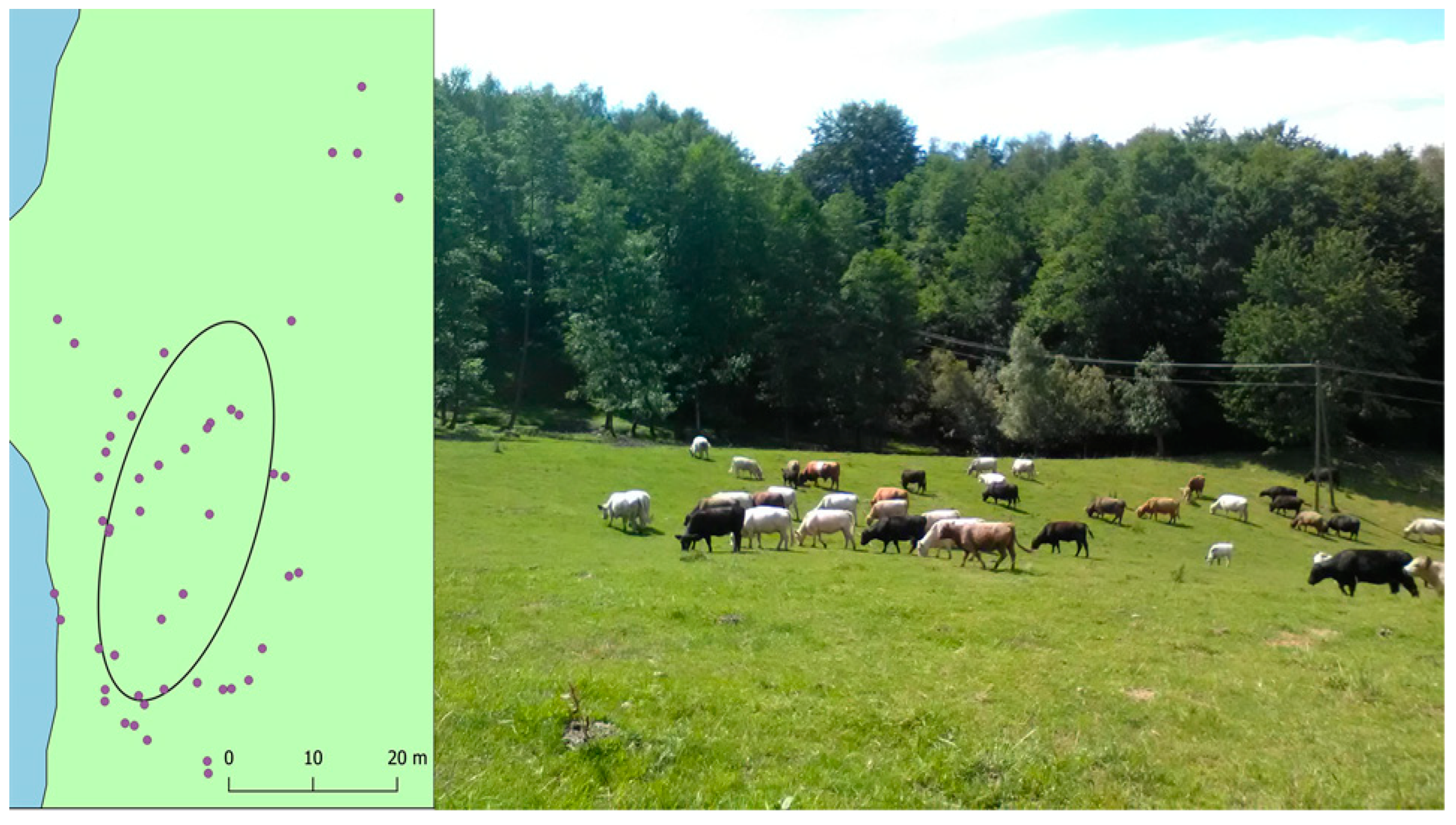
From July 7th to 9th, the herd was discreetly observed from 7 AM to 5 PM, and the temporary cattle groupings (TCGs, including bulls, cows, and heifers, excluding calves), occupying distinct wood-pasture areas (Figure 3: right) were mapped with the use of Field-Map (IFER) system coupled with Trupulse 360B (Laser Technology) range-finder and their durations were assessed. On the basis of the multiple-point groupings, “standard deviational ellipses” [35] were calculated with the QGIS plugin – a synthetic spatial representation of TCGs (Figure 3). Thereafter, we calculated the livestock temporal impact on herbaceous forages (LTI) in the wood-pasture as:
LTI = ni × ti × ASDE(i)−1, where ni – number of animals in the i-th TCG, ti – the i-th TCG duration, ASDE(i) – the area of the i-th standard deviational (SD) ellipse.
Kruskal-Wallis test was used to assess the differences in the size of TCGs (i.e., the number of animals in TCGs and the SD ellipses’ areas) between the three observation days, while cattle preferences for the five habitats were assessed by tests of single proportion and multiple proportions [36].
Figure 3.
Exemplary representation of a particular cattle grouping (on July 9, around 3 PM) with the standard deviational ellipse.
Figure 3.
Exemplary representation of a particular cattle grouping (on July 9, around 3 PM) with the standard deviational ellipse.
2.3.4. Cattle feeding pattern on arboreal forages
To understand how cattle feed on fresh arboreal forages, we selected fourteen trees as representative sources in the wood-pasture: birch (2 trees), alder (1), oak (3), wild cherry (3), sycamore maple (3), and hornbeam (2). Nine of them were pollarded on July 2nd and five on July 7th. While the branches of each of the first nine trees were cut-off and placed at 3-5 m away from each tree as multiple-species stacks (Figure 4), the whole crowns of the latter five trees were felled, and left close to each tree without cutting the branches (Figure 4). The stacks/crowns were represented by their perimeter polygons extended to the 2-m-wide buffers, an average length of a cow. To collect data on attractiveness of the arboreal forages to cattle, immediately after having established the forage posts, Acorn Ecotone phototraps were installed on neighbouring trees to film the attracted animals. The phototraps were uninstalled after five days of the exposure, when the accessible branches had been defoliated and the spots were no longer visited by cattle (Figure 4). The phototraps collected 348 digital photographs and 607 short movie clips. We extracted the date and time of the animals’ appearance, maximal number of animals consuming the arboreal forages, and the stack/crown abandonment time from the graphic data. Thereafter, the stack/crown-specific timelines of livestock temporal impact on arboreal forages (LTIa) were calculated as:
LTIa = ni × ti × Ai−1, where ni – number of animals at the i-th stack/pollarded crown, ti – the i-th grouping duration, Ai – the area of the 2-m-wide buffer of a stack/pollarded crown projection on the ground.
Figure 4.
Cattle browsing of the pollarded crown. Left: freshly pollarded young oak (July 7, after 12 PM); Right: the same pollarded crown almost 24 hours later, with six cows/heifers; the dashed line refers to the 2-m-wide buffer of the crown’s projections used in the LTIa calculation.
Figure 4.
Cattle browsing of the pollarded crown. Left: freshly pollarded young oak (July 7, after 12 PM); Right: the same pollarded crown almost 24 hours later, with six cows/heifers; the dashed line refers to the 2-m-wide buffer of the crown’s projections used in the LTIa calculation.

2.4. Nutritional characteristics of the wood-pasture fodder
To compare the nutritional potentials of tree leafage to that of the grassland herbaceous forage, we analysed two collective samples (of above-ground parts, including leaves, flowers, stems, of herbaceous plants sickled at 1 cm aboveground) harvested on July 1st in two fenced-off parts of the pasture’s open ryegrass-crested dogtail (Lolio-Cynosuretum) grassland of six tree species, i.e., common oak, sycamore maple, hornbeam, silver birch, wild cherry, and alder, divided into leaves and twigs sub-samples (Figure 2; Fagus sylvatica trees were not analysed because they are relatively old and tall). Crude protein, crude fat, crude fibre, ash, and nitrogen-free extracts (NFE) were determined by AOAC methods [37]. Following a documented procedure [38], we determined the content of the following macro- and microelements: Ca, K, Mg, P, S, Mn, Zn, Fe, and Sr (see supplementary material, SM2, for details).
3. Results
3.1. Vegetation structure of the wood-pasture
The wood-pasture was dominated by open grassland (OGR; 49% of the area; Figure 2), mainly constituted by perennial ryegrass (Lolium perenne, on average of 14% cover contribution), bentgrass (Agrostis capillaris, 11%), white clover (Trifolium repens, 11%), blue grass (Poa pratensis, 6%), red fescue (Festuca rubra, 5%), dandelion (Taraxacum officinale, 5%), creeping buttercup (Ranunculus repens, 4%), moneywort (Lysimachia nummularia, 4%), heal-all (Prunella vulgaris, 3%) among other species (SM1). As many as 41 (56%) of species in OGR had a phytosociological affiliation in grassland classes in the wood-pasture (see SM1). The herbaceous layer of semi-open treed grassland (SOW), covering 19% of pasture, had a very similar structure to OGR. Its major physiognomic difference was the presence of scattered trees as well as spiny and thorny shrubs, in particular, blackberries (Rubus sp.), dog rose (Rosa canina), and hawthorn (Crataegus sp.). Although the overall abundance of plants was three-fold lower to open grassland, species groups’ structure and actual species composition were similar to those of OGR. Bentgrass (11%), rough blue grass (Poa trivialis, 10%), perennial ryegrass (9%), moneywort (9%), dandelion (6%), blue grass (6%), creeping buttercup (5%) dominated the SOW, and 33 out of all 44 SOW species had a phytosociological affiliation in grassland classes, as in OGR (Figure 4, SM1). Equally high participation of grasslands-affiliated species (72%) occurred in pioneering groves (PIG), covering 13% of the wood-pasture. The most abundant species in PIG was bentgrass (22%) and among the next nine most abundant taxons, seven were mentioned above as OGR and SOW co-dominating species, accompanied here by yarrow (Achillea millefolium, 6%) and ribwort plantain (Plantago lanceolata, 3%). The riparian woods vegetation (RIW), occupying 12% of the wood-pasture, revealed substantial differences to the former categories. Although the most abundant species in RIW was rough blue grass, which contributed 28% of the overall species abundance, the next most abundant species were either nitrophilous or characteristic for wetland vegetation. Some of these species include mannagrass (Glyceria fluitans, 8%), wood clubrush (Scirpus sylvaticus, 7%), creeping buttercup (6%), ground elder (Aegopodium podagraria, 6%), wood avens (Geum urbanum, 5%), and herb-Robert (Geranium robertianum, 5%). The smallest share of the wood-pasture (6%) was occupied by the close canopy woods (CCW), which had the poorest herbaceous layer (i.e., 28 species compared to 73, 44, 43, 34 in, OGR, SOW, PIG, RIW, respectively). CCW was not only dominated by forest species sanicle (Sanicula europaea, 18%), wood anemone (Anemone nemorosa, 14%), broad-leaved enchanter’s nightshade (Circaea lutetiana, 7%), and wild strawberry (Fragaria vesca, 6%), but also ruderal taxa [i.e., wood avens (7%), herb-Robert (6%)], and grassland species [i.e., common self-heal (Prunella vulgaris, 6%) and moneywort (5%)] (Figure 2 and Figure 4; SM1).
Figure 5.
Vegetational composition of five habitat categories in the wood-pasture; Woodland, Grassland, Ruderal, Shrubland species belonging to forest (coniferous and deciduous), grassland, ruderal, and shrubland phytosociological classes, respectively (for details see SM1); the vegetation habitat structure differs significantly between the habitats even when the most outlier CCW was not considered (Pearson’s Chi-squared test, χ2 = 37.703, df = 9, p-value < 0.0001); for habitat categories see Figure 2.
Figure 5.
Vegetational composition of five habitat categories in the wood-pasture; Woodland, Grassland, Ruderal, Shrubland species belonging to forest (coniferous and deciduous), grassland, ruderal, and shrubland phytosociological classes, respectively (for details see SM1); the vegetation habitat structure differs significantly between the habitats even when the most outlier CCW was not considered (Pearson’s Chi-squared test, χ2 = 37.703, df = 9, p-value < 0.0001); for habitat categories see Figure 2.
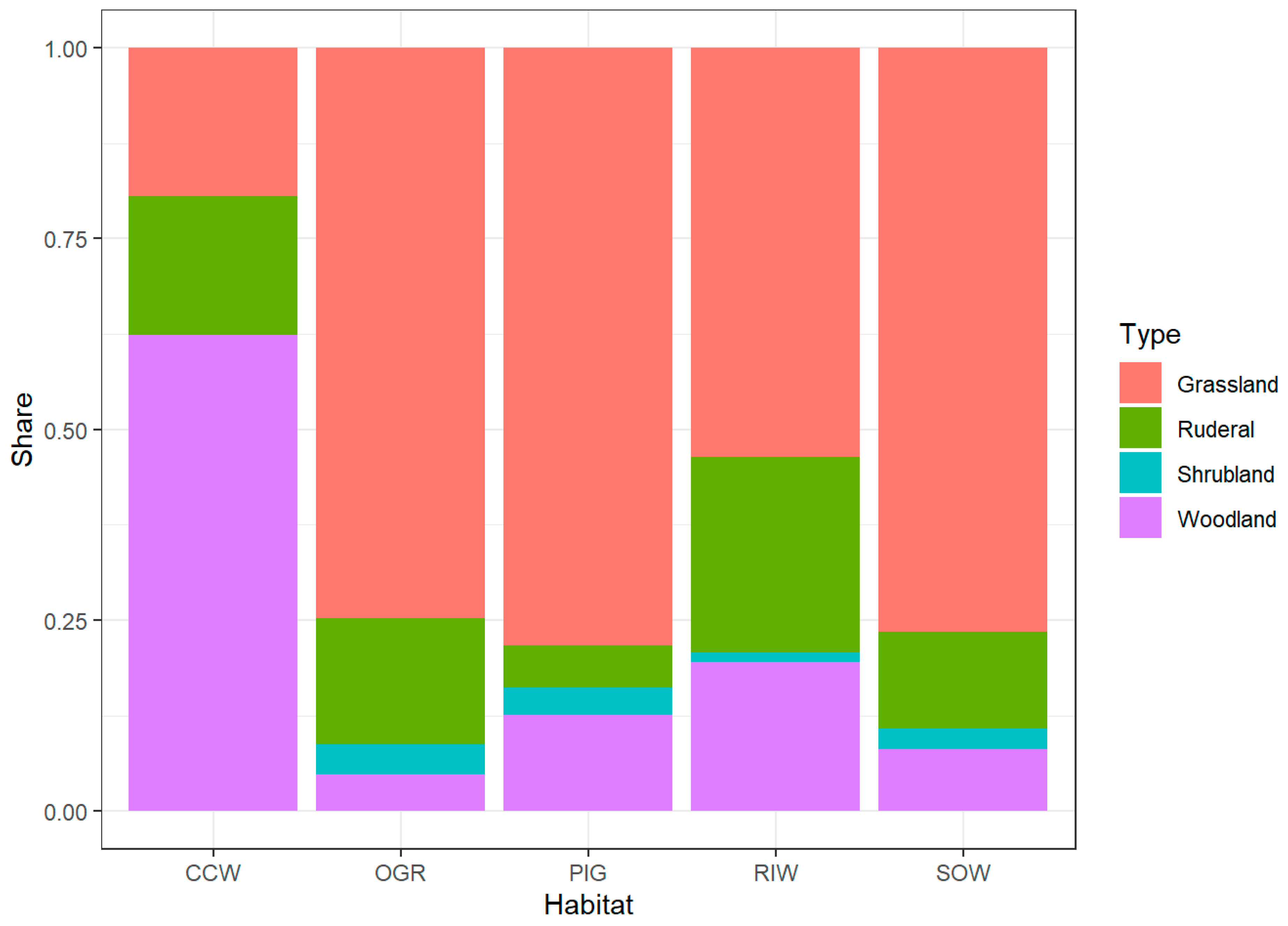
The multiple proportion test revealed significant differences of proportions of woodland to non-woodland herbaceous species among all five habitat categories (χ2 = 154.97, df = 4, p-value < 0.0001) as well as among particular pairs of habitats. In the latter case, the CCW was the most distinct category, significantly different from any other habitat category. The most conspicuous difference occurred between CCW and OGR (2-sample test for equality of proportions with continuity correction χ2 = 128.9, df = 1, p-value < 0.0001). Considering the species affiliated in woodland plant communities as “native” to CCW (while the non-woodland ones as “alien”) and the species affiliated in non-woodland communities as “native” to OGR (while the woodland ones as “alien”), the alien-to-native ratio was 0.60 in CCW and only 0.05 in OGR (Table 1; SM1).
3.2. Spatiotemporal use of the wood-pasture by cattle
3.2.1. Habitats’ preferences of the tracked cows and temporary cattle groupings
In order to investigate preferences of the tracked cows and cattle groupings, we have compared the time spent in each habitat with a surface share of each habitat. If there are no preferences then the cows should graze along random routes over pasture. In such case the distribution of time spent should be similar to the distribution of the surface of habitats under study. Since the share of surface is not random, we used single proportion test. The four tagged-cows’ visits in RIW and CCW were less than their shares (RIW: p=0.057; CCW: p=0.009) in the wood-pasture, while no significant differences were observed for OGR (p=0.694), Pion PIG (p=0.106) and SOW (p=0.133) (Figure 6). The order of the cows’ visits was consistent with the order of the share of the five habitats (OGR> SOW> PIG> RIW> CCW) in the wood-pasture (Figure 6).
As many as 44 temporary cattle groupings (TCGs), totalling 1119 animals, were mapped on 12 on July 7, 16 on July 8, and 16 on July 9 (Figure 7). Particular TCGs consisted of 8 to 51 animals, extending from 280 m2 to ~13800 m2, and lasting from 5 to 130 minutes. Overall, there were no significant differences between the daily number of animals in the TCGs and the daily areas of their SD-ellipses (Figure 7). The relative frequency of the use of CCW, RIW and SOW habitats by TCGs were significantly lower than their shares (CCW: p=0.054, RIW: p=0.017 and SOW: p=0.062), while there were no significant differences for OGR (p=0.670) and PIG (p=0.197) (Figure 6). However, the order of habitats’ uses by the TCGs differed (OGR> PIG> SOW> CCW> RIW) from that of the habitats’ spatial shares (OGR> SOW> PIG> RIW> CCW).
3.2.2. Livestock temporal impact on herbaceous and arboreal forages
The total livestock temporal impact on herbaceous forages (LTI) ranged from close to 0 to 4.8 individual-hours per 100 m-2, and there was no significant difference between the daily LTI values (Figure 8). The dynamics of foraging (LTIa) on the branches’ stacks and the whole crowns were similar (Arb1 vs. Arb2; Figure 8). Most of the cattle’s foraging-related presence around the two forms of arboreal forages occurred during the daytime, with two distinguishable peaks – one between 5 and 8 AM and the other one between 10 AM and 2 PM (SM3). Generally, either the first or the second visit had the highest LTIa value, which decreases with subsequent visits due to the fodder depletion (SM3). Overall, the LTIa was substantially higher than the LTI recorded during the three days of observation (Figure 8).
3.3. Comparison of nutritional composition of herbaceous and arboreal forages
Tree leaves were significantly richer in proteins, fat, and nitrogen-free extracts (NFE) than the herbaceous forages (Table 2; fourth column). Tree twigs had a higher content of fibre than both tree leaves and herbaceous forages (Table 2; fifth column). The herbaceous forages, however, had a substantially higher content of ash than the arboreal forages. Except for ash, the highest medians of the contents of nutritional components were found in the leaves of alder (proteins, fat) and hornbeam (NFE), and in hornbeam twigs (fibre; Table 2).
Both the tree leaves and twigs had higher calcium and strontium contents than the herbaceous forages (Table 3). The differences between the contents of other elements in the tree leaves versus herbaceous forages were either negligible or insignificant, except for smaller content of iron in tree leaves. Overall, twigs had smaller content of the elements than the herbaceous or tree leaves tissues. Of all the ten elements, only P and Fe were higher in the herbal tissue than in the leaf tissue of trees (Table 3).
4. Discussion
The Subcarpathian region, SE Poland, is the second most forested region of the country (forest cover contributes to 40% of the region’s land base). One fourth of its contemporary woodlands emerged after WWII, due to the massive depopulation of the countryside and land abandonment [18]). Although there are relatively large fragments of natural old-growth forests in Eastern Carpathians (e.g., [40,41]), numerous pockets of old trees in the foothill zone have developed from the semi-open grove enclaves or wider woodland-grassland ecotones in traditional agricultural landscapes [23,34]. Their abandonment, in particular, cessation of livestock grazing of wooded margins, has led to a radical change in the ecological characteristics of the woodland-farmland interface, transformed from the semi-open ecotone to the narrow, sharp ecological barriers, sealing off woods from neighbouring open habitats [34].
Our study demonstrates various aspects of livestock grazing in a treed landscape mosaic, which has almost entirely vanished in the Central Europe’s countrysides due to the intensification of animal production and the legal ban on forest grazing. We assessed the vegetation structure and silvopastoral potential of a wood-pasture, which developed during twelve years of free-range grazing of a feral post-agricultural landscape that was abandoned twenty years earlier. Most of such post-agricultural lands in the proximity of Rzeszów either continue succession towards woodland or, being commodified, undergo peri-urban, mostly residential, development [11].
The vegetation inventory proved that twelve years of cattle grazing has turned an apparently amorphic abandoned farmland (“a shrubby mess” – D. Jaworski, personal communication) into a well-established mosaic of diverse patches of five distinguished types of vegetation: treeless grassland, pioneering groves, riparian woods, treed grassland, and close-canopy woods. While the birch- and aspen-dominated groves emerged on the plough field farrows, treed grassland, with a higher tree species and age diversity, imply a different tree recruitment mechanism. This likely involved a gradual and slow process of grassland colonisation by heavy-seeded, zoochorically dispersed tree species, such as oak, hornbeam, and wild cherry (cf. [42]). Despite the limited acreage of the close-canopy wood habitat, the presence of ancient woodland, poorly dispersible herbaceous species (e.g., Anemone nemorosa and Oxalis acetosella in the herbaceous layer [43,44]) indicate that visiting cattle was not detrimental to the woodland aspect of those habitats. Quite the opposite, their relatively poor ground layer, despite the tree canopy’s heavy shade, is being substantially enriched by shade-intolerant grassland and ruderal species, most likely spread by cattle. As the present close canopy woods’ position corresponds with the forest area depicted on the Austro-Hungarian military map of the mid-1800s [45], it may imply that the present cattle visits in the woods resembles what was common in the foothills’ silvopastoral landscape of 150 years ago (cf. [23]). Although the share of wooded communities (in particular, dominated by pioneering species of birch, aspen, and alder) substantially increased in the local landscape during the first two decades following the collapse of farming, the re-introduction of grazing in 2008 prevented further succession of the woods and stabilised the landscape’s mosaic structure. That proves intensive cattle grazing to be an efficient inhibitor of the succession of the woody vegetation (H1).
In general, the structure of the cattle’s use of habitats did not differ from the expected pattern derived from the overall landscape’s mosaic composition. Among the few exceptions, pioneering birch woods were more often (than expected) visited by larger groups of cattle, while treed grassland and riparian woods were less visited. The attractiveness of the pioneering stands might be related to their ambient half-shadow microclimate, providing shelter from summer heat, and, simultaneously, securing better growing conditions for the herb layer than close-canopy woods. The riparian habitat, due to steep scarps and soft wet floodplain bottom, was probably a water source rather than flocks’ grazing area. Relatively low use of treed grassland might be explained by its transitory characteristics, between woodland and grassland, benefiting the visits of single, dispersed cows rather than of larger flocks. In general, the revealed cattle’s preferences for the habitat use supported our hypothesis (H2), anticipating a higher livestock’s impact on open grassland and scarcely treed habitats than on the close-canopy woods.
Although the study did not specifically address the effect of browsing on tree regeneration, in the south-eastern part of the wood-pasture, we observed a “brush” of the middle-aged hornbeam saplings, evenly “coppiced” by cattle, just in the way the wild ungulates browse this species in the Białowieża Forest [46,47]. In general, except for the locally occurring sprouts and saplings, and a small grove of the young alder-birch thicket in the south eastern part of the wood-pasture, the livestock had almost no unaided access to the arboreal forage. Our experimental feeding revealed a conspicuous appeal of such a diet to Galloway cattle. However, except for the cattle’s preferences for sycamore maple foliage than the neighbouring black alder, we did not detect any selection preferences amongst the assessed trees (see Figure S1). Overall, the intensity of consumption of the experimentally provided arboreal forages by the cattle was 4.5 folds higher than that of the herbaceous forages. Thus, our experiment revealed high overall attractiveness of the arboreal fodder supplement to the regular herbaceous diet of cattle. We are, however, aware a new study would be needed for ranking the cattle’s preference for the arboreal forage of different tree species.
There is plenty evidence indicating beneficial effects of the arboreal forages on both livestock health and environment. However, despite the well documented history of silvopastoralism [48], the arboreal forage use in Europe has been marginalised due to the intensification of animal production and its strict separation from wooded areas [23,49]. Perhaps the most important immediate effect of solitary trees and scattered canopy of treed grassland is the shade shelter, benefiting both herbaceous vegetation and animals, contributing to their better physiological performance [50,51]. During prolonged droughts, the role of arboreal forages may shift from diet supplement to a primary fresh forage [52,53].
It is important to remember that the arboreal forage of silvopastoral woods may compensate for the herbaceous forage scarcity resulting either from the natural soil’s poverty, its degradation, desiccation, or overgrazing. Trees, with their roots penetrating the soil far deeper than accompanying herbaceous plants, play the role of efficient “pumps” of elements and water to the green biomass benefiting both wildlife and livestock. Our analyses proved the tree leaves are richer in Ca, Sr, Mn, protein, fat, and NFE (i.e., primarily sugars) than the herbaceous forage. Although the tree twigs were poorer (compared to herbaceous vegetation) in most of the analysed elements (except for Sr and Zn), they contained higher quantities of NFE and fibre. Our findings partially corroborate with the data reported from tropical and subtropical regions, emphasising the tree foliage to be a rich source of Ca, protein, and fat [54,55,56]. In addition, the arboreal forages are important sources of compounds responsible for the ruminant’s health and self-medication. For instance, the fibre abundant in twigs is necessary for the performance of their digestive-intestinal system [57] and condensed tannins (abundant in both Alnus glutinosa and Quercus robur tissues [58]) protect animals against parasites [59,60], reduce nitrogen volatility [61] and methane emission [61 and therein referred authors]. The microelement strontium, relatively abundant in the analysed plant tissues (especially in hornbeam leaves), promotes the uptake of calcium, which strengthens animals’ bones and minimises the risk of osteoporosis [63,64].
The beneficial effect of oak leafage on the cattle’s health has been confirmed by the Borówki’s cattle keeper. According to his account, after the long stay of Galloway in the shed during the cold and wet 2023 spring, the cattle suffered from severe diarrhoea, of which the pharmaceutical treatment would be very expensive. Thus, as a first-aid measure, the leafy branches shredded from the local oaks and left under the trees was provided to the livestock. As we were informed, the self-medication proved to be 100% efficient, bringing all ill cattle to swift recovery (S. Pękala – personal communication).
To recapitulate, despite the very limited scale of this study, it provides an important insight into the potential of cattle grazing as a successful driver of transformation from early successional post-agricultural, overgrown abandoned land to working silvopastoral landscape. In our view, letting Galloway cattle to graze and shape the piece of feral landscape was a crucial decision, determining the spectrum of immediate or potentially available ecosystem services (Figure 9). Allowing further spontaneous successional development, encroachment and growth of woody vegetation, would homogenise the landscape structure, losing openness and the associated traits and functions. In temperate Europe, dense forest is not a scarce habitat, as it is shown by maps of the Potential Natural Vegetation (sensu [15]) that the dominating Europe’s PNVs are forests (e.g., [17]). As such, secondary succession, following agricultural abandonment, would inevitably lead to development of more forest communities, narrowing the spectrum of ecosystem services to the ones specifically dependent on woodlands (Figure 9). Although such a process would fit the strongly lobbied postulate of land sparing to natural processes and afforestation [1,2], we argue that it would not be a universally beneficial solution with respect to the optimal provision of the ecosystem values. We believe that a more diverse and dynamic landscape, such as working wood-pasture, would be a far better response to environmental and human challenges of the future (Figure 9).
5. Conclusions
This study proved that a robust cattle grazing can transform partly-wooded post-agricultural land into a dynamic mosaic landscape, consisting of interconnected open grasslands and various facets of woodland, which hardly occur in contemporary high forests. Due to their different dynamics, spatial patterns, and adaptation to environmental conditions, silvopastoral habitats should be appreciated as invaluable enrichment of simplified and homogenised vegetation, such as modern croplands, treeless grasslands, or dense forests. Increasing tracts of abandoned and growing feral farmlands could be considered as the future integrated silvopastoral landscapes.
We argue against the use of abandoned farmlands as a mere land base for any alternative projects, such as residential development, wind or photovoltaic farms, schematic tree planting, or leaving for natural succession. Abandoned farmlands, in particular, in suboptimal growing conditions of cultural landscapes shaped by traditional, small-holder farming – such as in the Carpathian foothills – inherit the rich legacy of landscape structures and its particular components (including open grown trees, semi-open woods, groves, and copses). Such habitats, patterns, and related processes can best be preserved if incorporated into a multipurpose, land-use model that benefits from that inherited diversity.
Although we do not provide a universal solution for all particular cases, our results present strong premises indicating that transformation of abandoned farmlands into working wood-pastures would be a feasible response to multiple environmental, conservation, and socio-economic challenges, and to the increasing demand for various ecosystem services. The development of such working wooded landscapes would require a knowledge sharing and cooperation platform between farmers, foresters, and field ecologists. Such a cooperation is necessary to secure a trade-off between grazing intensity and the fodder (both herbaceous and arboreal) regenerative capacity. Besides the wood-pastures’ “target” animal and wood production, their cooperative management should provide a wide range of regulatory and social services, including carbon sequestration, high biodiversity, pollination, seed dispersal, weed control, and scenic beauty.
Supplementary Materials
Any source data will be provided at request.
Author Contributions
Conceptualization, A.B., Ł.K.; methodology, A.B, Ł.K., P.W., R.T.-S., G.Z., and S.W.; software, S.W.; validation, A.B, Ł.K., P.W., R.T.-S., G.Z., and S.W.; formal analysis, A.B, Ł.K., P.W., R.T.-S., G.Z., T.O.M., and S.W.; software, S.W.; investigation, A.B, A.D., Ł.K., P.W., R.T.-S., and G.Z.; resources, A.B., R.T.-S., and G.Z.; data curation, A.B., P.W., and S.W.; writing—original draft preparation, A.B., T.O.M.; writing—review and editing, A.B., T.O.M.; visualization, A.B., S.W.; supervision, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the College of Natural Sciences, University of Rzeszów.
Data Availability Statement
Any source data will be provided at request.
Acknowledgments
The authors express their appreciation to Chmielnik-Zdrój S.A. and its staff for opening the wood-pasture for this research, their friendly support and invaluable pieces of advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- EC (European Commission). A European Green Deal: Striving to be the first climate-neutral continent. 2021. https://ec.europa.eu/info/strategy/priorities-2019-2024/european-green-deal/delivering-european-green-deal_en.
- Navarro, L.M.; Pereira, H.M. Rewilding abandoned landscapes in Europe. Ecosystems 2012, 15, 900–912. [Google Scholar] [CrossRef]
- MacDonald, D.; Crabtree, J.R.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. Journal of Environmental Management 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Brambilla, M.; Gustin, M.; Vitulano, S.; Falco, R.; Bergero, V.; Negri, I.; Bogliani, G.; Celada, C. Sixty years of habitat decline: Impact of land-cover changes in northern Italy on the decreasing ortolan bunting Emberiza hortulana. Regional Environmental Change 2017, 17, 323–333. [Google Scholar] [CrossRef]
- Biró, M.; Molnár, Z.; Öllerer, K.; Lengyel, A.; Ulicsni, V.; Szabados, K.; Kiš, A.; Perić, R.; Demeter, L.; Babai, D. Conservation and herding co-benefit from traditional extensive wetland grazing. Agriculture, Ecosystems & Environment 2020, 300, 106983. [Google Scholar] [CrossRef]
- Lomba, A.; Moreira, F.; Klimek, S.; Jongman, R.H.G.; Sullivan, C.; Moran, J.; Poux, X.; Honrado, J.P.; Pinto-Correia, T.; Plieninger, T.; McCracken, D.I. Back to the future: Rethinking socioecological systems underlying high nature value farmlands. Frontiers in Ecology and the Environment 2019, 18, 36–42. [Google Scholar] [CrossRef]
- Liu, J.; Dietz, T.; Carpenter, S.R.; Alberti, M.; Folke, C.; Moran, E.; Pell, A.N.; Deadman, P.; Kratz, T.; Lubchenco, J.; Taylor, W.W.; et al. Complexity of Coupled Human and Natural Systems. Science 2007, 317, 1513–1516. [Google Scholar] [CrossRef]
- Kirwan, M.C. Ecological embeddedness: An interrogation and refinement of the concept within the context of alternative food networks in the UK. Journal of Rural Studies 2011, 27, 322–330. [Google Scholar] [CrossRef]
- Rotherham, I.D. Cultural landscapes and problems associated with the loss of tradition and custom: An introduction and overview. In Cultural Severance and the Environment. Environmental History; Rotherham, I., Ed.; Springer: Dordrecht, 2013; Volume 2. [Google Scholar] [CrossRef]
- Vos, W.; Meekes, H. Trends in European cultural landscape development: Perspectives for a sustainable future. Landscape and Urban Planning 1999, 46, 3–14. [Google Scholar] [CrossRef]
- Bobiec, A.; Paderewski, J.; Gajdek, A. Urbanisation and globalised environmental discourse do not help to protect the bio-cultural legacy of rural landscapes. Landscape and Urban Planning 2021, 208, 104038. [Google Scholar] [CrossRef]
- Aswani, S.; Lemahieu, A.; Sauer, W.H.H. Global trends of local ecological knowledge and future implications. PLoS ONE 2018, 13, e0195440. [Google Scholar] [CrossRef]
- Barthel, S.; Crumley, C.; Svedin, U. Bio-cultural refugia—Safeguarding diversity of practices for food security and biodiversity. Global Environmental Change 2013, 23, 1142–1152. [Google Scholar] [CrossRef]
- Pérez-Caselles, C.; Brugarolas, M.; Martínez-Carrasco, L. Traditional varieties for local markets: A sustainable proposal for agricultural SMEs. Sustainability 2020, 12, 4517. [Google Scholar] [CrossRef]
- Tüxen, R. Die heutige potentielle natürliche vegetation als gegenstand der vegetationskartierung. Angewandte Pflanzensoziologie 1956, 13, 5–55. [Google Scholar]
- Matuszkewicz, W. Przewodnik do oznaczania zbiorowisk roślinnych Polski. [The guide to identification of plant communities of Poland] Vademecum Geobotanicum 3, Wyd. Nauk. PWN, Warszawa, 2008. [in Polish].
- Matuszkiewicz, J. Potential natural vegetation of Poland. Institute of Geography and Spatial Organization, Polish Academy of Sciences (IGSO PAS), Warsaw, 2008. https://www.igipz.pan.pl/potential-vegetation-dge.html.
- Zarzycki, J.; Korzeniak, J.; Perzanowska, J. Impact of land use changes on the diversity and conservation status of the vegetation of mountain grasslands (Polish Carpathians). Land 2022, 11. [Google Scholar] [CrossRef]
- Affek, A.N.; Wolski, J.; Zachwatowicz, M.; Ostafin, K.; Radeloff, V.C. Effects of post-WWII forced displacements on long-term landscape dynamics in the Polish Carpathians. Landscape and Urban Planning 2021, 214, 104164. [Google Scholar] [CrossRef]
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest adaptation to climate change—Is non-management an option? Annals of Forest Science 2019, 76, 48. [Google Scholar] [CrossRef]
- Olff, H.; Vera, F.; Bokdam, J.; Bakker, E.; Gleichman, J.; De Maeyer, K.; Smit, R. Associational resistance of plants to herbivory may lead to shifting mosaics in grazed woodlands. Plant Biology 1999, 1, 127–137. [Google Scholar] [CrossRef]
- Öllerer, K. The ground vegetation management of wood-pastures in Romania – Insights in the past for conservation management in the future. Applied Ecology and Environmental Research 2014, 12, 549–562. [Google Scholar] [CrossRef]
- Bobiec, A.; Podlaski, R.; Ortyl, B.; Korol, M.; Havryliuk, S.; Öllerer, K.; Ziobro, J.M.; Pilch, K.; Dychkevych, V.; Dudek, T.; Mázsa, K.; Varga, A.; Angelstam, P. Top-down segregated policies undermine the maintenance of traditional wooded landscapes: Evidence from oaks at the European Union’s eastern border. Landscape and Urban Planning 2019, 189, 247–259. [Google Scholar] [CrossRef]
- Hector, A.; Hautier, Y.; Saner, P.; Wacker, L.; Bagchi, R.; Joshi, J.; Scherer-Lorenzen, M.; Spehn, E.M.; Bazeley-White, E.; Weilenmann, M.; Loreau, M.; et al. General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding. Ecology 2010, 91, 2213–2220. [Google Scholar] [CrossRef]
- Muraina, T.O.; Xu, C.; Yu, Q.; Yang, Y.; Jing, M.; Jia, X.; Jaman, M.S.; Dam, Q.; Knapp, A.K.; Collins, S.L.; Smith, M.D.; et al. Species asynchrony stabilises productivity under extreme drought across Northern China grasslands. Journal of Ecology 2021, 109, 1665–1675. [Google Scholar] [CrossRef]
- Gámez-Virués, S.; Perović, D.J.; Gossner, M.M.; Börschig, C.; Blüthgen, N.; de Jong, H.; Simons, N.K.; Klein, A.-M.; Krauss, J.; Westphal, C.; et al. Landscape simplification filters species traits and drives biotic homogenization. Nature Communications 2015, 6, 8568. [Google Scholar] [CrossRef] [PubMed]
- Dass, P.; Houlton, B.Z.; Wang, Y.; Warlind, D. Grasslands may be more reliable carbon sinks than forests in California. Environmental Research Letters 2018, 13, 074027. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Global Change Biology 2018, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Horak, J.; Vodka, S.; Kout, J.; Halda, J.P.; Bogusch, P.; Pech, P. Biodiversity of most dead wood-dependent organisms in thermophilic temperate oak woodlands thrives on diversity of open landscape structures. Forest Ecology and Management 2014, 315, 80–85. [Google Scholar] [CrossRef]
- Miklín, J.; Čížek, L. Erasing a European biodiversity hot-spot: Open woodlands, veteran trees and mature forests succumb to forestry intensification, succession, and logging in a UNESCO Biosphere Reserve. Journal for Nature Conservation 2014, 22, 35–41. [Google Scholar] [CrossRef]
- Bruun, H.H.; Fritzbøger, B. The past impact of livestock husbandry on dispersal of plant seeds in the landscape of Denmark. AMBIO: A Journal of the Human Environment 2002, 31, 425–431. [Google Scholar] [CrossRef]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodiversity and Conservation 2010, 19, 2995–3014. [Google Scholar] [CrossRef]
- Coutinho, P.; Simões, M.; Pereira, C.; Paiva, T. Sustainable local exploitation and innovation on meat products based on the autochthonous bovine breed Jarmelista. Sustainability 2021, 13, 2515. [Google Scholar] [CrossRef]
- Wolański, P.; Bobiec, A.; Ortyl, B.; Makuch-Pietraś, I.; Czarnota, P.; Ziobro, J.; Korol, M.; Havryliuk, S.; Paderewski, J.; Kirby, K. The importance of livestock grazing at woodland-grassland interface in the conservation of rich oakwood plant communities in temperate Europe. Biodiversity and Conservation 2021, 30, 741–760. [Google Scholar] [CrossRef]
- Braun-Blanquet, J.; Pavillard, J. Vocabulaire de sociologie végétale. Roumégous et Déhan, Montpellier, 1925.
- Wang, B.; Shi, W.; Miao, Z. Confidence analysis of standard deviational ellipse and its extension into higher dimensional euclidean space. PLoS ONE 2015, 10, e0118537. [Google Scholar] [CrossRef] [PubMed]
- Marascuilo, L.A. Large-sample multiple comparisons. Psychological Bulletin 1966, 65, 280–290. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists (AOAC). 2020. http://www.eoma.aoac.org/.
- Zaguła, G.; Zardzewiały, M.; Saletnik, B.; Bajcar, M.; Czernicka, M.; Grabek-Lejko, D.; Kasprzyk, I.; Puchalski, C. Effects of fertiliser use and pre-sowing seed stimulation with a magnetic field on the mineral content and yield of three varieties of sugar beet roots. Journal of Elementology 2017, 22, 1401–1414. [Google Scholar] [CrossRef]
- Jaworski, A.; Kołodziej, Z.; Porada, K. Structure and dynamics of stands of primeval character in selected areas of the Bieszczady National Park. Journal of Forest Science 2002, 48, 185–201. https://www.agriculturejournals.cz/publicFiles/285822.pdf. [CrossRef]
- Spracklen, B.D.; Spracklen, D.V. Old-Growth Forest Disturbance in the Ukrainian Carpathians. Forests 2020, 11, 151. [Google Scholar] [CrossRef]
- Bobiec, A.; Reif, A.; Öllerer, K. Seeing the oakscape beyond the forest: A landscape approach to the oak regeneration in Europe. Landscape Ecology 2018, 33, 513–528. [Google Scholar] [CrossRef]
- Peterken, G.F.; Game, M. Historical factors affecting the number and distribution of vascular plant species in the woodlands of Central Lincolnshire. Journal of Ecology 1984, 72, 155–182. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Gawroński, S. The role of woodland fragments, soil types, and dominant species in secondary succession on the western Carpathian foothills. Vegetatio 1994, 111, 149–160. [Google Scholar] [CrossRef]
- Arcanum-maps. Europe in the XIX. Century. 2022. https://tinyurl.com/arcanummaps.
- Bobiec, A.; Kuijper, D.P.J.; Niklasson, M.; Romankiewicz, A.; Solecka, K. Oak (Quercus robur L.) regeneration in early successional woodlands grazed by wild ungulates in the absence of livestock. Forest Management and Ecology 2011, 262, 780–790. [Google Scholar] [CrossRef]
- Kowalczyk, R.; Kamiński, T.; Borowik, T. Do large herbivores maintain open habitats in temperate forests? Forest Ecology and Management 2021, 494, 119310. [Google Scholar] [CrossRef]
- Vandermeulen, S.; Ramírez-Restrepo, C.A.; Beckers, Y.; Claessens, H.; Bindelle, J. Agroforestry for ruminants: A review of trees and shrubs as fodder in silvopastoral temperate and tropical production systems. Animal Production Science 2018, 58, 767–777. [Google Scholar] [CrossRef]
- Slotte, H. Harvesting of leaf-hay shaped the Swedish landscape. Landscape Ecology 2001, 16, 691–702. [Google Scholar] [CrossRef]
- Treydte, A.C.; Heitkönig, I.M.A.; Prins, H.H.T.; Ludwig, F. Trees improve grass quality for herbivores in African savannas. Perspectives in Plant Ecology, Evolution and Systematics 2007, 8, 197–205. [Google Scholar] [CrossRef]
- Van Laer, E.; Tuyttens, F.A.M.; Ampe, B.; Sonck, B.; Moons, C.P.H.; Vandaele, L. Effect of summer conditions and shade on the production and metabolism of Holstein dairy cows on pasture in temperate climate. Animal 2015, 9, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.M.; Barry, T.N.; Cameron, P.N.; Lopez-Villalobos, N.; Cameron, D.J. Willow (Salix sp.) as a supplement for grazing cattle under drought conditions. Animal Feed Science and Technology 2003, 104, 1–11. [Google Scholar] [CrossRef]
- Papanastasis, V.P.; Yiakoulaki, M.D.; Decandia, M.; Dini-Papanastasi, O. Integrating woody species into livestock feeding in the Mediterranean areas of Europe. Animal Feed Science and Technology 2008, 140, 1–17. [Google Scholar] [CrossRef]
- Gowda, N.K.S.; Ramana, J.V.; Prasad, C.S.; Singh, K. Micronutrient content of certain tropical conventional and unconventional feed resources of southern India. Tropical Animal Health and Production 2004, 36, 77–94. [Google Scholar] [CrossRef]
- Bakshi, M.; Wadhwa, M. Comparative nutritional evaluation of forest tree leaves and wild grasses of semi hilly arid zone in Punjab. Indian Journal of Animal Sciences 2012, 82. [Google Scholar]
- Tefera, S.; Mlambo, V. Macro and trace elements of 26 browse species growing in semi-arid grazing lands of Eastern Cape, South Africa. African Journal of Range & Forage Science 2017, 34, 209–217. [Google Scholar] [CrossRef]
- Mateos, G.; Fondevila, G.; Cámara, L. Chapter 3: The importance of the fibre fraction of the feed in non-ruminant diets. 2019, pp. 61–83. [CrossRef]
- González-Hernández, M.P.; Karchesy, J.; Starkey, E.E. Research observation: Hydrolyzable and condensed tannins in plants of northwest spain forests. Journal of Range Management 2003, 56, 461–465. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Ruminant Research 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Lisonbee, L.D.; Villalba, J.J.; Provenza, F.D.; Hall, J.O. Tannins and self-medication: Implications for sustainable parasite control in herbivores. Behavioural Processes 2009, 82, 184–189. [Google Scholar] [CrossRef]
- Grainger, C.; Clarke, T.; Auldist, M.J.; Beauchemin, K.; McGinn, S.; Waghorn, G.C.; Eckard, R. Potential use of Acacia mearnsii condensed tannins to reduce methane emissions and nitrogen excretion from grazing dairy cows. Canadian Journal of Animal Science 2009, 89, 241–251. [Google Scholar] [CrossRef]
- Altermann, E.; Reilly, K.; Young, W.; Ronimus, R.S.; Muetzel, S. Tailored nanoparticles with the potential to reduce ruminant methane emissions. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef]
- Marie, P.J.; Ammann, P.; Boivin, G.; Rey, C. Mechanisms of action and therapeutic potential of strontium in bone. Calcified Tissue International 2001, 69, 121–129. [Google Scholar] [CrossRef]
- Nielsen, S.P. The biological role of strontium. Bone 2004, 35, 583–588. [Google Scholar] [CrossRef]
Figure 1.
Study area location (A), four zones of tree stem density (number of stems per 2000 m2 and white rectangular area in the north indicating the fenced farmstead enclave (B), LiDAR terrain model (source: MassGIS Data: Lidar DEM and Shaded Relief – https://www.mass.gov/info-details/massgis-data-lidar-dem-and-shaded-relief (C), and treescape (D).
Figure 1.
Study area location (A), four zones of tree stem density (number of stems per 2000 m2 and white rectangular area in the north indicating the fenced farmstead enclave (B), LiDAR terrain model (source: MassGIS Data: Lidar DEM and Shaded Relief – https://www.mass.gov/info-details/massgis-data-lidar-dem-and-shaded-relief (C), and treescape (D).
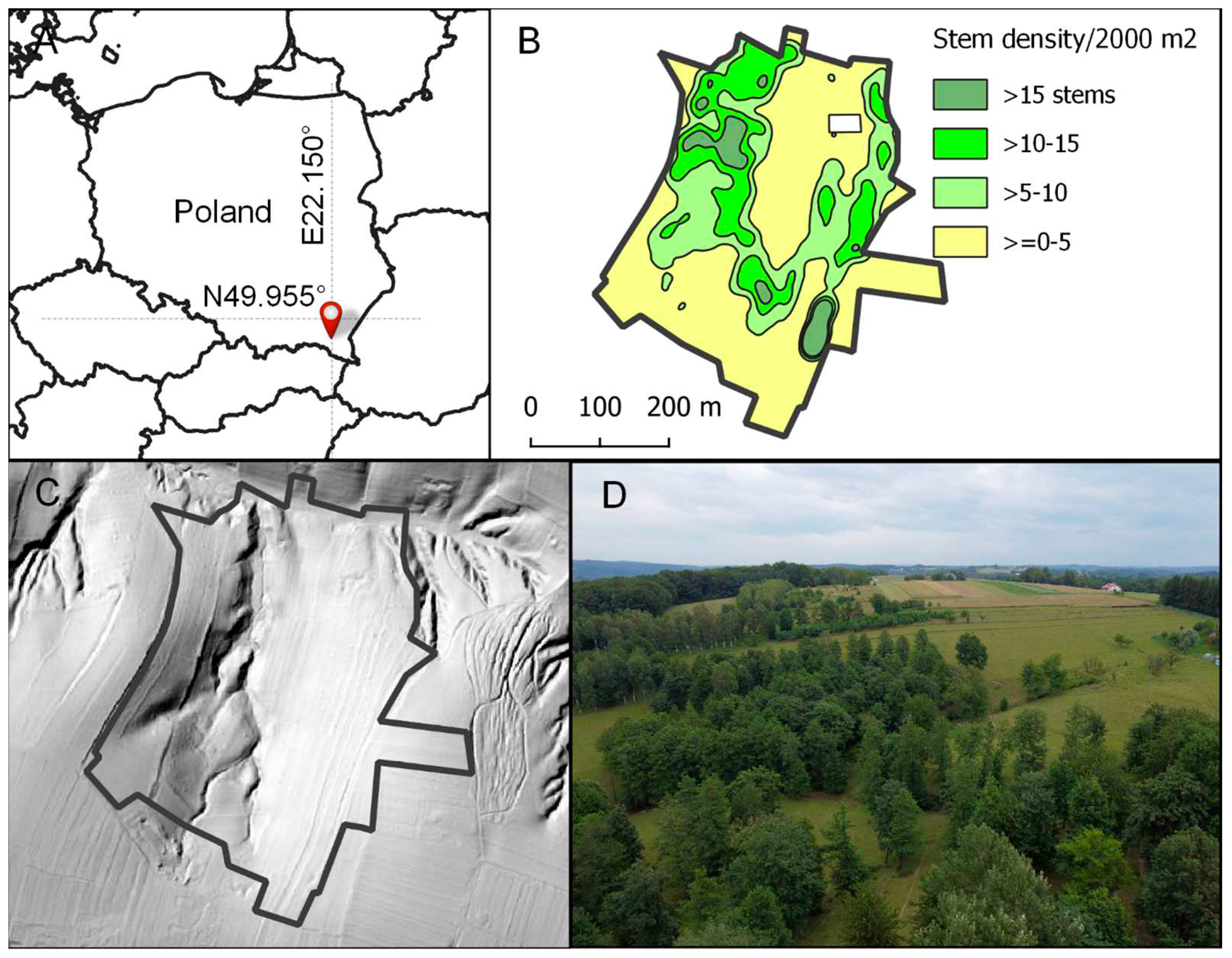
Figure 2.
Wood-pasture as a variegated landscape. A: the distribution of stems of the most frequent tree species; Quro – Quercus robur, Ceav – Cerasius avium, Acps – Acer pseudoplatanus, Cabe – Carpinus betulus, Bepe – Betula pendula, Algl – Alnus glutinosa, Fasy – Fagus sylvatica B: the major habitat categories; OGR – open grassland; SOW – treed grassland; PIG – pioneering birch (Betula pendula)/alder (Alnus glutinosa) groves; RIW – riparian wood with alder; CCW – close-canopy wood, dominated by shade-tolerant tree species; red dots – centroids of tree fodder stacks.
Figure 2.
Wood-pasture as a variegated landscape. A: the distribution of stems of the most frequent tree species; Quro – Quercus robur, Ceav – Cerasius avium, Acps – Acer pseudoplatanus, Cabe – Carpinus betulus, Bepe – Betula pendula, Algl – Alnus glutinosa, Fasy – Fagus sylvatica B: the major habitat categories; OGR – open grassland; SOW – treed grassland; PIG – pioneering birch (Betula pendula)/alder (Alnus glutinosa) groves; RIW – riparian wood with alder; CCW – close-canopy wood, dominated by shade-tolerant tree species; red dots – centroids of tree fodder stacks.
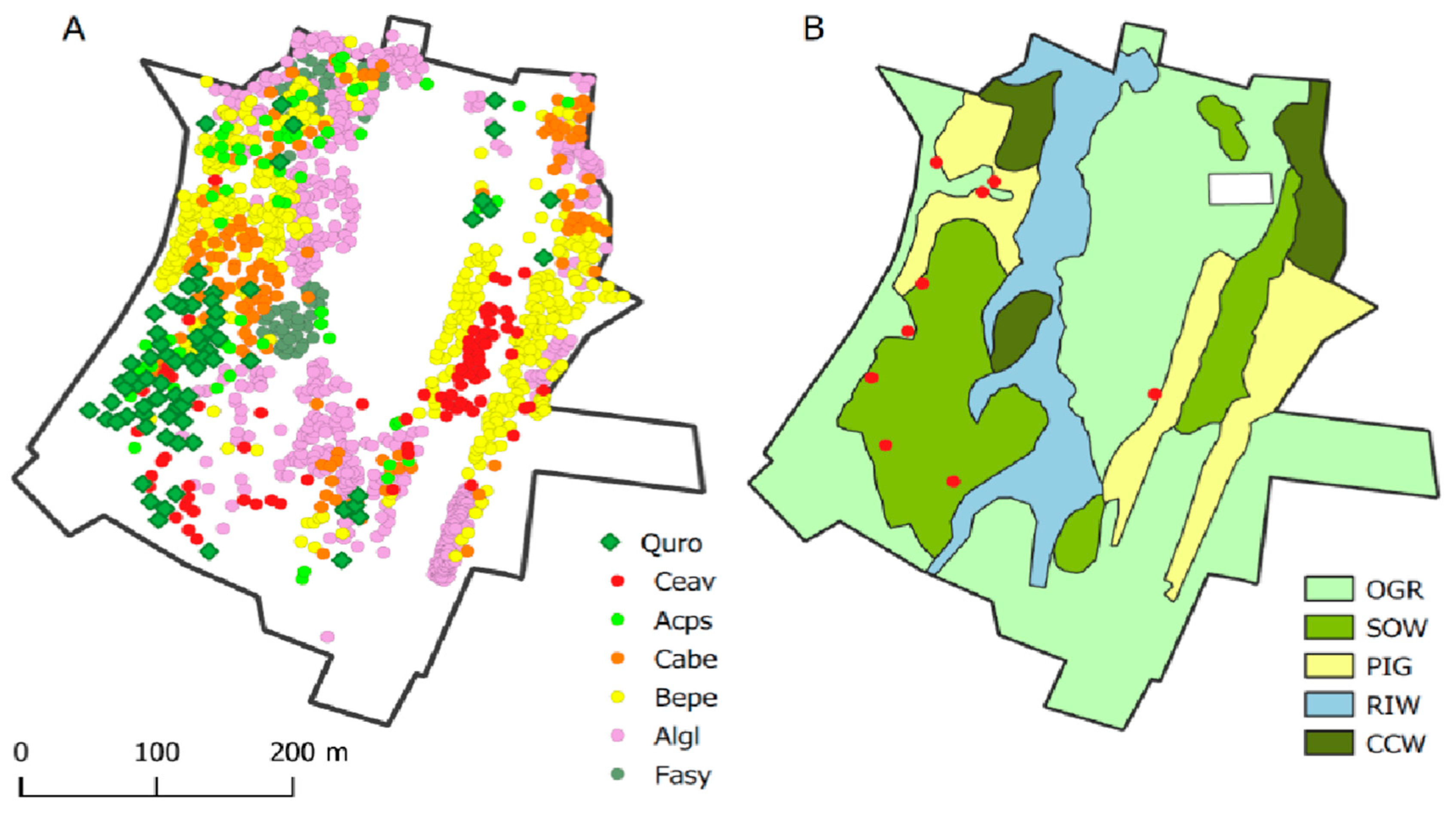
Figure 6.
Comparison of the average proportions of the habitats use by the cattle to the shares (%) of these habitats in the whole wood-pasture area; the numbers above the first and second bars are the p-values of the single proportion test. The “4 cows” indicate the four tagged-cows, while “groupings” represent the percentage of the total temporary cattle gatherings of bulls, cows, and heifers in the wood-pasture; for habitat categories see Figure 2.
Figure 6.
Comparison of the average proportions of the habitats use by the cattle to the shares (%) of these habitats in the whole wood-pasture area; the numbers above the first and second bars are the p-values of the single proportion test. The “4 cows” indicate the four tagged-cows, while “groupings” represent the percentage of the total temporary cattle gatherings of bulls, cows, and heifers in the wood-pasture; for habitat categories see Figure 2.
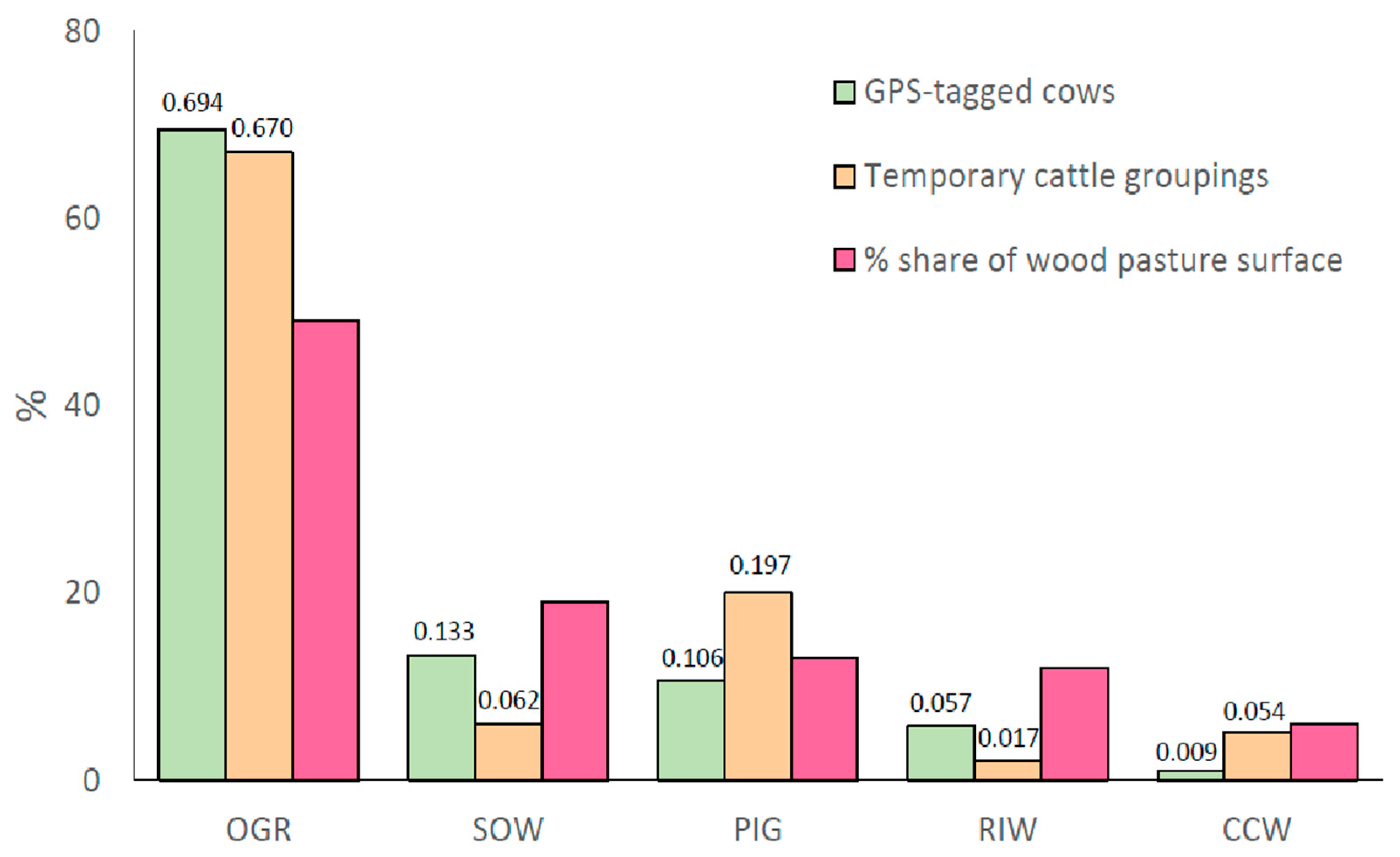
Figure 7.
Comparison of the temporary groupings’ parameters in three consecutive days (the tiny boxes in the big boxes represent medians; lower and upper edges of each big box represent Q1 and Q3 respectively; lower and upper bars across each big box show the min and max values in each set respectively; p represents the p-value of Kruskal-Wallis test; N means number; and SD ellipses means standard deviational ellipses).
Figure 7.
Comparison of the temporary groupings’ parameters in three consecutive days (the tiny boxes in the big boxes represent medians; lower and upper edges of each big box represent Q1 and Q3 respectively; lower and upper bars across each big box show the min and max values in each set respectively; p represents the p-value of Kruskal-Wallis test; N means number; and SD ellipses means standard deviational ellipses).
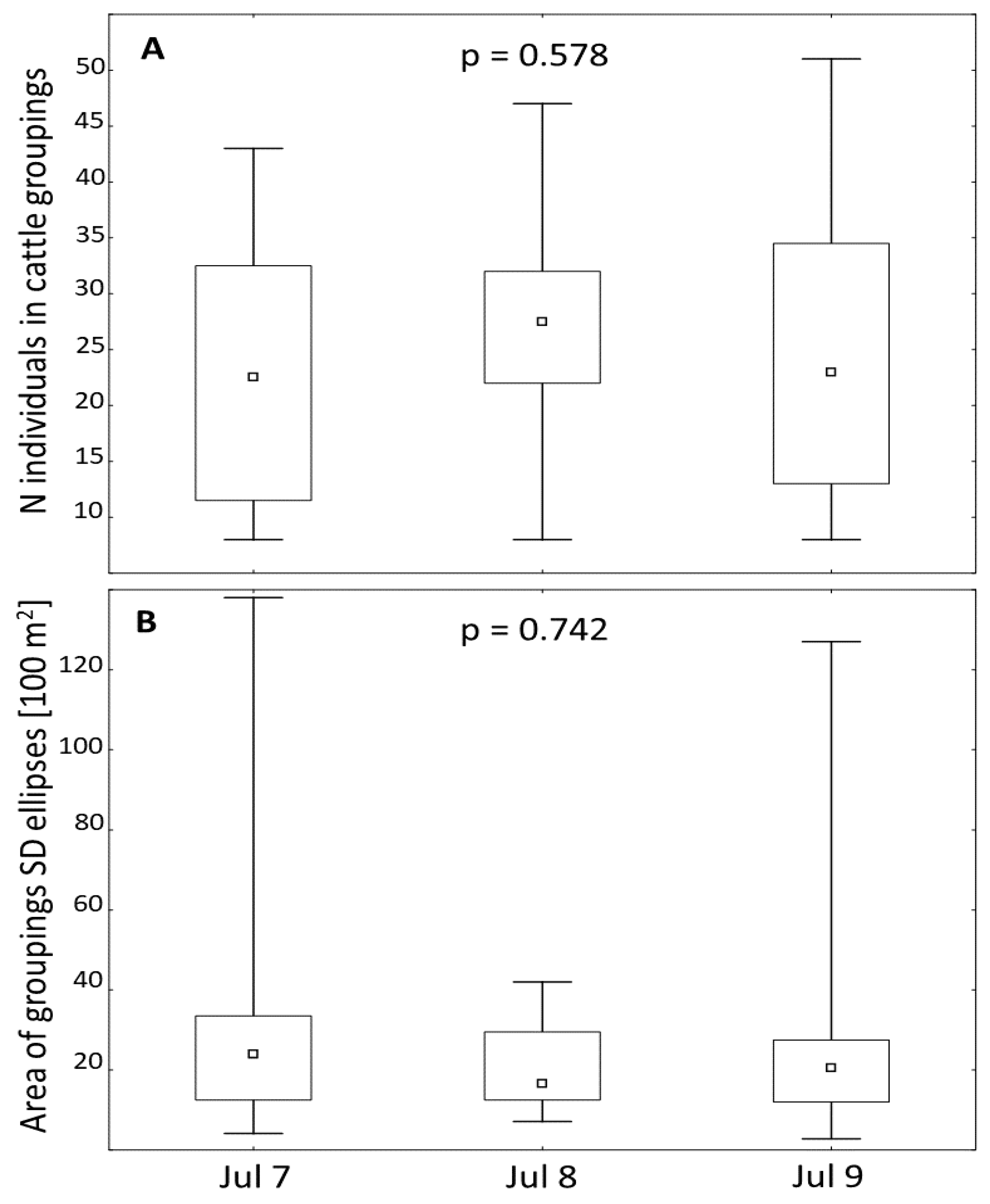
Figure 8.
Comparison (medians, Q1-Q3, min-max) of the LTI as observed in natural cattle groupings on herbaceous forages during three consecutive days (Jul 7-9; LTI) to LTIa base on phototrap record of the cattle groupings on arboreal forages (Arb1 – stacks of branches, Arb2 – felled whole crowns; LTIa); table in the top: p-values of the multiple comparisons on ranks of five independent samples; Kruskal-Wallis test: H (4, N = 69) = 24.024, p = 0.0001).
Figure 8.
Comparison (medians, Q1-Q3, min-max) of the LTI as observed in natural cattle groupings on herbaceous forages during three consecutive days (Jul 7-9; LTI) to LTIa base on phototrap record of the cattle groupings on arboreal forages (Arb1 – stacks of branches, Arb2 – felled whole crowns; LTIa); table in the top: p-values of the multiple comparisons on ranks of five independent samples; Kruskal-Wallis test: H (4, N = 69) = 24.024, p = 0.0001).
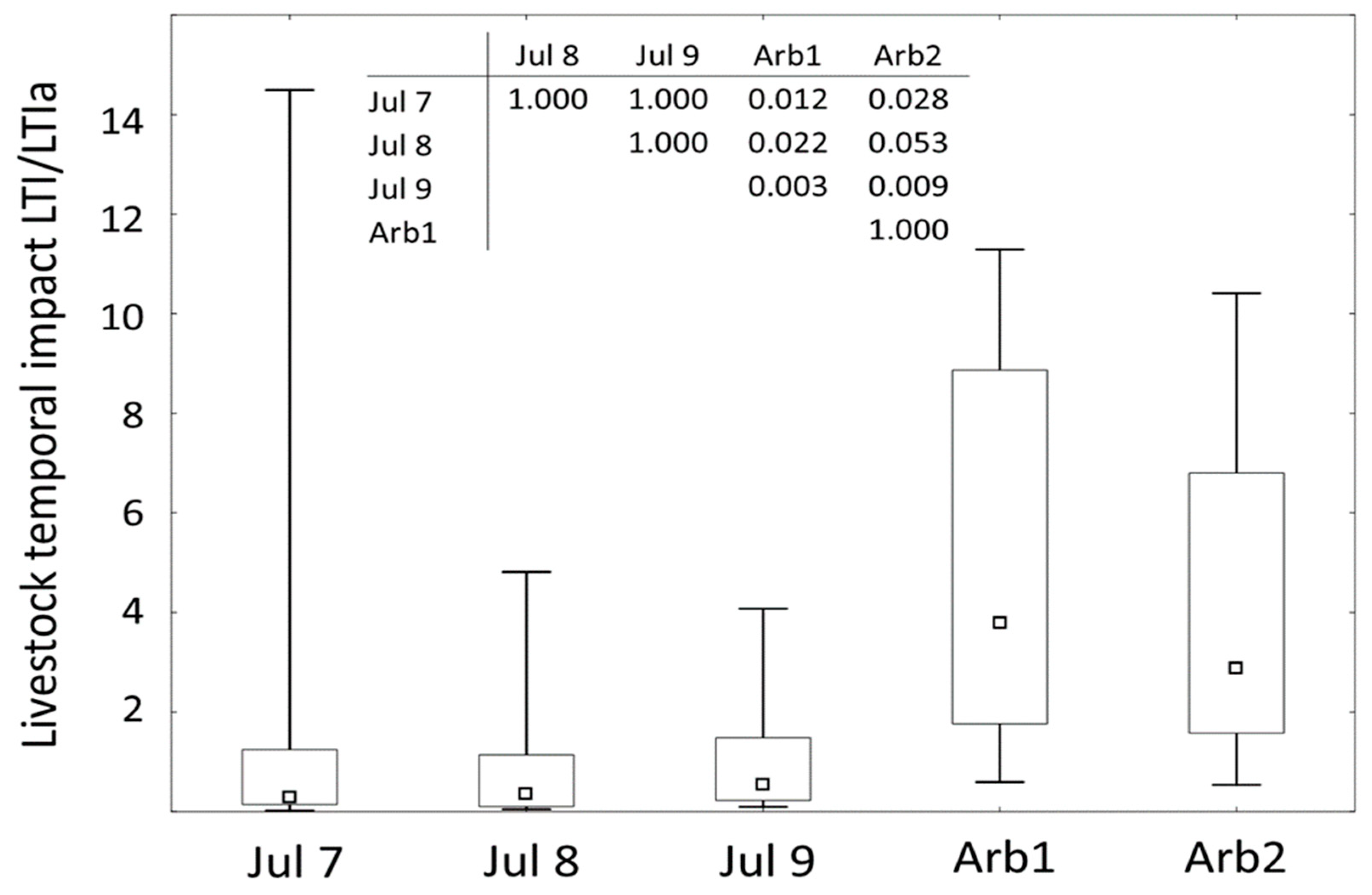
Figure 9.
Conceptual model of the alternative changes in land-use following agricultural abandonment, with respect to the ecosystem services structure.
Figure 9.
Conceptual model of the alternative changes in land-use following agricultural abandonment, with respect to the ecosystem services structure.
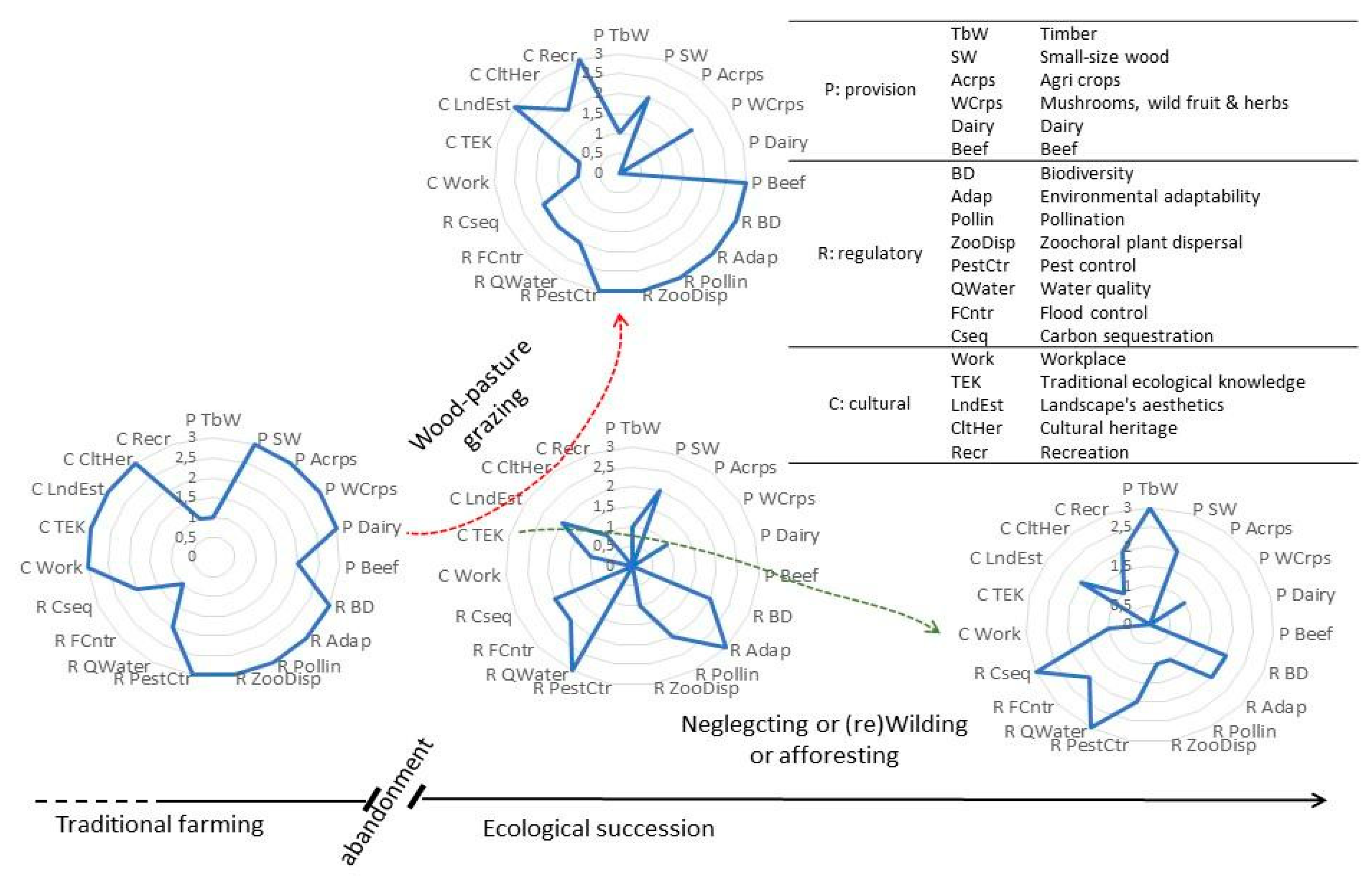

Table 1.
Total number of herbaceous species recorded in all relevés representing particular habitat categories, broken down by species affiliation in woodland and non-woodland plant communities.
Table 1.
Total number of herbaceous species recorded in all relevés representing particular habitat categories, broken down by species affiliation in woodland and non-woodland plant communities.
| Species affiliation | CCW | OGR | PIG | RIW | SOW |
| Woodland communities | 48 | 13 | 14 | 16 | 9 |
| Grassland and ruderal communities | 29 | 249 | 93 | 65 | 99 |
Table 2.
Comparison of nutritional composition of herbaceous forages (including leaves, flowers, and aboveground stems regarded as ‘Herbs’), tree leaves (Leaves), and tree twigs (Twigs).
Table 2.
Comparison of nutritional composition of herbaceous forages (including leaves, flowers, and aboveground stems regarded as ‘Herbs’), tree leaves (Leaves), and tree twigs (Twigs).
| Nutrient | K-W p | Leaves | Twigs | Herbs | Mmin | Mmax | |
|---|---|---|---|---|---|---|---|
| Protein | 0.000 | Herbs | 0.021↑ | 0.023↓ | Bepe.t | Algl.l | |
| Leaves | 0.000↓ | 10.732 | 5.117 | 19.860 | |||
| Fat | 0.000 | Herbs | 0.001↑ | 1.000 | Acps.t | Algl.l | |
| Leaves | 0.000↓ | 2.204 | 0.722 | 6.657 | |||
| Fibre | 0.000 | Herbs | 0.002↓ | 0.210↑ | Ceav.l | Cabe.t | |
| Leaves | 0.000↑ | 33.886 | 16.700 | 45.334 | |||
| Ash | 0.000 | Herbs | 0.092↓ | 0.000↓ | Bepe.t | ||
| Leaves | 0.000↓ | 9.523 | 2.691 | 9.523 | |||
| NFE | 0.000 | Herbs | 0.000↑ | 0.002↑ | Cabe.l | ||
| Leaves | 0.049↓ | 43.784 | 43.784 | 55.512 |
Note: NFE- nitrogen-free extracts; K-W p- Kruskal-Wallis test’s p-values; values in ‘Leaves’ and ‘Twigs’ columns indicate p-values of the multiple comparisons on ranks with ‘Herbs’, and ‘Leaves’ samples (third column); ↑ and ↓ respectively indicate superiority and inferiority of an element content median in ‘Leaves’ and ‘Twigs’ compared to ‘Herbs’ and ‘Leaves’; values in the ‘Herbs’ column indicate median of an element content; Mmin and Mmax respectively denote maximum and minimum median values attributable to ‘Herbs’ or leaves (.l) or twigs (.t) of particular tree species. Ca – Cerasus avium, Bp – Betula pendula, Ag – Alnus glutinosa, Cb – Carpinus betulus, Ap – Acer pseudoplatanus, Qr – Quercus r.obur
Table 3.
Comparison of herbaceous forage (including leaves, flowers, and aboveground stems referred as ’Herbs’), tree leaves (Leaves), and tree twigs (Twigs) with regards to the content of selected elements (g kg-1 of dry mass).
Table 3.
Comparison of herbaceous forage (including leaves, flowers, and aboveground stems referred as ’Herbs’), tree leaves (Leaves), and tree twigs (Twigs) with regards to the content of selected elements (g kg-1 of dry mass).
| Element. | K-W p | Leaves | Twigs | Herbs | Mmin | Mmax | |
|---|---|---|---|---|---|---|---|
| Ca | 0.000 | Herbs | 0.000↑ | 0.000↑ | Ca.l | ||
| Leaves | 0.996↓ | 5.482 | 5.482 | 16.630 | |||
| K | 0.000 | Herbs | 0.139↓ | 0.000↓ | Bp.t | Ca.l | |
| Leaves | 0.000↓ | 16.962 | 8.696 | 17.164 | |||
| Mg | 0.000 | Herbs | 0.409↑ | 0.001↓ | Ag.t | Ca.l | |
| Leaves | 0.000↓ | 2.128 | 0.606 | 4.063 | |||
| P | 0.000 | Herbs | 1.000 | 0.000↓ | Ca.t | ||
| Leaves | 0.000↓ | 2.247 | 0.896 | 2.247 | |||
| S | 0.000 | Herbs | 1.000 | 0.000↓ | Ca.t | Ag.l | |
| Leaves | 0.000↓ | 1.634 | 0.430 | 1.753 | |||
| Mn | 0.008 | Herbs | 0.144↑ | 1.000 | Ca.t | Cb.l | |
| Leaves | 0.010↓ | 0.321 | 0.116 | 1.987 | |||
| Fe | 0.066 | Herbs | 0.066↓ | 0.133↓ | Ap.t | ||
| Leaves | 1.000 | 0.175 | 0.050 | 0.175 | |||
| Al | 0.000 | Herbs | 0.194↓ | 0.000↓ | Qr.t | Cb.l | |
| Leaves | 0.003↓ | 0.177 | 0.023 | 0.443 | |||
| Sr | 0.000 | Herbs | 0.008↑ | 0.000↑ | Cb.l | ||
| Leaves | 0.005↑ | 0.020 | 0.020 | 0.043 | |||
| Zn | 0.020 | Herbs | 1.000 | 0.052↑ | Ca.l | Bp.l | |
| Leaves | 0.082↑ | 0.024 | 0.013 | 0.122 |
See footnote of Table 1 for the explanation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



