Submitted:
01 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
Antimicrobial peptides provide a powerful toolkit for combating multidrug resistance. Combatting eukaryotic pathogens is complicated by the fact, the intracellular drug targets in the eukaryotic pathogen are frequently homologs of cellular structures of vital importance in the host organism. The entomopathogenic bacteria, (symbionts of entomopathogenic nematode species), release a series of non-ribosomal templated antimicrobial peptides, some may be potential drug candidates. The ability of an entomopathogenic nematode /entomopathogenic bacterium symbiotic complex to survive in a given polyxenic milieu is a coevolutionary product, explaining that those gene complexes which are responsible for the biosynthesis of different non-ribosomal templated antimicrobial protective peptides (including those which are potently capable to inactivate the protist mammalian -pathogen Leishmania and the gallinaceous bird pathogen Histomonas) are co-regulated. Our approach is based on comparative antimicrobial bioassays of the culture media of the wild-type and regulatory mutant strains. We concluded that Xenorhabdus budapestensis and X. szentirmaii are excellent sources of non-ribosomal templated antimicrobial peptides, that are efficient antagonists of the mentioned pathogens. Data on selective cytotoxicity of different cell-free culture media encourages us to forecast that the recently discovered “easy-PACId” research strategy is suitable for constructing EPB strains producing and releasing single, harmless, non-ribosomal templated antimicrobial peptides with considerable drug, (probiotic) -candidate potential.
Keywords:
non-ribosomal antimicrobial peptides (NR-AMP)
; entomopathogenic nematode-symbiont bacteria (EPB)
; Xenorhabdus
; (X. budapestensis
; X. szentirmaii
; X. innexii)
; Photorhabdus (P. luminescens TT01)
; CFCM
; (cellfree conditioned culture media)
; intracellular mammalian parasite
; Leishmania
; extracellular avian pathogens
; Histomonas
; probiotic potential
1. Introduction
Multidrug resistance (MDR) is a great, and rapidly increasing challenge for scientists, in latest 8 decades. The different forms of MDR appearing in pathogen organisms of clinical and veterinary, (as well as of agricultural) importance partly due to the consequence of the astonishingly unprofessional applications in inaccurate doses providing optimal selective conditions and selective advantages for MDR organisms to original (wild-type) ones of the same species, [1] (Fodor et al., 2020). One attempt to overcome resistance problems is to test the general and specific antimicrobial potential to apply antimicrobial peptides (AMPs) [2] (Ötvös and Wade, 2014). AMPs are of great potential to combat MDR prokaryotes because of several reasons. First, there is an option of taking advantage from the option of using the remarkable toolkit called quantitative structure-activity relation (QSAR) [3] (Karelson, Lobanov, Katriczki) 1996); [4] (Gini, 2022) technology in designing novel antimicrobial-active sister molecules of a newly discovered natural drug-candidate AMP molecule. The drug designer has a powerful way to produce efficient, or even-more powerful (but physiologically less harmful analogs of AMPs which may overcome resistance problems in such a way. Furthermore, according to recent data, antibiotic-resistant bacteria tend to perform collateral sensitivity to AMPs, [5] (Lázár et al., 2018), and the mobility patterns of traditional AMP-resistance genes are different from that of the antibiotics-resistance genes, [6] (Kintses et al., 2019), [1] (Fodor et al., 2020).
We are interested in testing the antipathogenic potential of AMPs in controlling MDR eukaryotic pathogens. As for the two selected targets: they are Leishmania. donovani [7] (Chaoulitch, 1954); a mammalian pathogen, is transmittable by insects like cave-dwelling sand flies [8] (Dutra-Rêgo et al., 2022), and Histomonas. meleagridis is the causative factor (or etiological agent) of the life-threatening histomoniasis (or blackhead) disease) of birds (turkeys, chicken pheasants, first described in 1893 [9] (McDougald, 1993), [10] (McDougald, 2005), and have been studied in detail in the laboratory of [11] (Hess et al., 2015).
When planning experiments aiming at applying AMPs against eukaryotic pathogens, we have to take into consideration that the strategy of designing antimicrobials to control eukaryotic parasites (pathogens) must be different from that of designing antimicrobials to control prokaryotic pathogens, because targettable intracellular structures of a eukaryotic parasite (pathogen) biochemically might be similar to some intracellular structure of vital function in the eukaryote host to be protected.
We aim to learn whether any of those biosynthetic NR_AMPs (like fabclavine, [12], (Fuchs et al., 2012), [13] (Fuchs et al., 2014), phenazine, ˙[14] (Shi et al., 2019, or something else), produced by those entomopathogenic-nematode symbiont bacteria (EPB) we have been working on [15] Lengyel et al., et al., 2005), [16] (Fodor et al., 2022), were suitable for applying anti-protist drug candidates either against the intracellular parasite Leishmania, or the extracellular parasite Histononas. The arguments for choosing in vitro EPB liquid cultures as appropriate sources of efficient AMPs for this purpose are as follows: Obligate bacterial symbionts of entomopathogenic nematode species synthesize and release several non-ribosomal hybrid peptides [17] (Akhurst, 1982), of large target specificities which mainly serve to provide well-balanced pathobiome conditions for this symbiosis in polyxenic soil,- and cadaver environments [18] (Ogier, et al., 2020). These peptides are also considered potential sources of potent natural anti-microbial compounds, [17] (Akhurst, 1982). The motivation for choosing in vitro EPB liquid cultures as a source of efficient AMPs for this purpose are as follows: proven active on Gram-positive [19 (Furgani), [20] (Fodor et al., 2018), Gram-negative [22] (Böszörményi, et al., 2009) pathogenic bacteria, and oomycetes and different other plant pathogens, [23], (Fodor et al., 2010), [24] (Vozik et a., 2015) in our previous studies. We have reasons to believe that some of the autoclaved Xenohabdus cultures may not be harmful or toxic for toxic when added as a food supplement while some antimicrobial ingredients are still active, [21] (Fodor et al., 2023), we may not give up the option of considering potential probiotic applications, at least at experimental level.
As for the eukaryotic targets used in this study, the genus Leishmania belongs to the Order Trypanosomatida, the Class Kinetoplastea, and the Phylum: Euglenozoa, [25] (Fernández-Arévalo et al., 2020), and is considered a blood parasite. It is the causative agent of leishmaniasis in humans, [26] (McGwire and Satoskar, 2014), and other mammals like dogs, [27] (Alvar et al., 2004); [28] (Dantas-Torres, et al., 2019].
The Leishmania pathogens act as intracellular parasites [29] (Walker et al., 2014). They retained the ability to produce extracellular vesicles. Both human,- and canine (caused by Leishmania infantum) can appear in a severe form called visceral leishmaniasis (VL), or kala-azar, [30] (Burza et al., 2018), [31] (Safavi et al, 2021). It has become to-endemic diseases have become global due to human migration, climate change, and anthropogenic disturbance, causing significant worldwide health and economic burden [32] (Altamura et al., 2022). As for the existing defense strategies: despite promising research activities, no clinically usable vaccine is available. In the absence of an effective vaccine, [33] (Mauël, 2002), the control of leishmaniasis relies exclusively on chemotherapy, [34] (Ibarra-Meneses et al., 2022). Each used clinical chemotherapy is based on very few chemotherapeutic agents, mainly antimonial derivatives (Sb). [35] Douanne et al, 2020]. As for available chemotherapies: Amphotericin B (AmB), a polyene macrolide antibiotic derived from actinomycetes, [36] (Golenser and Domb, 2006) has routinely been used in cases of visceral and canine leishmaniasis It had been selected from 200 known polyene agents, because of its moderate toxicity. It has been permitted for intravenous administration, [36] (Golenser and Domb, 2006). AmB acts in a membrane-destroying manner, [37] (Ellis, 2004), and has not provoked clinical resistance up to 1956, [36] (Golenser, and Domb, 2006). Since then severe drug resistance to both AmB and the other routinely used drug, miltefosine, (especially in L. infantum strains), appeared and continues spreading at alarming rates [34] (Ibarra-Meneses et al., 2022). Leishmania has always become an exceptional t model for studying microbial virulence, [38 [Chang, 2003). To outline the fascinating resistance mechanisms developed by Leishmania would require a long detailed review, so we want to refer to some publications instead ([35] Douanne et al, 2020], [20] (Golenser and Domb, 2006; [34] (Ibarra-Meneses et al., 2022). [39] (Segovia, 1994),
The other chosen target, H. meleagridis belongs to the genus Histomonas, a member of the family Monocercomonadidae within the class Trichomonada, in the class Trichomonadida of Phylum Metamonda. Infections in turkeys may cause nearly 100% mortality. Outbreaks in chickens are more often marked by morbidity and subsequent recovery. The outbreaks in broiler breeder pullets at that time caused excessive losses from mortality (5 to 20%) culling, and overall poor flock performance. Histomonas parasites are carried by the eggs of the cecal worm Heterakis gallinarum enabling them to survive for long periods in the soil as a source of infection, [9] (McDougald, 1998), [10]] (McDougald, 1998), [11] (Hess et al., 2015). In the U.S. there were never any drug products available for the treatment of blackhead disease, [9] (McDougald, 1998), [10]] (McDougald, 1998), [11] (Hess et al., 2015). As for molecular taxonomy, and identification, the 5.8S, ITS-1, and ITS-2 rRNA regions were first sequenced [40] (Lollis et al. 2011). The most preferable remarkable reference gene to investigate gene expression as a symptom of the infection in selected organs from turkeys and chickens (called 60S ribosomal protein L13) has been identified [41] (Mitra et al., 2016), and used different infection models related to immunization, and vaccination, [42,43] (Kidane et al, 2016; (Mitra et al., 2018). The complete, annotated Histomonas reference genome sequence has been available, [44] (Palmieri et al., 2021). Histomoniasis is a fastidious disease in turkeys, with pathological lesions in the caeca and liver, sometimes with high mortality. In chickens, the disease is less fatal, and lesions are often confined to the caeca. In Europe, the disease was well controlled by applying nitroimidazoles and nitrofurans for therapy or prophylaxis, [11] (Hess et al., 2015). It infects a wide range of birds including chickens, turkeys, peafowl, quail, and pheasants, causing infectious enterohepatic lesions. Since their introduction into the market in the middle of the previous century, further research nearly ceased as outbreaks of histomoniasis occurred only very rarely.
With the ban of these drugs in the last two decades in North America, the European Union, and elsewhere, in combination with the changes in animal husbandry, the disease re-emerged, [11] (Hess et al., 2015). But at that time the previously applied drugs were banned. Consequently, research programs were set up in various places focusing on different features of the parasite and the disease. For the first time studies were performed to elucidate the molecular repertoire of the parasite. In addition, research has been started to investigate the parasite's interaction with its host, [11] (Hess et al., 2015). New diagnostic methods and tools were developed and tested with samples obtained from field outbreaks or experimental infections. Some of these studies aimed to clarify the introduction of the protozoan parasite into a flock and the transmission between birds, [45] (Hauck et al., 2010). Finally, a strong focus was placed on research concentrated on the development of new treatments and prophylactic strategies, urgently needed to combat the disease, [11] (Hess et al., 2015).
As for vaccination, studies on the immune response against histomoniasis focus mainly on different traits of the adaptive immune system. Activation of toll-like receptors (TLR) leads to the interplay between cells of innate and adaptive immunity with consequences on B and T cell clonal expansion. Efforts for successful vaccinations are on the way for years, [46] (Lagler et al., 2021), but various challenges need to be addressed until the first ever developed vaccine based upon live flagellates in human or bird medicine can be marketed. Summarizing: the poultry industry works without approved prophylactics, therapeutics, or vaccines to combat histomonosis, [47] (Beer et al., 2022).
Meanwhile, numerous chemical compounds were also tested for their efficacy against H. meleagridis with varying outcomes. One of the explanations for these half-failures / half-successes may be the absence of reliable and reproducible in vitro Histomonas culturing techniques. Escherichia coli strongly supports the growth of H. meleagridis in a monoxenic culture, without influencing its pathogenicity, [48] (Ganas et al., 2012). Based on clonal cultures of H. meleagridis, monoxenic cultures have, to our knowledge for the first time, been established in a liquid medium. The fecal flora was exchanged for defined bacterial strains by selective destruction of the initial bacteria with a variety of antibiotics, keeping the flagellate alive. The growth of the protozoan parasite was found to depend on the bacteria, especially on their energy metabolism. E. coli was found to strongly support the growth of the parasite, whereas Salmonella enterica serovar Typhimurium and Pseudomonas aeruginosa were less efficient. Confocal laser microscopy showed that H. meleagridis could take up green fluorescent protein-tagged E. coli DH5α, suggesting that bacteria served as a food supply for the protozoa. By exchanging the bacterial flora for E. coli strain DH5α in H. meleagridis cultures that underwent continuous in vitro passages, it was possible to show that the in vivo attenuation process was independent of the bacteria. Furthermore, the gut flora in infected turkeys had no negative effect on the protozoan's virulence. Consequently, attenuation depends not on the bacteria in the culture but on the in vitro passages. Finally, the experiments provided evidence that the infection of turkeys with H. meleagridis enabled infection of the liver with E. coli, [48] (Ganas et al., 2012)..
In the following study identical monoxenic settings for cultures of the same H. meleagridis clonal strain in its virulent low passage and attenuated high passage form enabled a comparative analysis of parasite characteristics. For the first time, it could be shown that long-term in vitro cultivation led to a severe shift in cell morphology, with the occurrence of a very distinct phenotype expressing a flagellated and highly amoebic cell morphology. Furthermore, the attenuated parasites showed better growth rates and a higher tenacity when confronted with adverse conditions. During these experiments up to 100% of the parasites, both virulent and attenuated, assumed a completely rounded morphology elucidated by electron microscopy. The findings indicated that such previously reported cyst-like stages were a defense strategy of H. meleagridis, independent of the passage level in vitro and pathogenicity in vivo. In conclusion, long-term in vitro passaging of H. meleagridis led not only to an attenuation of the parasite, as previously demonstrated but also to a shift in the parasite's phenotype regarding morphology, growth behavior, and a higher level of tenacity, [49] (Gruber et al., 2017).
The option of the monogenic culturing technique opens the door to, preliminary attempts to protect birds with cultured attenuated histomonads forecasting the possibility of vaccination, (50] (Liebhart et al., 2017). Whether the special feature of the parasite's intricate interplay with bacteria in vitro and in vivo, can be considered mutualistic or a predator-prey one is a philosophical question, [51] (Bilic, and Hess, 2020). As for drug resistance, the first documented example of drug-resistant H. meleagridis appeared in the literature, and the antimicrobial potential of alkaloids (harmaline and harmalol) in crude extracts of the medical plant Peganum harmala, was tested against it, ˙[52] (Arshad et al., 2008).
After banning all previously used prophylactic and therapeutic drugs against H. meleagridis in the European Union, some benzimidazoles were only antiparasitic drugs, licensed for use in food-producing animals. [33] (Hauck and, Hafez, 2009). Benzimidazoles act on beta-tubulin, and the beta-tubulin sequence allows predictions about the efficacy of benzimidazoles. When sequenced and analyzed a part of the beta-tubulin gene of five H. meleagridis strains was found. all histomonal amino acid sequences predicted susceptibility to benzimidazoles. However, when the efficacy of five benzimidazoles, (albendazole, fenbendazole, flubendazole, mebendazole, and nocodazole), were tested on the 5 H. meleagridis strains in vitro, all tested drugs showed no efficacy, [38] (Hauck and, Hafez, 2009). Reduced sensitivity of H. meleagridis to nitarsone in vitro and in vivo. [54] [Abraham et al., 2014). Parasites are highly prevalent in poultry; thus, the management of parasites is a key component in the profitable production of poultry. The most common nematode parasite of poultry, Heterakis gallinarum, typically causes no direct pathology but is the vector of Histomonas meleagridis, a highly pathogenic protozoan parasite that causes blackhead disease. There are no approved treatments for H. meleagridis, making control reliant on controlling the helminth vector, [55] (Collins et al., 2022).
Herein we compared the spectrum of in vitro antimicrobial activity of cell-free conditioned culture media (CFCM) from three EPB species together with testing against Leishmania donovanii, Trypanosoma cruzi, Histomonas meleagridis, and as well as against panel of clinical bacterial and fungal isolates, as positive controls. Considering the roles of the bacterial symbiont in the EPN/EPB symbiosis, we suppose that ani-protozoal compounds must be coexisting with other antimicrobial compounds in the CFCM of EPBs. Therefore for reliable bioassays, we should use appropriate controls. Two main aspects ought to be taken into consideration: (A) To be able to distinguish between the specified antiprotist activity and general cytotoxicity of the future drug-candidate antimicrobial compounds in the CFCM. (B) To be able to identify the special anti-protist compounds. In this paper, we described the very first steps toward d these goals
As for (A), one has to take into consideration that the lack of uniformity regarding the choice of cell types for cytotoxicity assays may lead to incomparable and inconclusive data. In vitro assays relying solely on non-phagocytic cell models may not represent a realistic result as the effect of an antileishmanial agent should ideally be presented based on its cytotoxicity profile against reticuloendothelial system cells. [56] (Brioschi et al., 2022). In the Leishmania studies, we used macrophage cell line J774A.1 [57] (Tominaga et al., 1998), [58] (Andreu et al., 2017), [59]. (Geroldinger et al., 2019), as cytopathogenecity control test organism, (see Materials and Methods).
In the Histomonas studies, we used permanent chicken liver cells (LMH (leghorn male hepatoma, LMH) cell line [60] (Kawaguchi et al., 1987), [61] (Amin et al., 2012), [21] (Fodor et al., 2023), as cytopathogenecity control test-organism, (see Materials and Methods).
As for (B), one has to take into consideration the following points: each NR_AMP-producing EPB cell is in the so-called primary (10) phase, or Phase 1, [62] (Forst and Nealson, 1996). In other terms, the phenotypic precondition for the antibiotic production (as well as for the capability of symbiosis) of the EPB species is the primary phase, [63] (Akhurst, 1980), [64] (Akhurst, 1982), [65]( Boemare and Akhurst., 1994). From the aspect of NR_AMP production, it means, that in a liquid culture of the primary (10) cells (of each of the studied EPB species, strain, isolate) more than one biosynthetic AMPs are present at the same time. Each expresses antimicrobial (maybe antiprotist) activity, so the antimicrobial (antiprotist) activity of a CFCM sample represents a cumulative activity. But in an optimal test system, one would prefer to determine the antimicrobial activity of each of the single antimicrobial compounds one by one.
Since the discovery that the activity (that is, the „switch-on” / „switch-off” states) of those group genes (operons, biosynthetic gene complexes, BGCs) which are responsible for the primary- secondary phenotypic phase shift in of each known EPB strain, not only work under the control of the versatile regulator hexA, [66] (Joyce and Clarke, 2003), [67] (Langer et al., 2017) but also are coordinately regulated at a higher level (via hfq-gene controlled sRNS/ HexA-mRNA base pairing, more precizely, the Hfq-dependent sRNA, ArcZ, directly base-pairs with the HexA-encoding mRNA, [68] (Tobias et al., 2016), [69]´(Tobias et al., 2017), [70] (Neubacher, 2020)) it is possible to construct double-mutant strains from each EPB species (or isolate) following the genuine strategy called easyPACId (easy Promoter Activated Compound Identification) approach, [71] (Bode et al., 2019), each of which biosynthesizes and releases only one single NR_AMP molecule into the culture media.
This discovery may be a starting point of the renaissance of Xenorabdus antimicrobial peptide research, [72] (Wenski et al., 2019), [73] (Wenski et al., 2020), [74] (Cimen et al., [75] 2022), (Gulsen et al., 2022).
We also managed to reconstruct the “easyPACId” hfq deletion mutant strains from EMA and EMC (Boros et al., in preparation) and publish here some information ab including data about their antimicrobial potential). In this paper, we give an account of the detail of constructing our hfq-del mutants from our EMA and EMC [16] (Fodor et al., 2022) strains, and provide phenotypic descriptions of them. Our easyPACId strains will be available for cooperation with fellow scientists. all over the world.
2. Materials and Methods
As for “living materials”, in this study plasmids, prokaryotic, and eukaryotic cellular organisms were used. Plasmids used in this study [69] (Tobias et a., 2017) , [76,77,78,79,80,81,82,83,84] (see Table A1 in Appendix A) served as molecular tools, so did some bacterium strains. Some prokaryotes (EPB cells) as antimicrobial producers. Others, like Gram-positive, and Gram-negative pathogenic clinical isolates, served as test organisms, and so did those eukaryotes (protists, fungi, and tissue culture cells) we mention.. Plasmids with a temperature-sensitive pSC101 replication system were growth at 30°C with antibiotic selection applied while curing these plasmids was carried out by growth at 30°C without antibiotic selection, followed by testing the resistance pattern of individual colonies for selecting plasmid-cured clones, [76] (Cherepanov and Wackernagel, 1995).. For cloning purposes, E. coli TG1 [77] (Gibson, 1984) was used as the host strain. For plasmid mobilization into Xenorhabdus, E. coli S17-1λ pir [78] (Simon, Priefer, & Pühler, 1983a) was applied. E. coli strains were routinely grown in LB at 37°C with appropriate antibiotic selection.
2.1. NR-AMP Producing EPB Strains, Test Organisms. Prepatatno and Biassays of CFCMs
2.1.1.The anti-intracellular-parasite protist activities of both the native (that is: centrifuged, and sterile-filtered (through 0.22 µm filter), and heat-sterilized (that is centrifuged, sterile-filtered (through 0.22 µm filter), and then (autoclaved) CFCMs of liquid cultures of 3 antimicrobial-producing EPB strains, EMA, EMC and EMK, representing 3 species, Xenorhabdus budapestensis nov. DSM16342(T) [15] (Lengyel et al., 2005), Xenorhabdus szentirmaii [15] (Lengyel et al., 2005)), and X. innexii nov. DSM16342(T) [15] ( Lengyel et al., 2005), [15] (Lengyel, et al., 2005). [16] (Fodor, at al, 2022), were bioassayed on Leishmania donovani and Trypanosoma cruzi human pathogenic clinical isolates, as well on expectedly antibiotic-susceptible (indicator control ) organisms, such as clinical isolates of Gram-positive (methicillin-sensitive (MSSA), and -resistant (MRSA) Staphylococcus aureus, and on Gram-negative (Francisella novicida, Escherichia coli, Salmonella typhimurium, and Pseudomonas aeruginosa) bacteria, and the commensal pathogenic fission yeast, Candida albicans,).The J744 murine macrophage cells served, [59] (Geroldinger et al., 2019), as a control for measuring the general eukaryotic cytopathogenicity of the CFCMs.
2.1.2 The anti-extracellular-parasite protist activities of both the native (that is: centrifuged, and sterile-filtered (through 0.22 µm filter), and heat-sterilized (that is centrifuged, sterile-filtered (through 0.22 µm filter), and then (autoclaved) CFCMs of liquid cultures of 2 antimicrobial-producing EPB strains, Pl.-„Yellow” and Pl.-„RED”), (both grown in Medium 199 containing Earle’s Salts, L-glutamine, 25 mM HEPES, L-amino acids, 15% heat-inactivated fetal bovine serum FBS and 0.22% rice starch for 65 hours at 30 °C.), representing the type strains of 1 EPB species Photorhbdus luminescens TT01, [85] (Waterfield t al., (2005). The latter species were represented by 2 colony-color variants (yellow and red) distinguishable on MacConkey [86] (Rahaman et al., 1975) agar plates..
2.2. Bioassays
2.2.1. Bioassays on Leishmania and Trypanosoma Cruzi Species, as well as on Control Test-Organisms.
The bioassays were extended to clinical pathogenic protozoa using the previously described routine methods [87] (McGwire, and Kulkarni 2010). CFCM of EMA and EMC (stored in a cold room, 4 oC) for two years) were taken for these experiments. Both native and autoclaved CFCM were tested in each bioassay. Samples of EMA-CFCM and EMC-CFCM were also fractionated using Centricon spin columns as described by McGwire (unpublished) and fractions of different size ranges were assayed for activity. The bioassays were carried out in 96-hole tissue culture plates, in 200 ul volumes, containing 95 µl of the respective liquid media, 100 µl cell-free respective CFCM, and 5 µl of overnight LB cultures of the respective test organisms in duplicate. Briefly, colony count assays were used to measure activity against clinical isolates of bacteria (and Candida). Overnight incubation in 25-100 microliters of LB medium was followed by plating on a solid medium. MTT utilization assays were performed after overnight incubation of cultures up to stationary phase pro- or epimastigotes, respectively with CFCM (1:1 ratio) for 4hrs for anti-leishmanial and- trypanosomal activity. Each case's control was of untreated microbes assayed in parallel with the LB medium alone. A reduction of CFUs (for bacteria or Candida) or MTT activity (for protozoa) of at least 50% was considered + for antimicrobial activity. In our Leishmania studies, macrophages, macrophage culture were used as cytopathogenecity control test-organism: the macrophage cell line J774A.1 (TIB-67; ATCC, Manassas, VA, USA) was used for all present study experiments. J774 cells [62] (Tominaga et al., 1998), [63] (Andreu et al., 2017) were grown in DMEM with high glucose (25 mM) and 1.5 g/L NaHCO3, 50,000 U/L penicillin 50 mg/L streptomycin, and 10% FCS (heat-inactivated). Cells were grown in bioreactor tubes in an incubator at 37 °C and 5% CO2. The J774 cells were passaged twice a week, similar to those described in more detail later,˙[64] (Geroldinger et al., 2019).
2.2.2. Bioassays on.H. Meleagridis Grown in Monoxenic Culture with / E. coli
Each of the above-mentioned CFCMs were tested on Histomonas cells grown in vitro monoxenic culture with Escherichia coli DH5α cells.
The monoxenic culture of Histomonas meleagridis Turkey/Austria/ 2922-C6/04 with Escherichia coli DH5α which was grown in Medium 199 containing Earle’s Salts, L-glutamine, 25 mM HEPES, L-amino acids, 15% heat-inactivated fetal bovine serum FBS and 0.22% rice starch for 72 hours at 40 °C, as described before, [48.] (Ganas et al., 2012), [61] (Amin et al.,2012), [49] (Gruber et al., 2017), [51] (Bilic and Hess , 2020).
Consequently, the anti-E.coli activities also had to be determined. In the first experiment (see Supplementary Material) the anti-histomonal tests were cell-free filtrates from cultures of P. luminescens TT01 yellow and red variants grown in Medium 199 containing Earle’s Salts, L-glutamine, 25 mM HEPES, L-amino acids, 15% heat-inactivated fetal bovine serum FBS and 0.22% rice starch for 65 hours at 30 °C were tested on E.coli and then the treated culture s were plated, while the effects on the Histomonas were measured by using a scoring system .The applied scoring system [61] (Amin et al. (2012), as well as the suggested scale we used are presented in Table 2). For scoring the degree of cell destruction, the following system was applied: 0 = intact layer; 1 = up to 25% of the cell-layer destructed; 2 = 25-50% of the cell-layer destructed; 3 = 50-75% of the cell-layer destructed; 4 = more than 75% of the cell-layer destructed). In these experiments, 60% dilution of EMA-CFCM and 80% dilution of EMC-CFCM were used as stocks and serially diluted further. In the second experiment, serially diluted CFCMs were tested. For more details, see Supplementary Material.
2.3. Constrauction and Bioassay of the Antimicrobial Potential of hfq(del) Mutants of EMA and EMC
2.3.1. Construction of hfq(del) Mutants of EMA and EMC (for Detais, se Appendix A)
The Δhfq mutant EMC strain was generated according to the one-step gene inactivation method [79] (Datsenko & Wanner, 2000), which was adapted to Photorhabdus [80] (Yin et al., 2015) then to Xenorhabdus.
The Δhfq mutant EMC strain was generated as follows: The apropriately modified plasmids (As for the relevant features of plasmids, see Table A1 in Appendix A,) were introduced into X. szentirmaii (EMC) by electroporation in our lab. Electrocompetent cells were prepared for introduction of plasmids as follows: 1 ml of X. szentirmaii (EMC) culture grown at 30°C in LB broth supplemented with Ap (150 μg/ml) to OD600~1.1-1.2 was centrifuged for 30 s at 9000 rpm at 2°C, then resuspended in ice-cold wash buffer SMG (0.5 M sorbitol, 0.5 M mannitol, 10 % glycerol) and centrifuged again. The washing step was repeated two times, then the cells were resuspended in 60μl SMG buffer. Approximately. 1 μg plasmid DNA purified by the QIAGen Plasmid Midi Kit according to the manufacturer’s protocol was added to 40μl electrocompetent cells. Electroporation was carried out using 2-mm gap electroporation cuvettes and BTX Electro Cell Manipulator 600 with the setting 129 ohm 24 kV/cm. Transformants were selected on LB plates supplemented with the appropriate antibiotics (Km - 120 μg/ml, Sm – 100μg/ml, Gm – 50 μg/ml, and Ap - 150 μg/ml).
For the hfq KO recombination, EMC cells containing plasmid pBZs7 were grown in LB broth supplemented with Gm and Ap to a late logarithmic (OD600~1.1-1.2) phase, then the culture was 2x diluted with fresh LB broth, L-arabinose was added in a final concentration of 1% and grown further in a shaker for 1.5 h at 30°C. The electro-competent cells were prepared as described above. The KO PCR fragment was amplified using pKD4 template plasmid and the primers hfq_EMCdelfor and hfq_EMCdelrev. The amplicon was precipitated in 96% ethanol, dried, and resuspended in 10 mM Tris pH 8.0. For electroporation, approx. 1 μg purified DNA fragment was used as described. After electroporation, cells were grown for 5 h at 30°C, then spread on LB agar plates supplemented with 120 μg/ml Km and incubated at 30°C for 2 days. The Δhfq::KmR recombinant was tested by colony PCRs. For deletion of the KmR cassette from the Δhfq::KmR mutant, pBZs1 was electroporated. A SmRApRKmR transformant colony was grown overnight at 30°C in LB broth supplemented with Ap, spread on LB+Ap agar plates, and incubated for 24 h at 30°C, then the colonies were individually tested for the loss of the KmR gene, and the pBZs1 plasmid on LB+Ap, LB+Km, and LB+Sm plates. The KmS Δhfq strains were tested by colony PCR using the primer pair hfqseqfor-hfqseqrev.
The Δhfq mutant EMA strain was generated according to [69] (Tobias et al., 2017). The 769-bp upstream and the 617 bp downstream region of the hfq gene of EMA were amplified with primer pairs hfq_upforBam-hfq_uprev and hfqEMA_downfor-hfq_downrevSac, respectively. The two amplicons were assembled in the following PCR: The assembled amplicon was purified using the Illustra GFX PCR DNA and Gel Band Purification Kit (GE Healthcare), digested with BamHI and SacI, and ligated into the BamHI-SacI site of pBluescript II SK(+) (pBZS13). After sequencing, the BamHI-SacI fragment from pBZS13 was transferred into the BamHI-SacI site of the R6Kγ-based plasmid pBZs12, leading to the SacB-bearing mobilizable recombination vector pBZs17.
pBZs17 was mobilized into EMA from E. coli S17-1 λ pir. The ON culture of S17-1 λ pir/pBZs17 was 50× diluted with fresh LB broth supplemented with Km and Sm and grown 3 h to OD600~0.5, while the EMA recipient was grown in LB+Ap ON to OD600~1.0. 0.1 ml of donor and 1 ml of recipient cultures were mixed, centrifuged, and washed twice with 0.9% NaCl solution, then spread onto LB agar plates.
After 4h incubation at 30°C, the bacterial lawn was resuspended in 3 ml 0.9% NaCl solution, and the cells were centrifuged and spread onto LB+KmAp agar plates. After four days of incubation at 30°C, the KmRGmRApR recombinant colonies were tested by colony PCRs using primer pairs EMAhfqseqfor-trn5neo3 and pUCfor24-hfqseqrev.
To gain Δhfq recombinants, the EMA::pBZs17 strain was grown for 18 h at 30°C in LB+Ap broth, and 50, 100, and 200 μl of the culture was spread onto LB+Ap+5% sucrose agar plates. After 48h incubation at 30°C, the individual SacR colonies were tested for KmS, GmS phenotype, then, the appropriate colonies were tested by colony PCR using primers EMAhfqseqfor and hfqseqrev. To verify the hfq deletion, the amplicon was sequenced.
2.3.2. Bioassy of the Antimicrobial Activities of Wild-Type and hfq(del) Mutants of EMA and EMC
The bioassays were carried out in 96-hole (8 rows and 12 columns) tissue culture plates, in 200 ul volumes, containing 95 µl Mueller-Hinton liquid media, 100 µl cell-free respective CFCM, and 5 µl of overnight LB cultures of the respective test organisms in duplicate. Each test was carried out in duplicates, in the following way: rows AB: contained 100 µl wild-type CFCMs, 95 µl Mueller-Hinton (MH)-media, and 5 µl of stationery phase test organisms from 37 oC overnight cultures in each hole. Rows C and D rows contained 100 µl of the hfq-del mutant CFCMs of the same species (EMA, or EMC), 95 µl Mueller-Hinton (MH)-media, and 5 µl of stationery phase culture of the respective test organism (from 37 oC overnight cultures) in each hole. Row E: 100 µl wild-type CFCMs, 100 µl Mueller-Hinton liquid (MH)-media. Row F: 100 µl the hfq-del mutant CFCMs of the same species (EMA, or EMC), 100 µl Mueller-Hinton (MH)-media. Row G: 195 µl of 50% Mueller-Hinton (MH)-media + 5 µl of stationery phase test organisms (from 37 oC overnight cultures) in each hole. Row H: Row G: 200 ul of 50% Mueller-Hinton (MH)-media 5 µl of stationery phase test organisms( from 37 oC overnight culture) in each hole.
2.4. Statistical Analysis
ANOVA was carried out using the respective propositions of the SAS 9.4, see Acknowledgment section Software mainly due to unbalanced data set. The significant differences (α = 0.05) between treatment means were assessed using the Least Significant Difference (LSD).
3. Results
3.1. Results on Anti-Leishmanial Potential
Antileishmanial activities of the cell-free conditioned media (CFCM) of three species of the entomopathogenic nematode-symbiont bacterium genus Xenorhabdus, X. budapestensis sp. nov (Lengyel) DSM16342 (T) EMA, X. szentirmaii sp. nov.,(Lengyel) DSM 16338 (T) EMC, and X. innexii sp. nov., (Lengyel) DSM16336 (T) EMK were determined, and compared with antibacterial and antifungal potential. Each of the three (EMA, EMC, EMK) examined CFCM showed significant anti-leishmanial activities, but EMA and EMC exerted stronger anti-microbial potential in general than that EMK. Data are summarized in Table 1.
Each of the three CFCMs strongly antagonized each of the tested clinical isolates of Gram-positive (methicillin-sensitive (MSSA), and resistant (MRSA)) Staphylococcus aureus. Similarly, these CFCMs were active against clinical isolates of Gram-negative clinical isolates of Francisella novicida, Escherichia coli, Salmonella Typhimurium, as well as Pseudomonas aeruginosa, and a clinical isolate of Candida albicans (Table 1).
Autoclaved CFCM samples of EMA and EMC were as active as the native ones. As for EMK, only native CFCM samples were tested. The results demonstrated the thermostability of the active components in CFCM of EMA and EMC since autoclaving of these samples did not affect the antimicrobial activity against any of the organisms.
As for the molecular sizes of the active natural fractions of the CFCM, interestingly the antimicrobial activity against all microbes was retained in different size fractions of EMA (below 3kDa) and EMC (above 3kDa
3.2. Results on Anti-Histomonal Potential
To learn whether the NR_AMP molecules were produced by strong AMP-producing EPB species, the antihistomonal potential CFCMs from 4 different cultures were compared, see Materials and Methods. Each CFCM was tested on Histomonas cells which, (as discovered by Petra Ganas el (2012) [33]) can be in vitro cultured reproducibly in monoxenic culture with Escherichia coli DH5α cells. Consequently, the anti-Gram-negative bacterial activities were also determined. As for the Photorhabdus data, they are presented in Figure 1, (effects on Histomonas), and Table 2 (effects on Escherichia coli DH5α) cells. The results indicated that there is no difference between the antimicrobia/anti-Histomonal potential of the colony color variants as both exerted equally strong effects on both partners of the monoxenic culture. In the cultures supplemented with either the red or the yellow colony color variants of TT01, CFCM, the cell propagation ceased and the Histomonas cells - earlier or later – died.
From Figure 1, and Table 2 we concluded that the tested CFCM of each of the two colony-color variants of the EPB P. luminescence exerted cytotoxic activity not only on the prokaryotic (E. coli DH5 @ (alpha), but the eukaryotic (H. meleagridis Turkey/Austria/ 2922-C6/0 pathogen) partners as well. The growth curve of the monoxenic culture from the non-supplemented, (that is, only FKS-containing M199) control media, (green), could be characterized as performing a perfect log phase, after a moderate growth period. It starts at 24h and reaches a peak between 48–72hrs, after which it declines. The growth curve of the monoxenic culture grown in the „completely supplemented” M199 control media (blue) can be characterized by a log phase that starts right after the start of incubation (at 0 h), and then reaches a peak at 48 h, and then it rapidly declines. can be observed. Each of the exogenous EPB CFCM drastically killed the Histomonas cells. CFCM of the red variant acts immediately, while that of the yellow variants after 24 hrs. Even in the latter case, the size „growth peak” at 24h was less than half of the 4h value of the respective control cultures. As expected, the CFCM of each colony color variant was almost totally killing all the cells of the EMA a and monoxenic accompanying prokaryotic species E. coli DH5 alpha (Table 2).
3.3 . Antimicrobial Activities of the hfq(del) Mutants EMA, and EMC
We have constructed hfq-del regulatory mutants similar to those of [67] (Bode et al., 2019) from both EMA and EMC strains. The relevant publication has been in preparation, the method we constructed can be seen in Appendix A. Wethen bioassayed the antimicrobial activities of the respective CFCMs on several targets. We intend to apply the recently discovered, and elaborated ingenious method called the easyPACId method [k1] d to identify antiprotozoal compounds in the future, similarly as it had been done by Prof. Selcuk Hazir, and his associates at Menderes University in Ankara, Turkey, [70] (Gulsen et al., 2022). his technique has been discovered and elaborated by Edna Bode and her associates in Frankfurt; Germany, [66] (Bode et al., 2019). Briefly, ∆hfq mutants of two studied EPB bacterial species X. budapestensis and X. szentirmaii were constructed, and then the CFCMs of the wt and hfq mutants were used to test different pathogen organisms. But before doing that we intend to forecasts the limitations of the potential use of this approach. To learn how this fact influences the reproducibility of the method we tested the antimicrobial potential of the hfq mutant of both EMA and EMC species, and the data are presented in Table 3, Table 4, and Table 5
These data indicate that the strong anti-Gram-negative potential of the EMA strain severely but not probably not exclusively is under the control of the hfq-govern genetic regulation [68] (Tobias, et al., 2016). The anti-Gram-positive antibacterial potential of EMC is limited in comparison to the EMA. It is similarly true concerning the anti-Gram-positive antibacterial potential, see Table 3.
These data indicate that the extremely strong anti-Gram-positive potential of the EMA strain severely but probably not exclusively is under the control of the hfq-govern genetic regulation [68] (Tobias, et al., 2016). The anti-Gram-positive antibacterial potential of EMC is stronger than the anti-Gram-negative potential, but, in comparison to that of the EMA is weaker. Note that the EMC-resistant Gram-positive species are not the same that those which are resistant to the antimicrobials present in the CFCM of the hfq-del regulatory mutant of EMA. To see how the intraspecific variability modifies the picture, we compared the resistance/sensitivity patterns of different geographical isolations of the unambiguously EMA,-sensitive Gram-positive, Paeibacillus larvae [88] (Ebeling et l., 2016). bacterium species. The P. larvae a rapidly spreading deadly pathogen organism for honey bee larvae are a significant pest of agricultural significance.
These data indicate that although the Paeibacillus larvae species are uniformly, and unambiguously sensitive to the antimicrobial(s) present in the CFCM of EMA wild-type strain, there is a significant intraspecific variability concerning partly the resistance//sensitivity issue to EMC-produced antimicrobials, partly resistance/sensitivity issue to the antibiotic potential if EMA hfq-del regulatory mutants. These data are of key importance when drawing our conclusion concerning the usefulness and the limitation of the potential use of the easyPACId [71] (Bode et al., 2019) approach in our future research. These data also confirm that the extremely strong anti-Gram-positive potential of the EMA strain severely but probably not exclusively is under the control of the hfq-govern genetic regulation [68] (Tobias, et al., 2016). As for our anti-histomonal stufies, we follow this way..
In (Figure 2 one can see the difference between the colonies of the wild. type and the hfq-( the regulatory mutant of the X. szentirmaii DSM 16338 (EMC). The spectacular difference is the presence (in the wild-type, left, a) and the mutant (right, b)
4. Discussion
This paper is about the anti-protist potential of antimicrobial compounds produced by entomopathogenic bacterium species (EMA, EMC, EMK Photorhabdus luminexscens TT01 [15] (Lengyel et al.,2005; [16] Fodor et al., 2022); Fodor et al.,2023, [85] (Waterfield, 2005) the obligate symbionts of certain entomopathogenic nematodes, (EPN), into their conditioned culture media (CFCM) in vitro, As for their chemical nature, of these antcrovials, they are non-ribosomal antimicrobial peptides, (NR-AMPs) which are enzymatically synthetized through more than one steps, in their respective biosynthetic pathways. Those genes encoding for the enzymes forming the respective biosynthetic pathway of a given NR-AMP are clustered in a single operon [92] (Jacob et al., 1960), also coalled as biosynthetic gene complex (BGC), [93] (Gualtieri et al., 2022), .One of the natural role of the procryiote partner in the EPN/EPB symbiosis is to protect of the given monoxenic symbitic association in a given polyxenic milieu, producing a chemical toolkit (e.g. a particular set of NR-AMPs) providing competitiveness against the prokaryotic and eukaryotic competitors, including protists. The biosynthesis of the given sets set of these effective antimicrobials are synchronized by unique, genuine hierarcchical genetic regulation mechanism called primary-secondary phase shift [62] (Nealson and Forst, 1996) with the hfq gene on the top, [66,67,68,69,70]. In laboratory liquid culture conditions (as well is in the active symbiotic stage) these compounds are produced abundantly providing an unexhausstible “golden mine” to the antimicrobial searching scientist, The bioassays of the CFCMs are the tools find e best sources and optimize the culture conditions, We had found that those strains (EMA and EMC) which were discovered and further developed not only by our international team of a laboratory without wallbot in those famous laboratories as those of Maxime Guatier in France and Helge Bode in Germany proving that these two strains are probably best what have eves found so far. and other laboartories are probably really excellent sources. These modest data presented here on an anti-Leishmanial and anti-Histonal potentials are seem to confirm this conclusion. The next step is to find a way to get mutant (not “genetically manipulated”, but mutant strains which are produce only single NR_AMP molecules instead of the banch of them. Te method to do that was recently discovered in the Bode laboratory [71,72,73,74,75] (Bode et al., 2019). We demonstrated heret hat we have strarted this project as well by constructing and characterizing hfq-del mutants from both EPB species EMA, and EMC). Our goals is to targeting to pathogens of veterinary and agricultural significance.
We have been interested in the drug potential of the NR-AMP of EMA and EMC against eukaryotic parasites and pathogens of eukaryotic (mammal and bird) hosts. We wonder whether a chemotherapeutic approach based on applying Xenorhabdus NR-AMPs to prevent and controlling of flagellate parasitic pathogen protozoan species like the intracellular mammalian (and human) pathogenic Leishmania donovani (Laveran & Mesnil, 1903) species complex, [25] (Fernández-Arévalo et al., 2020) and the extracellular avian pathogenic Histomonas meleagridis, (Smith, 1895), [89] (Gerbod et al., 2001), are a realistic possibility. Both target species are flagellated protozoon, however, Leishmania is intracellular, while Histomonas is an extracellular parasite, and both own strong, unique, and efficient resistance mechanisms. The strategy for targeting antimicrobials to the prokaryotic or eukaryotic parasite is entirely different because of the similarities between the intracellular structures of eukaryotic hosts and the eukaryotic parasites (pathogens). Our research conception is that we suppose that some Xenorhabdus NR_AMP (phenazine, fabclavine, or something else) may be capable of dysregulating mitochondrial function and stimulating apoptosis, [90] (Rochford et al., 2020).
Leishmania ssp. is one of the most problem-causing pathogens considering its sophisticated resistance mechanisms. Still, at the same time, it is an excellent model for analyzing the mechanisms of drug resistance in eukaryotes [39] (Segovia 1994) since 1956 when the first cases of clinical resistance have been appeared in [91] (Ponte-Sucre et al., 2017). From an application point of view, the medical or veterinary perspectives of natural compounds are at stake several aspects have to be taken into consideration: A) the antimicrobial potential; (B) the durability, thermotolerance, and bioavailability; (C) cytotoxicity, and unwanted side effects, especially if one group of target organisms are of eukaryotic (protist) pathogens.
We have demonstrated that Xenorhabdus CFCM has broad-spectrum antimicrobial activity against several - human microbial pathogens. However, it is unclear what the precise nature of the antimicrobial effects is. In other terms, the question is whether or if the same active molecules are responsible for the killing of each organism or just a special group of organisms. The CFCMs of all three Xenorhabdus species proved active against Leishmania promastigotes in our experiments, but neither of them was efficient against not Trypanosoma cruzi epimastigotes. Other researchers [75] (Gulsen et al., 2022) found that EMA and EMK CFCMs were equally efficient against both Leishmania and Trypanosoma. They stated that X. budapestensis, X. cabanillasii, X. hominickii, X. indica, X. innexii, and X. stockiae supernatants caused 100% mortality at the highest tested centration (10%) against the promastigote form of L. tropica. No differences occurred between this treatment group and positive control (P > 0.05). (N-methyl meglumine was used as positive control). They did not publish data on EMC in the quoted article.
The number of alternative explanations of these discrepancies between the data of the Hazír and those of our team, maybe more than one. One of the explanations may be related to the differences in the surface compositions between the two parasites that either confer resistance to the active compounds or the inactivated penetration intracellularly to act on specific molecular targets.
The identification of the antimicrobial component(s) present in the CFCMs is important to understand, as well as their mechanisms of action. Another possible alternative explanation is that the CFCM of the hfq-del mutants used in the experiments in Ankara exerted some antagonistic effects on Trypanosoma.
We decided to compare the antimicrobial potential of each of the used hfq-del mutants before using it as the source of double mutants producing single NR-AMPs. We focus only on EMA and EMC species. and Our data in Tables 35 suggest to beware of the potential basic antimicrobial activities of the hfq-del mutants because they may have unexpected antimicrobial potential. It would not be a surprise considering that several previous studies of different labs have revealed that approximately 7.5% of the total genes in Xenorhabdus bacteria are dedicated to secondary metabolite biosynthesis, and probably that majority of these products are NR-AMPs.
Calculating with about an average of 2.4Mb-size Xenorhabdus genomes, 1 kb locus-size, and about 10 ORF per operon, then we have to think about 2,400 genes and 240 respective operons (BGS) per genome. It compares with the average genome of Staphylococcus, we may count on about 225, operons. In EMA we are interested in 3, and in EMC and EMC in 12 operons only, and we cannot rule out that are several other operons encoding for enzymes to synthesize compounds with antimicrobial potential. We intend to establish and analyze RNA-seq from both the wild-type and the hfq-strains.
However, we suppose that the situation is probably somewhat simpler since the hfq probably regulates a well-defined group of operons providing the genetic background of the so-called “primary-secondary phase shift” [63] (Akhurst, 1980), (“phenotypic phase variation”, the term of Professor S. A. Forst) scenario. That is, only a relatively small set of natural product (NP) genes are involved, and most of the antimicrobial-synthetic genes are among them.
Toxicity studies using J774 macrophages [57] (Tominaga, 1998), [58] (Andreu et al., 2017) indicated that only the CFCM of EMA was able to cause significant killing of these cells. In contrast, the CFCMs of EMC or EMK had no effect. This experiment confirmed our previous unpublished observations that EMA exerts stronger cytotoxicity on a range of eukaryotic cells, which may cause limitations concerning future medical applications. But this selective cytotoxicity may provide an option to select for and find specific CFCMs against different eukaryotic parasites of different eukaryotic (animal, plant) hosts.
To put the right place the significance of our anti-leishmanial results, we have to realize that the current chemotherapies used to handle these dangerous diseases have rather limited efficacy and Sb- resistance [94] ( Kazemi-Rad et al., 2013) has ben spreading. As for the future, the main trend may be based upon the complete genome sequencing of multiple strains, taking advantage of the application of CRISPR/Cas9 technology, and in vivo, bioluminescence-based imaging has set the stage for advancing target-based drug discovery, [32] (Altamura et al., 2022). But, new drugs are urgently needed anyway.
As for Histomonas experiments, we concluded that in different degrees, but each studied EPB CFCM exerted rather strong cytopathogenic effects on the monoxenic culture of H. meleagridis Turkey/Austria/ 2922-C6/04 cells co-cultured eukaryotic targets as well, similarly to the Eimeria (data not shown). Considering the cytopathogenic effects were also unambiguously detected on a confluent monolayer of LMH cells (permanent chicken hepatocarcinoma liver cell line) [21] (Fodor et al, 2023), we have to work on finding a CFCM with strong antihistomonal, and low cytotoxic activity. To do that we intended to compare the anti-histomonal, and cytotoxic potential of the wild, the hfq-delated, and different single antibiotics.-producer double mutants (hfq-del, and reactivated induced promoter-carrying single antibiotic-producer EMA and EMC strains. Considering the estimated selective cytotoxic potential of different strains we intend to use not only manganous but normal cell lines in the cytotoxicity studies. As mentioned in the Introduction, in vitro assays relying solely on non-phagocytic cell models may not represent a realistic result as the effect of an antileishmanial agent should ideally be presented based on its cytotoxicity profile against reticuloendothelial system cells. [59]. (Brioschi et al., 2022).
As for the results in the control (bacterium, fungi and other strains), previous studies have identified several anti-microbial molecules in the CFCM of Xenorhabdus, including fabclavine, [12] (Fuchs et al., 2012). [13] (Fuchs et al., 2014) a specialized metabolite polypeptide/polyketide hybrid molecule that is synthesized via non-ribosomal peptide synthesis. Many different-sized derivatives of fabclavines can be produced by the X. szentirmaii species [72] (Wenski et al., 2019), and can vary between entomopathogenic nematode-symbiont (Xenorhabdus, Photorhabdus) bacterium species [72] (Wenski et al., 2019). These have a wide-ranging anti-microbial activity against various microbes [16] (Fodor, et al., 2022).
Multiple other antimicrobial molecules can also be produced in CFCMs, including xenofuranone A and B, [95] (Brachmann, et al., 2006) cabanillasin [96] (Houard, et al 2013), PAX peptides [97 (Fuchs et al., 2011), odilorhabdins, [98] (Racine, and Gualtieri, 2019), [93] (Lanois-Nouri et al., 2022) xenematides [99] (Xi et al, 2019), anti-oomycete peptides [100] (Xiao et al., 2012)¸ xenortide [101] (Reimer, 2014), rhabdoopeptided/xenortide-like peptides, [102] (Zhao, 2018), cyclic depsipeptide xenematide F, and G rhabdopeptides [103] (Bi et l., 2018), szentiamids [104] (Ohlendorf, et al., 2011), [105] (Nollmann, et al, 2012). These latter peptides have earlier been published to be active against Trypanosoma brucei, but we cannot confirm that.
Whether any of them, can be considered as a potential antihistomonal drug candidate will be learned if we had a chance to screen both the single mutant hfq-del stains of EMA and EMC and those double mutants (3 from EMA and 11 of EMC) in which the hfq is deleted end only one single operon is activated
5. Conclusions
The recently discovered hierarchical regulatory systems following signal integration provides a great metabolic versatility, that results in excellent adaptability and metabolic optimization in antibiotic- producing bacteria [106] (Monteagudo-Cascales et al., 2022). like EPBs. To emphasize the recent importance of these researches, we have to conclude that he delineation of specific anti-microbial products in CFCM can now be achieved using the “easyPACId” approach (easy Promoter Activated Compound Identification) which takes advantage of the alteration of regulation of coordinate gene clusters which products are responsible for their biosynthesis [71] (Bode et al., 2019). This discovery may revolutionize the Xenorhabdus/Photorhabdus researches, which in the age of multidrug resistance maybe important.
The point is that different biosynthetic gene clusters (BGCs, that is operons) are coordinately regulated [68] (Tobias et al., 2016) [69] (Tobias et al., 2017),by the gene encoding the RNA chaperone, Hfq, [671 (Bode et al., 2019). The experimental system is a deletion mutant of the regulatory gene Hfq [107,108,109,110,111] of the respective species. This kind of regulatory mutant strain does not produce any biosynthetic AMP, but the respective individual promoters upstream of the respective biosynthetic gene cluster can be changed to inducible promotors and this recombinant strain will produce only a single respective AMP molecule. This is further facilitated by the completed genome sequencing of several Xenorhabdus species including X. szentirmaii [102]) (Gualtieri et al., 2014), and X. budapestensis, [113] (Li et al., 2021) making possible the identification of the promoter regions of each biosynthetic gene cluster (BGC, operon). Creating bacterial mutants that synthesize individual antimicrobial products into CFCM can facilitate their identification purification and bioassays of antimicrobial activity. [74] Ozkan, et al., 2019), [75] (Cimen et al., 2021).
Author Contributions
Conceptualization, András Fodor, János Kiss, Anna Sebestyén, Bradford McGwire and Michael Hess; Data curation, László Pál and Bradford McGwire; Formal analysis, Claudia Hess; Funding acquisition, Károly Dublecz, Michael Klein, Bradford McGwire, Tibor Vellai and Michael Hess; Investigation, András Fodor, Claudia Hess, Petra Ganas, Zsófia Boros, János Kiss, László Makrai, László Pál , Manjusha Kulkarni and Bradford McGwire; Methodology, Petra Ganas, Zsófia Boros, János Kiss, László Makrai, Károly Dublecz, Anna Sebestyén and Manjusha Kulkarni; Project administration, Tibor Vellai and Michael Hess; Resources, Tibor Vellai and Michael Hess; Software, Zsófia Boros, János Kiss and László Makrai; Supervision, László Fodor, Michael Klein, Bradford McGwire, Tibor Vellai and Michael Hess; Validation, Claudia Hess, Petra Ganas, Károly Dublecz, Manjusha Kulkarni and Michael Hess; Visualization, László Makrai and Manjusha Kulkarni; Writing – original draft, András Fodor, László Fodor and Anna Sebestyén; Writing – review & editing, Claudia Hess, János Kiss, Károly Dublecz, Michael Klein, Eustachio Tarasco, Bradford McGwire, Tibor Vellai and Michael Hess. All authors will be informed about each step of manuscript processing including submission, revision, revision reminder, etc. via emails from our system or assigned Assistant Editor.
Funding
This research received no external funding, except for some parts of this study TÁMOP-4.2.2/B-10/1-2010-0025 project based on the financial support of the State of Hungary and the EU. The CFCM preparations for Histomonas experiments were supported by the Hess-Dublecz project, (recently a joint unit of the Hungarian University of Agriculture and Life Sciences (MATE). The Leishmania experiments were supported by the bench money of Bradford McGwire in Columbus, OH. All the molecular studies were supported by the Department of Genetics, Eövös University. (Prof, Vellai, OTHK Grant: Number:),
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All of the data are available from corresponding author. All data generated from this study are included in this article. and some information in the Supplementary Materials is strongly inseparable from information published in our previous paper, Fodor A, Vellai T, Hess C, Makrai L, Dublecz K, Pál L, Molnár A, Klein MG, Tarasco E, Józsa S, Ganas P, Hess M. XENOFOOD-An Autoclaved Feed Supplement Containing Autoclavable Antimicrobial Peptides-Exerts Anticoccidial GI Activity, and Causes Bursa Enlargement, but Has No Detectable Harmful Effects in Broiler Cockerels despite In Vitro Detectable Cytotoxicity on LHM Cells. Pathogens. 2023 Mar 14;12(3):458. https://doi.org/10.3390/pathogens12030458. PMID: 36986380; PMCID: PMC10059668.
Acknowledgments
This work has been done in a “laboratory without walls”, that is within a frame of a voluntary international research cooperation n Budapest, we thank Professor Tior Vellai, Head of the Department of the Genetics of Eötvös University, for welcoming us back that project, and supporting it financially and morally as well as technically. (This project was born 20 years earlier in that Department.). In Vienna, at the Clinic for Poultry and Fish Medicine, Department for Farm Animals and Veterinary Public Health, University of Veterinary Medicine (Vetmeduni), Vienna), Austria; we would like to express thanks and appreciation not only for their professional technical but the intellectual help to the whole research team of the Hess laboratory, especially to Aziza Amin, and Irina Prokofieva. . In Keszthely, at the Institute of Physiology and Nutrition, Georgikon Campus of Hungarian University of Agriculture and Life Sciences, (MATE), Hungary, where the previous (and probably the future) in vivo feeding experiments have been running. going on, we would like to express thanks and appreciation not only for their professional technical but the intellectual help of the Dublecz team. Many thanks for the For professional and financial help in the molecular and bioinformatical work I the team of János Kiss (especially Mónika Szabó). We would like to express the invaluable help in the laboratories of Dr. Michael Klein, Joe Hogan, and Brad McGwire concerning the Leishmania work in Ohio.
Conflicts of Interest
The authors declare no conflict of interest. We sincerely consider those fellow scientists working in the same field as potential cooperators, rather than competitors. Furthermore, the authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Appendix A
Expeimental details of the Construction of hfq(del) mutants of EMA and EMCsttains .
The Δhfq mutant EMC strain was generated according to the one-step gene inactivation method [107] (Datsenko & Wanner, 2000), which was adapted to Xenorhabdus. The λ Red recombinase was replaced by the Photorhabdus luminescence-derived homolog encoded by the Plu2934/Plu2935/Plu2936 operon [108] (Yin et al., 2015). The plasmid pSC101-BAD-plu2934/2935/2936-gamma-amp containing a thermosensitive pSC101-derived replication system and the Plu2934/Plu2935/Plu2936 operon with the λ Redγ gene under the control of ParaBAD was a kind gift from Jun Fu. Plasmids were introduced into X. szentirmaii (EMC) by electroporation. Electrocompetent cells were prepared for introduction of plasmids as follows: 1 ml of X. szentirmaii (EMC) culture grown at 30°C in LB broth supplemented with Ap (150 μg/ml) to OD600~1.1-1.2 was centrifuged for 30 s at 9000 rpm at 2°C, then resuspended in ice-cold wash buffer SMG (0.5 M sorbitol, 0.5 M mannitol, 10 % glycerol) and centrifuged again. The washing step was repeated two times, then the cells were resuspended in 60μl SMG buffer.
Approx. 1 μg plasmid DNA purified by the QIAGen Plasmid Midi Kit according to the manufacturer’s protocol was added to 40μl electrocompetent cells. Electroporation was carried out using 2-mm gap electroporation cuvettes and BTX Electro Cell Manipulator 600 with the setting 129 ohm 24 kV/cm. Transformants were selected on LB plates supplemented with the appropriate antibiotics (Km - 120 μg/ml, Sm – 100μg/ml, Gm – 50 μg/ml, and Ap - 150 μg/ml).
For the hfq KO recombination, EMC cells containing plasmid pBZs7 were grown in LB broth supplemented with Gm and Ap to a late logarithmic (OD600~1.1-1.2) phase, then the culture was 2x diluted with fresh LB broth, L-arabinose was added in a final concentration of 1% and grown further in a shaker for 1.5 h at 30°C. The electro-competent cells were prepared as described above. The KO PCR fragment was amplified using pKD4 template plasmid and the primers hfq_EMCdelfor and hfq_EMCdelrev. The amplicon was precipitated in 96% ethanol, dried, and resuspended in 10 mM Tris pH 8.0. For electroporation, approx. 1 μg purified DNA fragment was used as described. After electroporation, cells were grown for 5 h at 30°C, then spread on LB agar plates supplemented with 120 μg/ml Km and incubated at 30°C for 2 days. The Δhfq::KmR recombinant was tested by colony PCRs using the primer pairs hfqseqfor-trn5neo5, rn5neo3-hfqseqrev, and hfqseqfor-hfqseqrev.
For deletion of the KmR cassette from the Δhfq::KmR mutant, pBZs1 was electroporated. A SmRApRKmR transformant colony was grown overnight at 30°C in LB broth supplemented with Ap, spread on LB+Ap agar plates, and incubated for 24 h at 30°C, then the colonies were individually tested for the loss of the KmR gene, and the pBZs1 plasmid on LB+Ap, LB+Km, and LB+Sm plates. The KmS Δhfq strains were tested by colony PCR using the primer pair hfqseqfor-hfqseqrev.
The Δhfq mutant EMA strain was generated according to [109] (Tobias et al., 2017). The 769-bp upstream and the 617 bp downstream region of the hfq gene of EMA were amplified with primer pairs hfq_upforBam-hfq_uprev and hfqEMA_downfor-hfq_downrevSac, respectively. The two amplicons were assembled in the following PCR:
Using 0.5 μl of the upstream and 2 μl of the downstream amplicons, 60 s denaturation at 94 °C followed by 15 cycles of 94°C 20 s, 55°C 30 s, 72°C 2 min, then 0.5 μl of primers hfq_upforBam and hfq_downrevSac (10 μM each) were added and the cycling was continued for further 35 cycles. The amplicon was purified using the Illustra GFX PCR DNA and Gel Band Purification Kit (GE Healthcare), digested with BamHI and SacI, and ligated into the BamHI-SacI site of pBluescript II SK(+) (pBZS13). After sequencing, the BamHI-SacI fragment from pBZS13 was transferred into the BamHI-SacI site of the R6Kγ-based plasmid pBZs12, leading to the SacB-bearing mobilizable recombination vector pBZs17.
pBZs17 was mobilized into EMA from E. coli S17-1 λ pir. The ON culture of S17-1 λ pir/pBZs17 was 50× diluted with fresh LB broth supplemented with Km and Sm and grown 3 h to OD600~0.5, while the EMA recipient was grown in LB+Ap ON to OD600~1.0. 0.1 ml of donor and 1 ml of recipient cultures were mixed, centrifuged, and washed twice with 0.9% NaCl solution, then spread onto LB agar plates. After 4h incubation at 30°C, the bacterial lawn was resuspended in 3 ml 0.9% NaCl solution, and the cells were centrifuged and spread onto LB+KmAp agar plates. After four days of incubation at 30°C, the KmRGmRApR recombinant colonies were tested by colony PCRs using primer pairs EMAhfqseqfor-trn5neo3 and pUCfor24-hfqseqrev.
To gain Δhfq recombinants, the EMA::pBZs17 strain was grown for 18 h at 30°C in LB+Ap broth, and 50, 100, and 200 μl of the culture was spread onto LB+Ap+5% sucrose agar plates. After 48h incubation at 30°C, the individual SacR colonies were tested for KmS, GmS phenotype, then, the appropriate colonies were tested by colony PCR using primers EMAhfqseqfor and hfqseqrev. To verify the hfq deletion, the amplicon was sequenced.
Restriction enzymes and DNA polymerases were purchased from Thermo Scientific. Colony PCRs were carried out using Dream Taq polymerase, while for the amplification of fragments for cloning or KO experiments, a Phusion polymerase was applied.
For maintaining the plasmids with a temperature-sensitive pSC101 replication system, growth at 30°C with antibiotic selection was applied,while curing these plasmids was carried out by growth at 30°C without antibiotic selection, followed by testing the resistance pattern of individual colonies for selecting plasmid-cured clones.
For cloning purposes, E. coli TG1 (Gibson, 1984) was used as host strain. For plasmid mobilization into Xenorhabdus, E. coli S17-1λ pir (Simon, Priefer, & Pühler, 1983a) was applied. E. coli strains were routinely growth in LB at 37°C with appropriate antibiotic selection.Restriction enzymes and DNA polymerases were purchased from Thermo Scientific.Colony PCRs were carried out using Dream Taq polymerase, while aplification of fragments for cloning or KO experiments, Phusion polymerase was applied.
Relevant features of plasmids used in this work
| Name | Relevant features | References |
| pBluescript II SK (+) | pMB1-based ApR cloning vector. | (Short, Fernandez, Sorge, & Huse, 1988) |
| pKD4 | KmR,ApR r6kγ-based PCR template plasmid for one-step recombination gene-KO | (Datsenko & Wanner, 2000)(Datsenko & Wanner, 2000) |
| pBZs1 | SmR,CmR,ApS, derivative of pCP20 (Cherepanov & Wackernagel, 1995) containing also the oriT of RP4, thermo-inducible FLP recombinase expression (λ pR::FLP), temperature-sensitive pSC101 replication system, λ cI857, mobilization by S17-1λ pir host strain | this work |
| pBZS7 | KmR,ApS derivative of pSC101-BAD-plu2934/2935/2936-gamma-amp (Yin et al., 2015), L-ara-inducible expression of λ Red recombinase homologue from P. luminescens and λ Redγ protein, temperature-sensitive pSC101 replication system | this work |
| pBZs12 | pJQ200-SK derivative, where the SspI-NheI fragment carrying the p15A origin of replication was replaced by the ClaI-NheI fragment of pSG76-K containing the KmR gene and R6Kγ origin of replication. KmR, GmR, RP4 oriT, R6Kγ oriV, SacB | this work |
| pBZs17 | pBZs12 derivative, containing the assembled upstream and downstream region of the hfq gene of EMA. KmR, GmR, RP4 oriT, R6Kγ oriV, SacB | this work |
| pSG76-K | R6Kγ-based KmR replicon | (Pósfai, Koob, Kirkpatrick, & Blattner, 1997) |
| pJQ200-SK | A SacB-bearing p15A-based vector. GmR, RP4 oriT, p15A oriV, SacB. | (Quandt & Hynes, 1993) |
Relevant Features of bacterial strains used.
| TG1 | supE hsdΔ5 thi Δ(lac-proAB) F’[traD36 proAB+ lacIq lacZΔM15] | (Gibson, 1984) |
| S17-1 λpir | S17-1 λpir, a λ lysogen derivative of S17-1 (pro thi recA hsdR (r- m+) TpR SmR KmS [Ω RP4-2-Tc::Mu-Km::Tn7]) expressing Π protein from pir gene of R6K | (Simon, Priefer, & Pühler, 1983b) |
| X. budapestensis DSM16342 | wt isolate | Lengyel, et al., 2005 |
| X. szentirmaii DSM 16338 | wt isolate |
List of oligonucleotides used.
| Primer | Sequence (5’-3’) | Reference |
| hfq_EMCdelfor | gaaccaaaaggttcttagttaaaacaacaaaataaggaaaatatagaatgGTGTAGGCTGGAGCTGCTTC | this work |
| hfq_EMCdelrev | aagtctccctacctatgtttttatttacttcaggtatggtcactgatttaCATATGAATATCCTCCTTAGTTC | this work |
| hfqEMAseqfor | tgtggtcttatctcgcgggc | this work |
| hfqseqfor | tctcatgatgagatggtttatcgtg | this work |
| hfqseqrev | gcacaggagaaacacctgcgg | this work |
| trn5neo3 | tcgccttctatcgccttcttga | this work |
| trn5neo5 | aatagcctctccacccaagcgg | this work |
| hfq_upforBam | aaggatccactcctggttggcggaaccatgc | this work |
| hfq_uprev | caaagattgccccttagccattctatattttccttattttg | this work |
| hfqEMA_downfor | caaaataaggaaaatatagaatggctaaggggcaatctttgatcgctgaataaatcagtgataatgcctgg | this work |
| hfq_downrevSac | aagagctctttgtcacgcagcatacggcggtc | this work |
| pUCfor24 | cgccagggttttcccagtcacgac | (Nagy, Szabó, Hegyi, & Kiss, 2021) |
References
- Fodor, A.; Abate, B.A.; Deák, P.; Fodor, L.; Gyenge, E.; Klein, M.G.; Koncz, Z.; Muvevi, J.; Ötvös, L.; Székely, G.; et al. Multidrug resistance (MDR) and collateral sensitivity in bacteria, with special attention to genetic and evolutionary aspects and to the perspectives of antimicrobial peptides-A Review. Pathogens 2020, 9, 522. [Google Scholar] [CrossRef] [PubMed]
- Ötvös L Jr, Wade JD. Current challenges in peptide-based drug discovery. Front Chem. 2014 Aug 8;2:62. PMID: 25152873; PMCID: PMC4126357. [CrossRef] [PubMed] [PubMed Central]
- Karelson M, Lobanov VS, Katritzky AR. Quantum-Chemical Descriptors in QSAR/QSPR Studies. Chem Rev. 1996 May 9;96(3):1027-1044. [CrossRef] [PubMed]
- Gini G. QSAR Methods. Methods Mol Biol. 2022;2425:1-26. [CrossRef]
- Lázár V, Martins A, Spohn R, Daruka L, Grézal G, Fekete G, Számel M, Jangir PK, Kintses B, Csörgő B, Nyerges Á, Györkei Á, Kincses A, Dér A, Walter FR, Deli MA, Urbán E, Hegedűs Z, Olajos G, Méhi O, Bálint B, Nagy I, Martinek TA, Papp B, Pál C. Antibiotic-resistant bacteria show widespread collateral sensitivity to antimicrobial peptides. Nat Microbiol. 2018 Jun;3(6):718-731. Epub 2018 May 24. [CrossRef] [PubMed] [PubMed Central]
- Kintses B, Jangir PK, Fekete G, Számel M, Méhi O, Spohn R, Daruka L, Martins A, Hosseinnia A, Gagarinova A, Kim S, Phanse S, Csörgő B, Györkei Á, Ari E, Lázár V, Nagy I, Babu M, Pál C, Papp B. Chemical-genetic profiling reveals limited cross-resistance between antimicrobial peptides with different modes of action. Nat Commun. 2019 Dec 16;10(1):5731. [CrossRef] [PubMed] [PubMed Central]
- Chaoulitch, SP. Leishmania donovani Laveran et Mesnil, 1903 parasite aussä les érythrocytes [Leishmania donovani Laveran and Mesnil 1903 also parasitic in erythrocytes]. Bull Soc Pathol Exot Filiales. 1954;47(2):244-6. Undetermined Language. [PubMed]
- Dutra-Rêgo F, Freire ML, Carvalho GML, Andrade-Filho JD. Revisiting the cave-dwelling sand flies (Diptera, Psychodidae, Phlebotominae) from Brazil: Diversity and potential role in the transmission of Leishmania Ross, 1903 (Kinetoplastida: Trypanosomatidae). Med Vet Entomol. 2022 Dec;36(4):408-423. Epub 2022 May 18. [CrossRef] [PubMed]
- McDougald, LR. Blackhead disease (histomoniasis) in poultry: a critical review. Avian Dis. 2005 Dec;49(4):462-76. [CrossRef] [PubMed]
- McDougald, LR. Intestinal protozoa important to poultry. Poult Sci. 1998 Aug;77(8):1156-8. [CrossRef] [PubMed]
- Hess M, Liebhart D, Bilic I, Ganas P. Histomonas meleagridis--new insights into an old pathogen. Vet Parasitol. 2015 Feb 28;208(1-2):67-76. Epub 2014 Dec 23. [CrossRef] [PubMed]
- Fuchs SW, Sachs CC, Kegler C, Nollmann FI, Karas M, Bode HB. Neutral loss fragmentation pattern based screening for arginine-rich natural products in Xenorhabdus and Photorhabdus. Anal Chem. 2012 Aug 21;84(16):6948-55. Epub 2012 Aug 6. [CrossRef] [PubMed]
- Fuchs SW, Grundmann F, Kurz M, Kaiser M, Bode HB. Fabclavines: bioactive peptide-polyketide-polyamino hybrids from Xenorhabdus. Chembiochem. 2014 Mar 3;15(4):512-6. Epub 2014 Feb 13. [CrossRef] [PubMed]
- Shi YM, Brachmann AO, Westphalen MA, Neubacher N, Tobias NJ, Bode HB. Dual phenazine gene clusters enable diversification during biosynthesis. Nat Chem Biol. 2019 Apr;15(4):331-339. Epub 2019 Mar 18. [CrossRef] [PubMed]
- Lengyel, K.; Lang, E.; Fodor, A.; Szállás, E.; Schumann, P.; Stackebrandt, E. Description of four novel species of Xenorhabdus, family Enterobacteriaceae: Xenorhabdus budapestensis sp. nov., Xenorhabdus ehlersii sp. nov., Xenorhabdus innexi sp. nov., and Xenorhabdus szentirmaii sp. nov. Syst. Appl. Microbiol. 2005, 28, 115–122. Erratum in: Syst. Appl. Microbiol. 2007, 30, 83. [CrossRef]
- Fodor, A. ; Gualtieri, M. ; Zeller, M. ; Tarasco, E. ; Klein, M.G. ; Fodor, A.M. ; Haynes, L. ; Lengyel, K. ; Forst, S.A. ; Furgani, G.M.;et al. Type Strains of Entomopathogenic Nematode-Symbiotic Bacterium Species, Xenorhabdus szentirmaii (EMC) and X. budapestensis (EMA), Are Exceptional Sources of Non-Ribosomal Templated, Large-Target-Spectral, Thermotolerant Antimicrobial Peptides, and Iodinin (EMC). Pathogens 2022, 11, x. [CrossRef]
- Akhurst, R.J. Antibiotic activity of Xenorhabdus spp., bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J. Gen. Microbiol. 1982, 128, 3061. [CrossRef]
- .Ogier, J.C.; Pagès, S.; Frayssinet, M.; Gaudriault, S. Entomopathogenic nematode-associated microbiota: monoxenic paradigm to pathobiome. Microbiome 2020, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Furgani, G.; Böszörményi, E.; Fodor, A.; Máthé-Fodor, A.; Forst, S.; Hogan, J.S.; Katona, Z.; Klein, M.G.; Stackebrandt, E.; Szentirmai, A.; et al. Xenorhabdus antibiotics: A comparative analysis and potential utility for controlling mastitis caused by bacteria. J. Appl. Microbiol. 2008, 104, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Fodor, A.; Makrai, L. , Fodor L.., Venekei, I., Husvéth, F., Pál, L., Molnár A., Dublecz, K., Pintér, Cs., Józsa, S., Klein, M. G. Anti-Coccidiosis Potential of Autoclaveable Antimicrobial Peptides from Xenorhabdus budapestensis Resistant to Proteolytic (Pepsin, Trypsin) Digestion Based on In vitro Studies. Microbiology Research Journal International, Page 1-17. Published: 11 January 2018. [CrossRef]
- ]Fodor A, Tibor Vellai, Claudia Hess, László Makrai, Károly Dublecz, László Pál, Andor Molnár, Michael G. Klein, Eustachio Tarasco, Sándor Józsa, Petra Ganas, Michael Hess XENOFOOD—An Autoclaved Feed Supplement Containing Autoclavable Antimicrobial Peptides—Exerts Anticoccidial GI Activity, and Causes Bursa Enlargement, but Has No Detectable Harmful Effects in Broiler Cockerels despite In Vitro Detectable Cytotoxicity on LHM Cells. 2023Pathogens 12(3):458. LicenseCC BY 4. [CrossRef]
- Böszörményi, E.; Érsek, T.; Fodor, A.; Fodor, A.M.; Földes, L.S.; Hevesi, M.; Hogan, J.S.; Katona, Z.; Klein, M.G.; Kormány, A.; et al. Isolation and activity of Xenorhabdus antimicrobial compounds against the plant pathogens Erwinia amylovora and Phytophthora nicotianae. J. Appl. Microbiol. 2009, 107, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Fodor, A.; Varga, I.; Hevesi, M.; Máthé-Fodor, A.; Racsko, J.; Hogan, J.A. Novel anti-microbial peptides of Xenorhabdus 57. origin against multidrug resistant plant pathogens. In: V. Bobbarala (Ed.) A Search for Antibacterial Agents. London, United Kingdom, IntechOpen, 2012 [Online]. Available from: https://www.intechopen.com/books/2129 https://doi.org/10.5772/1085. pp. 3–32.
- Vozik, D.; Bélafi-Bakó, K.; Hevesi, M.; Böszöményi, E.; Fodor, A. Effectiveness of a peptide-rich fraction from Xenorhabdus budapestensis culture against fire blight disease on apple blossoms. Not. Bot. Horti Agrobo 2015, 43, 547–553 Available online: https://wwwnotulaebotanicaero/ ndexphp/index (accessed on February 22 2022). [Google Scholar] [CrossRef]
- Fernández-Arévalo A, El Baidouri F, Ravel C, Ballart C, Abras A, Lachaud L, Tebar S, Lami P, Pratlong F, Gállego M, Muñoz C. The Leishmania donovani species complex: A new insight into taxonomy☆. Int J Parasitol. 2020 Nov;50(13):1079-1088. Epub 2020 Sep 1. [CrossRef] [PubMed]
- McGwire BS, Satoskar AR. Leishmaniasis: clinical syndromes and treatment. QJM. 2014 Jan;107(1):7-14. Epub 2013 Jun 5. [CrossRef] [PubMed] [PubMed Central]
- Alvar J, Cañavate C, Molina R, Moreno J, Nieto J. Canine leishmaniasis. Adv Parasitol. 2004;57:1-88. [CrossRef] [PubMed]
- Dantas-Torres F, Miró G, Baneth G, Bourdeau P, Breitschwerdt E, Capelli G, Cardoso L, Day MJ, Dobler G, Ferrer L, Irwin P, Jongejan F, Kempf VAJ, Kohn B, Lappin M, Little S, Madder M, Maggi R, Maia C, Marcondes M, Naucke T, Oliva G, Pennisi MG, Penzhorn BL, Peregrine A, Pfeffer M, Roura X, Sainz A, Shin S, Solano-Gallego L, Straubinger RK, Tasker S, Traub R, Wright I, Bowman DD, Gradoni L, Otranto D. Canine Leishmaniasis Control in the Context of One Health. Emerg Infect Dis. 2019 Dec;25(12):1-4. [CrossRef] [PubMed] [PubMed Central]
- Walker DM, Oghumu S, Gupta G, McGwire BS, Drew ME, Satoskar AR. Mechanisms of cellular invasion by intracellular parasites. Cell Mol Life Sci. 2014 Apr;71(7):1245-63. Epub 2013 Nov 13. [CrossRef] [PubMed] [PubMed Central]
- Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet. 2018 Sep 15;392(10151):951-970. Epub 2018 Aug 17. [CrossRef] [PubMed]
- Safavi M, Eshaghi H, Hajihassani Z. Visceral Leishmaniasis: Kala-azar. Diagn Cytopathol. 2021 Mar;49(3):446-448. Epub 2020 Nov 22. [CrossRef] [PubMed]
- Altamura F, Rajesh R, Catta-Preta CMC, Moretti NS, Cestari I. The current drug discovery landscape for trypanosomiasis and leishmaniasis: Challenges and strategies to identify drug targets. Drug Dev Res. 2022 Apr;83(2):225-252. Epub 2020 Apr 6. [CrossRef] [PubMed]
- Mauël, J. Vaccination against Leishmania infections. Curr Drug Targets Immune Endocr Metabol Disord. 2002 Oct;2(3):201-26. [CrossRef] [PubMed]
- Ibarra-Meneses AV, Corbeil A, Wagner V, Beaudry F, do Monte-Neto RL, Fernandez-Prada C. Exploring direct and indirect targets of current antileishmanial drugs using a novel thermal proteomics profiling approach. Front Cell Infect Microbiol. 2022 Aug 3;12:954144. [CrossRef] [PubMed] [PubMed Central]
- Douanne N, Wagner V, Roy G, Leprohon P, Ouellette M, Fernandez-Prada C. MRPA-independent mechanisms of antimony resistance in Leishmania infantum. Int J Parasitol Drugs Drug Resist. 2020 Aug;13:28-37. Epub 2020 Apr 2. [CrossRef] [PubMed] [PubMed Central]
- Golenser J and Domb, A. New formulations and derivatives of amphotericin B for treatment of leishmaniasis. Mini Rev Med Chem. 2006 Feb;6(2):153-62. [CrossRef] [PubMed]
- Ellis, D. Amphotericin B: spectrum and resistance. J Antimicrob Chemother. 2002 Feb;49 Suppl 1:7-10. [CrossRef] [PubMed]
- Chang KP, Reed SG, McGwire BS, Soong L. Leishmania model for microbial virulence: the relevance of parasite multiplication and pathoantigenicity. Acta Trop. 2003 Mar;85(3):375-90. [CrossRef] [PubMed]
- Segovia, M. Leishmania gene amplification: a mechanism of drug resistance. Ann Trop Med Parasitol. 1994 Apr;88(2):123-30. [CrossRef] [PubMed]
- Lollis L, Gerhold R, McDougald L, Beckstead R. Molecular characterization of Histomonas meleagridis and other parabasalids in the United States using the 5.8S, ITS-1, and ITS-2 rRNA regions. J Parasitol. 2011 Aug;97(4):610-5. Epub 2011 Feb 3. [CrossRef] [PubMed]
- Mitra T, Bilic I, Hess M, Liebhart D. The 60S ribosomal protein L13 is the most preferable reference gene to investigate gene expression in selected organs from turkeys and chickens, in context of different infection models. Vet Res. 2016 Oct 20;47(1):105. [CrossRef] [PubMed] [PubMed Central]
- Kidane FA, Bilic I, Mitra T, Wernsdorf P, Hess M, Liebhart D. In situ hybridization to detect and localize signature cytokines of T-helper (Th) 1 and Th2 immune responses in chicken tissues. Vet Immunol Immunopathol. 2016 Jul;175:51-6. Epub 2016 May 12. [CrossRef] [PubMed] [PubMed Central]
- Mitra T, Kidane FA, Hess M, Liebhart D. Unravelling the Immunity of Poultry Against the Extracellular Protozoan Parasite Histomonas meleagridis Is a Cornerstone for Vaccine Development: A Review. Front Immunol. 2018 Nov 2;9:2518. [CrossRef] [PubMed] [PubMed Central]
- Palmieri N, de Jesus Ramires M, Hess M, Bilic I. Complete genomes of the eukaryotic poultry parasite Histomonas meleagridis: linking sequence analysis with virulence / attenuation. BMC Genomics. 2021 Oct 21;22(1):753. [CrossRef] [PubMed] [PubMed Central]
- Hauck R, Balczulat S, Hafez HM. Detection of DNA of Histomonas meleagridis and Tetratrichomonas gallinarum in German poultry flocks between 2004 and 2008. Avian Dis. 2010 Sep;54(3):1021-5. [CrossRef] [PubMed]
- Lagler J, Schmidt S, Mitra T, Stadler M, Patricia Wernsdorf, Grafl B, Hatfaludi T, Hess M, Gerner W, Liebhart D. Comparative investigation of IFN-γ-producing T cells in chickens and turkeys following vaccination and infection with the extracellular parasite Histomonas meleagridis. Dev Comp Immunol. 2021 Mar;116:103949. Epub 2020 Nov 28. [CrossRef] [PubMed]
- Beer LC, Petrone-Garcia VM, Graham BD, Hargis BM, Tellez-Isaias G, Vuong CN. Histomonosis in Poultry: A Comprehensive Review. Front Vet Sci. 2022 May 6;9:880738. [CrossRef] [PubMed] [PubMed Central]
- Ganas P, Liebhart D, Glösmann M, Hess C, Hess M. Escherichia coli strongly supports the growth of Histomonas meleagridis, in a monoxenic culture, without influence on its pathogenicity. Int J Parasitol. 2012 Sep;42(10):893-901. Epub 2012 Aug 17. [CrossRef] [PubMed]
- Gruber J, Ganas P, Hess M. Long-term in vitro cultivation of Histomonas meleagridis coincides with the dominance of a very distinct phenotype of the parasite exhibiting increased tenacity and improved cell yields. Parasitology. 2017 Aug;144(9):1253-1263. Epub 2017 May 8 . [CrossRef] [PubMed]
- Liebhart D, Ganas P, Sulejmanovic T, Hess M. Histomonosis in poultry: previous and current strategies for prevention and therapy. Avian Pathol. 2017 Feb;46(1):1-18. Epub 2016 Dec 7. [CrossRef] [PubMed]
- Bilic I, Hess M. Interplay between Histomonas meleagridis and Bacteria: Mutualistic or Predator-Prey? Trends Parasitol. 2020 Mar;36(3):232-235. Epub 2020 Jan 22. [CrossRef] [PubMed]
- Arshad N, Zitterl-Eglseer K, Hasnain S, Hess M. Effect of Peganum harmala or its beta-carboline alkaloids on certain antibiotic resistant strains of bacteria and protozoa from poultry. Phytother Res. 2008 Nov;22(11):1533-8. [CrossRef] [PubMed] [PubMed Central]
- Hauck R, Hafez HM. Partial sequence of the beta-tubulin of Histomonas meleagridis and the activity of benzimidazoles against H. meleagridis in vitro. Parasitol Res. 2009 Apr;104(5):1183-9. Epub 2008 Dec 20. [CrossRef] [PubMed]
- Abraham M, McDougald LR, Beckstead RB. Blackhead disease: reduced sensitivity of Histomonas meleagridis to nitarsone in vitro and in vivo. Avian Dis. 2014 Mar;58(1):60-3. [CrossRef] [PubMed]
- Collins JB, Jordan B, Vidyashankar A, Bishop A, Kaplan RM. Fenbendazole resistance in Heterakis gallinarum, the vector of Histomonas meleagridis, on a broiler breeder farm in South Carolina. Vet Parasitol Reg Stud Reports. 2022 Nov;36:100785. Epub 2022 Sep 22. [CrossRef] [PubMed]
- Brioschi MBC, Coser EM, Coelho AC, Gadelha FR, Miguel DC. Models for cytotoxicity screening of antileishmanial drugs: what has been done so far? Int J Antimicrob Agents. 2022 Aug;60(2):106612. Epub 2022 Jun 9. [CrossRef] [PubMed]
- Tominaga T, Suzuki M, Saeki H, Matsuno S, Tachibana T, Kudo T. Establishment of an activated macrophage cell line, A-THP-1, and its properties. Tohoku J Exp Med. 1998 Oct;186(2):99-119. [CrossRef] [PubMed]
- Andreu N, Phelan J, de Sessions PF, Cliff JM, Clark TG, Hibberd ML. Primary macrophages and J774 cells respond differently to infection with Mycobacterium tuberculosis. Sci Rep. 2017 Feb 8;7:42225. [CrossRef] [PubMed] [PubMed Central]
- Geroldinger G, Rezk M, Idris R, Gruber V, Tonner M, Moldzio R, Staniek K, Monzote L, Gille L. Techniques to study phagocytosis and uptake of Leishmania tarentolae by J774 macrophages. Exp Parasitol. 2019 Feb;197:57-64. [CrossRef] [PubMed] [PubMed Central]
- Kawaguchi, T.; Nomura, K.; Hirayama, Y.; Kitagawa, T. Establishment and characterization of a chicken hepatocellular carcinoma, cell line, LMH. Cancer Res. 1987, 47, 4460–4464. [Google Scholar] [PubMed]
- Amin A, Bilic I, Berger E, Hess M. Trichomonas gallinae, in comparison to Tetratrichomonas gallinarum, induces distinctive cytopathogenic effects in tissue cultures. Vet Parasitol. 2012 May 25;186(3-4):196-206. Epub 2011 Nov 20. [CrossRef] [PubMed]
- Forst S, Nealson K. Molecular biology of the symbiotic-pathogenic bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol Rev. 1996 Mar;60(1):21-43. [CrossRef] [PubMed] [PubMed Central]
- Akhurst, R. J. (1980). Morphological and functional dimorphism in Xenorhabdus spp., bacteria symbiotically associated with the insect pathogenic nematodes Neoaplectana and Heterorhabditis. Journal of General Microbiology 121, 303-309.
- Akhurst, R.J. Antibiotic activity of Xenorhabdus spp., bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J. Gen. Microbiol. 1982, 128, 3061.
- Boemare, N., and R. J. Akhurst. 1988. Biochemical and physiological characterization of colony form variants in Xenorhabdus spp. (Enterobacteriaceae). J. Gen. Microbiol. 134:751–761.
- Joyce SA, Clarke DJ. A hexA homologue from Photorhabdus regulates pathogenicity, symbiosis and phenotypic variation. Mol Microbiol. 2003 Mar;47(5):1445-57. [CrossRef] [PubMed]
- Langer A, Moldovan A, Harmath C, Joyce SA, Clarke DJ, Heermann R. HexA is a versatile regulator involved in the control of phenotypic heterogeneity of Photorhabdus luminescens. PLoS One. 2017 Apr 27;12(4):e0176535. [CrossRef] [PubMed] [PubMed Central]
- Tobias NJ, Mishra B, Gupta DK, Sharma R, Thines M, Stinear TP, Bode HB. Genome comparisons provide insights into the role of secondary metabolites in the pathogenic phase of the Photorhabdus life cycle. BMC Genomics. 2016 Aug 3;17:537. [CrossRef] [PubMed] [PubMed Central]
- Tobias, N.J.; Heinrich, A.K.; Eresmann, H.; Wright, P.R.; Neubacher, N.; Backofen, R.; Bode, H.B. Photorhabdus-nematode symbiosis is dependent on hfq-mediated regulation of secondary metabolites. Environ. Microbiol. 2017, 19, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Neubacher N, Tobias NJ, Huber M, Cai X, Glatter T, Pidot SJ, Stinear TP, Lütticke AL, Papenfort K, Bode HB. Symbiosis, virulence and natural-product biosynthesis in entomopathogenic bacteria are regulated by a small RNA. Nat Microbiol. 2020 Dec;5(12):1481-1489. Epub 2020 Nov 2. [CrossRef] [PubMed] [PubMed Central]
- Bode E, Heinrich AK, Hirschmann M, Abebew D, Shi YN, Vo TD, Wesche F, Shi YM, Grün P, Simonyi S, Keller N, Engel Y, Wenski S, Bennet R, Beyer S, Bischoff I, Buaya A, Brandt S, Cakmak I, Çimen H, Eckstein S, Frank D, Fürst R, Gand M, Geisslinger G, Hazir S, Henke M, Heermann R, Lecaudey V, Schäfer W, Schiffmann S, Schüffler A, Schwenk R, Skaljac M, Thines E, Thines M, Ulshöfer T, Vilcinskas A, Wichelhaus TA, Bode HB. Promoter Activation in Δhfq Mutants as an Efficient Tool for Specialized Metabolite Production Enabling Direct Bioactivity Testing. Angew Chem Int Ed Engl. 2019 Dec 19;58(52):18957-18963. Epub 2019 Dec 12. [CrossRef] [PubMed] [PubMed Central]
- Wenski SL, Kolbert D, Grammbitter GLC, Bode HB. Fabclavine biosynthesis in X. szentirmaii: shortened derivatives and characterization of the thioester reductase FclG and the condensation domain-like protein FclL. J Ind Microbiol Biotechnol. 2019 Mar;46(3-4):565-572. Epub 2019 Jan 4. [CrossRef] [PubMed]
- Wenski SL, Cimen H, Berghaus N, Fuchs SW, Hazir S, Bode HB. Fabclavine diversity in Xenorhabdus bacteria. Beilstein J Org Chem. 2020 May 7;16:956-965. [CrossRef] [PubMed] [PubMed Central]
- Cimen H, Touray M, Gulsen SH, Hazir S. Natural products from Photorhabdus and Xenorhabdus: mechanisms and impacts. Appl Microbiol Biotechnol. 2022 Jun;106(12):4387-4399. Epub 2022 Jun 20. [CrossRef] [PubMed]
- Gulsen SH, Tileklioglu E, Bode E, Cimen H, Ertabaklar H, Ulug D, Ertug S, Wenski SL, Touray M, Hazir C, Bilecenoglu DK, Yildiz I, Bode HB, Hazir S. Antiprotozoal activity of different Xenorhabdus and Photorhabdus bacterial secondary metabolites and identification of bioactive compounds using the easyPACId approach. Sci Rep. 2022 Jun 24;12(1):10779. [CrossRef] [PubMed] [PubMed Central]
- Cherepanov, P. P. , & Wackernagel, W. (1995). Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene, 158(1), 9–14. [CrossRef]
- Gibson, T. J. (1984). Studies on the Epstein-Barr virus genome. Thesis. Cambridge. U /Also cited in Appendix A).
- Simon, R. , Priefer, U., & Pühler, A. (1983b). A Broad Host Range Mobilization System for In Vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Bio/Technology, 1(9), 784–791. Also cited in Appendix A). [CrossRef]
- Datsenko, K. A. , & Wanner, B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proceedings of the National Academy of Sciences, 97(12), 6640–6645. Also cited in Appendix A). [CrossRef]
- Yin, J., Zhu, H., Xia, L., Ding, X., Hoffmann, T., Hoffmann, M., … Zhang, Y. (2015). A new recombineering system for Photorhabdus and Xenorhabdus. Nucleic Acids Research, 43(6), e36. Also cited in Appendix A). 6. [CrossRef]
- Short, J. M. , Fernandez, J. M., Sorge, J. A., & Huse, W. D. (1988). Lambda ZAP: A bacteriophage lambda expression vector with in vivo excision properties. Nucleic Acids Research, 16(15), 7583–7600. Also cited in Appendix A). [CrossRef]
- Pósfai, G., Koob, M. D., Kirkpatrick, H. A., & Blattner, F. R. (1997). Versatile insertion plasmids for targeted genome manipulations in bacteria: Isolation, deletion, and rescue of the pathogenicity island LEE of the Escherichia coli O157:H7 genome. Journal of Bacteriology, 179(13), 4426–4428. Also cited in Appendix A). [CrossRef]
- Quandt, J., & Hynes, M. F. (1993). Versatile suicide vectors which allow direct selection for gene replacement in gram-negative bacteria. Gene, 127, 15–21. Also cited in Appendix A). [CrossRef]
- Nagy, I., Szabó, M., Hegyi, A., & Kiss, J. (2021). Salmonella Genomic Island 1 requires a self-encoded small RNA for mobilization. Molecular Microbiology, 116(6), 1533–1551. Also cited in Appendix A). [CrossRef]
- Waterfield N, Kamita SG, Hammock BD, ffrench-Constant R. The Photorhabdus Pir toxins are similar to a developmentally regulated insect protein but show no juvenile hormone esterase activity. FEMS Microbiol Lett. 2005 Apr 1;245(1):47-52. [CrossRef] [PubMed]
- Rahaman MM, Huq I, Dey CR. Superiority of MacConkey's agar over salmonella-shigella agar for isolation of Shigella dysenteriae type 1. J Infect Dis. 1975 Jun;131(6):700-3. [CrossRef] [PubMed]
- McGwire BS, Kulkarni MM. Interactions of antimicrobial peptides with Leishmania and trypanosomes and their functional role in host parasitism. Exp Parasitol. 2010 Nov;126(3):397-405. Epub 2010 Feb 14. [CrossRef] [PubMed]
- Ebeling J, Knispel H, Hertlein G, Fünfhaus A, Genersch E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl Microbiol Biotechnol. 2016 Sep;100(17):7387-95. [CrossRef] [PubMed]
- Gerbod D, Edgcomb VP, Noël C, Zenner L, Wintjens R, Delgado-Viscogliosi P, Holder ME, Sogin ML, Viscogliosi E. Phylogenetic position of the trichomonad parasite of turkeys, Histomonas meleagridis (Smith) Tyzzer, inferred from small subunit rRNA sequence. J Eukaryot Microbiol. 2001 Jul-Aug;48(4):498-504. [CrossRef] [PubMed]
- .Rochford G, Molphy Z, Kavanagh K, McCann M, Devereux M, Kellett A, Howe O. Cu(ii) phenanthroline-phenazine complexes dysregulate mitochondrial function and stimulate apoptosis. Metallomics. 2020 Jan 29;12(1):65-78. [CrossRef] [PubMed]
- Ponte-Sucre A, Gamarro F, Dujardin JC, Barrett MP, López-Vélez R, García-Hernández R, Pountain AW, Mwenechanya R, Papadopoulou B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl Trop Dis. 2017 Dec 14;11(12):e0006052. [CrossRef] [PubMed] [PubMed Central]
- Jacob F, Perrin D, Sanchez C, Monod J. [Operon: a group of genes with the expression coordinated by an operator]. C R Hebd Seances Acad Sci. 1960 Feb 29;250:1727-9. French. [CrossRef]
- Lanois-Nouri A, Pantel L, Fu J, Houard J, Ogier JC, Polikanov YS, Racine E, Wang H, Gaudriault S, Givaudan A, Gualtieri M. The Odilorhabdin Antibiotic Biosynthetic Cluster and Acetyltransferase Self-Resistance Locus Are Niche and Species Specific. mBio. 2022 Feb 22;13(1):e0282621. Epub 2022 Jan 11. [CrossRef] [PubMed] [PubMed Central]
- Kazemi-Rad E, Mohebali M, Khadem-Erfan MB, Saffari M, Raoofian R, Hajjaran H, Hadighi R, Khamesipour A, Rezaie S, Abedkhojasteh H, Heidari M. Identification of antimony resistance markers in Leishmania tropica field isolates through a cDNA-AFLP approach. Exp Parasitol. 2013 Oct;135(2):344-9. Epub 2013 Aug 6. [CrossRef] [PubMed]
- Brachmann, A.O.; Forst, S.; Furgani, G.M.; Fodor, A.; Bode, H.B. Xenofuranones A and B: Phenylpyruvate dimers from Xenorhabdus szentirmaii. J. Nat. Prod. 2006, 69, 1830–1832. [Google Scholar] [CrossRef] [PubMed]
- Houard J, Aumelas A, Noël T, Pages S, Givaudan A, Fitton-Ouhabi V, Villain-Guillot P, Gualtieri M. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J Antibiot (Tokyo). 2013 Oct;66(10):617-20. Epub 2013 Jun 12. [CrossRef] [PubMed]
- Fuchs SW, Proschak A, Jaskolla TW, Karas M, Bode HB. Structure elucidation and biosynthesis of lysine-rich cyclic peptides in Xenorhabdus nematophila. Org Biomol Chem. 2011 May 7;9(9):3130-2. Epub 2011 Mar 21. PMID: 21423922. [CrossRef] [PubMed]
- Racine, E.; Gualtieri, M. From worms to drug candidate: The story of odilorhabdins, a new class of antimicrobial agents. Front. Microbiol. 2019, 10, 2893. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.; Lu, X.; Zhang, X.; Bi, Y.; Li, X.; Yu, Z. Two novel cyclic depsipeptides Xenematides F and G from the entomopathogenic bacterium Xenorhabdus budapestensis. J. Antibiot. 2019, 2, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Meng, F.; Qiu, D.; Yang, X. Two novel antimicrobial peptides purified from the symbiotic bacteria Xenorhabdus budapestensis NMC-10. Peptides 2012, 35, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Reimer D, Nollmann FI, Schultz K, Kaiser M, Bode HB. Xenortide Biosynthesis by Entomopathogenic Xenorhabdus nematophila. J Nat Prod. 2014 Aug 22;77(8):1976-80. [CrossRef] [PubMed]
- Zhao, L.; Kaiser, M.; Bode, H.B. Rhabdopeptide/Xenortide-like peptides from Xenorhabdus innexi with terminal amines showing potent antiprotozoal activity. Org. Lett. 2018, 20, 5116–5120. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Gao, C.; Yu, Z. Rhabdopeptides from Xenorhabdus budapestensis SN84 and their nematicidal activities against Meloidogyne incognita. J. Agric. Food Chem. 2018, 66, 3833–3839. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf, B.; Simon, S.; Wiese, J.; Imhoff, J.F. Szentiamide, an N-formylated cyclic depsipeptide from Xenorhabdus szentirmaii DSM 16338T. Nat. Prod. Commun. 2011, 6, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Nollmann, F.I.; Dowling, A.; Kaiser, M.; Deckmann, K.; Grösch, S.; Ffrench-Constant, R.; Bode, H.B. Synthesis of szentiamide, a depsipeptide from entomopathogenic Xenorhabdus szentirmaii with activity against Plasmodium falciparum. Beilstein J. Org. Chem. 2012, 8, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Cascales E, Santero E, Canosa I. The Regulatory Hierarchy Following Signal Integration by the CbrAB Two-Component System: Diversity of Responses and Functions. Genes (Basel). 2022 Feb 18;13(2):375. [CrossRef] [PubMed] [PubMed Central]
- Ishihama, A. Modulation of the nucleoid, the transcription apparatus, and the translation machinery in bacteria for stationary phase survival. Genes Cells. 1999 Mar;4(3):135-43. [CrossRef] [PubMed]
- Vogel J, Luisi BF. Hfq and its constellation of RNA. Nat Rev Microbiol. 2011 Aug 15;9(8):578-89. [CrossRef] [PubMed] [PubMed Central]
- Cech GM, Szalewska-Pałasz A, Kubiak K, Malabirade A, Grange W, Arluison V, Węgrzyn G. The Escherichia Coli Hfq Protein: An Unattended DNA-Transactions Regulator. Front Mol Biosci. 2016 Jul 28;3:36. [CrossRef] [PubMed] [PubMed Central]
- King KA, Caudill MT, Caswell CC. A comprehensive review of small regulatory RNAs in Brucella spp. Front Vet Sci. 2022 Dec 1;9:1026220. [CrossRef] [PubMed] [PubMed Central]
- Ponath F, Hör J, Vogel J. An overview of gene regulation in bacteria by small RNAs derived from mRNA 3' ends. FEMS Microbiol Rev. 2022 Sep 2;46(5):fuac017; PMCID: PM. [CrossRef] [PubMed]
- Gualtieri, M.; Ogier, J.C.; Pagès, S.; Givaudan, A.; Gaudriault, S. Draft genome sequence and annotation of the entomopathogenic bacterium Xenorhabdus szentirmaii Strain DSM16338. Genome Announc. 2014, 2, e00190–14. [Google Scholar] [CrossRef] [PubMed]
- Li, B, Qiu, D.; Wang, S. Complete genome sequence data of Xenorhabdus budapestensis strain C72, a candidate biological control agent from China. Plant Dis. 2021, 105, 3276–3278. [CrossRef]
Figure 1.
Monoxenic culture: H. meleagridis (Turkey/Austria/ 2922-C6/04) cells were co-cultured with E. coli DH5 @ (alpha) cells: (a) in not-supplemented M199 tissue culture media, which contained FKS (green, or (b) in supplemented M199 tissue culture media (blue) (see Materials and Methods). The cells were viable, and propagating up to 72h in (a) (green), or at least 48 hours in (b) (blue), at 40°C in the CFCM-free cultures. In those cultures however, which were supplemented with the sterile filtered CFCM (either of the red or the yellow) TT01 colony-color variants), the cell propagation quickly ceased, both in the supplemented M199 media, (red, earlier) and in the non-supplemented (but FKS containing) M199 media, a little later) and then the Histomonas cells died. (Data are given in the Supplementary Material).
Figure 1.
Monoxenic culture: H. meleagridis (Turkey/Austria/ 2922-C6/04) cells were co-cultured with E. coli DH5 @ (alpha) cells: (a) in not-supplemented M199 tissue culture media, which contained FKS (green, or (b) in supplemented M199 tissue culture media (blue) (see Materials and Methods). The cells were viable, and propagating up to 72h in (a) (green), or at least 48 hours in (b) (blue), at 40°C in the CFCM-free cultures. In those cultures however, which were supplemented with the sterile filtered CFCM (either of the red or the yellow) TT01 colony-color variants), the cell propagation quickly ceased, both in the supplemented M199 media, (red, earlier) and in the non-supplemented (but FKS containing) M199 media, a little later) and then the Histomonas cells died. (Data are given in the Supplementary Material).
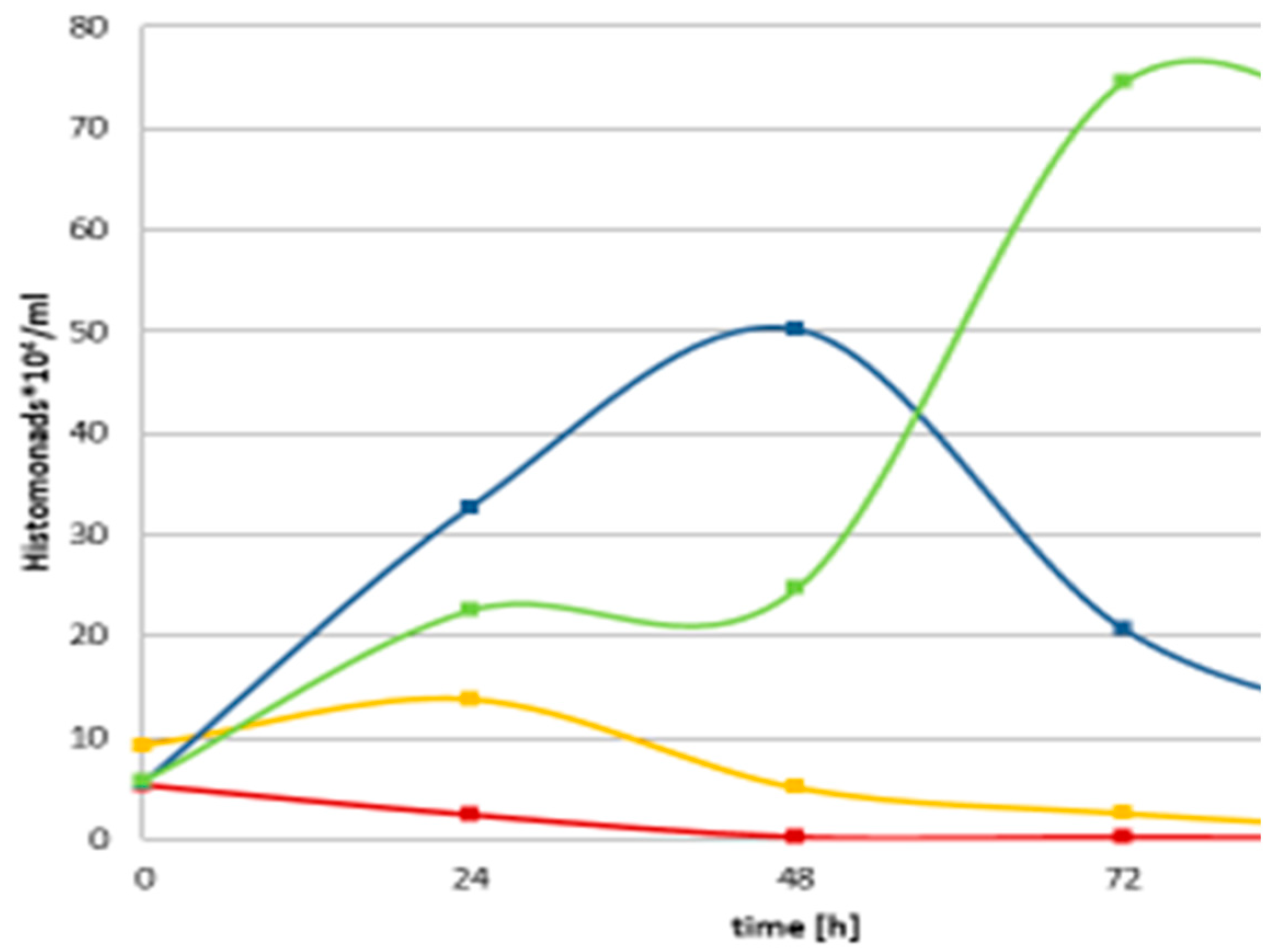
Figure 2.
Colony of the wild-type (left, B, and down, C), and the hfq-del regulatory mutant (left, A) of Xenorhabdus szentirmaii DSM16339 (EMC). The spectacular iodinine crystals, (the end product) of the phenazine biosynthesis are missing, indicating that the phenazine-operon encoded biosynthesis pathway is switched off. in the hfq-deleted regulatory mutant.
Figure 2.
Colony of the wild-type (left, B, and down, C), and the hfq-del regulatory mutant (left, A) of Xenorhabdus szentirmaii DSM16339 (EMC). The spectacular iodinine crystals, (the end product) of the phenazine biosynthesis are missing, indicating that the phenazine-operon encoded biosynthesis pathway is switched off. in the hfq-deleted regulatory mutant.


Table 1.
Antimicrobial Activities of 5-day Mid-Stationary-Phase Cell-Free Conditioned Culture Media (CFCM) of Xenorhabdus species.
Table 1.
Antimicrobial Activities of 5-day Mid-Stationary-Phase Cell-Free Conditioned Culture Media (CFCM) of Xenorhabdus species.
| BIOCHEMISTRY | ANTIMICROBIAL ACTIVITY | TOXICITY | ||||||||||||||||||
| Bacteria | ||||||||||||||||||||
| CFCM | Heat | PK | Centricon | Gram +) | (Gram -) | Protist parasite | Fungus | TOX | ||||||||||||
| MS | MS | FN | ST | EC | PA | LA | TC | CA | ||||||||||||
| EMA | + | - | <3 kDa | + | + | + | + | + | + | + | - | + | ||||||||
| EMC | + | + | >3 kDa | + | + | + | + | + | + | + | - | + | ||||||||
| EMK | ND | ND | ND | + | + | + | + | + | + | + | - | + | ||||||||
Table footer: Abbreviations: CM = 5-day Mid-Stationary-Phase Cell-Free Conditioned Culture Media of Xenorhabdus species: EMA = X budapestensis nov. DSM16342T; EMC = X. szentirmaii nov. DSM16338T; EMK = X. innexii nov. DSM16336T; Heat: autoclaved in standard conditions (15 min, 121oC, 1 Atm) and then tested; Centricon = indicates the molecular size of the separated fraction showing all antimicrobial activities; Tox = host toxicity, tested on determined microscopically and also determining MTT metabolism of J744-line murine macrophages (serving as checking for unspecific general cytotoxicity), exposed to the respective CM at 37oC for 72h. Abbreviations of the name of the tested microorganisms: MS and MR: methicillin-sensitive and -resistant Staphylococcus aureus, respectively; FN = Francisella novicida; ST = Salmonella typhimurium; EC = Escherichia coli; PA = Pseudomonas aeruginosa; LA = Leishmania amazonensis; TC = Trypanosoma cruzi; CA= Candida albicans. Activity assays were performed on cultured cells of the respective microorganisms incubated in 25-100 microliter of Luria Bertani-based, and appropriately supplemented media overnight. Each was then plated on the appropriate medium to CFU reduction assays (concerning bacterial or fungal pathogens) or using MTT utilization
Table 2.
Effect of Photorhabdus CFCMs on the prokaryote partner of H .meleagidis in monoxenic cuulture.
Table 2.
Effect of Photorhabdus CFCMs on the prokaryote partner of H .meleagidis in monoxenic cuulture.
| E. coli DH5 alpha | |||||||
| Sample | Hours | 0 | 24 | 48 | 72 | 96 | |
| 1 | TT01 Yellow | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 2 | TT01 Yellow | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 3 | TT01 Yellow | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 4 | TT01 Red | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 5 | TT01 Red | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 6 | TT01 Red | Single colonies | Single colonies | Single colonies | Single colonies | ||
| 7 | M199+15%MKS | Cell layer | Cell layer | Cell layer | Cell layer | ||
| 8 | M199+15%MKS | Cell layer | Cell layer | Cell layer | Cell layer | ||
| 9 | M199+15%MKS | Cell layer | Cell layer | Cell layer | Cell layer | ||
| 10 | Unchanged | Cell layer | Cell layer | Cell layer | Cell layer | ||
| 11 | Unchanged | Cell layer | Cell layer | Cell layer | Cell layer | ||
| 12 | Unchanged | Cell layer | Cell layer | Cell layer | Cell layer | ||
1 100 ul samples had been taken from H. meleagridis / E. coli monoxenic culture (grown in M199 media), and plated on McConkey agar afterward, where only the E. coli cells can grow, while the H. meleagridis cells cannot.
Table 3.
Anti-Gra-negative activities of CFCMs of wild-type n hfq (del) regulatory mutant cells of Xenorhabdus budapestensis DSM16342, and X. szentirmaii DSM 16338 (EMC).
Table 3.
Anti-Gra-negative activities of CFCMs of wild-type n hfq (del) regulatory mutant cells of Xenorhabdus budapestensis DSM16342, and X. szentirmaii DSM 16338 (EMC).
| TEST ORGANIS? | EMA (WT) | EMA (hfq-del) | EMC (WT) | EMC hfq-del) |
|---|---|---|---|---|
| GRAM NEGATIVE TARGETSS | ||||
| E. coli | - | - | - | + |
| Klebsiella pneumoniae | - | - | +/- | + |
| Salmonella enterica | - | + | + | + |
| Pasteurella multocida | - | - | + | + |
| Mannheimia haemolytica | - | + | + | + |
| Ps. aeruginosa VB(növ) | - | -/+ | + | + |
| Actinobacillus equuli | - | + | + | + |
| Enterobacter cloacae | - | +/- | + | + |
| Serratia marcescens | - | + | + | + |
| Bordetella bronchiseptica | - | - | + | + |
* It can be seen that each of the tested Gram-negative bacteria of veterinary significance proved completely sensitive to (one or more antimicrobial compounds in the CFCM of t wild-type Xenorhabdus budapestensis DSM16342 (EMA) strain. The majority of them (except for E.coli, Salmonella, and Bordetella) proved resistant to the antimicrobial active compound(s) present in the CFCM of the hfq (del) regulatory mutant of the same species (EMA). All but E. coli and Klebsiella proved)sensitive to the antimicrobial ingredient present in the CFCM of the wild-type X. szentirmaii DSM 16338 (EMC), but each of the proved fully resistant to the antimicrobial active compound(s) present in the CFCM of the hfq (del) regulatory mutant. Abbreviations: -: the test organism grew in the given CFCM, which is resistant to the antimicrobials present; +:: the test organism did not succeed in the given CFCM, which is sensitive to the antimicrobials present; +/-: the test organism grew at least one in one of the replicates, the given CFCM, that may be resistant to the antimicrobials present..
Table 4.
Anti-Gram-positive activities of CFCMs of wild-type n hfq (del) regulatory mutant cells of Xenorhabdus budapestensis DSM16342, and X. szentirmaii DSM 16338 (EMC).
Table 4.
Anti-Gram-positive activities of CFCMs of wild-type n hfq (del) regulatory mutant cells of Xenorhabdus budapestensis DSM16342, and X. szentirmaii DSM 16338 (EMC).
| TEST ORGANISMS | EMA (WT) | EMA (hfq-del) | EMC (WT) | EMC hfq-del) |
|---|---|---|---|---|
| GRAM POSITIVE | ||||
| Bacillus cereus | - | - | - | NT |
| Staphylococcus pseudintermedius | - | - | + | NT |
| Staphylococcus aureus (MRSA) | - | - | + | NT |
| Staphylococcus hyicus | - | - | NT | |
| Streptococcus suis | - | + | - | NT |
| Streptococcus agalactiae | - | - | - | NT |
| Listeria monocytogenes | - | + | - | NT |
| Erysipelothrix rhusiopathiae | - | - | - | NT |
| Corynebacterium pseudotuberculosis | - | - | - | NT |
| Rhodococcus equi | - | - | - | NT |
* It can be seen that each of the tested Gram-positive bacteria of veterinary significance is completely sensitive to (one or more antimicrobial compounds in the CFCM of t wild-type Xenorhabdus budapestensis DSM16342 (EMA) strain. All but two 2 (Streptococcus suis, and Lysteria monocytiogene) Gram-positive test bacteria proved also susceptible to the antimicrobial active compound(s) present in the CFCM of the hfq (del) regulatory mutant of the same species (EMA). All but (Uaniher) two Gram-positive bacteria (Staphylococcus aureus (MRSA) and Staphylococcus pseudintermedius)sensitive to the antimicrobial ingredient present is the CFCM of the wild-type X. szentirmaii DSM 16338 (EMC). Abbreviations: -: the test organism grew in the given CFCM, which is resistant to the antimicrobials present; +:: the test organism did not succeed in the given CFCM, which is sensitive to the antimicrobials present; +/-: the test organism grew at least one in one of the replicates, the given CFCM, that may be resistant to the antimicrobials present.
Table 5.
Intraspecific variability within the Gram-positive Paeibacillus larvae concerning semsitivities toward antimicrobial active compound of the CFCM of wild-types hfq (del) regulatory mutants strains of Xenorhabdus szentirmaii (EMC) EPB species.
Table 5.
Intraspecific variability within the Gram-positive Paeibacillus larvae concerning semsitivities toward antimicrobial active compound of the CFCM of wild-types hfq (del) regulatory mutants strains of Xenorhabdus szentirmaii (EMC) EPB species.
| TEST ORGANISMS | EMA (WT) | EMA (hfq-del) | EMC (WT) | EMC hfq-del) |
|---|---|---|---|---|
| GRAM POSITIVE | ||||
| Paenibaillus larvae isolates | ||||
| ML-234 | - | + | + | + |
| ML-236 | - | + | + | + |
| ML-237 | - | + | + | + |
| ML-238 | - | - | + | + |
| ML-239 | - | + | + | + |
| ML-248 | - | - | - | + |
| ML-250 | - | - | - | + |
| ML-252 | - | - | - | + |
| ML-263 | - | - | - | + |
*It can be seen that each of the tested geographical isolates of the Gram-positive, (deadly pathogen for honeybee larvae) Paeibacillus larvae [88] (Ebeling et l., 2016) bacterium species seems to be completely sensitive to (one or more) antimicrobial compounds present in the CFCM of t wild-type Xenorhabdus budapestensis DSM16342 (EMA). Six of the nine isolates, however, seem also to be sensitive to the antimicrobial active compound(s) present in the CFCM of the hfq (del) regulatory mutant of the same species (EMA). About half of them (mainly those which are resistant to the antimicrobials of the hfq (del) regulatory EMA mutant) proved also resistant to the wild-type EMC CFCM., but each of them is resistant to the antimicrobials present in the CFCM of the hfq /del) a regulatory mutant of EMC.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



