
Submitted:
05 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
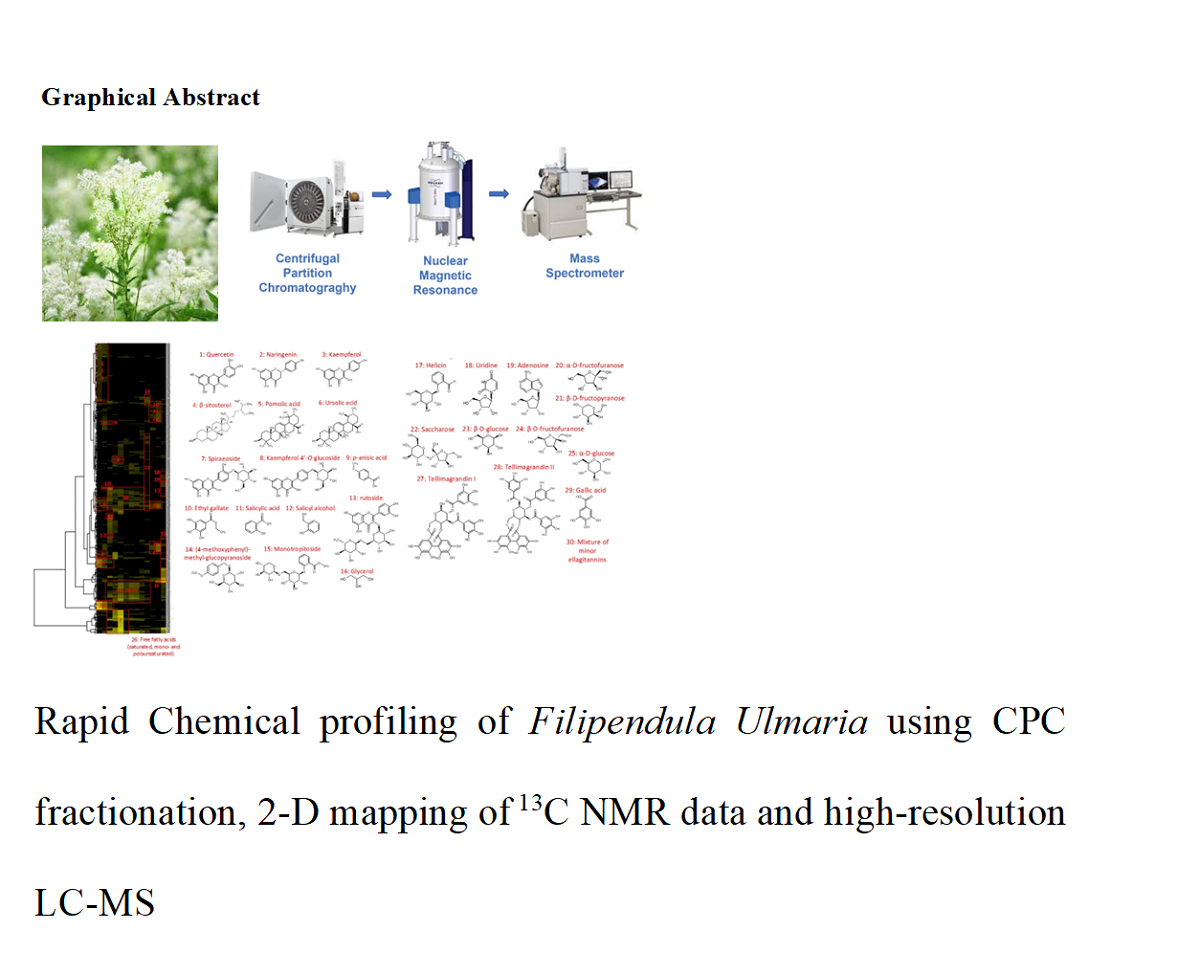
Abstract: Filipendula ulmaria, commonly known as meadowsweet, is a wild herbaceous flowering plant widely distributed in Europe. A range of salicylic acid derivatives and flavonol glycosides have been previously associated with the antirheumatic and diuretic properties of F. ulmaria. In the present work, an hydroalcoholic extract of F. ulmaria aerial parts was extensively profiled using an efficient NMR-based dereplication strategy. The approach involves fractionation of the crude extract by centrifugal partition chromatography (CPC), 13C NMR analysis of the fractions, 2D-cluster mapping of the whole NMR dataset and finally, structure elucidation using a natural metabolite database, validated by 2D NMR data interpretation and liquid chromatography coupled to mass spectrometry. The chemodiversity of the aerial parts was quite broad, with 28 compounds unambiguously identified, belonging to various biosynthetic classes. The F.ulmaria extract and CPC fractions were screened for their potential to improve skin epidermal barrier function and also skin renewal properties using in vitro assays performed on Normal Human Epidermal Keratinocytes. Fractions containing quercetin, kaempferol glycosides, ursolic acid, pomolic acid, naringenin, β-sitosterol, and tellimagrandins I and II were found to be responsible for the up-regulation of genes related to skin barrier function, epidermal renewal and stress responses. This research is significant as it could provide a natural solution for improving hydration and skin renewal properties.
Keywords:
Filipendula ulmaria
; Centrifugal Partition Chromatography
; NMR-based dereplication
; Liquid Chromatography Mass Spectrometry
; Epidermal barrier renewal
1. Introduction
Filipendula ulmaria (L.) Maxim., syn. Spiraea ulmaria L. (Meadowsweet), a wild herbaceous flower belonging to the Rosaceae family, is largely distributed in wet European roads [1,2,3]. The medicinal use of F. ulmaria dates back to the late 16th and 17th centuries and has been well documented in the British Herbal Pharmacopoeia [5,6] as a stomachic, mild urinary antiseptic, antirheumatic, and antacid. In addition, the British Herbal Compendium describes the action of Filipendula herb as an anti-inflammatory. The herb and its flowers have been traditionally used as a diuretic and antirheumatic in the treatment of inflammatory diseases. A number of secondary metabolites belonging to phenolic and flavonol glycosides, namely, Spiraeoside, Hyperoside, Rutoside, Xyloglycoside of methyl salicylate, and the rare salicylic aldehyde have been previously reported in Filipendula species [8,9,10]. These secondary metabolites and related phytochemical structures could possibly be responsible for the biological properties, described above. Interestingly, recent studies have reported that α-hydroxy acids, salicylic acid and its derivatives can improve skin hydration by gently peeling the corneocytes of the upper epidermis, softening the skin, while displaying a significant decrease in stratum corneum cohesion and minimally disrupting the skin barrier to water diffusion [11,12,13]. The aim of the present study was to delve deeper into the chemical profile of F. ulmaria and particular focus on its ability to improve skin barrier functions. This research is significant as it could provide a natural and effective solution for improving skin hydration and skin renewal properties.
Recently, the rapid development of powerful analytical platforms based on nuclear magnetic resonance (NMR) and high-resolution mass spectrometry (MS), have enabled faster separation, characterization, detection and quantification of chemically diverse secondary metabolites [14]. Here, a powerful chemical profiling strategy was deployed based on a combination of chromatographic, analytical, and data mining tools. From the ariel parts of F. ulmaria, 13 fractions were produced by centrifugal partition chromatography (CPC) from which, 28 secondary metabolites were identified by nuclear magnetic resonance (NMR), hierarchical clustering analysis (HCA) and a natural metabolite database, followed by liquid chromatography mass spectrometry to complete the identification process. The fractions were then screened for their potential to improve skin epidermal barrier function and skin renewal properties using appropriate in vitro assays. Reported below are the results of our investigation of this chemically prolific European plant.
2. Results and Discussion
2.1. Chemical Profiling of the Filipendula ulmaria extract
The chemical profile of the hydroalcoholic extract of F. ulmaria aerial parts was deciphered using a dereplication strategy named “CARAMEL” (CARActérisation des MELanges in French), which is based on a metabolomic workflow combining centrifugal partition chromatography, nuclear magnetic resonance, and computational treatments [15]. This strategy enables the direct identification of simplified mixtures of secondary metabolites (CPC fractions) without the need to purify individual constituents.
As a first step, F. ulmaria extract was fractionated by CPC, to produce a series of fractions without loss of biomass during separation. For this purpose, a biphasic solvent system of medium polarity composed of methyl ter-butyl ether, acetonitrile, and water (4/1/5, v/v) was selected. This system was employed in the ascending mode to tentatively separate F. ulmaria specialized metabolites during a single elution step while retaining the most hydrophilic compounds (mainly simple sugars) within the CPC column. A total of 13 final fractions were obtained based on TLC profile similarities (Figure 1).
The total recovery was estimated at >94.6 %. Fractions F01 to F10, recovered during the elution step represented 41.4 % of the injected F. ulmaria extract and exhibited a very diverse chemical diversity. The most polar fractions F11-F13 obtained at the end of the fractionation process by extrusion of the CPC column, represented 53.2 % of the extract mass (w/w). In a second step, all CPC fractions were directly analyzed by 1D and 2D NMR. The mass of each recovered fraction was largely sufficient to achieve NMR analyses (with 15 mg each except 8 mg for fraction F10), while also keeping aside fractions for biological evaluation. Automatic peak picking was performed on 13C NMR spectra, and the collected peaks were aligned across the fraction series using a bucketing strategy. The resulting table was made of 13 columns corresponding to the CPC fractions and 307 rows corresponding to the chemical shift buckets (Δ 0.3 ppm) for which a 13C peak was detected in at least one fraction. This table was subjected to Hierarchical Clustering Analysis (HCA) for the recognition of similarities between the 13C fingerprints obtained in the successive CPC fractions. In this way, 13C NMR signals belonging to the same compounds were aggregated as “chemical shift clusters” in a heat map which is illustrated in Figure 2.
An in-house database containing predicted NMR chemical shifts of natural metabolites was used to tentatively identify the metabolites corresponding to each cluster of the heat map. Database proposals were confirmed for all atom positions, and when experimental data did not exactly match with predicted data, the chemical structures detected in the extract were completed, or further elucidated by manual interpretation of 2D NMR data (HSQC, HMBC and COSY spectra). The NMR results were also cross-checked with LC-MS analyses to reinforce the identification and characterization process (Table 1).
Clusters 1, 2 and 3 were detected in the least polar fractions F01-F02 and were assigned to the non-glycosylated flavonoids quercetin (1), naringenin (2), and kaempferol (3), respectively. LC-MS analysis of the crude extract also revealed molecular ions at m/z 301.0348, m/z 271.0606, and m/z 285.0399 with retention times corresponding to Rt 14.9, 16.1 and 16.2 min, respectively, thereby corresponding to their respective parent ions [M-H]-. These three flavonoids are well known metabolites of F. ulmaria flower extracts [16,17]. Cluster 4 corresponded to β-sitosterol (4), a very common sterol occurring in many plant species, and clusters 5 and 6 comprised NMR signals belonging to pentacyclic triterpenes which were unambiguously confirmed as ursolic acid (5) and pomolic acid (6) based on interpretation of 2D NMR data. These two triterpenes have been previously reported to be present in the roots and aerial parts of F. ulmaria [16,18]. Cluster 7 corresponded to a group of intense NMR chemical shifts, which were assigned to spiraeoside (7), a biologically active 4’-O-monoglycosilated flavonol typically occurring in F. ulmaria flowers [19,20]. This compound was largely present in the major fractions, F04 and F05 that together represented ~17 % of the crude extract dry weight, therefore identified as an important biomarker of F. ulmaria. Its presence was also confirmed by the LC-MS with an intense molecular ion m/z 463.0882 [M-H]- with a retention time of Rt 12.7 min. Cluster 8 corresponded to kaempferol 4’-O-glucoside (8), also detected in fractions F04 and F05 but as a minor chemical constituent. This compound was also detected by LC-MS at a retention time of Rt 13.0 min with m/z 447.0922 [M-H]-. Clusters 9 and 10 were detected in fractions F02-F03 and were therefore easily assigned to the phenolic acid derivatives, p-anisic acid (9) and ethyl gallate (10). Furthermore, Clusters 11 and 12 were detected in F03-F04 and corresponded to the closely related compounds salicylic acid (11) and salicyl alcohol (12), that are characteristic constituents of the genus Filipendula [21]. Cluster 13 was detected only in the polar fraction F10 and was assigned to rutoside (13), while clusters 14 to 16 were assigned to phenol glycosides, (4-methoxyphenyl)-methyl-glucopyranoside (14), monotropitoside (15), and helicin (16). The presence of rutoside in the extract was confirmed by LC-MS with the detection of a molecular ion corresponding to m/z 609.1450 [M-H]- and a retention time of Rt 11.0 min. Cluster 17 was identified as glycerol (17), while Clusters 18 and 19 that were detected in the last fraction F13 corresponded to two nucleosides, uridine (18) and adenosine (19) which are universal metabolites distributed in the cells of all living species. Clusters 20 to 25 were unambiguously assigned to mono- and disaccharides, α-D-fructofuranose (20), β-D-fructopyranose (21), saccharose (22), β-D-glucose (23), β-D-fructofuranose (24), and α-D-glucose (25). These simple sugars were the major constituents of the most polar fractions F11-F12 which together represented ~45 % of the extract dry weight. Among the sugars, only saccharose was detected by LC-MS with a retention time of Rt 2.2 min with the corresponding molecular ion observed at m/z 341.1089 [M-H]-. Clusters 26 and 27 were assigned to tellimagrandin I (26) and tellimagrandin II (27) which have been described to be biologically active constituents of several Filipendula species [19,22,23]. These ellagitannins were found to be highly concentrated in fractions F04-F07, which together represented ~23 % of the dry extract weight, and therefore were also significant constituents of the extract. Two abundant isomers of tellimagrandin I were detected by LC-MS in the extract with intense peaks at m/z 785.0839 and m/z 785.0840 [M-H]-, corresponding to Rt 8.0 min and 8.9 min, respectively, while Tellimagrandin II was confirmed also by LC-MS with a molecular ion peak at m/z 937.0955 [M-H]- and Rt 10.4 min. Free gallic acid (28) was also unambiguously identified by NMR in these fractions and detected by LC-MS at Rt 5.2 min with m/z 169.0137 [M-H]-. To summarize, a total of 28 metabolites were unambiguously identified in the F. ulmaria extract using the NMR-based CARAMEL dereplication platform. Additional minor metabolites were detected by LC-MS in the extract (Figure 3), including for instance, several isomers of mono-, di- and tri-galloyl hexosides, several glycosylated derivatives of quercetin and kaempferol, quinic acid, syringic acid, as well as a diversity of tannins tentatively assigned to chebulagic acid, casuarinin, rugosin A and rugosin D in accordance with phytochemical data related to F. ulmaria. As an interesting corollary, the secondary metabolites identified using the LC-MS platform assisted in the determination of their molecular formula, although, not fully confirmed through NMR assignments, one can still consider putative isomers as possible chemical structures.
2.2. Biological Results for the Crude Extract and CPC Fractions and Assignment of the Metabolites Responsible for Skin barrier function Improvement
In an attempt to further explore the cosmetic applications of F. ulmaria on epidermal skin barrier, the crude extract was evaluated for the expression of different sets of genes involved in epidermal proliferation, differentiation, barrier function, epidermal renewal and stress response in normal human epidermal keratinocytes. The selection of normal human epidermal keratinocytes was done to eliminate alterations in gene expression due to cellular senescence induced as a result of prolonged culturing of immortalized cell lines like HaCaT [24,25]. The F. ulmaria extract was found to be cytotoxic at concentrations above 0.002 g/L, and was therefore tested at the maximal safe concentration of 0.002 g/L and at a lower concentration of 0.0004 g/L. The extract showed moderate modulation of gene expressions at the selected concentrations. However, several secondary metabolites reported from F. ulmaria such as salicylic acid, quercetin and its glycosides, or tannins are known to affect the skin barrier function and epidermal regulation. Quercetin is known to down-regulate the Epiregulin Growth Factor Receptor (EGFR) expression levels [26,27,28], while kaempferol is known to enhance the Claudin 1 gene expression and enhance the intercellular tight junction capacity in the skin [29]. Naringenin and β-sitosterol are involved in Glutathione peroxidase regulation [30,31] and ursolic acid and pomolic acid are known to restore the skin barrier function of the epidermis by preventing the trans-epidermal water loss, differentiation of keratinocytes, regulation of peroxisome proliferator-activated receptor-α and promoting the synthesis of hyaluronic acid and collagen [32,33]. Tellimagrandins present in F. ulmaria extracts are also reported previously for enhancing the cornified envelope formation and fillagrin mRNA expression in the HaCaT Cell lines [34].
A total of 13 fractions were obtained from the crude F. ulmaria extract. The secondary metabolite distribution in the fractions is given in Figure 1. Fractions F1, F3, F4, F6 and F9 were then subjected to the Epidermal gene expression modulation tests in the Normal Human Epidermal Keratinocytes for Epidermal proliferation, differentiation, barrier function, Epidermal renewal and stress response. Fractions F2, F5, F7, F8, F10, F11, F12 and F13 were not subjected to these tests due to paucity of the fractions. Fractions F1, F3, F4, F6, F9 were evaluated at the maximal concentration of 1 g/L and at the minimal dose of 0.2 g/L and the modulation of gene expressions are shown in Figure 3.
Fraction F1 of the F. ulmaria extract (4% of the crude extract mass) was found to be rich in quercetin, naringenin, kaempferol, ursolic acid, pomolic acid and β-sitosterol as major constituents. As expected, this fraction showed strong modulation of cornifelin, a tight junction protein, as well as epiregulin, due to the presence of quercetin [32,33,34]. Additionally, it was found to enhance the expression of genes involved in hyaluronic acid synthesis 3 and heme oxygenase 1 at both minimal and maximal concentrations. These results demonstrate the high efficacy of this fraction in reinforcing the skin barrier function and renewing the epidermis. Moreover, fraction F1 showed a significant upregulation of the Glutathione peroxidase 2 gene, likely due to the presence of naringenin and β-sitosterol, which have been reported to be involved in regulating glutathione peroxidase activity [36,37,38]. Fraction F3, which mainly comprises the glycosides of quercetin (Spiraeoside) and kaempferol (kaempferol 4'-O-glucoside), along with salicylic acid, salicyl alcohol, ethyl gallate, p-anisic acid, and tellimagrandins, accounts for approximately 5.7% of the total crude extract of F. ulmaria. This fraction shows a similar response in gene expression modulation for barrier function reinforcement and epidermal renewal as fraction F1, but only at the maximal concentration of 1g/L. Its efficacy at the minimal dose was not significant. Fraction F4, which makes up 8% of the total crude extract, contains spiraeoside and tellimagrandins as major constituents, along with small amounts of salicylic acid, salicyl alcohol, and gallic acid. This fraction exhibits strong modulation of cornifelin, kallikrein-related peptidase 7, heparin-binding EGF-like growth factor, keratin 19, and keratin 10 genes at a lower concentration of 0.2 g/L. No significant activity was observed at the maximal dose for fractions containing spiraeoside and tellimagrandin, suggesting that these compounds are effective in keratinocyte differentiation, as well as in reinforcing and renewing the epidermal barrier. Fractions F6 and F9, which make up 3% and 2% of the crude extract, respectively, contain mainly tellimagrandins and a mixture of minor Ellagitannins. Both fractions demonstrated efficacy in epidermal renewal and restoration of barrier function at a lower concentration of 0.2 g/L, indicating that ellagitannins play an important role in their efficacy at lower concentrations. This finding is consistent with previously reported activity of tellimagrandins in promoting the formation of cornified envelope [39], as mentioned earlier in this research paper.
3. Materials and Methods
3.1. Materials and reagents
The ariel parts of F. ulmaria were collected in 2019 from the Somerset County in Southwest of United Kingdom and were authenticated by an external consultant. A Voucher specimen has been deposited at the Herbarium facility at L'Oreal (Advanced Research, Bangalore, India), under the Voucher specimen number ARI 032063/E/B-1/1. HPLC-MS-grade acetonitrile and methanol were purchased from Merck. Laboratory grade chemicals were obtained from Sigma-Aldrich Chemical Co. (St Louis, MO, USA) and Merck Millipore (Darmstadt, Germany). Milli-Q Integral 15 system (Merck Millipore) was used to prepare the HPLC-grade water.
3.2. Extraction Procedure
The ariel parts of F. Ulmaria were powdered using the IKA® Pilotina dry milling system and sieved through 100 µm mesh to afford a coarse powder. The resulting powder (200 g) was extracted using 70% aqueous ethanol (1:10 m/v) for three consecutive cycles at 60°C. After three cycles of extraction, each was filtered using GFD filter and the filtrate was combined under reduced pressure and concentrated to afford a dry extract with a yield of 60 g, containing 4.5% of Spiraeoside. The solid crude extract was further defatted using petroleum ether at 40°C for 1 h to afford a dry residue containing 6% Spiraeoside. To this crude extract, 2 L of 10% ethanol was added and the solution was filtered through hyflo supercell. The subsequent clear filtrate was then passed through a HP-20 macroporous resin and eluted with 10 bed volumes of demineralized water and further eluted using increasing percentage of ethanol ranging from 40% ethanol (5 BV), 50% ethanol (5 BV) and finally 100% ethanol. The 50% eluate was concentrated and dried, to afford a dry powder highly enriched in Spiraeoside.
3.3. Centrifugal Partition Chromatography (CPC)
CPC was performed on a lab-scale FCPE300® column (Rousselet Robatel Kromaton, Annonay, France) of 301 mL capacity, containing seven circular partition disks and engraved with a total of 231 partition twin cells (≈ 1 mL each twin cell). The liquid phases were pumped with a KNAUER Preparative 1800 V7115 pump (Berlin, Germany). A two-phase solvent system (3 L in total) was prepared by mixing methyl ter-butyl ether, acetonitrile, and water in the proportions 4/1/5 (v/v) in a separating funnel. After decantation, the column was filled with the lower phase used as the stationary phase at 50 mL/min and 500 rpm. The column rotation speed was then set at 1200 rpm. The extract (1.015 g) was dissolved in a 80/20 (v/v) mixture of lower and upper phases and injected into the CPC column with a 35 mL loop. The upper phase used as the mobile phase was pumped at a flow rate of 20 mL/min in the ascending mode for 55 min, and then the column was extruded by switching the mode selection valve for 10 min. Fractions of 20 mL were collected over the whole experiment (elution and extrusion) by a Pharmacia Superfrac collector (Uppsala, Sweden), and combined according to their thin layer chromatography (TLC) profiles. TLC was performed with a CAMAG® Automatic TLC Sampler 4 (ATS4), a CAMAG® Automatic Developing Chamber 2 (ADC2), and a CAMAG® TLC Visualizer 2. Fractions were deposited on pre-coated silica gel 60 F254 Merck plates, eluted with the migration solvent system toluene/ethyl acetate/formic acid/acetic acid (30/70/11/11, v/v), and revealed at 254 nm, at 360 nm, and by spraying the dried plates with 50% H2SO4 and vanillin followed by heating. As a result, 13 final fractions were obtained, and their mass distribution and TLC profile are given in Figure 1.
3.4. NMR analyses and Metabolite Identification
All CPC fractions F01-F13 were dried under vacuum with a rotary evaporator. An aliquot (up to 15 mg when possible) was dissolved in 600 µL of DMSO-d6 and analyzed by 1H, 13C, HSQC, HMBC, and COSY NMR at 298 K on a Bruker Avance III 600 spectrometer (Karlsruhe, Germany) equipped with a cryogenic probe. The Bruker TopSpin 4.0.5 software was used for NMR data acquisition and processing. For 13C NMR analyses, a standard zgpg pulse sequence was used with an acquisition time of 0.9 s and a relaxation delay of 3 s. For each sample, 512 scans were co-added to obtain a satisfactory signal-to-noise ratio. The spectral width was 240 ppm and the receiver gain was set to the highest possible value. Spectra were manually phased, baseline corrected, and calibrated on the central resonance of DMSO-d6 (δ 39.8 ppm). The absolute intensities of 13C NMR signals were collected by automatic peak picking, and the peak list obtained for each fraction was exported as a text file. Then a bucketing was performed using a script written in Python, consisting in the division of 13C NMR spectral width into chemical shift buckets of 0.3 ppm and association of the absolute intensity of each peak to the corresponding bucket. The resulting table was submitted to HCA using the PermutMatrix 1.9.3 software (LIRMM, Montpellier, France) for data visualization. In parallel, a literature survey was performed to obtain the structures of a maximum of metabolites already reported in F.ulmaria (n ≈ 55). The 13C NMR chemical shifts of these metabolites were predicted (NMR Workbook Suite 2012, ACD/Labs, Ontario, Canada) and stored into an in-house database comprising ≈ 8300 chemical structures of natural molecules. The chemical shift clusters obtained by HCA were submitted to this database for metabolite identification. 2D NMR analyses (HSQC, HMBC, and COSY) were also interpretated to validate or complete the structural elucidation of the metabolites proposed by the database.
3.5. Liquid Chromatography – Mass Spectrometry analyses of the extract (LC/MS)
The crude F. ulmaria extract was also analyzed by LC-MS in the negative ion mode to tentatively confirm the identification of a maximum of metabolites. A 5 mg aliquot of each dried fraction was dissolved in 1 mL of MeOH/H2O (1:1, v/v) and analysed on a SYNAPT G2-Si High Resolution Mass Spectrometer from Waters (St Quentin en Yvelines, France). The chromatographic separation was performed at 1 mL/min on a RP18 reversed-phase column (Uptisphere C-18 ODB 150*4.6mm, 5µm, Interchim) with an injection volume of 5 µL. The column temperature was maintained at 35 °C. The solvents were formic acid 0.1% in ultrapure water (A) and formic acid 0.1% in LC/MS grade acetonitrile (B). The gradient started at 0% (B), increased up to 26% (B) in 9.9 min, to 65% (B) at 18.5 min, and then reached 100% (B) at 18.7 min and remained for 5 min. After that, the gradient returned back to 0% (B) in 1 min and remained for 2 min. The electrospray source operated in the negative mode with the following parameters: capillary voltage 3 kV, sampling cone 40 V, extraction cone 4 V, source temperature 150 °C, desolvation 650 L/h, collision energy 4 V. Accurate mass was ensured using a solution of leu-enkephalin as a standard compound in the internal lockmass. Ions were detected from m/z 50 to 2000 with scans of 0.2 s. Data were processed with the MassLynx software from Waters. The resulting BPI chromatogram and summarized LC/MS data are given in Figure 4.
3.6. Biological Assays
A Roche LightCycler480 instrument for 384-well microplates was used for this evaluation test of gene expression involved in keratinocytes epidermal physiology. Normal Human Epidermal Keratinocytes were grown and amplified to produce cells for the evaluation. 48-well microplates were seeded with the cells (50,000 cells per well) and incubated for 48 hours in a temperature, humidity and CO2 controlled environment. Samples were added on the cells while renewing the culture medium and were further incubated for 24 hours. After incubation, the cells were washed and frozen at -80°C to preserve the RNA. The RNAs were then extracted, quantified and their quality was checked before performing their reverse transcription into cDNA. An RT-qPCR was finally performed for each experimental condition for the quantification of the expression of a set of 16 selected genes related to the barrier function (Claudin 1, Cornifelin, Desmoglein 1, Kallikrein-related peptidase 7, Tight junction protein 1), epidermal renewal (Epiregulin, Hyaluronic acid synthase 3, Heparin-binding EGF-like growth factor, Keratin 19), keratinocyte differentiation (Keratin 10, Small prolin-rich protein A1, Transglutaminase 1) and stress response (Glutathione peroxidase 2, Heme oxygenase 1) in keratinocytes. The gene expression was measured on the highest non-toxic dose of each tested sample with a maximal dose of 0.2 g/L or 0.2 mM. The maximum non-cytotoxic dose was determined prior to the gene expression testing at a dose of 0.2 g/L using a biological model under the same incubation conditions. All samples were evaluated at the same concentration in addition to a five times lower dose in a one-step protocol. The fold changes (FC) were calculated after a double normalization against the housekeeping genes and non-treated condition. Fold changes of gene expression were considered as modulated over 1.5 (induction) or under 0.5 (repression).
4. Conclusions
The application of CPC fractionation, 13C NMR de-replication, and liquid chromatography hyphenated with mass spectrometry proved to be a valuable strategy for rapid identification of 28 secondary metabolites from the ariel parts of F. ulmaria. This study showcased the agile capability of our metabolomics workflow as a sensitive and convenient way for conducting both qualitative and quantitative analysis by using the same instrumentation. This new analytical approach offers a disruptive, innovative route for discovering and developing new cosmetic bio-actives with biologically active secondary metabolites from F. Ulmaria. By subjecting CPC fractions of F. Ulmaria to in vitro screening, quercetin, kaempferol glycosides, ursolic acid, pomolic acid, naringenin, β-sitosterol, and tellimagrandins I and II were identified as key secondary metabolites, involved in up-regulating genes related to skin barrier function, epidermal renewal, and stress responses in normal human epidermal keratinocytes. This research is significant as it could provide a natural solution for improving skin hydration with epidermal renewal properties. This new screening strategy, without the need for any purification step should find widespread application where plants have not been widely studied for the discovery of new natural products, as these platforms serve as a future strategic discovery tool.
Author Contributions
Conceptualization, Steve Thomas Pannakal, Joan Eilstein, Jane Hubert; Methodology and Software, Jane Hubert, Alexis Kotland; Validation, Amelie Gueguiniat-Prevot, Franck Juchaux; Formal Analysis and Investigation, Steve Thomas Pannakal, Joan Eilstein, Jane Hubert; Writing – Original Draft Preparation, Steve Thomas Pannakal; Writing – Review & Editing, Arpita Prasad; Visualization, Joan Eilstein, Jane Hubert; Project Administration, Floriane Beaumard, Ganapaty Seru, Sherluck John and Dhimoy Roy; Funding Acquisition, Sherluck John and Dhimoy Roy.
Funding
Financial support was provided by L’Oreal for the project orchestra no:2019-5126/A of the Innovative Ingredients department (D2I).
Acknowledgments
The Institute of Molecular Chemistry of the University of Reims Champagne Ardenne is also acknowledged for providing access to the analytical platform PlAneT for NMR and high-resolution liquid chromatography mass spectrometry analyses. The authors would like to warmly thank Dr. Santhan for sourcing of the ariel parts of Filipendula ulmaria from Europe.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are available from the authors.
Abbreviations
| CPC | Centrifugal Partition Chromatography |
| NMR | Nuclear Magnetic Resonance |
| HCA | Hierarchical Clustering Analysis |
| BV | Bed Volume |
| TLC | Thin Layer Chromatography |
| HSQC | Heteronuclear single quantum coherence |
| HMBC | Heteronuclear multiple-bond coherence |
| COSY | Correlated Spectroscopy |
| LC-MS | Liquid Chromatography Mass Spectra |
| FC | Fold Changes |
| CLDN1 | Claudin 1 |
| CNFN | Cornifelin |
| DSG1 | Desmoglein 1 |
| KLK7 | Kallikrein-related peptidase 7 |
| TJP1 | Tight junction protein 1 |
| EREG | Epiregulin |
| HAS3 | Hyaluronic acid synthase 3 |
| HBEGF | Heparin-binding EGF-like growth factor |
| KRT19 | Keratin 19 |
| KRT10 | Keratin 10 |
| SPRR1A | Small prolin-rich protein A1 |
| TGM1 | Transglutaminase 1 |
| GPX2 | Glutathione peroxidase 2 |
| HMOX1 | Heme oxygenase 1 |
| PPAR | peroxisome proliferator-activated receptor-α |
References
- Ball P.W., I.N. Miller, T.G. Tutin, V.H. Heywood, N.A. Burges, D.M. Moore, D.H. Valentine, S.M. Walters, D.A. Webb, Flora Europaea, Vol 2. Roseaceae to Umbelliferae, Cambridge University Press, London, pp. 6-7.
- Šarić-Kundalić B., C. Dobeš, V. Klatte-Asselmeyer, J. Saukel, Ethnobotanical survey of traditionally used plants in human therapy of east, north and north-east Bosnia and Herzegovina, Journal of Ethnopharmacology, 133 (2011) 1051-1076. [CrossRef]
- Vogl S., P. Picker, J. Mihaly-Bison, N. Fakhrudin, A.G. Atanasov, E.H. Heiss, C. Wawrosch, G. Reznicek, V.M. Dirsch, J. Saukel, B. KopP, Ethnopharmacological in vitro studies on Austria’s folk medicine-An unexplored lore in vitro anti-inflammatory activities of 71 Austrian traditional herbal drugs, Journal of Ethnopharmacology, 149 (2013) 750-771. [CrossRef]
- Halkes S.B.A., Filipendula ulmaria - A study on the immunomodulatory activity of extracts and constituents, PhD Thesis, University of Utrecht (1998).
- British Herbal Pharmacopoeia, British Herbal Medicine Association, (1983) 91-92.
- British Herbal Pharmacopoeia, British Herbal Medicine Association, 1 (1990) 65-6.
- Bradley PR editor. British Herbal Compendium, British Herbal Medicine Association, 1 (1992) 158-160.
- Bespalov, V.G., V.A. Alexandrov, A.L. Semenov, E.G. Kovan’ko, S.D. Ivanov, G.I. Vysochina, V.A. Kostikova, D.A. Baranenko, The inhibitory effect of meadowsweet (Filipendula ulmaria) on radiation-induced carcinogenesis in rats. International Journal of Radiational Biology, 93 (2018) 394–401. [CrossRef]
- Zeylstra, H., Filipendula ulmaria Br, Journal of Phytotherapy, 5 (1998) 8-12.
- Filipendulaeulmaria herba – Meadowsweet, ESCOP Monographs, European Scientific Cooperative on Phytotherapy, editor, Thieme, 2 (2003,) 157-161.
- Berardesca E., F. Distante, G. P. Vignoli, C. Oresajo, and B. Green, Alpha hydroxyacids modulate stratum corneum barrier function, British Journal of Dermatology, 137 (1997) 934–938. [CrossRef]
- Bashir S. J., F. Dreher, A. L. Chew, H. Zhai, C. Levin, R. Stern, H. I. Maibach, Cutaneous bioassay of salicylic acid as a keratolytic, International Journal of Pharmaceutics, 292 (2005) 187–194. [CrossRef]
- Ademola J., C. Frazier, S. J. Kim, C. Theaux, and X. Saudez, Clinical evaluation of 40% urea and 12% ammonium lactate in the treatment of xerosis, The American Journal of Clinical Dermatology, 3 (2002) 217–222. [CrossRef]
- Thomas Pannakal S., Eilstein J., Prasad A., et al. Comprehensive characterization of naturally occurring antioxidants from the twigs of mulberry (Morus alba) using on-line high-performance liquid chromatography coupled with chemical detection and high-resolution mass spectrometry. Phytochemical Analysis. 2022;33(1):105-114. [CrossRef]
- Hubert, J., J.M. Nuzillard, S. Purson, M. Hamzaoui, N. Borie, R. Reynaud, J.H. Renault, Identification of natural metabolites in mixture: a pattern recognition strategy based on (13)C NMR, Analytical Chemistry, 86 (2014) 2955-2962. [CrossRef]
- Bijttebier S., Auwera A., Voorspoels S., Noten B., Hermans N., Pieters L., ApersS., A First Step in the Quest for the Active Constituents in Filipendula ulmaria (Meadowsweet): Comprehensive Phytochemical Identification by Liquid Chromatography Coupled to Quadrupole-Orbitrap Mass Spectrometry, Planta Medica, 82 (2016) 559-572. [CrossRef]
- Gainche, M., Ogeron, C., Ripoche, I., Senejoux, F., Cholet, J., Xanthine Oxidase Inhibitors from Filipendula ulmaria (L.) Maxim. and Their Efficient Detections by HPTLC and HPLC Analyses, Molecules, 26 (2021) 1939-1950. [CrossRef]
- Halles, S.B.A., Filipendula ulmaria – a study on the immunomodulatory activity of extracts and constituents [dissertation] Universiteit Utrecht, (1998).
- Samardžić, S., J. Arsenijević, D. Božić, M. Milenković, V. Tešević, Z. Maksimović, Antioxidant, anti-inflammatory and gastroprotective activity of Filipendula ulmaria (L.) Maxim. and Filipendula vulgaris Moench, Journal of Ethnopharmacology, 213 (2018) 132-137. [CrossRef]
- Katanic, J., T. Boroja, V. Mihailovic, S. Nikles, S.P. Pan, G. Rosic, D. Selakovic, J. Joksimovic, S. Mitrovic, R. Bauer, In vitro and in vivo assessment of meadowsweet (Filipendula ulmaria) as anti-inflammatory agent, Journal of Ethnopharmacology, 193 (2016) 627-636.
- Harbourne, N., J.C. Jacquier, D. O’Riordan, Optimisation of the aqueous extraction conditions of phenols from meadowsweet (Filipendula ulmaria L.) for incorporation into beverages, Food Chemistry, 116 (2009) 722-727.
- Nitta, Y., H. Kikuzaki, T. Azuma, Y. Ye, M. Sakaue, Y. Higuchi, H. Komori, H. Ueno, Inhibitory activity of Filipendula ulmaria constituents on recombinant human histidine decarboxylase, Food Chemistry, 138 (2013) 1551-1556.
- Pukalskienė, M., Venskutonis, R., Pukalskas, A., Phytochemical Characterization of Filipendula ulmaria by UPLC/Q-TOF-MS and Evaluation of Antioxidant Activity, Records of Natural Products 9 (2015) 451-455.
- Rockwell G. A., Johnson, G., Sibatani, A., In vitro senescence of human keratinocyte cultures. Cell Structure & Functions, 12 (1987) 539–548.
- Katayama, S., Skoog, T., Jouhilahti, E., Siitonen, H. A., Nuutila, K., Gene expression analysis of skin grafts and cultured keratinocytes using synthetic RNA normalization reveals insights into differentiation and growth control, BMC Genomics 16 (2015) 476-489. [CrossRef]
- Jung, J. H., Lee J. O., Kim, J. H., Lee, S. K., You G. A., Park, S. H., Park, J. M., Kim, E. K., Suh, P. G., An, J. K., Kim, H. S., Quercetin Suppresses HeLa Cell Viability via AMPK-Induced HSP70 and EGFR Down-Regulation, Journal of Cellular Physiology, 223 (2010) 408–414. [CrossRef]
- Huang, Y.T., Hwang, J. J., Effects of luteolin and quercetin, inhibitors of tyrosine kinase, on cell growth and metastasis-associated properties in A431 cells overexpressing epidermal growth factor receptor, British Journal of Pharmacology, 128 (1999) 999 – 1010.
- Cuevas, M. J., Tieppo, J., Marroni, N. M., Suppression of Amphiregulin/Epidermal Growth Factor Receptor Signals Contributes to the Protective Effects of Quercetin in Cirrhotic Rat, Journal of Nutrition, 141 (2011) 1299–1305. [CrossRef]
- Kim, J., Cho, N., Kim, E. M., Park, K. S., Cudrania Tricuspidata Leaf Extracts and Its Components, Chlorogenic Acid, Kaempferol, And Quercetin, Increase Claudin 1 Expression in Human Keratinocytes, Enhancing Intercellular Tight Junction Capacity, Applied Biological Chemistry, 63 (2020) 23. [CrossRef]
- Al-Roujayee, A. S., Naringenin improves the healing process of thermally-induced skin damage in rats, Journal of International Medical Research, 45 (2017) 570–582. [CrossRef]
- Ferreira, M. S., Lobo, J. M. S., Almeida, I. F., Sensitive skin: Active ingredients on the spotlight, International Journal of Cosmetic Science, 44 (2022) 56–73. [CrossRef]
- Lim, S. W., Hong, S. P., Jeong, S. W., Simultaneous effect of ursolic acid and oleanolic acid on epidermal permeability barrier function and epidermal keratinocyte differentiation via peroxisome proliferator-activated receptor-α, Journal of Dermatology, 34 (2007) 625–634. [CrossRef]
- Tan, H., Sonam T., Shimizu, K., The Potential of Triterpenoids from Loquat Leaves (Eriobotrya japonica) for Prevention and Treatment of Skin Disorder, International Journal of Molecular Science, 18 (2017) 1030. [CrossRef]
- Yin, J., Hwang, I. H., and Lee, M. N., Anti-acne vulgaris effect including skin barrier improvement and 5α-reductase inhibition by Tellimagrandin I from Carpinus tschonoskii, BMC Complementary and Alternative Medicine, 19 (2019) 323. [CrossRef]
- Kim, J., Cho,N., Kim, E. M., Park, K. S., Cudrania Tricuspidata Leaf Extracts And Its Components, Chlorogenic Acid, Kaempferol, And Quercetin, Increase Claudin 1 Expression In Human Keratinocytes, Enhancing Intercellular Tight Junction Capacity, Applied Biological Chemistry, 63 (2020) 23. [CrossRef]
- Al-Roujayee, A. S., Naringenin improves the healing process of thermally-induced skin damage in rats, Journal of International Medical Research, 45 (2017) 570–582. [CrossRef]
- Ferreira, M. S., Lobo, J. M. S., Almeida, I. F., Sensitive skin: Active ingredients on the spotlight, International Journal of Cosmetic Science, 44 (2022) 56–73. [CrossRef]
- Lim, S. W., Hong, S. P., Jeong, S. W., Simultaneous effect of Ursolic acid and Oleanolic acid on Epidermal permeability barrier function and epidermal keratinocyte differentiation via peroxisome proliferator-activated receptor-α, Journal of Dermatology, 34 (2007) 625–634. [CrossRef]
- Yin, J., Hwang, I. H., and Lee, M. N., Anti-acne vulgaris effect including skin barrier improvement and 5α-reductase inhibition by tellimagrandin I from Carpinus tschonoskii, BMC Complementary and Alternative Medicine, 19 (2019) 323. [CrossRef]
Figure 1.
HPTLC profile of the 13 CPC fractions - a) 254 nm; b) 366 nm; c) visible after vanillin/H2SO4 reagent spraying.
Figure 1.
HPTLC profile of the 13 CPC fractions - a) 254 nm; b) 366 nm; c) visible after vanillin/H2SO4 reagent spraying.
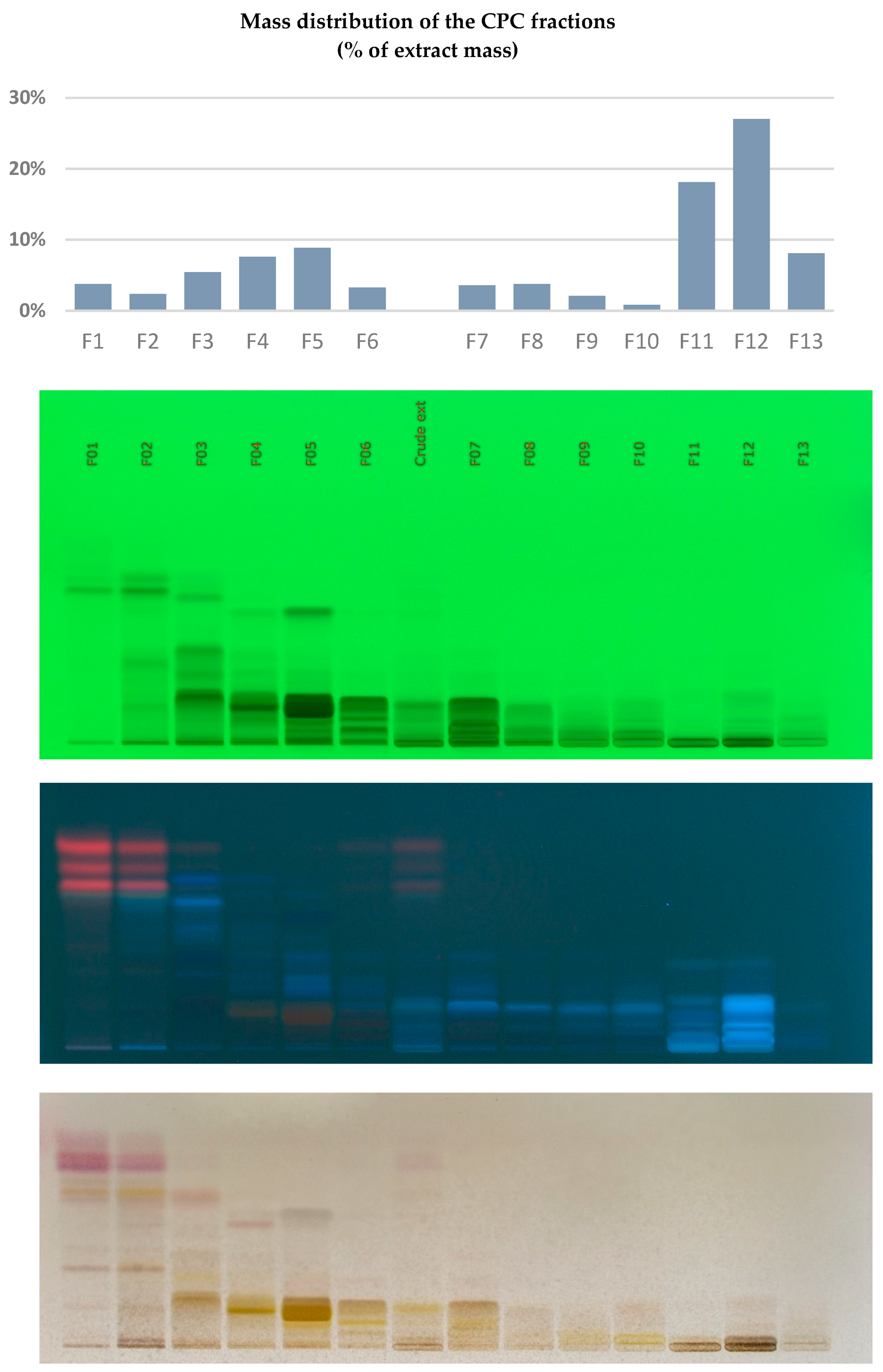
Figure 2.
Iterative Cluster Analysis using 13C NMR Resonance signals from CPC fractions of the Filipendula ulmaria extract, and identification of 28 Secondary Metabolites.
Figure 2.
Iterative Cluster Analysis using 13C NMR Resonance signals from CPC fractions of the Filipendula ulmaria extract, and identification of 28 Secondary Metabolites.
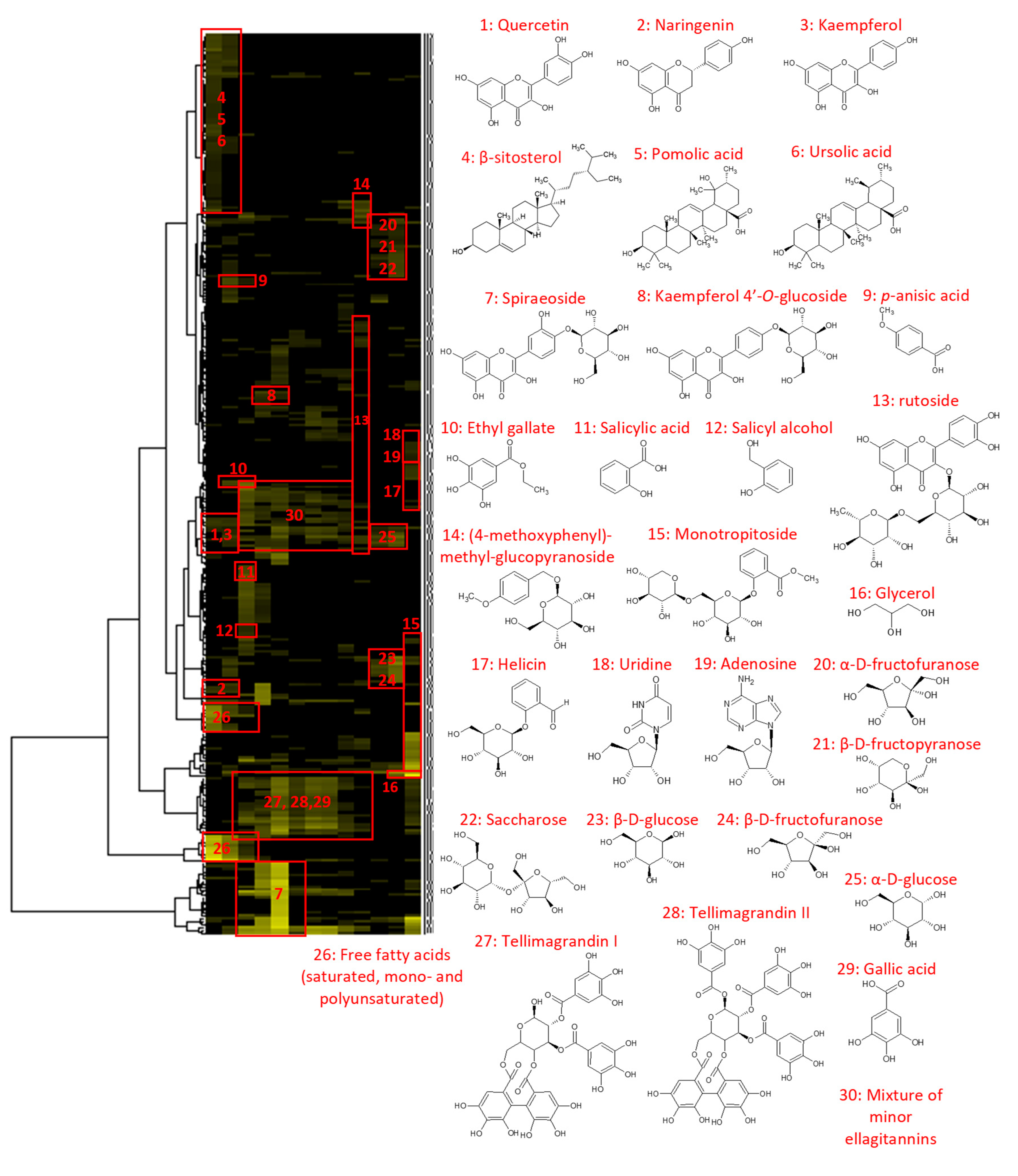
Figure 3.
Effect of F. ulmaria base extract, Fractions F01, F03, F04, F06 and F09 on Epidermal barrier function, Epidermal renewal, Keratinocyte differentiation and Stress response: (A) RT-qPCR on CNFN*; (B) RT-qPCR on EREG*; (C) RT-qPCR on HAS3*; (D) RT-qPCR on KRT10*; (E) RT-qPCR on GPX2* and (F) RT-qPCR on HMOX1*. * All evaluations were done in duplicates and therefore the P-values could not be calculated.
Figure 3.
Effect of F. ulmaria base extract, Fractions F01, F03, F04, F06 and F09 on Epidermal barrier function, Epidermal renewal, Keratinocyte differentiation and Stress response: (A) RT-qPCR on CNFN*; (B) RT-qPCR on EREG*; (C) RT-qPCR on HAS3*; (D) RT-qPCR on KRT10*; (E) RT-qPCR on GPX2* and (F) RT-qPCR on HMOX1*. * All evaluations were done in duplicates and therefore the P-values could not be calculated.
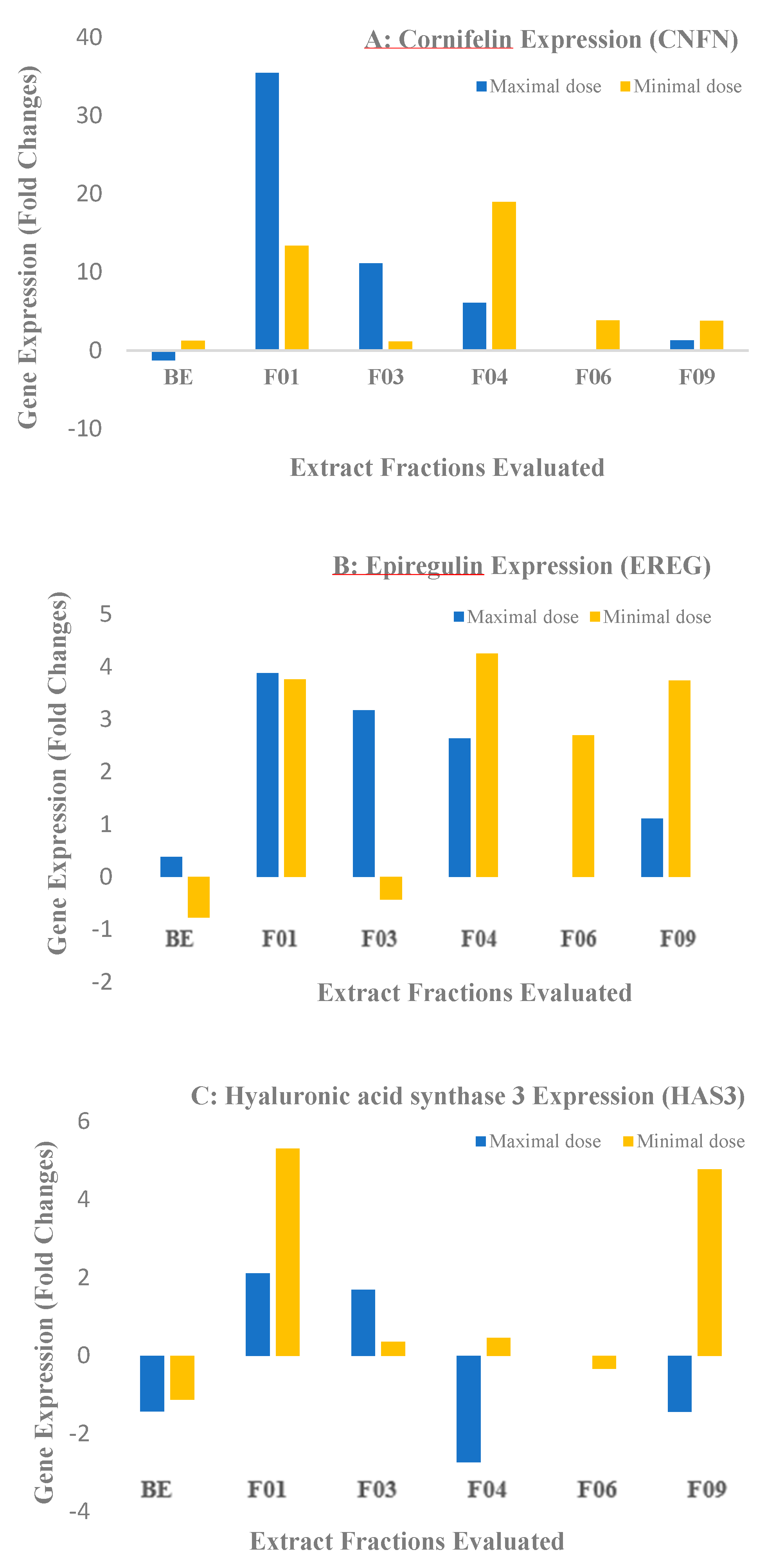
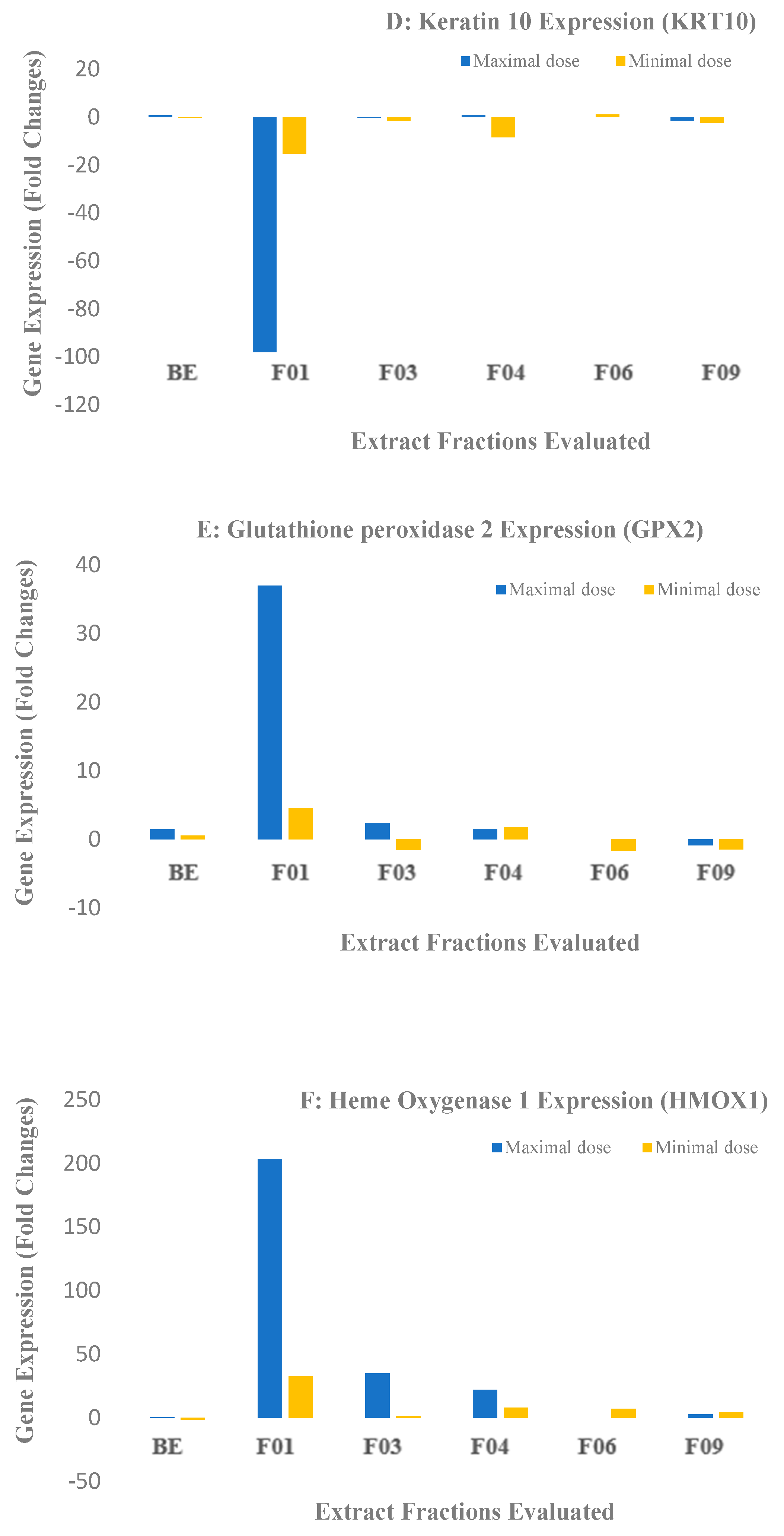
Figure 4.
LC/MS data of the crude extract of Filipendula Ulmaria in negative ion mode.
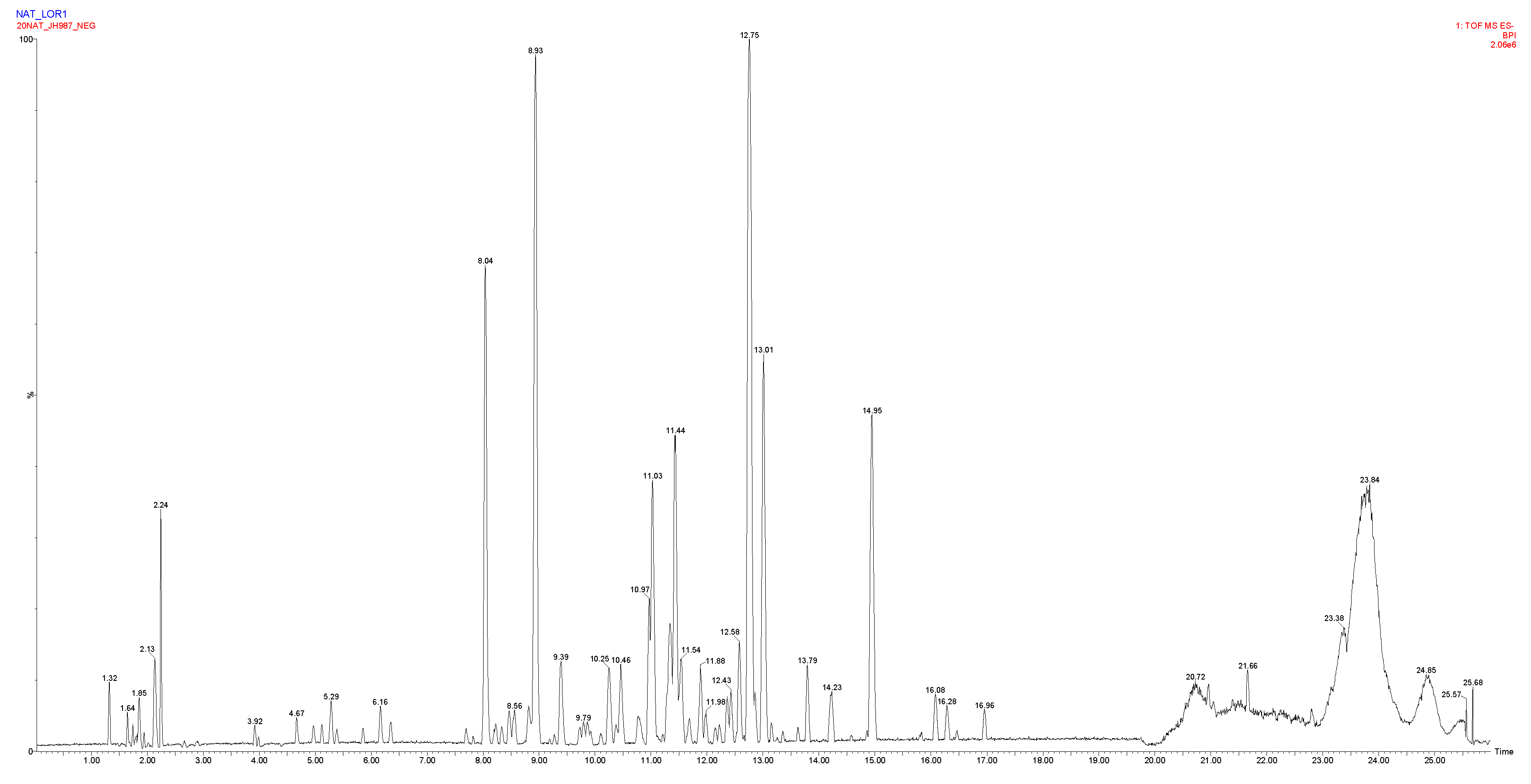

Table 1.
Overview of LC/MS Analytical descriptors relevant to the Identification of Secondary metabolites from the Crude Extract of Filipendula Ulmaria.
Table 1.
Overview of LC/MS Analytical descriptors relevant to the Identification of Secondary metabolites from the Crude Extract of Filipendula Ulmaria.
| Retention time (min) | Observed m/z | Elemental composition | Δppm | Tentative identification |
|---|---|---|---|---|
| 1.6 | 285.0815 | C9H17O10 | -2.5 | Not assigned |
| 1.8 | 195.0505 [M-H]- | C6H11O7 | 0.0 | Hexonic acid |
| 1.9 | 191.0555 [M-H]- | C7H11O6 | -0.5 | Quinic acid |
| 2.2 | 341.1089 [M-H]- | C12H21O11 | 1.5 | Saccharose* |
| 4.6 | 331.0664 [M-H]- | C13H15O10 | 0.0 | Mono-O-galloyl-hexoside isomer 1 |
| 4.9 | 331.0665 [M-H]- | C13H15O10 | -0.3 | Mono-O-galloyl-hexoside isomer 2 |
| 5.1 | 339.1292 | C13H23O10 | 0.3 | Not assigned |
| 5.2 | 169.0137 [M-H]- | C7H5O5 | 0.0 | Gallic acid* |
| 5.4 | 483.0783 [M-H]- | C20H19O14 | 1.7 | Di-O-galloyl-hexoside isomer 1 |
| 5.8 | 331.0667 [M-H]- | C13H15O10 | 0.6 | Mono-O-galloyl-hexoside isomer 3 |
| 6.1 | 483.0775 [M-H]- | C20H19O14 | 0.2 | Di-O-galloyl-hexoside isomer 2 |
| 6.3 | 315.0715 [M-H]- | C13H15O9 | -0.3 | Dihydroxybenzoic acid O-hexoside |
| 7.7 | 319.0423 [M-H]- | C15H11O8 | -9.7 | Dihydromyricetin |
| 7.8 | 483.0775 [M-H]- | C20H19O14 | 0.0 | Di-O-galloyl-hexoside isomer 3 |
| 8.0 | 785.0839 [M-H]- | C34H25O22 | 0.3 | Tellimagrandin I* or isomer |
| 8.2 | 635.0889 [M-H]- | C27H23O18 | 0.8 | Tri-O-galloyl-hexoside |
| 8.3 | 451.1010 [M-H]- | C24H19O9 | -4.2 | Coumaroylepigallocatechin |
| 8.4 | 375.0694 191.0556 quinic acid fragment |
C18H15O9 C7H11O6 |
-5.9 0.0 |
Not assigned |
| 8.5 | 289.0714 909.0999, 785.0842, 454.0461 |
C15H13O6 | 0.7 | Catechin |
| 8.8 | 953.0895 [M-H]- 909.0999 [M-COOH]- 785.0837, 465.0367, 454.0460 |
C41H29O27 C40H29O25 C34H25O22 |
-0.1 0.0 |
Chebulagic acid or isomer |
| 8.9 | 785.0840 [M-H]- | C34H25O22 | 0.4 | Tellimagrandin I* or isomer |
| 9.3 | 319.0431 | C15H11O8 | Not assigned | |
| 9.7 | 339.0718 | C15H15O9 | 0.6 | Not assigned |
| 9.8 | 359.0745 337.0925 coumaroylquinic acid 191.0556 quinic acid fragment |
C18H15O8 C16H17O8 C7H11O6 |
Not assigned | |
| 9.9 | 785.0845 [M-H]- 481.1118, 491.1403, 625.1407 |
C34H25O22 |
1.0 | Minor isomer of tellimagrandin I |
| 10.1 | 935.0803 [M-H]- 467.0357 [M-H-3galloyl]- |
C41H27O26 |
1.3 | Casuarinin or Casuarictin |
| 10.2 | 1105.1012 [M-H]- 1061.1110 fragment of Rugosin D 936.0874 [M-2H]2- 530.0513 fragment of rugosin A 541.0423 |
C48H33O31 C47H33O29 |
0.5 0.2 |
Rugosin A Rugosin D |
| 10.4 | 937.0955 [M-H]- 959.0774, 479.0345, 468.0435 |
C41H29O26 | 0.9 | Tellimagrandin II* |
| 10.8 | 935.0800 [M-H]- 787.1003 [M-H-galloyl]- 467.0357 [M-H-3galloyl]- 303.0485 [M-H-4galloyl]- |
C41H27O26 | 1.0 | Casuarinin or Casuarictin |
| 10.9 | 687.3029 [M-H]- | xx | xx | Not assigned |
| 11.0 | 609.1450 [M-H]- | C27H29O16 | -1.0 | Rutoside* |
| 11.3 | 197.0454 [M-H]- | C9H9O5 | 2.0 | Syringic acid |
| 11.4 | 463.0877 [M-H]- | C21H19O12 | 0.0 | Quercetin O-hexoside isomer 1 |
| 11.5 | 463.0876 [M-H]- 301.0348 quercetin fragment |
C21H19O12 | -0.2 | Quercetin O-hexoside isomer 2 |
| 11.9 | 593.1505 [M-H]- 1087.0900 [2M-H]- 285.0396 kaempferol fragment |
C27H29O15 | -0.2 | Kaempferol O-hexoside-rhamnoside |
| 11.9 | 433.0771 [M-H]- | C20H17O11 | 0.0 | Quercetin O-pentoside |
| 12.1 | 447.0930 [M-H]- | C21H19O11 | 0.0 | Quercetin O-rhamnoside |
| 12.2 | 477.1034 [M-H]- | C22H21O12 | 0.2 | Methyl-quercetin O-hexoside |
| 12.3 | 433.0772 [M-H]- 301.0353 quercetin fragment |
C20H17O11 | 0.2 | Quercetin O-pentoside |
| 12.4 | 447.0927 [M-H]- | C21H19O11 | 0.0 | Quercetin O-rhamnoside |
| 12.5 | 477.1031 [M-H]- | C22H21O12 | -0.4 | Methyl-quercetin O-hexoside |
| 12.7 | 463.0882 301.0353 quercetin fragment |
C21H19O12 | 1.1 | Spiraeoside* (Quercetin O-hexoside isomer 3) |
| 12.8 | 601.0827 [M-H]- 301.0347 quercetin fragment |
C27H21O16 | -0.5 | Quercetin O-galloyl-pentoside |
| 13.0 | 447.0922 [M-H]- 285.0389 kaempferol fragment |
C21H19O11 | -1.1 | Kaempferol 4’-O-glucoside* |
| 13.2 | 519.1136 465.1031 |
C24H23O13 C21H21O12 |
-0.6 -0.4 |
Not assigned |
| 13.4 | 615.0984 [M-H]- | C28H23O16 | -0.3 | Quercetin O-galloyl-hexoside |
| 13.8 | 585.0880 [M-H]- 301.0350 quercetin fragment |
C27H21O15 C15H9O7 |
0.9 0.7 |
Quercetin O-galloyl-arabinoside |
| 14.2 | 297.0399 [M-H]- | C16H9O6 | -7.1 | Not assigned |
| 14.9 | 301.0348 [M-H]- | C15H9O7 | 1.0 | Quercetin* |
| 16.1 | 271.0606 [M-H]- | C15H11O5 | 0.4 | Naringenin* |
| 16.2 | 285.0399 [M-H]- | C15H9O6 | 1.1 | Kaempferol* |
| 16.5 | 329.2329 | C18H33O5 | 0.3 | Tri-HOME (trihydroxyyoctadecenoic acid) |
| 16.9 | 287.222 | C16H31O4 | 1.5 | Dihydroxypalmitic acid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



