Submitted:
22 June 2023
Posted:
24 June 2023
You are already at the latest version
Abstract
Cruor, the main component responsible for the red color of blood in mammals, contains 90% hemoglobin, a protein that is considered a rich source of bioactive peptides. The aim of the present study is to evaluate the potential of human hemoglobin as a source of bioactive peptides, compared to bovine hemoglobin, which has been widely studied in recent years. More specifically, the study focused on neokyotorphin, a natural antimicrobial peptide and meat preservative, which corresponds to the α137-141 fragment of bovine hemoglobin produced by enzymatic hydrolysis. Firstly, the results of the in-silico analysis showed that the two types of hemoglobin have a high similarity for the α and β chains (identities of 88% and 91%, respectively). Secondly, both types of hemoglobin were subjected to similar enzymatic hydrolysis conditions, which showed that the hydrolysis of human hemoglobin followed the same reaction mechanism as the hydrolysis of bovine hemoglobin, the 'Zipper' mechanism. The hydrolysis conditions (23°C, pH 3.5) made it possible to obtain α137-141 in significant quantities to allow for significant valorization from an industrial point of view. The hydrolyses were able to take place at high initial substrate concentrations, allowing for faster and more efficient processing of the co-product in the same amount of time, with over 65% of this production achieved in just 30 min. Increasing the substrate concentration (from 1% to 10% (w/v)) resulted in a proportional increase in α137-141 production. Regardless of the concentrations tested, the kinetic pattern remained similar. Finally, the results of the mass spectrometry analysis revealed the presence of several bioactive peptides in cattle and humans. Although some were known before, new bioactive peptides were discovered in human hemoglobin, such as four antibacterial peptides (α37-46 PTTKTYFPHF; α36-45 FPTTKTYFPH; α137-141 TSKYR; α133-141 STVLTSKYR), four opioid peptides (α137-141 TSKYR; β31-40 LVVYPWTQRF; β31-37 LVVYPWT), an ACE inhibitor (β129-135 KVVAGVA), an anticancer agent (β33-39 VVYPWTQ) and an antioxidant (α137-141 TSKYR). These peptides have never been found in human hemoglobin before, according to our knowledge. These results highlight the potential of human hemoglobin as a source of bioactive peptides useful for the food or pharmaceutical industry.
Keywords:
Bovine hemoglobin
; Human hemoglobin
; Bioactives peptides
; Neokyotorphin
1. Introduction
Blood, whether it comes from animal slaughterhouses or human blood donations, is a coproduct very rich in proteins. Indeed, the food and/or pharmaceutical industry is now turning to the valorization of these raw materials and seeking to develop products with high added value. Namely, only 30% of blood is valorized [1] in slaughterhouses, the rest is considered as waste although it is rich in proteins and constitutes an ideal substrate for proteolysis. It contains about 17.8% in cattle and 20-25% of all proteins encoded by the human genome are known to be secreted there [2]. The potential of these many co-products remains underutilized. These raw materials currently constitute new resources with high potential for valorization.
Bovine blood has been very successful commercially both in relation to its collection and processing, unlike most animal co-products [3]. Its wide availability and relative homogeneity make it a promising case study for the production and valorization of bioactive compounds[4,5]. Moreover, in humans also this body fluid is available in blood transfusion centers or collected through blood donation unlike saliva or tears.
Blood is essentially subdivided into two parts after centrifugation in vertebrates. The colorless part, plasma, represents about 60% of the volume of blood and can be used in pharmacy because of its richness in thrombins, fibrinogen or serum albumin [4,6] of the red color of blood and represents up to 40% of the blood volume. It is called "cruor" and contains mainly hemoglobin, 90%.
In recent studies, the role of these dietary proteins as a source of biologically active peptides is widely described. Hemoglobin represents more than half of the proteins in many databases that reference the various activities on active peptides [6,7].
Hemoglobin (H) in vertebrates is a tetramer of two α-chains and two β -globin chains, each of which contains a prosthetic heme [8].
For example, the hydrolysis of bovine hemoglobin by pepsin is the source of many peptides with several biological activities such as opioids [9,10], hematopoietic [11]and antihypertensive [12]. Nevertheless, the most described aspect remains the antimicrobial activity [13,14,15,16]. According to a study by Przybylski et al. in 2016, α137-141 peptide (also known as neokyotorphin, 653 Da, Pi of 10.5) is an antimicrobial peptide that has been identified as a natural preservative for meat during storage or distribution. This peptide has been shown to reduce rancidity time by 60% and inhibit microbial growth after 14 days in the refrigerator. The study found that α137-141 peptide had similar effectiveness to butylated hydroxytoluene (BHT), which is commonly used as a meat preservative. This discovery has the potential to revolutionize the food industry and provide a natural alternative to synthetic preservatives.
The most commonly used procedure to generate α137-141 peptide from hemoglobin is enzymatic hydrolysis [12,14,17]. Yet, in the same way as bovine hemoglobin, human hemoglobin too has a high protein richness. Indeed, as mentioned above, it is an ideal substrate for proteolysis generating bioactive peptides that could turn out to be an ingenious strategy for the process of discovery and development of natural potential candidates or molecularly targeted drug candidates for the substitution of synthetic compounds [18,19].
The laboratory has developed a patented strategy to obtain active peptides by hydrolysis of hemoglobin. Indeed, obtaining the latter remains extremely sensitive because it remains an irreversible process generating varied peptide populations according to plural operating parameters (pH, temperature...) which themselves influence the selectivity and specificity of the enzyme for its substrate [20,21].
In this perspective, the current work aims to demonstrate the feasibility of enzymatic hydrolysis of human hemoglobin to produce α 137-141. To do so, a comparative study of human hemoglobin versus bovine hemoglobin by bioinformatics study was first performed. Then, the peptic hydrolysis of human hemoglobin was compared to the peptic hydrolysis of bovine hemoglobin, the laboratory model, in order to define the similarities in the generated peptides and the presence of bioactive peptides.
This study had three specific objectives. The first was to identify the enzymatic mechanism involved in the hydrolysis of human hemoglobin. The second objective was to quantify the production of α137-141. Finally, the third objective was to characterize, identify the population of peptides and more particularly the active peptides resulting from the hydrolysis process.
2. Results and Discussion
2.1. In Silico Comparative Analysis of Bovine (Bos taurus) and Human (Homo sapiens) Hemoglobin
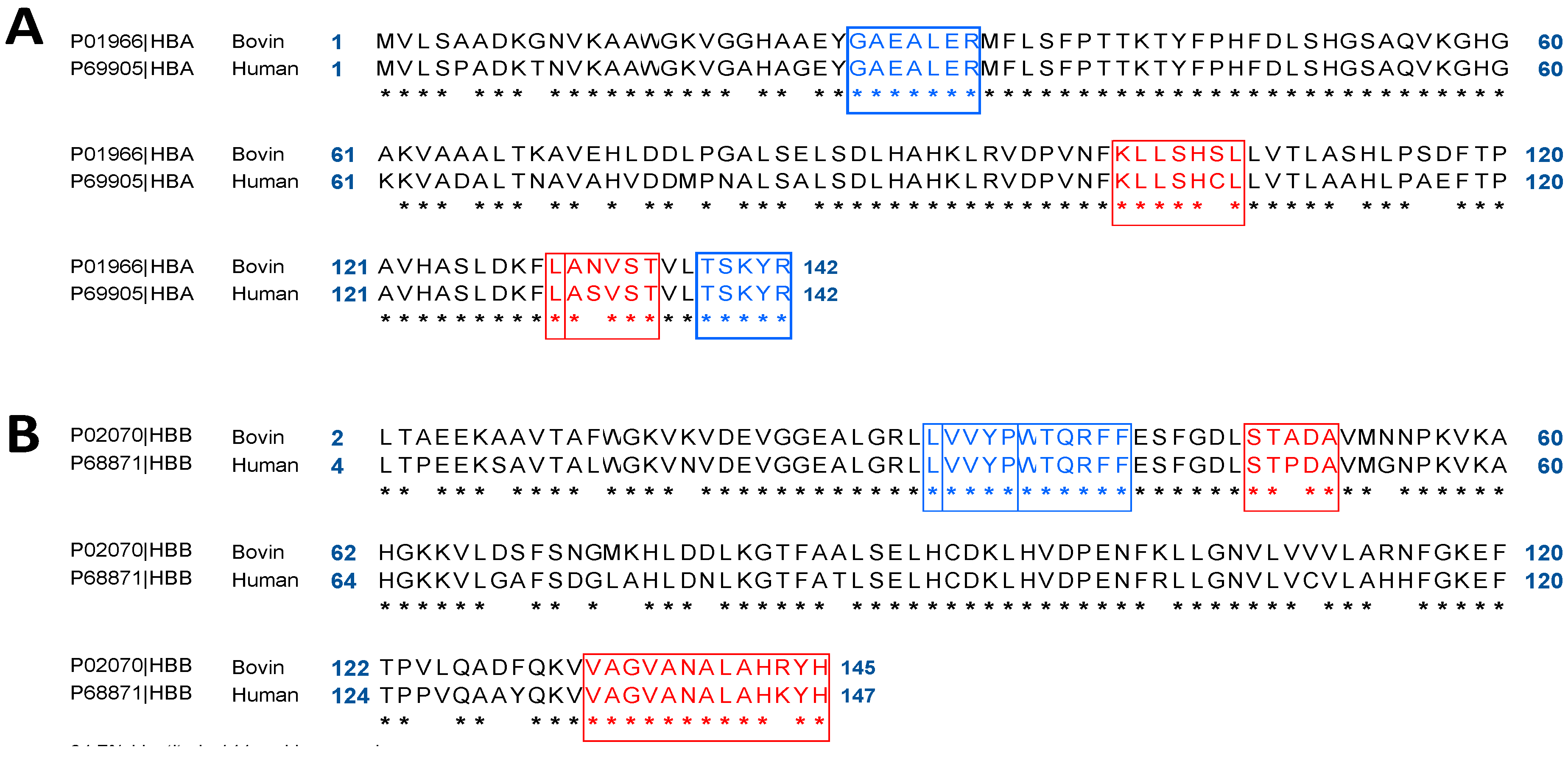
Human, with its scientific name Homo sapiens. For the domestic cattle, it is its scientific name Bos taurus. The two alpha and beta chains of bovine or human hemoglobin are identical two by two. The blast Figure 2 was therefore performed with two programs: (1) blastp from NCBI and (2) Local Similarity Program (SIM) from Expasy; to compare Homo sapiens (GI: 4504345) and Bos taurus (GI: 116812902) alpha and beta hemoglobin.
The two α-chains of bovine and human hemoglobin each consist of 142 amino acids, so there are no gaps when aligning the sequences. The Score obtained with blastp is 252, it corresponds to the statistical measure of the validity of the alignment. A high score indicates that the two sequences compared are similar. Below 50, the result is considered unreliable. The Expect value (e-value) corresponds to the value for which we consider that the matching is not due to chance. The closer the value is to zero, the better the result. Here it is 7.10-93 so the result is correct.
The alpha chains BosTaurus and Homo sapiens (Figure2A, SIM program) have many similarities in their amino acid sequences: 125 of the 142 are strictly aligned; the value of the identity obtained is 88%. Considering the similar amino acids, this positive value rises to 92%. This alignment of hemoglobin alpha chains allows us to observe that the peptides GAEALER (hematopoietic) and TSKYR (opioid, antimicrobial, and antioxidant) are conserved [22,32,33,34]. Contrarily, the peptides LANVST (Analgesic and Bradykinin Potentiator), ANVST (DPP-IV Inhibitor), and KLLSHSL (Antihypertensive) are only present in Bos taurus hemoglobin [12,35].
The same study was performed to align the amino acid sequences of the two β chains of hemoglobins Bos Taurus (GI: 27819608) and Homo sapiens (GI: 4504349). The β-chain of Bos Taurus contains 145 amino acids compared with 147 for the β-chain of homo sapiens. After sequence alignment, 147 amino acids were compared. The score obtained with blastp (254), the expectation value (2.10-93) and the gap value (0%) allows us to conclude that the alignment result is correct. The identity between these two β-chains is 85% and rises to 91% when similar amino acids are taken into account (Figure 2B, SIM program). The VVYP (lipid lowering) and LVVYPWTQRFF (opioid) peptides are conserved, as shown by this alignment of hemoglobin β-chains [32; 35]. Also, it should be noted that the human hemoglobin peptide VAGVANALAHKYH (coronary constrictor) has been modified to the peptide VAGVANALAHRYH in the β-chain of bovine hemoglobin. Furthermore, the STPDA peptide was modified from the STADA (bacterial growth stimulant) peptide of Bos taurus hemoglobin in the chain of human hemoglobin [29].
Figure 2.
Blast of hemoglobin alpha chain (A) and beta chain (B) from Bos taurus (bovine hemoglobin) and Homo sapiens (human hemoglobin). (A) Conservation of bioactive peptides GAEALER (hematopoietic) and TSKYR in blue (opioid, antioxidant, antimicrobial); Loss of peptides LANVST (Analgesic and Bradykinin Potentiator), ANVST (DPP-IV Inhibitor), and KLLSHSL (Antihypertensive) in red. (B) Conservation of the bioactive peptides VVYP (lipid-lowering), LVVYPWTQRFF (opioid) in blue and loss of the peptides VAGVANALAHKYR (coronary constrictor) and STADA (bacterial growth stimulator) in red. Blast were performed using Local Similarity Program (SIM) from Expasy.
Figure 2.
Blast of hemoglobin alpha chain (A) and beta chain (B) from Bos taurus (bovine hemoglobin) and Homo sapiens (human hemoglobin). (A) Conservation of bioactive peptides GAEALER (hematopoietic) and TSKYR in blue (opioid, antioxidant, antimicrobial); Loss of peptides LANVST (Analgesic and Bradykinin Potentiator), ANVST (DPP-IV Inhibitor), and KLLSHSL (Antihypertensive) in red. (B) Conservation of the bioactive peptides VVYP (lipid-lowering), LVVYPWTQRFF (opioid) in blue and loss of the peptides VAGVANALAHKYR (coronary constrictor) and STADA (bacterial growth stimulator) in red. Blast were performed using Local Similarity Program (SIM) from Expasy.
2.2. Bioinformatics Approach to Predict Peptides Derived from the Pepsic Hydrolysis of Bovine Hemoglobin Bos Taurus and Human Hemoglobin Homo sapiens
Bos taurus hemoglobin’s alpha and beta chain sequences (GI: 116812902; GI: 27819608) and human hemoglobin’s (GI: 4504345; GI: 4504349) were submitted into the Peptide Cutter software (Expasy). Figure 3A illustrates the predicted cleavage sites for peptic hydrolysis at pH higher than 2 of the alpha chains of two hemoglobins. They number 43 and 39, respectively. When the comparable amino acids are considered, Blast reports a 92% identity between the alpha chains of Homo sapiens and the Bos Taurus. Peptide Cutter did identify 4 distinct cleavage sites, which are indicated by red arrows. Bottom arrows pointed to several different cleavage sites. Note that for the alpha chain of human hemoglobin, one of the missing cleavages is the one that obtains the peptide VAAA (DPP-IV inhibitor) [12,35]. By prediction, this peptide will not be recovered during the hydrolysis of human hemoglobin.
Figure 3.
Peptide Cutter, Potential cleavage sites of the alpha (A) and beta (B) chains of bovine (Bos taurus) and human (Homo sapiens) hemoglobin by pepsin at pH > 2. The black arrows indicate the same predicted cleavage sites and in red differ only for one of the two hemoglobin chains. Blast were performed using Local Similarity Program (SIM) from Expasy and cleavage sites were predicted by Peptide Cutter from Expasy.
Figure 3.
Peptide Cutter, Potential cleavage sites of the alpha (A) and beta (B) chains of bovine (Bos taurus) and human (Homo sapiens) hemoglobin by pepsin at pH > 2. The black arrows indicate the same predicted cleavage sites and in red differ only for one of the two hemoglobin chains. Blast were performed using Local Similarity Program (SIM) from Expasy and cleavage sites were predicted by Peptide Cutter from Expasy.
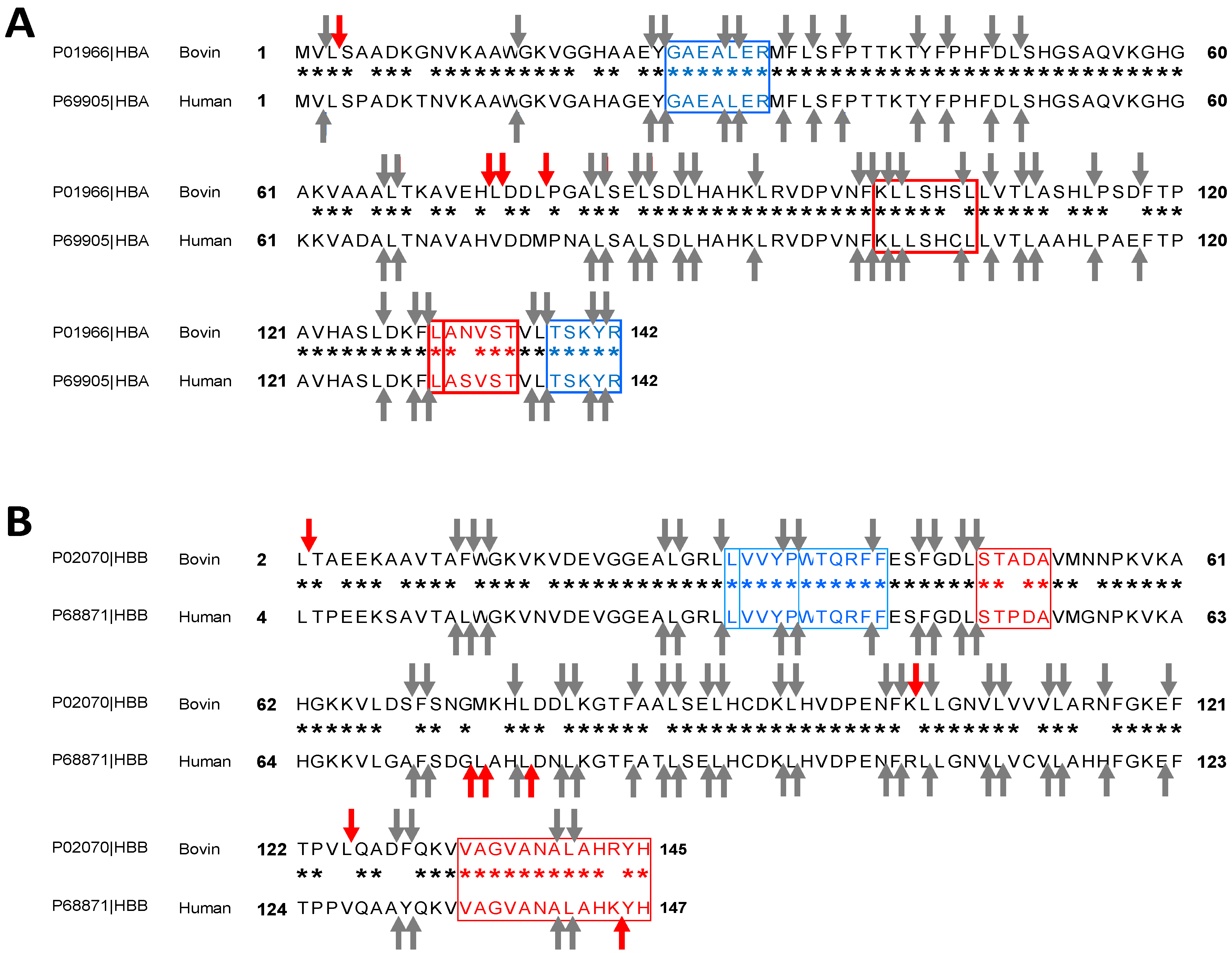
The cleavage sites predicted by Peptide Cutter, for the peptic hydrolysis, at pH higher than 2, of the beta chains of are presented in Figure 3B. 41 and 42 potential cleavages were identified for the beta chains of bovine Bos Taurus and human Homo sapiens hemoglobin respectively. However, we note that the cleavages are not identical: 10 different cleavages sites were identified by bottom arrows. Considering the similar amino acids, the identity between these two beta chains Bos taurus and Homo sapiens given by Blast rises to 89%. For example, for the bovine hemoglobin beta chain, one of the cleavages results in a cut of the bacterial growth stimulating peptide STADA [29], which is not the case for the Homo sapiens beta chain. By prediction, this peptide will not be found during the hydrolysis of human hemoglobin. On the other hand, the peptides VVYP (lipid-lowering), LVVYPWTQRFF (opioid) are conserved during peptic hydrolysis at pH above 2 according to Peptide Cutter [12,29,36].
2.3. Enzymatic Kinetics and Mechanism of Action Involved
2.3.1. Determination of the Degree of Hydrolysis
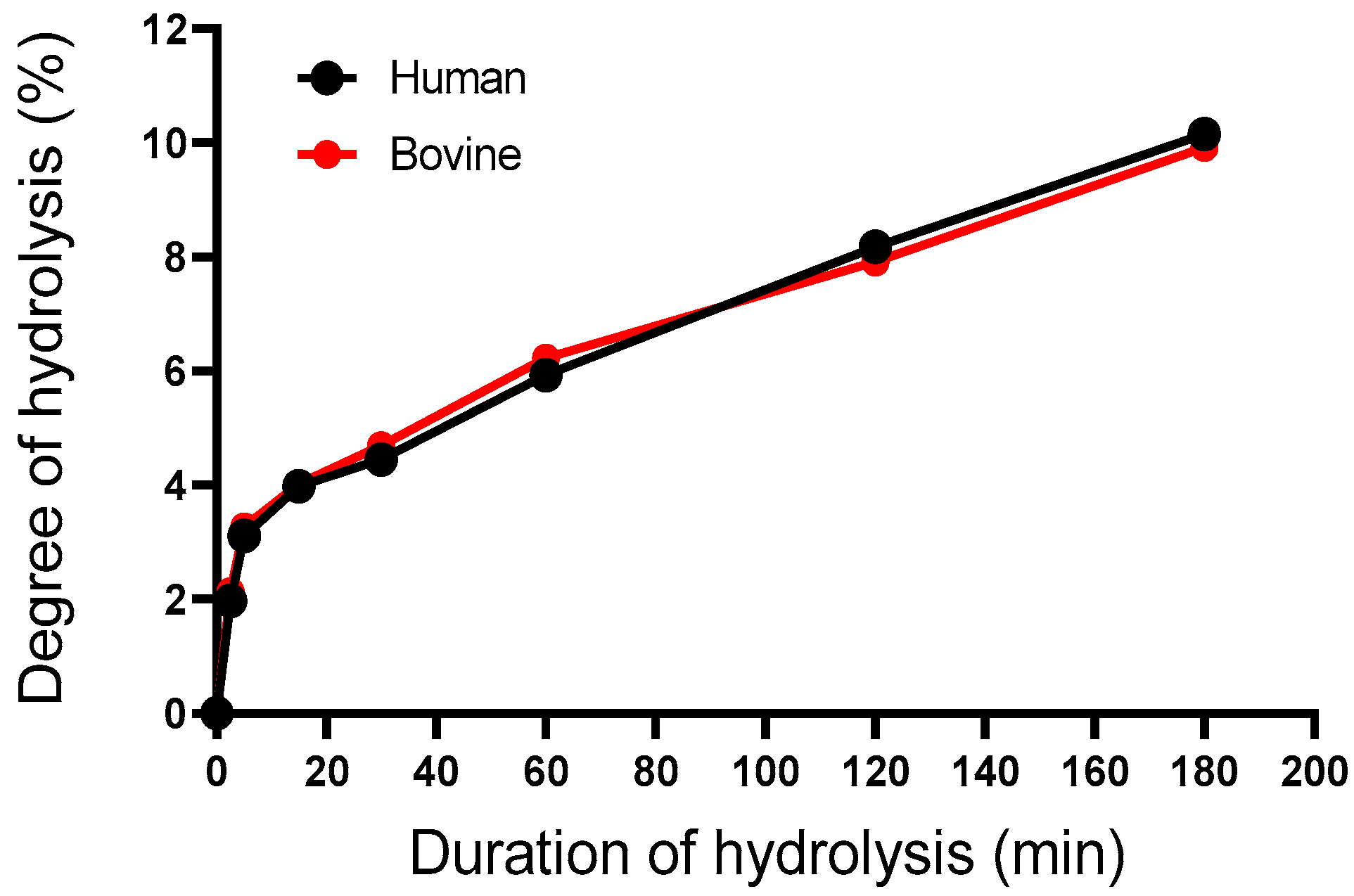
The primary objective of this study was to make a comparison between the hydrolysis of bovine hemoglobin and the hydrolysis of human hemoglobin. It would therefore be interesting to compare the different DHs under the applied hydrolysis conditions (23°C, pH 3.5) and at a CBH; CHH of 1% (w/v).
The degree of hydrolysis (DH) did not show a linear relationship with time in both species (Figure 4). As time progressed, the evolution of DH was identical, which is illustrated in Figure, both curves have the same shape. The curves had the same shape as those that had previously been reported in the literature [16]. Respectively, the DH was 2, 3, 4.5, 6, 8, and 10% when the hydrolysis time was 2.5, 5, 15, 30 min, 1, 2, and 3h under these conditions. This is indeed an indicator of the progress of the enzymatic reaction.
Figure 4.
Evolution of the degree of hydrolysis of bovine and human hemoglobin during its hydrolysis treated with porcine pepsin for 180 min (3h) (pH 3.5, 23°C, E/S = 1/11, CBH/CHH= 1%, w/v).
Figure 4.
Evolution of the degree of hydrolysis of bovine and human hemoglobin during its hydrolysis treated with porcine pepsin for 180 min (3h) (pH 3.5, 23°C, E/S = 1/11, CBH/CHH= 1%, w/v).
2.3.2. Identification of α137-141 by Waters Software
The identification of the α137-141 peptide was performed with the Waters software. Several parameters are then considered: the retention time of the peptide, its real-time spectrum, and the estimation of the peak purity, all three described in detail previously (see §2.5.2). This method has been shown to be effective in identifying and quantifying α137-141 in bovine cruor previously published in the literature [16].
The standard α137-141 had a retention time of approximately 5.3-5.5min. Next, the peptide from the human and bovine hydrolysate was analyzed by comparing them to the standard spectrum. With the four parameters (MA, MT, PA, and PT) and retention time, the peak at time a little over 5min was identified as α137-141. All measurements showed that PA was lower than PT and MA was lower than MT, highlighting good purity of the peak and the similarity of the spectrum to α137-141.
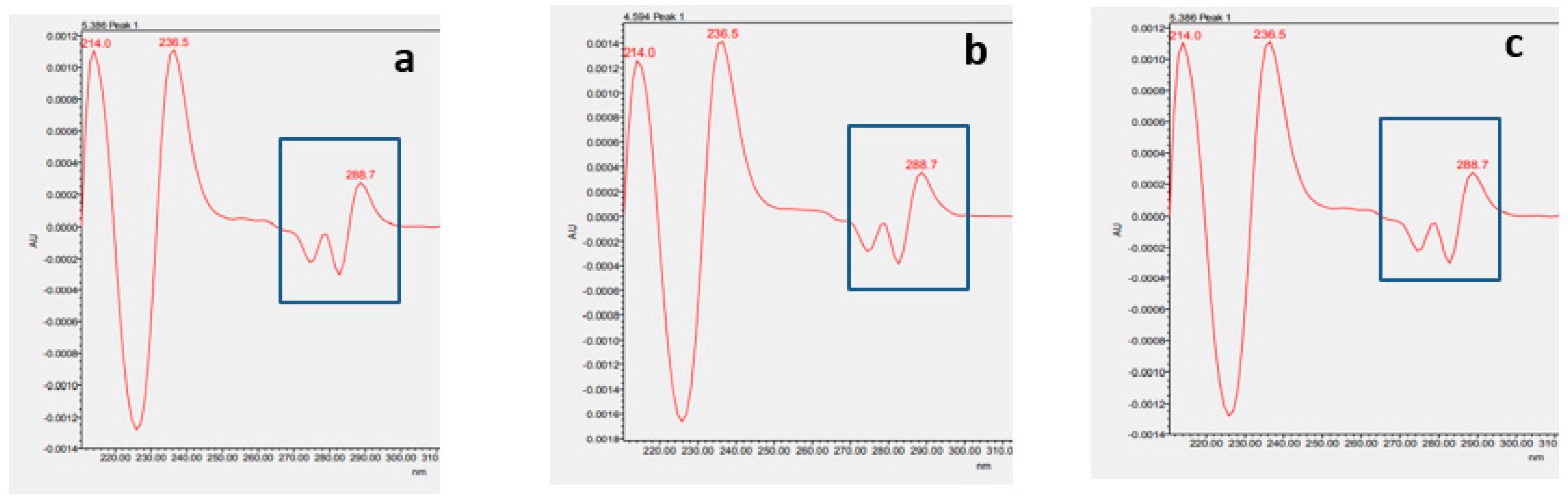
In addition, a comparison of second derivative spectra (Figure 5) between standard α137-141 peptide (a) and obtained α137-141 peptide from the hydrolysis of bovine hemoglobin (b) or human hemoglobin (c) by RP-UPLC was made. This analysis allows the estimation of the presence or absence of aromatic amino acid in the peak [30]. In this case, the presence of a tyrosine was assessed, which is in agreement with the sequence of the α137-141 peptide [23]. The specific maxima at 288.7 nm indicated the presence of this amino acid. Their excellent similarity thus allowed the peak at a retention time of 5min to be identified within the human and bovine hydrolysates as α137-141.
Figure 5.
Comparison of second derivative spectra between (a) standard α137-141 peptide and (b) obtained α137-141 peptide from bovine hemoglobin hydrolysisand (c) obtainedα137-141 peptide from human hemoglobin hydrolysis.
Figure 5.
Comparison of second derivative spectra between (a) standard α137-141 peptide and (b) obtained α137-141 peptide from bovine hemoglobin hydrolysisand (c) obtainedα137-141 peptide from human hemoglobin hydrolysis.
2.3.3. Reaction Mechanism of the Enzymatic Hydrolysis of Human Hemoglobin by Pepsin
To understand the mechanism of enzymatic hydrolysis of human hemoglobin, the RP-UPLC chromatographic profiles obtained as a function of time were analyzed. Figure 6 presents the chromatograms obtained for identical hydrolysis times on the one hand with bovine hemoglobin, on the other hand with human hemoglobin, both for a CBH and CHH equal to 1% (w/v) at 23°C, pH 3.5. At first sight, it can be noticed that for a given DH, the chromatograms were almost the same. This first element ensured that the kinetics of obtaining peptides from bovine hemoglobin and that of man were subject to the same reaction mechanisms. The accessibility of pepsin for its substrate thus remained essentially the same. This can be explained by the high similarity between the amino acids of the two species, as mentioned above in the in-silico study, for the α and β chains with identities of 88% and 91%, respectively. The first known fraction was α137-141 and it was present in all degrees of hydrolysis.
Before the addition of pepsin, three peaks are shown at T0:α-chain, β-chain, and heme. The hemoglobin subunits were quickly hydrolyzed once the enzyme was added, resulting in intermediate peptides that were then converted into final peptides, which are generally more hydrophilic and smaller than the intermediate peptides. This enzymatic mechanism, called "Zipper", was first described by Linderstrom-Lang (1953) [21]. This is explained by the fact that hemoglobin (bovine or human) was initially in its native form called "globular". The latter was denatured by lowering the pH to 3.5 by adding 2M HCl. It is thus found in an acidic environment in its "expanded" form. This favored the attack of the alpha and beta chains by the enzyme. This favors the enzymatic mechanism "Zipper" to the detriment of the mechanism "one-by-one", observed for higher pH [31]. At the end of the reaction, a set of peptides in solution were analyzed.
The same mechanism has been observed in other studies of hemoglobin hydrolysis and described by Nedjar-Arroume et al, (2008) [14,16,31].
Figure 6.
Chromatographic hydrolysis profiles of bovine hemoglobin compared to human hemoglobin at 215 nm by RP-UPLC, analyzed by C4 type column (pH 3.5, 23°C, E/S = 1/11, CBH = 1%, CHH= 1%, w/v) (A) bovine hemoglobin; (B) human hemoglobin.
Figure 6.
Chromatographic hydrolysis profiles of bovine hemoglobin compared to human hemoglobin at 215 nm by RP-UPLC, analyzed by C4 type column (pH 3.5, 23°C, E/S = 1/11, CBH = 1%, CHH= 1%, w/v) (A) bovine hemoglobin; (B) human hemoglobin.
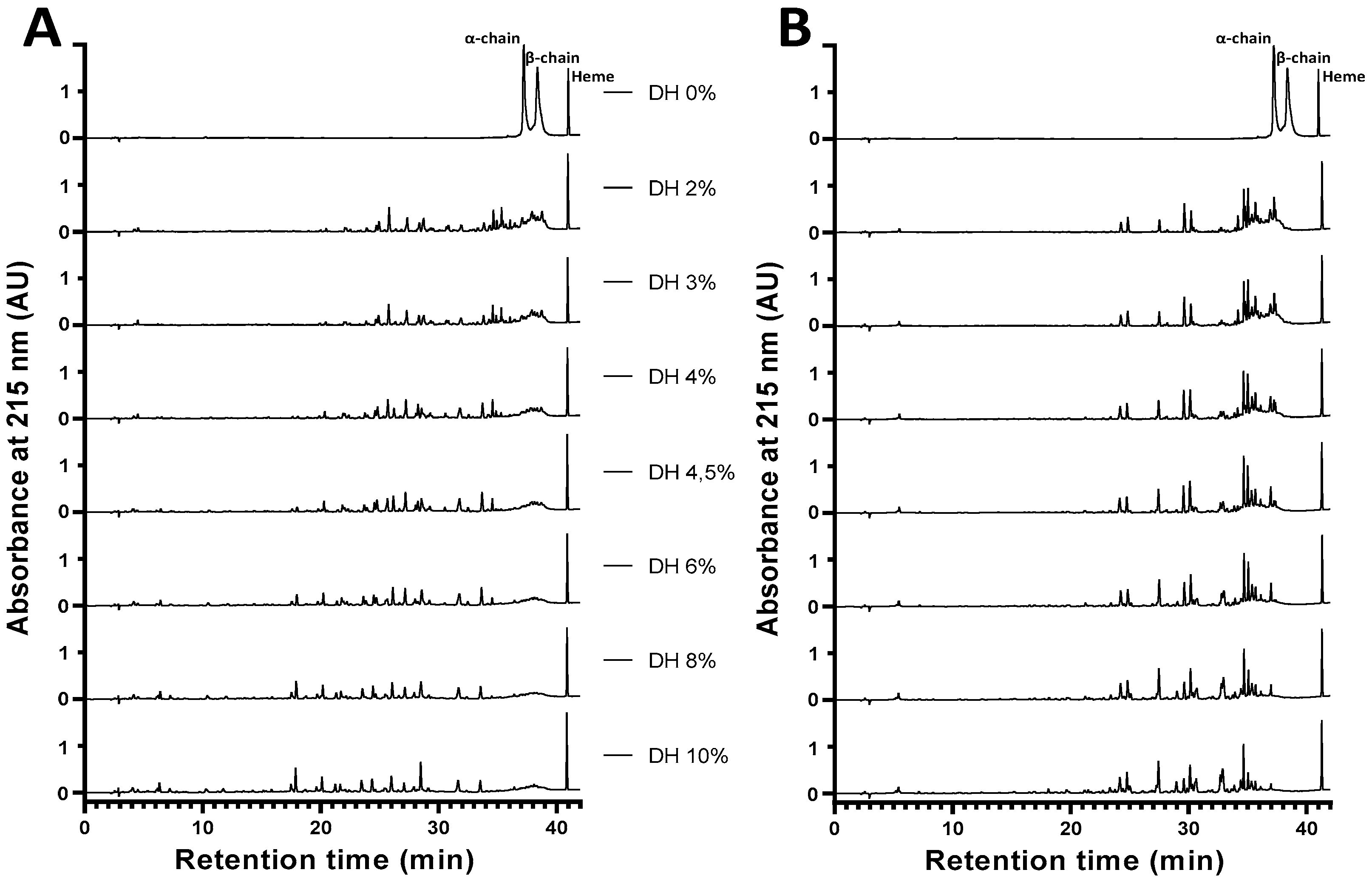
Under the hydrolysis conditions used here (23 °C; pH 3.5; CBH and CHH of 1% w/v), the production of α137-141 was rapid. This peptide appeared early, a mere 2 minutes after the start of hydrolysis i.e. DH= 1% and its concentration increased throughout the hydrolysis, so α137-141 accumulated during the process. It was therefore a final peptide: it was not hydrolyzed into a smaller peptide, in agreement with Lignot, Froidevaux, Nedjar-Arroume and Guillochon (1999) [9,14]who specified that α137-141 was a final peptide.
On the chromatographic profiles (Figure 6), other fractions were observed containing numerous intermediate peptides that were progressively hydrolyzed to final peptides throughout the hydrolysis of hemoglobin.
In conclusion, the hydrolysis of human hemoglobin showed the same enzymatic mechanism as the hydrolysis of bovine hemoglobin. It is indeed a potential source to produce active peptides such as antimicrobial peptides.
2.4. Obtaining the Active Peptide α137-141 during the Peptide Hydrolysis of Human and Bovine Hemoglobins
2.4.1. Effect of Increasing the Initial Concentration of Bovine Hemoglobin on the Enzymatic Obtention of Active Peptide α137-141
Since the peak of the peptide of interest, α137-141 was pure; the determination of the production of this peptide during hydrolysis could therefore be performed by RP-UPLC, while respecting the parameters previously defined (see Materials and Methods). It was necessary to increase the CBH and CHH in order to evaluate the production of the peptide on a larger scale and to verify the production of the peptide at sufficiently high initial concentrations. For this purpose, the assay was performed for all CBH and CHH studied from hemoglobin (1, 2, 8, and 10%, w/v) and for all DHs (0 to 10%). Chromatographic profiles of bovine and human hydrolysis for all CBH and CHH tested are shown, respectively in Supplementary Data File S1. It is then observed that whatever the concentration of human/bovine hemoglobin initially used, the hydrolysis mechanism, of zipper type, remained identical. Moreover, the RP-UPLC analyses indicated that the identification of the peptide was not impacted by the increase of CHB / CHH.
These measurements not only verified the production of α137-141 at higher hemoglobin concentration, in addition, it confirmed the ability of pepsin to hydrolyze at high substrate concentrations. However, a concentration higher than 10% (w/v) could be problematic because in this case, the initial medium was no longer homogeneous if the solubilization of hemoglobin was performed simply in water, without addition of salt.
2.4.2. Quantification of the Production of the Active Peptide α137-141
Figure 7 shows the kinetics of α137-141 peptide appearance as a function of DH and the different CBH and CHH tested. Looking at the peptic hydrolysis of human hemoglobin, at a CHH of 1% (w/v) and a DH of 10%, Cα137-141 was 69.70 ±6,76 mg L−1. Doubling the initial CHH to 2% (w/v), Cα137-141 was also increased by a factor of 2 to reach a final concentration of 138.73 ±6.57 mg.L-1. With 8% (w/v) CHH, hydrolysis yielded a final α137-141 concentration equal to 562.03±106.26 mg. L-1, i.e., a concentration approximately eight times higher than that obtained at 1% (w/v). Finally, hydrolysis with CHH equal to 10% (w/v) yielded about ten times that of 1% (w/v), i.e. Cα137–141 = 687.98±75.77 mg.L-1. Exactly the same findings were made in the pepsic hydrolysis of bovine hemoglobin. This is in accordance with what was described by Przybylski, R et al; (2016)[16].These results once again affirm that the enzyme maintains the same hydrolysis mechanism, of human hemoglobin that bovine hemoglobin.
Figure 7.
Appearance kinetics of the α137-141 peptide during pepsic hydrolysis of bovine hemoglobin BH (A) and human hemoglobin HH (B) hydrolysis at different bovine (CBH) and human (CHH) hemoglobin concentrations (w/v), (pH 3.5, 23°C, E/S = 1/11). A comparative study was performed between the different graphs in the same condition and showed no significant difference (ns) between human hemoglobin and bovine hemoglobin (multiple t test comparisons).
Figure 7.
Appearance kinetics of the α137-141 peptide during pepsic hydrolysis of bovine hemoglobin BH (A) and human hemoglobin HH (B) hydrolysis at different bovine (CBH) and human (CHH) hemoglobin concentrations (w/v), (pH 3.5, 23°C, E/S = 1/11). A comparative study was performed between the different graphs in the same condition and showed no significant difference (ns) between human hemoglobin and bovine hemoglobin (multiple t test comparisons).
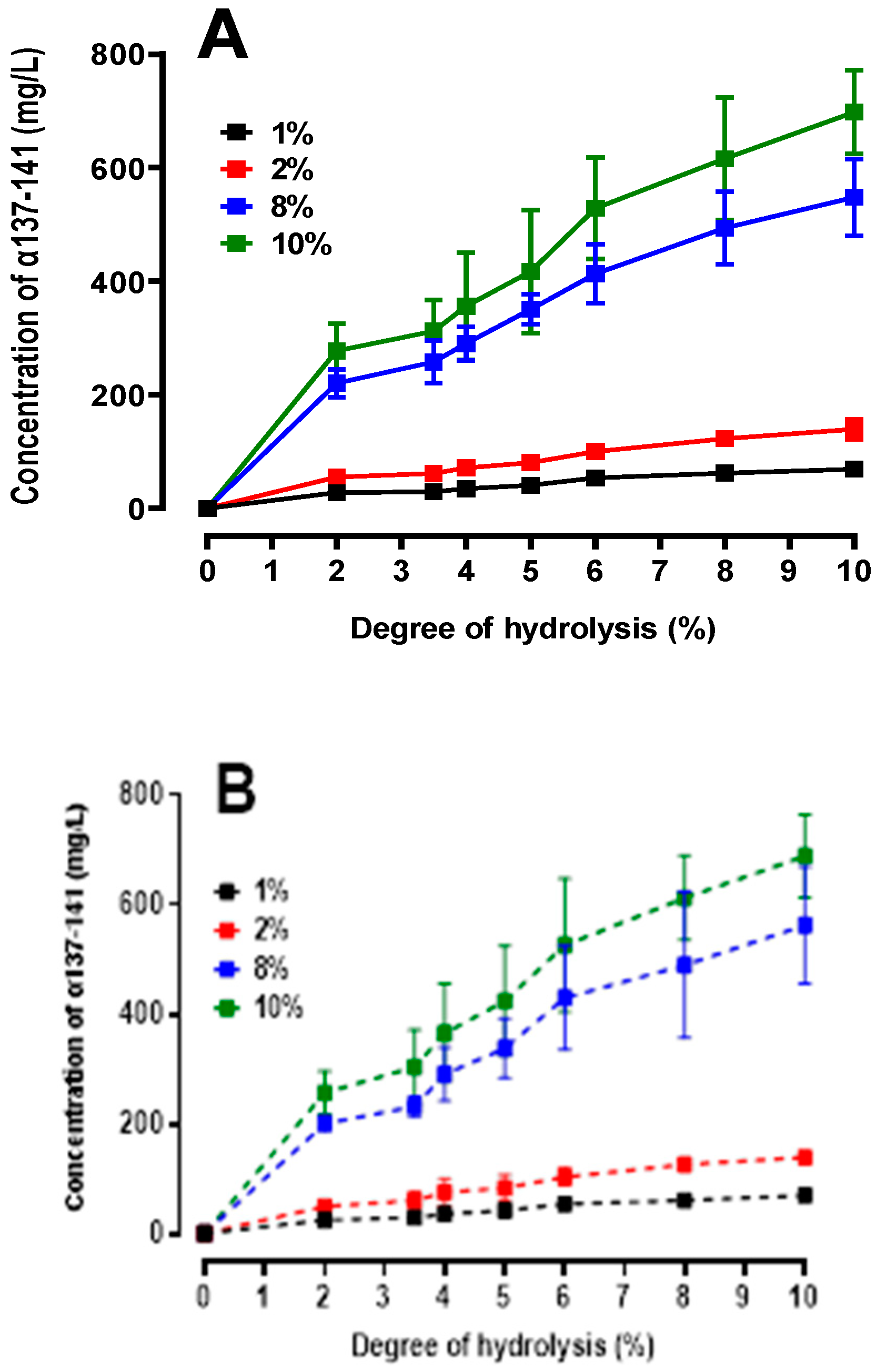
In addition, it is noted that the mechanism is rapid because α137-141 appears very early. Regardless of the CHH / CBH tested, the shape of the kinetics remained the same.
The appearance of α137-141 slows down and the kinetics appear to reach a plateau. These data ensure that α137-141 was a final peptide under these hydrolysis conditions in both human and bovine [9,16].
In an effort to complement the previous observations, it was of interest to evaluate the yield of α137-141 production in both species. Figure 8 shows the α137-141 production yields for all CHH/ CBHtested. The yield was calculated using the following formula:
Where Cα137-141 is the α137-141 concentration measured in UPLC for a given DH and CBH/HH and Cα137-141max being the theoretical maximum α137-141 concentration that can be obtained for a given CBH/HH.
Figure 8.
Production yields of α137-141 during the peptic hydrolysis of bovine (A) human (B) hemoglobin at different concentrations (w/v) (pH 3.5, 23°C, E/S = 1/11). A comparative study was performed between the two histograms and showed no significant difference (ns) between human hemoglobin and bovine hemoglobin (multiple t-test comparisons).
Figure 8.
Production yields of α137-141 during the peptic hydrolysis of bovine (A) human (B) hemoglobin at different concentrations (w/v) (pH 3.5, 23°C, E/S = 1/11). A comparative study was performed between the two histograms and showed no significant difference (ns) between human hemoglobin and bovine hemoglobin (multiple t-test comparisons).
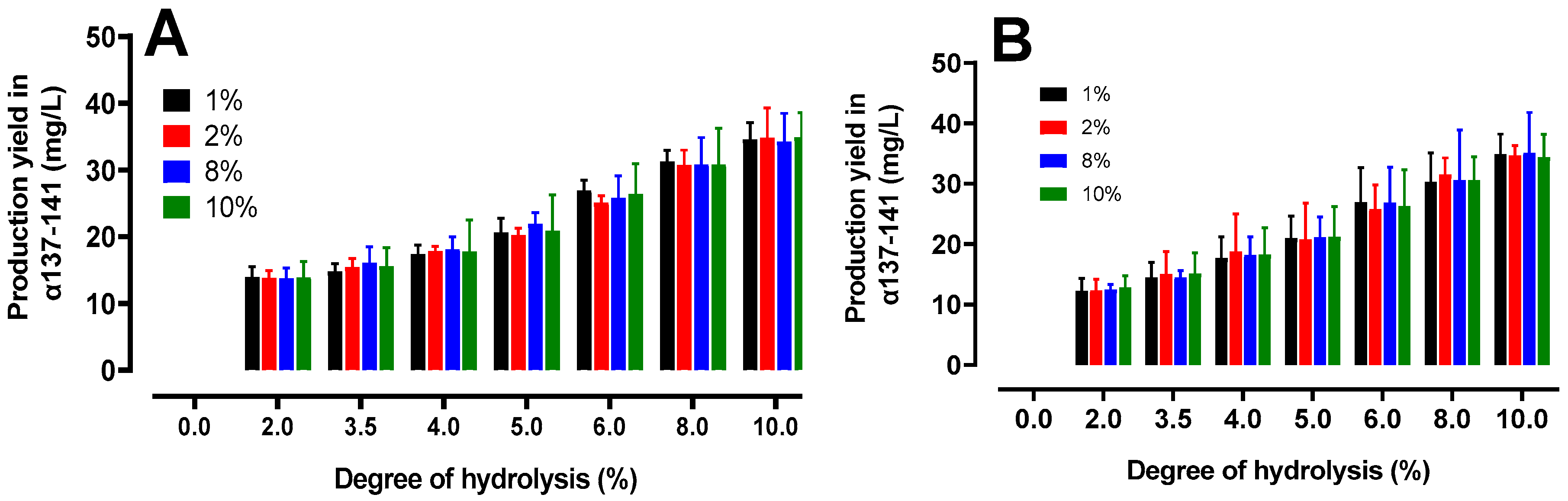
At a final DH of 10%, the average yield of α137-141 was around 34 %. Considering as maximum the production of α137-141 at 3 hours, it could be estimated that around only 30min of hydrolysis (i.e. a DH of 5% is equivalent around to 21%), 63% of the finally generated peptide is already generated. This could be justified by the generation of peptides during hydrolysis that compete as new substrates on the active site of the enzyme, thus slowing down the hydrolysis rate [20,37]. As previously observed, the production of the peptide of interest progressed rapidly, correlating the trend towards the plateau at the end of the reaction. There were no significant differences between bovine and human hemoglobin peptide hydrolysis. These data could allow a reflection in terms of profitability compared to the valorization of this active peptide.
2.5. Peptidomics Approach to Characterizing the Peptide Populations.
The peptide heterogeneity after 3 hours of pepsic hydrolysis of bovine and human hemoglobin was determined using a peptidomic approach. Triplicate RP-HPLC-MS/MS runs of both hydrolysates were performed, and MS and MS/MS data were recorded. As illustrated Figure 9A, irrespective to the species, the number of MS (around 6,000) and MS/MS (around 10,600) scans (compared two-by-two) were not significantly different (p <0.05), indicating that the MS and MS/MS data were comparable between species. Therefore, Peaks® Studio XPro identified 217 and 189 unique peptides from bovine and human hemoglobin hydrolysates, respectively. Among these latter, only 57 are unique peptides strictly common between species (Figure 9B).
Figure 9.
Peptidomics analysis, performed by RP-HPLC-MS/MS and bioinformatics, from bovine and human hemoglobin after a 3-hour hydrolysis period. (A) Number of MS and MS/MS scans. (B) Venn diagram showing the number of distinct and common hemoglobin peptides identified between the two hydrolysates. (C) and (D) 2D- and 3D-heat maps highlighting the occurrence of detected peptides along the amino acid sequences for hemoglobin subunit alpha (C) and subunit beta (D). The color code, corresponding to the peptides containing the amino acids that have been identified, is as follow: white color, no peptide containing these amino acids has been identified; yellow color, between 1 to 10 peptides; orange color, between 11 to 20 peptides; red color, between 21 to 30 peptides; black color, more than 31 peptides containing these amino acids have been identified. The grey bars under the 2D-heat maps correspond to unique peptide sequences identified. Heat maps using the 3D modeling were performed with SWISS-MODEL bioinformatics web-server. Heme is depicted in grey color. The hydrolysates were analyzed in triplicate. N-terminal extremity, (Nter) and C-terminal extremity (Cter).
Figure 9.
Peptidomics analysis, performed by RP-HPLC-MS/MS and bioinformatics, from bovine and human hemoglobin after a 3-hour hydrolysis period. (A) Number of MS and MS/MS scans. (B) Venn diagram showing the number of distinct and common hemoglobin peptides identified between the two hydrolysates. (C) and (D) 2D- and 3D-heat maps highlighting the occurrence of detected peptides along the amino acid sequences for hemoglobin subunit alpha (C) and subunit beta (D). The color code, corresponding to the peptides containing the amino acids that have been identified, is as follow: white color, no peptide containing these amino acids has been identified; yellow color, between 1 to 10 peptides; orange color, between 11 to 20 peptides; red color, between 21 to 30 peptides; black color, more than 31 peptides containing these amino acids have been identified. The grey bars under the 2D-heat maps correspond to unique peptide sequences identified. Heat maps using the 3D modeling were performed with SWISS-MODEL bioinformatics web-server. Heme is depicted in grey color. The hydrolysates were analyzed in triplicate. N-terminal extremity, (Nter) and C-terminal extremity (Cter).
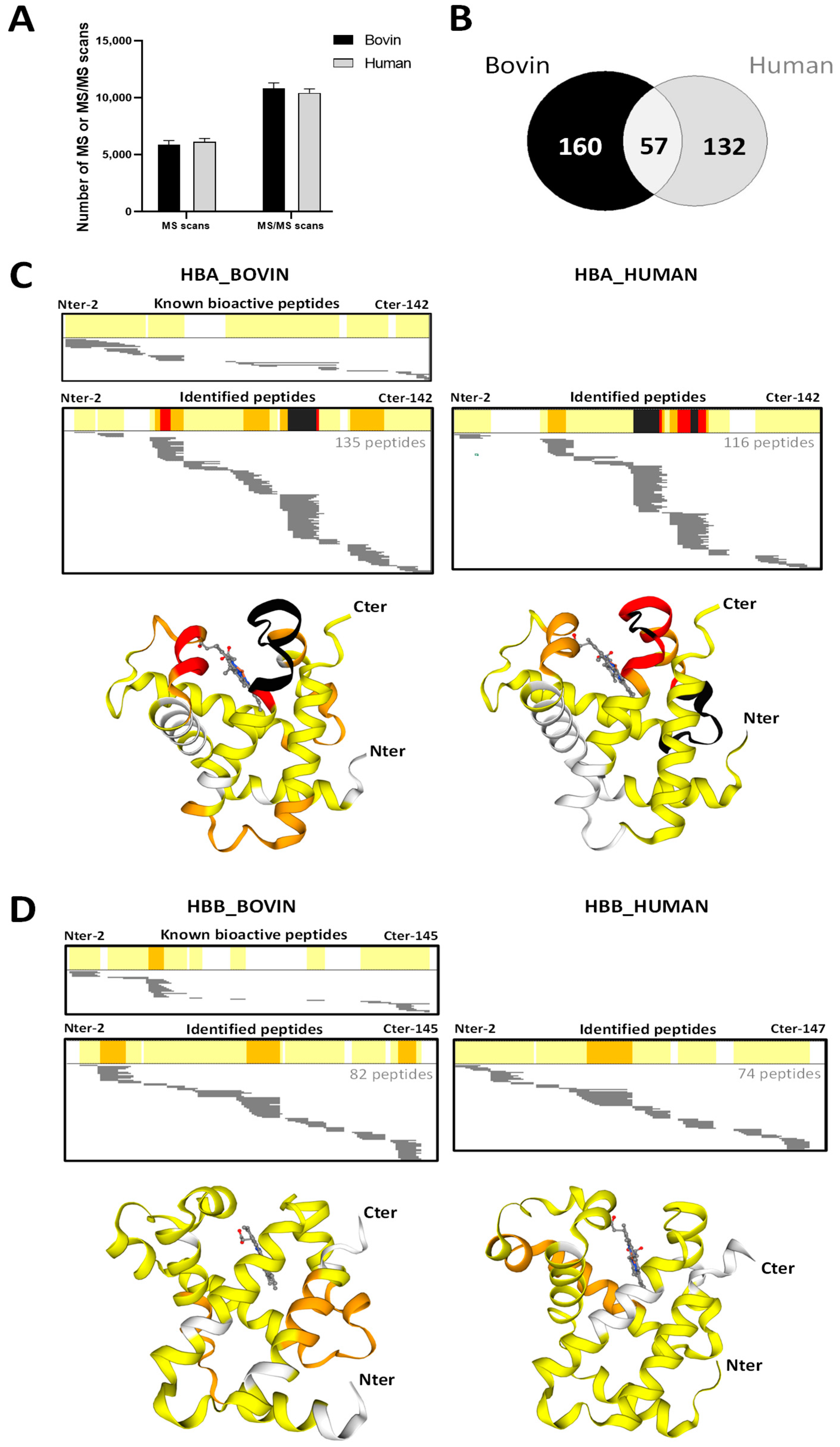
As illustrated in the Figure 9C, among the 217 unique peptides identified from bovine hemoglobin hydrolysate, 135 and 82 unique peptides came from the alpha- and beta-chains, respectively. In the same way, from the 189 unique peptides from human one, 116 and 74 came from the alpha- and beta-chains (Figure 9D), respectively. The Figure 9C,D illustrate, according to the same color code through the 2D- and 3D-heatmaps, the identified-peptide repartition all along the amino acid backbone for each hemoglobin chain. The higher the occurrence, the more the peptide zone tends towards red then black. Therefore, these heat maps underline the high abundance of detected peptides through the red- to black-colored protein regions, where the proteolysis is limited, and the low abundance of detected peptides in the non-colored protein regions, where the proteolysis is important.
Respective to species, the 2D-heat maps of alpha-chains (Figure 9C) mainly highlight three to four protein regions (in orange, red and black) where the peptides are resistant to pepsin hydrolysis due to the high number of unique peptides identified in these regions. For the bovine hemoglobin alpha-chain, pepsin-resistant protein regions are sources of bioactive peptides (see Figure 9C, upper panel). Unfortunately, the same comparison cannot be done for the human one due to the absence of bioactive peptides descripted from the human hemoglobin. More interesting, probably due to the high similarity between the 3D-structures of alpha-chains of both species and despite to the acid denaturation of 3D-protein structures, three pepsin-resistant protein regions are qualitatively identical between species (see the 3D-structure regions colored either in orange, red or black), and only quantitative differences related to the number of peptides identified, exist. Note that two of the three pepsin-resistant protein regions surround the heme (depicted in grey color). Concomitantly, only one pepsin-resistant protein region of the bovine hemoglobin alpha-chain (the bottom alpha-helix) is not recovered from the human one.
It is interesting to note that the three-dimensional structure of hemoglobin subunits has been remarkably conserved in all vertebrates since the emergence of the ancestral molecule, which likely dates back around 600 to 700 million years. As a result, the sequence of hemoglobin is also predominantly conserved between these two species.
Inversely, the 2D-heat maps of the hemoglobin beta-chains (Figure 9D) appear globally distinct since three pepsin-resistant protein regions (colored in orange) are highlighted from the bovine beta-chain compared to only one for the human one. By comparison with the 2D-heat map of bioactive peptides (Figure 9D, upper panel) from the bovine beta-chain, only the third pepsin-resistant protein region is source of bioactive peptides. Surprisingly, the 3D-heat maps reveal that no matching between the 3D-structures of the three pepsin-resistant protein regions of the bovine hemoglobin beta-chain and the human one. In this regard, it should be noted that our search did not find any studies specifically focused on enzymatic hydrolysis of hemoglobins that already present peptidomics results combined with 3D structures. Although other papers have already been published with such results on other types of proteins [38,39], we suspect that this might be the first time for hemoglobin.
2.6. Focus on Bioactive Peptides from Bovine and Human Hemoglobin Hydrolysis
By performing enzymatic hydrolysis using pepsin on bovine and human hemoglobin, a large population of bioactive peptides was produced. Some peptides identified by UPLC-MS/MS displayed antimicrobial activities [12,14,31], hematopoietic activities [11], opioid activitys [9,10], bradykinin potentiating activity [9,12], coronary constrictor activity [40], antihypertensive activity [12], antioxidant activity [16] and anticancer [41].
The Table 1 below presents the bioactive peptide sequences identified by peptidomics after hydrolysis of bovine and human hemoglobin. These results allowed for the comparison of peptides between the two species and highlighted similarities or differences in their composition. Indeed, the majority of bioactive peptides identified in bovine were also present in human without modifications. However, some antimicrobial peptides present in bovine hemoglobin were not found as such in human hemoglobin, probably due to differences in cleavage sites or modifications of the peptide sequence. For example, the antimicrobial peptide KLLSHSL located at α99-105 became KLLSHCL in human hemoglobin. Similarly, the growth-stimulating peptide STADA located at β48-52 became STPDA in humans. These observations directly corroborate the results of the bioinformatics approach previously performed. Following these observations, the presence of several new bioactive peptides in human hemoglobin was highlighted. Although some of these peptides were already known in bovine hemoglobin, this study allowed for the discovery of new bioactive peptides in human hemoglobin such as antibacterial peptides (α37-46, PTTKTYFPHF; α36-45, FPTTKTYFPH; α137-141, TSKYR, α133-141 STVLTSKYR), opioid peptides (α137-141, TSKYR; β31-40, LVVYPWTQRF; β31-37 LVVYPWT), an ACE inhibitor (β129-135, KVVAGVA), an anticancer agent (β33-39, VVYPWTQ), and an antioxidant (α137-141, TSKYR). These peptides have never been identified in human hemoglobin before, to our knowledge. These results highlight the potential of human hemoglobin as a source of bioactive peptides useful for the food or pharmaceutical industry.

Table 1.
Bioactive peptide sequences, identified by peptidomics, resulting from hydrolysis of bovine and human hemoglobin.
Table 1.
Bioactive peptide sequences, identified by peptidomics, resulting from hydrolysis of bovine and human hemoglobin.
| Biological Activity | Position | Sequence | Monoisotopic Molecular (Da) | Bovine hemoglobin | Human hemoglobin |
|---|---|---|---|---|---|
| Antimicrobial | α34-46 | LSFPTTKTYFPHF | 1584.787 | + | - |
| α33-46 | FLSFPTTKTYFPHF | 1731.855 | + | - | |
| α37-46 | PTTKTYFPHF | 1237.602 | + | + | |
| α36-45 | FPTTKTYFPH | 1237.602 | + | + | |
| α137-141 | TSKYR | 653.339 | + | + | |
| α133-141 | STVLTSKYR | 1053.571 | + | + | |
| α99-105 | KLLSHSL | 796.470 | + | KLLSHCL | |
| α100-105 | LLSHSL | 668.375 | + | - | |
| α99-106 | KLLSHSLL | 909.554 | + | KLLSHCLL | |
| β140-145 | LAHRYH | 795.403 | + | LAHKYH | |
| Hematopoietic | α76-82 | LPGALSE | 685.354 | + | MPNALSA |
| β115-122 | RNFGKEFT | 997.487 | + | HHFGKEFT | |
| Opioid | α137-141 | TSKYR | 653.339 | + | + |
| β32-40 | VVYPWTQRF | 1194.608 | + | + | |
| β31-40 | LVVYPWTQRF | 1307.692 | + | + | |
| β31-37 | LVVYPWT | 876.464 | + | ||
| Analgesic and Potentiator of bradykinin | α129-134 | LANVST | 603.312 | + | LASVST |
| β 129-134 | QKVVAG | 600.349 | + | - | |
| dipeptidyl-peptidase Inhibitor | α130-134 | ANVST | 490.228 | + | ASVST |
| β6-10 | KAAVT | 488.285 | + | KSAVT | |
| ACE inhibition | β129-135 | KVVAGVA | 642.395 | + | + |
| Antihypertensive | α99-105 | KLLSHSL | 796.470 | + | KLLSHCL |
| Antioxidant | α137-141 | TSKYR | 653.339 | + | + |
| Bacterial growth stimulator | β48-52 | STADA | 463.180 | + | STPDA |
| anticancer | β33–39 | VVYPWTQ | 891.438 | + | + |
The study focuses on the analysis of antimicrobial peptides derived from the hydrolysis of hemoglobin. However, it is important to note that different peptide populations are obtained at different degrees of hydrolysis (DH)[16]. Generally, a peptide population has a lower molecular weight when the DH is higher. In this study, hydrolysis was carried out for three hours, corresponding to a DH of 10. To further investigate the observations, a study was conducted on all the antimicrobial peptides identified during our experiments, as well as those reported in the literature [11,13,30,41,42,43]. The results of this study are presented in the following Table 2.
Several studies have highlighted the classification of peptides resulting from the hydrolysis of hemoglobin by pepsin into different families [9]. Antimicrobial peptides derived from bovine and human hemoglobin hydrolysis have thus been grouped into four distinct families, three in the α chain and one in the β chain of hemoglobin. This discovery suggests a possible structural and functional similarity between these different families of peptides.
Table 2.
Antimicrobial peptides sequences identified by UPLC-MS/MS resulting from the hydrolysis of bovine and human hemoglobin.
Table 2.
Antimicrobial peptides sequences identified by UPLC-MS/MS resulting from the hydrolysis of bovine and human hemoglobin.
| Position | Sequence | Monoisotopic Molecular (Da) | Bovine hemoglobin | Human hemoglobin | Ref |
| The first family is situated on the N-terminal end of the α-chain, with the active portion found between residues 1 and 23. | |||||
| α1-40 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERMFLSFPTTK | 4249 | - | + | b |
| α1-33 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERMF | 3474 | - | + | b |
| α1-32 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERM VLSAADKGNVKAAWGKVGGHAAEYGAEALERM (bovine) | 3327 3257 |
+ + |
+ - |
a |
| α1 -31 | VLSPADKTNVKAAWGKVGAHAGEYGAEALER | 3196 | - | + | b |
| α 1-29 | VLSPADKTNVKAAWGKVGAHAGEYGAEAL VLSAADKGNVKAAWGKVGGHAAEYGAEAL (bovine) |
2911 2841 |
- + |
+ - |
b a |
| α1-28 | VLSAADKGNVKAAWGKVGGHAAEYGAEA | 2728 | + | - | a |
| α1-27 | VLSAADKGNVKAAWGKVGGHAAEYGAE | 2656 | + | - | a |
| α1-23 | VLSAADKGNVKAAWGKVGGHAAE | 2237 | + | + | a |
| α1 -20 | VLSPADKTNVKAAWGKVGAH | 2949 | - | + | b |
| α18-44 | VGAHAGEYGAEALERMFLSFPTTKTYF | 2994 | - | + | b |
| A second family of peptides located between residues 32 and 98, with an active sequence between residues 36 and 46. | |||||
| α32-41 | FLSFPTTKTY | 1204 | - | + | c |
| α33-46 | FLSFPTTKTYFPHF | 1731 | + | + | a/e |
| α34-46 | LSFPTTKTYFPHF | 1585 | + | + | a/e |
| α36-45 | FPTTKTYFPH | 1238 | + | + | a/e |
| α37-46 | PTTKTYFPHF | 1238 | + | + | a/e |
| α33- 98 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 7151 | + | - | a |
| α33-97 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVN | 7004 | + | - | a |
| α34-98 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 7004 | + | - | a |
| α36-97 | SFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVN | 6744 | + | - | a |
| α37-98 | PTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 6657 | + | - | a |
| α 33-83 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL | 5422 | + | - | a |
| α34-83 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL | 5274 | + | - | a |
| α33-66 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAAL | 3632 | + | - | a |
| α34-66 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAAL | 3484 | + | - | a |
| α35-56 | SFPTTKTYFPHFDLSHGSAQVK | 2495 | - | + | b |
| α35-80 | SFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDDMPNAL | 4922 | - | + | b |
| α35-77 | SFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDMP | 4624 | - | + | b |
| Position | Sequence | Monoisotopic Molecular (Da) | Bovine hemoglobin | Human hemoglobin | Ref |
| The third family is located on the c-terminal side of the α-chain. | |||||
| α110-131 | AAHLPAEFTPAVHASLDKFLAS | 2293 | + | - | a |
| α107-141 | VTLASHLPSDFTPAVHASLDKFLANVSTVLTSKYR | 3788 | + | - | a |
| α107-136 | VTLASHLPSDFTPAVHASLDKFLANVSTVL | 3152 | + | - | a |
| α107-133 | VTLASHLPSDFTPAVHASLDKFLANVS | 2838 | + | - | a |
| α133-141 | STVLTSKYR | 1055 | + | + | a/e |
| α137-141 | TSKYR | 654 | + | + | a/e |
| α99-105 | KLLSHSL (bovine) KLLSHCL |
796 813 |
+ - |
- + |
a e |
| α100-105 | LLSHSL | 668 | + | - | a |
| α99-106 | KLLSHSLL KLLSHCLL |
910 926 |
+ - |
- + |
a e |
| The last family of peptides is located in the c terminal region of the β chain of hemoglobin | |||||
| β1-30 | MLTAEEKAAVTAFWGKVKVDEVGGEALGRL (bovine) MVH LTPEEKSA VTALWGKVNVDEVGGEALG |
3176 3137 |
+ | + | a |
| β1-55 | MVHLTPEEKSAVTALWGKVNVDEVGGEALGRLLVVYPWTQRFFESFGDLSTPDAV | 6063 | - | + | d |
| β56–146 | MGNPKVKAHGKKVLGAFSDGLAHLDNLKGTFATLSELHCDKLHVDPENFRLLGNVLVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 9815 | - | + | d |
| β116-146 | LLGNVLVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 4375 | - | + | d |
| β56–72 | MGNPKVKAHGKKVLGAF | 1782 | - | + | d |
| β43-83 | RFFESFGDLSTPDAVMGNPKVKAHGKKVLGAFSDGLAHLDNLK | 4616 | - | + | b |
| β111-146 | LVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 3878 | - | + | b |
| β115-146 | LAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 3463 | - | + | b |
| β114-145 | ARNFGKEFTPVLQADFQKVVAGVANALAHRYH | 3556 | + | - | a |
| β121-145 | FTPVLQADFQKVVAGVANALAHRYH | 2753 | - | + | b |
| β126-145 | QADFQKVVAGVANALAHRYH | 2196 | + | - | b |
| β1-13 | MLTAEEKAAVTAF | 1381 | + | - | a |
| β140-145 | LAHRYH (bovine) LAHKYH |
795 767 |
+ - |
- + |
a/e e |
The first family located on the N-terminal side of the α chain of bovine hemoglobin is composed of peptides α1-32, α1-29, α1-28, α1-27, and α1-23. The active part of these peptides is therefore located between residues 1 and 23. However, most of these antimicrobial peptides have also been isolated from the hydrolysis of human hemoglobin [42]. Among these pure peptides, peptides α1-40, α1-33, α1-32, α1-31, α1-29, and α1-20 have been identified. In this region of the α chain, there are only four amino acids (AAs) that differ between the two sequences (Figure 10). This shows that the change of four AAs (Proline to Alanine, Threonine to Glycine, Alanine to Glycine, and Glycine to Alanine) does not result in a total loss of antimicrobial activity.
Figure 10.
Amino acid sequence of the 1-40 region of the α chain of bovine (black) and human (blue) hemoglobin. The red color represents the variable amino acids.
Figure 10.
Amino acid sequence of the 1-40 region of the α chain of bovine (black) and human (blue) hemoglobin. The red color represents the variable amino acids.
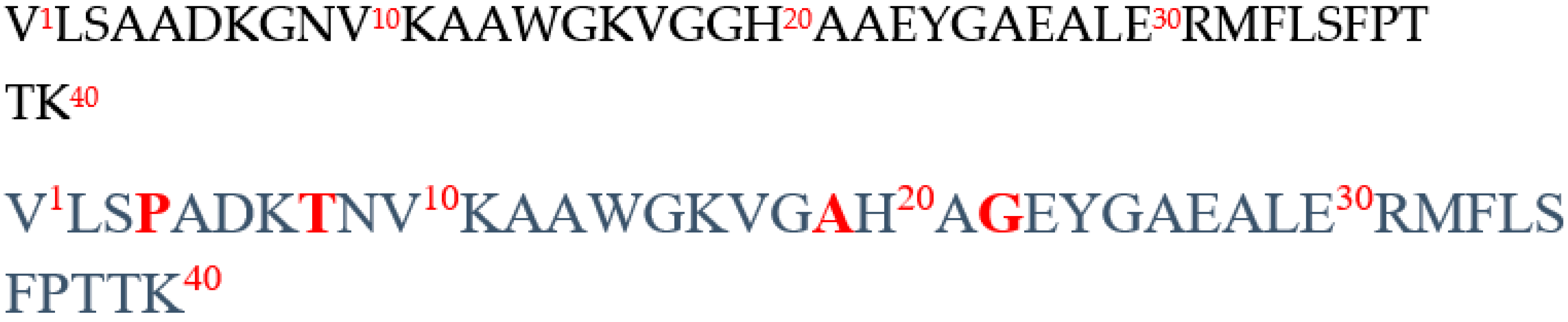
The second family located between residues 33 and 98 includes peptides α33-98, α33-97, α34-98, α36-97, α37-98, α33-83, α34-83, α33-66, α34-66 in cattle and α35-56, α35-80, α35-77 in humans. The following peptides are present in both species without modification: α33-46, α34-46, α36-45, and α37-46. The active sequence is located between residues 36 and 46. Indeed, this active sequence is present in both humans and cattle.
The last family located on the C-terminal side of the α chain is composed, on the one hand of the following peptides: α107-141, α107-136, α107-133 (bovine), α110-131 (human), α137-141, and α133-141 (human and bovine). On the other hand, peptides α99-105, α100-105, and α99-106 are listed in both species with only one AA that differs.
On the β chain side of bovine and human hemoglobin, peptide β1-13 (bovine), β43-83 (human), and β1-30 (in both species) have been identified, as well as a family of peptides located in the C-terminal region of the β chain. The first peptide forming this family corresponds to peptide β114-145, which then generates peptide β121-145, itself a precursor of peptide β126-145. Similarly, in humans, peptides β111-146 and β115-146 are part of this family, and finally, peptide β140-145 is present in both species. It is then possible to specify the active part within these peptides, which is located between residues 126 and 146.
In summary, the comparative study of the hydrolysis of human and bovine hemoglobin has made it possible to classify the vast majority of human and bovine antimicrobial peptides into the same families due to the presence of common active sites. This suggests a possible structural and functional similarity.
4. Materials and Methods
2.1. Comparison of Peptide Sequences by Bioinformatics Approach
The software used for this study is the Protein Basic Local Alignment Search Tool (BLAST®), which is freely available online at the following site: https://blast.ncbi.nlm.nih.gov/Blast.cgi.
The two proteins studied are : Bovine hemoglobin, model protein of our laboratory, the Charles Violette Institute; BioEcoAgro laboratory and Human hemoglobin.
The algorithm used by blast is the Smith Waterman algorithm, which allows a fast alignment while maintaining a good sensitivity. The reference amino acid sequences were obtained from the following database: https://www.ncbi.nlm.nih.gov/.
2.2. Prediction of Clipping Sites by Bioinformatics Approach
Peptide Cutter software from ExPASy was used for the prediction of the peptide population resulting from the pepsin hydrolysis of bovine or human hemoglobin. It is accessible free of charge by following the link: https://web.expasy.org/peptide_cutter/. A protein sequence generated by proteases can be predicted to contain possible cleavage sites using the Peptide Cutter tool. This software delivers the query sequence along with a table of the positions of potential cleavage sites that can be mapped to it.
2.3. Materials: Reagents, Solvents and Standards Used
All of the solvents and chemicals used in this study were of analytical grade and were sourced from commercial suppliers such as Sigma-Aldrich (Saint-Quentin Fallavier, France) or Flandre Chimie (Villeneuve d’Ascq, France). A Milli-Q system was used in the lab to prepare the ultrapure water.
- -
- - Purified bovine hemoglobin powder (H2625), dark brown and purified human hemoglobin (H7379), dark red, were obtained from Sigma-Aldrich. Hemoglobins were stored at 4°C before use.
- -
- - Pepsin, a lyophilized powder sourced from porcine gastric mucosa, purchased from Sigma-Aldrich (P6887). Its activity was measured at 3250 AU/mg protein using a protocol established by the supplier. To maintain the stability of the pepsin, it was stored at a -20°C.
Standard neokyotorphin (α137-141) was purchased from GeneCust (Boynes, France) and stored at -20°C before use.
2.4. Preparation of Hydrolysates
The classical enzymatic hydrolysis by pepsin in a beaker, patented by the laboratory, was used to compare bovine hemoglobin with human hemoglobin. Bovine hemoglobin is considered as the "control" since numerous enzymatic hydrolysis studies have proven it the production of active peptides and especially the peptide α137-141 with various biological activities [14,16,22,23]. All results were compared to this control.
The α137-141 peptide is a sequence of five amino acids, namely Thr-Ser-Lys-Tyr-Arg (TSKYR), located at the C-terminus of the α-chain of bovine hemoglobin. It has a monoisotopic molar mass of 653.339 Da and an isoelectric point (pI) of 10.5. It is therefore charged +2 at pH7[23].
2.4.1. Preparation of the Stock Solution
A stock solution was prepared by adding 15g of bovine hemoglobin (BH) or human hemoglobin (HH) in 100 mL of ultrapure water. The supernatant was recovered after centrifugation at 4000 min-1 for 30 min (Eppendorf AG, 22331 Hamburg, Germany; Centrifuge 5804 R, Brinkmann Instruments, Westbury, NY, USA). The Drabkin method established by Crosby, Munn and Furth (1954) was used to determine the real concentrations of BH (CBH) and HH (CHH) (1954). This is a spectrophotometric method used to quantitatively determine hemoglobin concentration in whole blood. For this purpose, 20 µL of the sample was added to 10 mL of Drabkin’s reagent D5941, Sigma-Aldrich, for 15 min at room temperature and protected from light. Then, the absorbance was then measured using a UV spectrophotometer (Chem Station UV-Vis spectrophotometer 8453A, Agilent Technologies, Santa Clara, CA, USA) at a wavelength of 540 nm. The measurements were reported on the calibration curve. From this concentration C of the bovine or human stock solution, several hemoglobin solutions were prepared by dilution to the following precise concentrations CBH and CHH: 1, 2, 8 and 10% (w/v).
2.4.2. Hydrolysis Process
Initially the stock solution of human or bovine hemoglobin was in their native, so-called "globular" form, with two alpha, beta chains and heme. It was denatured by lowering the pH to 3.5 by adding 2M HCl. It is found in an acidic environment in an "expanded" form. This will promote the attack of the alpha and beta chains by the enzyme. The hydrolysis process was carried out at a pH of 3.5 through the gradual addition of porcine pepsin (Sigma-Aldrich, - EC 3.4.23.1, 3200-4500 units mg-1 protein) that had been previously dissolved in ultrapure water. An enzyme/substrate ratio of 1/11 (mole/mole) was used. Porcine pepsin is a type of endopeptidase, which belongs to the aspartyl protease family. It selectively cleaves hydrophobic or aromatic amino acid residues in peptide bonds, as described by Dunn in 2002[24]. Samples were taken for the following hydrolysis times: 0; 2.5; 5; 15; 30; 60; 120 and 180 min, corresponding to different degrees of hydrolysis. Then the hydrolysis reaction was stopped by adding 5 M NaOH until a final pH of 9, pH which deactivates the enzyme. The pH was measured with a fine tip pH electrode (Mettler Toledo, LE422, micro, 3 mm, Leicester, UK).
Samples were stored at -20 °C to avoid basic hydrolysis. Finally, three copies were made for each species.
2.5. Determination of the Degree of Bovine/Human Hemoglobin Hydrolysis
The DH, which stands for degree of hydrolysis, is a measure of the ratio of the number of peptide bonds cleaved by an enzyme to the total number of substrate (protein) bonds. It is calculated using equation (3):
Where DH is expressed as a percentage (%), h is the number of hydrolyzed peptide bonds in the substrate, and h0 is the total number of peptide bonds in the protein substrate.
DH is quantified here by the ortho-phthaldialdehyde (OPA) technique described by Church et al. (1983)and Spellman et al. (2003) [26,27]. To determine the number h of hydrolyzed bonds, equation (4) was used :
Where ∆ Abs is the difference between the absorbance at 340 nm of the hydrolyzed sample and the absorbance of the unhydrolyzed sample, M the monoisotopic masses of the protein (for bovine hemoglobin: 64.500 Da and for human hemoglobin: 64.458 Da), DF the dilution factor of the sample, ε the molar extinction coefficient of the OPA reagent at 340 nm (6000 mol-1 -cm-1 according to Church et al.,1985[28]. CBH and CHH the concentration of bovine/human hemoglobin (10 g.L-1 equivalent to 1%).
The reagent was prepared at minimum one hour before use and stored in the dark. Its composition to obtain a final volume of 50 mL was as follows: 25 mL sodium tetraborate solution (0.1 M) (Fisher Chemicals, Belgium), 5 mL 10% (w/v) sodium dodecyl sulfate (SDS) (Fisher Scientific in Branchburg, NJ, USA), 10 μL β-mercaptoethanol (Sigma-Aldrich), and 80 mg OPA (Sigma-Aldrich) dissolved in 1 mL methanol (Fisher Chemicals, Belgium). The linearity of the reagent was checked by assaying L-leucine (Sigma-Aldrich). To perform the assay, a volume of 100 μL of test sample was added to 2 mL of reagent. After incubating for two minutes at ambient temperature, the absorbance at 340 nm was measured.
2.6. Analysis of the Peptide Hydrolysate by RP-UPLC and RP-HPLC
2.6.1. Hardware, Software and Protocol Used
Analysis by RP-UPLC
All samples were analyzed by reverse phase ultra-performance liquid chromatography (RP-UPLC) using a Watters ACQUITY UPLC H-Class PLUS (Waters Corporation, Milford, Massachusetts, USA) system adjusted to 215 nm.
The hydrolysis fractions were then injected into a C4 Colum (2.6 µm 150X2.1 mm) after filtration at 0.22 µm with polyvinylidene difluoride (PVDF) filters. The injection volume was 2.5 μL. The flow rate was set at 0.3 mL.min-1.
The elution program was as follows: The mobile phases were ultrapure water/trifluoroacetic acid (99:1, v/v) as solvent A, and acetonitrile/trifluoroacetic acid (99:1, v/v) as solvent B. A gradient was applied with solvent B increasing from 5% to 30% in 30 min, then to 60% for 10 min and held until 47min at 95%, then back to initial conditions. UV absorbance scans were performed between 200 and 390 nm at a rate of one spectrum per second with a resolution of 4.8 nm. Chromatographic acquisition and analysis were performed with Empower (Version 3 Waters) software. Each sample analysis was performed in triplicate to ensure technical reproducibility.
2.6.2. Identification of the α137-141 Peptide
Peak purity evaluation and spectral comparison as may have been described previously for the identification of hemorphinsand the α137-141 peptide, neokyotorphin in bovine cruor [15,16,29,30], an initial rapid identification of peptides from bovine and human hemoglobin peptic hydrolysate was carried out by comparison of UV spectra. The spectrum of the standard peptide was stored in the digital library of the software. Secondly, two other mathematical analyses are then performed. Firstly, a comparison of the UV spectra between the standard peptide and the peptide present in the peak is made and secondly, the purity of the peak is estimated.
For the spectral comparison, two parameters are studied. The first was the Match Angle (MA), or "angle of encounter" that was considered. The latter represented the measure of the difference in spectral patterns between the standard peptide and the peptide to be identified. The MA ranged from 0 to 90 degrees. The smaller this value was, the more similar the spectra were considered to be. Conversely, values approaching 90 degrees indicated large differences between the peptides from a spectral perspective.
The second measure was the Match Threshold (MT) parameter. It indicated the sensitivity of the measurement method, allowing quantification by an angle that ranged from 0 to 180 degrees. The larger the value, the more sensitive the method was found to be. However, if the MA was greater than the MT, it could show that the two spectra were different. On the other hand, if the MA was lower than the MT, there was a good chance that the two spectra being compared were similar or even the same [30]. Therefore, identical spectra can identify expected peptides.
A second measurement is performed to estimate the purity of a peak from the chromatographic profile in addition to the spectral comparison.
To do this, two criteria were expressed, the Purity Angle (PA), represented the relative spectral homogeneity within a peak. This value can range from 0 to 90 degrees, with 0 indicating perfect spectral homogeneity. Next, the purity threshold (PT) includes background, high sample concentration, photometric error and/or solvent. This can result in a higher PA than it actually was.
In the end, if the PA was lower than the PT, the measured spectra could be considered similar considering the inherent interferences of the method. But this does not prove chemical purity.
For each measurement, the lowest values of each of the parameters were sought, respecting the MA lower than the MT and the PA lower than the PT. In this way, the α137-141 assay was the most realistic and reproducible possible between different samples while avoiding the inclusion of other compounds or peptides in the measurements [29,31].
2.6.3. Quantification of α137-141 in Hydrolysates
A standard range was performed by injecting the standard α137-141 peptide at concentrations ranging from 0 to 5mg. mL-1 (Figure 1). The areas under the peaks were extracted from the chromatographic profiles at 215 nm. A linear relationship (r² = 1) was established between the concentration of α137-141 and the peak area of the peptide of interest within the hydrolysates, using the following equation:
where Cα137-141 is the concentration of α137-141 present in the sample (expressed in mg.L-1) and Aα137-141 being the peak area (μV.s).In addition, injection of the standard allowed identification of α137-141 at a retention time of approximately 5 min under the elution conditions described previously.
𝐶∝137-141 = 4419.6 × 𝐴∝137-141
Figure 1.
Standard Range of α137-141 by RP-UPLC assay.
2.7. Peptidomics Approach
After centrifugation for 10 min at 8,000 x g, aliquots of 10 µL of human and bovine hydrolysates, at a concentration of 30 mg.mL-1, were analyzed in triplicate by RP-HPLC-MS/MS as described in Hamed et al, 2022 with some minor changes. Peptides were separated using a C18 column (150 × 3.0 mm, 2.6 µm, Uptisphere CS EVOLUTION, Interchim, France). The mobile phases consisted of solvent A (0.1% (v/v) formic acid/99.9% (v/v) water) and solvent B (0.1% (v/v) formic acid /99.9% (v/v) acetonitrile (ACN)). The ACN gradient (flow rate 0.5 mL.min-1) was as follows: from 5% to 30% solvent B over 40, from 30% to 100% solvent B over 10 min followed by washing and equilibrating procedures using successively 100% and 1% solvent B for 5 min each. The eluate was directed into the electrospray ionization source of the qTOF Synapt G2-Si™ (Waters Corporation, Manchester, UK) previously calibrated using a sodium format solution. Mass spectrometry (MS) measures were performed in sensitivity, positive ion and data dependent analysis (DDA) modes using the proprietary Mass Lynx software (Waters). The source temperature was set at 150 °C and the capillary and cone voltages were set to 3000 and 60 V.MS data were collected for m/z values in the range of 50 and 2000 Da with a scan time of 0.2 s. A maximum of 10 precursor ions were chosen for MS/MS analysis with an intensity threshold of 10,000. MS/MS data were collected under the collision-induced dissociation (CID) fragmentation mode and a scan time of 0.1 with specified voltages ranging from 8 to 9 V and from 40 to 90 V for the lower and higher molecular mass ions, respectively. The leucin +enkephalin ([M + H]+ of 556.632) was injected in the system every 2 min for 0.5 s to follow and to correct the measure error during all the time of analysis.
Protein database searches to identify the chromatographed peptides were performed in the UniProt data bases restricted to Bos taurus and Homo sapiens (access online on July 2022) via PEAKS® Studio XPro (v 10.6) (Bioinformatics Solutions Inc., Waterloo, Canada). A mass tolerance of 35 ppm and an MS/MS tolerance of 0.2 Da were allowed. The data searches were performed with notifying pepsin as hydrolysis enzyme. The relevance of protein and peptide identities was judged according to their identification score in the research software (using a p-value< 0.05 and a false discovery rate<1%).
2.8. Two-Dimensional (2D) and Tri-Dimensional (3D) Heatmaps
For each hydrolysate, the peptide abundance all along the amino acid sequence was assessed using a home-built Microsoft Excel heat map giving the amino acid occurrences in the proteins (from the N-terminal amino acid (left) to the C-terminal amino acid (right). For each amino acid of the protein sequence, a score was calculated according to its occurrence in each unique peptide sequence identified. The higher the occurrence, the more the peptide zone tends towards red then black. The 3D structures of bovine and human hemoglobin alpha- and beta-chains were uploaded (access on March 2023) from RSCB Protein Databank (PDB) using the SWISS-MODEL bioinformatics web-server (PDBID were 1hda.1.C, 6ihx.1.D, 1bz1.1.A, 6kye.1.D for bovine alpha- and beta-chains and human alpha- and beta-chains, respectively).
2.9. RP-UPLC Statistical Analysis
Three independent replications of each condition were run, and each analysis was carried out in triplicate. One-way or two-way analyses of variance were performed on the data (ANOVA). GraphPad Prism 8.0.2(263) was used to conduct multiple t test comparisons tests on the data to identify the treatment that was statistically different from the others at a probability level of 0.05 (p < 0.05).
5. Conclusion
In conclusion, the bioinformatics approach confirmed the comparability of the two proteins, encouraging research on peptides such as the α137-141 peptide with antimicrobial and antioxidant properties. The hydrolysis of human hemoglobin follows the same enzymatic mechanism as bovine hemoglobin, allowing for rapid production of the antimicrobial peptide α137-141 with a maximum concentration reached in just 3 h, with over 63% generated in 30 minutes. Through the identification of bioactive peptides performed by peptidomics, it was observed that most of the active peptides (especially antimicrobial peptides) identified in cattle were also present in human without modification or belonged to the same family, due to the presence of common active sites. This study also revealed the presence of new bioactive peptides in human hemoglobin, including antibacterial peptides, opioid peptides, an ACE inhibitor, an anticancer agent, and an antioxidant. Some of these peptides were already known, but others had never been reported before in human hemoglobin. These results emphasize the untapped potential of human hemoglobin as a source of bioactive peptides for the food or pharmaceutical industry.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, NN., AO and BE.; methodology, NN, AO, BD, MAD., CF.; software, AO CFC.; validation, NN., AO. and BE.; formal analysis, NN., AO., CF; investigation, AO.; resources, BE., BG., NN; data curation, NN., AO., MAD., BD., writing—original draft preparation, NN., AO., BE.; writing—review and editing, NN, AO, BD., CFC., visualization, BE., BG.; NN supervision, NN., BE; project administration, BE., NN.; All authors have read and agreed to the published version of the manuscript.”.
Acknowledgments
The authors are grateful to the Advanced High Throughput Technologies Platform for Biorefineries Catalysts Design “REALCAT” funded by a French governmental subsidy managed by the French National Research Agency (ANR) within the frame of the “Future Investments program (ANR-11- EQPX-0037)”. The Hauts-de-France region and the FEDER, the Ecole Centrale de Lille and the Centrale Initiatives Foundation are also warmly acknowledged for their financial contributions to the acquisition of REALCAT platform equipment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gatnau, R.; Polo, J.; Robert, E. Plasma protein antimicrobial substitution at negligible risk, Feed Manufacturing in the Mediterranean Region. Improving Safety: From Feed to Food 2001, 54, 141–150. [Google Scholar]
- Mora, L.; Reig, M.; Toldrá, F. Bioactive peptides generated from meat industry by-products. Food Research International. 2014, 65, 344–349. [Google Scholar] [CrossRef]
- Gómez-Juárez, C.; Castellanos, R.; Ponce-Noyola, T.; Calderón, V.; Figueroa, J. Protein recovery from slaughterhouse wastes. Bioresource Technology 1999, 70, 129–133. [Google Scholar] [CrossRef]
- Bah, C.S.; Bekhit, A.E.-D.A.; Carne, A.; McConnell, M.A. Slaughterhouse blood: an emerging source of bioactive compounds. Comprehensive Reviews in Food Science and Food Safety 2013, 12, 314–331. [Google Scholar] [CrossRef]
- Lafarga, T.; Hayes, M. Bioactive peptides from meat muscle and by-products: generation, functionality and application as functional ingredients. Meat Science 2014, 98, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Aristoy, M.-C.; Mora, L.; Reig, M. Innovations in value-addition of edible meat by-products. Meat Science 2012, 92, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-H.P.; Ofori, J.A. Blood-derived products for human consumption. Revelation and Science 2011, 1. [Google Scholar]
- Sasakawa, S. Studies on Hemoglobin VI. Amino Acid Compositions of the Fractionated Bovine Globin α and β. The Journal of Biochemistry 1961, 50, 345–351. [Google Scholar] [CrossRef]
- Lignot, B.; Froidevaux, R.; Nedjar-Arroume, N.; Guillochon, D. Solvent effect on kinetics of appearance of neokyotorphin, VV-haemorphin-4 and a bradykinin-potentiating peptide in the course of peptic hydrolysis of bovine haemoglobin. Biotechnology and Applied Biochemistry 1999, 30, 201–207. [Google Scholar]
- Zhao, Q.; Molina, P.; Piot, J.M. Peptic peptide mapping by HPLC, on line with photodiode array detection, of a hemoglobin hydrolysate produced at pilot-plant scale from an ultrafiltration process. Journal of Liquid Chromatography & Related Technologies 1997, 20, 1717–1739. [Google Scholar]
- Vercaigne-Marko, D.; Kosciarz, E.; Nedjar-Arroume, N.; Guillochon, D. Improvement of Staphylococcus aureus-V8-protease hydrolysis of bovine haemoglobin by its adsorption on to a solid phase in the presence of SDS: peptide mapping and obtention of two haemopoietic peptides. Biotechnology and Applied Biochemistry 2000, 31, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Adje, E.Y. Hydrolyse ménagée de l’hémoglobine bovine par la pepsine porcine en mélanges hydroalcooliques et obtention d’une nouvelle famille de peptides antimicrobiens, Lille 1, 2010.
- Daoud, R.; Dubois, V.; Bors-Dodita, L.; Nedjar-Arroume, N.; Krier, F.; Chihib, N.-E.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. New antibacterial peptide derived from bovine hemoglobin. Peptides 2005, 26, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Nedjar-Arroume, N.; Dubois-Delval, V.; Adje, E.Y.; Traisnel, J.; Krier, F.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. Bovine hemoglobin: an attractive source of antibacterial peptides. Peptides 2008, 29, 969–977. [Google Scholar] [CrossRef]
- Choisnard, L.; Froidevaux, R.; Nedjar-Arroume, N.; Lignot, B.; Vercaigne-Marko, D.; Krier, F.; Dhulster, P.; Guillochon, D. Kinetic study of the appearance of an anti-bacterial peptide in the course of bovine haemoglobin peptic hydrolysis. Biotechnology and Applied Biochemistry 2002, 36, 187–194. [Google Scholar] [CrossRef]
- Przybylski, R.; Firdaous, L.; Châtaigné, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse by-product and its potential application on meat as preservative. Food Chemistry 2016, 211, 306–313. [Google Scholar] [CrossRef]
- de Melo, J.G.; de Sousa Araújo, T.A.; de Almeida e Castro, V.T.N.; de Vasconcelos Cabral, D.L.; Rodrigues, M.D.D.; Nascimento, S.C.D.; de Amorim, E.L.C.; De Albuquerque, U.P. Antiproliferative activity, antioxidant capacity and tannin content in plants of semi-arid northeastern Brazil. Molecules 2010, 15, 8534–8542. [Google Scholar] [CrossRef]
- Sakanaka, S.; Tachibana, Y.; Okada, Y. Preparation and antioxidant properties of extracts of Japanese persimmon leaf tea (kakinoha-cha). Food Chemistry 2005, 89, 569–575. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. VFDB 2012 update: toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Research 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- Saadi, S.; Saari, N.; Anwar, F.; Hamid, A.A.; Ghazali, H.M. Recent advances in food biopeptides: Production, biological functionalities and therapeutic applications. Biotechnology Advances 2015, 33, 80–116. [Google Scholar] [CrossRef]
- Linderstrom-Lang, K. Les phases initiales de la dégradation des protéines par les enzymes. Bulletin de La Société de Chimie Biologique 1953, 100–116. [Google Scholar]
- Takagi, H.; Shiomi, H.; Fukui, K.; Hayashi, K.; Kiso, Y.; Kitagawa, K. Isolation of a novel analgesic pentapeptide, neo-kyotorphin, from bovine brain. Life Sciences 1982, 31, 1733–1736. [Google Scholar] [CrossRef]
- Catiau, L.; Traisnel, J.; Delval-Dubois, V.; Chihib, N.-E.; Guillochon, D.; Nedjar-Arroume, N. Minimal antimicrobial peptidic sequence from hemoglobin alpha-chain: KYR. Peptides 2011, 32, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.M. Structure and mechanism of the pepsin-like family of aspartic peptidases. Chemical Reviews 2002, 102, 4431–4458. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. Journal of Dairy Science 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. Journal of Dairy Science 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. International Dairy Journal 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Church, F.C.; Porter, D.H.; Catignani, G.L.; Swaisgood, H.E. An o-phthalaldehyde spectrophotometric assay for proteinases. Analytical Biochemistry 1985, 146, 343–348. [Google Scholar] [CrossRef]
- Zhao, Q.; Sannier, F.; Piot, J.M. Kinetics of appearance of four hemorphins from bovine hemoglobin peptic hydrolysates by HPLC coupled with photodiode array detection. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology 1996, 1295, 73–80. [Google Scholar] [CrossRef]
- Zhao, Q.; Sannier, F.; Ricart, G.; Piot, J.M. A rapid detection and identification of hemorphins released from bovine hemoglobin enzymatic hydrolysis by use of HPLC coupled with photodiode array detector. Journal of Liquid Chromatography & Related Technologies 1995, 18, 93–103. [Google Scholar]
- Dubois, V.; Nedjar-Arroume, N.; Guillochon, D. Influence of pH on the appearance of active peptides in the course of peptic hydrolysis of bovine haemoglobin. Preparative Biochemistry and Biotechnology 2005, 35, 85–102. [Google Scholar] [CrossRef]
- Ivanov, V.; Andrei, T.; Karelin, A.; Mikhaleva, I.; Vaskovsky, B.V.; Sviryaev, V.L.; Nazimov, I.V. Isolation, structure and properties of endogenous peptides. Russian Journal of Bioorganic Chemistry 1992, 1271–1311. [Google Scholar]
- Piot, J.-M.; Zhao, Q.; Guillochon, D.; Ricart, G.; Thomas, D. Isolation and characterization of two opioid peptides from a bovine hemoglobin peptic hydrolysate. Biochemical and Biophysical Research Communications 1992, 189, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Abou-Diab, M.; Thibodeau, J.; Deracinois, B.; Flahaut, C.; Fliss, I.; Dhulster, P.; Nedjar, N.; Bazinet, L. Bovine Hemoglobin Enzymatic Hydrolysis by a New Ecoefficient Process—Part I: Feasibility of Electrodialysis with Bipolar Membrane and Production of Neokyotorphin (α137-141). Membranes 2020, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Cudennec, B.; Domenger, D.; Belguesmia, Y.; Flahaut, C.; Kouach, M.; Lesage, J.; Goossens, J.-F.; Dhulster, P.; Ravallec, R. Simulated GI digestion of dietary protein: Release of new bioactive peptides involved in gut hormone secretion. Food Research International 2016, 89, 382–390. [Google Scholar] [CrossRef]
- Kagawa, K.; Matsutaka, H.; Fukuhama, C.; Watanabe, Y.; Fujino, H. Globin digest, acidic protease hydrolysate, inhibits dietary hypertriglyceridemia and Val-Val-Tyr-Pro, one of its constituents, possesses most superior effect. Life Sciences 1996, 58, 1745–1755. [Google Scholar] [CrossRef]
- Sousa, R., Jr.; Lopes, G.P.; Tardioli, P.W.; Giordano, R.L.C.; Almeida, P.I.F.; Giordano, R.C. Kinetic model for whey protein hydrolysis by alcalase multipoint-immobilized on agarose gel particles. Brazilian Journal of Chemical Engineering 2004, 21, 147–153. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, M.; Zhang, J.; Zhao, D.; Sun, J. Peptidomics insights into the interplay between the pre-digestion effect of mixed starters and the digestive pattern of sausage proteins. Food Research International 2022, 162, 111963. [Google Scholar] [CrossRef]
- Albuquerque, W.; Ghezellou, P.; Lee, K.-Z.; Schneider, Q.; Gross, P.; Kessel, T.; Omokungbe, B.; Spengler, B.; Vilcinskas, A.; Zorn, H. Peptidomics as a Tool to Assess the Cleavage of Wine Haze Proteins by Peptidases from Drosophila suzukii Larvae. Biomolecules 2023, 13, 451. [Google Scholar] [CrossRef]
- Barkhudaryan, N.; Oberthuer, W.; Lottspeich, F.; Galoyan, A. Structure of hypothalamic coronaro-constrictory peptide factors. Neurochemical Research 1992, 17, 1217–1221. [Google Scholar] [CrossRef]
- Blishchenko, E.Y.; Mernenko, O.A.; Mirkina, I.I.; Satpaev, D.K.; Ivanov, V.S.; Tchikin, L.D.; Ostrovsky, A.G.; Karelin, A.A.; Ivanov, V.T. Tumor cell cytolysis mediated by valorphin, an opioid-like fragment of hemoglobin β-chain. Peptides 1997, 18, 79–85. [Google Scholar] [CrossRef]
- Mak, P.; Wójcik, K.; Wicherek, Ł.; Suder, P.; Dubin, A. Antibacterial hemoglobin peptides in human menstrual blood. Peptides 2004, 25, 1839–1847. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.A.; Jiang, H.; Tokiwa, Y.; Berova, N.; Nakanishi, K.; McCabe, D.; Zuckerman, W.; Xia, M.M.; Gabay, J.E. Broad-spectrum antimicrobial activity of hemoglobin. Bioorganic & Medicinal Chemistry 2001, 9, 377–382. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



