
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
Lack of water and salinity are common problems in many parts of the world. Therefore, some types of cacti can represent as promising crops. Therefore, the ability of cactus species to survive and adapt under natural stress conditions should be evaluated. In this study, it was aimed to evaluate the effect of salt stress on germination and emergence of pitaya seeds obtained from different fruit flesh colors, priming with plant growth regulators, namely salicylic acid (SA), oxalic acid (OA) and mepiquat chloride (MC). The experiment had a completely randomized design with a 2×4×3 factorial scheme corresponding to two pitaya cultivars (white and red fleshed), four NaCl concentrations (0, 2500, 5000 and 10000 ppm), and three PGRs (150 ppm/MC, SA, OC). The following variables were analyzed; germination percentage, mean germination time (MGT), germination stress tolerance ındex (GSTI), shoot length, seedling fresh weight and seedling dry weight ratio. According to the results, it was determined that the growing environment of the seed was important in salt stress resistance, while seeds matured in the environment with red fruit flesh were more tolerant to salt stress than white. Although Pitaya species are relatively salt tolerant, growth (about 30%) was significantly reduced above 2500 ppm and germination (about 45%) above 5000 ppm. MC was found promising as a priming agent in seeds obtained from fruits with white fruit flesh, and OA was found promising as a priming agent in seeds obtained from fruits with red fruit flesh in reducing NaCl-induced stress. Germination percentage stood out as the most important trait determining seed quality and was found to have positive effects on germination stress tolerance index (r: 0.63), seedling length (r: 0.74) and fresh seedling weight (r: 0.56). It is thought that the results obtained may contribute to pitaya cultivation and breeding.
Keywords:
Cactaceae
; seed viability
; seedling quality
; salinity
; PGRs
; Hylocereus sp.
1. Introduction
Pitaya, in the genus Hylocereus, is popularly consumed due to its aroma. In addition, in the current pandemic, the consumer’s interest in pitaya fruit, which is rich in chemicals with high antioxidant effects such as phenols, organic acids, minerals and vitamins, has increased [1,2,3]. This antioxidative effect has been proven to prevent many diseases [4,5]. The fruit peel of pitaya, which contains more than 50% unsaturated fatty acids (linoleic and linolenic) in its seeds [6], is a source of pectin and is very important in food and textile dyeing with the natural colorants it contains [7]. The demands of pitaya, which has the potential for many different uses for organoleptic, nutraceutical and industrial purposes, has increased, reflected in its production. As a matter of fact, all countries where pitaya is cultivated emphasize that the amount of production has increased compared to the previous year [8].
Sustainable and economic continuation of this high demand is only possible if breeders develop genotypes that meet the demands of producers, consumers and industry. For this reason, recently, efforts have been accelerated to develop early, middle and late varieties with high yield and attractiveness, resistant to biotic and abiotic stress conditions in combination, self-fertile, rich in chemicals with high antioxidant effect, suitable for different cultivation systems (open/greenhouse) [9]. Due to the quantitative nature of the desired traits and the complexity of inheritance mechanisms, hybrid breeding is still the most widely used method [10]. However, information on the factors affecting the germination of cactus seeds from the genus Hylocereus is very limited [11].
Germination depends on the chemical composition and testa permeability of the seed and favorable environmental conditions [12]. Salinity is a major environmental problem worldwide, especially in arid and semi-arid regions, limiting or making cultivation economically unfeasible. With global climate change, saline areas are expected to increase due to irrigation irregularity and high evaporation [13]. It has been pointed out that cactus species such as pitaya, which are particularly adaptable to different soil conditions, may be an alternative cultivation option for utilizing these areas [14,15].
By acting on the osmotic potential of the substrate, salts reduce the potential gradient between the substrate and the seed surface, limiting water uptake by the seed. They disrupt ion balance and negatively affect stomatal movement and photosynthesis [16]. Enzyme activity is impaired, and metabolic activity is restricted. Increased reactive oxygen species disrupt cell integrity and homeostasis [17,18,19]. Due to these reasons, it is not possible to maintain production sustainably. Although salt stress occurs in all plants, tolerance levels and reduction in growth and development rates differ among species and phenological stages [20]. Studies on the germination response of seeds exposed to artificial stress conditions are important ecophysiological tools that can be used to understand the ability of species to survive and adapt to natural stress conditions. These tools are also used to assess the sensitivity of these species and their ability to survive and adapt to suppression when exposed to unfavorable and novel environments [21]. Therefore, although salinity has been extensively analyzed in cultivated species, the effects of NaCl in cacti are poorly documented.
The positive effects of exogenously applied growth regulators on sustainability have been demonstrated in studies with different species to increase plant tolerance to various abiotic stress factors [22]. Salicylic acid (SA), oxalic acid (OA) or mepiquat chloride (MC) applied to plants or seeds regulate stomatal movements and contribute to ion uptake and transport as important signaling molecules [23,24,25]. They reduce the negative effects of reactive oxygen species (ROS) by protecting the structure and function of the cell membrane and increasing the synthesis of antioxidant-derived compunds. This allows the maintenance of osmolytes and homeostasis [26]. They promote germination and development by maintaining photosynthetic activity [27,28,29], regulate the expression of genes involved in synthesizing vital amino acids under stress [30] and restrict the development of different pathogens and have inducing effects on the acquisition of resistance [31,32,33,34].
This study evaluated the mitigating effect of some plant growth regulators on the performance of different pitaya species under different salt stress conditions during seed germination and seedling emergence periods under controlled laboratory conditions.
2. Materials and Methods
2.1. Plant Materials
Ripe fruits of Siam Red and Vietnam White pitaya varieties with different flesh colors were used in the study. The fruits were peeled, and the seeds were separated from the pulp manually and washed with tap water several times to remove mucilages and the pulp left over. Seeds were treated with 2% sodium hypochlorite for 3 min for surface sterilization.
2.2. Preparing Solutions and Performing Seed Priming
The dried seeds were separated according to the treatment groups, weighed and placed in Petri dishes. Salicylic acid, mepiquat chloride or oxalic acid solution with a concentration of 150 ppm was added to the petri dishes at five times the seed weight. The Petri dishes were wrapped with aluminum foil and kept in a growth chamber at 25˚C for 24 hours.
2.3. Experimental Layout
Following the priming application, the seeds were counted and grouped. The study carried out as a laboratory study, was designed with three replicates and each replicate consisted of 50 seeds. The seeds of each replicate were homogeneously distributed in 3 layers of Whatman filter paper, two on the bottom and one on top of the seeds, and 7 ml one of salt solution (2500 ppm, 5000 ppm, 10 000 ppm) or pure water (control) was applied for each filter paper (21 ml for each replicate at start). The homogeneously soaked filter papers were folded into zip lock bags to minimize moisture loss and placed in a growth chamber set at 25˚C, ~650 lux and 12/12 h photoperiod for germination. For 20 days, counts were made every day, and germination percentage (%), mean germination time and germination stress tolerance index were calculated at the end of the 20th day. On the 45th day following the establishment of the experiment, ten seedlings randomly selected from each replicate were measured for seedling shoot length (mm) using a caliper (Traceable - 6, VWR International, Milan, Italy) sensitive 0.01 and seedling fresh weight (mg) using a precision balance (Sartorius - CPA 16001S, Göttingen, Germany) sensitive to 0.001 g. Then, the samples were kept in an oven at 70 ˚C for 48 hours, and their dry weights were determined with the help of same precision balance. As a result of the ratio of dry weight to fresh weight multiplied by 100, the amount of dry matter per unit amount was found to be proportional (%) [35].
2.4. Germination-Related Traits
Seeds with radicles reaching 2 mm in length were considered for all parameters. The germination percentage was determined according to ISTA [36] rules.
Mean germination time (MGT) was calculated to evaluate the speed of germination as defined by ISTA [36] with followin formula. MGT= Σ(Dn)/Σn, where n is the seed number germinated on day D, and D is the number of days from the beginning of the germination test.
Germination stress tolerance index (GSTI, %) = [nd2 (1.00) + nd4 (0.75) + nd6 (0.5) + nd8 (0.25) of stressed seeds / nd2 (1.00) + nd4 (0.75) + nd6 (0.5) + nd8 (0.25) of control seeds] × 100, where n is the number of seeds germinated at day d [37].
2.5. Statistical Analysis
The experimental design was a 3-factor factorial, arranged in a completely randomized design with three replicates. Analysis of variance and comparison of means was performed by the MSTAT-C program (Michigan State University v. 2.10). Determination of the relations among the characteristics was revealed with correlation analysis by using Minitab-17 (Minitab Inc., State College, Pennsylvania, USA) and results were expressed with correlations coefficients.
3. Results and Discussion
Agricultural production depends on the suitability of ecological factors, and sometimes it is necessary to cultivate under different stress conditions [38]. In these cases, it can be critical to know the tolerance of the material to the prevailing stress conditions or the practices to counteract this stress [39]. Due to the lower water flux caused by the osmotic effect, salt stress affects seed metabolism and can lead to the inhibition of reserve mobilization [40]. The study’s results confirmed this, as the increase in salt concentration slowed the seed germination and caused a decrease in all germination and seedling traits except dry matter content (Table 1). Some species/varieties/genotypes can develop resistance or tolerance to certain stress factors through the mechanisms they possess. In this study, the seeds obtained from the Siam Red cultivar with red fruit flesh showed significantly higher performance than the Vietnam White with white fruit flesh color in terms of the examined traits (Table 1). Besides, using PGRs that regulate different physiological events can successfully alleviate stress effects [41,42]. OA used as a priming agent, was effective in terms of germination stress tolerance, germination percentage, mean germination time and fresh seedling weight. At the same time, MC was effective regarding seedling length and dry matter accumulation (Table 1). In addition, in some of the traits examined, double or triple interactions of the factors also showed significant effects.
Findings on the effect of the flesh color of the fruit from which the seed was obtained, varying levels of salt stress and priming agents (SA, OA and MC) on germination percentage and mean germination time of pitaya seeds are given in Table 2.
An increase in salt concentration increases the osmotic pressure of the seed and inhibits water uptake. Disruption of ion balance and increased reactive oxygen species lead to cell integrity and homeostasis disruption. It acts on different chemical components of the seed, disrupting the metabolic cycle. All these conditions lead to loss of seed viability or reduced performance [16,17,18,19]. The study’s findings were similar to the previous studies. In parallel with the increase in salt concentration, there was a decrease in the germination rate and an increase in the mean germination time. While the germination rate, determined as 98.44% in the control, was not affected much by 2500 ppm (90.00%) and 5000 ppm (89.50%) salt concentrations, it decreased sharply to 55.03% at 10 000 ppm. There are reports of decreased seed germination at increasing salt concentrations in different species such as cotton [37] and cucumber [43], including pitaya [44]. Ascending salt concentrations not only prevents seed germination but also extends the germination time by delaying the start of germination [45].
When all the study factors were considered together, the germination percentage (85.33%) of the seeds of the Siam Red was higher than that of Vietnam White (81.16%). In contrast, the mean germination time decreased from 11.24 days in Vietnam White to 7.33 days in Siam Red. In studies examining pitaya varieties with different flesh colors, the juices of varieties with red flesh color were found to have higher values in terms of pH and antioxidant activity [46,47]. The better performance of the Siam Red under saline conditions may be because its seeds are realized in a halophytic environment from the formation and development stage. Indeed, in Anabasis setifera, the maternal environment has been proven to affect resistance to salt stress significantly [48]. In addition, the high level of antioxidant activity may have contributed to lower negative effects caused by ROS. Seeds of different pitaya species were germinated under salt stress [35], and 33% decrease in the germination rate of H. undatus with white fruit flesh was observed, while no decrease was reported in H. polyrhizus with red fruit flesh. Similarly, the reduction in the germination rate of seeds from white fruit flesh (62%) was higher than that of red (46%). In conclusion, it was emphasized that H. polyrhizus is more tolerant to salt stress than H. Undatus similar with current study.
Priming practices may allow sustainable cultivation in areas where soil or irrigation water has high salinity before sowing. In these areas where ecological factors are unsuitable, it is vital to determine species-specific priming agents [22,37]. In this study, no statistically significant differences were detected between SA, OA and MC applied for this purpose regarding germination rate and germination speed. At the same time, all three PGRs were observed to make a positive contribution. In both Siam Red (99.33%) and Vietnam White (98.33%) cultivars, SA treatment was prominent in triggering germination under non-stress conditions. However, when germination occurred under saline conditions, OA (48.67%) was favored in Vietnam White and MC (72.00%) in Siam Red, and the same was observed for the mean germination time (Table 2). It has been reported that seeds of different species, including pitaya, showed positive effects on germination speed and rate after priming with SA, OA or MC. These positive effects have been proven to be because these chemicals act as signaling agents, maintain ion homeostasis, maintain membrane stability, suppress ROS and are involved in different physiological process [22,23,24,25,26].
The results of how the shoot length, fresh seedling weight and seedling dry weight ratio of the seedlings were affected by the combined effect of the factors are given in Table 3. According to the results, regardless of the treatments, an increase in seedling dry matter content and decreases in shoot length and fresh seedling weight were observed in parallel with the increase in salt concentration. At the same time, the values varied between 2.87% - 7.34%, 15.26 mm - 41.56 mm and 8.78 mg - 22.85 mg, respectively. Our results are in agreement with other reports conducted on different species, such as sunflower (Kaya et al., 2019), sugar beet [49] and pitaya [35]. Increased salt concentration in the seed interferes with the conversion of macro-molecules, which are a source for respiration, limiting the formation of soluble sugars necessary for embryo development. This disruption of the energy flow causes germination not to occur at all, to stop at a certain stage or to retard development [16,17,22].
Genotypes show high variation in salt tolerance through different adaptation pathways. Shoot length, fresh seedling weight and seedling dry matter content, which are defined as seedling development and quality criteria, were higher in Siam Red (31.48 mm, 21.23 mg and 4.65%, respectively) than in Vietnam White (27.90 mm, 12.23 mg and 4.44%, respectively), suggesting that H. polyrhizus is more tolerant to salt stress. Studies have shown that pitaya seeds with red fruit flesh have higher oil and sugar reserves [50]. Betalain derivatives, a natural osmolyte and antioxidant, K, which has a high ion exchange potential with Na, hydrolytic enzymes that catalyze metabolic events and antioxidants that eliminate the negative effects caused by excessive ROS are found at higher levels in H. polyrhizus with red fruit flesh [51,52,53]. The cumulative effect of both seed composition and maternal environment factors that favor tolerance to salt stress allowed for better results in H. polyrhizus seedlings growing under salt stress.
When factor effects were analyzed together, there was no difference between PGRs in terms of shoot length and seedling dry matter content, while OA (17.04 mg) and MC (16.96 mg) were more effective than SA (16.28 mg) in terms of fresh seedling weight. At the highest salt stress dose of 10 000 ppm, OA was found to be the highest in shoot length (15.62 mm), fresh seedling weight (9.14 mg) and seedling dry matter content (4.60%). The prominence of OA, an important osmoregulatory and antioxidant known to be positively correlated with K availability in stress conditions [54], may be interpreted as the seed trying to maintain ion homeostasis primarily to relieve salt stress. In the remediation of different heavy metals, exogenous application of OA to plants increased tolerance, enabling higher levels of heavy metal accumulation. In addition, among the grapevine rootstocks, it was emphasized that the genotype with the highest resistance to Na stress was the one containing the highest level of OA, and it was reported to show significant effects in eliminating Na stress [55].
The results of the stress tolerance index, which is the cumulative effect of the traits summarized above, detailed according to the factors, are given in Figure 1. When the effect of other factors is ignored, the increase in salt concentration causes the stress tolerance index to decrease, which was also reported by [37] and [43]. However, the fact that seeds obtained from fruits with red fruit flesh were more resistant to salinity-induced stress can be interpreted as the effect of the maternal environment. Maternal environment, defined as the mother’s effects other than gene effects, has shown significant effects on resistance to different biotic and abiotic stress conditions [10,48]. It was observed that all PGRs applied as priming agents were effective in reducing stress conditions but they were ranked as OA>MC>SA. However, the interactions of the factors on the stress tolerance index were found to be significant. According to the results, MC was more effective as a priming agent in seeds obtained from fruits with white fruit flesh, while a similar effect was found in seeds with red fruit flesh due to OA treatment (Figure 1). SA, OA and MC are widely used to alleviate the negative effects caused by different stress conditions [27,28,29,33,34].
The results of the relations between the traits examined in the study are given in Figure 2. Since the development in the plumule leads to an increase in biomass, there was a high positive correlation between seedling length and fresh seedling weight (r: 0.84). However, since the development in tissues caused an increase in intercellular spaces, a high negative correlation was found between shoot length and seedling dry matter content (r: -0.89). It is known that an increase in intercellular spaces leads to a decrease in the amount of dry matter accumulated per unit area [56]. Mean germination time is one of the important criteria for determining seed strength. If the time is prolonged, a decrease in germination rate and developmental retardation in germinated seedlings are observed [45]. The results of the study were in this direction, and strong negative relationships were found between mean germination time and germination percentage (r: -0.83), seedling length (r: -0.88) and fresh seedling weight (r: -0.81). Conversely, the increase in germination stress tolerance positively affected shoot length (r: 0.90), fresh seedling weight (r: 0.80) and germination percentage (r: 0.63), which are expressed as quality seed parameters, while shortening the mean germination time (r: -0.84). However, a negative correlation (r: -0.77) was calculated between GSTI and SDMR due to the increase in intercellular space in seedlings with vigorous growth. Germination percentage, which is one of the most important parameters determining seed quality, contributed to the increase in stress tolerance index (r: 0.63) and plant biomass (r: 0.56) while decreasing the mean germination time (r: -0.84). The results are consistent with previous studies [37,49].
5. Conclusions
This study investigated the effect of some plant growth regulators on the performance of seeds obtained from pitaya fruits with different fruit flesh colors under different salt stress conditions during germination and seedling emergence periods. According to the results, it was determined that the environment in which the seed was grown was important in salt stress resistance, while seeds matured in the environment with red fruit flesh were more tolerant to salt stress than white. Although pitaya species are relatively salt tolerant, growth of seedlings was significantly reduced above 2500 ppm and germination percentage was started to decrease with 5000 ppm. In reducing the stress caused by NaCl, MC can be used as a priming agent in seeds obtained from fruits with white fruit flesh, and OA can be used as a priming agent in seeds obtained from fruits with red fruit flesh.
Author Contributions
Design of the research was done by KM and NK. KM carriedt out the research. KM and MSD made the statistical analyzes and interpretation of the results. AMÇ, BBK and KM wrote the original draft. BBK and KM edited the final version of the article. All authors read and approved the final draft of the manuscript.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crane, JH.; Balerdi, CF. Pitaya growing in the Florida home landscape. 2005, IFAS Extension. Florida, United States.
- Wu, LC. ; Hsu, HW. ; Chen, YC. ; Chiu, CC. ; Lin, YI.; Ho, JAA. Antioxidant and antiproliferative activities of red pitaya. Food Chem. 2006, 95, 319–327. [CrossRef]
- Esquivel, P.; Stintzing, FC.; Carle, R. Comparison of morphological and chemical fruit traits from different pitaya genotypes (Hylocereus sp.) grown in Costa Rica. J. Appl. Bot. Food Qual. 2007, 81, 7–14. [Google Scholar]
- Cheok, A.; Xu, Y.; Zhang, Z.; Caton, PW.; Rodriguez-Mateos, A. Betalain-rich dragon fruit (pitaya) consumption improves vascular function in men and women: A double-blind, randomized controlled crossover trial. The American journal of clinical nutrition. 2022, 115, 1418–1431. [Google Scholar] [CrossRef] [PubMed]
- Nishikito, DF.; Borges, ACA. ; Laurindo, LF.; Otoboni, AMB.; Direito, R.; Goulart, RDA.; Nicolau, CCT.; Fiorini, AMR.; Sinatora, SM.; Barbalho, SM. Anti-inflammatory, antioxidant, and other health effects of dragon fruit and potential delivery systems for ıts bioactive compounds. Pharmaceutics. 2023, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Ariffin, AA.; Bakar, J.; Tan, CP.; Rahman, RA.; Karim, R.; Loi, CC. Essential fatty acids of pitaya (dragon fruit) seed oil. Food chem. 2009, 114, 561–564. [Google Scholar] [CrossRef]
- Mahmud, MH.; Raihan, MT.; Shakhik, MTZ.; Khan, FT.; Islam, MT. Dragon fruit (Hylocereus polyrhizus): A green colorant for cotton fabric. Colorants, 2023, 2, 230–244. [CrossRef]
- Mercado-Silva, EM. Pitaya—Hylocereus undatus (haw). Exotic fruits. 2018, 339–349. [Google Scholar]
- Tel-Zur, N. Pitahayas: Introduction, agrotechniques, and breeding. In VII International Congress on Cactus Pear and Cochineal. 2010, 995, 109–115. [Google Scholar] [CrossRef]
- Evrenosoğlu, Y.; Mertoğlu, K.; Bilgin, NA.; Misirli, A.; Özsoy, AN. Inheritance pattern of fire blight resistance in pear. Scientia horticulturae. 2019, 246, 887–892. [Google Scholar] [CrossRef]
- Suarez-Roman, RS.; Caetano, CM.; Ramírez, H.; Morales, J.G. Caracterización morfoanatómica y fisiológica de semilla sexual de pitahaya amarilla Selenicereus megalanthus (Haw. ) Britt & Rose. Revista de la Asociación Colombiana de Ciencias Biológicas. 2012, 24, 97–111. [Google Scholar]
- Carvalho, NM.; Nakagawa, J. Sementes: ciência, tecnologia e produção [Seeds: science, technology and production], 2012, Funep. 5th ed. Jaboticabal, Brazil.
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi journal of biological sciences. 2015, 22(2), 123–131. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, Y.; Nerd, A.; Sitrit, Y. New Fruits for arid climates. In Janick J, Whipkey A (eds.) Trends in new crops and new uses. 2002, ASHS Press, Alexandria. Virginia, United States.
- Tel-Zur, N.; Abbo, S.; Bar-Zvi, D.; Mizrahi, Y. Genetic relationships among Hylocereus and Selenicereus vine cacti (Cactaceae): Evidence from hybridization and cytological studies. Ann. Bot. 2004, 94, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Parihar, P.; Singh, S.; Singh, R.; Singh, VP.; Prasad, SM. Effect of salinity stress on plants and its tolerance strategies: A review. Environmental science and pollution research, 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Oliveira, AB.; Alencar, NLM. ; Gallão, MI.; Gomes Filho, E. Avaliação citoquímica durante a germinação de sementes de sorgo envelhecidas artificialmente e osmocondicionadas, sob salinidade [Cytochemical evaluation during the germination of artificial aged and primed sorghum seeds under salinity]. Rev. Ciênc. Agron. 2011, 42, 223–231. [Google Scholar]
- Singh, A.L.; Hsriprasanna, K.; and Chaudhari, V. Differential nutrients absorption an important tool for screening and identification of soil salinity tolerant peanut genotypes. Indian Journal of Plant Physiology. 2016, 21, 83–92. [Google Scholar] [CrossRef]
- Isayenkov, SV.; Maathuis, FJ. Plant salinity stress: Many unanswered questions remain. Frontiers in plant science, 2019, 10, 80. [Google Scholar] [CrossRef]
- Parida, AK.; Das, AB. Salt tolerance and salinity effects on plants: a review. Ecotoxicology and Environmental Safety. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Pereira, MRR. ; Martins, CC.; Souza, GSF.; Martins, D. Influência do estresse hídrico e salino na germinação de Urochloa decumbens e Urochloa ruziziensis [Influence of saline and water stress on germination of Urochloa decumbens and Urochloa ruziziensis]. Biosci. J. 2012, 28, 537–545. [Google Scholar]
- Hussain, M.; Farooq, S.; Hassan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management throughout breeding and agronomic alternatives. Agric Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Shaar-Moshe, L.; Blumwald, E.; Peleg, Z. Unique physiological and transcriptional shifts under combinations of salinity, drought, and heat. Plant Physiol. 2017, 174, 421–434. [Google Scholar] [CrossRef]
- Prasad, R.; Shivay, YS. Oxalic acid/oxalates in plants: From self-defence to phytoremediation. Current science. 2017, 1665–1667. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Pál, M. Salicylic acid signalling in plants. Int. J. Mol. Sci. 2020, 21, 2655. [Google Scholar] [CrossRef] [PubMed]
- Sakouhi, L.; Kharbech, O.; Massoud, MB.; Munemasa, S.; Murata, Y.; Chaoui, A. Oxalic acid mitigates cadmium toxicity in Cicer arietinum L. germinating seeds by maintaining the cellular redox homeostasis. Journal of Plant Growth Regulation. 2022, 1–13. [Google Scholar] [CrossRef]
- Srivastava, L.M. Gibberellins. In: Srivastava, L.M. Plant growth and development Academic Press, New York, USA. 2022, pp. 172-181.
- Siebert, J.D.; Stewart, A.M. Influence of plant density on cotton response to mepiquat chloride application. Agron. J. 2006, 98, 1634–1639. [Google Scholar] [CrossRef]
- Mahdavian, K.; Ghorbanli, M.; Kalantari, K.M. Role of salicylic acid in regulating ultraviolet radiation ınduced oxidative stress in pepper leaves. Russ. J. Plant Physiol. 2008, 55, 560–563. [Google Scholar] [CrossRef]
- Tung.
- Lehner, A.; Meimoun, P.; Errakhi, R.; Madiona, K.; Barakate, M.; Bouteau, F. Toxic and signaling effects of oxalic acid: natural born killer or natural born protector? Plant Signaling and Behavior, 2008, 3, 746–748. [Google Scholar] [CrossRef]
- Wang, Q.; Lai, T.; Qin, G.; Tian, S. Response of jujube fruits to exogenous oxalic acid treatment based on proteomic analysis. Plant Cell Physiology. 2009, 50(2), 230–242. [Google Scholar] [CrossRef]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef]
- Zhang, QC.; Deng, XX.; Wang, JG. The effects of mepiquat chloride (DPC) on the soluble protein content and the activities of protective enzymes in cotton in response to aphid feeding and on the activities of detoxifying enzymes in aphids. BMC Plant Biology 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Carvalho, SMC. ; Paiva, EPD.; Torres, SB.; Neta, MLDS.; Leite, MDS.; Benedito, CP.; Albuquerque, CCD.; Sá, FVDS. Pre-germination treatments in pitaya (Hylocereus spp.) seeds to attenuate salt stress. Revista Ciência Agronômica, 2022, 53. [Google Scholar]
- ISTA, (2003): International Rules for Seed Testing. – International Seed Testing Association, Switzerland.
- Ergin, N.; Kulan, E.; Gözükara, M.; Kaya, M.; Çetin, Ş.; Kaya, MD. Response of germination and seedling development of cotton to salinity under optimal and suboptimal temperatures. Kahramanmaraş Sütçü İmam Üniversitesi Tarım ve Doğa Dergisi. 2021, 24, 108–115. [Google Scholar] [CrossRef]
- Ibrahim, EA. Seed priming to alleviate salinity stress in germinating seeds. Journal of plant physiology. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, AC.; Coutinho, G. Nitrogênio no desenvolvimento inicial de mudas de pitaya vermelha. Global Science Technology. 2019, 12, 32–43. [Google Scholar]
- Freire, MHDC. ; Sousa, GGD.; de Souza, MV.; de Ceita, ED.; Fiusa, JN.; Leite, KN. Emergence and biomass accumulation in seedlings of rice cultivars irrigated with saline water. Revista Brasileira de Engenharia Agrícola e Ambiental. 2018, 22, 471–475. [Google Scholar] [CrossRef]
- Guirra, KS.; Torres, SB.; Leite, MDS. ; Guirra, BS.; Nogueira Neto, FA.; Rêgo, AL. Phytohormones on the germination and initial growth of pumpkin seedlings under different types of water. Revista Brasileira de Engenharia Agrícola e Ambiental. 2020, 24, 827–833. [Google Scholar] [CrossRef]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, CV.; Van Breusegem, F.; Gechev, T. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnology advances. 2020, 40, 107503. [Google Scholar] [CrossRef]
- Demir, I.; Kuzucu, CO.; Ermis, S.; Öktem, G. Radicle emergence as seed vigour test estimates seedling quality of hybrid cucumber (Cucumis sativus L.) cultivars in low temperature and salt stress conditions. Horticulturae, 2022, 9, 3. [Google Scholar] [CrossRef]
- Ortiz, TA.; Gomes, GR.; Takahashi, LUSA. ; Urbano, MR.; Strapasson, E. Water and salt stress in germinating seeds of pitaya genotypes (Hylocereus spp.). African Journal of Agricultural Research, 2014, 9, 3610–3619. [Google Scholar]
- Kaya, MD.; Akdoğan, G.; Kulan, EG.; Dağhan, H.; Sari, A. Salinity tolerance classification of sunflower (Helianthus annuus L.) and safflower (Carthamus tinctorius L.) by cluster and principal component analysis. Applied Ecology and Environmental Research. 2019, 17, 3849–3857. [Google Scholar] [CrossRef]
- Escribano, J.; Pedreño, MA.; García-Carmona, F.; Muñoz, R. Characterization of the antiradical activity of betalains from Beta vulgaris L. roots. Phytochemical Analysis: An International Journal of Plant Chemical and Biochemical Techniques, 1998, 9, 124–127. [Google Scholar] [CrossRef]
- Tamby Chik, C.; Bachok, S.; Baba, N.; Abdullah, A.; Abdullah, N. Quality characteristics and acceptability of three types of pitaya fruits in a consumer acceptance test. Journal of Tourism, Hospitality & Culinary Arts (JTHCA). 2011, 3, 89–98. [Google Scholar]
- El-Keblawy, A.; Gairola, S.; Bhatt, A. Maternal salinity environment affects salt tolerance during germination in Anabasis setifera: A facultative desert halophyte. Journal of Arid Land. 2016, 8, 254–263. [Google Scholar] [CrossRef]
- Kulan, EG.; Arpacıoğlu, A.; Ergin, N.; Kaya, MD. Evaluation of germination, emergence and physiological properties of sugar beet cultivars under salinity. Trakya University Journal of Natural Sciences. 2021, 22, 263–274. [Google Scholar] [CrossRef]
- Wichienchot, S.; Jatupornpipat, M.; Rastall, RA. Oligosaccharides of pitaya (dragon fruit) flesh and their prebiotic properties. Food chem. 2010, 120, 850–857. [Google Scholar] [CrossRef]
- Nizamlıoğlu, NM.; Ünver, A.; Kadakal, Ç. Mineral content of pitaya (Hylocereus polyrhizus and Hylocereus undatus) seeds grown in Turkey. Erwerbs-Obstbau. 2021, 63, 209–213. [Google Scholar] [CrossRef]
- Paśko, P.; Galanty, A.; Zagrodzki, P.; Ku, YG.; Luksirikul, P.; Weisz, M.; Gorinstein, S. Bioactivity and cytotoxicity of different species of pitaya fruits–A comparative study with advanced chemometric analysis. Food Bioscience. 2021, 40, 100888. [Google Scholar] [CrossRef]
- Attar, ŞH. ; Gündeşli, MA.; Urün, I.; Kafkas, S.; Kafkas, NE.; Ercisli, S.; Ge, C.; Mlcek, J.; Adamkova, A. Nutritional analysis of red-purple and white-fleshed pitaya (Hylocereus) species. Molecules. 2022, 27, 808. [Google Scholar]
- Rubio, JS.; García-Sánchez, F.; Rubio, F.; García, AL.; Martínez, V. The importance of K+ in ameliorating the negative effects of salt stress on the growth of pepper plants. European Journal of Horticultural Science. 2010, 75, 33–41. [Google Scholar]
- Guo, SH.; Niu, YJ.; Zhai, H.; Han, N.; Du, YP. Effects of alkaline stress on organic acid metabolism in roots of grape hybrid rootstocks. Scientia Horticulturae. 2018, 227, 255–260. [Google Scholar] [CrossRef]
- Çolak, AM.; Mertoğlu, K.; Alan, F.; Esatbeyoglu, T.; Bulduk, İ.; Akbel, E.; Kahramanoğlu, I. Screening of naturally grown european cranberrybush (Viburnum opulus L.) genotypes based on physico-chemical characteristics. Foods. 2022, 11, 1614. [Google Scholar] [CrossRef]
Figure 1.
Variation of germination stress tolerance index according to factors (LSD (%): Seed C: 0.51, Salt C: 0.69, Horm: 0.64, Seed CxSalt C: 0.98, Seed CxHorm: 0.96, Salt CxHorm: 1.20, Seed CxSalt CxHorm: 1.70, F Values (%): Seed C: 3223.38**, Salt C: 30224.38**, Horm: 521.50**, Seed CxSalt C: 1458.50**, Seed CxHorm: 193.87**, Salt CxHorm: 48.00**, Seed CxSalt CxHorm: 30.80**).
Figure 1.
Variation of germination stress tolerance index according to factors (LSD (%): Seed C: 0.51, Salt C: 0.69, Horm: 0.64, Seed CxSalt C: 0.98, Seed CxHorm: 0.96, Salt CxHorm: 1.20, Seed CxSalt CxHorm: 1.70, F Values (%): Seed C: 3223.38**, Salt C: 30224.38**, Horm: 521.50**, Seed CxSalt C: 1458.50**, Seed CxHorm: 193.87**, Salt CxHorm: 48.00**, Seed CxSalt CxHorm: 30.80**).
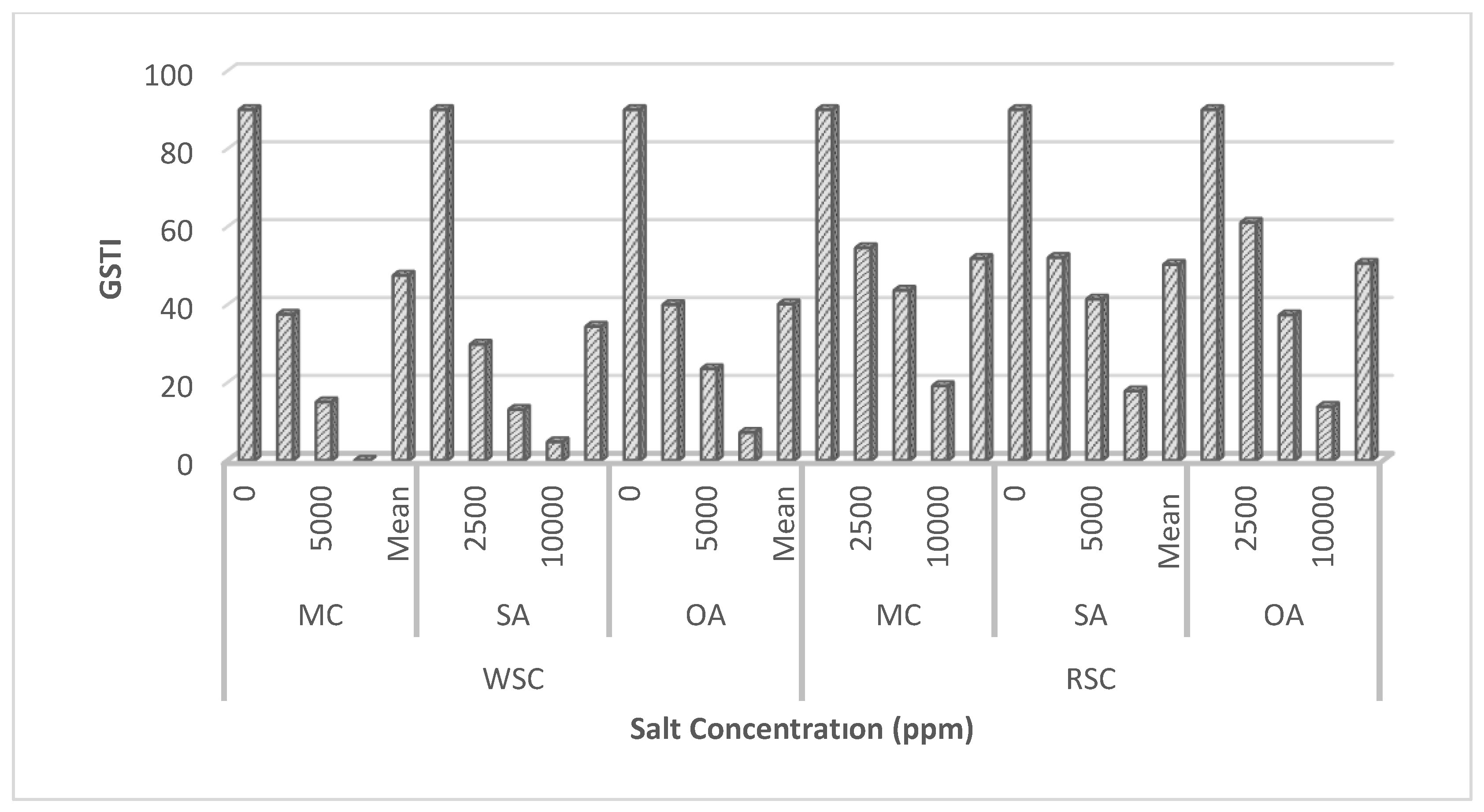
Figure 2.
Correlations among investigated characteristics.
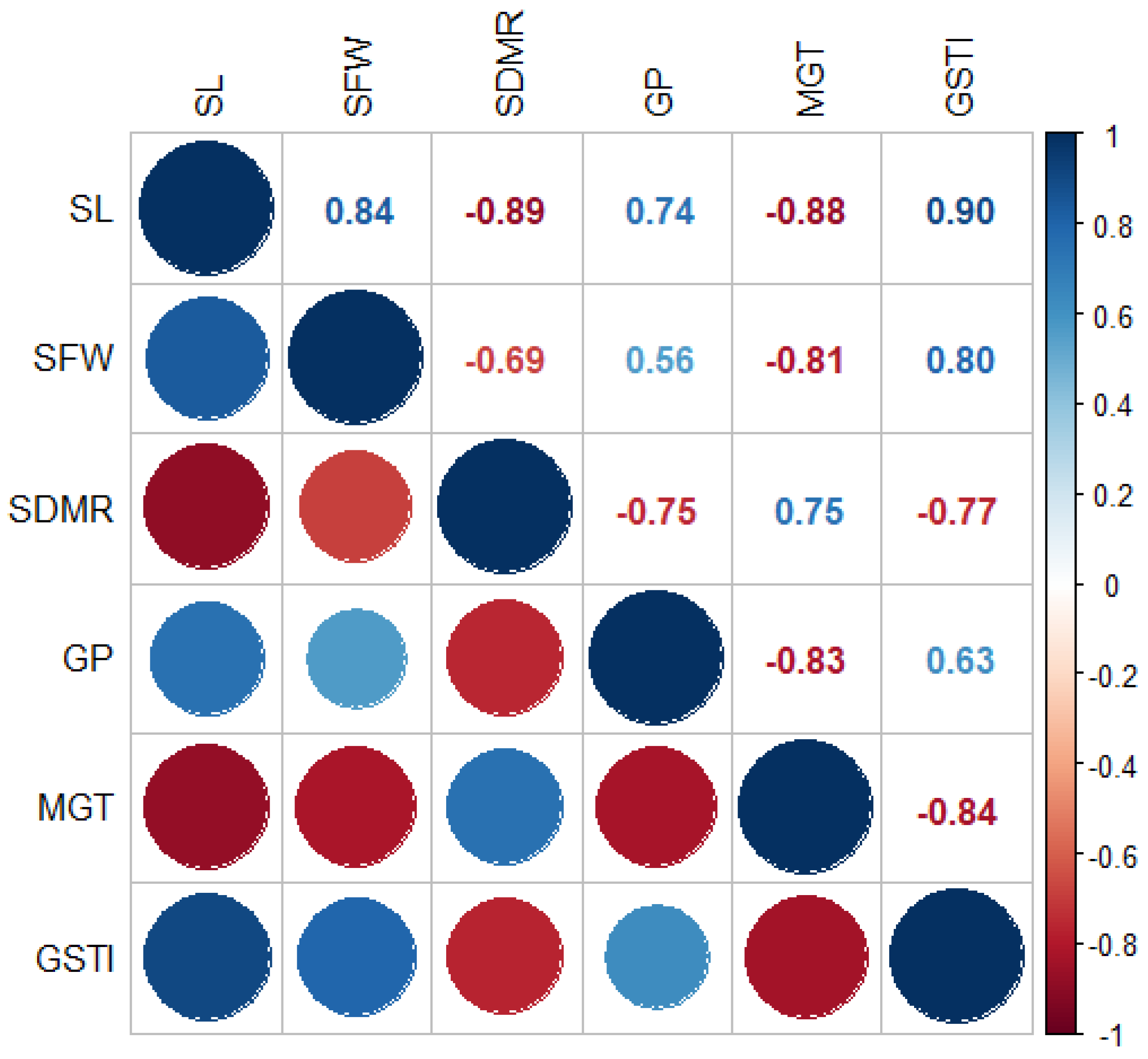

Table 1.
Distribution of germination and emergence performances according to factors.
| Flesh Color (FC) | Germination (%) | MGT (day) | Shoot length (cm) | Fresh seedling weight (mg) | Seedling dry matter ratio (%) | GSTI (%) |
|---|---|---|---|---|---|---|
| White | 81.17 b | 11.24 a | 27.90 b | 12.29 b | 4.44 b | 36.08 b |
| Red | 85.33 a | 7.33 b | 31.49 a | 21.23 a | 4.65 a | 54.91 a |
| Salt Concentrations (SC) | ||||||
| Control | 98.44 a | 4.74 d | 41.56 a | 22.85 a | 2.87 d | 100.0 a |
| 2500 ppm | 90.00 b | 7.06 c | 35.78 b | 20.76 b | 3.54 c | 51.32 b |
| 5000 ppm | 89.50 b | 9.80 b | 26.18 c | 14.64 c | 4.45 b | 25.97 c |
| 10 000 ppm | 55.06 c | 15.55 a | 15.26 d | 8.79 d | 7.33 a | 4.68 d |
| Plant Growth Regulators (PGRs) | ||||||
| MC | 83.54 ns | 9.16 b | 30.09 ns | 16.96 a | 4.46 ns | 46.05 b |
| SA | 82.21 | 9.62 a | 29.37 | 16.28 b | 4.58 | 43.22 c |
| OA | 84.00 | 9.08 b | 29.63 | 17.04 a | 4.60 | 47.21 a |
| ANOVA Significance levels | ||||||
| FC | ** | *** | *** | *** | * | *** |
| SC | *** | *** | *** | *** | *** | *** |
| PGR | ns | * | ns | ** | ns | *** |
| FC*SC | *** | *** | ns | *** | *** | ** |
| FC*PGR | ns | ns | ns | * | *** | *** |
| SC*PGR | * | ns | ns | ** | *** | *** |
| FC*SC*PGR | ns | ns | ns | ** | ** | *** |
*: Means statistical difference respectively at; 0.05, 0.01 and 0.001, ns: non-significant.
Table 2.
Variation of germination percentage and mean germination time according to factors.
| Germination percentage (%) | |||||
|---|---|---|---|---|---|
| Flesh Color | Salt Concentrations | Hormones | |||
| MC | SA | OA | Mean | ||
| White | 0 | 97.67 | 98.33 | 98.00 | 98.00A |
| 2500 | 94.67 | 89.33 | 93.33 | 92.44A | |
| 5000 | 94.67 | 92.33 | 93.33 | 93.44A | |
| 10000 | 39.33 | 34.27 | 48.67 | 40.76B | |
| Mean | 81.58 | 78.57 | 83.33 | 81.16B | |
| Red | 0 | 98.67 | 99.33 | 98.67 | 98.89A |
| 2500 | 83.33 | 88.67 | 90.67 | 87.56B | |
| 5000 | 88.00 | 89.33 | 79.33 | 85.56B | |
| 10000 | 72.00 | 66.00 | 69.92 | 69.31C | |
| Mean | 85.50 | 85.83 | 84.65 | 85.33A | |
| Means of Flesh Colors | 0 | 98.17A | 98.83A | 98.33A | 98.44A |
| 2500 | 89.00A | 89.00A | 92.00A | 90.00B | |
| 5000 | 91.33A | 90.83A | 86.33A | 89.50B | |
| 10000 | 55.67AB | 50.14B | 59.29A | 55.03C | |
| Mean | 83.54 | 82.20 | 83.99 | 83.24 | |
| Mean germination time (day) | |||||
| Flesh Color | Salt Concentrations | Hormones | |||
| MC | SA | OA | Mean | ||
| White | 0 | 5.31 | 5.25 | 5.30 | 5.29D |
| 2500 | 8.20 | 8.79 | 7.958 | 8.35C | |
| 5000 | 11.78 | 12.32 | 11.90 | 12.00B | |
| 10000 | 19.51 | 20.10 | 18.39 | 19.33A | |
| Mean | 11.22 | 11.61 | 10.89 | 11.24A | |
| Red | 0 | 4.20 | 4.25 | 4.15 | 4.20 D |
| 2500 | 5.73 | 6.11 | 5.46 | 5.77 C | |
| 5000 | 7.18 | 7.54 | 8.06 | 7.59 B | |
| 10000 | 11.28 | 12.56 | 11.46 | 11.77A | |
| Mean | 7.10 | 7.62 | 7.28 | 7.33B | |
| Means of Flesh Colors | 0 | 4.75 | 4.75 | 4.72 | 4.74D |
| 2500 | 7.02 | 7.45 | 6.71 | 7.06C | |
| 5000 | 9.48 | 9.93 | 9.98 | 9.78B | |
| 10000 | 15.40B | 16.33A | 14.92B | 15.55A | |
| Mean | 9.16 | 9.62 | 9.08 | 9.29 | |
Table 3.
Variation of shoot length and fresh seedling weight and seedling dry matter ratio according to factors.
Table 3.
Variation of shoot length and fresh seedling weight and seedling dry matter ratio according to factors.
| Shoot length (mm) | |||||
|---|---|---|---|---|---|
| Flesh Color | Salt Concentrations | Plant Growth Regulator | |||
| MC | SA | OA | Mean | ||
| White | 0 | 39.83A | 38.83A | 41.00A | 39.89A |
| 2500 | 33.00A | 31.92A | 33.92A | 32.94B | |
| 5000 | 25.00A | 25.17A | 24.00A | 24.72C | |
| 10000 | 14.67A | 13.33A | 14.17A | 14.06D | |
| Mean | 28.12 | 27.31 | 28.27 | 27.90B | |
| Red | 0 | 43.67A | 43.83A | 42.17A | 43.22A |
| 2500 | 39.75A | 39.75A | 36.33B | 38.61B | |
| 5000 | 29.08A | 25.50B | 28.33AB | 27.64C | |
| 10000 | 15.75A | 16.58A | 17.08A | 16.47D | |
| Mean | 32.06 | 31.42 | 30.98 | 31.48A | |
| Means of Flesh Colors | 0 | 41.75 | 41.33 | 41.58 | 41.56A |
| 2500 | 36.37 | 35.83 | 35.12 | 35.78B | |
| 5000 | 27.04 | 25.33 | 26.17 | 26.18C | |
| 10000 | 15.21 | 14.96 | 15.62 | 15.26D | |
| Mean | 30.09 | 29.36 | 29.62 | 29.69 | |
| Fresh seedling weight (mg) | |||||
| Flesh Color | Salt Concentrations | Plant Growth Regulator | |||
| MC | SA | OA | Mean | ||
| White | 0 | 16.89A | 16.50A | 17.06A | 16.82A |
| 2500 | 14.71A | 14.18A | 14.48A | 14.45B | |
| 5000 | 11.39A | 11.97A | 10.51A | 10.96C | |
| 10000 | 7.07A | 6.71A | 6.97A | 6.92D | |
| Mean | 12.51A | 12.09A | 12.25A | 12.29B | |
| Red | 0 | 28.94A | 29.03A | 28.69A | 28.89A |
| 2500 | 28.28A | 25.58B | 27.36A | 27.07B | |
| 5000 | 18.89A | 16.14B | 19.92A | 18.32C | |
| 10000 | 9.53B | 11.11AB | 11.31A | 10.65D | |
| Mean | 21.41A | 20.46B | 21.82A | 21.23A | |
| Means of Flesh Colors | 0 | 22.92A | 22.76A | 22.87A | 22.85A |
| 2500 | 21.50A | 19.88B | 20.92A | 20.76B | |
| 5000 | 15.14A | 13.56B | 15.21A | 14.64C | |
| 10000 | 8.30A | 8.91A | 9.14A | 8.78D | |
| Mean | 16.96A | 16.28B | 17.04A | 16.76 | |
| Seedling dry matter ratio (%) | |||||
| Flesh Color | Salt Concentrations | Plant Growth Regulator | |||
| MC | SA | OA | Mean | ||
| White | 0 | 2.99A | 3.01A | 2.97A | 2.99D |
| 2500 | 3.46A | 3.66A | 3.78A | 3.64C | |
| 5000 | 4.51A | 4.39A | 3.90B | 4.27B | |
| 10000 | 7.06A | 7.37A | 6.19B | 6.88A | |
| Mean | 4.51A | 4.61A | 4.21B | 4.44B | |
| Red | 0 | 2.76A | 2.77A | 2.69A | 2.74D |
| 2500 | 3.10B | 3.48AB | 3.76A | 3.45C | |
| 5000 | 4.90A | 4.54A | 4.46A | 4.63B | |
| 10000 | 8.00A | 7.43B | 7.92AB | 7.78A | |
| Mean | 4.69A | 4.56A | 4.71A | 4.65A | |
| Means of Flesh Colors | 0 | 2.88A | 2.89A | 2.83A | 2.87D |
| 2500 | 3.28B | 3.57AB | 3.77A | 3.54C | |
| 5000 | 4.71A | 4.47AB | 4.18B | 4.45B | |
| 10000 | 7.53A | 7.40A | 7.06B | 7.34A | |
| Mean | 4.46 | 4.58 | 4.60 | 4.55 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



