
Submitted:
04 August 2023
Posted:
07 August 2023
You are already at the latest version
Abstract
Avenanthramides (Avns) and its derivatives, a group of polyphenolic compounds found abundantly in Oats (Avena sativa Linn.) have emerged as a promising candidate for neuroprotection due to their immense anti-oxidant, anti-inflammatory, and anti-apoptotic properties. Neurodegenerative diseases (NDDs), characterized by progressive degeneration of neurons, present a significant global health burden with limited therapeutic options. The phosphoinositide 3-kinase (PI3K) signalling pathway plays a crucial role in cell survival, growth, and metabolism, making it an attractive target for therapeutic intervention. Dysregulation of PI3K signalling has been implicated in the pathogenesis of various NDDs including Alzheimer's and Parkinson's disease. Avns has been shown to modulate PI3K/AKT signalling, leading to increased neuronal survival, reduced oxidative stress, and improved cognitive function. This review explores the potential of Avns polyphenols, as modulators of the PI3K signalling pathway focusing on their beneficial effects against NDDs. Further, we outlined the need for clinical exploration to elucidate the specific mechanisms of Avns action on the PI3K/AKT pathway and its potential interactions with other signalling cascades involved in neurodegeneration. Based on the available literature using relevant keywords from Google Scholar, PubMed, Scopus, Science Direct, and Web of Science, our review emphasizes the potential of Avns as a therapeutic strategy for NDDs and warrants further investigation and clinical exploration.
Keywords:
Avenanthramide
; AKT-Protein kinase B
; Alzheimer's disease
; Parkinson's disease
; oxidative stress
; neuroprotection
1. Introduction
Since ancient times, cultivated oats has been an important cereal crop for human consumption worldwide. Recently, interest in oats has been increased due to their nutritional properties and associated health benefits [1]. In the world grain ranking, oats were placed sixth for their nutritional values and regarded as most suitable for cultivation under variable climatic and soil conditions [2,3]. Cereal oats contain numerous bioactive compounds with high nutritive values including cellulose, arabinoxylan, β-glucans, proteins, unsaturated fatty acids, vitamins, minerals, phenols and dietary fibre [4,5]. Studies suggest that regular consumption of oat grains is associated with a lower risk of various chronic diseases [6]. Meta-analysis studies also revealed that regular oat intake lowers blood cholesterol, improves insulin sensitivity and post-prandial glycaemic control [7,8]. The water-soluble β-glucan contained in oat seed mainly lowers cholesterol levels, leading to a reduced risk of heart disease in humans [9,10]. Further, pharmacological studies indicated that the polyphenolic active constituents in oats exhibited strong anti-inflammatory, anti-bacterial, anti-carcinogenic, cytotoxic, and anti-proliferative properties [11,12]. For this reason, the human demand for oat consumption has been drastically increased.
In the central nervous system (CNS), different signalling pathways regulates various cellular activities and physiological functions. In particular, the protective role of the phosphoinositide 3-kinase (PI3K) signalling and Protein kinase B (AKT) has been widely reported in neurodegenerative conditions. PI3K/AKT signalling pathway is functional in many CNS processes such as synaptic plasticity, neurogenesis, proliferation and differentiation, aging, and neuronal autophagy [13]. In addition, PI3K also plays a predominant role in molecular trafficking [14]. The PI3K/AKT pathway constitutes a major signalling cascade, which consists of PI3K, a multifaceted protein and, its downstream molecules including glycogen synthase kinase-3 beta (GSK-3β), mammalian target of rapamycin (mTOR), and nuclear factor erythroid 2-related factor (Nrf) [15].
In a growing number of studies, neuroprotective agents from natural products are involved in targeting PI3K/AKT signalling, thus contributing to the prevention and treatment of neurodegenerative diseases (NDDs) including Alzheimer’s disease (AD) and Parkinson’s disease (PD) [16,17]. Although great efforts have been made to understand the pathogenesis of NDD and the design of effective treatment to delay progression, there is still no potential therapy available. Thus, the interest in derived natural compounds with potential benefits in CNS disorders has been substantially growing. Since the mechanisms of actions of natural compounds are variable, suggesting that they could be highly effective and specific in improving neuroprotective capacity.
Avenanthramides (Avns, Figure 1) are the major water-soluble phenolamides obtained exclusively from oats. The therapeutic potential of Avns has been reported in numerous literatures which include the protective role of Avns against several chronic diseases such as cardiovascular diseases, cancer, diabetes, and skin disorders [1]. Recently, Avns and its derivatives were reported to possess neuroprotective effects in experimental models and also improve AD and PD pathologies including memory and behavioural impairments [18,19,20,21].
Among the 25 polyphenolic Avns identified, avenanthramide c (Avn-c), an amide conjugate of anthranilic acid and hydroxycinnamic acid is found to be protective in experimental models of cerebral ischemia [22,23,24]. However, the role of Avns and their derivatives against NDDs and their mechanistic basis have not been fully understood. In the present review, we highlight the potential of Avns as novel therapeutics for the treatment of NDDs and the influence of Avns on the regulation of the PI3K signalling pathway in the pathophysiology of NDDs including AD and PD. The relevant literature using the keywords “avenanthramide”, “PI3K/AKT”, “neurodegenerative diseases”, “AD” and “PD” from Google Scholar, PubMed, Scopus, Science Direct, and Web of Science were selected. The most appropriate and recent articles were prioritized based on the purpose of this review.
2. Modulation of PI3K signalling pathway in NDDs
It is well documented that the dysregulation of PI3K signal transduction significantly contributes in the pathogenesis of various NDDs including AD, PD, and Huntington’s disease [25]. Further, signal transduction involving AKT along with PI3K is known to mediate neuronal survival. Therefore, understanding the role and regulatory aspects of PI3K and AKT pathway might help in the development of suitable therapeutic agents against NDDs targeting PI3K/AKT signalling. Although the role of PI3K/Akt in NDDs was thoroughly reviewed [25], in the following sections the role of PI3K/AKT activation and other related signalling molecular pathways in relation to NDDs was focused.
2.1. PI3K
According to the structure, regulation, and substrate specificity, the PI3Ks have been divided into three subcategories: class I, II, and III [26]. Among this, class I isoform has been studied extensively, which are activated by surface receptors, composed of regulatory and catalytic subunits. These are further subcategorized into class IA and class IB based on the mode of regulation. Class II PI3K enzymes require an additional signal for activation whereas class III PI3K are vacuolar protein sorting 34 (Vps34) required mainly for membrane trafficking [27].
2.2. AKT-Protein kinase B (PKB)
AKT has been considered as a core effector of the PI3K downstream signalling pathway. AKT is a serine/threonine kinase, categorized into three homologous isoforms, AKT1/PKBα, AKT2/PKBβ, and AKT3 [28,29]. It consists of three different domains pleckstrin homology (PH) domain at N-terminal responsible for membrane translocation after activation by PI3K, the central fragment, and the regulatory domain at C-terminal contains the phosphorylation site required for activation of AKT. Among the isoforms, AKT3/PKBγ is crucial for the development of brain and microglial survival.
2.3. Activation of PI3K/AKT pathway
The signalling pathway begins with the interaction between the ligand and transmembrane receptor mainly, tyrosine kinase (RTK), which results in receptor dimerization and autophosphorylation of the intracellular tyrosine domain, leading to the recruitment of PI3. The P85, a regulatory domain on class I PI3K binds to the phosphorylated tyrosine residue following recruitment of p110, a catalytic domain on PI3K responsible for complete activation of PI3K enzyme. The p110 also recruits the inactive AKT and phosphatidylinositol-dependent protein kinase 1 (PDK1) from cytoplasm on the cell membrane that brings conformational changes in AKT exposing phosphorylation site on serine (473) and threonine (308). The activated PI3K also phosphorylates phosphatidylinositol 4,5 biphosphate (PtdIns,4,5, P2) to phosphatidylinositol 3,4,5 triphosphate (PtdIns,3,4,5, P3), which gets dephosphorylated with phosphatase and tensin homologue on chromosome 10 (PTEN), which regulates inactivation of AKT [30].
The phosphorylated PI3K/AKT-Ser473 forms complex with a wide range of downstream signalling molecules including GSK-3β, mTOR, ERK, NF-κB, Hsp, etc to execute diverse cellular activities [16,31]. The downstream target mTOR, regulates the metabolism. Activation of PI3K/AKT also inhibits the process of apoptosis by interacting with different signalling molecules including Bcl2-antagonist of cell death (Bad), bcl-2—like protein 11 (BIM) and caspase-9. NF-κB, one of the downstream modulators of PI3K/AKT regulates the inflammatory response by modulating the expression of inflammatory markers IL-1β, IL-6, TNF-α, iNOS, and COX-2. At the resting state, the activity of NF-κB was repressed by inhibitory kappa B (IκB). The PI3K/AKT interacts with nuclear factor erythrocyte two related factors (Nrf2) to regulate oxidative stress [16]. In NDDs including AD and PD, the PI3K/AKT signalling is altered, resulting in disruption of cellular function including autophagy and synaptic plasticity, thus indicating that targeting the PI3K/AKT pathway or its downstream regulator could be a novel strategy to treat neurodegenerative diseases.
3. PI3K/AKT pathway in neurodegeneration
Neurodegenerative disorders including AD and PD are the category of conditions with selective and progressive loss of structure or function of neurons. These disorders are characterized by the gradual degeneration of neurons resulting in behavioural, learning, emotional, and cognitive abnormalities [32]. The prevalence report suggested a steady increase in the number of people suffering from neurodegenerative conditions. The treatment is targeted to alleviate symptoms and to prevent progression to improve the quality of life. Many studies have been carried out to understand the pathophysiological cause of the progressive neurodegeneration associated with AD and PD, which highlighted several possible targets for the development of neuroprotective strategies for effective treatment [33]. In this section, we review the evidence for the involvement of PI3K/AKT downstream targets in AD and PD. In addition, we highlight the potential targets of the PI3K/AKT pathway for the treatment of NDDs.
Reactive oxygen species (ROS) are produced in normal physiological settings as a bioproduct of cellular metabolism. However, ROS levels are firmly regulated via redox homeostasis. When the production of ROS gets increased in certain pathophysiological conditions due to several risk factors including, environmental stress, mutation, or genetic factors, failing to regulate the excessive production of ROS causes structural, functional, and loss of cellular function. Neurons are particularly vulnerable to oxidative stress conditions due to high consumption of oxygen, low antioxidant levels, high polyunsaturated membrane fatty acids as well as post-mitotic high accumulation of oxidized molecules. In NDDs such as AD and PD in particular, oxidative stress has been considered to be crucial risk factors. Thus increased oxidative stress markers as well as deficient antioxidant defence systems are considered to be common hallmarks in these conditions [34,35,36,37].
One of the main neuroprotective factors that regulate antioxidant and anti-inflammatory genes is Nrf2. Nrf2 modulates oxidative stress by regulating the synthesis of antioxidant enzymes through binding with the Antioxidant Response Element (ARE). Under a physiological setting, the activity of Nrf2 is repressed by a protein, Kelch-like ECH-associated protein 1 (Keap1), an adaptor for E3 ligase in ubiquitin-proteasome pathway [38,39]. However, under stressful conditions Keap1 is inactivated due to post-translational modification leading to migration and accumulation of active Nrf2 inside the nucleus leading to heterodimerization with small protein designated as sMaf, which stimulates expression of antioxidant through binding at ARE on target gene [40,41]. Besides Keap1, activation of the PI3K/AKT pathway can also indirectly regulates Nrf2-ARE signalling through a serine/threonine protein kinase designated as glycogen synthase kinase 3β (GSK-3β) via phosphorylating Nrf2 [42,43]. Mounting evidence suggests that negative regulation of GSK-3β via PI3K/AKT signalling can regulate the activity of Nrf2 through stabilizing and regulating the gene expression of Nrf2 [44,45,46]. Under physiological conditions, activation of PI3K/AKT inhibits GSK-3β through phosphorylation at Ser21-GSK-3α or Ser9-GSK-3β [47].
Several studies investigated that PI3K/AKT/Nrf2/ GSK-3β pathway has been impaired in AD patients and pre-clinical mouse models [48,49,50,51]. Oxidative stress in AD causes downregulation of phosphorylated PI3K, which results in the inactivation of PI3K/AKT pathway leading to GSK-3β activation, which translocates Nrf2 from the nucleus into cytosol resulting in low levels of antioxidant enzymes [52,53,54,55]. In addition, amyloid-β oligomers found to activate GSK-3β, blocking PI3K/AKT/mTOR signalling, which increases phosphorylation of Tau, inducing neurofibrillary tangle formation, a pathophysiological hallmark in AD. These findings suggest the involvement of different downstream molecules of PI3K/AKT in the pathophysiology of AD.
Oxidative stress-induced neuronal death also contributes to the pathology of PD. The PI3K/AKT signalling influences oxidative stress by modulating downstream molecules such as GSK-3β and mTOR. Abnormal expression of GSK-3β has been reported in PD. Several findings indicated that Nrf2 signalling is compromised with age, thus reduced Nrf2 expression in different brain regions has been associated with increased age [56]. Though the expression of Nrf2 has been suggested to be restricted in astrocytes in substantia nigra (SN), still sufficient to protect neurons from 1-Methyl-4-phenyl1,2,3,6-tetrahydropyridine (MPTP) induce neurotoxicity [57]. In addition, data from post-mortem brains indicated Nrf2 dysfunction in PD [58,59]. Also, the levels of downstream targets in PI3K/AKT/mTOR signalling were found to be significantly reduced in PD patients. In PD, the imbalance of oxidative stress has been reported due to disrupted FoxO3a, a downstream target of mTOR [60,61]. Thus, the molecules that activate the PI3K/AKT pathway through activation of p-GSK-3β (Ser9) or mTOR can prevent oxidative stress-induced neuronal death, which can be beneficial in preserving neuronal structure and function.
4. Neuroprotective role of Avn
Avns derived from oats have been shown to exert neuroprotective effects by modulating signaling pathways involved in cell survival, neuronal growth, synaptic plasticity, apoptosis and neuroinflammation. These mechanisms can enhance neuronal resilience and support the maintenance of cognitive function. Oat phenols have been well reported for their pharmacological benefits including antioxidant activities. The polyphenolic combination with different groups as phenolamide tends to potentiate its action [62,63,64]. Avns have been found to exhibit anti-oxidant and anti-inflammatory effects in the CNS, which can be beneficial for reducing stress and neuroinflammation associated with various neurological conditions.
Preclinical studies have suggested that Avns possess neuroprotective properties that could be beneficial in cerebral stroke. Avns can reduce area of tissue damage and improve neurological outcomes following cerebral stroke [19]. These effects have been attributed to the ability of Avns to enhance blood flow, inhibit inflammatory responses, and protect against oxidative damage. Furthermore, Avns have been found to promote neurogenesis and enhance synaptic plasticity, which are crucial for brain repair and functional recovery after stroke. While the research on Avns in neurological conditions is still emerging, the available evidence suggests their potential as neuroprotective agents. In the following sections we discussed the beneficial role of Avns againgt NDDs including AD and PD pathology.
4.1. Alzheimer’s disease
Globally, most geriatric people are suffering from this common condition which is associated with dementia, memory loss, and cognition impairment [65]. According to the annual report of the Alzheimer’s Association, about 6 million American populations belonging to the age group 65-75 years most commonly have AD, and also the incidence will be increasing day by day. Nerve cell destruction, extracellular amyloid plaques, and neurofibrillary tangles (NFTs) within the cell seem to be the markers of this complex NDD. These plaques are composed of amyloid beta (Aβ), a cleavage by-product of amyloid precursor protein (APP) [66]. Moreover, the gradual formation of oligomers, fibrils, and insoluble amyloid plaques from Aβ monomers results in a reduction in the plasticity of neurons in the synaptic domain [67]. In addition, tau protein has been hyperphosphorylated which forms NFTs. In healthy conditions, tau encourages microtubule stabilization. But as paired helical filaments connect, hyperphosphorylated tau builds up and eventually produces NFTs. Accumulation of Aβ leads to dysregulation of synaptic and neuronal activities, which further generates intracellular conditions for NFT production and ultimately results in neuronal death and disruption of neurotransmitter functions [68].
The antioxidant potential of Avn has been confirmed through a variety of in vivo and in vitro studies. The hydroxy and amide group in its structure are mainly responsible for scavenging free radical species formed during a variety of physiological as well as pathological conditions. This phenolamide is also able to induce the synthesis of a variety of cytoprotective enzymes. In addition, Avn contains an unsaturated amide group, which mainly interacts with the cysteine residue in Keap1, thus effectively preventing the phosphorylation and accumulation of Nrf2 in the cytosol and promoting upregulation of transcriptive action of Nrf2 [69,70].
Synaptic dysfunction has a major contribution to memory deficits in AD pathophysiology. Thus, molecules that strengthen the synapse could be beneficial for the treatment of AD. Alteration in synaptic plasticity due to the accumulation of amyloid-β leading to memory loss and dementia is a key feature of AD. The methanolic extract of Avns was found to restore and alter long-term potentiation in CA3 to CA1 region of the hippocampus in the Tg2576 AD mouse brain [71]. In AD, amyloid-β aggregates were found to act through activating GSK3β of PI3K/AKT pathway [72,73]. The study revealed that Avn-C, in particular, modulates the S9-GSKβ leading to improvement in brain function altered due to amyloid-β accumulation. Inhibition of S9-GSKβ effectively upregulates Nrf2-ARE activity, which may be responsible for the neuroprotective action of Avn-C in Tg2576 mice [74]. In addition, it was also reported that Avn-C acts as an amyloid inhibitor thereby preventing Aβ protein aggregation [75].
The study showed that Avn-C improved the memory deficits associated with the Aβ and was reported to strengthen the synapse indicated by improvement in LTP. The study also revealed that Avn-C effectively reduced the caspase-3 concentration and thereby inhibited the activation of pro-inflammatory markers, and in contrast increased the levels of anti-inflammatory markers. In addition, it activates the S9GSK3β and improved the antioxidant defence through activation of Nrf2 in different model of AD. A possible role of Avn in modulation of PI3K/AKT/ Nrf2/ GSK-3β signalling pathway was shown in Figure 2.
4.2. Parkinson’s disease
PD is the second most common neurodegenerative condition, affecting 1% of the old age population. Clinical presentation in PD is associated with motor disturbances including bradykinesia, rigidity, and tremors [76]. Majorly, PD is characterized by progressive loss of dopaminergic neurons in SN, but the neuropathology indicates a widespread association between different regions of the brain where involvement of SN occurs in the middle stage of the disease. The pathological hallmark of the disease involves the deposition of Lewy bodies, which are round eosinophilic protein aggregates composed of α-synuclein (α-syn) and synphililin-1 [77,78]. A variety of factors are responsible for the alteration in transcription in the sporadic form of PD such as environmental factors, oxidative stress, exposures, and, aging [79,80], whereas the familial type of PD is associated with a mutation in genes encoding α-synuclein [81,82,83,84,85]. Studies have suggested that among the list of factors oxidative stress and ROS are mainly responsible for neuronal loss in the SN region in PD pathophysiology. Bisavenanthramide-B (Bis-B), a synthetic analog of Avn-C appears to protect from oxidative stress. The molecule has been discovered as a product formed during the reaction of Avn with ROS. Bis-B contains an electrophilic group designated as Michael acceptor which is capable of covalently conjugating with cysteine residues of Keap1, thereby inducing conformational changes leading to translocation of Nrf2 in the nucleus, which results in cytoprotective gene expression through Nrf2-ARE interaction [86].
The oxidative stress induced via dysfunctional mitochondria has been a well-known pathology in neurodegenerative diseases. These synthetic Avn has also shown a neuroprotective effect against rotenone/oligomycin-induced oxidative damage, which was considerably similar to PD pathology. Thus, Bis-Avns could be the effective therapeutic approach in preventing selectively SN dopaminergic degeneration in PD. The same study has revealed that Avn has excellent neuroprotective action through direct and indirectly scavenging ROS by Nrf2 stimulation against okadaic acid-induced Tau hyperphosphorylation and oxidative stress in SH-SY5Y cells, which has been linked critically with the neurodegenerative tauopathies in AD and PD [87,88]. In addition, Avn-C can directly protect from oxidative stress through the activation of Nrf2-ARE [89]. The Avn-2c, isoform efficiently translocation of Nrf2 into the nucleus in PC12 cells and thereby stimulates the expression of cytoprotective enzymes, thus could be beneficial in the treatment of neurodegenerative diseases. The available literature on the therapeutic potential of Avn and its derivatives was shown in Table 1.
5. Modulation of other targets by Avns
Phytochemicals are mainly associated with neuroprotective benefits against numerous risk factors including chemical toxicity and oxidative stress in neurodegenerative conditions. Khaled and Yasmin, 2016 indicated that oat extract enriched with Avns possesses a neuroprotective role against toxicity and oxidative stress induced by titanium dioxide nanoparticles (Tio2NPs) in rats. The biochemical and histopathological studies revealed that the combination of Avns with thymoquinone exerts antioxidant and anti-inflammatory action, thus preventing neuronal degeneration induced with Tio2NPs [96]. The Avns improved the deleterious effect by altering the oxidative stress and improving antioxidant concentration.
Numerous studies suggested that the levels of pro-inflammatory markers including TNF-α, IL-1β and IL-6 were significantly increased in cerebrospinal fluid of AD patients and were correlated with impairment in LTP in animal models of AD [97,98,99]. Thus, inhibition of the inflammatory pathway would be an effective approach to improve memory impairment. Ramasamy et al. reposted that treatment with Avn-C significantly improves memory impairment in Tg2576 and 5XFAD mice brains [20]. In addition, the study also reported that Avn-C reduced the level of inflammatory markers, which could be a potential mechanism for its action. Several studies also suggested that the levels of TNF-α were significantly reduced with Avn-c treatment [100,101,102].
In addition, Avns were found to reduce inflammation via altering TNF-α concentration. The Avns were found to limit the TNF-α induced neurogenic inflammation via modulating the NF-κB signalling and IL-8 levels [103,104]. Thus, the anti-inflammatory mechanism of Avn-c could be beneficial in the treatment of neurodegenerative conditions (Figure 3). The levels of the neurotransmitter acetylcholine are reduced due to hydrolysis due to overstimulation of acetylcholinesterase in AD patients. Thus targeting acetylcholinesterase could be beneficial in AD treatment [105]. Avn was also found to inhibit acetylcholinesterase and thus can improve memory in neurodegenerative conditions like AD [106,107].
6. Future perspectives
Oats are a useful grain with distinct constituents and a unique source of Avns possessing abundant nutritional benefits. Recently, focus has been given to the biological activities of naturally produced Avns as well as their derivatives in the treatment of NDDs. Avenanthramides' anti-inflammatory effects make them attractive candidates for mitigating neuroinflammation and its detrimental impact on the progression of these diseases. Future studies could focus on investigating their specific anti-inflammatory mechanisms and assessing their effectiveness in reducing neuroinflammation. Further, the antioxidant properties of Avns make them potential candidates for managing oxidative stress and preventing neuronal damage. Research could explore their potential in slowing or halting the progression of NDDs including AD and PD.
While Avns show promise, it is important to note that further research is necessary to fully understand their mechanisms of action, optimal dosages, and long-term effects in the context of NDDs. Further, the beneficial effects of Avns using synthetic Avn analogs and Avns obtained by recombinant techniques should also be explored in treatments of NDDs. For this purpose, collaboration between researchers and clinicians will be crucial in translating the potential of Avns into effective therapeutic interventions for NDDs.
7. Conclusions
The PI3K/AKT signalling pathway in NDDs has been proven to be a successful hypothesis for the development of an effective treatment approach and more detailed research is being conducted to understand the involvement of downstream targets for the treatment of NDDs. Avn holds potential as a treatment for NDDs caused by oxidative stress. The protective effects of Avns are mediated mainly through activation of the Nrf2, a downstream target of PI3K/AKT which upregulates transcription genes encoding antioxidant enzymes. Avn-C and its synthetic analog were also found to modulate other targets PI3K/AKT including GSKβ and NF-κB in CNS. The molecule also possesses anti-inflammatory and anti-apoptotic action in wide pathological states including cancer and skin inflammation indicating its mechanism could be beneficial for the treatment of NDDs. In conclusion, the need for in depth pre-clinical studies in various models of neurodegeneration and clinical exploration to elucidate the specific mechanisms of Avns action and its potential interactions with other signalling cascades involved in neurodegeneration is quite essential.
Author Contributions
N.L.W., M.B.K., A.K.B., M.M.A., S.R.K. and S.K. conceptualized, designed, and wrote the manuscript; N.L.W., M.B.K., S.K., B.G.T., R.V.T., M.J.U., A.K., P.W. and S.RK., revised the manuscript; N.L.W., M.B.K., S.K. and S.R.K. participated in drafting the article and revising it critically; S.K. and S.R.K. were involved in funding acquisition. All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This work was supported by Sejong university, South Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perrelli, A.; Goitre, L.; Salzano, A.M.; Moglia, A.; Scaloni, A.; Retta, S.F. Biological Activities, Health Benefits, and Therapeutic Properties of Avenanthramides: From Skin Protection to Prevention and Treatment of Cerebrovascular Diseases. Oxidative Medicine and Cellular Longevity 2018, 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Strychar, R.; Webster, F.H.; Wood, P.J. World Oat Production, Trade, and Usage. In Oats: Chemistry and Technology; 2011; pp. 77–94. [Google Scholar]
- Hoffmann, L.A. World Production and Use of Oats. In The Oat Crop Production and Utilization; 1995; pp. 34–61. [Google Scholar]
- Singh, R.; De, S.; Belkheir, A. Avena Sativa (Oat), A Potential Neutraceutical and Therapeutic Agent: An Overview. Critical Reviews in Food Science and Nutrition 2013, 53, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Gonzalez, T.; Ferruzzi, M.; Jackson, E.; Winderl, D.; Watson, J. Oats—from Farm to Fork. In Advances in food and nutrition research; 2016; Volume 77, pp. 1–55. [Google Scholar]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate Quality and Human Health: A Series of Systematic Reviews and Meta-Analyses. The Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.V.T.; Sievenpiper, J.L.; Zurbau, A.; Blanco Mejia, S.; Jovanovski, E.; Au-Yeung, F.; Jenkins, A.L.; Vuksan, V. The Effect of Oat β -Glucan on LDL-Cholesterol, Non-HDL-Cholesterol and ApoB for CVD Risk Reduction: A Systematic Review and Meta-Analysis of Randomised-Controlled Trials. British Journal of Nutrition 2016, 116, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate Quality and Human Health: A Series of Systematic Reviews and Meta-Analyses. The Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef]
- Maki, K.C.; Galant, R.; Samuel, P.; Tesser, J.; Witchger, M.S.; Ribaya-Mercado, J.D.; Blumberg, J.B.; Geohas, J. Effects of Consuming Foods Containing Oat β-Glucan on Blood Pressure, Carbohydrate Metabolism and Biomarkers of Oxidative Stress in Men and Women with Elevated Blood Pressure. European Journal of Clinical Nutrition 2007, 61, 786–795. [Google Scholar] [CrossRef]
- Ripsin, C.M.; Keenan, J.M.; Jacobs, D.R.; Elmer, P.J.; Welch, R.R.; Horn, L. Van; Liu, K.; Turnbull, W.H.; Thye, F.W.; Kestin, M.; et al. Oat Products and Lipid Lowering: A Meta-Analysis. JAMA 1992, 267, 3317–3325. [Google Scholar] [CrossRef]
- Sang, S.; Chu, Y. Whole Grain Oats, More than Just a Fiber: Role of Unique Phytochemicals. Molecular Nutrition & Food Research 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Singh, R.; De, S.; Belkheir, A. Avena Sativa (Oat), A Potential Neutraceutical and Therapeutic Agent: An Overview. Critical Reviews in Food Science and Nutrition 2013, 53, 126–144. [Google Scholar] [CrossRef]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef]
- Johansen, K.H.; Golec, D.P.; Thomsen, J.H.; Schwartzberg, P.L.; Okkenhaug, K. PI3K in T Cell Adhesion and Trafficking. Frontiers in Immunology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Bilanges, B.; Posor, Y.; Vanhaesebroeck, B. PI3K Isoforms in Cell Signalling and Vesicle Trafficking. Nature Reviews Molecular Cell Biology 2019, 20, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Long, H.Z.; Cheng, Y.; Zhou, Z.W.; Luo, H.Y.; Wen, D.D.; Gao, L.C. PI3K/AKT Signal Pathway: A Target of Natural Products in the Prevention and Treatment of Alzheimer’s Disease and Parkinson’s Disease. Frontiers in Pharmacology 2021, 12, 619. [Google Scholar] [CrossRef]
- Chen, Y.; Hsu, C.; Chen, X.; Zhang, H.; Peng, W. Editorial: Regulation of PI3K/Akt Signaling Pathway: A Feasible Approach for Natural Neuroprotective Agents to Treat Various Neuron Injury-Related Diseases. Frontiers in Pharmacology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Peng, S.; Song, Z.; Bai, F.; Li, X.; Fang, J. Oat Polyphenol Avenanthramide-2c Confers Protection from Oxidative Stress by Regulating the Nrf2-ARE Signaling Pathway in PC12 Cells. Archives of Biochemistry and Biophysics 2021, 706, 108857. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Kim, H.; Choi, J.-I.; Bae, H.-B.; Jeong, S. Avenanthramide C Prevents Neuronal Apoptosis via PI3K/Akt/GSK3β Signaling Pathway Following Middle Cerebral Artery Occlusion. Brain Sciences 2020, 10, 878. [Google Scholar] [CrossRef]
- Ramasamy, V.S.; Samidurai, M.; Park, H.J.; Wang, M.; Park, R.Y.; Yu, S.Y.; Kang, H.K.; Hong, S.; Choi, W.S.; Lee, Y.Y.; et al. Avenanthramide-C Restores Impaired Plasticity and Cognition in Alzheimer’s Disease Model Mice. Molecular Neurobiology 2020, 57, 315–330. [Google Scholar] [CrossRef]
- Lee, Y.-Y.; Wang, M.; Son, Y.; Yang, E.-J.; Kang, M.-S.; Kim, H.-J.; Kim, H.-S.; Jo, J. Oat Extract Avenanthramide-C Reverses Hippocampal Long-Term Potentiation Decline in Tg2576 Mice. Molecules 2021, 26, 6105. [Google Scholar] [CrossRef]
- Peterson, D.M.; Hahn, M.J.; Emmons, C.L. Oat Avenanthramides Exhibit Antioxidant Activities in Vitro. Food Chemistry 2002, 79, 473–478. [Google Scholar] [CrossRef]
- Helnæs, A.; Kyrø, C.; Andersen, I.; Lacoppidan, S.; Overvad, K.; Christensen, J.; Tjønneland, A.; Olsen, A. Intake of Whole Grains Is Associated with Lower Risk of Myocardial Infarction: The Danish Diet, Cancer and Health Cohort. The American Journal of Clinical Nutrition 2016, 103, 999–1007. [Google Scholar] [CrossRef]
- Sang, S.; Chu, Y. Whole Grain Oats, More than Just a Fiber: Role of Unique Phytochemicals. Molecular Nutrition & Food Research 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S. Sen; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotoxicity Research 2019, 35, 775–795. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Kiger, A.A. Classes of Phosphoinositide 3-Kinases at a Glance. Journal of Cell Science 2014, 127, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Taksande, B.G.; Gawande, D.Y.; Chopde, C.T.; Umekar, M.J.; Kotagale, N.R. Agmatine Ameliorates Adjuvant Induced Arthritis and Inflammatory Cachexia in Rats. Biomedicine and Pharmacotherapy 2017, 86, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Hanada, M.; Feng, J.; Hemmings, B.A. Structure, Regulation and Function of PKB/AKT—A Major Therapeutic Target. In Proceedings of the Biochimica et Biophysica Acta - Proteins and Proteomics; 2004; Volume 1697, pp. 3–16. [Google Scholar]
- Long, H.Z.; Cheng, Y.; Zhou, Z.W.; Luo, H.Y.; Wen, D.D.; Gao, L.C. PI3K/AKT Signal Pathway: A Target of Natural Products in the Prevention and Treatment of Alzheimer’s Disease and Parkinson’s Disease. Frontiers in Pharmacology 2021, 12, 619. [Google Scholar] [CrossRef]
- Matsuda, S.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.; Nakanishi, A.; Murai, T. Implications of PI3K/AKT/PTEN Signaling on Superoxide Dismutases Expression and in the Pathogenesis of Alzheimer’s Disease. Diseases 2018, 6, 28. [Google Scholar] [CrossRef]
- Kucuksayan, H.H.; Akgun, S.S. Pl3K/Akt/NF-ΚB Signalling Pathway on NSCLC Invasion. Medicinal chemistry 2016, 06. [Google Scholar] [CrossRef]
- Katsnelson, A.; De Strooper, B.; Zoghbi, H.Y. Neurodegeneration: From Cellular Concepts to Clinical Applications. Science Translational Medicine 2016, 8. [Google Scholar] [CrossRef]
- Peplow, P.V.; Martinez, B.; Gennarelli, T.A. Prevalence, Needs, Strategies, and Risk Factors for Neurodegenerative Diseases. Neuromethods 2022, 173, 3–8. [Google Scholar] [CrossRef]
- Jenner, P. Oxidative Stress in Parkinson’s Disease. Annals of Neurology 2003, 53, S26–S38. [Google Scholar] [CrossRef]
- Gan, L.; Johnson, J.A. Oxidative Damage and the Nrf2-ARE Pathway in Neurodegenerative Diseases. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2014, 1842, 1208–1218. [Google Scholar] [CrossRef]
- Andersen, J.K. Oxidative Stress in Neurodegeneration: Cause or Consequence? Nature Medicine 2004, 10, S18–S25. [Google Scholar] [CrossRef] [PubMed]
- Aslan, M.; Ozben, T. Reactive Oxygen and Nitrogen Species in Alzheimers Disease. Current Alzheimer Research 2004, 1, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Mimura, J.; Yamamoto, M. Discovery of the Negative Regulator of Nrf2, Keap1: A Historical Overview. Antioxidants & Redox Signaling 2010, 13, 1665–1678. [Google Scholar] [CrossRef]
- Suzuki, T.; Motohashi, H.; Yamamoto, M. Toward Clinical Application of the Keap1–Nrf2 Pathway. Trends in Pharmacological Sciences 2013, 34, 340–346. [Google Scholar] [CrossRef]
- Yamazaki, H.; Tanji, K.; Wakabayashi, K.; Matsuura, S.; Itoh, K. Role of the Keap1/Nrf2 Pathway in Neurodegenerative Diseases. Pathology International 2015, 65, 210–219. [Google Scholar] [CrossRef]
- Gan, L.; Johnson, J.A. Oxidative Damage and the Nrf2-ARE Pathway in Neurodegenerative Diseases. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2014, 1842, 1208–1218. [Google Scholar] [CrossRef]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 Is Controlled by Two Distinct β-TrCP Recognition Motifs in Its Neh6 Domain, One of Which Can Be Modulated by GSK-3 Activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef]
- Hayes, J.D.; Chowdhry, S.; Dinkova-Kostova, A.T.; Sutherland, C. Dual Regulation of Transcription Factor Nrf2 by Keap1 and by the Combined Actions of β-TrCP and GSK-3. Biochemical Society Transactions 2015, 43, 611–620. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Sternberg, P.; Cai, J. Essential Roles of the PI3 Kinase/Akt Pathway in Regulating Nrf2-Dependent Antioxidant Functions in the RPE. Investigative Opthalmology & Visual Science 2008, 49, 1671. [Google Scholar] [CrossRef]
- Armagan, G.; Sevgili, E.; Gürkan, F.T.; Köse, F.A.; Bilgiç, T.; Dagcı, T.; Saso, L. Regulation of the Nrf2 Pathway by Glycogen Synthase Kinase-3β in MPP+-Induced Cell Damage. Molecules 2019, 24, 1377. [Google Scholar] [CrossRef] [PubMed]
- Sotolongo, K.; Ghiso, J.; Rostagno, A. Nrf2 Activation through the PI3K/GSK-3 Axis Protects Neuronal Cells from Aβ-Mediated Oxidative and Metabolic Damage. Alzheimer’s Research & Therapy 2020, 12, 13. [Google Scholar] [CrossRef]
- Beurel, E.; Grieco, S.F.; Jope, R.S. Glycogen Synthase Kinase-3 (GSK3): Regulation, Actions, and Diseases. Pharmacology & Therapeutics 2015, 148, 114–131. [Google Scholar] [CrossRef]
- Singh, A.K.; Kashyap, M.P.; Tripathi, V.K.; Singh, S.; Garg, G.; Rizvi, S.I. Neuroprotection Through Rapamycin-Induced Activation of Autophagy and PI3K/Akt1/MTOR/CREB Signaling Against Amyloid-β-Induced Oxidative Stress, Synaptic/Neurotransmission Dysfunction, and Neurodegeneration in Adult Rats. Molecular Neurobiology 2017, 54, 5815–5828. [Google Scholar] [CrossRef]
- Luo, J. GSK3β in Ethanol Neurotoxicity. Molecular Neurobiology 2009, 40, 108–121. [Google Scholar] [CrossRef]
- Le Belle, J.E.; Orozco, N.M.; Paucar, A.A.; Saxe, J.P.; Mottahedeh, J.; Pyle, A.D.; Wu, H.; Kornblum, H.I. Proliferative Neural Stem Cells Have High Endogenous ROS Levels That Regulate Self-Renewal and Neurogenesis in a PI3K/Akt-Dependant Manner. Cell Stem Cell 2011, 8, 59–71. [Google Scholar] [CrossRef]
- Ali, T.; Kim, M.O. Melatonin Ameliorates Amyloid Beta-Induced Memory Deficits, Tau Hyperphosphorylation and Neurodegeneration via PI3/Akt/GSk3β Pathway in the Mouse Hippocampus. Journal of Pineal Research 2015, 59, 47–59. [Google Scholar] [CrossRef]
- Kitagishi, Y.; Nakanishi, A.; Ogura, Y.; Matsuda, S. Dietary Regulation of PI3K/AKT/GSK-3β Pathway in Alzheimer’s Disease. Alzheimer’s Research & Therapy 2014, 6, 35. [Google Scholar] [CrossRef]
- Ali, T.; Yoon, G.H.; Shah, S.A.; Lee, H.Y.; Kim, M.O. Osmotin Attenuates Amyloid Beta-Induced Memory Impairment, Tau Phosphorylation and Neurodegeneration in the Mouse Hippocampus. Scientific Reports 2015, 5, 11708. [Google Scholar] [CrossRef]
- Ali, T.; Kim, T.; Rehman, S.U.; Khan, M.S.; Amin, F.U.; Khan, M.; Ikram, M.; Kim, M.O. Natural Dietary Supplementation of Anthocyanins via PI3K/Akt/Nrf2/HO-1 Pathways Mitigate Oxidative Stress, Neurodegeneration, and Memory Impairment in a Mouse Model of Alzheimer’s Disease. Molecular Neurobiology 2018, 55, 6076–6093. [Google Scholar] [CrossRef]
- Erdogdu, Ö.; Nathanson, D.; Sjöholm, Å.; Nyström, T.; Zhang, Q. Exendin-4 Stimulates Proliferation of Human Coronary Artery Endothelial Cells through ENOS-, PKA- and PI3K/Akt-Dependent Pathways and Requires GLP-1 Receptor. Molecular and Cellular Endocrinology 2010, 325, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Silva-Palacios, A.; Ostolga-Chavarría, M.; Zazueta, C.; Königsberg, M. Nrf2: Molecular and Epigenetic Regulation during Aging. Ageing Research Reviews 2018, 47, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-C.; Vargas, M.R.; Pani, A.K.; Smeyne, R.J.; Johnson, D.A.; Kan, Y.W.; Johnson, J.A. Nrf2-Mediated Neuroprotection in the MPTP Mouse Model of Parkinson’s Disease: Critical Role for the Astrocyte. Proceedings of the National Academy of Sciences 2009, 106, 2933–2938. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, J.; Dong, M. Nrf2 as a Potential Target for Parkinson’s Disease Therapy. Journal of Molecular Medicine 2021, 99, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox Regulation by NRF2 in Aging and Disease. Free Radical Biology and Medicine 2019, 134, 702–707. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, L.; Zhang, Q.; Li, X.; Xia, X.-J.; Liu, Y.-Y.; Yang, Q.-S. Lentiviral Vector-Mediated SHC3 Silencing Exacerbates Oxidative Stress Injury in Nigral Dopamine Neurons by Regulating the PI3K-AKT-FoxO Signaling Pathway in Rats with Parkinson’s Disease. Cellular Physiology and Biochemistry 2018, 49, 971–984. [Google Scholar] [CrossRef]
- Jia, Y.; Mo, S.-J.; Feng, Q.-Q.; Zhan, M.-L.; OuYang, L.-S.; Chen, J.-C.; Ma, Y.-X.; Wu, J.-J.; Lei, W.-L. EPO-Dependent Activation of PI3K/Akt/FoxO3a Signalling Mediates Neuroprotection in In Vitro and In Vivo Models of Parkinson’s Disease. Journal of Molecular Neuroscience 2014, 53, 117–124. [Google Scholar] [CrossRef]
- Kabanda, M.M. A Theoretical Study of the Antioxidant Properties of Phenolic Acid Amides Investigated through the Radical-Scavenging and Metal Chelation Mechanisms. European Food Research and Technology 2015, 241, 553–572. [Google Scholar] [CrossRef]
- Ha, H.C.; Sirisoma, N.S.; Kuppusamy, P.; Zweier, J.L.; Woster, P.M.; Casero, R.A. The Natural Polyamine Spermine Functions Directly as a Free Radical Scavenger. Proceedings of the National Academy of Sciences 1998, 95, 11140–11145. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free Radical Biology and Medicine 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Palop, J.J.; Mucke, L. Amyloid-β–Induced Neuronal Dysfunction in Alzheimer’s Disease: From Synapses toward Neural Networks. Nature Neuroscience 2010, 13, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Uddin, Md.S.; Kabir, Md.T.; Tewari, D.; Mamun, A. Al; Mathew, B.; Aleya, L.; Barreto, G.E.; Bin-Jumah, M.N.; Abdel-Daim, M.M.; Ashraf, G.M. Revisiting the Role of Brain and Peripheral Aβ in the Pathogenesis of Alzheimer’s Disease. Journal of the Neurological Sciences 2020, 416, 116974. [Google Scholar] [CrossRef] [PubMed]
- Festa, G.; Mallamace, F.; Sancesario, G.M.; Corsaro, C.; Mallamace, D.; Fazio, E.; Arcidiacono, L.; Garcia Sakai, V.; Senesi, R.; Preziosi, E.; et al. Aggregation States of Aβ1–40, Aβ1–42 and Aβp3–42 Amyloid Beta Peptides: A SANS Study. International Journal of Molecular Sciences 2019, 20, 4126. [Google Scholar] [CrossRef] [PubMed]
- Mamun, A.; Uddin, M.; Mathew, B.; Ashraf, G. Toxic Tau: Structural Origins of Tau Aggregation in Alzheimer’s Disease. Neural Regeneration Research 2020, 15, 1417. [Google Scholar] [CrossRef]
- Fu, J.; Zhu, Y.; Yerke, A.; Wise, M.L.; Johnson, J.; Chu, Y.; Sang, S. Oat Avenanthramides Induce Heme Oxygenase-1 Expression via Nrf2-Mediated Signaling in HK-2 Cells. Molecular Nutrition & Food Research 2015, 59, 2471–2479. [Google Scholar] [CrossRef]
- Di Maso, M.J.; Nepomuceno, G.M.; St. Peter, M.A.; Gitre, H.H.; Martin, K.S.; Shaw, J.T. Synthesis of (±)-Bisavenanthramide B-6 by an Anionic Anhydride Mannich Reaction. Organic Letters 2016, 18, 1740–1743. [Google Scholar] [CrossRef]
- Lee, Y.; Wang, M.; Son, Y.; Yang, E.; Kang, M.; Kim, H.; Kim, H.; Jo, J. Oat Extract Avenanthramide-C Reverses Hippocampal Long-Term Potentiation Decline in Tg2576 Mice. Molecules 2021, 26, 6105. [Google Scholar] [CrossRef]
- Jo, J.; Whitcomb, D.J.; Olsen, K.M.; Kerrigan, T.L.; Lo, S.-C.; Bru-Mercier, G.; Dickinson, B.; Scullion, S.; Sheng, M.; Collingridge, G.; et al. Aβ1–42 Inhibition of LTP Is Mediated by a Signaling Pathway Involving Caspase-3, Akt1 and GSK-3β. Nature Neuroscience 2011, 14, 545–547. [Google Scholar] [CrossRef]
- Llorens-Martín, M.; Jurado, J.; Hernández, F.; Ávila, J. GSK-3β, a Pivotal Kinase in Alzheimer Disease. Frontiers in Molecular Neuroscience 2014, 7, 46. [Google Scholar] [CrossRef]
- Lee, Y.; Wang, M.; Son, Y.; Yang, E.; Kang, M.; Kim, H.; Kim, H.; Jo, J. Oat Extract Avenanthramide-C Reverses Hippocampal Long-Term Potentiation Decline in Tg2576 Mice. Molecules 2021, 26, 6105. [Google Scholar] [CrossRef]
- Quiroz Vazquez, M.G.; Montiel Condado, D.; Gonzalez Hernandez, B.; Gonzalez-Horta, A. Avenanthramide-C Prevents Amyloid Formation of Bovine Serum Albumin. Biophysical Chemistry 2020, 263, 106391. [Google Scholar] [CrossRef] [PubMed]
- Wirdefeldt, K.; Adami, H.-O.; Cole, P.; Trichopoulos, D.; Mandel, J.; Wirdefeldt, K.; Adami, H.-O.; Trichopoulos, Á.D.; Cole, P.; Mandel, J. Epidemiology and Etiology of Parkinson’s Disease: A Review of the Evidence. Eur J Epidemiol 2011, 26, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiology of Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. α-Synuclein in Filamentous Inclusions of Lewy Bodies from Parkinson’s Disease and Dementia with Lewy Bodies. Proceedings of the National Academy of Sciences 1998, 95, 6469–6473. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Kordower, J.H. Age-Associated Increases of α-Synuclein in Monkeys and Humans Are Associated with Nigrostriatal Dopamine Depletion: Is This the Target for Parkinson’s Disease? Neurobiology of Disease 2007, 25, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.-K.; Nodet, G.; Kim, H.-Y.; Jensen, M.R.; Bernado, P.; Fernandez, C.O.; Becker, S.; Blackledge, M.; Zweckstetter, M. Structural Characterization of α-Synuclein in an Aggregation Prone State. Protein Science 2009, 18, 1840–1846. [Google Scholar] [CrossRef]
- Maraganore, D.M.; Andrade, M. de; Elbaz, A.; Farrer, M.J.; Ioannidis, J.P.; Krüger, R.; Rocca, W.A.; Schneider, N.K.; Lesnick, T.G.; Lincoln, S.J.; et al. Collaborative Analysis of α-Synuclein Gene Promoter Variability and Parkinson Disease. JAMA 2006, 296, 661–670. [Google Scholar] [CrossRef]
- Chartier-Harlin, M.C.; Kachergus, J.; Roumier, C.; Mouroux, V.; Douay, X.; Lincoln, S.; Levecque, C.; Larvor, L.; Andrieux, J.; Hulihan, M.; et al. α-Synuclein Locus Duplication as a Cause of Familial Parkinson’s Disease. The Lancet 2004, 364, 1167–1169. [Google Scholar] [CrossRef]
- Singleton, A.B.; Farrer, M.; Johnson, J.; Singleton, A.; Hague, S.; Kachergus, J.; Hulihan, M.; Peuralinna, T.; Dutra, A.N.R.; Lincoln, S.; et al. [Alpha]-Synuclein Locus Triplication Causes Parkinson’s Disease. Science 2003, 302, 841–842. [Google Scholar] [CrossRef]
- Chu, Y.; Kordower, J.H. Age-Associated Increases of α-Synuclein in Monkeys and Humans Are Associated with Nigrostriatal Dopamine Depletion: Is This the Target for Parkinson’s Disease? Neurobiology of Disease 2007, 25, 134–149. [Google Scholar] [CrossRef]
- Cho, M.-K.; Nodet, G.; Kim, H.-Y.; Jensen, M.R.; Bernado, P.; Fernandez, C.O.; Becker, S.; Blackledge, M.; Zweckstetter, M. Structural Characterization of α-Synuclein in an Aggregation Prone State. Protein Science 2009, 18, 1840–1846. [Google Scholar] [CrossRef] [PubMed]
- Cores, Á.; Abril, S.; Michalska, P.; Duarte, P.; Olives, A.I.; Martín, M.A.; Villacampa, M.; León, R.; Carlos Menéndez, J. Bisavenathramide Analogues as Nrf2 Inductors and Neuroprotectors in in Vitro Models of Oxidative Stress and Hyperphosphorylation. Antioxidants 2021, 10, 941. [Google Scholar] [CrossRef] [PubMed]
- Xiong, N.; Xiong, J.; Jia, M.; Liu, L.; Zhang, X.; Chen, Z.; Huang, J.; Zhang, Z.; Hou, L.; Luo, Z.; et al. The Role of Autophagy in Parkinson’s Disease: Rotenone-Based Modeling. Behavioral and Brain Functions 2013, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Alavi Naini, S.M.; Soussi-Yanicostas, N. Tau Hyperphosphorylation and Oxidative Stress, a Critical Vicious Circle in Neurodegenerative Tauopathies? Oxidative Medicine and Cellular Longevity 2015, 2015, 1–17. [Google Scholar] [CrossRef]
- Hou, Y.; Peng, S.; Song, Z.; Bai, F.; Li, X.; Fang, J. Oat Polyphenol Avenanthramide-2c Confers Protection from Oxidative Stress by Regulating the Nrf2-ARE Signaling Pathway in PC12 Cells. Archives of Biochemistry and Biophysics 2021, 706, 108857. [Google Scholar] [CrossRef]
- Ramasamy, V.S.; Samidurai, M.; Park, H.J.; Wang, M.; Park, R.Y.; Yu, S.Y.; Kang, H.K.; Hong, S.; Choi, W.S.; Lee, Y.Y.; et al. Avenanthramide-C Restores Impaired Plasticity and Cognition in Alzheimer’s Disease Model Mice. Molecular Neurobiology 2020, 57, 315–330. [Google Scholar] [CrossRef]
- Quiroz Vazquez, M.G.; Montiel Condado, D.; Gonzalez Hernandez, B.; Gonzalez-Horta, A. Avenanthramide-C Prevents Amyloid Formation of Bovine Serum Albumin. Biophysical Chemistry 2020, 263, 106391. [Google Scholar] [CrossRef]
- Cores, Á.; Abril, S.; Michalska, P.; Duarte, P.; Olives, A.I.; Martín, M.A.; Villacampa, M.; León, R.; Carlos Menéndez, J. Bisavenathramide Analogues as Nrf2 Inductors and Neuroprotectors in in Vitro Models of Oxidative Stress and Hyperphosphorylation. Antioxidants 2021, 10, 941. [Google Scholar] [CrossRef]
- Hou, Y.; Peng, S.; Song, Z.; Bai, F.; Li, X.; Fang, J. Oat Polyphenol Avenanthramide-2c Confers Protection from Oxidative Stress by Regulating the Nrf2-ARE Signaling Pathway in PC12 Cells. Archives of Biochemistry and Biophysics 2021, 706, 108857. [Google Scholar] [CrossRef]
- Ma, Z.; Ma, Y.; Cao, X.; Zhang, Y.; Song, T. Avenanthramide-C Activates Nrf2/ARE Pathway and Inhibiting Ferroptosis Pathway to Improve Cognitive Dysfunction in Aging Rats. Neurochemical Research 2023, 48, 393–403. [Google Scholar] [CrossRef]
- Hassanein, K.M.A.; El-Amir, Y.O. Protective Effects of Thymoquinone and Avenanthramides on Titanium Dioxide Nanoparticles Induced Toxicity in Sprague-Dawley Rats. Pathology - Research and Practice 2017, 213, 13–22. [Google Scholar] [CrossRef]
- Hassanein, K.M.A.; El-Amir, Y.O. Protective Effects of Thymoquinone and Avenanthramides on Titanium Dioxide Nanoparticles Induced Toxicity in Sprague-Dawley Rats. Pathology - Research and Practice 2017, 213, 13–22. [Google Scholar] [CrossRef]
- Brosseron, F.; Krauthausen, M.; Kummer, M.; Heneka, M.T. Body Fluid Cytokine Levels in Mild Cognitive Impairment and Alzheimer’s Disease: A Comparative Overview. Molecular Neurobiology 2014, 50, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Swardfager, W.; Lanctôt, K.; Rothenburg, L.; Wong, A.; Cappell, J.; Herrmann, N. A Meta-Analysis of Cytokines in Alzheimer’s Disease. Biological Psychiatry 2010, 68, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Magaki, S.; Mueller, C.; Dickson, C.; Kirsch, W. Increased Production of Inflammatory Cytokines in Mild Cognitive Impairment. Experimental Gerontology 2007, 42, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.F.; Wise, M.L.; Gulvady, A.A.; Chang, T.; Kendra, D.F.; Jan-Willem Van Klinken, B.; Shi, Y.; O’Shea, M. In Vitro Antioxidant Capacity and Anti-Inflammatory Activity of Seven Common Oats. Food Chemistry 2013, 139, 426–431. [Google Scholar] [CrossRef]
- El Amir, Y.O.; Omar, W.; Khabrani, A.Y.; Jahfali, A.E.; Alhakami, S.M.; Dobab, N.M. Protective Effect of Avenanthramides against Cisplatin Induced Nephrotoxicity in Rats. Journal of advanced veterinary and animal research 2019, 6, 521–527. [Google Scholar] [CrossRef]
- Sur, R.; Nigam, A.; Grote, D.; Liebel, F.; Southall, M.D. Avenanthramides, Polyphenols from Oats, Exhibit Anti-Inflammatory and Anti-Itch Activity. Archives of Dermatological Research 2008, 300, 569–574. [Google Scholar] [CrossRef]
- Kang, C.; Shin, W.S.; Yeo, D.; Lim, W.; Zhang, T.; Ji, L.L. Anti-Inflammatory Effect of Avenanthramides via NF-ΚB Pathways in C2C12 Skeletal Muscle Cells. Free Radical Biology and Medicine 2018, 117, 30–36. [Google Scholar] [CrossRef]
- Sur, R.; Nigam, A.; Grote, D.; Liebel, F.; Southall, M.D. Avenanthramides, Polyphenols from Oats, Exhibit Anti-Inflammatory and Anti-Itch Activity. Archives of Dermatological Research 2008, 300, 569–574. [Google Scholar] [CrossRef]
- Taqui, R.; Debnath, M.; Ahmed, S.; Ghosh, A. Advances on Plant Extracts and Phytocompounds with Acetylcholinesterase Inhibition Activity for Possible Treatment of Alzheimer’s Disease. Phytomedicine Plus 2022, 2, 100184. [Google Scholar] [CrossRef]
- Yang, J.; Ou, B.; Wise, M.L.; Chu, Y. In Vitro Total Antioxidant Capacity and Anti-Inflammatory Activity of Three Common Oat-Derived Avenanthramides. Food Chemistry 2014, 160, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Cores, Á.; Abril, S.; Michalska, P.; Duarte, P.; Olives, A.I.; Martín, M.A.; Villacampa, M.; León, R.; Carlos Menéndez, J. Bisavenathramide Analogues as Nrf2 Inductors and Neuroprotectors in in Vitro Models of Oxidative Stress and Hyperphosphorylation. Antioxidants 2021, 10, 941. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of Avenanthramides.
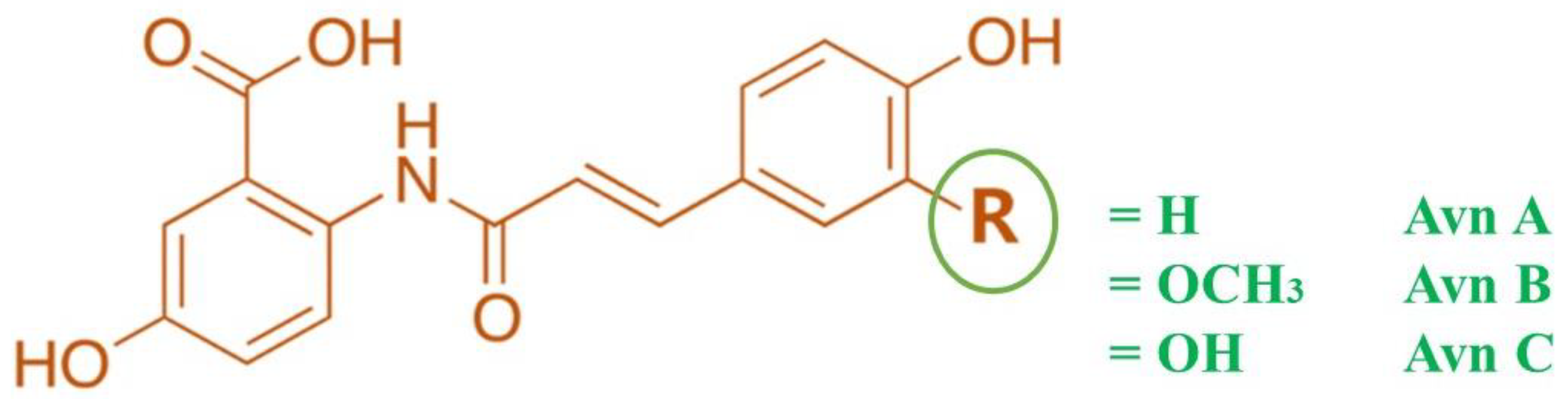
Figure 2.
Modulation of PI3K/AKT/ Nrf2/ GSK-3β signalling pathway by Avn. PI3K: Phosphatidylinositol-3-kinases (PI3Ks); AKT: Ak strain transforming-Phosphokinase B; GSK3β: Glycogen synthase kinase 3β; Nrf2: Nuclear factor erythroid 2-related factor 2; ARE: Antioxidant Response Element; Keap1: Kelchlike ECH-associated protein 1; RTK: Receptor tyrosine kinase. Oxidative stress limits the action of PI3K/AKT and thereby activates GSK3β thus inhibiting Nrf2 translocation into the nucleus, leading to downregulation of expression of cytoprotective enzymes. Avn-C acts as an electrophile that interacts with Keap, resulting in loss of inhibitory control and translocation of Nrf2 into the nucleus and activating ARE for transcription of the antioxidant enzyme. Avn-C can also activate Nrf2 indirectly by inhibiting GSK3β.
Figure 2.
Modulation of PI3K/AKT/ Nrf2/ GSK-3β signalling pathway by Avn. PI3K: Phosphatidylinositol-3-kinases (PI3Ks); AKT: Ak strain transforming-Phosphokinase B; GSK3β: Glycogen synthase kinase 3β; Nrf2: Nuclear factor erythroid 2-related factor 2; ARE: Antioxidant Response Element; Keap1: Kelchlike ECH-associated protein 1; RTK: Receptor tyrosine kinase. Oxidative stress limits the action of PI3K/AKT and thereby activates GSK3β thus inhibiting Nrf2 translocation into the nucleus, leading to downregulation of expression of cytoprotective enzymes. Avn-C acts as an electrophile that interacts with Keap, resulting in loss of inhibitory control and translocation of Nrf2 into the nucleus and activating ARE for transcription of the antioxidant enzyme. Avn-C can also activate Nrf2 indirectly by inhibiting GSK3β.
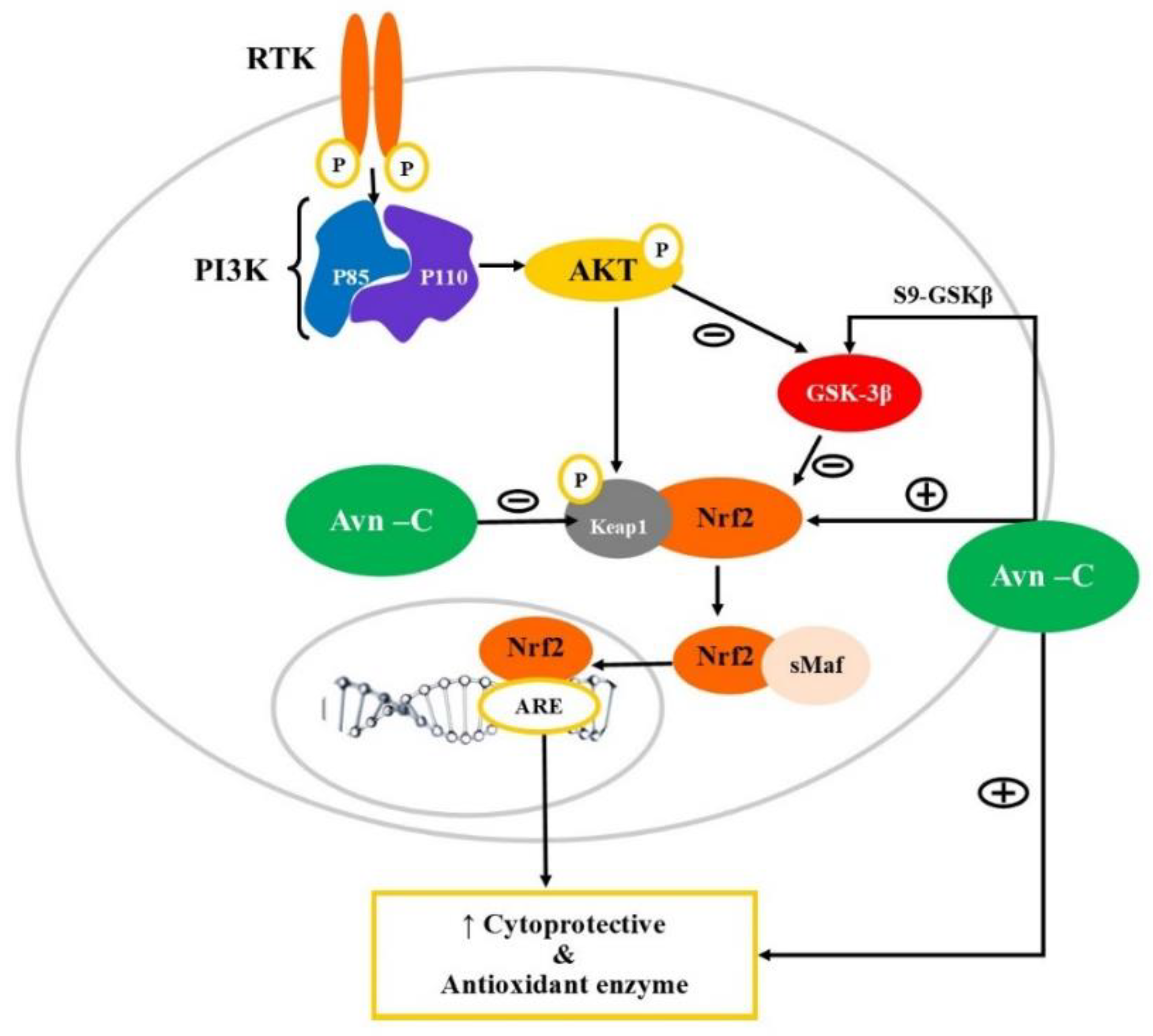
Figure 3.
Modulation of PI3K/AKT/NF-κB signalling pathway by Avn-c. PI3K: Phosphatidylinositol-3-kinases (PI3Ks); AKT: Ak strain transforming-Phosphokinase B; RTK: Receptor tyrosine kinase; NF-κB: Nuclear factor kappa B; IKK- Inhibitor of NF-κB kinase; IκB: inhibitor of nuclear factor-κB; TNFα: Tumor necrosis factor; IL-1β: Interleukin 1β; IL6: Interleukin 6. Avn-C downregulates the expression of inflammatory cytokines by inhibiting NF-κB and prevents its translocation into the nucleus thereby. Avn-C can directly inhibit the action of inflammatory mediators including TNFα, IL6, and IL1β.
Figure 3.
Modulation of PI3K/AKT/NF-κB signalling pathway by Avn-c. PI3K: Phosphatidylinositol-3-kinases (PI3Ks); AKT: Ak strain transforming-Phosphokinase B; RTK: Receptor tyrosine kinase; NF-κB: Nuclear factor kappa B; IKK- Inhibitor of NF-κB kinase; IκB: inhibitor of nuclear factor-κB; TNFα: Tumor necrosis factor; IL-1β: Interleukin 1β; IL6: Interleukin 6. Avn-C downregulates the expression of inflammatory cytokines by inhibiting NF-κB and prevents its translocation into the nucleus thereby. Avn-C can directly inhibit the action of inflammatory mediators including TNFα, IL6, and IL1β.
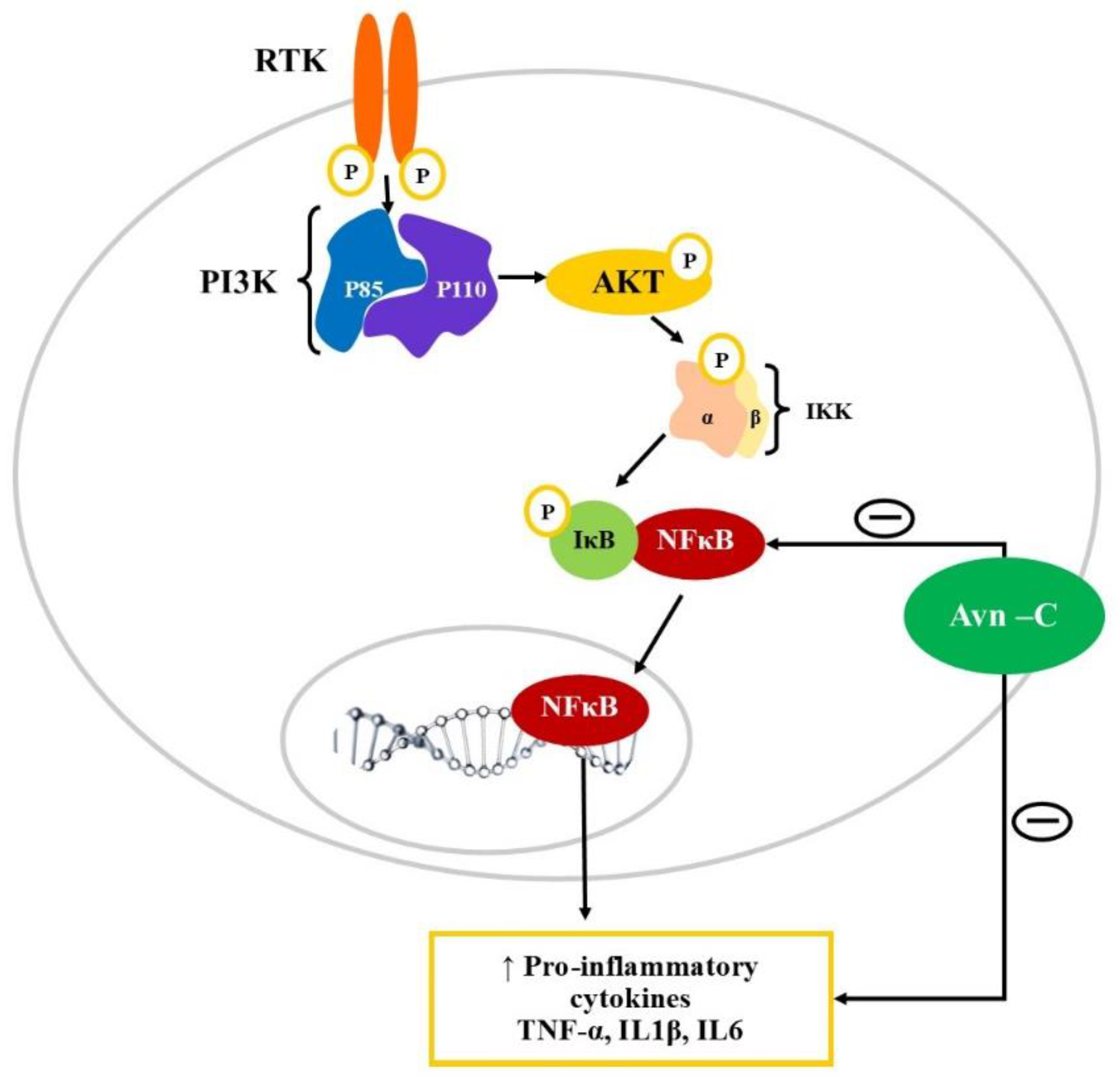

Table 1.
Therapeutic potential of Avenanthramide in Neurodegenerative disease.
| Sr. No. | Study design | Mechanism | Methodology | Reference |
|---|---|---|---|---|
| 1 | Electrophysiological study on Hippocampal LTP in Tg2576 male mice | Alteration in p-GSK-3β-S9 levels and reduction in Caspase 3 | Avn-C extracted from germinated oats using column chromatography | [74] |
| 2 | Evaluation of neuro-pathologies and behavioural impairments associated with AD | Modulates p-GSK-3β- S9 Reduced caspase 3 and neuroinflammation Binds to α1A adrenergic receptors to stimulate phospho-AMPK levels |
For electrophysiological studies the hippocampal slices of WT, Tg2576 and 5XFAD mice were treated with Aβ42 oligomers in the presence of Avn-C (10, 25, and 50 μM) | [90] |
| Tg2576 and 5XFAD mice were administered with Avn-C (2, 4, and 6 mg/kg. p.o.) for evaluation of long-term potentiation | ||||
| 3 | Effect on protein aggregation using spectroscopy techniques | Inhibition of BSA oligomerization showing anti-amyloid effect | Protein aggregation in bovine serum albumin was initiated by incubating the protein monomer at an elevated temperature and the aggregation kinetics was monitored by incubating with Avn-C (100, 250 and 500 μM) and was analysed using ThT-fluorescence assay. |
[91] |
| 4 | Evaluation of Bisavenanthramide analogues Nrf2 inductors and neuroprotectors in in-vitro models | Nrf2-ARE dependent protein expression | The antioxidant activity was measured using DPPH scavenging assay, FRAP Method and AChE inhibition assay was performed. Neuroprotection potential against tau hyperphosphorylation was evaluated using SH-SY5Y cell line | [92] |
| 5 | Evaluation of cytoprotective activity against oxidative stress-induced PC12 cell injuries | Activating Nrf2-ARE pathway | Rat PC12 were used to study antioxidant effect of Avns. The antioxidant and cytoprotective activity of Avn-2c, Avn-2f and Avn-2p was measured in-vitro using the ABTS•+ and DPPH scavenging assay, MTT, LDH release assay. | [93] |
| 6 | Cognitive dysfunction induced by repeated propofol anaesthesia in aging rats | Activating Nrf2/ARE pathway | Aging rat model was established by continuous 200 mg/kg propofol anaesthesia | [94] |
| 7 | Protective effect on titanium dioxide nanoparticles induced neurotoxicity in SD rats | Decreases oxidative stress and TNF-α Increases the total antioxidant and GSH levels |
TiO2 NPs (150 mg/kg b.w.) was administered orally for six weeks and Avn was administered daily at a dose of 20 mg/kg by gastric tube. | [95] |
Abbreviations: Avns, Avenanthramides; LTP, Long-term potentiation; AMPK, AMP-activated protein kinase; GSK-3β, Glycogen Synthase Kinase 3 Beta; TNF-α, Tumour Necrosis Factor alpha; WT, Wild type; SD, Sprague-Dawley; GSH, Glutathione; Nrf2/ARE, NF-E2-related factor 2/ antioxidant response element; TiO2 NPs, Titanium dioxide nanoparticles; BSA, Bovine serum albumin; PC12, pheochromocytoma cells; DPPH, 2,2-diphenylpicrylhydrazyl; MTT, 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide; LDH, L actate dehydrogenase; SH-SY5Y, neuroblastoma cell line; ABTS, 2,2'-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid; AChE, Acetylcholinesterase; FRAP, Ferric Reducing Antioxidant Power; ThT, thioflavin T fluorescence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



