Submitted:
30 June 2023
Posted:
03 July 2023
You are already at the latest version
Abstract
Mast cells play a critical role in allergies and inflammation via secretion of numerous vasoactive, pro-inflammatory and neuro-sensitizing mediators. Secretion may utilize different modes that involve the cytoskeleton, but our understanding of the molecular mechanisms regulating secretion is still not well understood. We previously showed that the ability of the so called mast cell “stabilizer” disodium cromoglycate (cromolyn) to inhibit secretion from rat mast cells closely paralleled the phosphorylation of a 78 kDa protein, and subsequently showed this protein to be moesin, a member of the Ezrin/Radixin/Moesin (ERM) family of proteins, which are involved in linking cell surface-initiated signaling to the actin cytoskeleton. Unlike phosphorylation on the C-terminus Thr558 associated with activation of ERMs, including secretion from macrophages and platelets, we showed that phosphorylation of moesin during inhibition of secretion was on the N-terminal Ser56/74 and Thr66 residues. This phosphorylation pattern could lock moesin in its inactive state and remain inaccessible to bind to the Soluble NSF attachment protein receptors (SNAREs) and synaptosomal associated proteins (SNAPs). Using Confocal microscopic imaging, we showed moesin to colocalize with actin and cluster around secretory granules during inhibition of secretion, In conclusion, the phosphorylation pattern and localization of moesin may be important in the regulation of mast cell secretion and could be targeted for the development of effective inhibitors of secretion from mast cells.
Keywords:
ERMs
; flavonoids
; luteolin
; mast cells
; mediators
; moesin
; phosphorylation
; secretion
; SNAREs
; SNAPs
; tryptase
1. Introduction
Mast cells are specialized hemopoietic cells that play an important role in health,1 and in allergies,2-12 but also in innate and in adaptive immune processes,13-16 antigen presentation,16, 17 regulation of T-cell responses,18-20 autoimmunity,21 and inflammation,10, 22-25 in response to allergic and immunologic,4, 26, 27 but also non-allergic stress and toxic stimuli.10, 28 Mast cells are increased in number and are more reactive in mastocytosis26 and Mast Cell Activation syndrome (MCAS),26, 29, 30 but can also participate other disorders,4, 10, 31-33 including neurotrauma, neuroinflammatory and neurodegenerative diseases.34-36
Mast cells are located in tissues at the interface with the external environment37 such as eyes, nose, lungs, skin and gastrointestinal tract. However, perivascular mast cells also sense the blood vessel lumen by extending filopodia through endothelial gaps and bind circulating immunoglobulin E (IgE).38 Mast cells are well known for their involvement in allergic and anaphylactic reactions via activation of the high-affinity surface receptor for IgE (FcεRI). Multivalent allergen binding leads to aggregation of FcεRI and influx of calcium ions thus initiating a cascade of downstream events that involve phosphorylation of phosphatidyl inositol (IP3) and various Tyr kinases.39-42 In addition to allergens, mast cells are also stimulated by a variety of triggers that include drugs, foods, pathogens, and “danger signals,”26 as well as certain neuropeptides especially substance P (SP)43 via activation of their high-affinity receptors. Mast cells are also stimulated/activated by several cytokines, chemokines, and hormones such as corticotropin-releasing hormone (CRH), toxins and extreme external environmental changes.23, 36, 44, 45
Upon stimulation, mast cells secrete multiple biologically active mediators,46 some of which are preformed and stored in as many as 1,000 secretory granules per cell such as β-hexosaminidase (β-hex), heparin, histamine, tumor necrosis factor (TNF) and the serine proteases chymase and tryptase tryptase through rapid (1-5 min) degranulation by exocytosis.47 Histamine and tryptase are the main mediators associated with mast cells.48 Chymase is found in all mast cells, but unlike mucosal mast cells (MMCs) that contain only chymase, connective tissue mast cells (CTMCs) contain both chymase and tryptase. Even though these proteases are considered to be stored in the same secretory granules, there is evidence that this may not be necessarily true. For instance, serum tryptase was not elevated in many patients with MCAS28 or in cutaneous mastocytosis.49 In one paper, it was shown that IgE-mediated degranulation of primary murine MMCs and CTMCs released phenotypically different extracellular vesicle (EV) populations depending on the stimulus.50 In particular, unstimulated mast cells constitutively released CD9+ EVs, while degranulation was accompanied by the release of CD63+ EVs that contained different proteases.50
Mast cells also release newly-synthesized phospholipid products such as prostaglandin D2 (PGD2) and leukotrienes (LTs),51-53 as well as numerous de novo synthesized protein mediators 6-24 hours after stimulation such as interleukins,54 including interleukin-1beta (IL-1β), 55 IL-6,45, 56 IL-31, 57 IL-3355 and TNF.43
Mast cells can secrete their numerous mediators25, 47, 58 utilizing different signaling11, 59-62 and secretory60, 63 64 pathways sometimes referred to as the “secretome”.65 The secretory pathways include degranulation by exocytosis, compound exocytosis, piecemeal degranulation, transgranulation, directed degranulation, vesicular (differential) release of mediators, extracellular microvesicles (exosomes), nanotubules.66 and antibody-dependent “immunologic synapses for dedicated secretion67, 68 (Table 1). The term “secretion” is used in this review to include both degranulation by exocytosis, which is the main means of secretion of granule-stored mediators,69 as well as differential release via which chemokines and cytokines are released without degranulation.59 For instance, we first reported that serotonin,45, 52, 56 and later vascular endothelial growth factor (VEGF),70 and IL-6 45, 56 could be secreted from mast cells without degranulation and without the release of histamine or tryptase.59 We had also reported that mast cells can release the content of individual secretory granules71 or individual mediators without degranulation.52 This process was distinct from “piece-meal degranulation”,72 granule-associated vesicle transport63 or the release of extracellular vesicles.67, 73-78
Moreover, mast cell mediators could have autocrine actions affecting the expression of receptors or the overall reactivity of mast cells. For instance, mast cells can release the “alarmin” IL-33, themselves.55 IL-33 then stimulated mast cells via activation of its own specific surface receptor ST2 and significantly increased the ability of substance P (SP) to secrete VEGF,79, 80 IL-31, 57 TNF43 and IL-1β.55 Mast cell-derived IL-1β or histamine further stimulated release of IL-1β from macrophages.81 IL-1β could, in turn, stimulate mast cells to release IL-6, which was shown to stimulate mast cell proliferation.82 The presence of the D816V-KIT mutation in mast cells was associated with constitutive release of IL-6.83 Serum levels of IL-6 were reported to be elevated in mastocytosis84-86 and correlated with disease severity. Mast cells could also undergo directional mast cell degranulation and secretion of TNF and possibly other pro-inflammatory mediators into the bloodstream.87 It is also important to note that mast cells exhibit different phenotypes including expression of different receptors depending on the tissue microenvironment.88 Moreover, different receptors may interact and increase mast cell reactivity,89 as shown for FcεRI and MRGPRX2, which were reported to have additive effect in stimulating degranulation in human skin mast cells.90
IL-33 increased the expression of the SP receptor neurokinin-1 (NK-1), while SP increased expression of the IL-33 receptor ST2.55 SP also induced expression of the receptor CRHR-1 for the key stress hormone CRH in human mast cells.91 Instead, SP downregulated expression of FcεRI in human mast cells.92 CRH stimulated mast cells to release VEGF without degranulation, an action that was augmented by the peptide neurotensin (NT);93 during this process, CRH stimulated the expression of the NT receptor NT3, while NT stimulated the expression of CRHR-1.94 These findings could help explain why many atopic patients worsen dramatically after a major stressful episode.95, 96.
Mast cell-derived mediators could also induce epigenetic effects as shown for tryptase, which could catalyze histone clipping97 and could regulate modification of histones in mast cell leukemia cells.98 Expression of Ten-eleven translocation-2 (TET2), an epigenetic regulator, was induced in response to activation of mast cells.99, 100 Hence, mast cells are very dynamic cells that respond not only to external but also to innate stimuli. Such findings have prompted the re-evaluation of the secretory processes and their regulation in mast cells.101
Our understanding of the regulation of mediator release via these different modes of secretion and its regulation is still poorly understood. Even though the stimulus-response coupling pathway has been well delineated for activation of the high-affinity surface receptor for IgE (FcεRI),42, 102, 103 and more recently of the low-affinity receptor for cationic peptides, Mas-Related G Protein-Coupled Receptor-X2 (MRGPRX2),104-108 there is still a lack of understanding of the molecular events regulating secretion, whether by degranulation, selective release of mediators or any other mode of secretion (Table 1). The mode and extent of mast cell responsiveness ultimately depend on the interplay between stimulatory and inhibitory signaling pathways, such as CD300109, 110 and Singlets,111 especially Siglec-7,112 and the β subunit of FcεRI (FcεRIβ).113
In spite of the advances briefly outlined above, there is still no effective inhibitor of mediator secretion from mast cells. Antihistamines interfere with histamine binding to its receptors after it has been secreted. There has been considerable progress in developing drugs that block tyrosine kinases involved in mast cell proliferation.114 As a result, our understanding of mast cell stimulation especially by non-IgE triggers remains poor and there are still no clinically effective inhibitors of mast cell activation and inflammatory mediator release.
2. Ezrin, Radixin, Moesin (ERM) Family of Proteins
Ezrin, radixin and moesin (ERMs) are fairly homologous proteins (73% amino acid identity) that link the actin cytoskeleton to the cytoplasmic tail of transmembrane proteins in the plasma membrane thus regulating the formation of F-actin-based structures.115-120 ERMs localize to cell surface protrusions such as microvilli, filopodia and cell–cell junctions ERMs are critical for signal transduction from the cell surface into the cell. Given the high degree of homology and their co-expression to various degrees in many cell types, overlapping or even compensatory functions have been proposed.
Ezrin was named after Ezra Cornell University where it was first isolated from microvilli in chicken intestinal epithelial cells, while radixin (from the Latin meaning root) was isolated from the adherens junctions of rat liver hepatocytes. Moesin (membrane-organizing extension spike protein) was isolated from smooth muscle cells of the bovine uterus. ERMs contain two functional domains connected through a long α-helix region (Fig. 1A): the N-terminal FERM (band 4.1 protein-ERM) domain, which is critical for the function of the ERMs, and the C-terminal ERM association domain (C-ERMAD). The FERM domain is composed of three subdomains (F1, a ubiquitin-like domain; F2, with four α-helices; and F3, a pleckstrin homology domain). The FERM domain and the C-ERMAD can bind each other in a head-to-tail manner, leading to a closed/inactive conformation (Fig. 1B).
Figure 1.
Diagrammatic representation of how differential phosphorylation of moesin could regulate secretion from mast cells. Phosphorylation of moesin at Thr558 in response to triggers opens up binding sites permitting granules to travel to the cell surface and secrete granule-stored mediators via degranulation. In contrast, phosphorylation of moesin at Ser56/Thr66 by cromolyn or flavonoids changes the conformational structure of moesin so that Ser558 is no longer accessible to bind to actin thus preventing secretion.
Figure 1.
Diagrammatic representation of how differential phosphorylation of moesin could regulate secretion from mast cells. Phosphorylation of moesin at Thr558 in response to triggers opens up binding sites permitting granules to travel to the cell surface and secrete granule-stored mediators via degranulation. In contrast, phosphorylation of moesin at Ser56/Thr66 by cromolyn or flavonoids changes the conformational structure of moesin so that Ser558 is no longer accessible to bind to actin thus preventing secretion.
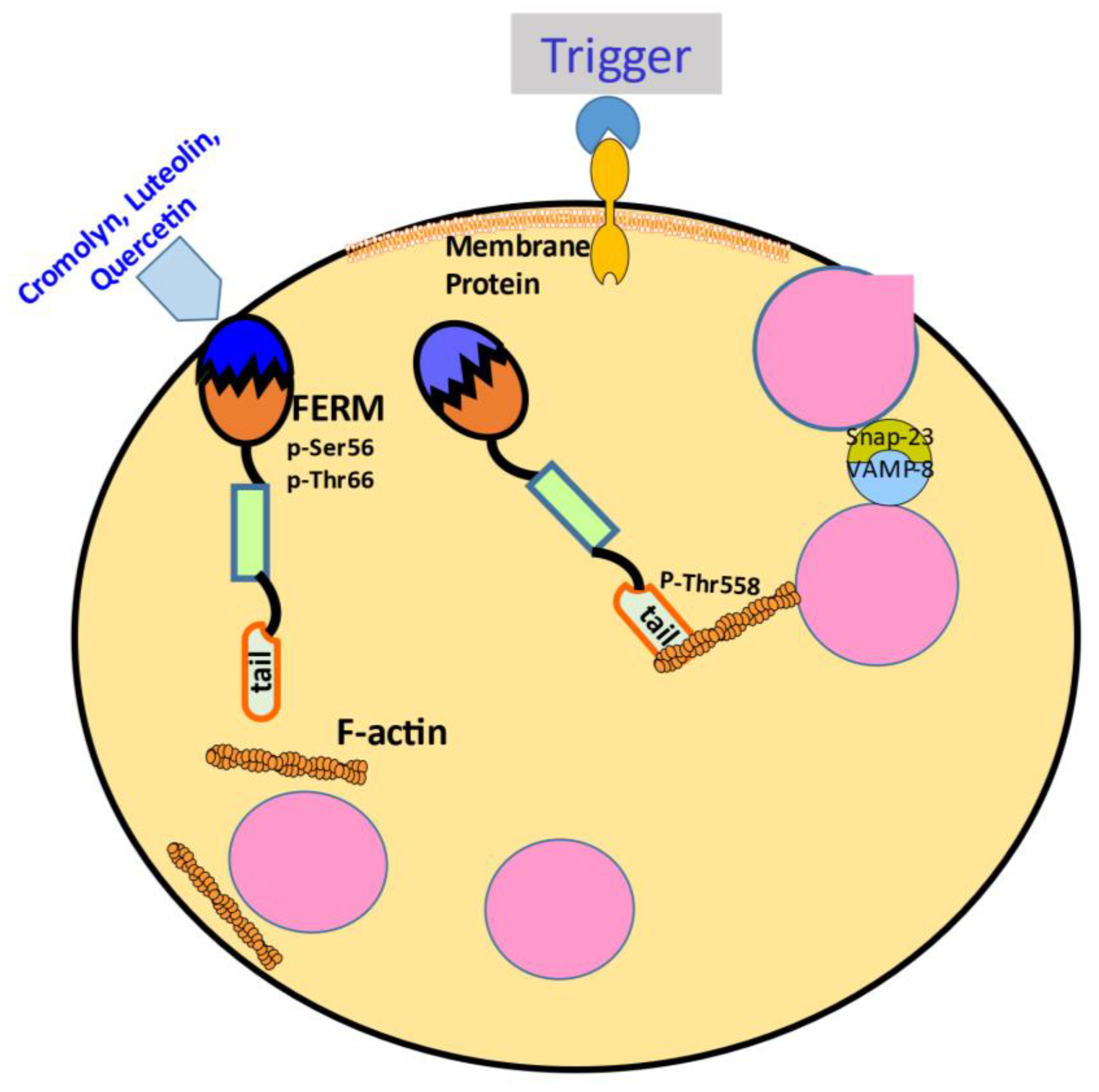
The release of the C-ERMAD from the FERM domain is necessary for the activation of ERMs, unmasking their F-actin- and PM-binding sites. Activation of ERMs occurs first by phosphatidylinositol 4,5-bisphosphate (PIP2) binding to the N-terminus and changing the 3-D structure exposing a C-terminal Threonine (Thr567 in ezrin, Thr564 in radixin and Thr558 in moesin) for phosphorylation116, 121 by the Rho family of GTPases (RhoA/Rac/Cdc42). This step transitions ERMs from a closed (inactive, Fig. B) to an open (active, Fig. 1A) conformation122 that exposes the C-terminal F-actin binding domain that cross-links plasma membrane proteins with actin filaments (Fig. 2).116, 119-122
Figure 2.
Moesin in Mast Cell Secretion.

3. Moesin in mast cells
The expression of particular ERM members varies among different cells. Moesin is mainly expressed in endothelial cells, ezrin in intestinal epithelial cells and radixin in hepatocytes. However, moesin is the most abundant ERM in leukocytes, whereas ezrin is less expressed and radixin is nearly absent;118 we found that the same was also true for mast cells.
Mast cells, like any other secretory cell, require the actin cytoskeleton123 that is necessary for signal-transduction and movement of secretory granules or vesicles destined for secretion to the cell surface. For instance, aggregation of IgE bound to FcεRI by a multivalent antigen stimulates mast cell secretion and rapidly depolymerizes actin filaments, with the actin-severing protein cofilin being dephosphorylated several minutes after stimulation.124In contrast, disaggregation of IgE terminates degranulation mediated by dephosphorylation of Syk associated with a decrease in intracellular Ca2+ concentration and rapid recovery of actin polymerization. Upon FcεRI stimulation, Dok-1(downstream of tyrosine kinase 1), undergoes Tyr phosphorylation, which negatively regulates Ras/Erk signaling and the subsequent secretion.125 Following FcεRI activation, Dok-1 is recruited to the plasma membrane, leading to Tyr phosphorylation. In contrast, phosphorylation of Dok-1 inhibited FcεRI-induced calcium influx and calcium-dependent disassembly of actin filaments, thus negatively regulating degranulation.125 It was previously shown that Rho GTPases regulate exocytosis and possibly secretory granule transport. One paper used live-cell imaging to analyze cytoskeleton assembly and secretory granule transport in real-time of mast cells or rat basophil cells (RBL-1) during antigen stimulation. This paper showed that granule transport to the cell periphery was coordinated by de novo microtubule formation and not F-actin since kinesore, which activates the microtubule motor kinesin-1 inhibited microtubule-granule association and significantly reduced degranulation.126 However, how F-actin or microtubules communicate with secretory granules (or vesicles) and the plasma membrane is still not well understood. Knockdown of the unconventional long-tailed myosin (MYO1F), which localizes with cortical F-actin by short hairpin RNA, reduced human mast cell degranulation stimulated by both IgE and MRGPRX2, and was accompanied by reduced reassembly of the cortical actin ring and fewer secretory granules localized close to the cell surface.127 Interestingly, MYO1F knockdown also resulted in fewer fissioned mitochondria and deficient mitochondria translocation to sites of degranulation by exocytosis.127 We had also shown that mitochondria fission accompanied secretion by degranulation, but not during secretion of de novo synthesized mediators from human mast cells stimulated by SP18 and also in skin biopsies from patients with atopic dermatitis.128 We further showed that stimulation of mast cells resulted in extracellular secretion of mitochondrial DNA (mtDNA) that acted as an “innate pathogen” and triggered an autoinflammatory response. Increased levels of mtDNA have been reported in patients with COVID-19,129-132 psoriasis,133 as well as in EVs from patients with myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS)134 and from children with autism spectrum disorder (ASD), in both cases of which mtDNA activated cultured human microglia to secrete IL-1β.135
We had shown that the ability of the so called “mast cell stabilizer” disodium cromoglycate (cromolyn) to inhibit secretion from rat mast cells in response to the cationic Compound 48/80 (C48/80) closely paralleled the phosphorylation of a 78 kDa protein,136-138 on the N-terminal Ser56, Ser74 and Thr66 residues (Fig. 1B).139 We subsequently cloned this protein from mast cells and showed it to be moesin,140 but we had called it Mast Cell Degranulation Inhibitory Agent=MACEDONIA.141 It is important to note that phosphorylation of at least the N-terminal Ser56/74 and Thr66 residues during inhibition is different than the well-known phosphorylation of C-ERMAD Thr558 associated with moesin activation, inhibition of mast cell secretion of histamine was associated with.139 In support of the involvement of additional phosphorylation sites than Thr558, there is evidence that at least in ezrin, Thr235 is phosphorylated by cyclin-dependent kinase 5 (CDK5) and cooperates with Thr576 for its full activation.142
Using Confocal microscopy and ultra cryo-immuno-electron microscopy to preserve the antigenicity of ERMs, we had shown that mast cells contain almost exclusively moesin, (with a small amount of ezrin), which was critically localized primarily at the plasma membrane and filopodia, with less around secretory granules; we further showed that cromolyn induced clustering of moesin around secretory granules.140 We hypothesized that conformational changes of moesin, regulated by phosphorylation/dephosphorylation, could possibly regulate mast cell secretion via positional rearrangements with respect to the membrane/cytoskeleton.140 We further hypothesized that moesin could, in fact, serve a dual function depending on its phosphorylation pattern that occurs after a trigger or an inhibitor interacts with the cell surface.143 In other words, moesin phosphorylation at C-terminal Thr558 would switch moesin to its active form (Fig. 1A) and permit mast secretory granules to move to the surface, fuse with the plasma membrane and undergo exocytosis (Fig. 2). In contrast, phosphorylation of N-terminal Ser/Thr sites would switch moesin to its inactive state (Fig. 1B) resulting in either: (a) prevention of phosphorylation of Thr558 and moesin activation, (b) interaction with secretory granules preventing them from moving to the cell surface or (c) affecting the structure of the cell cortex and block secretion indirectly (Fig. 2). However, it remains unknown how phosphorylation of moesin at different sites affects secretion from mast cells in response to different triggers, and how phosphorylation at the N-terminal sites mechanistically leads to inhibition of mast cell secretion. Moreover, it is not presently known if phosphorylation of moesin may affect modes of secretion other than degranulation by exocytosis. One paper identified a number of ser/thr phosphorylated proteins in activated mast cells, including moesin, but these were involved in different processes such as metabolism and cell structure.144 Even though ezrin has been mostly discussed for its involvement in cancer,145 it is not known if ezrin could compensate for moesin should the latter be absent or “incapacitated” in mast cells. In fact, ezrin, has been implicated in asthma.146 Phosphorylation of ezrin at Thr567 was associated with trophoblast motility.147
Interestingly, moesin knock-out mice were shown to have lymphopenia,148 but mast cell numbers were apparently intact; however, the authors did not investigate mast cell secretion.148 One X-linked moesin-associated immunodeficiency (X-MAID) has been identified and is characterized by a primary immunodeficiency associated with severe lymphopenia leading to recurrent infections. X-MAID is caused by a single point mutation leading to a R171W amino acid change in moesin (moesinR171W).149 In fact, a mouse model with global expression of moesinR171W exhibited lymphopenia, but was still characterized by systemic inflammation.149
Phosphorylation of moesin has also been studied in other secretory systems. Moesin was shown to be phosphorylated at Thr558 within seconds of thrombin-induced activation of platelets.150, 151 Instead, tyrosine phosphorylation of moesin was reported during the activation of platelets by arachidonic acid.152 These phosphorylation patterns are reversed by protein phosphatase 2C, which inactivates the F-actin binding site of activated platelets.153 Phosphorylation at Thr558 was also reported in activated RAW264.7 macrophages.154 ERM proteins have been shown to be involved in T-cell polarization and immune synapse formation.155 It is interesting that anti-moesin autoantibodies were isolated from patients with aplastic anemia156 and autoimmune vasculitis.157 However, their significance of these autoantibodies is not apparent nor is their potential presence in patients with allergies and inflammatory disorders.
4. SNAREs and SNAPs
One possible mechanism of how moesin may regulate mast cell secretion could involve the Soluble NSF attachment protein receptors (SNAREs) and synaptosomal associated proteins (SNAPs) discovered by Dr. J.E. Rothman, who was awarded the 2013 Nobel in Physiology and Medicine for delineating the principles for membrane fusion during secretory membrane fusion.158 The existence of distinct secretory vesicle calcium-sensitive proteins “snapping” with corresponding proteins on the plasma membrane during secretion of mast cells had actually been proposed much earlier by one of the authors (TCT) in his doctoral thesis examination at Yale University in 1974 with examiner being Dr. G. Palade who had just received the 1974 Nobel in Physiology and Medicine for his discovery that secreted proteins are carried from the endoplasmic reticulum (ER) to the cell surface in specialized compartments or transport vesicles.
SNAREs159-161 and synaptosomal associated protein of 23 kDa) (SNAP-23)162-167 have been shown to be involved in mast cell secretion (Fig. 2). In fact, there may be different mechanisms regulating exocytosis in mast cells168 and mast cell distinct secretory granule subsets may be regulated by different SNARE isoforms169 and different vesicle-associated membrane proteins (VAMPs), especially VAMP2- and VAMP8.170, 171
Mast cells express Munc18-2, which interacts with SNARE syntaxin 2 or 3, as well as Munc18-3, which interacts with syntaxin 4. Munc18-2 was localised to secretory granules, whereas Munc18-3 was found on the plasma membrane. Increased expression of Munc18-2 inhibited IgE-triggered exocytosis, while increased expression of Munc18-3 had no effect. Upon stimulation, Munc18-2 redistributed persisted on granules that were aligned along microtubules, but was excluded from F-actin ruffles, suggesting a role for Munc18-2 and the microtubule network in the regulation of secretion by degranulation in mast cells.172 In addition, a number of so-called 'adapters' have been reported to regulate secretion from mast cells by binding multiple signaling proteins and localizing them to specific cellular compartments. 40
It is of note that degranulation of different mast cell vesicle subsets was differentially and selectively regulated by various polyphenols via interfering with two SNARE complexes, Syn (syntaxin) 4/SNAP-23/VAMP2 and Syn4/SNAP23/VAMP8.173 Similarly, polyphenols were shown to interfere with “zippering” of SNARES in the neuron.174 The structure of the phenolic flavonol quercetin is somewhat similar to cromolyn,143 but is a more potent inhibitor than cromolyn.175 Quercetin inhibited rat mast cell degranulation,176, 177 possibly via inhibition of protein kinase C (PKC),138, 178 but it also induced phosphorylation of moesin.178 Quercetin also inhibited the release of pro-inflammatory cytokines,138 including IL-6,177 from cultured human mast cells. The quercetin-related flavone luteolin and the luteolin analogue tetramethoxyluteolin were even more potent inhibitors of both of degranulation,179 as well as of the release of TNF43 and IL-1β55 from human mast cells.
The ability of flavonoids to inhibit mast cell secretion via phosphorylation of moesin led to conjectures about design of more potent inhibitors.143
5. Neuroinflammation
Mast cells communicate with microglia180, 181 and can activate them181-184 via the release of mediators such as histamine185 and tryptase,186 leading to neuroinflammation.180, 182 Activation of mast cells and microglia in the brain187 could affect neurodevelopment,188 resulting in neuronal apoptosis,189 and lead to cognitive dysfunction.189 In fact activation of mast cells and microglia has been linked to the pathogenesis of autism spectrum disorder (ASD),190-194 neurodegenerative diseases35, 195 and traumatic brain injury (TBI).24, 196 It is, therefore, of interest that moesin has been reported to be involved in the activation of microglia.197 Moreover, the moesin pseudogene 1 antisense (MSNP1AS) was shown to decrease the number and length of neurites, reduce neural viability and promote apoptosis via inhibition of moesin protein expression, while moesin improved social interactions and reduced repetitive behaviors in BTBR mice.198
Moreover, one paper reported that ezrin, radixin and moesin had distinct roles of in maintaining the plasma membrane integrity and functions of the blood-brain barrier (BBB) transporters,199 which is important because mast cells can regulate the permeability of the BBB,200 disruption of which has been implicated in ASD,201 in Alzheimer’s disease33 and in neuro-COVID.202 In this context, it is relevant that flavonoids could have anti-inflammatory34, 203-209 and neuroprotective effects,210 as well as reduce cognitive dysfunction,211-215 especially brain fog.216-218 In particular, luteolin inhibited both microglia219-221 and mast cells.222, 223
One formulation containing liposomal luteolin in olive pomace (fruit) oil (NeuroProtek®) resulted in significant improvement of children with ASD224 with a concomitant decrease in serum inflammatory markers.225 Other papers reported the beneficial action of luteolin in Long-COVID-associated brain fog.216, 226 and neurotrauma.227
6. Conclusions
The studies reviewed indicate that moesin phosphorylation and localization may be important in the regulation of mast cell-derived secretion of at least secretory granule-associated mediators such as histamine, TNF and tryptase.
It would be important to investigate the expression of total and phosphorylated moesin in human mast cells of different degrees of reactivity/types, such as the leukemic human mast cell line-1 (HMC-1), the Laboratory of allergic diseases-2 (LAD2) and LADR mast cells,228 as well as primary human umbilical cord blood-derived cultured mast cells (hCBMCs), mast cells developed from pluripotent stem cells,229-231 but also mast cells from a cutaneous mastocytosis or urticaria lesions. Other future studies should investigate whether the knockdown of moesin using small interfering ribonucleic acid (siRNA) would affect the extent of secretion or interfere with the ability of flavonoids to inhibit mast cell secretion. Additionally, studies should also investigate which specific sites are phosphorylated in response to triggers or inhibitors of either degranulation or differential release of select mediators using trypsin-digested moesin peptides analyzed by mass spectrometry and validated with site-specific phospho-antibodies and point mutant analysis.
It would be important to investigate the possible presence of some innate molecule or identify novel molecules that keep moesin in its inactive state, that could be targeted for the development of new effective anti-allergic and anti-inflammatory drugs.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
Authors declare no conflict of interest.
References
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed]
- Parwaresch, M.; Homy, H.-P.; Lennert, K. Tissue mast cells in health and disease. Pathol. - Res. Pr. 1985, 179, 439–461. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G. Mast cell, the peculiar member of the immune system: A homeostatic aspect. Acta Microbiol. et Immunol. Hung. 2015, 62, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Siebenhaar, F.; Redegeld, F.A.; Bischoff, S.C.; Gibbs, B.F.; Maurer, M. Mast Cells as Drivers of Disease and Therapeutic Targets. Trends Immunol. 2017, 39, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.; Looareesuwan, S.; White, N.; Silamut, K.; Kietinun, S.; Warrell, D. Quinine pharmacokinetics and toxicity in pregnant and lactating women with falciparum malaria. Br. J. Clin. Pharmacol. 1986, 21, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Falduto, G.H.; Pfeiffer, A.; Luker, A.; Metcalfe, D.D.; Olivera, A. Emerging mechanisms contributing to mast cell-mediated pathophysiology with therapeutic implications. Pharmacol. Ther. 2020, 220, 107718. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, J.S.; Maurer, M.; Metcalfe, D.D.; Pejler, G.; Sagi-Eisenberg, R.; Nilsson, G. The ingenious mast cell: Contemporary insights into mast cell behavior and function. Allergy 2021, 77, 83–99. [Google Scholar] [CrossRef]
- Kolkhir, P.; Elieh-Ali-Komi, D.; Metz, M.; Siebenhaar, F.; Maurer, M. Understanding human mast cells: lesson from therapies for allergic and non-allergic diseases. Nat. Rev. Immunol. 2021, 22, 294–308. [Google Scholar] [CrossRef]
- Levi-Schaffer, F.; Gibbs, B.F.; Hallgren, J.; Pucillo, C.; Redegeld, F.; Siebenhaar, F.; Vitte, J.; Mezouar, S.; Michel, M.; Puzzovio, P.G.; et al. Selected recent advances in understanding the role of human mast cells in health and disease. J. Allergy Clin. Immunol. 2022, 149, 1833–1844. [Google Scholar] [CrossRef]
- Olivera, A.; Beaven, M.A.; Metcalfe, D.D. Mast cells signal their importance in health and disease. J. Allergy Clin. Immunol. 2018, 142, 381–393. [Google Scholar] [CrossRef]
- Sibilano, R.; Frossi, B.; Pucillo, C.E. Mast cell activation: A complex interplay of positive and negative signaling pathways. Eur. J. Immunol. 2014, 44, 2558–2566. [Google Scholar] [CrossRef]
- E Gallenga, C.; Pandolfi, F.; Caraffa, A.; Kritas, S.K.; Ronconi, G.; Toniato, E.; Martinotti, S.; Conti, P. Interleukin-1 family cytokines and mast cells: activation and inhibition. J. Biol. Regul. Homeost. agents 2019, 33, 1–6. [Google Scholar]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Toniato, E.; Frydas, I.; Robuffo, I.; Ronconi, G.; Caraffa, A.; Kritas, S.K.; Conti, P. Activation and inhibition of adaptive immune response mediated by mast cells. . 2017, 31, 543–548. [Google Scholar] [PubMed]
- Avila, M.; Gonzalez-Espinosa, C. Signaling through Toll-like receptor 4 and mast cell-dependent innate immunity responses. IUBMB Life 2011, 63, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P. Microbes taming mast cells: Implications for allergic inflammation and beyond. Eur. J. Pharmacol. 2016, 778, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Carroll-Portillo, A.; Cannon, J.L.; Riet, J.T.; Holmes, A.; Kawakami, Y.; Kawakami, T.; Cambi, A.; Lidke, D.S. Mast cells and dendritic cells form synapses that facilitate antigen transfer for T cell activation. J. Cell Biol. 2015, 210, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Weng, Z.; Sismanopoulos, N.; Asadi, S.; Therianou, A.; Alysandratos, K.-D.; Angelidou, A.; Shirihai, O.; Theoharides, T.C. Mitochondria Distinguish Granule-Stored from de novo Synthesized Tumor Necrosis Factor Secretion in Human Mast Cells. Int. Arch. Allergy Immunol. 2012, 159, 23–32. [Google Scholar] [CrossRef]
- Ishii, T.; Wang, J.; Zhang, W.; Mascarenhas, J.; Hoffman, R.; Dai, Y.; Wisch, N.; Xu, M. Pivotal role of mast cells in pruritogenesis in patients with myeloproliferative disorders. Blood 2009, 113, 5942–5950. [Google Scholar] [CrossRef] [PubMed]
- Mekori, Y.A.; Hershko, A.Y.; Frossi, B.; Mion, F.; Pucillo, C.E. Integrating innate and adaptive immune cells: Mast cells as crossroads between regulatory and effector B and T cells. Eur. J. Pharmacol. 2016, 778, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Christy, A.L.; Brown, M.A. The Multitasking Mast Cell: Positive and Negative Roles in the Progression of Autoimmunity. J. Immunol. 2007, 179, 2673–2679. [Google Scholar] [CrossRef] [PubMed]
- Hakim-Rad K, Metz M, Maurer M. Mast cells: makers and breakers of allergic inflammation. Curr Opin Allergy Clin Immunol 2009, 9, 427–430.
- Theoharides TC, Alysandratos KD, Angelidou A, Delivanis DA, Sismanopoulos N, Zhang B, Asadi S, Vasiadi M, Weng Z, Miniati A, Kalogeromitros D. Mast cells and inflammation. Biochim Biophys Acta 2012, 1822, 21–33.
- Kempuraj D, Ahmed ME, Selvakumar GP, Thangavel R, Dhaliwal AS, Dubova I, Mentor S, Premkumar K, Saeed D, Zahoor H, Raikwar SP, Zaheer S, Iyer SS, Zaheer A. Brain Injury-Mediated Neuroinflammatory Response and Alzheimer's Disease. Neuroscientist 2020, 26, 134–155.
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Theoharides TC, Valent P, Akin C. Mast Cells, Mastocytosis, and Related Disorders. N Engl J Med 2015, 373, 163–172.
- Theoharides, T.C. Atopic Conditions in Search of Pathogenesis and Therapy. Clin. Ther. 2013, 35, 544–547. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Leeman, S.E. Effect of IL-33 on de novo synthesized mediators from human mast cells. J. Allergy Clin. Immunol. 2019, 143, 451. [Google Scholar] [CrossRef]
- Akin, C. Mast cell activation disorders. J Allergy Clin Immunol Pract 2014, 2, 252–257 e251; quiz 258. [Google Scholar] [CrossRef]
- Theoharides TC, Tsilioni I, Ren H. Recent advances in our understanding of mast cell activation - or should it be mast cell mediator disorders? Expert Rev Clin Immunol 2019, 15, 639–656.
- Galli, S.J.; Gaudenzio, N.; Tsai, M. Mast Cells in Inflammation and Disease: Recent Progress and Ongoing Concerns. Annu. Rev. Immunol. 2020, 38, 49–77. [Google Scholar] [CrossRef]
- Kempuraj D, Selvakumar GP, Ahmed ME, Raikwar SP, Thangavel R, Khan A, Zaheer SA, Iyer SS, Burton C, James D, Zaheer A. COVID-19, Mast Cells, Cytokine Storm, Psychological Stress, and Neuroinflammation. Neuroscientist 2020, 1073858420941476.
- Kempuraj D, Mentor S, Thangavel R, Ahmed ME, Selvakumar GP, Raikwar SP, Dubova I, Zaheer S, Iyer SS, Zaheer A. Mast Cells in Stress, Pain, Blood-Brain Barrier, Neuroinflammation and Alzheimer's Disease. Front Cell Neurosci 2019, 13, 54. [CrossRef]
- Theoharides, T.C.B.; Conti, P.D.; Economu, M.M. Brain Inflammation, Neuropsychiatric Disorders, and Immunoendocrine Effects of Luteolin. J. Clin. Psychopharmacol. 2014, 34, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj D, Selvakumar GP, Thangavel R, Ahmed ME, Zaheer S, Raikwar SP, Iyer SS, Bhagavan SM, Beladakere-Ramaswamy S, Zaheer A. Mast Cell Activation in Brain Injury, Stress, and Post-traumatic Stress Disorder and Alzheimer's Disease Pathogenesis. Front Neurosci 2017, 11, 703. [CrossRef]
- Kempuraj, D.; Thangavel, R.; Selvakumar, G.P.; Zaheer, S.; Ahmed, M.E.; Raikwar, S.P.; Zahoor, H.; Saeed, D.; Natteru, P.A.; Iyer, S.; et al. Brain and Peripheral Atypical Inflammatory Mediators Potentiate Neuroinflammation and Neurodegeneration. Front. Cell. Neurosci. 2017, 11, 216. [Google Scholar] [CrossRef]
- Galli, S.J.; Grimbaldeston, M.; Tsai, M. Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nat. Rev. Immunol. 2008, 8, 478–486. [Google Scholar] [CrossRef]
- Cheng, L.E.; Hartmann, K.; Roers, A.; Krummel, M.F.; Locksley, R.M. Perivascular Mast Cells Dynamically Probe Cutaneous Blood Vessels to Capture Immunoglobulin E. Immunity 2013, 38, 166–175. [Google Scholar] [CrossRef]
- Metzger, H.; Eglite, S.; Haleem-Smith, H.; Reischl, I.; Torigoe, C. Quantitative aspects of signal transduction by the receptor with high affinity for IgE. Mol. Immunol. 2002, 38, 1207–1211. [Google Scholar] [CrossRef]
- Alvarez-Errico, D.; Lessmann, E.; Rivera, J. Adapters in the organization of mast cell signaling. Immunol. Rev. 2009, 232, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Ando T, Kitaura J. Tuning IgE. IgE-Associating Molecules and Their Effects on IgE-Dependent Mast Cell Reactions. Cells 2021, 10. [CrossRef]
- Nagata Y, Suzuki R. FcepsilonRI. A Master Regulator of Mast Cell Functions. Cells 2022, 11. [CrossRef]
- Taracanova, A.; Alevizos, M.; Karagkouni, A.; Weng, Z.; Norwitz, E.; Conti, P.; Leeman, S.E.; Theoharides, T.C. SP and IL-33 together markedly enhance TNF synthesis and secretion from human mast cells mediated by the interaction of their receptors. Proc. Natl. Acad. Sci. 2017, 114, E4002–E4009. [Google Scholar] [CrossRef]
- Theoharis, C.T.; Theoharides, T.C.; Konstantinidou, A.D. Corticotropin-releasing hormone and the blood-brain-barrier. Front. Biosci. 2007, 12, 1615–28. [Google Scholar] [CrossRef] [PubMed]
- Kandere-Grzybowska, K.; Letourneau, R.; Kempuraj, D.; Donelan, J.; Poplawski, S.; Boucher, W.; Athanassiou, A.; Theoharides, T.C. IL-1 Induces Vesicular Secretion of IL-6 without Degranulation from Human Mast Cells. J. Immunol. 2003, 171, 4830–4836. [Google Scholar] [CrossRef]
- Schwartz, L.B. Mediators of human mast cells and human mast cell subsets. Ann. Allergy 1987, 58, 226–35. [Google Scholar]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Uvnäs, B. Histamine storage and release. Fed. Proc. 1974, 33. [Google Scholar] [CrossRef]
- Awan, S.F.; Schwartz, L.B.; Maric, I.; Metcalfe, D.D.; Carter, M.C. Acute increases in total serum tryptase unassociated with hemodynamic instability in diffuse cutaneous mastocytosis. Ann. Allergy, Asthma Immunol. 2022, 129, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, T.G.; Arkesteijn, G.J.A.; van de Lest, C.H.A.; Geerts, W.J.C.; Goerdayal, S.S.; Altelaar, M.A.F.; Redegeld, F.A.; Hoen, E.N.M.N.; Wauben, M.H.M. Mast Cell Degranulation Is Accompanied by the Release of a Selective Subset of Extracellular Vesicles That Contain Mast Cell–Specific Proteases. J. Immunol. 2016, 197, 3382–3392. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; Giavina-Bianchi, P.; Mezzano, V.; Castells, M. Expanding Spectrum of Mast Cell Activation Disorders: Monoclonal and Idiopathic Mast Cell Activation Syndromes. Clin. Ther. 2013, 35, 548–562. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Bondy, P.K.; Tsakalos, N.D.; Askenase, P.W. Differential release of serotonin and histamine from mast cells. Nature 1982, 297, 229–231. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Cochrane, D.E. Critical role of mast cells in inflammatory diseases and the effect of acute stress. J. Neuroimmunol. 2004, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Solimando, A.G.; Desantis, V.; Ribatti, D. Mast Cells and Interleukins. Int. J. Mol. Sci. 2022, 23, 14004. [Google Scholar] [CrossRef] [PubMed]
- Taracanova A, Tsilioni I, Conti P, Norwitz ER, Leeman SE, Theoharides TC. Substance P and IL-33 administered together stimulate a marked secretion of IL-1beta from human mast cells, inhibited by methoxyluteolin. Proc Natl Acad Sci U S A 2018, 115, E9381–E9390.
- Gagari, E.; Tsai, M.; Lantz, C.S.; Fox, L.G.; Galli, S.J. Differential Release of Mast Cell Interleukin-6 Via c-kit. Blood 1997, 89, 2654–2663. [Google Scholar] [CrossRef]
- Petra, A.I.; Tsilioni, I.; Taracanova, A.; Katsarou-Katsari, A.; Theoharides, TC. Interleukin, 3. 3.; interleukin 4 regulate interleukin 31 gene expression secretion from human laboratory of allergic diseases 2 mast cells stimulated by substance, P.; /or immunoglobulin, E. Allergy Asthma Proc 2018, 39, 153–160. [Google Scholar]
- Theoharides, T.C.; Kalogeromitros, D. The Critical Role of Mast Cells in Allergy and Inflammation. Ann. New York Acad. Sci. 2006, 1088, 78–99. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kempuraj, D.; Tagen, M.; Conti, P.; Kalogeromitros, D. Differential release of mast cell mediators and the pathogenesis of inflammation. Immunol. Rev. 2007, 217, 65–78. [Google Scholar] [CrossRef]
- Xu, H.; Bin, N.-R.; Sugita, S. Diverse exocytic pathways for mast cell mediators. Biochem. Soc. Trans. 2018, 46, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef]
- Gaudenzio, N.; Sibilano, R.; Marichal, T.; Starkl, P.; Reber, L.L.; Cenac, N.; McNeil, B.D.; Dong, X.; Hernandez, J.D.; Sagi-Eisenberg, R.; et al. Different activation signals induce distinct mast cell degranulation strategies. J. Clin. Investig. 2016, 126, 3981–3998. [Google Scholar] [CrossRef]
- Crivellato, E.; Nico, B.; Gallo, V.P.; Ribatti, D. Cell Secretion Mediated by Granule-Associated Vesicle Transport: A Glimpse at Evolution. Anat. Rec. 2010, 293, 1115–1124. [Google Scholar] [CrossRef]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast Cell Mediators: Their Differential Release and the Secretory Pathways Involved. Front. Immunol. 2014, 5, 569–569. [Google Scholar] [CrossRef]
- Vukman, K.V.; Försönits, A.; Oszvald. ; Tóth, E.Á.; Buzás, E.I. Mast cell secretome: Soluble and vesicular components. Semin. Cell Dev. Biol. 2017, 67, 65–73. [Google Scholar] [CrossRef]
- Weng Z, Zhang B, Tsilioni I, Theoharides TC. Nanotube Formation: A Rapid Form of "Alarm Signaling"? Clin Ther 2016, 38, 1066–1072. [CrossRef]
- Carroll-Portillo, A.; Surviladze, Z.; Cambi, A.; Lidke, D.S.; Wilson, B.S. Mast cell synapses and exosomes: membrane contacts for information exchange. Front. Immunol. 2012, 3, 46. [Google Scholar] [CrossRef]
- Joulia, R.; Gaudenzio, N.; Rodrigues, M.; Lopez, J.; Blanchard, N.; Valitutti, S.; Espinosa, E. Mast cells form antibody-dependent degranulatory synapse for dedicated secretion and defence. Nat. Commun. 2015, 6, 6174. [Google Scholar] [CrossRef]
- Cochrane, D.E.; Douglas, W.W. Calcium-Induced Extrusion of Secretory Granules (Exocytosis) in Mast Cells Exposed to 48/80 or the Ionophores A-23187 and X-537A. Proc. Natl. Acad. Sci. 1974, 71, 408–412. [Google Scholar] [CrossRef]
- Asadi, S.; Theoharides, T.C. Corticotropin-releasing hormone and extracellular mitochondria augment IgE-stimulated human mast-cell vascular endothelial growth factor release, which is inhibited by luteolin. J. Neuroinflammation 2012, 9, 85–85. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Douglas, W.W. Secretion in Mast Cells Induced by Calcium Entrapped Within Phospholipid Vesicles. Science 1978, 201, 1143–1145. [Google Scholar] [CrossRef]
- Dvorak, A.M. Piecemeal Degranulation of Basophils and Mast Cells Is Effected by Vesicular Transport of Stored Secretory Granule Contents. 2005, 85, 135–184. [CrossRef]
- Skokos, D.; Le Panse, S.; Villa, I.; Rousselle, J.-C.; Peronet, R.; David, B.; Namane, A.; Mécheri, S. Mast Cell-Dependent B and T Lymphocyte Activation Is Mediated by the Secretion of Immunologically Active Exosomes. J. Immunol. 2001, 166, 868–876. [Google Scholar] [CrossRef]
- Skokos, D.; Goubran-Botros, H.; Roa, M.; Mécheri, S. Erratum to “Immunoregulatory properties of mast cell-derived exosomes” [Molecular Immunology 38 (16–18) (2002) 1359–1362]. Mol. Immunol. 2003, 39, 835. [Google Scholar] [CrossRef]
- Shefler, I.; Salamon, P.; Hershko, A.Y.; Mekori, Y.A. Mast Cells as Sources and Targets of Membrane Vesicles. Curr. Pharm. Des. 2011, 17, 3797–3804. [Google Scholar] [CrossRef] [PubMed]
- Lecce M, Molfetta R, Milito ND, Santoni A, Paolini R. FcepsilonRI Signaling in the Modulation of Allergic Response: Role of Mast Cell-Derived Exosomes. Int J Mol Sci 2020, 21. [CrossRef]
- Shefler, I.; Salamon, P.; Mekori, Y.A. Extracellular Vesicles as Emerging Players in Intercellular Communication: Relevance in Mast Cell-Mediated Pathophysiology. Int. J. Mol. Sci. 2021, 22, 9176. [Google Scholar] [CrossRef] [PubMed]
- Phukan, P.; Barman, B.; Chengappa, N.K.; Lynser, D.; Paul, S.; Nune, A.; Sarma, K. Diffusion tensor imaging analysis of rheumatoid arthritis patients with neuropsychiatric features to determine the alteration of white matter integrity due to vascular events. Clin. Rheumatol. 2022, 41, 3169–3177. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Zhang, B.; Kempuraj, D.; Tagen, M.; Vasiadi, M.; Angelidou, A.; Alysandratos, K.-D.; Kalogeromitros, D.; Asadi, S.; Stavrianeas, N.; et al. IL-33 augments substance P-induced VEGF secretion from human mast cells and is increased in psoriatic skin. Proc. Natl. Acad. Sci. USA 2010, 107, 4448–4453. [Google Scholar] [CrossRef]
- Cristinziano, L.; Poto, R.; Criscuolo, G.; Ferrara, A.L.; Galdiero, M.R.; Modestino, L.; Loffredo, S.; de Paulis, A.; Marone, G.; Spadaro, G.; et al. IL-33 and Superantigenic Activation of Human Lung Mast Cells Induce the Release of Angiogenic and Lymphangiogenic Factors. Cells 2021, 10, 145. [Google Scholar] [CrossRef]
- Conti P, Caraffa A, Tete G, Gallenga CE, Ross R, Kritas SK, Frydas I, Younes A, Di Emidio P, Ronconi G. Mast cells activated by SARS-CoV-2 release histamine which increases IL-1 levels causing cytokine storm and inflammatory reaction in COVID-19. J Biol Regul Homeost Agents 2020, 34, 1629–1632.
- Kaur, D.; Gomez, E.; Doe, C.; Berair, R.; Woodman, L.; Saunders, R.; Hollins, F.; Rose, F.; Amrani, Y.; May, R.; et al. IL-33 drives airway hyper-responsiveness through IL-13-mediated mast cell: airway smooth muscle crosstalk. Allergy 2015, 70, 556–567. [Google Scholar] [CrossRef]
- Tobío, A.; Bandara, G.; Morris, D.A.; Kim, D.; Connell, M.P.O.; Komarow, H.D.; Carter, M.C.; Smrz, D. Oncogenic D816V-KIT signaling in mast cells causes persistent IL-6 production. Haematologica 2020, 105, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Boucher, W.; Spear, K. Serum Interleukin-6 Reflects Disease Severity and Osteoporosis in Mastocytosis Patients. Int. Arch. Allergy Immunol. 2002, 128, 344–350. [Google Scholar] [CrossRef]
- Brockow, K.; Akin, C.; Huber, M.; Metcalfe, D.D. IL-6 levels predict disease variant and extent of organ involvement in patients with mastocytosis. Clin. Immunol. 2005, 115, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Mayado, A.; Teodosio, C.; Garcia-Montero, A.C.; Matito, A.; Rodriguez-Caballero, A.; Morgado, J.M.; Muñiz, C.; Jara-Acevedo, M.; Álvarez-Twose, I.; Sanchez-Muñoz, L.; et al. Increased IL6 plasma levels in indolent systemic mastocytosis patients are associated with high risk of disease progression. Leukemia 2015, 30, 124–130. [Google Scholar] [CrossRef]
- Dudeck, J.; Kotrba, J.; Immler, R.; Hoffmann, A.; Voss, M.; Alexaki, V.I.; Morton, L.; Jahn, S.R.; Katsoulis-Dimitriou, K.; Winzer, S.; et al. Directional mast cell degranulation of tumor necrosis factor into blood vessels primes neutrophil extravasation. Immunity 2021, 54, 468–483. [Google Scholar] [CrossRef]
- Lyons DO, Pullen NA. Beyond IgE. Alternative Mast Cell Activation Across Different Disease States. Int J Mol Sci 2020, 21. [CrossRef]
- Franke K, Wang Z, Zuberbier T, Babina M. Cytokines Stimulated by IL-33 in Human Skin Mast Cells: Involvement of NF-kappaB and p38 at Distinct Levels and Potent Co-Operation with FcepsilonRI and MRGPRX2. Int J Mol Sci 2021, 22. [CrossRef]
- Babina M, Wang Z, Li Z, Franke K, Guhl S, Artuc M, Zuberbier T. FcepsilonRI- and MRGPRX2-evoked acute degranulation responses are fully additive in human skin mast cells. Allergy 2022, 77, 1906–1909.
- Asadi, S.; Alysandratos, K.-D.; Angelidou, A.; Miniati, A.; Sismanopoulos, N.; Vasiadi, M.; Zhang, B.; Kalogeromitros, D.; Theoharides, T.C. Substance P (SP) Induces Expression of Functional Corticotropin-Releasing Hormone Receptor-1 (CRHR-1) in Human Mast Cells. J. Investig. Dermatol. 2012, 132, 324–329. [Google Scholar] [CrossRef] [PubMed]
- McCary C, Tancowny BP, Catalli A, Grammer LC, Harris KE, Schleimer RP, Kulka M. Substance P downregulates expression of the high affinity IgE receptor (FcepsilonRI) by human mast cells. J Neuroimmunol 2010, 220, 17–24.
- Donelan, J.; Boucher, W.; Papadopoulou, N.; Lytinas, M.; Papaliodis, D.; Dobner, P.; Theoharides, T.C. Corticotropin-releasing hormone induces skin vascular permeability through a neurotensin-dependent process. Proc. Natl. Acad. Sci. 2006, 103, 7759–7764. [Google Scholar] [CrossRef]
- Alysandratos, K.; Asadi, S.; Angelidou, A.; Zhang, B.; Sismanopoulos, N.; Yang, H.; Critchfield, A.; Theoharides, T.C. Neurotensin and CRH Interactions Augment Human Mast Cell Activation. PLOS ONE 2012, 7, e48934. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Effect of Stress on Neuroimmune Processes. Clin. Ther. 2020, 42, 1007–1014. [Google Scholar] [CrossRef]
- Theoharides, T.C. The impact of psychological stress on mast cells. Ann. Allergy, Asthma Immunol. 2020, 125, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.R.; Wallerman, O.; Paivandy, A.; Calounova, G.; Gustafson, A.-M.; Sabari, B.R.; Zabucchi, G.; Allis, C.D.; Pejler, G. Tryptase-catalyzed core histone truncation: A novel epigenetic regulatory mechanism in mast cells. J. Allergy Clin. Immunol. 2017, 140, 474–485. [Google Scholar] [CrossRef]
- Alanazi, S.; Melo, F.R.; Pejler, G. Tryptase Regulates the Epigenetic Modification of Core Histones in Mast Cell Leukemia Cells. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Monticelli, S.; Leoni, C. Epigenetic and transcriptional control of mast cell responses. F1000Research 2017, 6, 2064. [Google Scholar] [CrossRef]
- Rigo, R.; Chelbi, R.; Agopian, J.; Letard, S.; Griffon, A.; Ghamlouch, H.; Vernerey, J.; Ladopoulos, V.; Voisset, E.; De Sepulveda, P.; et al. TET2 regulates immune tolerance in chronically activated mast cells. J. Clin. Investig. 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Perlman, A.I.; Twahir, A.; Kempuraj, D. Mast cell activation: beyond histamine and tryptase. Expert Rev. Clin. Immunol. 2023, 19, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Blank, U.; Huang, H.; Kawakami, T. The high affinity IgE receptor: a signaling update. Curr. Opin. Immunol. 2021, 72, 51–58. [Google Scholar] [CrossRef]
- Li Y, Leung PSC, Gershwin ME, Song J. New Mechanistic Advances in FcepsilonRI-Mast Cell-Mediated Allergic Signaling. Clin Rev Allergy Immunol 2022, 63, 431–446.
- Babina, M.; Wang, Z.; Artuc, M.; Guhl, S.; Zuberbier, T. MRGPRX2 is negatively targeted by SCF and IL-4 to diminish pseudo-allergic stimulation of skin mast cells in culture. Exp. Dermatol. 2018, 27, 1298–1303. [Google Scholar] [CrossRef]
- Wang Z, Babina M. MRGPRX2 signals its importance in cutaneous mast cell biology: Does MRGPRX2 connect mast cells and atopic dermatitis? Exp Dermatol 2020, 29, 1104–1111.
- Ogasawara, H.; Noguchi, M. Therapeutic Potential of MRGPRX2 Inhibitors on Mast Cells. Cells 2021, 10, 2906. [Google Scholar] [CrossRef] [PubMed]
- Wang Z, Li Z, Bal G, Franke K, Zuberbier T, Babina M. beta-arrestin-1 and beta-arrestin-2 Restrain MRGPRX2-Triggered Degranulation and ERK1/2 Activation in Human Skin Mast Cells. Front Allergy 2022, 3, 930233.
- Wang Z, Franke K, Bal G, Li Z, Zuberbier T, Babina M. MRGPRX2-Mediated Degranulation of Human Skin Mast Cells Requires the Operation of G(alphai), G(alphaq), Ca++ Channels, ERK1/2 and PI3K-Interconnection between Early and Late Signaling. Cells 2022, 11. [CrossRef]
- Bulfone-Paus, S.; Nilsson, G.; Draber, P.; Blank, U.; Levi-Schaffer, F. Positive and Negative Signals in Mast Cell Activation. Trends Immunol. 2017, 38, 657–667. [Google Scholar] [CrossRef]
- Vitallé, J.; Terrén, I.; Orrantia, A.; Bilbao, A.; Gamboa, P.M.; Borrego, F.; Zenarruzabeitia, O. The Expression and Function of CD300 Molecules in the Main Players of Allergic Responses: Mast Cells, Basophils and Eosinophils. Int. J. Mol. Sci. 2020, 21, 3173. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B.S.; O'Sullivan, J.A.; Chang, A.T.; Youngblood, B.A. Siglecs in allergy and asthma. Mol. Asp. Med. 2023, 90, 101104. [Google Scholar] [CrossRef]
- Mizrahi, S.; Gibbs, B.F.; Karra, L.; Ben-Zimra, M.; Levi-Schaffer, F. Siglec-7 is an inhibitory receptor on human mast cells and basophils. J. Allergy Clin. Immunol. 2014, 134, 230–233. [Google Scholar] [CrossRef]
- Arthur GK, Cruse G. Regulation of Trafficking and Signaling of the High Affinity IgE Receptor by FcepsilonRIbeta and the Potential Impact of FcepsilonRIbeta Splicing in Allergic Inflammation. Int J Mol Sci 2022, 23. [CrossRef]
- Gamperl, S.; Stefanzl, G.; Peter, B.; Smiljkovic, D.; Bauer, K.; Willmann, M.; Valent, P.; Hadzijusufovic, E. Effects of ibrutinib on proliferation and histamine release in canine neoplastic mast cells. Veter- Comp. Oncol. 2019, 17, 553–561. [Google Scholar] [CrossRef]
- Ponuwei, G.A. A glimpse of the ERM proteins. J. Biomed. Sci. 2016, 23, 1–6. [Google Scholar] [CrossRef]
- Tsukita, S.; Yonemura, S. Cortical Actin Organization: Lessons from ERM (Ezrin/Radixin/Moesin) Proteins. J. Biol. Chem. 1999, 274, 34507–34510. [Google Scholar] [CrossRef]
- Neisch AL, Fehon RG. Ezrin, Radixin and Moesin: key regulators of membrane-cortex interactions and signaling. Curr Opin Cell Biol 2011, 23, 377–382.
- García-Ortiz, A.; Serrador, J.M. ERM Proteins at the Crossroad of Leukocyte Polarization, Migration and Intercellular Adhesion. Int. J. Mol. Sci. 2020, 21, 1502. [Google Scholar] [CrossRef] [PubMed]
- Iontcheva, I.; Amar, S.; Zawawi, K.H.; Kantarci, A.; Van Dyke, T.E. Role for Moesin in Lipopolysaccharide-Stimulated Signal Transduction. Infect. Immun. 2004, 72, 2312–2320. [Google Scholar] [CrossRef] [PubMed]
- Lopez JP, Turner JR, Philipson LH. Glucose-induced ERM protein activation and translocation regulates insulin secretion. Am J Physiol Endocrinol Metab 2010, 299, E772–E785.
- Ben-Aissa, K.; Patino-Lopez, G.; Belkina, N.V.; Maniti, O.; Rosales, T.; Hao, J.-J.; Kruhlak, M.J.; Knutson, J.R.; Picart, C.; Shaw, S. Activation of Moesin, a Protein That Links Actin Cytoskeleton to the Plasma Membrane, Occurs by Phosphatidylinositol 4,5-bisphosphate (PIP2) Binding Sequentially to Two Sites and Releasing an Autoinhibitory Linker. J. Biol. Chem. 2012, 287, 16311–16323. [Google Scholar] [CrossRef]
- Matsui, T.; Maeda, M.; Doi, Y.; Yonemura, S.; Amano, M.; Kaibuchi, K.; Tsukita, S.; Tsukita, S. Rho-Kinase Phosphorylates COOH-terminal Threonines of Ezrin/Radixin/Moesin (ERM) Proteins and Regulates Their Head-to-Tail Association. J. Cell Biol. 1998, 140, 647–657. [Google Scholar] [CrossRef]
- Lazki-Hagenbach, P.; Klein, O.; Sagi-Eisenberg, R. The actin cytoskeleton and mast cell function. Curr. Opin. Immunol. 2021, 72, 27–33. [Google Scholar] [CrossRef]
- Suzuki, R.; Inoh, Y.; Yokawa, S.; Furuno, T.; Hirashima, N. Receptor dynamics regulates actin polymerization state through phosphorylation of cofilin in mast cells. Biochem. Biophys. Res. Commun. 2020, 534, 714–719. [Google Scholar] [CrossRef]
- Du, H.; Sun, N.; Han, S.; Song, R.; Che, H. Dok-1 regulates mast cell degranulation negatively through inhibiting calcium-dependent F-actin disassembly. Clin. Immunol. 2022, 238, 109008. [Google Scholar] [CrossRef]
- Ibanga, J.; Zhang, E.L.; Eitzen, G.; Guo, Y. Mast cell granule motility and exocytosis is driven by dynamic microtubule formation and kinesin-1 motor function. PLOS ONE 2022, 17, e0265122. [Google Scholar] [CrossRef]
- Navinés-Ferrer, A.; Ainsua-Enrich, E.; Serrano-Candelas, E.; Proaño-Pérez, E.; Muñoz-Cano, R.; Gastaminza, G.; Olivera, A.; Martin, M. MYO1F Regulates IgE and MRGPRX2-Dependent Mast Cell Exocytosis. J. Immunol. 2021, 206, 2277–2289. [Google Scholar] [CrossRef]
- Zhang, B.; Alysandratos, K.-D.; Angelidou, A.; Asadi, S.; Sismanopoulos, N.; Delivanis, D.-A.; Weng, Z.; Miniati, A.; Vasiadi, M.; Katsarou-Katsari, A.; et al. Human mast cell degranulation and preformed TNF secretion require mitochondrial translocation to exocytosis sites: Relevance to atopic dermatitis. J. Allergy Clin. Immunol. 2011, 127, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Storci, G.; Bonifazi, F.; Garagnani, P.; Olivieri, F.; Bonafè, M. The role of extracellular DNA in COVID-19: Clues from inflamm-aging. Ageing Res. Rev. 2020, 66, 101234–101234. [Google Scholar] [CrossRef] [PubMed]
- Andargie, T.E.; Tsuji, N.; Seifuddin, F.; Jang, M.K.; Yuen, P.S.; Kong, H.; Tunc, I.; Singh, K.; Charya, A.; Wilkins, K.; et al. Cell-free DNA maps COVID-19 tissue injury and risk of death and can cause tissue injury. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef]
- Costa TJ, Potje SR, Fraga-Silva TFC, da Silva-Neto JA, Barros PR, Rodrigues D, Machado MR, Martins RB, Santos-Eichler RA, Benatti MN, de Sa KSG, Almado CEL, Castro IA, Pontelli MC, Serra L, Carneiro FS, Becari C, Louzada-Junior P, Oliveira RDR, Zamboni DS, Arruda E, Auxiliadora-Martins M, Giachini FRC, Bonato VLD, Zachara NE, Bomfim GF, Tostes RC. Mitochondrial DNA and TLR9 activation contribute to SARS-CoV-2-induced endothelial cell damage. Vascul Pharmacol 2022, 142, 106946. [CrossRef]
- Edinger, F.; Edinger, S.; Koch, C.; Markmann, M.; Hecker, M.; Sander, M.; Schneck, E. Peak Plasma Levels of mtDNA Serve as a Predictive Biomarker for COVID-19 in-Hospital Mortality. J. Clin. Med. 2022, 11, 7161. [Google Scholar] [CrossRef] [PubMed]
- Therianou, A.; Vasiadi, M.; Delivanis, D.A.; Petrakopoulou, T.; Katsarou-Katsari, A.; Antoniou, C.; Stratigos, A.; Tsilioni, I.; Katsambas, A.; Rigopoulos, D.; et al. Mitochondrial dysfunction in affected skin and increased mitochondrial DNA in serum from patients with psoriasis. Exp. Dermatol. 2019, 28, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni I, Natelson B, Theoharides TC. Exosome-Associated Mitochondrial DNA from Patients with ME/CFS Stimulates Human Cultured Microglia to Release IL-1beta. Eur J Neurosci 2022. [CrossRef]
- Zhang, B.; Angelidou, A.; Alysandratos, K.-D.; Vasiadi, M.; Francis, K.; Asadi, S.; Theoharides, A.; Sideri, K.; Lykouras, L.; Kalogeromitros, D.; et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J. Neuroinflammation 2010, 7, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, W.; Theoharides, T.C.; Alper, S.L.; Douglas, W.W.; Greengard, P. Calcium-dependent protein phosphorylation during secretion by exocytosis in the mast cell. Nature 1978, 275, 329–331. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Sieghart, W.; Greengard, P.; Douglas, W.W. Antiallergic Drug Cromolyn May Inhibit Histamine Secretion by Regulating Phosphorylation of a Mast Cell Protein. Science 1980, 207, 80–82. [Google Scholar] [CrossRef]
- Kempuraj, D.; Madhappan, B.; Christodoulou, S.; Boucher, W.; Cao, J.; Papadopoulou, N.; Cetrulo, C.L.; Theoharides, T.C. Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. Br. J. Pharmacol. 2005, 145, 934–944. [Google Scholar] [CrossRef]
- Wang, L.; Correia, I.; Basu, S.; Theoharides, T.C. Ca2+ and phorbol ester effect on the mast cell phosphoprotein induced by cromolyn. Eur. J. Pharmacol. 1999, 371, 241–249. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Wang, L.; Pang, X.; Letourneau, R.; E Culm, K.; Basu, S.; Wang, Y.; Correia, I. Cloning and cellular localization of the rat mast cell 78-kDa protein phosphorylated in response to the mast cell "stabilizer" cromolyn. J. Pharmacol. Exp. Ther. 2000, 294. [Google Scholar]
- Theoharides, T.C. The mast cell: a neuroimmunoendocrine master player. Int. J. Tissue React. 1996, 18. [Google Scholar]
- Yang, H.-S.; Hinds, P.W. Phosphorylation of Ezrin by Cyclin-Dependent Kinase 5 Induces the Release of Rho GDP Dissociation Inhibitor to Inhibit Rac1 Activity in Senescent Cells. Cancer Res 2006, 66, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Alexandrakis, M.; Kempuraj, D.; Lytinas, M. Anti-inflammatory actions of flavonoids and structural requirements for new design. Int. J. Immunopathol. Pharmacol. 2003, 14. [Google Scholar]
- Olson, F.J.; Ludowyke, R.I.; Karlsson, N.G. Discovery and Identification of Serine and Threonine Phosphorylated Proteins in Activated Mast Cells: Implications for Regulation of Protein Synthesis in the Rat Basophilic Leukemia Mast Cell Line RBL-2H3. J. Proteome Res. 2009, 8, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Asano, S. Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin. Int. J. Mol. Sci. 2022, 23, 3246. [Google Scholar] [CrossRef]
- Zhao, S.; Luo, J.; Hu, J.; Wang, H.; Zhao, N.; Cao, M.; Zhang, C.; Hu, R.; Liu, L. Role of Ezrin in Asthma-Related Airway Inflammation and Remodeling. Mediat. Inflamm. 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Tabrizi, M.E.A.; Gupta, J.K.; Gross, S.R. Ezrin and Its Phosphorylated Thr567 Form Are Key Regulators of Human Extravillous Trophoblast Motility and Invasion. Cells 2023, 12, 711. [Google Scholar] [CrossRef]
- Doi, Y.; Itoh, M.; Yonemura, S.; Ishihara, S.; Takano, H.; Noda, T.; Tsukita, S.; Tsukita, S. Normal Development of Mice and Unimpaired Cell Adhesion/Cell Motility/Actin-based Cytoskeleton without Compensatory Up-regulation of Ezrin or Radixin in Moesin Gene Knockout. J. Biol. Chem. 1999, 274, 2315–2321. [Google Scholar] [CrossRef]
- Avery, L.; Robertson, T.F.; Wu, C.F.; Roy, N.H.; Chauvin, S.D.; Perkey, E.; Vanderbeck, A.; Maillard, I.; Burkhardt, J.K. A Murine Model of X-Linked Moesin-Associated Immunodeficiency (X-MAID) Reveals Defects in T Cell Homeostasis and Migration. Front. Immunol. 2022, 12, 726406. [Google Scholar] [CrossRef]
- Nakamura, F.; Amieva, M.R.; Furthmayr, H. Phosphorylation of Threonine 558 in the Carboxyl-terminal Actin-binding Domain of Moesin by Thrombin Activation of Human Platelets. J. Biol. Chem. 1995, 270, 31377–31385. [Google Scholar] [CrossRef] [PubMed]
- Shcherbina, A.; Kenney, D.M.; Bretscher, A.; Remold-O-Donnell, E. Dynamic association of moesin with the membrane skeleton of thrombin- activated platelets. Blood 1999, 93. [Google Scholar]
- Meyer, T.; Uher, T.; Schwartz, P.; Buchwald, A.B. Tyrosine Phosphorylation of Moesin in Arachidonic Acid-Stimulated Human Platelets. J. Thromb. Thrombolysis 1998, 6, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Hishiya, A.; Ohnishi, M.; Tamura, S.; Nakamura, F. Protein Phosphatase 2C Inactivates F-actin Binding of Human Platelet Moesin. J. Biol. Chem. 1999, 274, 26705–26712. [Google Scholar] [CrossRef]
- Nakamura, F.; Amieva, M.R.; Hirota, C.; Mizuno, Y.; Furthmayr, H. Phosphorylation of558T of Moesin Detected by Site-Specific Antibodies in RAW264.7 Macrophages. Biochem. Biophys. Res. Commun. 1996, 226, 650–656. [Google Scholar] [CrossRef]
- Charrin, S.; Alcover, A. Role of ERM (ezrin-radixin-moesin) proteins in T lymphocyte polarization, immune synapse formation and in T cell receptor-mediated signaling. Front. Biosci. 2006, 11, 1987–97. [Google Scholar] [CrossRef]
- Takamatsu H, Espinoza JL, Lu X, Qi Z, Okawa K, Nakao S. Anti-moesin antibodies in the serum of patients with aplastic anemia stimulate peripheral blood mononuclear cells to secrete TNF-alpha and IFN-gamma. J Immunol 2009, 182, 703–710. [CrossRef]
- Suzuki, K.; Nagao, T.; Itabashi, M.; Hamano, Y.; Sugamata, R.; Yamazaki, Y.; Yumura, W.; Tsukita, S.; Wang, P.-C.; Nakayama, T.; et al. A novel autoantibody against moesin in the serum of patients with MPO-ANCA-associated vasculitis. Nephrol. Dial. Transplant. 2013, 29, 1168–1177. [Google Scholar] [CrossRef]
- Südhof, T.C.; Rothman, J.E. Membrane Fusion: Grappling with SNARE and SM Proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef]
- Blank, U.; Cyprien, B.; Martin-Verdeaux, S.; Paumet, F.; Pombo, I.; Rivera, J.; Roa, M.; Varin-Blank, N. SNAREs and associated regulators in the control of exocytosis in the RBL-2H3 mast cell line. Mol. Immunol. 2002, 38, 1341–1345. [Google Scholar] [CrossRef]
- Lorentz, A.; Baumann, A.; Vitte, J.; Blank, U. The SNARE Machinery in Mast Cell Secretion. Front. Immunol. 2012, 3, 143. [Google Scholar] [CrossRef]
- Woska JR, Jr. , Gillespie ME. SNARE complex-mediated degranulation in mast cells. J Cell Mol Med 2012, 16, 649–656. [Google Scholar]
- Suzuki K, Verma IM. Phosphorylation of SNAP-23 by IkappaB kinase 2 regulates mast cell degranulation. Cell 2008, 134, 485–495.
- Janowicz, Z.A.; Melber, K.; Merckelbach, A.; Jacobs, E.; Harford, N.; Comberbach, M.; Hollenberg, C.P. Simultaneous expression of the S and L surface antigens of hepatitis B, and formation of mixed particles in the methylotrophic yeast,Hansenula polymorpha. Yeast 1991, 7, 431–443. [Google Scholar] [CrossRef]
- Frank, S.P.; Thon, K.-P.; Bischoff, S.C.; Lorentz, A. SNAP-23 and syntaxin-3 are required for chemokine release by mature human mast cells. Mol. Immunol. 2011, 49, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Hepp, R.; Puri, N.; Hohenstein, A.C.; Crawford, G.L.; Whiteheart, S.W.; Roche, P.A. Phosphorylation of SNAP-23 Regulates Exocytosis from Mast Cells. J. Biol. Chem. 2005, 280, 6610–6620. [Google Scholar] [CrossRef] [PubMed]
- Naskar, P.; Puri, N. Phosphorylation of SNAP-23 regulates its dynamic membrane association during Mast Cell exocytosis. Biol. Open 2017, 6, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kong, B.; Jung, Y.; Park, J.-B.; Oh, J.-M.; Hwang, J.; Cho, J.Y.; Kweon, D.-H. Soluble N-Ethylmaleimide-Sensitive Factor Attachment Protein Receptor-Derived Peptides for Regulation of Mast Cell Degranulation. Front. Immunol. 2018, 9, 725. [Google Scholar] [CrossRef]
- Gilliam FR, 3rd, Rivas PA, Wendt DJ, Starmer CF, Grant AO. Extracellular pH modulates block of both sodium and calcium channels by nicardipine. Am J Physiol 1990, 259, H1178–H1184.
- Puri, N.; Roche, P.A. Mast cells possess distinct secretory granule subsets whose exocytosis is regulated by different SNARE isoforms. Proc. Natl. Acad. Sci. 2008, 105, 2580–2585. [Google Scholar] [CrossRef] [PubMed]
- Paumet, F.; Le Mao, J.; Martin, S.; Galli, T.; David, B.; Blank, U.; Roa, M. Soluble NSF Attachment Protein Receptors (SNAREs) in RBL-2H3 Mast Cells: Functional Role of Syntaxin 4 in Exocytosis and Identification of a Vesicle-Associated Membrane Protein 8-Containing Secretory Compartment. J. Immunol. 2000, 164, 5850–5857. [Google Scholar] [CrossRef]
- Sander, L.E.; Frank, S.P.C.; Bolat, S.; Blank, U.; Galli, T.; Bigalke, H.; Bischoff, S.C.; Lorentz, A. Vesicle associated membrane protein (VAMP)-7 and VAMP-8, but not VAMP-2 or VAMP-3, are required for activation-induced degranulation of mature human mast cells. Eur. J. Immunol. 2008, 38, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Martin-Verdeaux, S.; Pombo, I.; Iannascoli, B.; Roa, M.; Varin-Blank, N.; Rivera, J.; Blank, U. Evidence of a role for Munc18-2 and microtubules in mast cell granule exocytosis. J. Cell Sci. 2003, 116, 325–334. [Google Scholar] [CrossRef]
- Yang, Y.; Oh, J.-M.; Heo, P.; Shin, J.Y.; Kong, B.; Shin, J.; Lee, J.-C.; Oh, J.S.; Park, K.W.; Lee, C.H.; et al. Polyphenols differentially inhibit degranulation of distinct subsets of vesicles in mast cells by specific interaction with granule-type-dependent SNARE complexes. Biochem. J. 2013, 450, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kim, S.-H.; Heo, P.; Kong, B.; Shin, J.; Jung, Y.-H.; Yoon, K.; Chung, W.-J.; Shin, Y.-K.; Kweon, D.-H. SNARE zippering is hindered by polyphenols in the neuron. Biochem. Biophys. Res. Commun. 2014, 450, 831–836. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Asadi, S.; Sismanopoulos, N.; Butcher, A.; Fu, X.; Katsarou-Katsari, A.; Antoniou, C.; Theoharides, T.C. Quercetin Is More Effective than Cromolyn in Blocking Human Mast Cell Cytokine Release and Inhibits Contact Dermatitis and Photosensitivity in Humans. PLOS ONE 2012, 7, e33805. [Google Scholar] [CrossRef]
- Fox, C.C.; Wolf, E.J.; Kagey-Sobotka, A.; Lichtenstein, L.M. Comparison of human lung and intestinal mast cells. J. Allergy Clin. Immunol. 1988, 81, 89–94. [Google Scholar] [CrossRef]
- Kandere-Grzybowska, K.; Kempuraj, D.; Cao, J.; Cetrulo, C.L.; Theoharides, T.C. Regulation of IL-1-induced selective IL-6 release from human mast cells and inhibition by quercetin. Br. J. Pharmacol. 2006, 148, 208–215. [Google Scholar] [CrossRef]
- Sieghart, W.; Theoharides, T.C.; Douglas, W.W.; Greengard, P. Phosphorylation of a single mast cell protein in response to drugs that inhibit secretion. Biochem. Pharmacol. 1981, 30, 2737–2738. [Google Scholar] [CrossRef]
- Patel, A.B.; Theoharides, T.C. Methoxyluteolin Inhibits Neuropeptide-stimulated Proinflammatory Mediator Release via mTOR Activation from Human Mast Cells. Experiment 2017, 361, 462–471. [Google Scholar] [CrossRef]
- Sandhu, J.K.; Kulka, M. Decoding Mast Cell-Microglia Communication in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 1093. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.S.; Redegeld, F.A. Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Giusti, P. Neuroinflammation, Microglia and Mast Cells in the Pathophysiology of Neurocognitive Disorders: A Review. CNS Neurol. Disord. Drug Targets 2014, 13, 1654–1666. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Zusso, M.; Giusti, P. Neuroinflammation, Mast Cells, and Glia: Dangerous Liaisons. Neurosci. 2017, 23, 478–498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Dong, H.; Xu, Y.; Zhang, S. Induction of Microglial Activation by Mediators Released from Mast Cells. Cell. Physiol. Biochem. 2016, 38, 1520–1531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, X.; Zhang, Y.; Qu, C.; Zhou, X.; Zhang, S. Histamine Induces Microglia Activation and the Release of Proinflammatory Mediators in Rat Brain Via H1R or H4R. J. Neuroimmune Pharmacol. 2019, 15, 280–291. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Yang, H.; Hu, G.; He, S. Mast Cell Tryptase Induces Microglia Activation via Protease-activated Receptor 2 Signaling. Cell. Physiol. Biochem. 2012, 29, 931–940. [Google Scholar] [CrossRef]
- Wang, Y.; Sha, H.; Zhou, L.; Chen, Y.; Zhou, Q.; Dong, H.; Qian, Y. The Mast Cell Is an Early Activator of Lipopolysaccharide-Induced Neuroinflammation and Blood-Brain Barrier Dysfunction in the Hippocampus. Mediat. Inflamm. 2020, 2020, 8098439. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Kavalioti, M.; Martinotti, R. Factors adversely influencing neurodevelopment. . 2020, 33, 1663–1667. [Google Scholar]
- Zhang, X.; Dong, H.; Li, N.; Zhang, S.; Sun, J.; Zhang, S.; Qian, Y. Activated brain mast cells contribute to postoperative cognitive dysfunction by evoking microglia activation and neuronal apoptosis. J. Neuroinflammation 2016, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kavalioti, M.; Tsilioni, I. Mast Cells, Stress, Fear and Autism Spectrum Disorder. Int. J. Mol. Sci. 2019, 20, 3611. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Ways to Address Perinatal Mast Cell Activation and Focal Brain Inflammation, including Response to SARS-CoV-2, in Autism Spectrum Disorder. J. Pers. Med. 2021, 11, 860. [Google Scholar] [CrossRef]
- Hu, C.; Li, H.; Li, J.; Luo, X.; Hao, Y. Microglia: Synaptic modulator in autism spectrum disorder. Front. Psychiatry 2022, 13, 958661. [Google Scholar] [CrossRef]
- Lampiasi, N.; Bonaventura, R.; Deidda, I.; Zito, F.; Russo, R. Inflammation and the Potential Implication of Macrophage-Microglia Polarization in Human ASD: An Overview. Int. J. Mol. Sci. 2023, 24, 2703. [Google Scholar] [CrossRef]
- Liao, X.; Chen, M.; Li, Y. The glial perspective of autism spectrum disorder convergent evidence from postmortem brain and PET studies. Front. Neuroendocr. 2023, 70, 101064. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Thangavel, R.; Selvakumar, G.P.; Ahmed, M.E.; Zaheer, S.; Raikwar, S.P.; Zahoor, H.; Saeed, D.; Dubova, I.; Giler, G.; Herr, S.; Iyer, S.S.; Zaheer, A. Mast Cell Proteases Activate Astrocytes Glia-Neurons Release Interleukin-33 by Activating, p. 3.8.; ERK1/2 MAPKs, N.F.-k.a.p.p.a.B. Mol Neurobiol 2019, 56, 1681–1693. [Google Scholar]
- Kempuraj, D.; Ahmed, M.E.; Selvakumar, G.P.; Thangavel, R.; Raikwar, S.P.; Zaheer, S.A.; Iyer, S.S.; Burton, C.; James, D.; Zaheer, A. Mast Cell Activation, Neuroinflammation, and Tight Junction Protein Derangement in Acute Traumatic Brain Injury. Mediat. Inflamm. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Okazaki, T.; Saito, D.; Inden, M.; Kawaguchi, K.; Wakimoto, S.; Nakahari, T.; Asano, S. Moesin is involved in microglial activation accompanying morphological changes and reorganization of the actin cytoskeleton. J. Physiol. Sci. 2020, 70, 1–11. [Google Scholar] [CrossRef]
- Luo T, Ou JN, Cao LF, Peng XQ, Li YM, Tian YQ. The Autism-Related lncRNA MSNP1AS Regulates Moesin Protein to Influence the RhoA, Rac1, and PI3K/Akt Pathways and Regulate the Structure and Survival of Neurons. Autism Res 2020, 13, 2073–2082.
- Hoshi Y, Uchida Y, Kuroda T, Tachikawa M, Couraud PO, Suzuki T, Terasaki T. Distinct roles of ezrin, radixin and moesin in maintaining the plasma membrane localizations and functions of human blood-brain barrier transporters. J Cereb Blood Flow Metab 2020, 40, 1533–1545.
- Theoharides, T. Mast cells: The immune gate to the brain. Life Sci. 1990, 46, 607–617. [Google Scholar] [CrossRef]
- Theoharides TC, Zhang B. Neuro-inflammation, blood-brain barrier, seizures and autism. J Neuroinflammation 2011, 8, 168.
- Theoharides, T.C.; Kempuraj, D. Role of SARS-CoV-2 Spike-Protein-Induced Activation of Microglia Mast Cells in the Pathogenesis of, N.e.u.r.o.-C.O.V.I.D. Cells 2023, 12. [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef] [PubMed]
- Jager, A.K.; Saaby, L. Flavonoids the, C. N.S. Molecules 2011, 16, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Calfio, C.; Gonzalez, A.; Singh, S.K.; Rojo, L.E.; Maccioni, R.B. The Emerging Role of Nutraceuticals and Phytochemicals in the Prevention and Treatment of Alzheimer’s Disease. J. Alzheimer's Dis. 2020, 77, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Thangavel, R.; Kempuraj, D.D.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Zaheer, S.A.; Iyer, S.S.; Govindarajan, R.; Chandrasekaran, P.N.; et al. Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. BioFactors 2020, 47, 190–197. [Google Scholar] [CrossRef]
- Calis, Z.; Mogulkoc, R.; Baltaci, A.K. The Roles of Flavonols/Flavonoids in Neurodegeneration and Neuroinflammation. Mini-Reviews Med. Chem. 2020, 20, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, TC. Could SARS-CoV-2 Spike Protein Be Responsible for Long-COVID Syndrome? MolNeurobiol 2022, 13, 1–12. [Google Scholar]
- Ashaari, Z.; Hadjzadeh, M.-A.; Hassanzadeh, G.; Alizamir, T.; Yousefi, B.; Keshavarzi, Z.; Mokhtari, T. The Flavone Luteolin Improves Central Nervous System Disorders by Different Mechanisms: A Review. J. Mol. Neurosci. 2018, 65, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Rezai-Zadeh K, Douglas SR, Bai Y, Tian J, Hou H, Mori T, Zeng J, Obregon D, Town T, Tan J. Flavonoid-mediated presenilin-1 phosphorylation reduces Alzheimer's disease beta-amyloid production. J Cell MolMed 2009, 13, 574–588.
- Yao, Z.-H.; Yao, X.-L.; Zhang, Y.; Zhang, S.-F.; Hu, J.-C. Luteolin Could Improve Cognitive Dysfunction by Inhibiting Neuroinflammation. Neurochem. Res. 2018, 43, 806–820. [Google Scholar] [CrossRef]
- Gratton G, Weaver SR, Burley CV, Low KA, Maclin EL, Johns PW, Pham QS, Lucas SJE, Fabiani M, Rendeiro C. Dietary flavanols improve cerebral cortical oxygenation and cognition in healthy adults. SciRep, 10, 19409. [CrossRef]
- Devi, S.A.; Chamoli, A. Polyphenols as an Effective Therapeutic Intervention Against Cognitive Decline During Normal and Pathological Brain Aging. 2020, 1260, 159–174. [CrossRef]
- Theoharides TC, Stewart JM, Hatziagelaki E, Kolaitis G. Brain "fog," inflammation and obesity: key aspects of 2 neuropsychiatric disorders improved by luteolin. FrontNeurosci 2015, 9, 225.
- Theoharides, T.C.; Cholevas, C.; Polyzoidis, K.; Politis, A. Long-COVID syndrome-associated brain fog and chemofog: Luteolin to the rescue. BioFactors 2021, 47, 232–241. [Google Scholar] [CrossRef]
- Stefano, G.B.; Büttiker, P.; Weissenberger, S.; Martin, A.; Ptacek, R.; Kream, R.M. Editorial: The Pathogenesis of Long-Term Neuropsychiatric COVID-19 and the Role of Microglia, Mitochondria, and Persistent Neuroinflammation: A Hypothesis. Experiment 2021, 27, e933015. [Google Scholar] [CrossRef]
- Hugon, J.; Msika, E.-F.; Queneau, M.; Farid, K.; Paquet, C. Long COVID: cognitive complaints (brain fog) and dysfunction of the cingulate cortex. J. Neurol. 2021, 269, 44–46. [Google Scholar] [CrossRef]
- Rezai-Zadeh, K.; Ehrhart, J.; Bai, Y.; Sanberg, P.R.; Bickford, P.; Tan, J.; Shytle, R.D. Apigenin and luteolin modulate microglial activation via inhibition of STAT1-induced CD40 expression. J. Neuroinflammation 2008, 5, 41–41. [Google Scholar] [CrossRef]
- Jang, S.; Kelley, K.W.; Johnson, R.W. Luteolin reduces IL-6 production in microglia by inhibiting JNK phosphorylation and activation of AP-1. Proc. Natl. Acad. Sci. 2008, 105, 7534–7539. [Google Scholar] [CrossRef]
- Patel, A.B.; Tsilioni, I.; Leeman, S.E.; Theoharides, T.C. Neurotensin stimulates sortilin and mTOR in human microglia inhibitable by methoxyluteolin, a potential therapeutic target for autism. Proc. Natl. Acad. Sci. 2016, 113, E7049–E7058. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Patel, A.B.; Panagiotidou, S.; Theoharides, T.C. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J. Allergy Clin. Immunol. 2014, 135, 1044–1052. [Google Scholar] [CrossRef]
- Seelinger, G.; Merfort, I.; Schempp, C.M. Anti-Oxidant, Anti-Inflammatory and Anti-Allergic Activities of Luteolin. Planta Medica 2008, 74, 1667–1677. [Google Scholar] [CrossRef] [PubMed]
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An Open-Label Pilot Study of a Formulation Containing the Anti-Inflammatory Flavonoid Luteolin and Its Effects on Behavior in Children With Autism Spectrum Disorders. Clin. Ther. 2013, 35, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Taliou, A.; Francis, K.; Theoharides, T.C. Children with autism spectrum disorders, who improved with a luteolin-containing dietary formulation, show reduced serum levels of TNF and IL-6. Transl. Psychiatry 2015, 5, e647–e647. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Guerra, L.; Patel, K. Successful Treatment of a Patient With Severe COVID-19 Using an Integrated Approach Addressing Mast Cells and Their Mediators. Int. J. Infect. Dis. 2022, 118, 164–166. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Kempuraj, D.D.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Zaheer, S.A.; Iyer, S.S.; Govindarajan, R.; Chandrasekaran, P.N.; et al. Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. BioFactors 2020, 47, 190–197. [Google Scholar] [CrossRef]
- Kirshenbaum, A.S.; Yin, Y.; Sundstrom, J.B.; Bandara, G.; Metcalfe, D.D. Description and Characterization of a Novel Human Mast Cell Line for Scientific Study. Int. J. Mol. Sci. 2019, 20, 5520. [Google Scholar] [CrossRef]
- Luo, Y.; Vallone, V.F.; He, J.; Frischbutter, S.; Kolkhir, P.; Moñino-Romero, S.; Stachelscheid, H.; Streu-Haddad, V.; Maurer, M.; Siebenhaar, F.; et al. A novel approach for studying mast cell–driven disorders: Mast cells derived from induced pluripotent stem cells. J. Allergy Clin. Immunol. 2021, 149, 1060–1068. [Google Scholar] [CrossRef]
- L'Homme L, Dombrowicz D. In vitro models of human mast cells: How to get more and better with induced pluripotent stem cells? J Allergy Clin Immunol 2022, 149, 904–906. [CrossRef]
- de Toledo, M.A.S.; Fu, X.; Maié, T.; Buhl, E.M.; Götz, K.; Schmitz, S.; Kaiser, A.; Boor, P.; Braunschweig, T.; Chatain, N.; et al. KIT D816V Mast Cells Derived from Induced Pluripotent Stem Cells Recapitulate Systemic Mastocytosis Transcriptional Profile. Int. J. Mol. Sci. 2023, 24, 5275. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Different Modes of Secretion of Mediators from Mast Cells.
| Degranulation (exocytosis) |
| Compound exocytosis |
| Piece meal degranulation |
| Transgranulation |
| Directed degranulation |
| Vesicular (differential) release of mediators |
| Extracellular microvesicles (exosomes) |
| Nanotubules |
| Immunologic synapses |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



