Submitted:
23 June 2023
Posted:
26 June 2023
You are already at the latest version
Abstract
Topoisomerases, common targets for anti-tumor therapeutics, are crucial enzymes for DNA replication, transcription, and many other aspects of DNA metabolism. The potential anti-cancer effect of thiosemicarbazones (TSC) and metal-TSC complexes has been demonstrated by targeting several biological processes, including DNA metabolism. Human topoisomerases were discovered among the molecular targets for TSCs, and metal-chelated TSCs specifically displayed significant inhibition. The processes by which metal-TSCs or TSCs inhibit topoisomerases are still being studied. In this brief review, we summarize the TSCs and metal-TSCs that inhibit various types of human topoisomerases.
Keywords:
topoisomerase
; thiosemicarbazone
; cancer
; DNA
1. Introduction
Topoisomerases (Top) are essential enzymes for genome stability that are involved DNA metabolism through the maintenance of DNA topology. All cells maintain DNA supercoiling in a dynamic process that is required for transcription, replication, and cell division to take place. Top are found across all domains of life and several viruses encode topoisomerases. Top are divided into two families: Type I and Type II.
Type I Top cuts and religates one strand of the double helix coupled with either a strand passage or controlled rotation mechanism to remove DNA positive (overwinding) and negative (underwinding) supercoiling. To modulate supercoiling, remove DNA knots, and unlink catenated DNA (intertwined DNA), Type II Top forms a double-stranded DNA break and passes a double-strand DNA section through the break before ligating the DNA back together. In both mechanisms, the DNA break is stabilized through a covalent intermediate with an active site tyrosine residue on the Top. There are six human Top including Top1B, mitochondrial Top1B, Top2α, Top2β, Top3α, Top3β [1].
There are two main Type I subfamilies IA (Top3α/β in humans) and IB (Top1B/mitochondrial Top1B in humans) [1,2]. The subfamilies differ both structurally and mechanistically, and these differences have been reviewed elsewhere [1,2]. There is a Type IC that is only found in an archaeal species.
There are two main Type II subfamilies, which include Type IIA and Type IIB depending on either a 4- or 2-base stagger between the cleaved positions on the DNA, respectively [1,3,4,5]. Human Top2α and Top2β both belong to the Type IIA subfamily [1,5]. While the names and structures for the archaeal and bacterial counterparts differ, this review will focus on the mammalian forms of the eukaryotic enzymes as named above.
During the catalytic cycle, Top form temporary single- or double-stranded DNA breaks [1,5]. These momentary DNA breaks may become permanent, leading to DNA damage and cell death [1,2,4]. Several widely-used anti-cancer therapeutics target either Type I or Type II Top, and in a similar manner, several antibacterial agents are used to target bacterial Top to fight off infections. Interfacial poisons and catalytic inhibitors are the two primary groups of inhibitors for Top enzymes [2,4,6]. Whereas catalytic inhibitors prevent Top from completing its catalytic cycle, interfacial poisons stabilize single-stranded or double-stranded breaks leading to further damage [2,4,6]. The general mechanism for catalytic inhibitors is interaction with the N-terminal ATPase domain, which is a clamp-like region at the “top” of the enzyme. Top poisons are thought to act by a “doorstop” mechanism where the drug molecule prevents ligation by slipping between the cleaved ends of the DNA during catalysis. In general, poisons tend to lead to accumulation of cleaved DNA while catalytic inhibitors do not. In addition, some agents are reactive and lead to inactivation of the enzyme and/or covalent adduction, which could occur either in the active site or at or near the ATPase domain [6,7].
Thiosemicarbazones (TSC) are a varied class of compounds with common N-N-S coordinates. They were discovered in the 1950s with anti-bacterial, anti-fungal, and anti-tumor activities [8,9]. TSCs are multi-target drugs and the molecular mechanisms involved metal chelation, DNA interference, topoisomerase inhibition and ribonucleotide reductase inhibition [10]. Many metal-TSC complexes have been synthesized and were found to be more effective in cell toxicity experiments than the ligand form of the TSC [10,11]. The cell toxicity of metal-TSCs included a decrease in de novo purine synthesis and inhibition of IMP dehydrogenase, DNA polymerase activity, and topoisomerase II activity [10,11,12]. Recent studies in yeast have uncovered other cellular mechanisms for metal-bis(TSC) which includes chromatin remodeling, cytoskeleton organization, mitochondrial function, and iron metabolism [13]. Due to the varied structures of TSCs, they impact multiple cellular targets. In this paper, we summarize the TSCs that are targeting Top with a particular emphasis on metal-TSCs.
2. Thiosemicarbazones as Inhibitors of Topoisomerases
2.1. TSC Ligand or Bis(TSC) Ligand Inhibition of Topoisomerases
TSCs have been studied for decades for antiviral, antifungal and antiproliferation activity. In recent years, Triapine (3-AP, 3-aminopyridine-2carboxaldehyde thiosemicarbazone), a member of TSC family, has been studied in clinical trial phase I and II [14,15,16,17]. The major molecular target of Triapine was identified as ribonucleotide reductase [17,18]. Triapine did show some inhibition of Top2A but only in the presence of Cu(II) [19]. Previous work on Triapine did not show any inhibition of relaxation or poisoning of plasmid DNA cleavage with Top2α. [20]
In a series of TSC compounds, Top2 was discovered as one of the molecular targets for TSC24 [21]. The pattern of TSC antiproliferative effects against the human cancer cell line (NCI-60) is similar to those found with recognized anticancer treatments [21]. These investigations revealed TSC24’s profile is comparable to known Top2 agents. TSC24 was further investigated and found that it hindered DNA relaxing and decatenation by inhibiting the Top2α ATPase domain [21].
Another TSC ligand that has been well studied was Dp44mT. In relaxation and cleavage experiments, Dp44mT was shown to preferentially target Top2α, with minimal effect on Top2β and little inhibition of Top1 [22]. But this result is controversial as an additional study found that the ligand Dp44mT did not inhibit Top2α or increase cellular cleavage complexes [20]. A more recent study on the ligand Dp44mT also found little effect of the compound alone, but they show inhibition when combined with Cu(II) [19]. It is unclear whether it was the presence of Cu(II) alone or if the Cu(II) formed a complex with the ligand leading to the effect in this case. As is discussed below, it is possible that the ligand forms alone show little activity, while metal-chelated forms have varying levels of activity depending on the metal ion.
Figure 1.
Structures of selected TSC compounds.
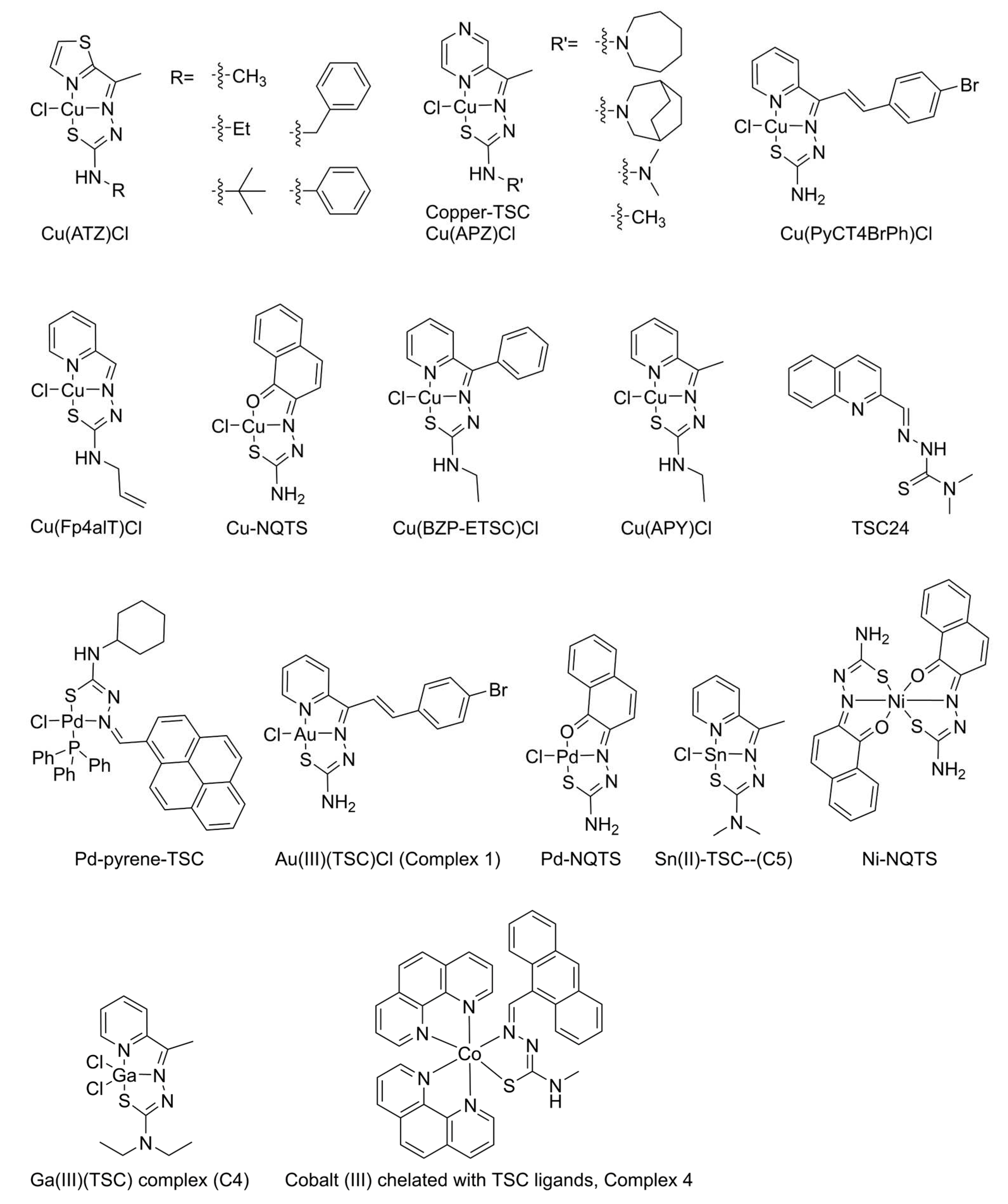
Computational docking and surface plasmon resonance studies support the ability of TSC24 to bind near the ATP binding pocket, but it is unclear if this is generalizable to other TSCs and whether this has been biochemically validated. Both TSC24 and Dp44mT appear to act as catalytic inhibitors, and there is an increase in DNA cleavage seen with Dp44mT [21,22]. TSC24 does not appear to increase strand breaks in cells, nor block the effects of VP16 [21].
A series of thiosemicarbazones and 4-thiazolindinones have been synthesized and some were identified with activity against Top2. The thiosemicarbazone (E)-2-(1H-indol-3ylmethylene)-N-(naphthalen-1-yl) hydrazinecarbothioamide (compound 2b) was shown to disrupt cell growth of HT29 cells and appear to weakly inhibit plasmid DNA relaxation by Top2α [23]. Compound 3a [(Z)-2-(Acridin-9-ylmethylene)-N-phenyl-hydrazinecarbothioamide)] and (compound 3h) [(Z)-2-(Acridin-9-ylmethylene)-N-(naphtalen-1-yl) hydrazine-carbothioamide] also had limited ability to inhibit relaxation [24]. Detailed studies on the structure-activity relationship of TSC against Top2α showed that most of TSC ligands inhibit Top2α very weakly or not at all [25,26,27,28].
Despite the fact that TSC ligands were initially found to target Top2, the majority of the ligands produced had little or no effect on Top1 [29] or Top2 [26,27,28,30,31]. Metal-TSCs, on the other hand, showed profound inhibition on Top, as discussed below.

Table 1.
TSC ligand inhibition on Tops.
| Name | Inhibition of Top | Reference |
|---|---|---|
| TSC24 | Inhibit Top2α DNA relaxation at 25 µM Inhibit Top2 at decatenation at 100 µM |
[21] |
| Dp44mT | Inhibit human Top2α in relaxation assay with 5’ labeled 161-bp fragment from pBluescript SK phagemid DNA Do not inhibit human Top2β or human TopoI |
[22] |
| Dp44mT | Do not inhibit Top2α in decatenation Do not increase cleavage complex by human Top2α |
[20] |
| Triapine | ||
| Compound 2b | Inhibit human Top2α weakly at 100 µM | [23] |
| Compound 3a | Inhibit human Top2α-mediated DNA relaxation at 100 µM | [24] |
| Compound 3h | ||
| Triapine | Do not inhibit Top2α-mediated DNA relaxation at 50 µM | [19] |
| Dp44mT | ||
| Compound 24 | Do not inhibit isolated Top2 from L1210 | [12] |
| Compound 36 | ||
| NQTS | Do not inhibit Top2α | [30] |
| HFp4mT, HFp4pyrrT |
Inhibit Top2α at 100 µM | [31] |
| HFp4eT, HFp4ipT, HFp4alT, HAp4mT, HAp4-eT, HFp4bzT, HFpT, HAp4alT, and HApz4mT. |
Do not inhibit Top2α at 100 µM | [31] |
| BZP-TSC ligands series | Inhibits Top2α slightly at 50 µM. | [14] |
| ATZ ligand series | Do not inhibit Top2α at 10µM, do not increase cleavage complex | [27] |
| BZP ligands series | ||
| APY | Do not inhibit Top2α at 100 µM | [26] |
| APY | Do not inhibit Top2β at 200 µM | [28] |
| APZ | Inhibit Top2 α at 100µM | [26] |
| BZP | Inhibit Top2β at 200µM weakly | [28] |
| HPyCT4BrPh | Do not inhibit human Top1B at 50 µM | [29] |
2.2. Metal-TSC or Metal-Bis(TSC) Inhibition of Topoisomerases
TSCs can chelate with various metal ions. Metal-TSC complexes have been produced from copper (Cu), nickel (Ni), palladium (Pd), ruthenium (Ru), tin (Sn), gallium (Ga), gold (Au), and cobalt (Co) [25,30,32,33,34,35,36]. Metal-TSCs inhibit Top more strongly than their TSC ligand [19,25,26,27,28,31]. As will be discussed more below, metal-TSCs appear to be active against both Type I and Type II Top.
2.2.1. Inhibition of Type I Top
dHuman Top1B belongs to the Type I Top family. It relaxes the DNA supercoil during DNA replication, recombination and transcription by cutting one strand of DNA [1,2]. Top1B inhibitors are a class of compounds that target the enzyme and preventing from relaxing DNA and leading to the accumulation of DNA damages. These inhibitors have potential as antitumor agents since tumor cells are under fast proliferation and DNA replication, making them more susceptible to DNA damage. Several classes of Top1B inhibitors have been developed, including camptothecin analogs and indolocarbazoles [1,2,37]. There are a couple of metal-TSC compounds that have been studied against Top1B activity.
Table 2.
Metal-TSC inhibition of TopI.
| Name | Inhibition of TopI | Reference |
|---|---|---|
| Cu(PyCT4BrPh)Cl | Inhibit TopI The inhibition is severe when pre-incubation of the compound to TopI Inhibited the cleavage step and partially inhibited religation. |
[29] |
| Pd-pyrene-TSC | Inhibited human Top1B at 12.5 µM | [38] |
| Ga(III)-TSC complex (C4) | Inhibited TopI | [36] |
| Au(III)(TSC)Cl (complex 1) | Inhibited human Top1B activity starting at 1.5 μM Pre-incubation of Top1B with Complex 1 increased the inhibition |
[25] |
| Ni-bis(TSC) | No inhibition of E. coli TopI | [39] |
| Nine copper complexes | Inhibited TopI | [40] |
| Cobalt (III)-TSC (Complex 4) | Inhibited TopI-induced DNA relaxation | [35] |
Cu(PyCT4BrPh)Cl [Cu(3-(4-bromophenyl)-1-pyridin-2-ylprop-2-en-1-one-thiosemi-carbazone)Cl] was studied against human Top1B [29]. It inhibited Topo1B by partially blocking ligation of the cleaved DNA [29]. The complex also reduced enzyme-DNA binding according to an EMSA assay [29]. But additional studies would be needed to clarify the exact mechanism.
In another study, pyrene TSCs were complexed with Pd (Complex 1) and examined for inhibition of human Top1B [38]. Pd-pyrene-TSC complexes inhibited relaxation of supercoiled plasmid by human Top1B at 12.5 µM [38]. Additionally, the Pd pyrene-TSC complex displayed the ability to inhibit ligation of cleaved DNA with Top1B, similar to Cu(PyCT4BrPh)Cl [38].
One group reported the use of a Ga(III)-TSC complex, [N,N-diethyl-2-[1-(2-pyridinyl) ethylidene]hydrazinecarbothioamide-N,N,S-gallium(III)]bis(chloride), referred to as C4 in the study [36]. Based upon their results, human Top1B cleavage activity was inhibited by the Ga(III)-TSC complex while the ligand alone did not show significant inhibition [36].
The Au(III) TSC complex [(3-(4-bromophenyl)-1-pyridin-2-ylprop-2-en-1-enone thiosemicarbazonato)chlorogold(III)] chloride, [Au(PyCT4BrPh)Cl]Cl, was studied with human Top1B and found to inhibit relaxation at 1.5 μM [25]. In contrast, HAuCl4·3H2O did not inhibit until 200 µM. Pre-incubation of Top1B with this compound increased the inhibition, which suggests Gold(III)-TSC binds and inhibits the activity of Topo1B [25].
In another study, Ni chelated with testosterone TSC to form a distorted square planar with ligand as a bidentate NS donor---Ni-bisTSC [39]. Ni-bis(TSC) did not inhibit E. coli TopI, but it showed DNA binding affinity similar to ethidium bromide [39].
In summary, the research on metal-TSCs inhibiting Top1B is limited. Some compounds displayed catalytic inhibition, such as [Au(PyCT4BrPh)Cl]Cl, others are interfacial poisons by inhibiting ligation, including Cu(PyCT4BrPh)Cl, Pd-Pyrene-TSC, and Ga(III)-TSC.
2.2.2. Inhibition of Type II Top
Type II Top is the primary target for studies of TSCs antitumor activity. Multiple metal-TSC complexes showed higher inhibition compared with their ligand counterpart. Cu-TSCs are the most studied and have demonstrated the highest inhibition of Top2.
- Ni chelated with bis(TSC)
Ni-TSCs were discovered to block a variety of metabolic pathways, including purine synthesis, DNA polymerase, PRPP-amino transferase, IMP dehydrogenase, dihydrofolate reductase, TMP-kinase, and thymidylate synthetase activities, against the L1210 cell line in 1997 [32]. Despite the fact that Ni-TSCs demonstrated several cellular pathways for inhibition, research suggests that Ni-TSCs do not efficiently inhibit Top enzymes [13,32]. Ni(II) coordinates with two TSC ligands, Ni-bis(TSC), which lack the essential square planar structure for Topo inhibition [26,32].
There are some controversies on the inhibition of Ni-TSCs against Top2. Ni-NQTS showed inhibition of Top2α-mediated DNA relaxation assay using a TopoGEN kit [30]. However, the data are inconsistent with other reported results. For example, a yeast screen did not show Ni-bisTSC interferes with Top [13]. Our unpublished results showed that when bisTSC chelate with metal ions, the metal-bisTSC compounds do not inhibit Top2α (Beckett and Jiang, unpublished). Ni-NQTS were also tested in DNA cleavage assays with Top2α [30]. The results seem to show that Ni-NQTS does not stabilize double-stranded DNA cleavage, but there was a low amount of nicking observed, though it was not quantified [30]. In another study, several Ni-TSC complexes were examined alongside Cu analogs discussed below [41]. Interestingly, Ni(L1)(HL1)Cl, Ni(HL2)2Cl2, Ni(L3)2, Ni(L4)2, and Ni(L5)2Cl2 did not appreciably inhibit Top2α from TopoGEN [41]. Although Ni-TSCs performed profound inhibition against cell proliferation, Top2 may not be the target (or the primary target) for Ni-TSCs [8,9,10].
- 2.
- Cu chelated TSCs
In cell toxicity studies, copper (Cu2+) chelated TSCs are one of the most active groups of metal-TSCs [8,10]. When Copper chelates with TSC, it forms a square planar structure with Cu in the middle, which seems to be the crucial structure element for Top2 inhibition [26,27,28], Cu(TSC)s demonstrated greater inhibition compared with their ligands. For example, Cu(TSC)Cl (Compound 1 and 2) inhibited Top2 while the corresponding TSC ligands (Compound 24 and 36) did not [12]. In general, Cu(TSC) complexes act on Top2 as catalytic inhibitors through inhibiting the ATPase function and inhibiting relaxation.
Another study showed that Cu(TSC)Cl complexes (Compounds 1-3) reduced DNA cleavage observed in the presence of etoposide, and these compounds alone did not show any stabilization of cleavage complexes [42]. Cu-NQTS inhibited Top2α-mediated DNA relaxation assays [30].
Cu(Fp4alT)Cl completely inhibits Top2α without promoting the formation of linear DNA products [31]. Similar results were observed with the other Cu(TSC)Cl complexes in the study [31]. Thus, Cu(Fp4alT)Cl and its family of Cu(TSC)Cl complexes are catalytic inhibitors of Top2α rather than poisons of the enzyme [31]. Cu(L1)Cl, Cu(L2)Cl, Cu(L3)Cl, Cu(L4)Cl, and Cu(L5)Cl2 all showed inhibition of Top2α from TopoGEN [41]. Cu(TSC) cation (Complex 1) increase DNA cleavage complex and inhibited DNA relaxation [43].
In another study, the complexes [Cu(S,R)-L] and [Cu(R,S)-L] showed inhibition of Top2α relaxation at 300 µM [44]. However, the concentration of inhibition is similar to ligand TSC and much higher (10-100+-fold) than other Cu(TSC)Cl [44].
Our collaboration works on a series of Cu(TSC)Cl complexes that demonstrated their inhibition of both human Top2α and Top2β [26,27,28]. The structure-activity relationship of metal-TSC showed that Cu(II) played a predominant role in inhibition of Top2 [26,27,28]. The mechanism of Cu(TSC)Cl inhibition on Top2 is complicated. Cu(TSC)Cl inhibited ATP hydrolysis and plasmid DNA relaxation by Top2α and Top2β, which is consistent with these compounds acting as catalytic inhibitors. However, unlike other catalytic inhibitors, Cu(TSC)Cl complexes stabilize the DNA cleavage complexes and increase levels of DNA cleavage, which is the characteristic of interfacial poisons [26,27,28]. In addition, the complexes we tested lead to higher levels of double-stranded breaks implying an increase in coordination between the two active sites [26]. The increase in DNA cleavage was not seen in a mutant lacking the ATPase domain [26]. Further, incubation of Cu(TSC)Cl complexes with Top2α or Top2β prior to DNA leads to a progressive inactivation of the enzyme [26,28]. Consistent with this data is the observation that the Cu(TSC)Cl stabilize a closed N-terminal region (ATPase domain) of Top2α or Top2β [28]. The significance of this particular aspect is that the ATPase domains of each half of the homodimer close around DNA in the presence of ATP. Our results demonstrate that the Cu(TSC)Cl complexes that were studied were able to induce closure of this N-terminal gate in a way that stabilized the gate similar to what is seen with a non-hydrolyzable ATP analog (AMP-PNP) [28].
Although Top2α has been widely used as the molecular target to study Cu(TSC)Cl inhibition, our research found that Cu(TSC)Cl complexes inhibited ATPase and relaxation activity of both Top2α and Top2β [28]. Taken together, the data support the idea that these Cu(TSC)Cl complexes act on or near the ATPase domain, which is highly similar between both isoforms. Using N-terminally and C-terminally truncated versions of Top2α or Top2β both resulted in a lack of increased DNA cleavage [26,28]. Interestingly, some Cu(TSC)Cl showed inhibition both of Top1 and Top2, as will be discussed below [40].
Other metal-TSC also showed inhibition of Top2. Pd-NQTS inhibited Top2α-mediated DNA relaxation assay [30]. When chelated with the same ligand, Pd(TSC)Cl seemed to be less active compared with its Copper counterpart [27]. Ru(TSC)Cl {[(η-6-p-cymene)Ru(EtATSC)Cl]+ cation}, with a big substrate ring structure inhibited human Top2α in relaxation assay [33]. Ruthenium complex of TSC has been tested in a Top2α-mediated DNA relaxation assay and found to inhibit relaxation [33]. Sn(II) chelated TSC complex (C5) inhibited Top2 at 20 µM [34].
Table 3.
Metal-TSC inhibition of Top2.
| Name | Inhibition of Top2 | Reference |
|---|---|---|
| Nine compounds and their copper complexes | Inhibited human Top2α | [40] |
| Cobalt (III) chelated with TSC ligands. Complex 4 | Inhibited human Topo2α-induced DNA relaxation | [35] |
| Ni-bis(TSC) complex 1 | Did not inhibit isolated Top2 from L1210 cells at 100 µM | [32] |
| Cu-TSCs (Compound 1 and 2) | Inhibited isolated Top2 from L1210 cells | [12] |
| Copper TSC | Inhibited Top2 of L1210 cells with IC50 value of 6.25-12.2 µM. antagonize the DNA break affect by etoposide. |
[42] |
| Ni-NQTS | Inhibited DNA relaxation | [30] |
| Cu-NQTS | Inhibited DNA relaxation (TOPOGEN kit) | [30] |
| Pd-NQTS | ||
| Cu(Fp4alT)Cl and its family of Cu(TSC)Cl |
Inhibited relaxation by Top2 at 10 µM Completely inhibits Top2α without promoting the formation of linear DNA products |
[31] |
| Ni-bis(TSC) | No effect in stabilizing DNA breaks | [45] |
| Cu(TSC) | Stabilizes DNA breaks | [45] |
| Ni(L1)(H1)Cl, Ni(HL2)2Cl2, Ni(L3)2,Ni(L4)2 Ni(L5)2Cl2 |
No inhibition of Top2 (TopoGEN) | [41] |
| Cu(L1)Cl, Cu(L2)Cl, Cu(L3)Cl, Cu(L4)Cl, Cu(L5)Cl2 | Inhibited Top2 (TopoGEN) | [41] |
| complex 1 (CuTSC cation) | inhibited DNA relaxation and increase DNA cleavage | [43] |
| Sn(II)-TSC—(C5) | Inhibited Topo2α at 20 µM. | [34] |
| Cu(S,R)L and Cu(R,S)L | Inhibited Top2α relaxation at 300 µM | [44] |
| Cu(APY)Cl] | Inhibited Top2α from 0.5 µM, Increased DNA cleavage Inhibited Top2α ATP hydrolysis No inhibition of ligation by Top2α Pre-incubate compounds with Top2α inactivate the enzyme |
[26] |
| Cu(APZ)Cl] | ||
| [Cu(APY)Cl] | Inhibited Top2β at 5 µM, Increased DNA cleavage by Top2β, Inhibited Top2β ATP hydrolysis, Inhibited ligation by Top2β Pre-incubate compounds with Top2β inactivate the enzyme Stabilized closure of N-terminal Top2α and Top2β clamp |
[28] |
| [Cu(BZP)Cl] | ||
| Cu(BZP)Cl series | Inhibited Top2α relaxation and increased DNA cleavage |
[27] |
| Cu(ATZ)Cl series | ||
| Pd(BZP)Cl series | Inhibited Top2α relaxation and increased DNA cleavage | [27] |
2.2.3. Inhibition of Type I and Type II Top
A few studies have compared the inhibition on Top1B and Top2α. Nine compounds and their copper complex were investigated against human Top1B and Top2α from TopoGEN [40]. Relaxation assays were quantified to generate an IC50. The Cu-TSC complexes were at least 10-fold more effective than the ligand alone [40]. They displayed greater inhibition of Top2α than Top1B [40]. Interestingly, the larger side chain substitutions generally displayed better inhibition of Top2α [40].
Complex 4 of Co(III)-TSC complexes inhibited Top1B-induced and Top2α-induced DNA relaxation, but the ligand nor its precursor were not able to inhibit either enzyme [35]. Complex 4 did not cause significant increase in DNA complexes with Top1B or Top2α, which suggests that Complex 4 is a catalytic inhibitor not a poison [35].
3. Discussion
TSCs are a broad group of compounds and thus many diverse TSCs have been synthesized and examined. While TSCs display a broad range of possible mechanisms of action, direct studies on purified Top enzymes have been very helpful in identifying the mechanism of inhibition. Cu-TSCs have been studied extensively against Top2α. The mechanism showed that Cu-TSCs are catalytic inhibitors that also exhibit some features of interfacial poisons (namely the increase in DNA cleavage levels). Some metal-TSCs inhibited DNA religation by Top1B, but religation of DNA by Top2 is not consistently inhibited by Cu-TSCs. The cleavage complexes created by Top2 are stabilized by the metal-TSC complexes may or may not involve inhibition of religation [26,27,28]. Further, Top2 enzymes generally are more impacted by metal-TSC compounds rather than the ligands alone. There remains a lack of a clear understanding on exactly where and how these compounds are acting. For example, biochemical evidence supports that these compounds can inhibit ATP hydrolysis, but some also increase DNA cleavage. These two mechanisms seem to contradict one another since the general thought is that catalytic inhibitors affect ATP hydrolysis but do not disturb DNA cleavage. Interestingly, these compounds do appear to stabilize the N-terminal clamp of Top2α and Top2β, which may help clarify the increase in DNA cleavage [28]. For instance, 1,4-benzoquinone is known to increase DNA cleavage and stabilize the N-terminal clamp likely through a covalent adduction mechanism [28,46,47,48]. Several computational studies indicated that TSC complexes can bind in or around the ATPase domain, which may be similar to the mechanism observed with 1,4-benzoquinone. Again, these data are consistent with the observation that N-terminally and C-terminally truncated mutants of Top2α or Top2β are not affected by metal-TSC complexes [26,28]. Together, these data suggest that there may be an increase in coordination between the two active sites of Top2 when metal-TSCs are present, which could result in increased DNA cleavage without true poisoning.
Although there are many molecular modeling studies to predict where TSC binds on Top, currently no NMR or crystal structure information is available on the exact location of where TSC or metal-TSC binds on Top. Further structural studies are urgently needed to elucidate the molecular structural information of metal-TSC inhibition on Top. The mode of interaction of the Cu(TSC)Cl complexes with Top may be inferred from the literature of TSCs interacting with other molecules. It has been well established that Cu(TSC)+ complexes have been easily synthesized and crystal structures have been done that demonstrate that the complexes can pick up water as a ligand and form 5-coordinate complexes [44,45]. Many of the Cu(TSC)Cl complexes are often found in the solid state as dimers, [Cu(TSC)Cl]2, which shows a weak bond, dissociable in solution, at the Cu center making it 5-coordinate. This structure then forms the square planar 4-cooordinate complex Cu(TSC)Cl in solution. Also, Cu(TSC)Cl complexes can dissociate the chloride ion (Cl-) in aqueous solution to replace Cl- with other ligands and water [46]. These observations indicate that Cu(TSC)Cl complexes can shed Cl- and bond directly with Top. The observation that they can become 5-coordinate indicates that the complex possibly forms two bonds with Top. This may suggest why structurally similar square planar Pd(TSC)Cl and Pt(TSC)Cl complexes may lose the Cl- ligand to bind to Top, but they cannot form 5-coordinate structures, and thus, are not as potent inhibitors of Top as the Cu(TSC)Cl complexes.
The ability of metal-TSCs to inhibit Top2α and Top2β suggests that these compounds may have therapeutic potential. However, there are additional considerations that must be addressed before these compounds can be used clinically. First, it is unclear whether inhibition of cell growth can be attributed to Top2 or to other possible mechanisms, known or unknown. Given that the ATPase domain of Top2 is similar to some other enzymes (GHKL ATPase/kinase superfamily), it is possible that metal-TSCs may impact other enzymes as well [49]. Second, the reactivity of these compounds must be explored to determine whether these compounds can covalently interact with proteins and the consequences of this must be considered. Previous data indicates that Top2 can be inactivated through incubation with Cu(TSC)Cl complexes, and the mechanism of this needs to be explored to determine whether this is a specific action against Top2 or could occur more generally leading to unforeseen collateral damage. Third, there is significant structure-activity relationship data available to focus on specific families and classes of TSCs for further development. Fourth, the delivery, bioavailability, and metabolism of these compounds will need to be established and could vary widely depending on the structure of the complex and the metal ion chosen. Much work toward this area has been done with the few compounds that have reached clinical trials, and these efforts should help guide the development of additional compounds. In spite of the challenges that lie ahead, these compounds represent a promising area of further research.
Author Contributions
Writing—original draft preparation, X.J. Writing—review and editing, J.E.D., E.L., L.F.; visualization, L.F., J.E.D., X.J.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pommier, Y.; Sun, Y.; Huang, S.N.; Nitiss, J.L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703-721. [CrossRef]
- Thomas, A.; Pommier, Y. Targeting Topoisomerase I in the Era of Precision Medicine. Clin. Cancer Res. 2019, 25, 6581-6589. [CrossRef]
- McKie, S.J.; Desai, P.R.; Seol, Y.; Allen, A.M.; Maxwell, A.; Neuman, K.C. Topoisomerase VI is a chirally-selective, preferential DNA decatenase. Elife 2022, 11. [CrossRef]
- Vann, K.R.; Oviatt, A.A.; Osheroff, N. Topoisomerase II Poisons: Converting Essential Enzymes into Molecular Scissors. Biochemistry 2021, 60, 1630-1641. [CrossRef]
- Deweese, J.E.; Osheroff, N. The DNA cleavage reaction of topoisomerase II: wolf in sheep's clothing. Nucleic Acids Res. 2009, 37, 738-749. [CrossRef]
- Murphy, M.B.; Mercer, S.L.; Deweese, J.E. Inhibitors and Poisons of Mammalian Type II Topoisomerases. In Advances in Molecular Toxicology, Fishbein, J.C., Heilman, J., Eds.; Academic Press: Cambridge, MA, 2017; Volume 11, pp. 203-240. [CrossRef]
- Gibson, E.G.; Deweese, J.E. Covalent poisons of topoisomerase II. Curr. Top. Pharm. 2013, 17, 1-12.
- Moorthy, N.S.; Cerqueira, N.M.; Ramos, M.J.; Fernandes, P.A. Aryl- and heteroaryl-thiosemicarbazone derivatives and their metal complexes: a pharmacological template. Recent Pat. Anticancer Drug Discov. 2013, 8, 168-182. [CrossRef]
- Bai, X.G.; Zheng, Y.; Qi, J. Advances in thiosemicarbazone metal complexes as anti-lung cancer agents. Front. Pharmacol. 2022, 13, 1018951. [CrossRef]
- Matesanz, A.I.; Herrero, J.M.; Quiroga, A.G. Chemical and Biological Evaluation of Thiosemicarbazone-Bearing Heterocyclic Metal Complexes. Curr. Top. Med. Chem. 2021, 21, 59-72. [CrossRef]
- Liberta, A.E.; West, D.X. Antifungal and antitumor activity of heterocyclic thiosemicarbazones and their metal complexes: current status. Biometals 1992, 5, 121-126. [CrossRef]
- Miller, M.C., 3rd; Stineman, C.N.; Vance, J.R.; West, D.X.; Hall, I.H. The cytotoxicity of copper(II) complexes of 2-acetyl-pyridyl-4N-substituted thiosemicarbazones. Anticancer Res. 1998, 18, 4131-4139.
- Baruffini, E.; Ruotolo, R.; Bisceglie, F.; Montalbano, S.; Ottonello, S.; Pelosi, G.; Buschini, A.; Lodi, T. Mechanistic insights on the mode of action of an antiproliferative thiosemicarbazone-nickel complex revealed by an integrated chemogenomic profiling study. Sci. Rep. 2020, 10, 10524. [CrossRef]
- Karp, J.E.; Giles, F.J.; Gojo, I.; Morris, L.; Greer, J.; Johnson, B.; Thein, M.; Sznol, M.; Low, J. A phase I study of the novel ribonucleotide reductase inhibitor 3-aminopyridine-2-carboxaldehyde thiosemicarbazone (3-AP, Triapine) in combination with the nucleoside analog fludarabine for patients with refractory acute leukemias and aggressive myeloproliferative disorders. Leuk. Res. 2008, 32, 71-77. [CrossRef]
- Ma, B.; Goh, B.C.; Tan, E.H.; Lam, K.C.; Soo, R.; Leong, S.S.; Wang, L.Z.; Mo, F.; Chan, A.T.; Zee, B.; et al. A multicenter phase II trial of 3-aminopyridine-2-carboxaldehyde thiosemicarbazone (3-AP, Triapine) and gemcitabine in advanced non-small-cell lung cancer with pharmacokinetic evaluation using peripheral blood mononuclear cells. Invest. New Drugs 2008, 26, 169-173. [CrossRef]
- Nutting, C.M.; van Herpen, C.M.; Miah, A.B.; Bhide, S.A.; Machiels, J.P.; Buter, J.; Kelly, C.; de Raucourt, D.; Harrington, K.J. Phase II study of 3-AP Triapine in patients with recurrent or metastatic head and neck squamous cell carcinoma. Ann. Oncol. 2009, 20, 1275-1279. [CrossRef]
- Finch, R.A.; Liu, M.C.; Cory, A.H.; Cory, J.G.; Sartorelli, A.C. Triapine (3-aminopyridine-2-carboxaldehyde thiosemicarbazone; 3-AP): an inhibitor of ribonucleotide reductase with antineoplastic activity. Adv. Enzyme Regul. 1999, 39, 3-12. [CrossRef]
- Finch, R.A.; Liu, M.; Grill, S.P.; Rose, W.C.; Loomis, R.; Vasquez, K.M.; Cheng, Y.; Sartorelli, A.C. Triapine (3-aminopyridine-2-carboxaldehyde- thiosemicarbazone): A potent inhibitor of ribonucleotide reductase activity with broad spectrum antitumor activity. Biochem. Pharmacol. 2000, 59, 983-991. [CrossRef]
- Posa, V.; Hajdu, B.; Toth, G.; Domotor, O.; Kowol, C.R.; Keppler, B.K.; Spengler, G.; Gyurcsik, B.; Enyedy, E.A. The coordination modes of (thio)semicarbazone copper(II) complexes strongly modulate the solution chemical properties and mechanism of anticancer activity. J. Inorg. Biochem. 2022, 231, 111786. [CrossRef]
- Yalowich, J.C.; Wu, X.; Zhang, R.; Kanagasabai, R.; Hornbaker, M.; Hasinoff, B.B. The anticancer thiosemicarbazones Dp44mT and triapine lack inhibitory effects as catalytic inhibitors or poisons of DNA topoisomerase IIα. Biochem. Pharmacol. 2012, 84, 52-58. [CrossRef]
- Huang, H.; Chen, Q.; Ku, X.; Meng, L.; Lin, L.; Wang, X.; Zhu, C.X.; Wang, Y.; Chen, Z.; Li, M.; et al. A series of α-heterocyclic carboxaldehyde thiosemicarbazones inhibit topoisomerase IIα catalytic activity. J. Med. Chem. 2010, 53, 3048-3064. [CrossRef]
- Rao, V.A.; Klein, S.R.; Agama, K.K.; Toyoda, E.; Adachi, N.; Pommier, Y.; Shacter, E.B. The iron chelator Dp44mT causes DNA damage and selective inhibition of topoisomerase IIalpha in breast cancer cells. Cancer Res. 2009, 69, 948-957. [CrossRef]
- de Oliveira, J.F.; Lima, T.S.; Vendramini-Costa, D.B.; de Lacerda Pedrosa, S.C.B.; Lafayette, E.A.; da Silva, R.M.F.; de Almeida, S.M.V.; de Moura, R.O.; Ruiz, A.; de Carvalho, J.E.; et al. Thiosemicarbazones and 4-thiazolidinones indole-based derivatives: Synthesis, evaluation of antiproliferative activity, cell death mechanisms and topoisomerase inhibition assay. Eur J Med Chem 2017, 136, 305-314. [CrossRef]
- da Silva Filho, F.A.; de Freitas Souza, T.; Ribeiro, A.G.; Alves, J.E.F.; de Oliveira, J.F.; de Lima Souza, T.R.C.; de Moura, R.O.; do Carmo Alves de Lima, M.; de Carvalho Junior, L.B.; de Almeida, S.M.V. Topoisomerase inhibition and albumin interaction studies of acridine-thiosemicarbazone derivatives. Int. J. Biol. Macromol. 2019, 138, 582-589. [CrossRef]
- Samia, L.B.; Parrilha, G.L.; Da Silva, J.G.; Ramos, J.P.; Souza-Fagundes, E.M.; Castelli, S.; Vutey, V.; Desideri, A.; Beraldo, H. Metal complexes of 3-(4-bromophenyl)-1-pyridin-2-ylprop-2-en-1-one thiosemicarbazone: cytotoxic activity and investigation on the mode of action of the gold(III) complex. Biometals 2016, 29, 515-526. [CrossRef]
- Wilson, J.T.; Jiang, X.; McGill, B.C.; Lisic, E.C.; Deweese, J.E. Examination of the Impact of Copper(II) alpha-(N)-Heterocyclic Thiosemicarbazone Complexes on DNA Topoisomerase IIalpha. Chem. Res. Toxicol. 2016, 29, 649-658. [CrossRef]
- Morris, W.H.; Ngo, L.; Wilson, J.T.; Medawala, W.; Brown, A.R.; Conner, J.D.; Fabunmi, F.; Cashman, D.J.; Lisic, E.C.; Yu, T.; et al. Structural and Metal Ion Effects on Human Topoisomerase IIalpha Inhibition by alpha-(N)-Heterocyclic Thiosemicarbazones. Chem. Res. Toxicol. 2019, 32, 90-99. [CrossRef]
- Keck, J.M.; Conner, J.D.; Wilson, J.T.; Jiang, X.; Lisic, E.C.; Deweese, J.E. Clarifying the Mechanism of Copper(II) alpha-(N)-Heterocyclic Thiosemicarbazone Complexes on DNA Topoisomerase IIalpha and IIbeta. Chem. Res. Toxicol. 2019, 32, 2135-2143. [CrossRef]
- Vutey, V.; Castelli, S.; D'Annessa, I.; Samia, L.B.; Souza-Fagundes, E.M.; Beraldo, H.; Desideri, A. Human topoisomerase IB is a target of a thiosemicarbazone copper(II) complex. Arch Biochem Biophys 2016, 606, 34-40. [CrossRef]
- Chen, J.; Huang, Y.; Liu, G.; Afrasiabi, Z.; Sinn, E.; Padhye, S.; Ma, Y. The cytotoxicity and mechanisms of 1,2-naphthoquinone thiosemicarbazone and its metal derivatives against MCF-7 human breast cancer cells. Tox. App. Pharm. 2004, 197, 40-48. [CrossRef]
- Zeglis, B.M.; Divilov, V.; Lewis, J.S. Role of metalation in the topoisomerase IIα inhibition and antiproliferation activity of a series of α-heterocyclic-N4-substituted thiosemicarbazones and their Cu(II) complexes. J. Med. Chem. 2011, 54, 2391-2398. [CrossRef]
- Hall, I.H.; Miller, M.C.; West, D.X. Antineoplastic and Cytotoxic Activities of Nickel(II) Complexes of Thiosemicarbazones. Met Based Drugs 1997, 4, 89-95. [CrossRef]
- Beckford, F.; Thessing, J.; Woods, J.; Didion, J.; Gerasimchuk, N.; Gonzalez-Sarrias, A.; Seeram, N.P. Synthesis and structure of [(eta(6)-p-cymene)Ru(2-anthracen-9-ylmethylene-N-ethylhydrazinecarbothioamide)Cl]Cl; biological evaluation, topoisomerase II inhibition and reaction with DNA and human serum albumin. Metallomics 2011, 3, 491-502. [CrossRef]
- Wu, J.; Yang, T.; Wang, X.; Li, W.; Pang, M.; Sun, H.; Liang, H.; Yang, F. Development of a multi-target anticancer Sn(ii) pyridine-2-carboxaldehyde thiosemicarbazone complex. Dalton Trans 2021, 50, 10909-10921. [CrossRef]
- Beebe, S.J.; Celestine, M.J.; Bullock, J.L.; Sandhaus, S.; Arca, J.F.; Cropek, D.M.; Ludvig, T.A.; Foster, S.R.; Clark, J.S.; Beckford, F.A.; et al. Synthesis, characterization, DNA binding, topoisomerase inhibition, and apoptosis induction studies of a novel cobalt(III) complex with a thiosemicarbazone ligand. J. Inorg. Biochem. 2020, 203, 110907. [CrossRef]
- Qi, J.; Zheng, Y.; Qian, K.; Tian, L.; Zhang, G.X.; Cheng, Z.; Wang, Y. Synthesis, crystal structure and antiproliferative mechanisms of 2-acetylpyridine-thiosemicarbazones Ga(III) with a greater selectivity against tumor cells. J. Inorg. Biochem. 2017, 177, 110-117. [CrossRef]
- Teicher, B.A. Next generation topoisomerase I inhibitors: Rationale and biomarker strategies. Biochem. Pharmacol. 2008, 75, 1262-1271. [CrossRef]
- Oliveira, C.G.; Romero-Canelon, I.; Silva, M.M.; Coverdale, J.P.C.; Maia, P.I.S.; Batista, A.A.; Castelli, S.; Desideri, A.; Sadler, P.J.; Deflon, V.M. Palladium(ii) complexes with thiosemicarbazones derived from pyrene as topoisomerase IB inhibitors. Dalton Trans 2019, 48, 16509-16517. [CrossRef]
- Heng, M.P.; Sinniah, S.K.; Teoh, W.Y.; Sim, K.S.; Ng, S.W.; Cheah, Y.K.; Tan, K.W. Synthesis of a DNA-targeting nickel (II) complex with testosterone thiosemicarbazone which exhibits selective cytotoxicity towards human prostate cancer cells (LNCaP). Spectrochim. Acta. A Mol. Biomol. Spectrosc. 2015, 150, 360-372. [CrossRef]
- Qi, J.; Zheng, Y.; Li, B.; Ai, Y.; Chen, M.; Zheng, X. Pyridoxal hydrochloride thiosemicarbazones with copper ions inhibit cell division via Topo-I and Topo-IIa. J. Inorg. Biochem. 2022, 232, 111816. [CrossRef]
- Bisceglie, F.; Musiari, A.; Pinelli, S.; Alinovi, R.; Menozzi, I.; Polverini, E.; Tarasconi, P.; Tavone, M.; Pelosi, G. Quinoline-2-carboxaldehyde thiosemicarbazones and their Cu(II) and Ni(II) complexes as topoisomerase IIα inhibitors. J. Inorg. Biochem. 2015, 152, 10-19. [CrossRef]
- Miller III, M.C.; Stineman, C.N.; Vance, J.R.; West, D.X.; Hall, I.H. Multiple mechanisms for cytotoxicity induced by copper(II) complexes of 2-acetylpyrazine-N-substituted thiosemicarbazones. Appl. Organomet. Chem. 1999, 13, 9-19. [CrossRef]
- Sandhaus, S.; Taylor, R.; Edwards, T.; Huddleston, A.; Wooten, Y.; Venkatraman, R.; Weber, R.T.; Gonzalez-Sarrias, A.; Martin, P.M.; Cagle, P.; et al. A novel copper(II) complex identified as a potent drug against colorectal and breast cancer cells and as a poison inhibitor for human topoisomerase IIalpha. Inorg. Chem. Commun. 2016, 64, 45-49. [CrossRef]
- Bacher, F.; Enyedy, E.; Nagy, N.V.; Rockenbauer, A.; Bognar, G.M.; Trondl, R.; Novak, M.S.; Klapproth, E.; Kiss, T.; Arion, V.B. Copper(II) complexes with highly water-soluble L- and D-proline-thiosemicarbazone conjugates as potential inhibitors of Topoisomerase IIalpha. Inorg. Chem. 2013, 52, 8895-8908. [CrossRef]
- Bisceglie, F.; Pinelli, S.; Alinovi, R.; Goldoni, M.; Mutti, A.; Camerini, A.; Piola, L.; Tarasconi, P.; Pelosi, G. Cinnamaldehyde and cuminaldehyde thiosemicarbazones and their copper(II) and nickel(II) complexes: a study to understand their biological activity. J. Inorg. Biochem. 2014, 140, 111-125. [CrossRef]
- Lindsey, R.H.; Bender, R.P.; Osheroff, N. Stimulation of topoisomerase II-mediated DNA cleavage by benzene metabolites. Chem. Biol. Interact. 2005, 153-154, 197-205. [CrossRef]
- Lindsey, R.H., Jr.; Bender, R.P.; Osheroff, N. Effects of benzene metabolites on DNA cleavage mediated by human topoisomerase IIα: 1,4-hydroquinone is a topoisomerase II poison. Chem. Res. Toxicol. 2005, 18, 761-770. [CrossRef]
- Lindsey, R.H., Jr.; Bromberg, K.D.; Felix, C.A.; Osheroff, N. 1,4-Benzoquinone is a topoisomerase II poison. Biochemistry 2004, 43, 7563-7574. [CrossRef]
- Dutta, R.; Inouye, M. GHKL, an emergent ATPase/kinase superfamily. Trends Biochem Sci 2000, 25, 24-28. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



