Submitted:
11 July 2023
Posted:
12 July 2023
You are already at the latest version
Abstract
The rise of antibiotic-resistant strains of clinically important pathogens is a major threat to global health security. The World Health Organization (WHO) has recognized the urgent need to develop alternative treatments to address the growing list of priority pathogens. Antimicrobial peptides (AMPs) rank among the various suggested options with proven activity and a high potential to be developed into effective agents. Many AMPs are naturally produced by living organisms and protect the host against pathogens as a part of their innate immunity. Various mechanisms associated with their antimicrobial actions include cell membrane disruption, cell wall weakening, protein synthesis inhibition, and interference in nucleic acid dynamics that can induce apoptosis and necrosis. Acinetobacter baumannii is considered a critical pathogen, and severe clinical problems have appeared with isolates resistant to current antibiotic treatments and conventional control procedures such as UV light, disinfectants, and drying. Here, we review the natural AMPs representing the primary candidates for new anti-A. baumannii drugs in a post-antibiotic era and present the use of computational tools to develop the next generation of AMPs with greater microbicidal activity and reduced toxicity.
Keywords:
AMP
; Acinetobacter baumannii
; resistance
; action mechanism
1. Introduction
The rise in antibiotic resistance is a major contributor to global mortality statistics and represents a pressing challenge across various sectors of society, including healthcare providers, government agencies, and the pharmaceutical industry, that also impinges on environmental concerns. The inability to develop new antibiotics to hinder the rapid rise in the emergence of drug-resistant pathogens suggests that the world is heading toward a post-antibiotic era [1,2]. For bacteria, three types of antimicrobial resistance have been described as intrinsic, acquired, and or adaptive resistance through changes in phenotype [3,4,5,6,7,8,9,10,11]. While many mechanisms lead to resistance, the exposure of microbes to antimicrobial drugs can trigger their evolution, which can be accelerated by the incorrect use of antibiotics through wrong choices, inadequate dosing, and poor adherence to treatment guidelines that all contribute to the selection of antimicrobial resistance [12,13].
There are several terminologies in the literature used to describe microbes based on the number of classes of antibiotics to which they are resistant. According to Magiorakos et al. (2012), a multidrug-resistant (MDR) strain shows resistance to at least one antimicrobial in more than three classes of antimicrobials, and extensively drug-resistant (XDR) strains display resistance to at least one antimicrobial in all classes of antimicrobials except two or fewer types while a pan drug-resistant (PDR) strain is resistant to all antimicrobial agents [14]. In hospital settings, ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, A. baumannii, Pseudomonas aeruginosa, Enterobacter species) describe the most opportunistic pathogens in nosocomial infections that "escape" the effects of antibiotics and conventional therapies that account for increased morbidity and mortality for improved resource utilization in healthcare [15,16].
Notably, Acinetobacter baumannii has globally emerged as a concerning hospital-related pathogen, frequently presenting MDR, XDR, and PDR phenotypes. Unfortunately, evidence has shown increased antibiotic resistance over time [17]. It is a gram-negative, non-motile, non-fermentative, and non-sporulated bacterium of the Moraxellaceae family [18], part of the Acinetobacter calcoaceticus–A. baumannii complex (Acb) that has long been associated with human disease [19]. Consisting of the species A. calcoaceticus, A. baumannii, A. pittii, A. nosocomialis, A. seifertii, and A. lactucae (a later heterotypic synonym of A. dijkshoorniae) [20,21], they differ in epidemiology, pathogenicity, and antimicrobial resistance [22]. While their genetic and physiological relatedness makes them difficult to distinguish phenotypically with standard laboratory methods [23], A. baumannii is the most widespread in hospitals causing various infections, including wounds, skin and urinary tracts, and diseases such as pneumonia, meningitis, and bacteremia [24,25]. All contribute to longer hospital stays, higher treatment costs, and increased morbidity and mortality risks [26].
Treatment options have proven limited for A. baumannii due to its extended virolome and resistome, evasion of host immune effectors, survival in extreme environmental conditions, growth in biofilms, and latent growth forms with a minimal metabolic rate [27,28]. The World Health Organization (WHO) recently highlighted the resistance of A. baumannii to carbapenems (CRAb) [29,30], which included its classification as a "priority 1 for research and develop new antibiotic treatments". As a "critical" pathogen [31], AMP have a high potential for research and development of anti-Acinetobacter drugs [32,33]. In this review, we sought to update knowledge about the potential and activity of antimicrobial peptides (AMP) to act against multidrug-resistant A. baumannii.
2. Antimicrobial peptides
Also known as host defense peptides, AMPs are naturally produced by living organisms as a part of their innate immune system against pathogens. They are amphipathic molecules of varying molecular weights containing 11-50 amino acids with an overall positive electric charge [34,35], classified as either α-helical, β-sheet, or extended peptides [36,37,38]. AMPs are essential in regulating immune processes such as inflammation and activating and recruiting immune system cells [34]. In addition, they can inhibit protein and nucleic acid synthesis that can, cause apoptosis and lead to necrosis [39,40]. Their activities begin with interactions with cell membranes through electrostatic interactions.
As polycationic peptides, their multiple positive amino acids drive electrostatic interactions with lipid membranes that are also influenced by hydrophobic interactions (Figure 1). Due to the inherent differences in the surface composition between bacteria and mammalian cells, there can be differences in the preferential association of AMPs with a cell surface, leading to an accumulation at the surface and self-assembly reaching a particular concentration [41,42]. At this stage, various models have been proposed to describe the mechanism of action (MOA) of AMPs.
Multiple MOAs have been proposed for AMPs during interactions with bacterial cell surfaces generalized as transmembrane pore and non-pore models (Figure 2). Within the pore model, the forms are differentiated as barrel-stave and toroidal, reflecting the bilayer's net arrangement. The barrel-stave shape preserves the bilayer organization and begins with AMPs oriented parallel to the surface before perpendicularly inserting into the lipid bilayer [43]. The amphipathic structure of α and/or β sheet peptides permits lateral peptide-peptide interactions between the hydrophilic amino acids to form the lumen, and the hydrophobic regions interact with bilayer lipids [44,45], which organize similarly to a protein ion channel (Figure 2A). A minimum length of 22 residues in an α-helical structure or eight residues in a β sheet is needed to span a lipid bilayer. Only a subset of known AMPs, such as alamethicin [46], pardaxin [47,48], and protegrins [43], have been shown to form barrel stave channels.
Toroidal pores also result from the perpendicular insertion of AMPs into the lipid bilayer but do not display lateral peptide-peptide interactions [46]. Rather, peptides disrupt the hydrophobic/hydrophilic arrangement of the bilayer and induce a local curvature in the lipid bilayer (Figure 2B). Pores are formed from a dynamic interaction between the inserted peptides and phospholipid head groups that create transient lipid-peptide supramolecules. In toroidal pores, the disruption in the hydrophobic and hydrophilic arrangement of the bills is temporary. Upon disintegration, some peptides are translocated to the inner cytoplasmic leaflet allowing cell entry to target intracellular components [49]. Several AMPs, such as magainin 2 [50], lacticin Q [50], aurein 2.2 [51], and melittin [46,50], have been shown to form toroidal pores. For aurein 2.2, lipid composition and thickness have been shown to influence pore formation [52,53]. In a 1:1 mixture of 1-palmitoyl-2-oleoyl-sn-glycerol-3-phospho-(1-rac-glycerol) with 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine toroidal pores are formed. In a membrane model of 1:1 1,2-dimyristoyl-sn-glycero-3-phospho-(1-rac-glycerol) with 1,2-dimyristoyl-sn-glyce rol-3-phosphocholine, aurein 2.2 does not form discrete pores. Other features of the toroidal pore include ion and size selectivity [54]. Both toroidal and barrel-stave pores ultimately lead to membrane depolarization and cell death.
The carpet model describes AMPs that do not insert into the lipid bilayer to form pores [50,54,55,56]; peptides adsorb to the cell surface (Figure 2C). Upon reaching a threshold concentration, membrane integrity is compromised by a detergent-like effect that leads to the formation of micelles (Figure 2D). As the results in the carpet model are not dependent on specific amino acid compositions, lengths, or interactions, it can describe the MOA of several AMPs at high concentrations due to their amphiphilic nature, such as cecropin [57], indolicidin [58], aurein 1.2 [56], and LL-37 [55]. It has been suggested that the carpet-like mechanism is a prerequisite step for the toroidal pore model [51]. Other models have been proposed, including interfacial activity, electroporation, and Shai-Huang-Matsazuki models [51]. However, in most cases, the results from model membrane systems. Only a few AMPs have been studied in whole bacterial cells to define their MOAs [59,60], suggesting that the results from model membranes describe potential MOAs and may need to explain their actions against pathogens fully.
Many AMPs are currently being studied to describe their therapeutic efficacy against A. baumannii strains. We have curated the online antimicrobial peptide database, APD3, to list many examples of AMPs under study (Table 1 and Table 2). These include both peptides produced by living organisms and novel peptides inspired by their activities.
2.1. Cathelicidins
Cathelicidins are a group of more than 30 cationic AMPs (CAMPs) identified in the immune system of several vertebrates. Their structure comprises two domains involved in their antimicrobial activity [195,196]. In comparison to the broad-spectrum antibiotics imipenem and meropenem of the carbapenem class that is often chosen to treat MDR A. baumannii (MIC = 16–32 mg/L) [197], these peptides have shown good activity.
2.1.1. Humans
The human cathelicidin LL-37 has an α-helical structure and is produced as a component of the innate immune response. It exhibits a broad-spectrum microbicidal activity against Gram-positive and Gram-negative bacteria observed as plasma-membrane disruption [198]. It also neutralizes lipopolysaccharide (LPS) and modulates the immune response through cellular activation, inflammation regulation, chemotaxis, and wound healing [61,199,200,201,202]. Against A. baumannii, LL-37 and its fragments KS-30 and KR-12 showed activity against a sensitive and four MDR clinical isolates [62]. LL-37 inhibited all sensitive and drug-resistant strains at concentrations between 16–32 μg/ml, while the minimum inhibitory concentration (MIC) for KS-30 and KR-12 was 8–16 and 128–256 μg/ml, respectively. In biofilms, LL-37 and the fragment KS-30 were observed to significantly inhibit and disperse the A. baumannii on abiotic surfaces at 32 and 64 μg/ml, respectively [62].
Against the ESKAPE pathogens P. aeruginosa, A. baumannii, and S. aureus, a group of synthetic peptides based on LL-37 AM showed potent microbicidal activity without selecting resistance and could eliminate persistent cells and biofilms at micromolar concentrations [63]. One peptide, SAAP-148, is an α-helical AMP that could inhibit the growth of A. baumannii MDR and prevent biofilm formation at a concentration of 6 μg/ml. An ex vivo human skin infection model and an in vivo murine skin infection model eliminated acute and biofilm-related infections at concentrations of ~5% [63]. Its anti-biofilm activity improved when incorporated into nanoparticles of Poly(lactic-co-glycolic) (PLGA) that gradually increased over time, suggesting a sustained local release of the peptide based on the dose-effect in vitro profile [64].
Another synthetic derivative of LL-37, P10, is cationic, which showed stronger activity than LL-37 [143,203]. Also, based on LL-37, the de novo pepD2 was designed with a trigonal distribution of positive charges in its helical structure and displayed a MIC value of 8 µg/mL against the ATCC strain of A. baumannii. WLBU2 (also called PLG0206) is an engineered cationic amphipathic α-helix derived from the LL-37 peptide that can insert into bacterial membranes leading to cell death as well as potent antibacterial effects against biofilms of MDR A. baumannii and K. pneumoniae [125]. MIC values for WLBU2 were reported to be 1.5-3.2 μM for an XDR A. baumannii [204], 7.484 μM for clinical isolates [128], and 7.943 μM for K. pneumoniae.
2.1.2. Snake
Various cathelicidins have been identified from snakes. Cathelicidin-BF (Cath-BF) was isolated from the venous glands of banded krait (Bungatus fasciatus) [205]. It is one of the best-known cathelicidins and presents an α-helical structure. Two mechanisms attributed to its antimicrobial activity: disrupting bacteria membranes and directly targeting intracellular targets [205]. It has proven highly active against drug-resistant clinical isolates of A. baumannii and can inhibit growth at a concentration of 12.8 μg/ml [64]. A disulfide bridge stabilized cyclic peptide derivative of Cath-BF, ZY4, also displayed activity against clinical MDR isolates of A. baumannii with MIC values ranging between 4.6 and 9.4 μg/mL. It killed bacteria by membrane permeabilization and could inhibit biofilm formation [65]. With a half-life of 1.8h in vivo, ZY4 displayed good stability and a low tendency to induce resistance. NA-CATH has an N-terminal α-helical structure with an unstructured C-terminal [66]. Identified in Chinese cobra (Naja atra) [206], it can inhibit the growth of drug-resistant strains of A. baumannii at 10 µg/ml [206]. Its antimicrobial MOA appears to be through membrane deformation and the formation of transient pores [207]. From the king cobra (Ophiophagus hannah), OH-CATH was identified [67]. Its analog, DOH-CATH30, exhibits microbicidal activity against MDR A. baumannii with a MIC of 1.56 to 12.5 μg/mL.
2.1.3. Alligator
The antibacterial activity of serum from American alligators (Alligator mississippiensis) can be attributed to the presence of CAMPs, and several have been identified [208]. AM-CATH36 inhibited the growth of both drug-resistant and sensitive A. baumannii at 2.5 µg/ml, while its two fragments, AM-CATH28 and AM-CATH21, inhibited at 10 µg/ml [66]. All three appear to permeabilize bacterial membranes. MDR clinical isolates appeared more sensitive to the fragments than full length. The recently identified As-CATH8 displayed in vitro activity profiles similar to the last-resort vancomycin and polymyxin B antibiotics. In a murine abscess model of high-density bacterial infections, As-CATH8 demonstrated good activity against A. baumannii (MIC = 0.6 µg/mL) and another ESKAPE pathogen, S. aureus [142].
2.1.4. Wallaby
WAM-1 is a cathelicidin present in the milk of marsupials and isolated from the mammary gland of Tammar wallaby (Macropus eugenii) with antimicrobial activity [74,209]. In studies on biofilms, WAM-1 inhibited biofilm formation in clinical isolates and could disperse 24h-old biofilms of most isolates tested that included MDR strains [75]. In comparison to LL-37, WAM-1 shows several desirable properties. It's in vitro activity was 12 to 80 times more effective than LL-37 at eliminating clinical isolates of A. baumannii, and its activity as a peptide is not reduced in the presence of total serum or high NaCl concentrations. While its MOA is unknown, it does not lead to hemolysis and has the potential for in vivo applications [75].
2.1.5. Hoofed animals
Domesticated animals have yielded several cathelicidins. The cytoplasmic granules of the bovine neutrophils contain indolicidin, a short tryptophan-rich cationic peptide that displaces divalent cations on the surface of cell membranes, forms pores and can inhibit DNA processing enzymes [78,210,211,212,213]. It displays potent anti-A. baumannii activity with a MIC of 4–32 for sensitive clinical isolates and 16 μg/ml against colistin-resistant strains [76]. In combination in vitro with antimicrobial agents, the MIC of indolicidin against 12 MDR clinical isolates was reported to be between 2–64 μg/ml [78]. Bactenecin is a cyclic, arginine-rich cationic AMP isolated from cows, sheep, and goats with a type I β-turn structure and a disulfide bond between cysteines at positions 3 and 11 [77,78]. It can permeabilize cell membranes and inhibit RNA and protein synthesis with a MIC of 16 and 64 µg/ml against sensitive and colistin-resistant A. baumannii, respectively [59,76]. Other studies of cathelicidins include bovine BMAP-27, sheep SMAP29, and goat minibactenecins [73], which have been shown to inhibit the growth of clinical MDR A. baumannii strains [68].
2.2. Defensins
Animals, plants, and fungi produce an ancient class of AMPs called defensins that contain six to eight conserved cysteine residues. Their MOA includes binding cell membranes and forming pores that kill pathogens [214]. Defensins have been categorized into α, β, and θ-defensins subfamilies [215].
2.2.1. Human α-Defensins
The CAMPs HNP-1 and HNP-2 are α-defensins produced in human neutrophils that differ by their N-terminal amino acid. They are components of the human neutrophil peptides contained in polymorphonuclear neutrophil granules that are released for secretion upon activation to microbes [216]. The standard strain of A. baumannii (ATCC 19606) was affected by concentrations of 50 μg/ml, while a colistin-resistant strain appeared more sensitive (MIC = 3.25 μg/ml) [76]. HD5, another human defensin, has little effect on A. baumannii effect (MIC = 320 μg/ml), buts its derivative HD5d5 showed a lower MIC (40 μg/ml) through cell membrane damage and cell entry that reduced the activities of superoxide dismutase and catalase [217,218].
2.2.2. β-Defensins
HBD-2 and HBD-3 are human β-defensins found in the epithelial lining of the respiratory system and urinary tracts. Interestingly, they appear more effective against MDR clinical isolates [219]. Another β-defensins, HBD-3, combines an α-helical segment with the β strand and can kill both non-MDR and MDR strains of A. baumannii in serum-free conditions [220]. It also displayed wound healing properties and has a potential application in wound dressings [83,221]. In A. mississippiensis, the AM23sk isoform of its β-Defensin showed in vitro antibacterial activity against A. baumannii (MIC = 2 μg / ml) [142].
2.2.3. Insect defensins
The insect defensin CL-defensin can partially permeabilize A. baumannii and, different from others, is predicted to have an N-terminal loop, an α-helix segment, and an antiparallel β-sheet according to circular dichroism spectroscopy [81].
2.3. Frog AMP
2.3.1. Magainin and pexiganan
The skin of the African clawed frog (Xenopus laevis) has two α-helical cationic amphipathic AMPs, Magainin-1 and 2 [222]. Their primary MOA against microbes is pore formation [85,223]. Magainin-2 shows higher activity against MDR strains of A. baumannii at 4.9–64 μg/ml and can both inhibit and eliminate biofilms [76,85]. It also offers greater stability in physiological conditions and low hemolytic activity. Interestingly, it shows anticancer potential with low toxicity against non-cancerous mammalian cells [85]. A synthetic analog of Magainin-2, Pexiganan, or MSI-78 also displays a broad potent action against the formation of toroidal pores [224,225,226]. Against A. baumannii, pexiganan can inhibit the growth of MDR clinical isolates at a concentration of 1–8 μg/ml [87,88,227]. Studies with the ATCC 196060 reference strain of A. baumannii confirmed pexiganan's antimicrobial and anti-biofilm activity [89].
2.3.2. Brevinin-2 related peptide
Skin secretions from both the mink frog (Rana septentrionalis) and carpenter frog (R. virgatipes) contain B2RP, a brevinin-related AMP with an α-helical structure that affects the organization of bacterial membranes [228,229]. It can inhibit sensitive strains of A. baumannii at a concentration of 29 μg/ml and MDR strains at 7–13.9 μg/ml [90]. However, its hemolytic properties limit its potential use [230]. Three analogs of B2RP (D4K, K16A, and L18K) showed reduced red blood cell toxicity and a two-fold increase in activity against A. baumannii growth. [90,91]. The D4K substitution also showed activity against colistin-resistant and XDR clinical isolates [92]. B2RP-ERa is a smaller cationic peptide structurally similar to B2RP found in the skin of the Asian frog (Hylarana erythraea) [93, 231]. It can inhibit sensitive Acinetobacter strain growth at concentrations of 8–32 and drug-resistant strains 8–64 μg/ml, respectively [91]. It shows anti-inflammatory characteristics without toxicity on peripheral blood mononuclear cells or red blood cells [231,232].
2.3.3. Alyteserins
Alyteserin-1c and Alyteserin-2a are two of the alyteserin class of cationic AMPs with anti-A. baumannii activity released in skin secretions following norepinephrine stimulation of the midwife toad (Alytes obstetricans) [94,95,233]. Against clinical isolates of MDR A. baumannii, Alyteserin-1c inhibited growth and caused death at concentrations between 11.3–22.6 μg/ml with low hemolytic activity [94]. The substitution E4K reduced effects on red blood cells further while improving growth inhibition of colistin-resistant and XDR isolates of A. baumannii isolates [92]. Structural changes of Alyteserin-2a also resulted in an analog with a 4-8 fold greater antimicrobial activity and lower hemolytic effects [95].
2.3.4. Peptide glycine-leucine-amide
The volcano-clawed frog (Xenopus amieti) produces PGLa-AM1, peptide glycine-leucine-amide, an AMP with anti-Acinetobacter AMP activity. It can kill sensitive and colistin-resistant A. baumannii isolates at 16–128 μg/ ml [91]. With low hemolytic activity, it is also active against other ESKAPE pathogens, including E. coli and S. aureus [91,93,100].
2.3.5. Caerulein precursor fragment
Also isolated from the volcano-clawed frog, caerulein precursor fragment (CPF-AM1) is a cationic AMP that binds LPS [92,93]. It inhibits the growth of sensitive and colistin-resistant strains with minimal fibroblast toxicity and hemolytic activity [101]. CPF-B1 was isolated from the Marsabit clawed frog (Xenopus borealis) that displays anti-A. baumannii activity at concentrations between 11.4–22.8 μg/ml with low hemolysis [99]. From the Peracca clawed frog (Xenopus clivii), CPF-C1 is another member of the caerulein family of peptides with proven activity against A. baumannii with inhibitory activity at concentrations as low as 5 μg/ml concentration [98].
2.3.6. Hymenochirins
Hymenochirin-1B was isolated from the Zaire Dwarf Clawed Frog (Hymenochirus boettgeri) and is the first member of hymenochirins class of AMPs of their host defense system [234,235]. It is an α-helical cationic peptide that can inhibit the growth of MDR isolates of A. baumannii properties with a MIC of 19.1 μg/ml [100]. In addition to its antimicrobial activity, it displays anticancer and immunomodulatory properties. Hymenochirin-1B, which was generated by E6K and D9K substitutions, showed a nearly 4-fold increase in activity against A. baumannii, including MDR and XDR isolates with reduced toxicity to human erythrocytes [100]. Hymenochirin-1Pa was isolated from Merlin's dwarf gray frog (Pseudhymenochirus merlini) that inhibited the growth of XDR clinical isolates of A. baumannii at a concentration between 7.5–15 μg/ml. However, it also showed moderate hemolytic activity [101, 235].
2.3.7. XT-7
Norepinephrine stimulation of the western clawed frog (Xenopus tropicalis) permitted isolation from skin secretions XT-7 [236], an AMP with anti-Acinetobacter activity against the Euroclone I NM8 strain with a MIC of 22.2 μg/ml [98]. A substitution, G4K, increased its therapeutic index [102], inhibiting sensitive and drug-resistant A. baumannii strains at concentrations as low as 4 μg/ml [91].
2.3.8. Buforins
The stomach of the Asian toad (Bufo gargarizans) yielded Burfoin I [237]. Its derivative, Buforin II, is a potent antimicrobial peptide that kills bacteria by crossing the membrane to bind intracellular targets, including DNA and RNA that inhibit cellular activities [103]. Against Acinetobacter, Buforin II can hinder the growth of sensitive and resistant isolates at concentrations between 0.25–39 μg/ml [84,85]. In combination with antibiotic treatments or alone, it showed good potential in a rat model of sepsis with A. baumannii [91].
2.3.9. Caerin 1.1 and 1.9
The host defense peptides caerin 1.1 and caerin 1.9 of the Australian tree frog(Litoria caerulea) were isolated from skin secretions. They are α-helical cationic amphipathic AMP with antiviral, antitumor, antimicrobial, and neuropeptide-type activities [238]. Each displayed anti-A. baumannii growth activity that was additive in combination [141].
2.3.10. Hyalin a1
Hylin a1 is an α-helical cationic amphipathic AMP isolated from the skin secretions of the white spotted tree frog (Hypsiboas albopunctatus) [239]. Its antimicrobial activity has been attributed to its action on bacterial membranes, but it also displays a strong hemolytic activity. Two analogs, Hylin a1-11K and Hylin a1-15K, showed good antimicrobial activity against carbapenem-resistant A. baumannii clinical isolates at 1–2 µM with no report on changes to the hemolytic activity [148].
2.4. Fish piscins
Fish possess a strong innate immune system that is the first line of defense against a broad spectrum of pathogens [240]. Various antimicrobial components can be found within the epidermal mucus, including AMPs, lysozyme, proteases, and lectins [241]. Piscidins are cationic AMPs expressed by fish mast cells [242], which comprise a family of structurally related mature peptides between 21 and 44 residues. They possess an amphipathic α-helical structure, which suggests that piscins have bactericidal activities against various microorganisms [243]. The piscidin family includes pleurocidin, moronecidin, chrysophsin, dicentracin, epinecidin-1, and myxinidin [244].
Pleurocidin is an amphipathic α-helical cationic peptide found in the gills, gut, and on the skin of winter flounder (Pseudopleuronectes americanus) [245], which is genetically similar to piscidin [246]. It displays a broad-spectrum activity against pathogenic bacteria and fungi such as K. pneumoniae, S. aureus, P. aeruginosa, and the opportunistic oral pathogen C. albicans [245,247]. Against MDR isolate strains of A. baumannii, this peptide can inhibit growth at concentrations between 16 and 8–32 [68]. Its MOA appears to be through membrane disruption through its binding [248], although it shows lower hemolysis compared with other natural peptides by in vitro toxicity studies [249].
Tilapia piscidin 2 (TP2) is an inactive antibacterial peptide found in Nile tilapia (Oreochromis niloticus) [250], which was modified to develop the peptides TP2-5 and TP2-6 that improved cationic and amphipathic balance [251]. The changes resulted in a significant improvement in their antimicrobial potential in normal media against the A. baumannii wild-type strains (MIC = 3.1 μg/ml) and MDR isolates (MIC = 1.6-12.5 μg/ml) [181]. Another AMP in Nile tilapia (TP4) displayed antimicrobial activity against sensitive and MDR A. baumannii strains between 16-32 μg/ml [68].
2.5. Hepcidin
First identified in human blood ultrafiltrate and urine samples and called a liver-expressed antimicrobial peptide (LEAP-1) [252,253], hepcidin is a cationic amphipathic peptide that functions in multiple vertebrates. It has been reported to be involved in iron metabolism, inflammation, and clearance of invading pathogens [254]. Since the first fish hepcidin was reported in hybrid striped bass in 2002 [255], many isoforms have been found across numerous fish species. Unlike a single gene in humans, many teleost fish have more than two hepcidin genes, most notably in Perciformes and Pleuronectiformes [256]. Fish hepcidin isoforms are currently phylogenetically classified into two groups, HAMP1-type and HAMP2-type [257,258,259]. From Japanese seabass (Lateolabrax japonicus), the peptide LJ-hep2 has been investigated using its recombinant precursor protein (rLJ-hep2) expressed in Pichia pastoris and a chemically synthesized mature peptide LJ-hep2(66–86) with LJ-hep2(66–86) displaying a stronger antimicrobial activity against several clinically isolated MDR A. baumannii (MIC = 1.5-3 μg/ml) [160].
2.6. Melittin
The cationic amphipathic α-helical AMP melittin was isolated from the European honeybee (Apis mellifera) 's venom, comprising nearly half its dry weight [260]. Numerous properties have been reported, including antibacterial [260], antiparasitic [261], and antifungal [262], along with anticancer and antiviral properties [263]. Its primary MOA is a carpet-like interaction with membranes, leading to pore formation and lysis [264]. Melittin displays potent antimicrobial activity against MDR and XDR clinical isolates of Acinetobacter at concentrations as low as 0.125 μg/ml [105,106]. In a mouse model of third-degree burns, topical application of melittin at 16 µg/ml concentrations eliminated 93.3% of an XDR isolate of A. baumannii, and doubling the dosage stopped the bacteria [105]. Importantly, the injured derma and surrounding tissue, including red blood cells, showed no toxicity. Brazilian clinical studies confirmed the activity of melittin against most strains except for one pan-drug-resistant strain [265].
2.7. Cecropins
Cecropin describes a class of AMPs with a primary MOA attributed to membrane lysis [268]. The founding member, cecropin A, was isolated from the hemolymph of the giant silk moth (Hyalophora cecropia) [269]. Initial results showed in vitro antibacterial and anticancer activity [270]. Viability studies in a Caenorhabditis elegans model for A. baumannii infections demonstrated that 15 cecropin or cecropin-like peptides displayed antimicrobial activity and improved survival [108]. Several studies have further defined the growth inhibition of individual peptides, including cecropin A against colistin-resistant strains and MDR clinical isolates [76,107], BR003-cecropin A from Aedes aegypti against MDR A. baumannii isolates [108], Musca domestica cecropin (Mdc) from housefly (Musca domestica) larvae against standard and MDR isolates [109], cecropin-4 from houseflies against MDR and XDR clinical isolates [151,152,153], cecropin P1 from pig roundworms (Ascaris suum) against colistin-sensitive A. baumannii [76]. Many cecropins also display anti-biofilm activity, such as myxinidin isolated from hagfish (Myxine glutinosa) [111] and the AMP complex FLIRP7 (Fly Larvae Immune Peptides 7) in blowfly (Calliphora vicina) larvae [112].
The fusion of cecropin A to endolysin ST01 has been shown to have increased bactericidal activity against ESKAPE pathogens, with A. baumannii (ATCC 17978) eliminated at a concentration of 0.25 [271]. Another hybrid of cecropin with melittin, CAMEL, rapidly kills A. baumannii [89]. OMN6 is a 40-amino acid synthetic cyclic peptide based on cecropin A that displays an increase in stability and a significant decrease in proteolytic degradation and low cytotoxicity against eukaryote cells. This peptide exerts a rapid bactericidal effect by causing the selective disruption of the bacterial membrane integrity [272], which is effective on A. baumannii laboratory strains (MIC = 8 μg/ml) and clinical isolates (MIC = 4-8 μg/ml) suggesting a low likelihood for the development of resistance [158].
2.8. Mastoparan
Mastoparan was isolated from the venom of hornets (Vespula lewisii) [218]. While it displays good activity against wild-type A. baumannii and colistin-resistant and pan-resistant clinical isolates [76,273], it also shows high hemolytic activity that would prevent its therapeutic applications [274]. Activity against MDR clinical isolates of A. baumannii was observed with mastoparan-AF isolated from the venom of another hornet (Vespa affinis) with a MIC between 2-16 μg/ml [109]. Improvements in serum stability (24h) have been achieved in analogs of mastoparan that maintain the growth inhibition of XDR clinical isolates [76]. Greater therapeutic efficacy against MDR clinical isolates has been acquired by conjugating mastoparan with chitin to form nanoconstructs [Afreenish hassan]. Improvements in hemolytic toxicity have not been reported.
2.9. Histatins
Histatins are a family of low-molecular-weight, histidine-rich cationic peptides isolated from salivary glands, which display antimicrobial activity through membrane disruption [222]. The only member that affects A. baumannii is histatin-8, a hemagglutination-inhibiting peptide [230]. It could inhibit the growth of colistin-sensitive and -resistant strains at a concentration of 32 μg/ml [76].
2.10. Dermcidin
The DCD gene in humans encodes dermcidin, an anionic AMP with two regions produced and secreted by eccrine sweat glands for transport to the skin surface [117,275]. While the N-terminal peptide is involved with neuronal cell survival in response to oxidative stress [117], the C-terminal fragment shows anti-Acinetobacter activity [276]. With a net charge of -2, DCD-1L can interact with negatively charged bacterial phospholipids. PDR clinical isolates of A. baumannii isolates show a two-fold increase in susceptibility than XDR isolates and the standard ATCC 19606 strain [118]. DCD-1L can also inhibit bacterial attachment and biofilm formation, which could affect infection initiation [118].
2.11. Tachyplesin III
The hemolymph of the Southeast Asian horseshoe crabs (Tachypleu gigas and Carcinoscorpius rotundicauda) contains tachyplesin III, an AMP of 17 amino acids. As opposed to an α-helical structure, this peptide presents a cyclic β-sheet with two disulfide bridges. Against an XDR clinical isolate of A. baumannii, tachyplesin III had a measured MIC of 8–16 μg/ml and could fully eliminate the bacteria at twice the MIC concentration [277]. However, it also displays high toxicity against mammalian cells, preventing therapeutic applications [277].
2.12. Spider peptides
Various AMPs have been isolated from the venom of spiders. Lycosin-I is a 23-amino acid peptide from the venom of the Chinese wolf spider (Lycosa singoriensis) that had MIC against MDR isolates of A. baumannii between 8-32 µg/ml [140,278]. In the venom of ant spiders (Lachesana tarabaevi), latarcins 2a also displayed potent antimicrobial activity against MDR clinical isolates A. baumannii at concentrations between 8–64 μg/ml [68]. Similar to Lycosin-I, LS-AMP-E1 and LS-AMP-F1 from the burrowing wolf spider (Lycosa sinensis) had different inhibitory activity against other clinical drug-resistant bacteria and could effectively inhibit the formation of biofilms with no obvious hemolytic effects. Among the ESKAPE pathogens, LS-AMP-F1 was the most effective against A. baumannii, with the lowest MIC of 3.1 µM [140]. LyeTx I was isolated from a wolf spider from Brazil (Lycosa erythrognatha) and showed inhibitory activity against several multidrug-resistant bacteria but also showed hemolytic and cytotoxic effects. Conjugating a derivative, LyeTx I-b, to PEG could eliminate these contradictory effects while maintaining the MIC values against A. baumannii, anti-biofilm formation, and did not induce resistance [155].
2.13. Scorpion
Many AMPs have been identified in the venom of scorpions that display antimicrobial activity against A. baumannii, such as Hp1404, ctriporin, and Im5 [144]. In many instances, these peptides also show harmful effects, such as hemolysis, that alterations in their sequences could remedy. Hp1404 was isolated from the venomous gland of the giant forest scorpion (Heterometrus petersii) and is an amphipathic α-helical peptide that exhibited antimicrobial activity against methicillin-resistant S. aureus along with cytotoxicity. Many analogs showed lower cytotoxic activity against MDR isolates of A. baumannii [145]. BmKn2 is another naturally occurring cationic α-helical AMP from the Chinese scorpion (Mesobuthus martensii Karsch) with antimicrobial and strong hemolytic activity. It merely shows against gram-positive bacteria, such as S. aureus. Its mutant BmKn2-7 has lower hemolytic activity and maintains a broadened antimicrobial spectrum [279]. Another analog, BmKn2-7K, is non-toxic at antimicrobial dosages and exhibits potent antimicrobial activity via a membrane-lytic mechanism against a series of clinically isolated antibiotic-resistant ESKAPE pathogens. For MDR A. baumannii, BmKn2-7K and BmKn2-7R (MIC = 2.5–5 µg/ml) showed potent and improved antimicrobial activity than that of BmKn2-7 (MIC = 5–10 µg/ml) [147].
2.14. Lynronne-1
Lynronne-1 is an α-helical cationic amphipathic peptide identified using a metagenomics investigation of the bovine rumen microbiome for the presence of novel AMPs. Although it is in vivo activity was lower than conventional antibiotics, it showed selectivity for bacterial cells with low hemolytic activity and minimal cytotoxicity against mammalian cells [280]. Against some common gram-positive and gram-negative pathogenic bacteria, Lynronne-1 displayed broad-spectrum activity, including methicillin-resistant S. aureus (MRSA) (MIC of 8–32 μg/mL) and A. baumannii (MIC of 4 μg/mL) [280].
2.15. Hybrid peptides
The combination of different AMPs offers a rational approach to developing non-natural AMP. The peptide PapMA consists of 18 amino acids that combine the first eight amino acids from papiliocin, a 37-residue AMP purified from larvae of the swallowtail butterfly (Papilio xuthus) with resides 4-12 of magainin 2, a 23-residue AMP purified from the skin of the African clawed frog (Xenopus laevis). A proline hinge joined the two fragments. While PapMA showed high antimicrobial activity, it was cytotoxic to mammalian cells [281]. The hybrid peptides P7A3 and A3P7 combined cathelicidin (P7) and aurein (A3) were obtained by a flipping technique [282]. The serial truncation of the C-terminal led to an optimal candidate, AP19, that was stabilized against proteolytic enzymes by a D-amino acid substitution (D-AP19). The final peptide rapidly killed A. baumannii ATCC 19.606 (MIC = 7.81 µg/mL) via membrane disruption and showed a low tendency to induce bacterial resistance. It also exhibited potent antibacterial activity against multidrug-resistant (MDR) and extensively drug-resistant (XDR) clinical isolates of A. baumannii (MIC = 3.91-15.63) [189]. BP214 is a cationic amphipathic all-D decapeptide developed from a short cecropin A-melittin hybrid peptide BP100 [283], which showed excellent activity against colistin-resistant A. baumannii and modest hemolytic properties [284].
3. Resistance to AMPS
Resistance to AMPs can be acquired by their degradation, sequestration, impedance by exopolymers or biofilm matrix molecules, alteration of the membrane lipid composition, and export mechanisms [285,286,287,288,289,290,291]. Resistance to colistin has been documented for A. baumannii following its long-term clinical application [292,293]. Resistance was also observed after the inactivation of one of the genes involved in LPS biosynthesis, resulting in loss. As colistin is a last-resort drug to treat MDR nosocomial pathogens, resistance is an important clinical issue [293,294,295]. Several nanocarriers have been developed to overcome low bioavailability, proteolysis, and toxicity associated with AMPs [296,297]. Changes in the molecular structure, modifications of biochemical characteristics, and their combination with common antibiotics have been reported to minimize AMP resistance [287].
4. Conclusions
Of the ESKCAPE pathogens, A. baumannii is a growing concern for nosocomial and community-acquired infections. With a high capacity to acquire resistance and form biofilms, there has been an alarming increase in the loss of antibiotic efficacy and a rise of MDR strains worldwide. Projections combined with the scarcity of new antibiotic treatments [13] show the need to transition to a "post-antibiotic era" by developing new therapeutics based on alternative approaches. AMPs have emerged as excellent candidates due to the breadth of natural peptides found as part of innate immune systems that demonstrate activity against many A. baumannii strains, including MDR and XDR clinical isolates. While many of these AMPs also display undesirable effects such as hemolysis and host toxicity, studies have demonstrated the ability to modify their sequences to improve performance. Future advances in bioinformatics combined with more studies on the sequence/structure/function relationship can generate synthetic AMPs to address a major health concern. Our review of AMPs highlights common characteristics such as cationic, α-helical structure, interactions with bacterial membranes, bilipid pore formation, and intracellular component targeting. Many instances of improved performance combined with traditional treatments and their use as bioconjugates show promise for future applications. In addition to their antimicrobial properties, many AMPs have demonstrated other beneficial activities such as anticancer, antioxidant, wound healing, and angiogenesis that further support additional research [34,61,298,299].
Author Contributions
KR and SGD-S also contributed to the elaboration of the article.
Funding
Please add: This research was funded by Carlos Chagas Filho Foundation for Research Support of the State of Rio de Janeiro/FAPERJ (#110.198-13) and the Brazilian Council for Scientific Research (CNPq, #467.488/2014-2 and 301744/2019-0). Funding was also provided by FAPERJ (#210.003/2018) through the National Institutes of Science and Technology Program (INCT) to Carlos M. Morel (INCT-IDPN).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- O’Neill, J. Tackling drug-resistance infections globally: Final report and recommendations. The Review on Antimicrobial Resistance. London, UK: Government of the United Kingdom; 2016, 84 p.
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new. antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells, dormancy, and infectious disease. Nat Rev Microbiol 2007, 5, 48–56. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat Rev Microbiol 2017 15, 453–464. [CrossRef]
- Fernández, L.; Breidenstein, E.B.M.; Hancock, R.E.W. Importance of adaptive and stepwise changes in the rise and spread of antimicrobial resistance. In: Keen, P.; Monforts, M. editors. Antimicrobial resistance in the environment. Hoboken, New Jersey, EUA: Wiley-Blackwell; 2011. pp. 43-71. ISBN: 978-1-118-15623-0.
- Olivares, J.; Bernardini, A.; Garcia-Leon, G.; Corona, F.; Sanchez, M.B.; Martinez, J.L. The intrinsic resistome of bacterial pathogens. Front Microbiol 2013, 30, 103. [Google Scholar] [CrossRef]
- Lewis, K.; Shan, Y. Persister awakening. Mol Cell 2016, 63, 3–4. [Google Scholar] [CrossRef]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; et al. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef]
- Shan, Y.; Brown Gandt, A.; Rowe, S.E.; Deisinger, J.P.; Conlon, B.P.; Lewis, K. ATP-dependent persister formation in Escherichia coli. mBio 2017, 8, 02267–e02216. [Google Scholar] [CrossRef]
- Magana, M.; Sereti, C.; Ioannidis, A.; Mitchell, C.A.; Ball, A.R.; Magiorkinis, E. Chatzipanagiotou, S.; Hamblin, M.R.; Hadjifrangiskou, M.; Tegos, G.P.; et al. Options and limitations in clinical investigation of bacterial biofilms. Clin Microbiol Ver 2018, 31, 00084–e00016. [Google Scholar] [CrossRef]
- Cameron, D.R.; Shan, Y.; Zalis, E.A.; Isabella, V.; Lewis, K. A genetic determinant of persister cell formation in bacterial pathogens. J Bacteriol 2018, 200, e00303–e00318. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global, multifaceted phenomenon. Pathog Glob Health 2015, 109, 309–318. [Google Scholar] [CrossRef]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pan drug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Rice LB. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE J Infect Dis 2008, 197, 1079-1081. [CrossRef]
- Friedman ND, Temkin E, Carmeli Y. The negative impact of antibiotic resistance. Clin Microbiol Infect. 2016, 22, 416. [CrossRef]
- Xie, R.; Zhang, X.D.; Zhao, Q.; Peng, B.; Zheng, J. Analysis of global prevalence of antibiotic resistance in Acinetobacter baumannii infections disclosed a faster increase in OECD countries. Emerg Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii biofilms: effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect Drug Resist 2018, 11, 2277–2299. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Biswas, I.; Veeraraghavan, B. Accurate identification of clinically important Acinetobacter spp.: An update. Future Sci OA 2019, 5, FSO395. [Google Scholar] [CrossRef]
- Cosgaya, C.; Mari-Almirall, M.; van Assche, A.; Fernandez-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int J Syst Evol Microbiol 2016, 66, 4105–4111. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus–Acinetobacter baumannii complex isolated from human clinical specimens. Int J Syst Evol Microbiol 2015, 63, 934–942. [Google Scholar] [CrossRef]
- Chen, T.L.; Lee, Y.T.; Kuo, S.C.; Yang, S.P.; Fung, C.P.; Lee, S.D. Rapid identification of Acinetobacter baumannii, Acinetobacter nosocomialis, and Acinetobacter pittii with a multiplex PCR assay. J Med Microbiol 2014, 63, 1154–1159. [Google Scholar] [CrossRef]
- Marí-Almirall, M.; Cosgaya, C.; Higgins, P.G.; Van Assche, A.; Telli, M.; Huys, G.; Lievens, B.; Seifert, H.; Dijkshoorn, L.; Roca, I.; et al. MALDI-TOF/MS identification of species from the Acinetobacter baumannii (ab) group revisited: Inclusion of the novel A. seifertii and A. dijkshoorniae species. Clin Microbiol Infect 2017, 23, 210.e1–210.e9. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol 2007, 5, 939–951. [Google Scholar] [CrossRef]
- Garnacho-Montero, J.; Timsit, J.F. Managing Acinetobacter baumannii infections. Curr Opin Infect Dis 2019, 32, 69–76. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef]
- Barth, V.C.J.; Rodrigues, B.Á.; Bonatto, G.D.; Gallo, S.W.; Pagnussatti, V.E.; Ferreira, C.A.S.; de Oliveira, S.D. Heterogeneous persister cells formation in Acinetobacter baumannii. PLoS ONE 2013, 8, e84361. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, B.; Gajic, I.; Dimkic, I.; Kekic, D.; Zornic, S.; Pozder, T.; Radisavljevic, S.; Opavski, N.; Kojic, M.; Ranin, L. The first nationwide multicenter study of Acinetobacter baumannii recovered in Serbia: Emergence of OXA-72, OXA-23 and NDM-1-producing isolates. Antimicrob Resist Infect Control 2020, 9, 101. [Google Scholar] [CrossRef]
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New treatment options against carbapenem-resistant Acinetobacter baumannii infections. Antimicrob Agents Chemother 2019, 63, e01110–e01118. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. Available at: https://www.who. int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf. [Accessed in June 2023].
- Domalaon R, Zhanel GG, Schweizer F. Short antimicrobial peptides and peptide scaffolds as promising antibacterial agents. Curr Top Med Chem 2016, 16, 1217-1230. [CrossRef]
- Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic resistance profiles, molecular mechanisms and innovative treatment strategies of Acinetobacter baumannii. Microorganisms 2020, 8, 935. [Google Scholar] [CrossRef]
- Fan, L.; Sun, J.; Zhou, M.; Zhou, J.; Lao, X.; Zheng, H.; Xu, H. DRAMP: A comprehensive data repository of antimicrobial peptides. Sci Rep 2016, 6, 24482. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sunkara, L.T. Avian antimicrobial host defense peptides: From biology to therapeutic applications. Pharmaceuticals (Basel) 2014, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.; Ortiz, C.; Guzman, F.; Fernandez-Lafuente, R.; Torres, R. Antimicrobial peptides: Promising compounds against pathogenic microorganisms. Curr Med Chem 2014, 21, 2299. [Google Scholar] [CrossRef]
- Govender, T.; Dawood, A.; Esterhuyse, A.J.; Katerere, D.R. Antimicrobial properties of the skin secretions of frogs. S Afr J Sci 2012, 108, 25–30. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim Biophys Acta. 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Andersson DI, Hughes D, Kubicek-Sutherland JZ. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist Updat 2016, 26, 43-57. [CrossRef]
- Ehrenstein, G.; Lecar, H. Electrically gated ionic channels in lipid bilayers. Quarterly Reviews of Biophysics 1977, 10, 1–34. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat Rev Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Breukink, E.; de Kruijff, B. The lantibiotic nisin, a special case or not? Biochim Biophys Acta. 1999, 1462, 223–234. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chem Biol 2010, 5, 905–917. [Google Scholar] [CrossRef]
- Rapaport, D.; Shai, Y. Interaction of fluorescently labeled pardaxin and its analogs with lipid bilayers. J Biol Chem 1991, 266, 23769–23775, PMID: 1748653. [Google Scholar] [CrossRef]
- Shai, Y.; Bach, D.; Yanovsky, A. Channel formation properties of synthetic pardaxin and analogs. J Biol Chem. 1990, 265, 20202–20209, PMID: 1700783. [Google Scholar] [CrossRef]
- Uematsu, N.; Matsuzaki, K. Polar angle as a determinant of amphipathic α-helix-lipid interactions: A model peptide study. Biophys J. 2000, 79, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Lee, T-H.; Hall, K.N.; Aguilar, M-I. Antimicrobial peptide structure and mechanism of action: A focus on the role of membrane structure. Curr TopMed Chem. 2016, 16, 25-39. [CrossRef]
- Cheng, J.T.J.; Hale, J.D.; Elliot, M.; Hancock, R.E.W.; Straus, S.K. Effect of membrane composition on antimicrobial peptides aurein 2.2 and 2.3 from Australian southern bell frogs. Biophys J 2009, 96, 552–565. [Google Scholar] [CrossRef]
- Sparr, E.; Ash, W.L.; Nazarov, P.V.; Rijkers, D.T.S.; Hemminga, M.A.; Tieleman, D.P.; et al. Self-association of transmembrane-helices in model membranes. J Biol Chem. 2005, 280, 39324–39331. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.T.J.; Hale, J.D.; Elliott, M.; Hancock, R.E.W.; Straus, S.K. The importance of bacterial membrane composition in the structure and function of aurein 2.2 and selected variants. Biochim Biophys Acta - Biomembranes 2011, 1808, 622–633. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol Rev 2003, 55, 27–55. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane-active antimicrobial peptides. Biopolymers. 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Fernandez, D.I.; Le Brun, A.P.; Whitwell, T.C.; Sani, M.-A.; James, M.; Separovic, F. The antimicrobial peptide aurein 1.2 disrupts model membranes via the carpet mechanism. Phys Chem Chem Phys 2012, 14, 15739. [Google Scholar] [CrossRef]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochimt Biophys Acta 1999, 1462, 29–54. [Google Scholar] [CrossRef]
- Rozek, A.; Friedrich, C.L.; Hancock, R.E. Structure of the bovine antimicrobial peptide indolicidin bound to dodecyl phosphocholine and sodium dodecyl sulfate micelles. Biochemistry 2000, 39, 15765–15774, PMID: 11123901. [Google Scholar]
- Gee, M.L.; Burton, M.; Grevis-James, A.; Hossain, M.A.; McArthur, S.; Palombo, E.A.; Wade, J.D.; Clayton, A.H. Imaging the action of antimicrobial peptides on living bacterial cells. Sci Rep 2013, 3, 1557. [Google Scholar] [CrossRef]
- Choi, H.; Rangarajan, N.; Weisshaar, J.C. Lights, camera, action! Antimicrobial peptide mechanisms imaging in space and time. Trends Microbiol 2016, 24, 111–122. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Akbari Eidgahi, M.R.; Chichaklu, A.H.; Movaqar, A.; Ghazvini, K. Review of antimicrobial peptides with anti-helicobacter pylori activity. Helicobacter 2019, 24, e12555. [Google Scholar] [CrossRef]
- Feng, X.; Sambanthamoorthy, K.; Palys, T.; Paranavitana, C. The human antimicrobial peptide LL-37 and its fragments possess both antimicrobial and antibiofilm activities against multidrug-resistant Acinetobacter baumannii. Peptides 2013, 49, 131–137. [Google Scholar] [CrossRef]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; De Boer, L.; Koning, R.I.; et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci Transl Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, M.; Akhavan, M.M.; Fallah, F.; Karimi, A. A recombinant snake cathelicidin derivative peptide: Antibiofilm properties and expression in Escherichia coli. Biomolecules 2018, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc Natl Acad Sci USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef]
- Barksdale, S.M.; Hrifko, E.J.; van Hoek, M.L. Cathelicidin antimicrobial peptide from Alligator mississippiensis has antibacterial activity against multi-drug resistant Acinetobacter baumanii and Klebsiella pneumonia. Dev Comp Immunol 2017, 70, 135–144. [Google Scholar] [CrossRef]
- Zhao, F.; Lan, X.Q.; Du, Y.; Chen, P.Y.; Zhao, J.; Zhao, F.; et al. King cobra peptide OH-CATH30 as a potential candidate drug through clinic drug-resistant isolates. Zool Res 2018, 9, 87–96. [Google Scholar] [CrossRef]
- Chiau-Jing, Jung.; You-Di, Liao.; ;Chih-Chieh, Hsu.; Ting-Yu, Huang.; Yu-Chung, C.; Jeng-Wei, C.; Yu-Min, K.; Jean-San, C. Identification of potential therapeutic antimicrobial peptides against Acinetobacter baumannii in a mouse model of pneumonia. Sci Rep 2021, 11, 7318. [CrossRef]
- Dekan, Z.; Headey, S.J.; Scanlon, M.; Baldo, B.A.; Lee, T.-H.; Aguilar, M.-I.; Deuis, J.R.; Vetter, I.; Elliott, A.G.; Amado, M.; et al. ∆-Myrtoxin-Mp1a is a helical heterodimer from the venom of the jack jumper ant that has antimicrobial, membrane-disrupting, and nociceptive activities. Angew Chem Int Ed Engl 2017, 56, 8495–8499. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari AK, Alamri MA, Almasoudi AS, Abbasmanthiri R, Mahfoud M. Evaluation of the in vitro antimicrobial activity of selected Saudi scorpion venoms tested against multidrug-resistant micro-organisms. J Glob Antimicrob Resist 2017,10, 14-18. [CrossRef]
- Domhan, C.; Uhl, P.; Kleist, C.; Zimmermann, S.; Umstätter, F.; Leotta, K.; Mier, W.; Wink, M. Replacement of L-amino acids by d-amino acids in the antimicrobial peptide ranalexin and its consequences for antimicrobial activity and biodistribution. Molecules 2019, 24, 2987. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, R.; Wiesner, J.; Marker, A.; Pfeifer, Y.; Bauer, A.; Hammann, P.E.; Vilcinskas, A. Profiling antimicrobial peptides from the medical maggot Lucilia sericata as potential antibiotics for MDR gram-negative bacteria. J Antimicrob Chemother 2019, 74, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Shamova, O.V.; Orlov, D.S.; Zharkova, M.S.; Balandin, S.V.; Yamschikova, E.V.; Knappe, D.; Hoffmann, R.; Kokryakov, V.N.; Ovchinnikova, T.V. Minibactenecins ChBac7.Nα and ChBac7. Nβ—Antimicrobial peptides from leukocytes of the goat Capra hircus. Acta Naturae 2016, 8, 136-146. PMID: 27795854. [PubMed]
- Wang, J.; Wong, E.S.W.; Whitley, J.C.; Li, J.; Stringer, J.M.; Short, K.R.; Renfree, M.B.; Belov, K.; Cocks, B.G. Ancient antimicrobial peptides kill antibiotic-resistant pathogens: Australian mammals provide new options. PloS One 2011, 6, e24030. [Google Scholar] [CrossRef]
- Spencer, J.J.; Pitts, R.E.; Pearson, R.A.; King, L.B. The effects of antimicrobial peptides WAM-1 and LL-37 on multidrug-resistant Acinetobacter baumannii. Patho Dis 2018, 76, fty007. [Google Scholar] [CrossRef]
- Vila-Farres, X.; De La Maria, C.G.; López-Rojas, R.; Pachón, J.; Giralt, E.; Vila, J. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin Microbiol Infec 2012, 18, 383–387. [Google Scholar] [CrossRef]
- Romeo, D.; Skerlavaj, B.; Bolognesi, M.; Gennaro, R. Structure and bactericidal activity of an antibiotic dodecapeptide purified from bovine neutrophils. J Biol Chem 1988, 263, 9573–9575, PMID: 3290210. [Google Scholar] [CrossRef]
- Falla, T.J.; Karunaratne, D.N.; Hancock, R.E. Mode of action of the antimicrobial peptide indolicidin. J Biol Chem 1996, 271, 19298–19303, PMID: 8702613. [Google Scholar] [CrossRef]
- Wu, M.; Hancock, R.E. Interaction of the cyclic antimicrobial cationic peptide bactenecin with the outer and cytoplasmic membrane. J Biol Chem 1999, 274, 29–35, PMID: 9867806. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, G.; Wang, S.; Chen, Y.; Gong, Y.; Chen, S.; Xu, Y.; Hu, M.; Wang, X.; Zeng, H.; et al. A simplified derivative of human defensin 5 with potent and efficient activity against multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother 2018, 62, e01504–e01517. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Gupta, K.; Van Hoek, M.L. Characterization of Cimex lectularius (bedbug) defensin peptide and its antimicrobial activity against human skin microflora. Biochem Biophys Res Commun 2016, 470, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Routsias, J.G.; Karagounis, P.; Parvulesku, G.; Legakis, N.J.; Tsakris, A. In vitro bactericidal activity of human β-defensin 2 against nosocomial strains. Peptides 2010, 31, 1654–1660. [Google Scholar] [CrossRef]
- Maisetta, G.; Batoni, G.; Esin, S.; Florio, W.; Bottai, D.; Favilli, F.; Campa, M. In vitro bactericidal activity of human beta-defensin 3 against multidrug-resistant nosocomial strains. Antimicrob Agents Chemother 2006, 50, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Del Prete, M.S.; Barchiesi, F.; Paggi, A.M.; Petrelli, E.; Scalise, G. Comparative activities of polycationic peptides and clinically used antimicrobial agents against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. J Antimicrob Chemother 2000, 46, 807–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kang, N.; Ko, S.J.; Park, J.; Park, E.; Shin, D.W.; Kim, S.H.; Lee, S.A.; Lee, J.I.; Lee, S.H.; et al. Antibacterial and antibiofilm activity and mode of action of Magainin 2 against drug-resistant Acinetobacter baumannii. Int J Mol Sci. 2018, 19, 3041. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc Natl Acad Sci U S A. 1987, 84, 5449–53. [Google Scholar] [CrossRef]
- Flamm, R.K.; Rhomberg, P.R.; Simpson, K.M.; Farrell, D.J.; Sader, H.S.; Jones, R.N. In vitro spectrum of pexiganan activity when tested against pathogens from diabetic foot infections and with selected resistance mechanisms. Antimicrob Agents Chemother 2015, 59, 1751–4. [Google Scholar] [CrossRef]
- Ge, Y.; MacDonald, D.L.; Holroyd, K.J.; Thornsberry, C.; Wexler, H.; Zasloff, M. In vitro antibacterial properties of pexiganan, an analog of magainin. Antimicrob Agents Chemother 1999, 43, 782–8. [Google Scholar] [CrossRef]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial activity of selected antimicrobial peptides against planktonic culture and biofilm of Acinetobacter baumannii. Probiotics Antimicrob Proteins. 2019, 11, 317–324. [Google Scholar] [CrossRef]
- Conlon, J.M.; Ahmed, E.; Condamine, E. Antimicrobial properties of brevinin-2-related peptide and its analogs: Efficacy against multidrug-resistant Acinetobacter baumannii. Chem Biol Drug Des. 2009, 74, 488–93. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Sonnevend, A.; Pál, T.; Vila-Farrés, X. Efficacy of six frog skin-derived antimicrobial peptides against colistin-resistant strains of the Acinetobacter baumannii group. Int J Antimicrob Agents. 2012, 39, 317–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.B.; Shan, B.; Bai, H.M.; Tang, J.; Yan, L.Z.; Ma, Y.B. Hydrophilic/hydrophobic characters of antimicrobial peptides derived from animals and their effects on multidrug resistant clinical isolates. Dongwuxue Yanjiu. 2015, 36, 41–7. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghaferi, N.; Kolodziejek, J.; Nowotny, N.; Coquet, L.; Jouenne, T.; Leprince, J.; Vaudry, H.; King, J.D.; Conlon, J.M. Antimicrobial peptides from the skin secretions of the South-East Asian frog Hylarana erythraea (Ranidae). Peptides 2010, 31, 548–54. [Google Scholar] [CrossRef]
- Conlon, J.M.; Ahmed, E.; Pal, T.; Sonnevend, A. Potent and rapid bactericidal action of alyteserin-1c and its [E4K] analog against multidrug-resistant strains of Acinetobacter baumannii. Peptides 2010, 31, 1806–10. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Arafat, K.; Attoub, S.; Sonnevend, A. Analogues of the frog skin peptide alyteserin-2a with enhanced antimicrobial activities against Gram-negative bacteria. J Pept Sci 2012, 18, 270–5. [Google Scholar] [CrossRef]
- McLean DT, McCrudden MT, Linden GJ, Irwin CR, Conlon JM, Lundy FT. Antimicrobial and immunomodulatory properties of PGLa-AM1, CPF-AM1, and magainin-AM1: potent activity against oral pathogens. Regul Pept. 2014, 194-195, 63-8. [CrossRef]
- Conlon, J.M.; Al-Ghaferi, N.; Ahmed, E.; Meetani, M.A.; Leprince, J.; Nielsen, P.F. Orthologs of magainin, PGLa, procaerulein-derived, and proxenopsin-derived peptides from skin secretions of the octoploid frog Xenopus amieti (Pipidae). Peptides. 2010, 31, 989–94. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Ahmed, E.; Leprince, J.; Vaudry, H.; King, J.D.; Takada, K. Purification and properties of antimicrobial peptides from skin secretions of the Eritrea clawed frog Xenopus clivii (Pipidae). Comp Biochem Physiol C Toxicol Pharmacol 2011, ;153, 350-4. [CrossRef]
- Mechkarska, M.; Ahmed, E.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D.; Conlon, J.M. Antimicrobial peptides with therapeutic potential from skin secretions of the Marsabit clawed frog Xenopus borealis (Pipidae). Comp Biochem Physiol C Toxicol Pharmacol. 2010, 152, 467–72. [Google Scholar] [CrossRef]
- Mechkarska, M.; Prajeep, M.; Radosavljevic, G.D.; Jovanovic, I.P.; Al Baloushi, A.; Sonnevend, A.; Lukic, M.L.; Conlon, J.M. An analog of the host-defense peptide hymenochirin-1B with potent broad-spectrum activity against multidrug-resistant bacteria and immunomodulatory properties. Peptides 2013, 50, 153–9. [Google Scholar] [CrossRef]
- Serra, I.; Scorciapino, M.A.; Manzo, G.; Casu, M.; Rinaldi, A.C.; Attoub, S.; Mechkarska, M.; Conlon, J.M. Conformational analysis and cytotoxic activities of the frog skin host-defense peptide, hymenochirin-1Pa. Peptides 2014, 61, 114–21. [Google Scholar] [CrossRef]
- Conlon, J.M.; Galadari, S.; Raza, H.; Condamine, E. Design of potent, non-toxic antimicrobial agents based upon the naturally occurring frog skin peptides, ascaphin-8 and peptide XT-7. Chem Biol Drug Des 2008, 72, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem Biophys Res Commun 1998, 244, 253–7. [Google Scholar] [CrossRef] [PubMed]
- Cirioni, O.; Silvestri, C.; Ghiselli, R.; Orlando, F.; Riva, A.; Gabrielli, E.; Mocchegiani, F.; Cianforlini, N.; Trombettoni, M.M.; Saba, V.; et al. Therapeutic efficacy of buforin II and rifampin in a rat model of Acinetobacter baumannii sepsis. Crit Care Med 2009, 37, 1403–7. [Google Scholar] [CrossRef] [PubMed]
- Pashaei, F.; Bevalian, P.; Akbari, R.; Pooshang Bagheri, K. Single dose eradication of extensively drug resistant Acinetobacter spp. In a mouse model of burn infection by melittin antimicrobial peptide. Microb Pathog 2019, 127, 60–69. [Google Scholar] [CrossRef]
- Akbar, R.; Hakemi-Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly synergistic effects of melittin with conventional antibiotics against multidrug-resistant isolates of Acinetobacter baumannii and Pseudomonas aeruginosa. Microb Drug Resist 2019, 25, 193–202. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; D'Amato, G.; Silvestri, C.; Del Prete, M.S.; Łukasiak, J.; Scalise, G. Comparative activities of cecropin A, melittin, and cecropin A-melittin peptide CA(1-7)M(2-9)NH2 against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. Peptides 2003, 24, 1315–8. [Google Scholar] [CrossRef]
- Jayamani, E.; Rajamuthiah, R.; Larkins-Ford, J.; Fuchs, B.B.; Conery, A.L.; Vilcinskas, A.; Ausubel, F.M.; Mylonakis, E. Insect-derived cecropins display activity against Acinetobacter baumannii in a whole-animal high-throughput Caenorhabditis elegans model. Antimicrob Agents Chemother 2015, 59, 1728–37. [Google Scholar] [CrossRef]
- Gui, S.; Li, R.; Feng, Y.; Wang, S. Transmission electron microscopic morphological study and flow cytometric viability assessment of Acinetobacter baumannii susceptible to Musca domestica cecropin. ScientificWorldJournal 2014, 2014, 657536. [Google Scholar] [CrossRef]
- Boman, H.G.; Agerberth, B.; Boman, A. Mechanisms of action on Escherichia coli of cecropin P1 and PR-39, two antibacterial peptides from pig intestine. Infect Immun. 1993, 61, 2978–84. [Google Scholar] [CrossRef]
- Han, H.M.; Ko, S.; Cheong, M.J.; Bang, J.K.; Seo, C.H.; Luchian, T.; Park, Y. Myxinidin2 and myxinidin3 suppress inflammatory responses through STAT3 and MAPKs to promote wound healing. Oncotarget. 2017, 8, 87582–87597. [Google Scholar] [CrossRef]
- Gordya, N.; Yakovlev, A.; Kruglikova, A.; Tulin, D.; Potolitsina, E.; Suborova, T.; Bordo, D.; Rosano, C.; Chernysh, S. Natural antimicrobial peptide complexes in the fighting of antibiotic resistant biofilms: Calliphora vicina medicinal maggots. PLoS One. 2017, 12, e0173559. [Google Scholar] [CrossRef] [PubMed]
- Vila-Farrés, X.; López-Rojas, R.; Pachón-Ibáñez, M.E.; Teixidó, M.; Pachón, J.; Vila, J.; Giralt, E. Sequence-activity relationship, and mechanism of action of mastoparan analogues against extended-drug resistant Acinetobacter baumannii. Eur J Med Chem. 2015, 101, 34–40. [Google Scholar] [CrossRef]
- Al-Khafaji, Z.; Al-Samaree, M. Design of synthetic antimicrobial peptides against resistant Acinetobacter baumannii using computational approach. Int J Pharmaceut Sci Res. 2017, 8, 2033–2039255. [Google Scholar] [CrossRef]
- Lin, C.H.; Lee, M.C.; Tzen, J.T.C.; Lee, H.M.; Chang, S.M.; Tu, W.C.; Lin, C.F. Efficacy of Mastoparan-AF alone and in combination with clinically used antibiotics on nosocomial multidrug-resistant Acinetobacter baumannii. Saudi J Biol Sci. 2017, 24, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Ikram, A.; Raza, A.; Saeed, S.; Zafar Paracha, R.; Younas, Z.; Khadim, M.T. Therapeutic potential of novel mastoparan-chitosan nanoconstructs against clinical MDR Acinetobacter baumannii: In silico, in vitro and in vivo studies. Int J Nanomedicine 2021, 16, 3755–3773. [Google Scholar] [CrossRef]
- Sağmak Tartar, A.; Özer Balin, Ş.; Akbulut, A.; Yardim, M.; Aydin, S. Roles of dermcidin, salusin-α, salusin-β and TNF-α in the pathogenesis of human brucellosis. Iran J Immunol 2019, 16, 182–189. [Google Scholar] [CrossRef]
- Farshadzadeh, Z.; Pourhajibagher, M.; Taheri, B.; Ekrami, A.; Modarressi, M.H.; Azimzadeh, M.; Bahador, A. Antimicrobial and anti-biofilm potencies of dermcidin-derived peptide DCD-1L against Acinetobacter baumannii: an in vivo wound healing model. BMC Microbiol 2022, 22, 25. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.J.; Kim, M.K.; Park, Y. Comparative antimicrobial activity of Hp404 peptide and its analogs against Acinetobacter baumannii. Int J Mol Sci. 2021, 22, 5540. [Google Scholar] [CrossRef]
- Neshani, A.; Sedighian, H.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Jahangiri, A. Antimicrobial peptides as a promising treatment option against Acinetobacter baumannii infections. Microb Pathog 2020, 146, 104238. [Google Scholar] [CrossRef]
- Morroni, G.; Simonetti, O.; Brenciani, A.; Brescini, L.; Kamysz, W.; Kamysz, E.; Neubauer, D.; Caffarini, M.; Orciani, M.; Giovanetti, E.; et al. In vitro activity of Protegrin-1, alone and in combination with clinically useful antibiotics, against Acinetobacter baumannii strains isolated from surgical wounds. Med Microbiol Immunol 2019, 208, 877–883. [Google Scholar] [CrossRef]
- Mohan, N.M.; Zorgani, A.; Jalowicki, G.; Kerr, A.; Khaldi, N.; Martins, M. Unlocking NuriPep 1653 from common pea protein: a potent antimicrobial peptide to tackle a pan-drug resistant Acinetobacter baumannii. Front Microbiol 2019, 18, 10:2086. [Google Scholar] [CrossRef]
- das Neves, R.C.; Mortari, M.R.; Schwartz, E.F.; Kipnis, A.; Junqueira-Kipnis, A.P. Antimicrobial and antibiofilm effects of peptides from venom of social wasp and scorpion on multidrug-resistant Acinetobacter baumannii. Toxins (Basel) 2019, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Robinson, A.; Davidov, Y.; Shulman, A.; Varvak, A.; Reyes, F.; Rahav, G.; Nissan, I. Delftibactin-A, a non-ribosomal peptide with broad antimicrobial activity. Front Microbiol 2019, 10, 2377. [Google Scholar] [CrossRef] [PubMed]
- Swedan, S.; Shubair, Z.; Almaaytah, A. Synergism of cationic antimicrobial peptide WLBU2 with antibacterial agents against biofilms of multi-drug resistant Acinetobacter baumannii and Klebsiella pneumoniae. Infect Drug Resist 2019, 12, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- 126 Mant, C.; Jiang, Z.; Gera, L.; Davis, T.; Nelson, K.L.; Bevers, S.; Hodges, R.S. De novo designed amphipathic α-helical antimicrobial peptides incorporating dab and dap residues on the polar face to treat the gram-negative pathogen, Acinetobacter baumannii. J Med Chem. 2019, 62, 3354–3366. [Google Scholar] [CrossRef]
- Kirkpatrick, C.L.; Broberg, C.A.; McCool, E.M.; Lee, W.J.; Chao, A.; McConnell, E.W.; Pritchard, D.A.; Hebert, M.; Fleeman, R.; Adams, J.; et al. The "PepSAVI-MS" pipeline for natural product bioactive peptide discovery. Anal Chem. 2017, 89, 1194–1201. [Google Scholar] [CrossRef]
- Eales, M.G.; Ferrari, E.; Goddard, A.D.; Lancaster, L.; Sanderson, P.; Miller, C. Mechanistic and phenotypic studies of bicarinalin, BP100 and colistin action on Acinetobacter baumannii. Res Microbiol. 2018, 169, 296–302. [Google Scholar] [CrossRef]
- Christiansen, S.H.; Murphy, R.A.; Juul-Madsen, K.; Fredborg, M.; Hvam, M.L.; Axelgaard, E.; Skovdal, S.M.; Meyer, R.L.; Sørensen, U.B.S.; Möller, A.; et al. The immunomodulatory drug glatiramer acetate is also an effective antimicrobial agent that kills gram-negative bacteria. Sci Rep 2017, 7, 15653. [Google Scholar] [CrossRef]
- Mahdi, L.; Mahdi, N.; Al-Kakei, S.; Musafer, H.; Al-Joofy, I.; Essa, R.; Zwain, L.; Salman, I.; Mater, H.; Al-Alak, S.; et al. Treatment strategy by lactoperoxidase and lactoferrin combination: Immunomodulatory and antibacterial activity against multidrug-resistant Acinetobacter baumannii. Microb Pathog 2018, 114, 147–152. [Google Scholar] [CrossRef]
- Defraine, V.; Schuermans, J.; Grymonprez, B.; Govers, S.K.; Aertsen, A.; Fauvart, M.; Michiels, J.; Lavigne, R.; Briers, Y. Efficacy of artilysin art-175 against resistant and persistent Acinetobacter baumannii. Antimicrob Agents Chemothe 2016, 60, 3480–8. [Google Scholar] [CrossRef]
- Khan, A.; Xu, M.; Wang, T.; You, C.; Wang, X.; Ren, H.; Zhou, H.; Khan, A.; Han, C.; Li, P. Catechol cross-linked antimicrobial peptide hydrogels prevent multidrug-resistant Acinetobacter baumannii infection in burn wounds. Biosci Rep. 2019, 39, BSR20190504. [Google Scholar] [CrossRef] [PubMed]
- Jenei, S.; Tiricz, H.; Szolomájer, J.; Tímár, E.; Klement, É.; Al Bouni, M.A.; Lima, R.M.; Kata, D.; Harmati, M.; Buzás, K.; et al. Potent chimeric antimicrobial derivatives of the medicago truncatula ncr247 symbiotic peptide. Front Microbiol. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.M.; Rathod, B.B.; Tiricz, H.; Howan, D.H.O.; Al Bouni, M.A.; Jenei, S.; Tímár, E.; Endre, G.; Tóth, G.K.; Kondorosi, É. legume plant peptides as sources of novel antimicrobial molecules against human pathogens. Front Mol Biosci. 2022, 9, 870460. [Google Scholar] [CrossRef]
- Chen, H.L.; Su, P.Y.; Kuo, S.C.; Lauderdale, T.Y.; Shih, C. Adding a c-terminal cysteine (ctc) can enhance the bactericidal activity of three different antimicrobial peptides. Front Microbiol 2018, 9, 1440. [Google Scholar] [CrossRef]
- Cai, Y.; Wang, R.; Liang, B.B.; An, M.M. In-vitro bactericidal activity of colistin against biofilm-associated Pseudomonas aeruginosa and Acinetobacter baumannii. J Hosp Infect 2009, 72, 368–70. [Google Scholar] [CrossRef] [PubMed]
- Thandar, M.; Lood, R.; Winer, BY.; Deutsch, D.R.; Euler, C.W.; Fischetti, V. A. novel engineered peptides of a phage lysin as effective antimicrobials against multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother 2016, 60, 671–9. [Google Scholar] [CrossRef]
- Irani, N.; Basardeh, E.; Samiee, F.; Fateh, A.; Shooraj, F.; Rahimi, A.; Shahcheraghi, F.; Vaziri, F.; Masoumi, M.; Pazhouhandeh, M.; et al. The inhibitory effect of the combination of two new peptides on biofilm formation by Acinetobacter baumannii. Microb Pathog 2018, 121, 310–317. [Google Scholar] [CrossRef]
- Bardbari, AM.; Arabestani, MR.; Karami, M.; Keramat, F.; Aghazadeh, H.; Alikhani, MY.; Bagheri, KP. Highly synergistic activity of melittin with imipenem and colistin in biofilm inhibition against multidrug-resistant strong biofilm producer strains of Acinetobacter baumannii. Eur J Clin Microbiol Infect Dis 2018 Mar;37(3):443-454. [CrossRef]
- Tan, H.; Wang, J.; Song, Y.; Liu, S.; Lu, Z.; Luo, H.; Tang, X. Antibacterial potential analysis of novel α-helix peptides in the chinese wolf spider Lycosa sinensis. Pharmaceutics 2022, 14, 2540. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, P.; Xiao, L.; Liu, Y.; Wu, K.; Ni, G.; Li, H.; Wang, T.; Wu, X.; Chen, G.; et al. Caerin 1.1 and 1.9 peptides from australian tree frog inhibit antibiotic-resistant bacteria growth in a murine skin infection model. Microbiol Spectr 2021, 9, e0005121. [Google Scholar] [CrossRef]
- Santana, F.L.; Arenas, I.; Haney, E.F.; Estrada, K.; Hancock, R.E.W.; Corzo, G. Identification of a crocodylian β-defensin variant from Alligator mississippiensis with antimicrobial and antibiofilm activity. Peptides 2021, 141, 170549. [Google Scholar] [CrossRef]
- Jahangiri, A.; Neshani, A.; Mirhosseini, SA.; Ghazvini, K.; Zare, H.; Sedighian, H. Synergistic effect of two antimicrobial peptides, Nisin and P10 with conventional antibiotics against extensively drug-resistant Acinetobacter baumannii and colistin-resistant Pseudomonas aeruginosa isolates. Microb Pathog 2021, 150, 104700. [Google Scholar] [CrossRef]
- Luo, X.; Ye, X.; Ding, L.; Zhu, W.; Zhao, Z.; Luo, D.; Liu, N.; Sun, L.; Chen, Z. Identification of the scorpion venom-derived antimicrobial peptide Hp1404 as a new antimicrobial agent against carbapenem-resistant Acinetobacter baumannii. Microb Pathog 2021, 157, 104960. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, M.K.; Park, Y. Comparative antimicrobial activity of Hp404 peptide and its analogs against Acinetobacter baumannii. Int J Mol Sci. 2021, 22, 5540. [Google Scholar] [CrossRef]
- Jayawant, E.S.; Hutchinson, J.; Gašparíková, D.; Lockey, C.; Pruñonosa Lara, L.; Guy, C.; Brooks, R.L.; Dixon, AM. Molecular basis of selectivity and activity for the antimicrobial peptide lynronne-1 informs rational design of peptide with improved activity. Chembiochem 2021, 22, 2430–2439. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Swain, S.S.; Behera, A.; Sahoo, G.; Mahapatra, P.K.; Panda, S.K. Antimicrobial peptides derived from insects offer a novel therapeutic option to combat biofilm: a review. Front Microbiol 2021, 10, 661195. [Google Scholar] [CrossRef]
- Park, H.J.; Kang, H.K.; Park, E.; Kim, M.K.; Park, Y. Bactericidal activities and action mechanism of the novel antimicrobial peptide Hylin a1 and its analog peptides against Acinetobacter baumannii infection. Eur J Pharm Sci 2022, 175, 106205. [Google Scholar] [CrossRef]
- Sacco, F.; Bitossi, C.; Casciaro, B.; Loffredo, M.R.; Fabiano, G.; Torrini, L.; Raponi, F.; Raponi, G.; Mangoni, ML. The antimicrobial peptide Esc(1-21) synergizes with colistin in inhibiting the growth and in killing multidrug resistant Acinetobacter baumannii strains. Antibiotics (Basel) 2022, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Bolatchiev, A. Antimicrobial peptides Epinecidin-1 and Beta-Defesin-3 are effective against a broad spectrum of antibiotic-resistant bacterial isolates and increase survival rate in experimental sepsis. Antibiotics (Basel) 2022, 11, 76. [Google Scholar] [CrossRef]
- Ali, M.; van Gent, M.E.; de Waal, A.M.; van Doodewaerd, B.R.; Bos, E.; Koning, R.I.; Cordfunke, R.A.; Drijfhout, J.W.; Nibbering, P.H. Physical and functional characterization of plga nanoparticles containing the antimicrobial peptide SAAP-148. Int J Mol Sci 2023, 24, 2867. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Long, H.; Liu, W.; Wu, Z.; Wang, T.; Zeng, Z.; Guo, G.; Wu, J. Antibacterial mechanism of peptide Cec4 against Acinetobacter baumannii. Infect Drug Resist 2019, 12, 417–2428. [Google Scholar] [CrossRef]
- Peng, J.; Wang, Y.; Wu, Z.; Mao, C.; Li, L.; Cao, H.; Qiu, Z.; Guo, G.; Liang, G.; Shen, F. Antimicrobial peptide Cec4 eradicates multidrug-resistant Acinetobacter baumannii in vitro and in vivo. Drug Des Devel Ther 2023, 30, 977–992. [Google Scholar] [CrossRef] [PubMed]
- Rajapaksha, D.C.; Jayathilaka, E.H.T.T.; Edirisinghe, S.L.; Nikapitiya, C.; Lee, J.; Whang, I.; De Zoysa, M. Octopromycin: Antibacterial and antibiofilm functions of a novel peptide derived from Octopus minor against multidrug-resistant Acinetobacter baumannii. Fish Shellfish Immunol 2021, 117, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.C.M.; Willliam, G.L.; Resende, J.M.; Assis, D.C.S.; Boff, D.; Cardoso, V.N.; Amaral, F.A.; Souza-Fagundes, E.M.; Fernandes, S.O.A.; Lima, M.E. Pegylated LyeTx I-b peptide is effective against carbapenem-resistant Acinetobacter baumannii in an in vivo model of pneumonia and shows reduced toxicity. Int J Pharm 2021, 609, 121156. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Yi, L.; Xu, J.; Gao, W.; Xu, C.; Chen, S.; Chan, KF.; Wong, KY. Investigation of antibiofilm activity, antibacterial activity, and mechanistic studies of an amphiphilic peptide against Acinetobacter baumannii. Biochim Biophys Acta Biomembr. 2021, 1863, 183600. [Google Scholar] [CrossRef] [PubMed]
- Howan, D.H.O.; Jenei, S.; Szolomajer, J.; Endre, G.; Kondorosi, É.; Tóth, G.K. Enhanced antibacterial activity of substituted derivatives of NCR169C peptide. Int J Mol Sci 2023, 24, 2694. [Google Scholar] [CrossRef] [PubMed]
- Mandel, S.; Michaeli, J.; Nur, N.; Erbetti, I.; Zazoun, J.; Ferrari, L.; Felici, A.; Cohen-Kutner, M.; Bachnoff, N. OMN6 a novel bioengineered peptide for the treatment of multidrug resistant Gram-negative bacteria. Sci Rep 2021, 11, 6603. [Google Scholar] [CrossRef]
- Michaeli, J.; Mandel, S.; Maximov, S.; Zazoun, J.; Savoia, P.; Kothari, N.; Valmont, T.; Ferrari, L.; Duncan, LR.; Hawser, S.; et al. In vitro and in vivo antimicrobial activity of the novel peptide OMN6 against multidrug-resistant Acinetobacter baumannii. Antibiotics (Basel) 2022, 11, 1201. [Google Scholar] [CrossRef]
- Gong, R.; An, Z.; Zhang, W.; Chen, F.; Wang, K.J. The antimicrobial peptide lj-hep2 from Lateolabrax japonicus exerting activities against multiple pathogenic bacteria and immune protection in vivo. Mar Drugs 2022, 20, 651. [Google Scholar] [CrossRef]
- Gorr, S.U.; Brigman, H.V.; Anderson, J.C.; Hirsch, E.B. The antimicrobial peptide DGL13K is active against drug-resistant gram-negative bacteria and sub-inhibitory concentrations stimulate bacterial growth without causing resistance. PLoS One 2022, 17, e0273504. [Google Scholar] [CrossRef]
- Moreno-Morales, J.; Guardiola, S.; Ballesté-Delpierre, C.; Giralt, E.; Vila, J. A new synthetic protegrin as a promising peptide with antibacterial activity against MDR Gram-negative pathogens. J Antimicrob Chemother 2022, 77, 3077–3085. [Google Scholar] [CrossRef]
- Li, J.; Prats-Ejarque, G.; Torrent, M.; Andreu, D.; Brandenburg, K.; Fernández-Millán, P.; Boix, E. In vivo evaluation of ecp peptide analogues for the treatment of Acinetobacter baumannii infection. Biomedicines 2022, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Hua, X.; Zhou, R.; Cui, J.; Wang, T.; Kong, F.; You, H.; Liu, X.; Adu-Amankwaah, J.; Guo, G.; et al. Antimicrobial and anti-inflammatory activities of MAF-1-derived antimicrobial peptide Mt6 and its D-enantiomer D-Mt6 against Acinetobacter baumannii by targeting cell membranes and lipopolysaccharide interaction. Microbiol Spectr 2022, 10, e0131222. [Google Scholar] [CrossRef] [PubMed]
- Jayathilaka, E.H.T.T.; Rajapaksha, D.C.; Nikapitiya, C.; De Zoysa, M.; Whang, I. Antimicrobial and anti-biofilm peptide octominin for controlling multidrug-resistant Acinetobacter baumannii. Int J Mol Sci 2021, 22, 5353. [Google Scholar] [CrossRef]
- Jayathilaka, E.H.T.T.; Nikapitiya, C.; De Zoysa, M.; Whang, I. Antimicrobial peptide octominin-encapsulated chitosan nanoparticles enhanced antifungal and antibacterial activities. Int J Mol Sci 2022, 23, 15882. [Google Scholar] [CrossRef] [PubMed]
- Hacioglu, M.; Oyardi, O.; Bozkurt-Guzel, C.; Savage, PB. Antibiofilm activities of ceragenins and antimicrobial peptides against fungal-bacterial mono and multispecies biofilms. J Antibiot (Tokyo) 2020, 73, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, F.W.; Wang, W.X.; Zhao, Y.Y.; Sun, S.Y.; Yu, J.H.; Vitek, M.P.; Li, G.F.; Ma, R.; Wang, S.; et al. Apolipoprotein E mimetic peptide COG1410 combats pandrug-resistant Acinetobacter baumannii. Front Microbiol 2022, 13, 934765. [Google Scholar] [CrossRef]
- Denardi, L.B.; de Arruda Trindade, P.; Weiblen, C.; Ianiski, LB.; Stibbe, PC.; Pinto, SC.; Santurio, J.M. In vitro activity of the antimicrobial peptides h-Lf1-11, MSI-78, LL-37, fengycin 2B, and magainin-2 against clinically important bacteria. Braz J Microbiol 2022, 53, 171–177. [Google Scholar] [CrossRef]
- Eshtiaghi, S.; Nazari, R.; Fasihi-Ramandi, M. Molecular docking, anti-biofilm & antibacterial activities and therapeutic index of mCM11 peptide on Acinetobacter baumannii strains. Curr Microbiol 2023, 80, 191. [Google Scholar] [CrossRef] [PubMed]
- Farzi, N.; Oloomi, M.; Bahramali, G.; Siadat, S.D.; Bouzari, S. Antibacterial properties and efficacy of LL-37 fragment GF-17D3 and scolopendin A2 peptides against resistant clinical strains of Staphylococcus aureus, Pseudomonas aeruginosa, and Acinetobacter baumannii in vitro and in vivo model studies. Probiotics Antimicrob Proteins 2023, 6. [Google Scholar] [CrossRef]
- Dalla Torre, C.; Sannio, F.; Battistella, M.; Docquier, J.D.; De Zotti, M. Peptaibol analogs show potent antibacterial activity against multidrug resistant opportunistic pathogens. Int J Mol Sci 2023, 24, 7997. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Hamed, M.I.; Panitch, A.; Seleem, M.N. Targeting methicillin-resistant Staphylococcus aureus with short salt-resistant synthetic peptides. Antimicrob Agents Chemother. 2014, 58, 4113–22. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.F.; Brezden, A.; Mohammad, H.; Chmielewski, J.; Seleem, M.N. A short D-enantiomeric antimicrobial peptide with potent immunomodulatory and antibiofilm activity against multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Sci Rep. 2017, 7, 6953. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Z.; Li, X.; Fan, Y.; He, G.; Wan, Y.; Yu, C.; Tang, J.; Li, M.; Zhang, X.; rt, al. In vitro and in vivo activities of antimicrobial peptides developed using an amino acid-based activity prediction method. Antimicrob Agents Chemother 2014, 58, 5342–9. [Google Scholar] [CrossRef]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug Des Devel Ther 2017, 11, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Z.; Tian, Y.; Yin, Q.; Cheng, X.; Lian, M.; Zhou, B.; Zhang, X.; Yang, L. Efficacy of antimicrobial peptide DP7, designed by machine-learning method, against methicillin-resistant Staphylococcus aureus. Front Microbiol. 2019, 10, 1175. [Google Scholar] [CrossRef]
- Nagarajan, D.; Roy, N.; Kulkarni, O.; Nanajkar, N.; Datey, A.; Ravichandran, S.; Thakur, C.T.S.; Aprameya, I.V.; Sarma, S.P.; Chakravortty, D.; et al. Ω76: A designed antimicrobial peptide to combat carbapenem- and tigecycline-resistant Acinetobacter baumannii. Sci Adv 2019, 5, eaax–1946. [Google Scholar] [CrossRef]
- Chen, S.P.; Chen, E.H.; Yang, S.Y.; Kuo, P.S.; Jan, H.M.; Yang, T.C.; Hsieh, M.Y.; Lee, KT.; Lin, C.H.; Chen, R.P. A systematic study of the stability, safety, and efficacy of the de novo designed antimicrobial peptide PepD2 and its modified derivatives against Acinetobacter baumannii. Front Microbiol 2021, 12, 678330. [Google Scholar] [CrossRef]
- Krishnan, M.; Choi, J.; Jang, A.; Yoon, YK.; Kim, Y. Antiseptic 9-Meric peptide with potency against carbapenem-resistant Acinetobacter baumannii infection. Int J Mol Sci 2021, 22, 12520. [Google Scholar] [CrossRef]
- Hazam, P.K.; Cheng, C.C.; Hsieh, C.Y.; Lin, W.C.; Hsu, P.H.; Chen, T.L.; Lee, Y.T.; Chen, J.Y. Development of bactericidal peptides against multidrug-resistant Acinetobacter baumannii with enhanced stability and low toxicity. Int J Mol Sci 2022, 23, 2191. [Google Scholar] [CrossRef]
- Ramalho, S.R.; de Sardi, J.C.O.; Júnior, E.C.; Marchetto, R.; Wender, H.; Vargas, L.F.P.; de Miranda, A.; Almeida, C.V.; de Oliveira Almeida, L.H.; de Oliveira, C.F.R.; et al. The synthetic antimicrobial peptide IKR18 displays anti-infectious properties in Galleria mellonella in vivo model. Biochim Biophys Acta Gen Subj 2022, 1866, 130244. [Google Scholar] [CrossRef]
- Gopal, R.; Kim, Y.G.; Lee, J.H.; Lee, S.K.; Chae, J.D.; Son, B.K.; Seo, C.H.; Park, Y. Synergistic effects and antibiofilm properties of chimeric peptides against multidrug-resistant Acinetobacter baumannii strains. Antimicrob Agents Chemother. 2014, 58, 1622–9. [Google Scholar] [CrossRef]
- Heulot, M.; Jacquier, N.; Aeby, S.; Le Roy, D.; Roger, T.; Trofimenko, E.; Barras, D.; Greub, G.; Widmann, C. The anticancer peptide TAT-RasGAP317-326 exerts broad antimicrobial activity. Front Microbiol 2017, 8, 994. [Google Scholar] [CrossRef] [PubMed]
- Vizzarro, G.; Jacquier, N. In vitro synergistic action of TAT-RasGAP317-326 peptide with antibiotics against Gram-negative pathogens. J Glob Antimicrob Resist 2022, 31, 295–303. [Google Scholar] [CrossRef]
- Heinonen, T.; Hargraves, S.; Georgieva, M.; Widmann, C.; Jacquier, N. The antimicrobial peptide TAT-RasGAP317-326 inhibits the formation and expansion of bacterial biofilms in vitro. J Glob Antimicrob Resist 2021, 25, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Taheri, B.; Mohammadi, M.; Momenzadeh, N.; Farshadzadeh, Z.; Roozbehani, M.; Dehghani, P.; Hajian, S.; Darvishi, S.; Shamseddin, J. Substitution of lysine for isoleucine at the center of the nonpolar face of the antimicrobial peptide, piscidin-1, leads to an increase in the rapidity of bactericidal activity and a reduction in toxicity. Infect Drug Resist. 2019, 12, 1629–1647. [Google Scholar] [CrossRef]
- Choi, J.; Jang, A.; Yoon, Y.K.; Kim, Y. development of novel peptides for the antimicrobial combination therapy against carbapenem-resistant Acinetobacter baumannii Infection. Pharmaceutics 2021, 13, 1800. [Google Scholar] [CrossRef]
- Jariyarattanarach, P.; Klubthawee, N.; Wongchai, M.; Roytrakul, S.; Aunpad, R. Novel D-form of hybrid peptide (D-AP19) rapidly kills Acinetobacter baumannii while tolerating proteolytic enzymes. Sci Rep 2022, 12, 15852. [Google Scholar] [CrossRef] [PubMed]
- Andersen, I.K.L.; Thomsen, T.T.; Rashid, J.; Bobak, T.R.; Oddo, A.; Franzyk, H.; Løbner-Olesen, A.; Hansen, PR. C-locked analogs of the antimicrobial peptide BP214. Antibiotics (Basel) 2022, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Lapuebla, A.; Landman, D.; Quale, J. In vitro and in vivo activity of a novel antisense peptide nucleic acid compound against multidrug-resistant Acinetobacter baumannii. Microb Drug Resist 2019, 25, 961–965. [Google Scholar] [CrossRef]
- Mourtada, R.; Herce, H.D.; Yin, D.J.; Moroco, J.A.; Wales, T.E.; Engen, J.R.; Walensky, L.D. Design of stapled antimicrobial peptides that are stable, nontoxic and kill antibiotic-resistant bacteria in mice. Nat Biotechnol 2019, 37, 1186–1197. [Google Scholar] [CrossRef]
- Sharma, D.; Choudhary, M.; Vashistt, J.; Shrivastava, R.; Bisht, G.S. Cationic antimicrobial peptide and its poly-N-substituted glycine congener: Antibacterial and antibiofilm potential against A. baumannii. Biochem Biophys Res Commun 2019, 518, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Ostorhazi, E.; Hoffmann, R.; Herth, N.; Wade, J.D.; Kraus, C.N.; Otvos, L., Jr. Advantage of a narrow spectrum host defense (antimicrobial) peptide over a broad spectrum analog in preclinical drug development. Front Chem 2018, 6, 359. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Duan, H.; Li, M.; Xu, W.; Wei, L. Characterization and mechanism of action of amphibian-derived wound-healing-promoting peptides. Front Cell Dev Biol 2023, 11, 1219427. [Google Scholar] [CrossRef]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: family of antimicrobial peptides. A review. Mol Biol Rep 2012, 39, 10957–70. [Google Scholar] [CrossRef] [PubMed]
- Dowzicky, M.J.; Chmelařová, E. Antimicrobial susceptibility of gram-negative and gram-positive bacteria collected from eastern Europe: results from the tigecycline evaluation and surveillance trial (T. E.S.T.), 2011-2016. J Glob Antimicrob Resist. 2019, 17, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Björstad, A.; Askarieh, G.; Brown, K.L.; Christenson, K.; Forsman, H.; Onnheim, K.; Li, HN.; Teneberg, S.; Maier, O.; Hoekstra, D.; et al. The host defense peptide LL-37 selectively permeabilizes apoptotic leukocytes. Antimicrob Agents Chemother 2009, 53, 1027–38. [Google Scholar] [CrossRef] [PubMed]
- De, Y.; Chen, Q.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37 the neutrophil granule-and epithelial cell-derived cathelicidin utilizes formyl peptide receptor-like 1 (FPRL1) as a receptor to chemoattract human peripheral blood neutrophils monocytes and T cells. J Exp Med 2000, 192, 1069–1074. [Google Scholar] [CrossRef]
- Nijnik, A.; Hancock, R.E.W. Host defense peptides: Antimicrobial and immunomodulatory activity and potential applications for tackling antibiotic-resistant infections. Emerg Health Threats J 2009, 2, e1. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Eidgahi, M.R.A.; Kakhki, R.K.; Safdari, H.; Khaledi, A.; Ghazvini, K. LL-37: A review of antimicrobial profile against sensitive and antibiotic-resistant human bacterial pathogens. Gene Rep 2019, 17, 100519. [Google Scholar] [CrossRef]
- Esfandiyari, R.; Halabian, R.; Behzadi, E.; Sedighian, H.; Jafari, R.; Fooladi, A.A.I. Performance evaluation of antimicrobial peptide ll-37 and hepcidin and β-defensin-2 secreted by mesenchymal stem cells. Heliyon 2019, 5, e02652. [Google Scholar] [CrossRef]
- Haisma, E.M.; de Breij, A.; Chan, H.; van Dissel, J.T.; Drijfhout, J.W.; Hiemstra, P.S.; El Ghalbzouri, A.; Nibbering, P.H. LL-37-derived peptides eradicate multidrug-resistant Staphylococcus aureus from thermally wounded human skin equivalents. Antimicrob Agents Chemother 2014, 58, 4411–4419. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Mietzner, T.A.; Montelaro, R.C. Rational design of engineered cationic antimicrobial peptides consisting exclusively of arginine and tryptophan, and their activity against multidrug-resistant pathogens. Antimicrob Agents Chemother 2013, 57, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Shan, B.; Qi, J.; Ma, Y. Systemic responses of multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii following exposure to the antimicrobial peptide cathelicidin-BF imply multiple intracellular targets. Front Cell Infect Microbiol 2017, 7, 466. [Google Scholar] [CrossRef]
- Zhao, H.; Gan, T.X.; Liu, X.D.; Jin, Y.; Lee, W.H.; Shen, J.H.; et al. Identification and characterization of novel reptile cathelicidins from elapid snakes. Peptides 2008, 29, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Samuel, R.L.; Massiah, M.A.; Gillmor, S.D. The structure and behavior of the NA-CATH antimicrobial peptide with liposomes. Biochim Biophy Acta 2015, 1848, 2394–2405. [Google Scholar] [CrossRef]
- Bishop, B.M.; Juba, M.L.; Devine, M.C.; Barksdale, S.M.; Rodriguez, C.A.; Chung, M.C.; et al. Bioprospecting the American alligator (Alligator mississippiensis) host defense peptidome. PLoS One 2015, 10, e0117394. [Google Scholar] [CrossRef]
- Daly, K.A.; Digby, M.R.; Lefévre, C.; Nicholas, K.R.; Deane, E.M.; Williamson, P. Identification characterization and expression of cathelicidin in the pouch young of Tammar wallaby (Macropus eugenii). Comp Biochem Physiol Part B, Biochem Mol Biol 2008, 149, 524-533. [CrossRef]
- Selsted, M.E.; Novotny, M.J.; Morris, W.L.; Tang, Y.Q.; Smith, W.; Cullor, J.S. Indolicidin is a novel bactericidal tridecapeptide amide from neutrophils. J Biol Chem 1992, 267, 4292–4295. [Google Scholar] [CrossRef]
- Végh, A.G.; Nagy, K.; Bálint, Z.; Kerényi, A.; Rákhely, G.; Váró, G.; Szegletes, Z. Effect of antimicrobial peptide-amide: Indolicidin on biological membranes. J Biomed Biotechnol 2011, 2011, 670589. [Google Scholar] [CrossRef]
- Hsu, C.H.; Chen, C.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide indolicidin: Evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res 2005, 33, 4053–4064. [Google Scholar] [CrossRef]
- Marchand, C.; Krajewski, K.; Lee, H.F.; Antony, S.; Johnson, A.A.; Amin, R.; Kvaratskhelia, M.; Pommier, Y. Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res 2006, 34, 5157–5165. [Google Scholar] [CrossRef]
- Knutelski, S.; Awad, M.; Łukasz, N.; Bukowski, M.; Śmiałek, J.; Suder, P.; Dubin, G.; Mak, P. Isolation, identification, and bioinformatic analysis of antibacterial proteins and peptides from immunized hemolymph of red palm weevil Rhynchophorus ferrugineus. Biomolecules 2021, 11, 83. [Google Scholar] [CrossRef]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schäfer-Korting, M.; Korting, H.C. Human defensins. J Mol Med(Berlin, Germany) 2005, 83, 587–595. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Lu, W. α-Defensins in human innate immunity. Immunol Rev 2012, 45, 84–112. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, G.; Wang, S.; Chen, Y.; Gong, Y.; Chen, S.; Xu, Y.; Hu, M.; Wang, X.; Zeng, H.; et al. A simplified derivative of human defensin 5 with potent and efficient activity against multidrug-resistant Acinetobacter baumannii. Antim Agents Chemoth 2018, 62, e01504–e01517. [Google Scholar] [CrossRef] [PubMed]
- Wanniarachchi, Y.A.; Kaczmarek, P.; Wan, A.; Nolan, E.M. Human defensin 5 disulfide array mutants: Disulfide bond deletion attenuates antibacterial activity against Staphylococcus aureus. Biochemistry 2011, 37, 8005–8017. [Google Scholar] [CrossRef] [PubMed]
- Marcelino-Pérez, G.; Ruiz-Medrano, R.; Gallardo-Hernández, S.; Xoconostle-Cázares, B. Adsorption of recombinant human β-defensin 2 and two mutants on mesoporous silica nanoparticles and its effect against Clavibacter michiganensis subsp. Michiganensis. Nanomaterials (Basel) 2021, 11, 8–2144. [Google Scholar] [CrossRef] [PubMed]
- Schibli, D.J.; Hunter, H.N.; Aseyev, V.; Starner, T.D.; Wiencek, J.M.; McCray, P.B.; Tack, B.F.; Vogel, H.J. The solution structures of the human beta-defensins lead to a better understanding of the potent bactericidal activity of HBD3 against Staphylococcus aureus. J Biol Chem 2002, 277, 8279–8289. [Google Scholar] [CrossRef]
- Hirsch, T.; Spielmann, M.; Zuhaili, B.; Fossum, M.; Metzig, M.; Koehler, T.; et al. Human beta defensin-3 promotes wound healing in infected diabetic wounds. J Gene Med 2009, 11, 220–228. [Google Scholar] [CrossRef]
- Zerweck, J.; Strandberg, E.; Kukharenko, O.; Reichert, J.; Bürck, J.; Wadhwani, P.; Ulrich, A.S. Molecular mechanism of synergy between the antimicrobial peptides PGLa and magainin 2. Sci Rep 2017, 7, 13153. [Google Scholar] [CrossRef]
- Tamba, Y.; Yamazaki, M. Magainin 2-induced pore formation in the lipid membranes depends on its concentration in the membrane interface. J Physical Chem B 2009, 113, 486–4852. [Google Scholar] [CrossRef]
- Maloy, W.L.; Kari, U.P. Structure-activity studies on magainins and other host defense peptides. Biopolymers 1995, 37, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Gottler, L.M.; Ramamoorthy, A. Structure membrane orientation mechanism and function of pexiganan-a highly potent antimicrobial peptide designed from magainin. Biochim Biophys Acta 2009, 1788, 1680–1686. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, A.; Thennarasu, S.; Lee, D.K.; Tan, A.; Maloy, L. Solid-state NMR investigation of the membrane-disrupting mechanism of antimicrobial peptides MSI-78 and MSI-594 derived from magainin 2 and melittin. Biophysical J 2006, 91, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, P.C.; Barry, A.L.; Brown, S.D. In vitro antimicrobial activity of MSI-78 a magainin analog. Antim Agents Chemoth 1998, 42, 1213–1216. [Google Scholar] [CrossRef]
- Bevier, C.R.; Sonnevend, A.; Kolodziejek, J.; Nowotny, N.; Nielsen, P.F.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin secretions of the mink frog (Rana septentrionalis). Comp Biochem Physiol C Toxicol Pharmacol 2004, 139, 31–38. [Google Scholar] [CrossRef]
- Conlon, J.M.; Abraham, B.; Sonnevend, A.; Jouenne, T.; Cosette, P.; Leprince, J.; et al. Purification and characterization of antimicrobial peptides from the skin secretions of the carpenter frog Rana virgatipes (Ranidae, Aquarana). Reg Peptides 2005, 13, 38–45. [Google Scholar] [CrossRef]
- Savelyeva, A.; Ghavami, S.; Davoodpour, P.; Asoodeh, A.; Los, M.J. An overview of Brevinin superfamily: Structure function and clinical perspectives. Adv Exp Med Biol 2014, 818, 197–212. [Google Scholar] [CrossRef]
- Popovic, S.; Urbán, E.; Lukic, M.; Conlon, J.M. Peptides with antimicrobial and anti-inflammatory activities that have therapeutic potential for treatment of acne vulgaris. Peptides 2012, 34, 275–282. [Google Scholar] [CrossRef]
- Popovic, S.; Djurdjevic, P.; Zaric, M.; Mijailovic, Z.; Avramovic, D.; Baskic, D. Effects of host defense peptides B2RP Brevinin-2GU D-Lys-Temporin Lys-XT-7 and DLys-Ascaphin-8 on peripheral blood mononuclear cells: Preliminary study. Periodicum Biologorum 2017, 119, 113–118. [Google Scholar] [CrossRef]
- Conlon, J.M.; Demandt, A.; Nielsen, P.F.; Leprince, J.; Vaudry, H.; Woodhams, D.C. The alyteserins: Two families of antimicrobial peptides from the skin secretions of the midwife toad Alytes obstetricans (Alytidae). Peptides 2009, 30, 1069–1073. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res 2015, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Prajeep, M.; Mechkarska, M.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D. Characterization of the host-defense peptides from skin secretions of Merlin's clawed frog Pseudhymenochirus merlini: Insights into phylogenetic relationships among the Pipidae. Comp. Biochem. Physiol. Part D Genomic Proteonomics 2013, 8, 352–357. [Google Scholar] [CrossRef]
- Ali, M.F.; Soto, A.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides isolated from skin secretions of the diploid frog Xenopus tropicalis (Pipidae). Biochim Biophys Acta 2001, 1550, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from Bufo bufo gargarizans. Biochem Biophy Res Commun 1996, 218, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Apponyi, M.A.; Pukala, T.L.; Brinkworth, C.S.; Maselli, V.M.; Bowie, J.H.; Tyler, M.J.; Booker, G.W.; Wallace, J.C.; Carver, J.A.; Separovic, F. Host-defence peptides of Australian anurans: structure, mechanism of action and evolutionary significance. Peptides 2004, 25, 1035–1054. [Google Scholar] [CrossRef]
- Castro, M.S.; Ferreira, T.C.G.; Cilli, E.M.; Crusca JR, E.; Mendes-Giannini, M.J.S.; Sebben, A.; Ricart, C.A.O.; Sousa, M.V.; Fontes, W. Hylin a1, the first cytolytic peptide isolated from the arboreal South American frog Hypsiboas albopunctatus (“spotted treefrog”). Peptides 2009, 30, 291–296. [Google Scholar] [CrossRef]
- Subramanian, S.; Ross, N.W.; Mackinnon, S.L. Comparison of the biochemical composition of normal epidermal mucus and extruded slime of hagfish (Myxine glutinosa L.). Fish Shellfish Immunol 2008, 25, 625–632. [Google Scholar] [CrossRef]
- Ellis, A.E. Innate host defense mechanisms of fish against viruses and bacteria. Dev Comp Immunol 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Silphaduang, U.; Noga, E.J. Peptide antibiotics in mast cells of fish. Nature 2001, 414, 268–269. [Google Scholar] [CrossRef]
- Noga, E.J.; Silphaduang, U.; Park, N.G.; Seo, J.K.; Stephenson, J.; Kozlowicz, S. Piscidin 4, a novel member of the piscidin family of antimicrobial peptides. Comp Biochem Physiol B Biochem Mol Biol 2009, 152, 299–305. [Google Scholar] [CrossRef]
- Peng, K.C.; Lee, S.H.; Hour, A.L.; Pan, C.Y.; Lee, L.H.; Chen, J.Y. Five diferent piscidins from Nile tilapia, Oreochromis niloticus: analysis of their expressions and biological functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J Biol Chem 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Tong, Z.; Lin, Y.; Xue, Y.; Wang, W.; Kuang, R.; Wang, P.; Tian, Y.; Ni, L. Antimicrobial and antibiofilm activity of pleurocidin against cariogenic microorganisms. Peptides 2011, 32, 1748–54. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, D.G. Structure–antimicrobial activity relationship between pleurocidin and its enantiomer. Exp Mol Med 2008, 40, 370–376. [Google Scholar] [CrossRef]
- Patrzykat, A.; Friedrich, C.L.; Zhang, L.; Mendoza, V.; Hancock, R.E.W. Sublethal concentrations of pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob Agents Chemother 2002, 46, 605–614. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Influence of the hydrophobic amino acids in the N- and Cterminal regions of pleurocidin on antifungal activity. J Microbiol Biotechnol 2010, 20, 1192–1195. [Google Scholar] [CrossRef]
- Hazam, P.K.; Cheng, C.C.; Lin, W.C.; Hsieh, C.Y.; Chen, J.Y. Strategic modification of low-active natural antimicrobial peptides confers ability to neutralize pathogens in vitro and in vivo. 2022; Unpublished.
- Hazam, P.K.; Chen, J.Y. Therapeutic utility of the antimicrobial peptide Tilapia Piscidin 4 (TP4). Aquac Rep 2020, 17, 100409. [Google Scholar] [CrossRef]
- Krause, A.; Neitz, S.; Mägert, H.J.; Schulz, A.; Forssmann, W.G.; Schulz-Knappe, P.; Adermann, K. LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett 2000, 480, 147–150. [Google Scholar] [CrossRef]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J Biol Chem 2001, 276, 7806–7810. [Google Scholar] [CrossRef]
- Barton, J.C.; Acton, R.T. Hepcidin, iron, and bacterial infection. Vitam Horm 2019, 110, 223–242. [Google Scholar] [CrossRef]
- Shike, H.; Lauth, X.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; Van Olst, J.C.; Shimizu, C.; Bulet, P.; Burns, J.C. Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge. Eur J Biochem 2002, 269, 2232–2237. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A.; Verghese, B. Evidence for positive Darwinian selection on the hepcidin gene of Perciform and Pleuronectiform fishes. Mol Divers 2007, 11, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Hilton, K.B.; Lambert, L.A. Molecular evolution and characterization of hepcidin gene products in vertebrates. Gene 2008, 415, 40–48. [Google Scholar] [CrossRef]
- Wang, K.J.; Cai, J.J.; Cai, L.; Qu, H.D.; Yang, M.; Zhang, M. Cloning and expression of a hepcidin gene from a marine fish (Pseudosciaena crocea) and the antimicrobial activity of its synthetic peptide. Peptides 2009, 30, 638–646. [Google Scholar] [CrossRef]
- Neves, J.V.; Caldas, C.; Vieira, I.; Ramos, M.F.; Rodrigues, P.N. Multiple hepcidins in a teleost fish, Dicentrarchus labrax: Different hepcidins for differen roles. J Immunol 2015, 195, 2696–2709. [Google Scholar] [CrossRef]
- Fennell JF, Shipman WH, Cole LJ. Antibacterial action of melittin, a polypeptide from bee venom. Proceed Soc Exp Biol Med 1968, 127, 707-710. [CrossRef]
- Pereira, A.V.; de Barros, G.; Pinto, E.G.; Tempone, A.G.; Orsi, R.O.; Dos Santos, L.D.; et al. Melittin induces in vitro death of Leishmania (Leishmania) infantum by triggering the cellular innate immune response. J Venom Animals Toxins Trop Dis 2016, 22, 1. [Google Scholar] [CrossRef]
- Park, J.; Kwon, O.; An, H.J.; Park, K.K. Antifungal effects of bee venom components on Trichophyton rubrum: A novel approach of bee venom study for possible emerging antifungal agent. Annals Dermatol 2018, 30, 202–210. [Google Scholar] [CrossRef]
- Kim, Y.W.; Chaturvedi, P.K.; Chun, S.N.; Lee, Y.G.; Ahn, W.S. Honeybee venom possesses anticancer and antiviral effects by differential inhibition of HPV E6 and E7 expression on cervical cancer cell line. Oncology Rep 2015, 33, 1675–1682. [Google Scholar] [CrossRef]
- Van den Bogaart, G.; Guzman, J.V.; Mika, J.T.; Poolman, B. On the mechanism of pore formation by melittin. J Biol Chem 2008, 283, 33854–33857. [Google Scholar] [CrossRef]
- Rangel, K.; Lechuga, G.C.; Almeida Souza, A.L.; Carvalho, J.P.R.S.; Villas-Bôas, M.H.S.; De Simone, S.G. Pan-drug resistant Acinetobacter baumannii but not other strains are resistant to the bee venom peptide melittin. Antibiotics (Basel) 2020, 149, 178. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Borovsky, D.; Rougé, P.; Shatters, R.G. Jr. Bactericidal properties of proline-rich Aedes aegypti trypsin modulating oostatic factor (AeaTMOF). Life (Basel) 2022, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial peptides, a mini review. Toxins (Basel). 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.; Hultmark, D.; Engström, Å.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Leung, K.; Chen, H.M. The combined effects of antibacterial peptide cecropin a and anticancer agents on leukemia cells. Anticancer Research 2002, 22, 2811–2816. [Google Scholar]
- Lim, J.; Hong, J.; Jung, Y.; Ha, J.; Kim, H.; Myung, H; Song, M.J. Bactericidal effect of Cecropin A fused endolysin on drug-resistant gram-negative pathogens. Microbiol Biotechnol 2022, 32, 816-823. [CrossRef]
- Mandel, S.; Michaeli, J.; Nur, N.; Erbetti, I.; Zazoun, J.; Ferrari, L.; Felici, A.; Cohen-Kutner, M.; Bachnoff, N. OMN6 a novel bioengineered peptide for the treatment of multidrug resistant Gram negative bacteria. Sci Rep 2021, 11, 6603. [Google Scholar] [CrossRef]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int J Biol Sci 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Burian, M.; Schittek, B. The secrets of dermcidin action. Int J Med Microbiol 2015, 305, 283–286. [Google Scholar] [CrossRef]
- Zeth, K.; Méndez-Vilas, A. Structure and mechanism of human antimicrobial peptide dermcidin and its antimicrobial potential microb pathog strateg combat them. Sci Technol Educ 2013, p.1333-1342.
- Liu, C.; Qi, J.; Shan, B.; Ma, Y. Tachyplesin causes membrane instability that kills multidrug-resistant bacteria by inhibiting the 3-ketoacyl carrier protein reductase FabG. Front Microbiol 2018, 9, 825. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.J.; Liu, Y.Y.; Li, H.; Guo, L.X.; Liu, Z.H.; Shi, X.L.; Hu, M. In vitro potential of Lycosin-I as an alternative antimicrobial drug for treatment of multidrug-resistant Acinetobacter baumannii infections. Antimicrob Agents Chemother 2014, 58, 6999–7002. [Google Scholar] [CrossRef]
- Cao, L.; Dai, C.; Li, Z.; Fan, Z.; Song, Y.; Wu, Y.; Cao, Z.; Li, W. Antibacterial activity and mechanism of a scorpion venom peptide derivative in vitro and in vivo. PLoS One 2012, 7, e40135. [Google Scholar] [CrossRef]
- Oyama, L.B.; Girdwood, S.E.; Cookson, A.R.; Fernandez-Fuentes, N.; Privé, F.; Vallin, H.E.; Wilkinson, T.J.; Golyshin, P.N.; Golyshina, O.V.; Mikut, R.; et al. The rumen microbiome: an underexplored resource for novel antimicrobial discovery. NPJ Biofilms Microbiomes 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Shin, A.; Lee, E.; Jeon, D.; Park, Y.G.; Bang, J.K.; Park, Y.S.; Shin, S.Y.; Kim, Y. Peptoid-substituted hybrid antimicrobial peptide derived from papiliocin and magainin 2 with enhanced bacterial selectivity and anti-inflammatory activity. Biochemistry 2015, 54, 3921–31. [Google Scholar] [CrossRef] [PubMed]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane-active mechanisms against Pseudomonas aeruginosa. Sci Rep 2021, 10, 9117. [Google Scholar] [CrossRef]
- Ferre, R.; Melo, M.N.; Correia, A.D.; Feliu, L.; Bardaji, E.; Planas, M.; Castanho, M. Synergistic effects of the membrane actions of cecropin-melittin antimicrobial hybrid peptide BP100. Biophys J 2009, 96, 1815–1827. [Google Scholar] [CrossRef]
- Oddo, A.; Thomsen, T.T.; Kjelstrup, S.; Gorey, C.; Franzyk, H.; Frimodt-Møller, N.; Løbner-Olesen, A.; Hansen, P.R. An all-D amphipathic undecapeptide shows promising activity against colistin-resistant strains of Acinetobacter baumannii and a dual mode of action. Antimicrob Agents Chemother 2016, 60, 592–599. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem Pharmacol 2017, 133, 117. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist Updat 2016, 26, 43–57. [Google Scholar] [CrossRef]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moghaddam, M.M. , Mirnejad R. Antimicrobial peptides: Features action and their resistance mechanisms in bacteria. Microb Drug Resist 2018, 24, 747. [Google Scholar] [CrossRef]
- Joo, H.S.; Fu, C.I.; Otto, M. Bacterial strategies of resistance to antimicrobial peptides. Phil Trans R Soc Lond B Biol Sci 2016, 371, 20150292. [Google Scholar] [CrossRef]
- Omardien, S.; Brul, S.; Zaat, S.A.J. Antimicrobial activity of cationic antimicrobial peptides against gram-positive: Current progress made in understanding the mode of action and the response of bacteria. Front Cell Develop Biol 2016, 4, 111. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front Microbiol 2012, 3, 408. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.U. Antimicrobial peptides: Mechanisms of action and resistance. J Dental Res 2017, 96, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to polymyxins in gram-negative organisms. Int. J Antimicr Ag 2017, 49, 526–535. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J, Doi Y, Tian G, Dong B, Huang X.; et al., Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect Dis 2016, 16, 161-168. [CrossRef]
- Paterson, D.L.; Harris, P.N.A. Colistin resistance: A major breach in our last line of defense. Lancet Infect Dis 2016, 16, 132–133. [Google Scholar] [CrossRef]
- Macnair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming mcr-1 mediated colistin resistance with colistin in combination with other antibiotics. Nature Commun 2018, 9, 458. [Google Scholar] [CrossRef]
- Gheorghe, I.; Saviuc, C.; Ciubuca, B.; Lazar, V.; Chifiriuc, M.C. Chapter 8—Nano drug delivery. In: Grumezescu, A.M, Ed. Nanomaterials for Drug Delivery and Therapy. Norwich NY USA: William Andrew Publishing; 2019. pp. 225-244. ISBN: 978-0-12-816505-8.
- Sun, B.; Wibowo, D.; Middelberg, A.P.J.; Zhao, C.X. Cost-effective downstream processing of recombinantly produced pexiganan peptide and its antimicrobial activity. AMB Express 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Akbari Eidgahi, M.R.; Khaledi, A.; Ghazvini, K. Epinecidin-1, a highly potent marine antimicrobial peptide with anticancer and immunomodulatory activities. BMC Pharmacol Toxicol 2019, 20, 33. [Google Scholar] [CrossRef]
- Neshani, A.; Tanhaeian, A.; Zare, H.; Eidgahi, M.R.A.; Ghazvini, K. Preparation and evaluation of a new biopesticide solution candidate for plant disease control using pexiganan gene and Pichia pastoris expression system. Gene Rep 2019, 17, 100509. [Google Scholar] [CrossRef]
Figure 1.
Interaction of cationic AMPs with eukaryotic and bacterial membranes. Images were created using BioRender.com.
Figure 1.
Interaction of cationic AMPs with eukaryotic and bacterial membranes. Images were created using BioRender.com.
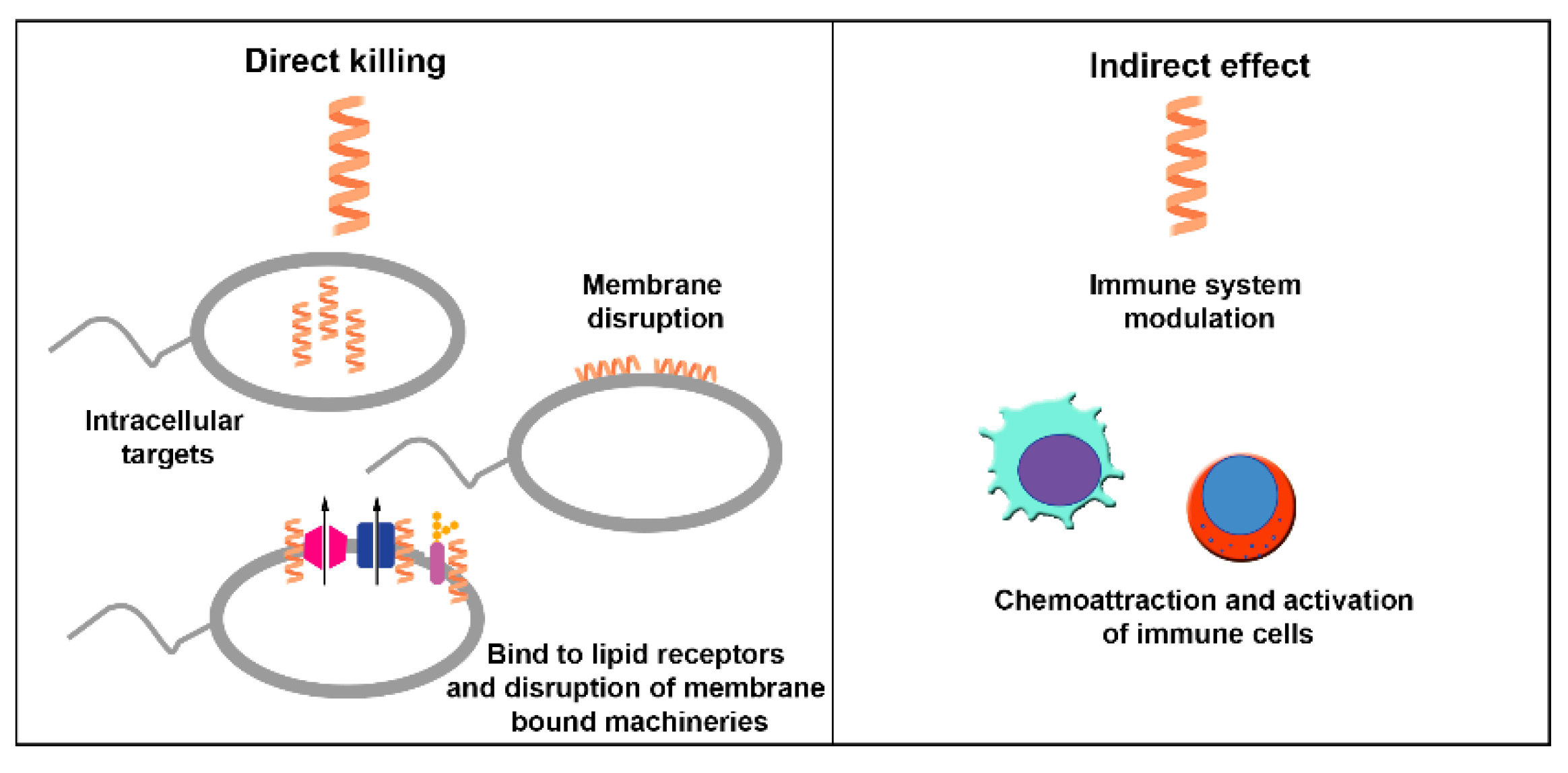
Figure 2.
Mechanisms of action of AMPs on the surface of bacteria. A) Barrel-stave model: accumulated AMPs inserted into the membrane bilayer and associated into a channel. (B) Toroidal pore model: accumulated AMPs inserted in vertical and bent orientations to form a pore. (C) Carpet model: AMPs accumulate on the surface until a critical concentration displays detergent behavior to form micelles (D). Images generated at BioRender.com.
Figure 2.
Mechanisms of action of AMPs on the surface of bacteria. A) Barrel-stave model: accumulated AMPs inserted into the membrane bilayer and associated into a channel. (B) Toroidal pore model: accumulated AMPs inserted in vertical and bent orientations to form a pore. (C) Carpet model: AMPs accumulate on the surface until a critical concentration displays detergent behavior to form micelles (D). Images generated at BioRender.com.
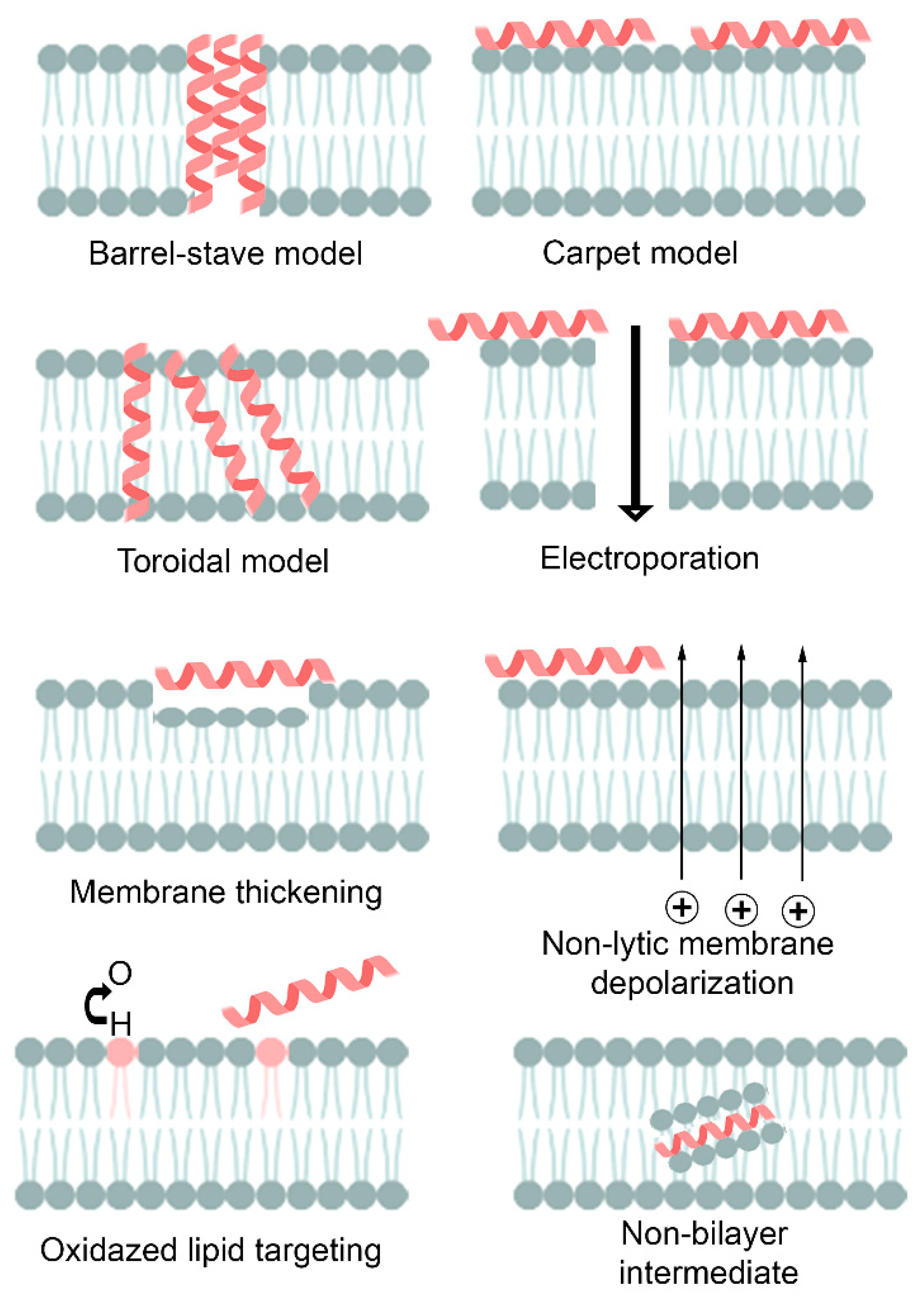

Table 1.
AMP produced by living organisms with activity anti-A. baumannii.
| Peptide | Source | Sequence (nº amino acid) | Structure | MIC against A. baumannii (μg/mL) | Ref. | |
|---|---|---|---|---|---|---|
| Antibiotic-susceptible | MDR | |||||
| LL-37 | H. sapiens | LLGDFFRKSKEKIGKEFKRIVQRIK DFLRNLVPRTES (37aa) | AH | 32 | 16–32 | [61,62] |
| KR-30 | H. sapiens | KSKEKIGKEFKRIVQRIKDFLRNLV PRTES (30aa) | AH | 16 | 8–16 | [62] |
| KR-20 | H. sapiens | KRIVQRIKDFLRNLVPRTES (20aa) | AH | 64 | 16–32 | [62] |
| KS-12 | H. sapiens | KRIVQRIKDFLR (12aa) | AH | 256 | 64–256 | [62] |
| SAAP-148 | H. sapiens | LKRVWKRVFKLLKRYWRQLKKPVR (24aa) | AH | — | 6 | [63] |
| CATH-BF derivative (Cath-A and OH- | Bungarus fasciatus (Snake venom) | KFFRKLKKSVKKRAKEFFKKPRVI GVSIPF(30aa) | AH | — | 8–32 | [64] |
| ZY4 cathelicidin-BF-15 derived | Bungarus fasciatus (Snake venom) | VCKRWKKWKR KWKKWCV-NH2 (17aa) | Cyclic SH-bridge | — | 4.6–9.4 | [65] |
| NA-CATH | Naja atra (Snake venom) | KRFKKFFKKLKNSVKKRAKKFFKK PKVIGVTFPF (34aa) | AH | 10 | 10 | [66] |
| OH-CATH30 | King cobra (Snake venon) | KFFKKLKNSVKKRAKKFFKKPRVI GVSIPF(30aa) | AH | 10 | 10 | [67] |
| DOH-CATH30 | King cobra (Snake venon) | KFFKKLKNSVKKRAKKFFKKPRVIGVSIPF (30aa) | AH | — | 1.56–12.5 | [67] |
| BMAP-27 | Bovine myeloid | GRFKRFRKKFKKLFKKLSPVIPLLHLG (27aa) | AH | 8-16 | 4-16 | [68] |
| SMAP29 | Sheep myeloid | RGLRRLGRKIAHGVKKYGPTVLRIIRIAG (29aa) | AH | 8 | 4-32 | [68] |
| Latarcin 2a | Winter flounder (Pleuronectes americanos) | H-GLFGKLIKKFGRKAISYAVKKARGKH-OH (26aa) | AH | 16 | 8-64 | [68] |
| Maximin H2 | Nile Tilapia (Oreochromis niloticus) | ILGPVLSMVGSALGGLIKKI-NH2 (20aa) | AH | 64 | 16-128 | [68] |
| NRC12 | Flatfish Genes | GWKKWFNRAKKVGKTVGGLAVDHYL-NH2 (25aa) | AH | 16 | 8-32 | [68] |
| Pilosulin | Ant venom (toxin pilosulin) | GLGSVFGRLARILGRVIPKV-NH2 (20aa) | AH | 16 | 8-16 | [68] |
| Pleurocidin | GWGSFFKKAAHVGKHVGKAALTHYL-NH2 (25aa) | AH | 16 | 8-32 | [68] | |
| TP4 | FIHHIIGGLFSAGKAIHRLIRRRRR (25aa) | AH | 16 | 8-32 | [68] | |
| D-Myrtoxin-Mp 1a (Mp1a) | Venom of Myrmecia pilosula | IDWKKVDWKKVSKKTCKVMXKACKEL-NH2 (26aa) | AH | 0.025* | — | [69] |
| Venon cocktail proteins | Leiurus quinquestriatus (Scorpion venom) | Cocktail | — | — | 50.6% of inhibition at 20 mg/mL of venom | [70] |
| Ranalexin | Rana catesbeiana (American bulfrog) | LGGLIKIVPAMICAVTKKC (19aa) | AH | — | 4–18 | [71] |
| Danalexin | Rana catesbeiana (American bulfrog) | LGGLIKIVPAMICAVTKKC (19aa) | AH | — | 4–16 | [71] |
| LS-sarcotoxin | Lucilla serricata | GWLKKIGKKIERVGQHTRDATIQTIGVAQQAANVAATLK-NH2 (39aa) | AH | 4 | 4–8 | [72] |
| LS-stomoxyn | Lucilla serricata | GFRKRFNKLVKKVKHTIKETANVSKDVAIVAGSGVAVGAAM-NH2 (41aa) | AH | 8 | 4–16 | [72] |
| Mini-ChBac7.5 Nα | Domestic goat (Capra hircus) | RRLRPRRPRLPRPRPRPRPRPR (22aa) | AH | — | 2* | [73] |
| Mini-ChBac7.5 Nβ | Domestic goat (Capra hircus) | RRLRPRRPRLPRPRPRPRPRP (21aa) | AH | — | 4* | [73] |
| AM-CATH36 | American alligator | GLFKKLRRKIKKGFKKIFKRLPPIG VGVSIPLAGKR (36aa) | AH | 5.2 | 5.2 | [66] |
| AM-CATH28 | American alligator | KIKKGFKKIFKRLPPIGVGVSIPLAGKR (28aa) | AH | 28 | 10 | [66] |
| AM-CATH21 | American alligator | GLFKKLRRKIKKGFKKIFKRL (21aa) | AH | 42 | 10 | [66] |
| WAM-1 | Tammar wallaby (Macropus eugenii) | KRGFGKKLRKRLKKFRNSIK KRLKNFNVVIPIPLPG (36aa) | AH | 8.12 | 4–64 | [74,75] |
| Indolicidin | Cytoplasmic granules of the bovine neutrophils | LPWKWPWWPWRR-NH2 (13aa) | Other structure | 4 | 2–64 | [75,76,77] |
| Bactenecin | Bovine neutrophil granules, Caprine | LCRIVVIRVCR (12aa) | B-turn structure Ciclyc | 64 | — | [75,78,79] |
| Histatin-8 | H. sapiens | KFHEKHHSHRGY (12aa) | AH | 8 | — | [77] |
| HNP-1 | H. sapiens (Polymorphonuclear neutrophil) | ACYCRIPACIAGERRYGTCIYQGRLWAFCC (30aa) | AH | 50 | — | [76] |
| HNP-2 | H.sapiens (Polymorphonuclear neutrophil) | CYCRIPACIAGERRYGTCIYQGRLWAFCC (29aa) | AH | 50 | — | [76] |
| HD5d5 | H.sapiens (Polymorphonuclear neutrophil) | ARARCRRGRAARRRRLRGVCRIRGRLRRLAAR (32aa) | AH | 40 | 40 | [80] |
| CL defensin | Cimex Lectularius (Bedbug) | ATCDLFSFQSKWVTPNHAACAAHCTARGNRGGRCKKAVCHCRK (43aa) | AH, antiparallel BS; N-terminal loop | — | — | [81] |
| HBD-2 | Epithelial lining of respiratory /urinary tracts | GIGDPVTCLKSGAICHPVFCPRRY KQIGTCGLPGTKCCKKP (41aa) | Beta | 3.90–9.35 | 3.25–4.5 | [82] |
| HBD-3 | Epithelial lining of respiratory /urinary tracts | GIINTLQKYYCRVRGGRCAVLSCL PKEEQIGKCSTRGRKCCRRKK (45aa) | AH + BS | 4 | 4 | [83] |
| Magainin-1 | Frog skin peptide | GIGKFLHSAGKFGKAFVGEIMKS (23aa) | AH | — | 256 | [84,85] |
| Magainin-2 | Frog skin peptide | GIGKFLHSAKKFGKAFVGEIMNS (23aa) | AH | 9.8–64 | 4.9–64 | [84,85,86] |
| Pexiganan | Frog skin peptide | GIGKFLKKAKKFGKAFVKILKK (22aa) | AH | 1–8 | 1–8 | [87,88,89] |
| Aurein 1,2 | Frog skin peptide | GLFDIIKKIAESF (13aa) | AH | 16 | — | [89] |
| Citropin 1.1. | Australian tree frog of the Litora genus | GLFDVIKKVASVIGGL-NH2 (16aa) | AH | 16 | — | [89] |
| Temporin A | European red frog Rana temporaria | FLPLIGRVLSGIL-NH2 (13aa) | AH | 128 | — | [89] |
| Brevinina 2 (B2RP) | Frog skin peptide | GIWDTIKSMGKVFAGKILQNL-NH2 (21aa) | AH | 29 | 7–13.9 | [90] |
| [D4K] B2RP | Frog skin peptide | GIWKTIKSMGKVFAGKILQNL-NH2 (21aa) | AH | 4–16 | 4–16 | [91,92] |
| B2RP-Era | Frog skin peptide | GVIKSVLKGVAKTVALGML-NH2 (19aa) | AH | 8–32 | 8–64 | [91,93] |
| Alytesirin-1c | Frog skin peptide | GLKEIFKAGLGSLVKGIAAHVAS-NH2 (23aa) | AH | — | 11.3–22.6 | [94] |
| [E4k] Alytesirin-1c | Frog skin peptide | GLKEIFKAGLGSLVKGIAAHVAS-NH2 (23aa) | AH | 4–16 | 4–16 | [91,92] |
| Tachyplesin III | Horseshoe crabs (Tachypleus gigas) and (Carcinoscorpius rotundicauda) | KWCFRVCYRGICYRKCR-NH2 (17aa) | BS 2 dissulfite bridges | — | 8-16 | [92] |
| [S7K, G11K] Alytesirrin-2a | Frog skin peptide | ILGKLLKTAAKLLSNL-NH2 (16aa) | AH | — | 8 | [95] |
| PGLa-AM1 | Frog skin peptide | GMASKAGSVLGKVAKVALKAAL-NH2 (22aa) | AH | 16–128 | 16–128 | [91,96] |
| CPF-AM1 | Frog skin peptide | GLGSVLGKALKIGANLL (19aa) | AH | 16–128 | 4–128 | [91,97,98] |
| CPF-B1 | Frog skin peptide | GLGSLLGKAFKIGLKTVGKMMGGAPREQ (28aa) | — | 11.4–22.8 | [99] | |
| CPF-C1 | Frog skin peptide | GFGSLLGKALRLGANVL (17aa) | 5 | — | [98] | |
| [E6k,D9k] Hymenochirin-1B | Frog skin peptide | LKLSPKTKDTLKKVLKGAIKGAIA IASMA-NH2 (29aa) | AH | — | 4.9 | [100] |
| Hymenochirin-1 Pa | Frog skin peptide | LKLSPKTKDTLKKVLKGAIKGAIAIASMA-NH2 (29aa) | AH | — | — | [101] |
| [G4K] XT7 | Frog skin peptide | GLLGPLLKIAAKVGSNLL-NH2 (18aa) | AH | 4–32 | 4–64 | [91,102] |
| Buforin II | Frog skin peptide | TRSSRAGLQFPVGRVHRLLRK (21aa) | AH | 8–19.5 | 0.25–39 | [84,85,103,104] |
| Melittin | European honeybee (Apis mellifera) | GIGAVLKVLTTGLPALISWIKRKRQQ (26aa) | AH | 0.25–4 | 0.25–25 | [76,105,106] |
| Cecropin A | Cecropia moth (Hyalophora cecropia) | KWKLFKKIEKVGQNIRDGIIKAGP AVAVVGQATQIAK (37aa) | AH | 32 | 0.5–32 | [76,107] |
| BR003-cecropin A | Aedes aegypti | GGLKKLGKKLEGAGKRVFNAAEK ALPVVAGAKALRK (36aa) | 5 | 5 | [108] | |
| Mdc | Housefly larvae | GWLKKIGKKIERVGQHTRDATIQ TIGVAQQANAVAATLKG (40aa) | 4 | 4 | [109] | |
| Cecropin P1 | Pig (Ascaris suum) | SWLSKTAKKLENSAKKRISEGIAIA IQGGPR (31aa) | 1.6 | — | [76,110] | |
| Myxinidin 2 | Myxine glutinosa L | KIKWILKYWKWS (12aa) | AH | — | 12.5 | [111] |
| Myxinidin 3 | Myxine glutinosa L | RIRWILRYWRWS (12aa) | BS | — | 6.3 | [111] |
| FLIP 7 | Calliphora vicina (Medicinal Maggots) | AH | — | 125-416 biofilm bactéria sensitivity | [112] | |
| Mastoparan | Vespula lewisii (Hornet venom) | INLKALAALAKKIL (14aa) | AH | 4 | — | [76,113,114] |
| Mastoparan-AF (EMP-AF) | Hornet venom (Vespa affinis) | INLKAIAALAKKLF-NH2 (14aa) | AH | 2–16 | 2–16 | [115] |
| Mastoparan-Chitosan Nanoconstruct | Wasp venom(Vespula lewissi) | INLKALAALAKKIL-NH2 (14aa) | AH | - | 2-4 | [116] |
| DCD-1 L | Eccrine sweat glands | SSLLEKGLDGAKKAVGGLGKLGKDAVEDLESVGKGAVHDVKDVLD SVL (48aa) | AH | 16 | — | [117,118] |
| Hp l404 analogs (A, K, V, L, I, W) | Venom gland scorpion (Heterometnus pertersii) | GILGKLWEGVKSIF-NH2 (14aa) | AH | 3.13–12.5 | 3.13–16.25 | [119,120] |
| Protegrin-1 | Cimex lectularius | RGGRLCYCRRRFCVCVGR-NH2 (18aa) | AH | — | 2-8 | [121] |
| Nuripep 1653 | Derived from the P54 nutrient reservoir protein (aa 271–292) pea protein from Pisum sativum | VRGLAPKKSLWPFGGPFKSPFN (22aa) | AH | — | 12 | [122] |
| Agelaia-MPI | Agelaia pallipes pallipes | INWLKLGKAIIDAL (14aa) | AH | 6.25 | 12.5–25 | [123] |
| Polybia-MPII | Pseudopolybia vespiceps testacea | INWLKLGKMVIDAL (14aa) | AH | 12.5 | 25 | [123] |
| Polydin-I | Polybia dimorpha (Social wasp) | AVAGEKLWLLPHLLKMLLTPTP (22aa) | AH | >25 | >25 | [123] |
| Con10 | Scorpion venoms (Opisthacanthus cayaporum) | FWSFLVKAASKILPSLIGGGDDNKSSS (27aa) | AH | 12.5 | 12.5 | [123] |
| NDBP5.8 | Scorpion venoms (Opisthacanthus cayaporum) | GILGKIWEGVKSLI (14aa) | AH | >25 | >25 | [123] |
| Delfibactin A | Gram-negative bactéria Delfia spp. | C40H68N14O18 | — | — | 16 | [124] |
| WLBU2- arginine-rich amphiphilic peptide | Skin wounds | RRWVRRVRRWVRRVVRVVRRWVRR (24aa) | — | ∼7.484 | ∼7.484 | [125] |
| α-Helical-26 (A12L/A20L) | D- and L-diastereomeric peptides | Ac-KWKSFLKTFKSLKKTVLHTLLKAISS-NH2 (26aa) | AH | — | 0.5–1.0 | [126] |
| Cy02 (cyclotide) | Viola odorata | GIPCGESCVWIPCISSAIGCSCKSKVCYRN (30aa) | BSs | — | 15* | [127] |
| Bicarinalin (YRTX-Tb1a) | Tetramorium bicarinatum venom | KIKIPWGKVKDFLVGGMKAV (20aa) | AH | — | 4 | [128] |
| Glatiramer acetate | Homo sapiens | EAYKAAEKAYAAKEAAKEAAKAKAEKKAAYAKAKAAKYEKKAKKAAAEYKKK (52aa) | — | Reduct viable cells | Reduct viable cells | [129] |
| Lactoperoxidase (Lpo) | Camel (Colostrum milk) | Large protein | complex | — | Inhibition effects, significant clearance of Ab in lung and BC | [130] |
| Lactoferrin (Lf) | Camel (Colostrum milk) | Large protein | complex | — | Inhibition effects, significant clearance of Ab in lung and BC | [130] |
| Artlysin Art-175 | Pseudomonas aeruginosa bacteriophage | Comprises a modified variant of endolysin KZ144 with an N-terminal fusion to SMAP-29 | — | — | 4–20 | [131] |
| Epsilon-poly L-lysine (EPL)-catechol | Streptomyces albulus derived | Complex | — | — | Reducing bacterial burden in vivo | [132] |
| Nodule-specific cysteine-rich (NCR) peptide and its derivatives | Medicago trunculata | RNGCIVDPRCPYQQCRRPLYCRRR (24aa) | AH | 1.6–25 MBC | — | [133,134] |
| D-150-177C, HBcARD derivative peptide | Hepatitis B virus | RRRGRSPRRRTPSPRRRRSQSPRR RRSC (28aa) | AH | 16 | 16–32 | [135] |
| Colistin (Polymyxin E) | Bacillus colistinus | C52H98N16O13 (cyclic compound) | — | Antibiofilm, side effects | — | [136] |
| PlyF307 (P307) | Phage Lysin | 146 aa (Access number KJ740396) | — | 750 | 750–2000 | [137] |
| P307SQ−8C | Hepatitis B virus | NAKDYKGAAAEFPKWNKAGGRV LAGLVKRRKSQSRESQC (39aa) | — | 125 | 62.5–125 | [137] |
| N10 | Blood biopanning | ACKDVNTSMCGGK (13aa) | AH | 500 | 500 | [138] |
| NB2 | Biofilm biopanning | ACERSIRTVCGGK (13aa) | AH | 500 | 500 | [138] |
| Melittin with imipenem (IPM) | European honeybee and antimicrobial | GIGAVLKVLTTGLPALISWIKRKR QQ (26aa) + IPM | AH | 0.31–0.37 | 0.12–0.25 | [139] |
| Melittin with colistin (COL) | European honeybee and antimicrobial | GIGAVLKVLTTGLPALISWIKRKR QQ (26aa) + COL | AH | 0.37–0.5 | 0.19–0.37 | [139] |
| LS-AMP-E1 | Chinese wolf spider (Lycosa sinensis) | AGMKNIIDAIKKKLGGKL (18aa) | AH | - | 25-100* | [140] |
| LS-AMP-F1 | Chinese wolf spider (Lycosa sinensis) | TGLGKIGYLMKKLLSKAKV(19aa) | AH | - | 3.1-12.5* | [140] |
| Caerin 1.1 | Australian tree frog | GLLSVLGSVAKHVLPHVVPVIAEHL-NH2 | AH | 7.5 | - | [141] |
| Caerin 1.9 | Australian tree frog | GLFGVLGSIAKHVLPHVVPVIAEKL-NH2 | AH | 3.75 | - | [141] |
| Caerin 1.1 + Caerin 1.9 | Australian tree frog | - | AH | 0.9375- 1.875 | - | [141] |
| Am23SK | Alligator mississippiensis | SCRFSGGYCIWNWERCRSGHFLVALCPFRKRCCK (34aa) | AH | - | 2 | [142] |
| Nisin | Probiotic bacterium (Lactococcus lactis) | (34aa) | - | 128 | 64-128 | [143] |
| P10 + Nisin in combination | - | - | - | 32 | 16-32 | [143] |
| Hp1404 | Scorpion venom gland (Heterometrus petersii) | GILGKLWEGVKSIF (14aa) | AH | 5 | 5-10 | [144,145] |
| VsCT1 | Scorpion venom gland (Heterometrus petersii) | FLKGIIDTVSNWL (13aa) | AH | >40 | - | [144] |
| BmKn1 | Scorpion venom gland (Heterometrus petersii) | FIGAVAGLLSKIF (13aa) | AH | >40 | - | [144] |
| Spiniferin | Scorpion venom gland (Heterometrus petersii) | ILGEIWKGIKDIL (13aa) | AH | >40 | - | [144] |
| Im4 | Scorpion venom gland (Heterometrus petersii) | FIGMIPGLIGGLISAIK (17aa) | AH | >40 | - | [144] |
| Ctriporin | Scorpion venom gland (Heterometrus petersii) | FLWGLIPGAISAVTSLIKK (19aa) | AH | 20 | 20-40 | [144] |
| Im5 | Scorpion venom gland (Heterometrus petersii) | FLGSLFSIGSKLLPGVIKLFQRKKQ (25aa | AH | 2.5 | 2.5-10 | [144] |
| Hp1404 analogs | Scorpion venom gland (Heterometrus petersii) | - | AH | 3.13-25* | - | [145] |
| Lynronne-1 | Bovine rumen microbiome | LPRRNRWSKIWKKVVTVFS (19aa) | AH | 4 | - | [146] |
| BmKn2 | Scorpion (Mesobuthus martensii Karsch) | FIGAIARLLSKIF-NH2 (13aa) | AH | 10 | 5-10 | [147] |
| Hylin a1 | American frog (Hypsiboas albopunctatus) | IFGAILPLALGALKNLIK-NH2 (18aa) | AH | 2* | 2-8* | [148] |
| Hylin a1-11K | American frog (Hypsiboas albopunctatus) | IAKAILPLALKALKNLIK-NH2 (19aa) | AH | 1-2* | 1-2* | [148] |
| aHylin a1-15K | American frog (Hypsiboas albopunctatus) | IAKAILPLALKALKKLIK-NH2 (19aa) | AH | 1-2* | 1-2* | [148] |
| Esc(1-21) | Frog-skin | GIFSKLAGKKIKNLLISGLKG-NH2 (21aa) | AH | - | 17.5-35 | [149] |
| Esc(1-21) + Colistina | Frog-skin | GIFSKLAGKKIKNLLISGLKG-NH2 (21aa) + Colistina | AH | - | 1.1-4.4 | [149] |
| HBD-3 | Epithelial cells | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK (45aa) | - | - | 4-16 | [150] |
| Epi-122–42 | Orange-spotted grouper (Epinephelus coioides) | GFIFHIIKGLFHAGKMIHGLV (21aa) | - | - | 4–32 | [150] |
Table 2.
Synthetic AMPs with activity anti-A. baumannii. c.
| Peptide | Source | Structure | MIC against A. baumannii (μg/mL) | Ref. | ||
|---|---|---|---|---|---|---|
| Antibiotic-susceptible | MDR | |||||
| GW-A2 | Synthetic peptide | GAKYAKIIYNYLKKIANALW (20aa) | AH | 32 | 8-32 | [68] |
| GW-H1a | Synthetic peptide | GYNYAKKLANLAKKFANALW-NH2 (20aa) | AH | 32 | 8-32 | [68] |
| GW-Q6 | Synthetic peptide | GIKIAKKAITIAKKIAKIYW (20aa) | AH | 16 | 8-16 | [68] |
| Omiganan | Synthetic peptide | ILRWPWWPWRRK-NH2 (12aa) | AH | 32 | — | [89] |
| r-Omiganan | Synthetic peptide | KRRWPWWPWRLI-NH2 (12aa) | AH | 16 | — | [89] |
| SAAP-148 NPs | Synthetic peptide | LKRVWKRVFKLLKRYWRQLKKPVR (24aa) + NPs | AH | — | — | [151] |
| Cecropin-4 | Synthetic peptide | GWLKKIGKKIERVGQNTRDATIQ AIGVAQQAANVAATLKG (40aa) | AH | 4 | 4 | [152,153] |
| Octopromycin | Synthetic peptide | N-RRLIRTDTGPIIYDYFKDQLLKKGMVILRESMKNLKGM-C (38aa) | AH | - | 50 | [154] |
| LyeTx I-bPEG | Synthetic peptide | AH | [155] | |||
| zp3 | Synthetic peptide | GIIAGIIIKIKK-NH2 (12aa) | AH | 4 | - | [156] |
| P10 | Synthetic derivated | LAREYKKIVEKLKRWLRQVLRTLR (24aa) | 4 | 8-32 | [143] | |
| P10 + Nisin in combination | - | - | 1 | 4-16 | [143] | |
| NCR169C and its substitution derivatives | Synthetic peptide | KSKKPLFKIWKCVENVCVLWYK | AH | 1.6-12.5 MBC | — | [157] |
| OMN6 | Synthetic peptide | H-M-C-KWKLFKKIEKVGQNIRDGIIKA-GP-AVAVVGQATQIAK-C-NH2(40aa) | AH | 8 | 4-8 | [158,159] |
| BmKn2-7 | Synthetic peptide | FIKRIARLLRKIF-NH2 (13aa) | AH | 5 | 5-10 | [147] |
| BmKn2-7R | Synthetic peptide | FIRRIARLLRRIF-NH2 (13aa) | AH | 2.5 | 2.5-5 | [147] |
| BmKn2-7K | Synthetic peptide | FIKKIAKLLKKIF-NH2 (13aa) | AH | 2.5 | 2.5-5 | [147] |
| AS-CATH8 | Synthetic peptide | KRVNWAKVGRTALKLLPYIFG (21aa) | AH | 0.6 | - | [142] |
| LJ-hep2(66–86) | Synthetic peptide | IKCKFCCGCCTPGVCGVCCRF (21aa) | - | 1.5-3 | [160] | |
| DGL 13K | Synthetic derived D-enantiomers of GL13K derived from the salivary protein BPIFA2 | GKIIKLKASLKLL-NH2 (13aa) | - | - | 8-32 | [161] |
| PLP-3 | Synthetic peptide derived from the innate immune system of vertebrates | Antiparallel BS | 1-2 | 1-2 | [162] | |
| ECPep-D | Synthetic peptide | RPFTRAQWFAIQHISPRTIAMRAINNYRWR (30aa) | - | 37.57 | - | [163] |
| ECPep-2D-Orn | Synthetic peptide | OPFTOAQWFAIQHISPOTIAMOAINNYOWO (30aa) | - | 17.53 | - | [163] |
| Mt6 | Synthetic peptide | KKFKKTAKWLIKSAWLLLKSLALKMK (26aa) | AH | 8 | - | [164] |
| D-Mt6 | Synthetic peptide | KFKKTAKWLIKSAWLLLKSLALKMK (25aa) | AH | 8 | - | [164] |
| Octominin, Octominin-CNPs | Synthetic derived, defensin 3 of Octopus minor | GWLIRGAIHAGKAIHGLIHRRRH (23aa) | AH | — | 5 | [165,166] |
| Ceragenins; CSA-192; CSA-131; D-150-177C; HBcARDderivative | Cholic acid synthetic mimics, | Steroids compounds | - | — | — | [167] |
| OG1410 | ApoE-based synthetic peptide | acetyl-ASAib-LRKL-Aib-KRLL-amide | AH | 16 | 16 | [168] |
| MSI-78 | Synthetic peptide, magainin analogue | GIGLPLLLALLPGLAPVLILLL-NH2 (22aa) | AH | - | 5 | [169] |
| mCM11, cecropin–melittin 11 | Synthetic peptide | NH2-WRLFRRILRVL-NH2 (11aa) | AH | 32 | < 4 - > 512 | [170] |
| Scolopendin A2 | Synthetic peptide | AGLQFKVGRIGRLLRK (16aa) | - | 16 | [171] | |
| Trichogin analogs | Synthetic peptide | 1-Oct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Aib-Gly-Ile-Lol | 2- >128 | - | [172] | |
| RR | Computationally designed | WLRRIKAWLRR (11aa) | AH | — | 25–99 | [173,174] |
| RR2 | Computationally designed | WIRRIKKWIRRVHK (14aa) | AH | — | 3–6 | [174] |
| RR-4 | Computationally designed | WLRRIKAWLRRIKA (14aa) | AH | — | 3–6 | [174] |
| DP7 | Computationally designed | VQWRIRVAVIRK (12aa) | AH | — | 4–16 | [175,176,177] |
| Omega 76-shuft1 | Computationally designed | AFLLKKKKGIIFFEKAKKGK (20aa) | AH | — | 4–16 | [178] |
| ′Ω17 family peptides | Computationally designed | RKKAIKLVKKLVKKLKKALK (20aa) | AH | 2 | 1–8 | [178] |
| ′Ω76 family peptides | Computationally designed | FLKAIKKFGKEFKKIGAKLK (20aa) | AH | 4 | 2–8 | [178] |
| pepD2 | Computationally designed | WKKLKKLLKKLKKL-NH2 (14aa) | AH | 8 | - | [179] |
| Pro9-3 | Computationally designed | RLWLAIWRR-NH2 (9aa) | AH | 16 | 8-64 | [180] |
| Pro9-3D | Computationally designed | RLWLAIWRR-NH2 (9aa) | AH | 8 | 4-16 | [180] |
| R-Pro9-3 | Computationally designed | RRWIALWLR-NH2 (9aa) | AH | 16 | 8-32 | [180] |
| R-Pro9-3D | Computationally designed | RRWIALWLR-NH2 (9aa) | AH | 8 | 4-16 | [180] |
| TP2-5 | Computationally designed | KKCIAKAILKKAKKLLKKLVNP (22aa) | AH | 3.125 | 1.56-3.125 | [181] |
| TP2-6 | Computationally designed | KKCIAKAILKKAKKLLKDLVNP (22ª) | AH | 3.125 | 3.125-12.5 | [181] |
| IKR18 | Computationally designed | IKRQYKRFFKLFKWFLKK (18aa) | AH | 1 | - | [182] |
| HP(2–9)-ME(1–12) (HPME) | Chimeric peptide | AKKVFKRLGIGAVLKVLTTG (20aa) | AH | 6.25 | 3.12–12.5 | [183] |
| HP(2–9)-MA(1–12) (HPMA) | Chimeric peptide | AKKVFKRLGIGKFLHSAKKF-NH2 (20aa) | AH | 6.25 | 3.12–6.25 | [183] |
| CA(1–8)-ME(1–12) (CAME) | Chimeric peptide | KWKLFKKIGIGAVLKVLTTG-NH2 (20aa) | AH | 3.12 | 3.12–12.5 | [183] |
| CA(1–8)-MA(1–12) (CAMA) | Chimeric peptide | KWKLFKKIGIGKFLHSAKKF-NH2 (20aa) | AH | 12.5 | 3.12–12.5 | [183] |
| TAT-RasGAP317–326 anticancer peptide | Chimeric (cell penetrating sequence + Src homology sequence) | G48RKKRRQRRR57 + W317MWVTNLRTD326 | AH | Growth inhibitory effect | — | [184,185] |
| TAT-RasGAP317–326 | Chimeric peptide | G48RKKRRQRRR57 (10aa) | - | - | - | [186] |
| Cecropin A (1–8) melittin (1–10) (CAME) | Hybrid peptide | KWKLFKKIGIGAVLKVLTTG-NH2 (20aa) | AH | 32 | 8-32 | [68] |
| BP100 | Hybrid peptide | KKLFKKILKYL (11aa) | AH | — | 4 | [128] |
| I16K-piscidin-1 and analogs | Hybrid striped bass Morone saxatilis x M. chrysops | FFHHIFRGIVHVGKTIHRLVTG (22aa) | — | — | 3.1 | [187] |
| PapMA | Hybrid peptide | RWKIFKKIPKFLHSAKKF-NH2 (18aa) | AH | 32 | 16-32 | [188] |
| D-AP19 | Hybrid peptide | RLFRRVKKVAGKIAKRIWK-NH2 (19aa) | 7.81 | 3.91-15.63 | [189] | |
| BP214 | Hybrid peptide | KKLFKKILRYL (11aa) | AH | 2 | - | [190] |
| BP214 analogs | Hybrid peptide | - | AH | 2- >64 | - | [190] |
| PNA (RXR)4 XB | Peptide nucleic acid conjugated to (RXR)4 Phosphorodiamidate Morpholino Oligomers | RXRRXRRXRRXRXB (14aa) | — | — | 1.25* | [191] |
| Stapled AMP Mag (i + 4)1, 15(A9 K, B21A, N22 K, S23 K) | NA, based in magainin 2 structure | Mag(i + 4)1,15(A9K,B21A,N22K,S23K) | complex | — | — | [192] |
| S4A | NA | IOWAGOLFOLFO-NH2 (12aa) | AH | 100 | 50 | [193] |
| SPO | NA | NINONWNANGNONLNFNONLNFNO-NH2 (22aa) | AH | 100 | 50 | [193] |
| Chex1-Arg20 amide (ARV-1502) | NA | H-Chex-Arg-Pro-Asp-Lys-Pro-Arg-Pro-Tyr-Leu-Pro-Arg-Pro-Arg-Pro-Pro-ArgPro-Val-Arg-NH2 | — | — | Reduction of bacterial load | [194] |
AH, alpha helical; BS, beta sheet; NA, unavailble; *, µM; aa, amino acid; NPs, nanoparticles.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



