Submitted:
19 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
Trypanosomatids are mainly responsible for leishmaniasis, sleeping sickness, and Chagas disease, which are the most challenging among the neglected tropical diseases due to the problem of drug resistance. Although problems of target deconvolution and polypharmacology are encountered, a target-based approach is a rational method for screening drug candidates targeting a biomolecule that causes diseases. The present study aims to summarize the latest information re-garding potential inhibitors of squalene synthase and farnesyl phosphate synthase with anti-Trypanosomatidae activity. The information was obtained by referencing textbooks and major scientific databases from their inception until April 2023. Based on in vitro experiments, more than seventy compounds were reported to inhibit squalene synthase and farnesyl diphosphate synthase. Among these compounds, more than 30 were found to be active in vitro against Trypanosomatidae, inferring that these compounds can be prospected as scaffolds for the development of new drugs against trypanosomatid-related infections. Over-all, natural and synthetic products can inhibit enzymes that are crucial for the survival and virulence of trypanosomatids. Moreover, in vitro experiments have confirmed the activity of more than half of these inhibitors using cell-based as-says. Nevertheless, additional studies on the cytotoxicity, pharmacokinetics, and lead optimization of potent anti-Trypanosomatid compounds should be investi-gated.
Keywords:
Trypanosomatids
; squalene synthase
; farnesyl diphosphate synthase
; target-based approach
; neglected tropical diseases.
1. Introduction
Neglected tropical diseases (NTDs) are a group of 20 pathological conditions that are prevalent in tropical and subtropical regions, where they affect more than 1 billion people who live in impoverished communities. Most of these diseases are parasitic with high endemicity in developing countries of Africa, Asia, and the Americas [1]. Leishmaniasis and trypanosomiases are among the NTDs that occur in several sub-Saharan African countries [2]. These diseases are caused by trypanosomatid parasites that interact with a wide range of insects and mammals to complete their life cycles. In general, Trypanosoma brucei gambiense and Trypanosoma brucei rhodesiense, Trypanosoma cruzi, and Leishmania spp. are the parasites causing, human African trypanosomiasis (HAT or sleeping sickness), Chagas disease, and cutaneous, mucocutaneous, and visceral leishmaniasis, respectively. These kinetoplastid diseases are maladies of poverty that have received limited funding for the discovery, development and delivery of new tools [3]. An estimated 6 to 7 million people worldwide are infected with T. cruzi, and the disease is mostly transmitted when humans come into contact with feces and/or urine of infected blood-sucking triatomine bugs (vector-borne transmission) [4]. The number of new cases of human African trypanosomiasis (HAT or sleeping sickness) has been reduced to the lowest level (<1000 new cases) in 2020, with 60-70 million people at risk of infection in 36 sub-Saharan African countries. This disease is transmitted by infected tsetse flies [5]. Regarding leishmaniasis, transmission occurs through bites by infected female phlebotomine sandflies, leading to estimates of approximately 700 000 to 1 million new cases annually [6]. Current treatments for human African trypanosomiasis include pentamidine, suramin, melarsoprol, eflornithine and nifurtimox, whereas nifurtimox and benznidazole are indicated for the treatment of Chagas disease [4,5]. Approved medications for the treatment of leishmaniasis include intravenous liposomal amphotericin B for VL and oral miltefosine for CL, ML, and VL caused by particular species [7]. However, these drugs suffer the limitations of toxicity, variable efficacy, requirements for parenteral administration and/or length of treatment regimens, in addition to the problem of drug resistance [2]. Thus, there is an urgent need to search for safer anti-trypanosomatidae therapies. Numerous studies have recently demonstrated the effectiveness of natural products vis-à-vis trypanosomatidae at preclinical and clinical levels. Some of these studies were focused on the identification of enzymes that are crucial for the survival and virulence of trypanosomatidae. These enzymes include farnesyl diphosphate synthase and squalene synthase [8].
In previous studies, 2-alkyl aminoethyl-1,1-bisphosphonic acids [9] and quinuclidine-based compounds [10,11] were reported to inhibit squalene synthase, whereas taxodione and arenarone inhibited the enzyme farnesyl diphosphate synthase [12]. Unlike humans, these parasites (T. cruzi, T. brucei, and Leishmania spp.) use a variety of sterols (e.g., ergosterol, 24-ethyl-cholesta-5,7,22-trien-3 beta ol, and its 22-dihydro analogs) rather than cholesterol in their cell membranes, so inhibiting endogenous s terol biosynthesis is an important therapeutic target [13].
The identification of inhibitors of these crucial enzymes might afford potentially active anti-Trypanosomatidae hit compounds that can serve as starting points for the discovery of new treatments against leishmaniases and trypanosomiases. Thus, reviews or monographs are needed to summarize and discuss the latest information regarding the possibility for inhibitors of farnesyl diphosphate synthase and squalene synthase to impede the growth of Trypanosoma and Leishmania species.
To date, only a few reports [13,14] have previously been published on this topic. Therefore, the present work aims to summarize up-to-date information regarding potential inhibitors of squalene synthase and farnesyl phosphate synthase with anti-Trypanosomatidae activity that could be used as scaffolds for the development of new and safe treatments for leishmaniasis and trypanosomiases.
2. Research methods
In this study, a comprehensive literature review on inhibitors of farnesyl diphosphate synthase and squalene synthase as potential anti-Trypanosomatidae hit compounds is presented and discussed.
2.1. Literature search
Literature information was obtained from published and unpublished materials, with a focus on natural products that are potentially active against leishmaniasis and American and African trypanosomiases. Databases, such as Science Direct, PubMed (National Library of Medicine), Web of Science, SciFinder, Scopus, Wiley, American Chemical Society (ACS), Springer, and Google Scholar, as well as dissertations, theses, and textbooks, were searched from their respective inception until April 2023 to obtain the relevant data. The search terms included “Farnesyl diphosphate synthase”; “Squalene synthase”; “Farnesyl diphosphate synthase” AND “Trypanosoma spp.” OR “Squalene synthase” AND “Trypanosoma spp.” OR “Farnesyl diphosphate synthase” AND “Leishmania spp.” OR “Squalene synthase” AND “Leishmania spp.”. Moreover, dissertations, theses, books, and reports from classic literature; articles published in peer-reviewed journals; and unpublished materials related to inhibitors of squalene synthase and farnesyl diphosphate synthase and leishmaniasis and American and African trypanosomiases were also examined and searched. Reference lists of all the included reviews and other archives of the publications were hand searched for additional relevant articles. No limitation was set for languages.
2.2. Data extraction and synthesis
Potentially eligible articles were assessed in full text, irrespective of the database. Study selection and data extraction were conducted by the first author and confirmed by the other authors. The extracted data were summarized in tables, and a narrative synthesis was used to provide a summary of the results. Graphical expression was used to present the structures of active potential inhibitors of farnesyl diphosphate synthase and squalene synthase.
2.3. Results of the literature search
From the database searches, 1225 (“Squalene synthase and Trypanosoma”: 342; “Farnesyl diphosphate synthase and Trypanosoma”: 247; “Squalene synthase and Trypanosoma”: 342; “Squalene synthase and Leishmania”: 323) potentially relevant records were identified, from which 1190 were excluded after screening the titles or abstracts. Thirty-five full-length research articles were exploited to gather relevant information. In addition, data from dissertations, theses, books, reports and unpublished materials were also included. Eventually, among the inhibitors of squalene synthase and farnesyl diphosphate synthase embodied in selected documents, only those having at least micromolar inhibition, preferably in a nanomolar range, were considered in this study.
3. Human African trypanosomiasis
Human African trypanosomiasis takes two forms, depending on the subspecies of the parasite involved, and includes Trypanosoma brucei gambiense and Trypanosoma brucei rhodesiense. This disease is transmitted by the tsetse fly (Glossina species), which is found only in sub-Saharan Africa [15,16]. The subspecies T.b. gambiense is mainly responsible for mostly chronic anthroponotic infections in West and Central Africa, accounting for approximately 95% of all HAT cases [16]. Fever, extreme fatigue, severe headaches, skin rash, swollen lymph nodes, irritability, and aching muscles and joints are common symptoms of human African trypanosomiasis. Progressive confusion, personality changes, and other neurologic problems occur after infection has invaded the central nervous system [17]. Trypanosomes proliferate in the bloodstream of mammalian hosts as morphologically slender forms. As parasite numbers increase in the bloodstream, differentiation to morphologically stumpy forms occurs. These are division-arrested forms preadapted for transmission to tsetse flies. Upon uptake in a tsetse blood meal, procyclic forms are generated, which are proliferative in the fly midgut. After establishment in the fly midgut, trypanosomes arrest in division and then migrate to the tsetse salivary gland, where they attach as epimastigote forms. These cells are proliferative and attached through elaboration of their flagellum. Eventually, these generate nonproliferative metacyclic forms in preparation for transmission to a new mammalian host [18]. For treatment, pentamidine and suramin have historically been used as the drugs of choice for the treatment of blood-stage gambiense and rhodesiense forms, respectively. For the treatment of second-stage infections, drugs that cross the blood‒brain barrier are needed, and melarsoprol has been effectively used for both forms of HAT in the past. Combination therapy of eflornithine and nifurtimox has long been used as the treatment of choice to cure second-stage infections [16]. Preventive measures include wearing protective clothing, including long-sleeved shirts and pants, using mosquito repellents, and avoiding tsetse flies’ bites [5].
4. Human American trypanosomiasis
Chagas disease, American trypanosomiasis, is caused by infection with the protozoan parasite Trypanosoma cruzi, which displays a complex life cycle involving human and animal hosts as reservoirs of disease and triatomine insects as vectors [4]. Transmission to humans occurs when humans come in contact with feces and/or urine of infected blood-sucking triatomine bugs (vector-borne transmission). From their ecological biotope, triatomine bugs become active at night, defecating close to the skin area that they have bitten. Next, the bug’s waste can be inadvertently smeared into the bite or another skin break, mouth and eyes. Food contaminated with waste from infected triatomine bugs can typically infect groups of people with higher frequency and severity. After being bitten by a triatomine bug, characteristic first visible signs can be a skin lesion or a purplish swelling of the lids of one eye. In addition, they can present headache, fever, muscle pain, difficulty breathing, enlarged lymph glands, pallor, swelling, and abdominal or chest pain. During the chronic phase, the parasites are hidden mainly in the heart and digestive muscle, which may lead to destruction of the heart muscle and nervous system, consequent cardiac arrhythmias or progressive heart failure and sudden death. An estimated 6-7 million people are infected with T. cruzi worldwide annually [4]. This disease manifests itself in two phases, including the acute phase and chronic phase. During the acute phase, symptoms are absent or mild and nonspecific, although a high number of parasites are under circulation in the blood [19]. The diagnosis and treatment of Chagas disease remain challenging [20]. There is a lack of effective vaccines against T. cruzi [21]. Early treatment is critical to manage the 28,000 new cases of AT estimated to occur every year [4]. In the acute stage of the disease, nifurtimox and benznidazole (nitroimidazole compounds) are the most effective drugs used; however, in the chronic stage, their efficacy is limited, and their use is still a matter of debate [22]. In any case, the disease should be treated in the acute stage to avoid complications that may arise at the chronic phase. To prevent this disease, it is necessary to avoid receiving blood transfusions and organ transplants in areas where Chagas disease is found, sleep under a bed net treated with insecticide, and practice safe food and water precautions [23].
5. Leishmaniases
Diseases that are caused by protozoan parasites from more than 20 Leishmania species are called leishmaniases. These parasites are transmitted to humans by the bite of an infected female phlebotomine sandfly, a tiny 2-3 mm long insect vector [6]. Cutaneous (CL), mucocutaneous (MCL) and visceral leishmaniasis (VL) are the three main forms of this disease. As most infected people do not develop disease symptoms, leishmaniasis should refer to the term of becoming sick owing to Leishmania infection and not being infected with the parasite [6]. Today, more than 1 billion people live in areas endemic for leishmaniasis and are at risk of infection. An estimated 30 000 new cases of VL and more than 1 million new cases of CL occur annually [6].
In cutaneous leishmaniasis, exposed parts of the body, such as arms, face, and legs, are affected by ulcers that produce many lesions that cause serious disability. Mainly seen in the Eastern hemisphere, Old World cutaneous leishmaniasis is caused by L. donovani, L. infantum, L. major, L. tropica and L. aethiopica, whereas New World cutaneous leishmaniasis (prevalent in Central and South America) is predominantly caused by L. braziliensis, L. panamensis, L. guyanensis, L. amazonensis, L. mexicana and L. peruviana [24]. In the case of mucocutaneous leishmaniasis, the mucous membranes of the mouth, nose, and throat cavities are affected. In visceral leishmaniasis, symptoms such as fever, weight loss, anemia, and swelling of the spleen and liver are observed [6]. Currently used antileishmanial treatments include pentavalent antimonials [25], amphotericin B [26], miltefosine [27], paromomycin [28] and topical and systemic azoles [29]. Regardless of the causative Leishmania species, antileishmanial treatment cannot provide a sterile cure, and the parasite remains in the human body and can cause a relapse when there is immunosuppression. Leishmania parasites have a complex life cycle that involves both vertebrate and invertebrate hosts and two developmental stages: promastigotes, the proliferative form found in the lumen of the female sand fly, and amastigotes, the proliferative form found inside several types of mammalian host cells [30]. Early diagnosis and effective prompt treatment reduce the prevalence of the disease and prevent disabilities and death. Vector control helps to reduce or interrupt the transmission of disease by decreasing the number of sandflies. Control methods include the use of insecticide-treated nets, insecticide spray, environmental management and personal protection [6].
6. Squalene synthase and farnesyl diphosphate synthase: targets for anti-Trypanosomatidae drug discovery
Squalene synthase catalyzes the first reaction of the branch of the isoprenoid metabolic pathway committed specifically to sterol biosynthesis [31]. In Leishmania parasites, this enzyme catalyzes the dimerization of two farnesyl pyrophosphate molecules to produce squalene, which then undergoes different enzymatic reaction steps to yield ergosterol, in contrast to cholesterol in humans [32]. The basic structure of sterol molecules (such as cholesterol, ergosterol, etc.) is derived from acetyl-CoA through two successive pathways, including the mevalonate and isoprenoid pathways (Figure 1) [33]. In the mevalonate pathway, there is condensation of two acetyl-CoA units to form acetoacetyl-CoA in the presence of the cytosolic enzyme acetoacetyl-CoA thiolase. This reaction is followed by the addition of a third acetyl-CoA unit to form 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA), which is then reduced by NADPH to give mevalonic acid by using two mitochondrial enzymes in trypanosomatids, i.e., HMG-CoA synthase and HMG-CoA reductase, respectively [34]. In humans, the enzymes involved in the mevalonate pathways have been reported to occur in the cytosol and endoplasmic reticulum, especially in peroxisomes and in glycosomes [33].
The isoprenoid pathway is initiated with the conversion of mevalonate into isopentenyl diphosphate (IPP) by two phosphorylation reactions followed by one decarboxylation. Afterward, dimethylallyl diphosphate (DMAPP) is obtained from the isomerization of IPP by isopentenyl diphosphate isomerase. Then, condensation of IPP with DMAPP and geranyl diphosphate (GPP) generates longer chains of isoprenoids (15 carbons), namely, farnesyl diphosphate (FPP), in two successive reactions catalyzed by farnesyl diphosphate synthase (FPPS). Farnesyl diphosphate is the direct precursor for the synthesis of sterols, dichols, prenylated proteins, etc. It is noteworthy that the enzymes required for the isoprenoid pathway are located in the cytosol (L. major), peroxisome (animals), and mitochondria, among others [33,35]. After the isoprenoid pathway, the next two reactions comprise the first committed step in sterol biosynthesis. Next to the isoprenoid pathway, there is a condensation of two molecules of farnesyl diphosphate that generate squalene in the presence of the enzyme squalene synthase. Plausible localization of squalene synthase in T. cruzi and Leishmania spp. is either in mitochondria, microsomes, or glycosomes [33]. Afterward, squalene epoxidase catalyzes the conversion of squalene to squalene epoxide, and this reaction mostly occurs in microsomes. Next, a 2,2-oxidosqualene cyclase cyclizes the 2,3-oxidosqualene to lanosterol, the initial precursor of all steroid structures formed by trypanosomatids, mammals, etc. This reaction is followed by a series of transformations to form cholesterol in mammals and ergosterol in trypanosomatids (Figure 2) [33]. One of the most important stages of ergosterol biosynthesis that does not exist in the synthesis of cholesterol is the addition of a methyl group at the C24 position in the sterol side chain.
7. Potential inhibitors of squalene synthase
Squalene synthase is an attractive target mainly because it catalyzes the conversion of farnesyl diphosphate to squalene in the first committed step of cholesterol biosynthesis [36]. Inhibitors of squalene synthase include alkoxy-aminobenzhydrol derivatives and fungus-derived zaragozic acids, but none are in clinical use due to unfavorable toxicity profiles [37,38]. Thus, there is still a need to search for new and safe inhibitors of squalene synthase.
In the last 3 decades, numerous studies have been reported on the identification of squalene synthase inhibitors, and a total of forty hit compounds (Table 1, Figure 3) are summarized in this subsection. An example is the study by Baxter et al. [39], which described the potential of squalestatin-1 (1) as a potent selective inhibitor of squalene synthase in vitro with an IC50 value of 12 nM [39,40]. Four years later, Ward et al. [41] demonstrated that a compound [3-(biphenyl-4-yl)-quinuclidine (2)] from the quinuclidine group exhibited in vitro inhibition of squalene synthase from human and marmoset liver microsomes with Ki values of 5 nM and 1300 nM, respectively. Moreover, McTaggart et al. [42] confirmed the inhibitory potential of 3-(biphenyl-4-yl) quinuclidine (BPQ) (2) and 3-(biphenyl-4-yl)-3-hydroxyquinuclidine (BPQ-OH) (3) by reporting their apparent inhibition constants (K’i) as 12 nM and 15 nM, respectively [42]. Likewise, zaragozic acid A (4) has also been reported as a potent inhibitor (a competitive inhibitor with a Ki value of <1.6 nM) of squalene synthase [43,44,45]. 3-Hydroxy-3-[4-(quinolin-6-yl)phenyl]-1-azabicyclo[2-2-2]octane dihydrochloride (5) is a potent inhibitor of rat liver microsomal squalene synthase, with an IC50 value less than 0.9 nM. Under in vivo conditions, oral administration of this compound (3-hydroxy-3-[4-(quinolin-6-yl)phenyl]-1-azabicyclo[2-2-2]octane dihydrochloride) (5) to rats inhibited de novo [14C]cholesterol biosynthesis from [14C]mevalonate in the liver with an ED50 value of 5 mg/kg [46]. A quinuclidin derivative ((E)-2-[2-uoro-2-(quinuclidin-3-ylidene) ethoxy]-9H-carbazole monohydrochloride) (6) was reported to inhibit squalene synthase from hepatic microsomes of several species, including rats, hamster, guinea pig, rhesus monkey and HepG2 cells (IC50: 90, 170, 46, 45, and 79 nM, respectively). Under in vivo conditions, this compound equally inhibited squalene synthase activities in hepatic microsomes and suppressed cholesterol biosynthesis in rats (ED50: 32 mg/kg) [47].
Zaragozic acids and quinuclidine derivatives are well-known inhibitors of squalene synthase; however, due to their unfavorable toxicity profiles, these promising compounds have failed to enter the clinical trial phase [38]. Furthermore, computer simulations indicated strong interactions between squalene synthase and zaragozic acid with a high docking score, confirming the in vitro enzymatic results [38].
Several nitrogen- and nonnitrogen-containing bisphosphonate derivatives (BPs) were reported as inhibitors of squalene synthase. One such example includes 5-(N-[2-butenyl-3-(2-methoxyphenyl)]-N-methylamino)-1,1-penthylidenebis(phosphonic acid) trisodium salt (7) (IC50: 3.6 nM), which selectively inhibited the activity of squalene synthase. In addition, a tripivaloyloxymethyl derivative (8) of compound 7 demonstrated inhibitory potential vis-à-vis squalene synthase with an IC50 value of 39 μM [48]. However, these nitrogen-containing BPs are not metabolized and are excreted unaffected via the kidney. Notably, BPs are typically administered under fasting conditions, as food reduces their bioavailability [49].
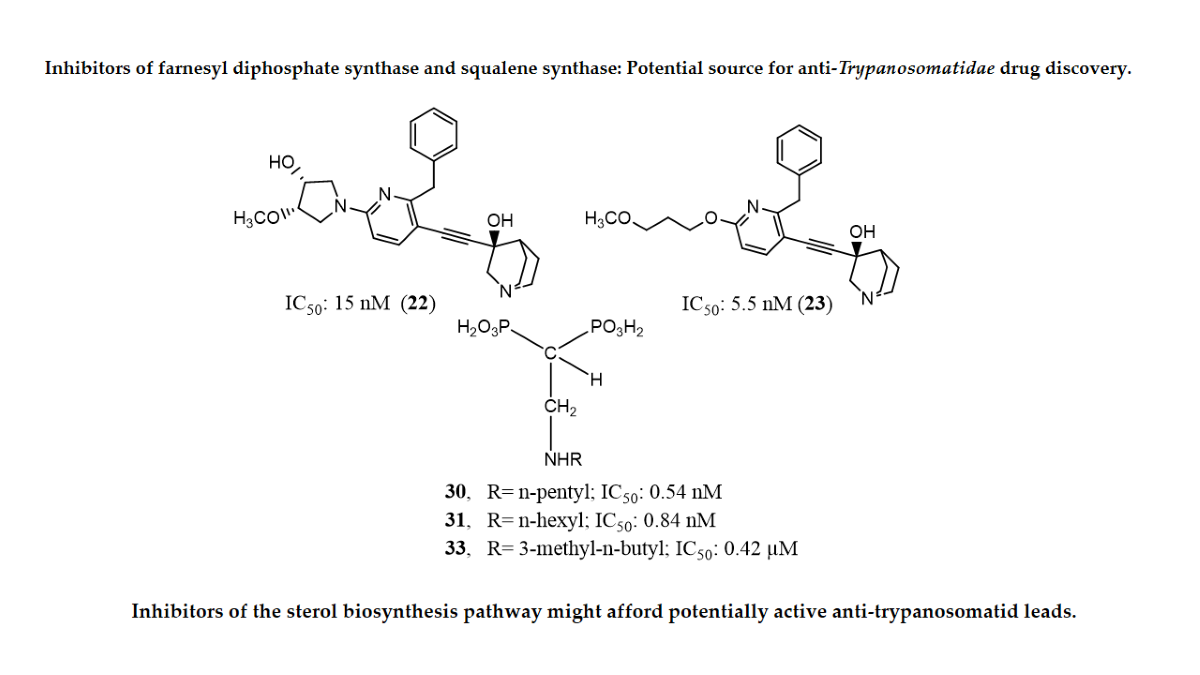
In another experiment, a series of 4,1-benzoxazepine derivatives [1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-(3-hydroxy-2,2-dimethylpropyl)-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (9); 1-[[(3R,5S)-1-(3-acetoxy-2,2-dimethylpropyl)-7-chloro5-(2,3-dimethoxyphenyl)-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (10); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(propionyloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (11); 1-[[(3R,5S)-1-[3-(butyryloxy)-2,2-dimethylpropyl]-7-chloro-5-(2,3-dimethoxyphenyl)-2-oxo-1,2,3,5-tetrahydro4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (12); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(isobutyryloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (13); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(pivaloyloxymethyloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (14)] were synthesized and evaluated for inhibition of squalene synthase and sterol synthesis in rat liver by Miki et al. [50] (Table 1). Under in vitro conditions, compounds 9-14 inhibited squalene synthase with IC50 values of 45, 76, 87, 93, 89 and 471 nM, respectively. Under in vivo conditions, the single oral administration of 10 mg/kg compounds 9-14 to [14C]acetate (50 µCi/kg)-induced Wistar rats inhibited sterol synthesis by 64, 81, 40, 30, 34 and 43%, respectively [50]. Although the exact mechanism of action of benzoxazepin derivatives has not yet been fully elucidated, they are believed to interact with multiple receptors, including serotonin, dopamine, and norepinephrine receptors [51], and to modulate the activity of certain enzymes, such as monoamine oxidase [52], or interact with certain ion channels, such as calcium and potassium channels [51,53]. Furthermore, Urbina et al. [10] verified and confirmed the T. cruzi epimastigote squalene synthase inhibitory (noncompetitive inhibitor) potential of 3-(biphenyl-4-yl)-3-hydroxyquinuclidine (3-biphenyl4-yl-1-aza-bicyclo[2.2.2]-octan-3-ol (BPQ-OH) (3) with a Ki value of 48 nM. Furthermore, in vitro anti-T. cruzi (amostigote forms) (IC50: 3 µM; SI: 33.33, Vero cells) and anti-L. mexicana (promastigotes) (IC50: 12 µM) experiments revealed low IC50 values. Although appropriate controls were not used for the assessment, the results of this study clearly indicated selective antiparasitic activity of the quinuclidine derivative BPQ-OH. A year later, Ishihara et al. [54] reported a series of five 3-ethylidenequinuclidine derivatives (compounds 15, 16, 17, 18 and 19) [(Z)-3-[2-(9H-Fluoren-2-yloxy)ethylidene]quinuclidine hydrochloride (15); (E)-3-[2-(9H-fluoren-2-yloxy)ethylidene]quinuclidine hydrochloride (16); 3-[2-(9H-fluoren-2-yloxy)ethyl]quinuclidine hydrochloride (17); (Z)-3-[2-(9H-fluoren-2-ylamino)ethylidene]quinuclidine hydrochloride (18); and (Z)-3-[2-[N-(9H-fluoren-2-yl)-N-methylamino]ethylidene] quinuclidine fumarate (19)] that inhibit squalene synthase from the hamster liver with IC50 values of 0.076, 0.15, 0.25, 0.27 and 0.56 µM, respectively. Compound 15, which was the most potent inhibitor, also demonstrated effective inhibition against human hepatoma cells (IC50: 48 nM). This research group also found that (Z)-2-[2-(quinuclidin-3-ylidene)ethoxy]-9H-carbazole hydrochloride (20) and (E)-2-[2-fluoro-2-(quinuclidin-3-ylidene)ethoxy]-9H-carbazole hydrochloride (21) inhibit squalene synthase derived from human hepatoma cells, with IC50 values of 160 and 79 nM, respectively [55]. In 2004, Urbina et al. [11] prepared two other quinuclidine-based derivatives [{(3R)-3-[[2-benzyl-6-[(3R,4S)-3-hydroxy-4-methoxypyrrolidin-1-yl]pyridin-3-yl]ethynyl]quinuclidin-3-ol monohydrate} (22) and {(3R)-3-[[2-benzyl-6-(3-methoxypropyloxy)pyridin-3-yl]ethynyl]quinuclidin-3-ol} (23)], which exhibited noncompetitive inhibition toward T. cruzi proteins with IC50 values of 5.4 and 7.2 nM, respectively, for glycosomal squalene synthase and 15 and 5.5 nM, respectively, for microsomal squalene synthase of T. cruzi epimastigotes. To verify the direct effect on the parasite in culture, further screening of compounds (22) and (23) afforded IC50 values of 10 nM and 0.4 to 1.6 nM against extracellular epimastigotes and intracellular amastigotes, respectively. Under in vivo conditions, 22 was able to provide full protection against death and completely arrested the development of parasitemia when given at a dose of 50 mg/kg of body weight for 30 days [11,56]. Other biphenyl quinuclidine derivatives (24, 25 and 26) were also evaluated for their inhibitory activity toward L. major squalene synthase. Compounds (3-(biphenyl-4-yl)-3-hydroxyquinuclidine) (24), (3-(biphenyl-4-yl)-2,3-dehydro quinuclidine) (25) and (3-(biphenyl-4-yl-40-hydroxy)-2,3-dehydro quinuclidine) (26) afforded IC50 values of 0.013, 0.243 and 0.096 µM, respectively. Against L. donovani intracellular amastigotes, compounds 24, 25 and 26 afforded ED50 values of 29.0, 74.3 and >108 µM, respectively, attesting to the enzymatic effectiveness of these compounds against L. major squalene synthase [57]. In 2006, Tavridou et al. [58] reported the inhibitory potential of two biphenyl derivatives, i.e., 2-(4-Biphenyl)-4-methyl-octahydro-1,4-benzoxazin-2-ol, hydrobromide (27) and 2-(4-biphenyl)-2-(3-nitrooxy propoxy)-4-methylmorpholine, hydrobromide (28) on squalene synthase in rabbit liver microsomes with IC50 values of 33 and 0.6 μM, respectively. In human liver microsomes, compounds 27 and 28 afforded IC50 values of 63 and 1 μM, respectively. Simvastatin (a standard squalene synthase inhibitor; IC50: 30 μM) inhibited the activity of the rabbit enzyme by 23% but had poor or no effect on the activity of the human enzyme (percent inhibition: 19% at 100 μM) [58]. Developed as cholesterol-lowering agents, two arylquinuclidine derivatives {(3R)-3-[[2-benzyl-6-(3-methoxypropyloxy)-pyridin-3-yl]ethynyl] quinuclidin-3-ol} (ER-119884) (23) and {(3R)-3-[[2-benzyl-6-[3R,4S)-3-hydroxy-4-methoxypyrrolidin-1-yl]pyridin-3-yl]ethynyl] quinuclidin-3-ol monohydrate} (22) were found to be potent noncompetitive inhibitors of native L. amazonensis squalene synthase (glycosomal SQS: Ki=6.4 and 6.9 nM for compounds 23 and 22, respectively; microsomal or mitochondrial SQS: Ki=5.5 and 14.8 nM for compounds 23 and 22, respectively). Furthermore, the antileishmanial activity of these compounds revealed IC50 values in the nanomolar range when tested against promastigotes (IC50 value: 14.7 and 1.7 nM for compounds 23 and 22, respectively) and intracellular amastigotes (IC50 value: 4.0 and 0.9 nM for compounds 23 and 22, respectively) (after 72 hours of incubation). Growth inhibition was strictly associated with the depletion of the parasite’s main endogenous sterols and the concomitant accumulation of exogenous cholesterol [59]. In addition, Rodrígues-Poveda et al. [9] synthesized a series of 2-alkyl aminoethyl-1,1-bisphosphonic acids (compounds 29-33 and 34) as potent inhibitors of T. cruzi squalene synthase with IC50 values of 39.0, 5.0, 21.4, 11.9, 22.0 and 30.0 nM for compounds 29-33 and 34, respectively. Further in vitro studies demonstrated that compounds 29-33 inhibit the growth of T. cruzi amastigotes with IC50 values of 4.8, 0.54, 0.84, 10.0 and 0.94 nM, respectively [9]. Huang et al. [60] described the in vitro squalene synthase inhibitory activity of four compounds, including compounds 35, 36, 37 and 38, with IC50 values of 1.7, 0.14, 191 and 63 nM, respectively. More recently, Macías-Alonso et al. [61] reported the inhibitory effect of an abietane-type diterpenoid (39) (carnosol; IC50: 17.6 μM) isolated from cultivated Salvia canariensis. In a computational study by Wadanambi and Mannapperuma [32], 3-O-methyl diplacol (40) strongly bound to Leishmania donovani squalene synthase with a binding energy of -9.00 kcal/mol (vs ancistrotanzanine B: -9.83 kcal/mol), inferring that this compound possesses squalene synthase inhibitory action [32].
From 1992 to 2021, a total of 40 compounds (Table 1, Figure 3) were reported to inhibit the enzyme squalene synthase in in vitro and in vivo studies. Although diseases caused by Trypanosomatidae have several treatment options, the therapy displays many problems, such as extensive toxicity, lack of efficacy, parenteral route of administration affecting compliance, high costs, and emerging drug resistance. In the last few decades, success in anti-Trypanosomatidae drug discovery has been acceptable; however, almost no further research beyond the academic results has been achieved. Indeed, various approaches (classical pharmacology, also known as phenotypic drug discovery, which is the historical basis of drug discovery, and reverse pharmacology or target-based drug discovery) have been used to identify lead compounds against the parasites responsible for leishmaniasis and trypanosomiasis [62]. However, there is room to improve anti-Trypanosomatidae drug discovery and development: validation of molecular targets, widening of the chemical space explored, increase in the predictive value of surrogate models, characterization of the effect of leishmaniasis on pharmacological properties of drugs and combinations, sustainability of the drug discovery and development process, an increase in funding and public‒private partnerships, among others [63]. It is worth noting that biochemical components localized in the parasite plasma membrane, such as proteins and sterols, play a pivotal role in the pathogenesis of Leishmania and Trypanosoma. Leishmania spp. lack the enzymes for cholesterol synthesis, and the dynamics of sterol acquisition and biosynthesis in parasite developmental stages are poorly understood [64]. The Trypanosomatidae family produces a special class of sterols, including ergosterol and other 24-methyl sterols, which are required for parasitic growth and viability but are absent from mammalian host cells [33]. These secondary metabolites are produced through a sequential process that includes pathways such as the isoprenoid pathway and sterol biosynthesis, which involve the key enzymes farnesyl diphosphate synthase and squalene synthase, respectively. As the production of sterols is crucial for the survival and virulence of Trypanosomatidae, inhibitors of squalene synthase and farnesyl diphosphate synthase can be prospected as scaffolds for the identification of anti-Trypanosomatidae compounds.
Among the inhibitors of squalene synthase, the in vitro activity of eight compounds were validated against Trypanosoma cruzi [(3-(biphenyl-4-yl)-3-hydroxyquinuclidine (3-biphenyl4-yl-1-aza-bicyclo[2.2.2]-octan-3-ol (BPQ-OH) (3); IC50: 3 µM; SI: 33.33, Vero cells, [10]; (compounds 29-33); IC50 values: 4.8, 0.54, 0.84, 10.0 and 0.94 nM, respectively; Rodrígues-Poveda et al., 2012) [9]; (compounds 22 and 23; IC50 values: 10 nM and 0.4 to 1.6 nM; extracellular epimastigotes and intracellular amastigotes, respectively, [11]; whereas the in vitro phenotypic screening of five compounds [24, 25 and 26; ED50s: 29.0, 74.3 and >108 µM, respectively; L. donovani intracellular amastigotes, [57]; compounds 22 and 23; IC50 values: 1.7 and 14.7 nM, respectively, [59] were substantiated against Leishmania spp. in culture.

Table 1.
Prospective inhibitors of squalene synthase.
| Serial number | Compound name | Model | Significant results | Reference |
|---|---|---|---|---|
| 1 | Squalestatin-1 (1) | SQS in rat liver microsomes | IC50: 12 nM | [39] |
| 2 | 3-(biphenyl-4-yl)-quinuclidine (2) | Microsomal SQS from human or marmoset liver | Ki: 5 nM and 1300 nM in microsomes prepared from human and marmoset liver, respectively | [40] |
| 3 & 4 | 3-(biphenyl-4-yl) quinuclidine (BPQ) (2) and 3-(biphenyl-4-yl)-3-hydroxyquinuclidine (BPQ-OH) (3) | T. cruzi epimastigote’s squalene synthase ; NS | Ki: 12 nM and 15 nM; Ki value: 48 nM |
[10,42] |
| 5 | 3-hydroxy-3-[4-(quinolin-6-yl)phenyl]-1-azabicyclo[2-2-2]octane dihydrochloride (5) | Rat liver microsomal squalene synthase | -In vitro studies: IC50 < 0.9 nM; -In vivo studies: Reduction of cholesterol synthesis with ED50 value of 5 mg/kg |
[46] |
| 6 | ((E)-2-[2-uoro-2-(quinuclidin-3-ylidene) ethoxy]-9H-carbazole monohydrochloride) (6) | Hepatic microsomes of rats, Hamster, guinea pig, Rhesus monkey and HepG2 cells |
In vitro studies: IC50: 90, 170, 46, 45, and 79 nM, respectively; In vivo studies: Reduction of cholesterol synthesis with ED50: 32 mg/kg |
[47] |
| 7 & 8 | 5-(N-[2-butenyl-3-(2-methoxyphenyl)]-N-methylamino)-1,1-penthylidenebis(phosphonic acid) trisodium salt (7) and tripivaloyloxymethyl derivative (8) | SQS in SD (Sprague Dawley) rat liver microsomes | IC50: 3.6 nM and 39 μM, respectively | [48] |
| 9-14 | 1-[[(3R,5S)-7-Chloro-5-(2,3-dimethoxyphenyl)-1-(3-hydroxy-2,2-dimethylpropyl)-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (9); 1-[[(3R,5S)-1-(3-Acetoxy-2,2-dimethylpropyl)-7-chloro5-(2,3-dimethoxyphenyl)-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (10); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(propionyloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (11); 1-[[(3R,5S)-1-[3-(Butyryloxy)-2,2-dimethylpropyl]-7-chloro-5-(2,3-dimethoxyphenyl)-2-oxo-1,2,3,5-tetrahydro4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (12); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(isobutyryloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (13); 1-[[(3R,5S)-7-chloro-5-(2,3-dimethoxyphenyl)-1-[2,2-dimethyl-3-(pivaloyloxymethyloxy)propyl]-2-oxo-1,2,3,5-tetrahydro-4,1-benzoxazepin-3-yl]acetyl]piperidine-4-acetic acid (14)] | Squalene synthase of rat liver | IC50 values: 45, 76, 87, 93, 89 and 471 nM, respectively. | [50] |
| 15-19 | [(Z)-3-[2-(9H-Fluoren-2-yloxy)ethylidene]quinuclidine hydrochloride (15); (E)-3-[2-(9H-fluoren-2-yloxy)ethylidene]quinuclidine hydrochloride (16); 3-[2-(9H-fluoren-2-yloxy)ethyl]quinuclidine hydrochloride (17); (Z)-3-[2-(9H-fluoren-2-ylamino)ethylidene]quinuclidine hydrochloride (18); (Z)-3-[2-[N-(9H-fluoren-2-yl)-N-methylamino]ethylidene] quinuclidine fumarate (19) | Squalene synthase from the hamster liver | IC50s: 0.076, 0.15, 0.25, 0.27 and 0.56 µM, for compounds 15-19, respectively | [54] |
| 20, 21 | (Z)-2-[2-(quinuclidin-3-ylidene)ethoxy]-9H-carbazole hydrochloride (20) and (E)-2-[2-fluoro-2-(quinuclidin-3-ylidene)ethoxy]-9H-carbazole hydrochloride (21) | Squalene synthase derived from human hepatoma cells | IC50s: 160 and 79 nM, respectively | [55] |
| 22, 23 | {(3R)-3-[[2-benzyl-6-[(3R,4S)-3-hydroxy-4-methoxypyrrolidin-1-yl]pyridin-3-yl]ethynyl]quinuclidin-3-ol monohydrate} (22) and {(3R)-3-[[2-benzyl-6-(3-methoxypropyloxy)pyridin-3-yl]ethynyl]quinuclidin-3-ol} (23) | Glycosomal and microsomal SQS of T. cruzi epimastigotes L. amazonensis squalene synthase |
-IC50 values: *glycosomal SQS: 5.4 and 7.2 nM, respectively *microsomal SQS: IC50 values of 15 and 5.5 nM, respectively; -in vivo studies: complete arrest of the development of parasitemia when compound E5700 was given at a dose of 50 mg/kg of body weight for 30 days; IC50s: -Glycosomal SQS: 6.4 and 6.9 nM for compounds 23 and 22, respectively; -Microsomal or mitochondrial SQS: 5.5 and 14.8 nM for compounds 23 and 22, respectively |
[11,56,59] |
| 24-26 | (3-(Biphenyl-4-yl)-3-hydroxyquinuclidine) (24); (3-(Biphenyl-4-yl)-2,3-dehydroquinuclidine); (25) and 5b (3-(Biphenyl-4-yl-40-hydroxy)-2,3-dehydroquinuclidine) (26) |
L. major squalene synthase | IC50s: 0.013, 0.243 and 0.096 µM, for compounds 24, 25, and 26, respectively; In vivo studies: Against L. donovani intracellular amastigotes, the following ED50 values were obtained: 29.0, 74.3 and >108 µM, respectively |
[57] |
| 27-28 | 2-(4-Biphenyl)-4-methyl-octahydro-1,4-benzoxazin-2-ol, hydrobromide (27) and 2-(4-biphenyl)-2-(3-nitrooxypropoxy)-4-methylmorpholine, hydrobromide (28) | Squalene synthase in rabbit liver microsomes | IC50s: of 33 and 0.6 μM for compounds 27 and 28, respectively | [58] |
| 29-34 | Compounds 29-33 and 34 | T. cruzi squalene synthase | IC50s: 39.0, 5.0, 21.4, 11.9, 22.0 and 30.0 nM, respectively | [9] |
| 35-38 | (R)-5-(5-(4-methoxy-3-methylphenyl)furan-2-yl)-2,2-dimethyl-2,3,5,6-tetrahydrobenzo[a]phenanthridin-4(1H)-one (35); (R)-N-(2-(2-((3-methoxybenzyl)oxy)phenyl)-1,4-dihydroquinazolin-3(2H)-yl)benzo[d][1,3]dioxole-5-carboxamide (36); (2S,3R)-8-((R)-1-(3,5-dihydroxyphenyl)ethyl)-3-hydroxy-2-methyl-2-(naphthalen-2-ylmethyl)-3,4,7,8-tetrahydro-2H,6H-pyrano[3,2-g]chromen-6-one (37) and 2-(2-(7-chloro-5-(naphthalen-1-yl)-1-neopentyl-2-oxo-1,2,3,5-tetrahydrobenzo[e][1,4]oxazepin-3-yl)acetamido)butanoic acid (38) | NS | IC50s: 1.7, 0.14, 191 and 63 nM for compounds 35, 36, 37 and 38, respectively |
[60] |
| 39 | Carnosol (39) |
NS | IC50: 17.6 μM | [61] |
SQS: squalene synthase.
8. Potential inhibitors of farnesyl diphosphate synthase
Farnesyl diphosphate synthase (FPPS) is an enzyme from the class of short chain (E)-prenyltransferases that catalyzes the condensation of two molecules of isopentenyl diphosphate (IPP, C5) with dimethylallyl diphosphate (DMAPP, C5) to generate the C15 product FPP (farnesyl pyrophosphate) [65].
According to the literature, a variety of compounds (Table 2, Figure 4) have been reported to inhibit the activity of farnesyl diphosphate synthase, an enzyme involved in several metabolic pathways, including sterol biosynthesis. Trypanosomatidae are parasite species that are responsible for leishmaniasis and trypanosomiasis in humans and strongly depend on metabolites, such as ergosterol and other proteins, for their growth and survival. It is therefore hypothesized that farnesyl diphosphate synthase is among the attractive targets for the discovery of anti-Trypanosomatidae hit compounds. In a previous study by Szajnman et al. [66], Trypanosoma cruzi farnesyl pyrophosphate synthase was used as an experimental model to assess the inhibitory potential of a series of nonnitrogen-containing bisphosphonates, including compounds 41-45, 46, 47, 48, 49 and 50 (IC50: 42.83, 1.94, 2.37, 9.36, 8.45, 5.71, 5.67, 4.54, 19.73 and 4.25 µM, respectively).
Upon incubation of compounds 41, 43, 45, and 49 with the amastigote forms of Trypanosoma cruzi, the inhibition was effective, yielding IC50 values of 21.4, 18.1, 65.8 and 22.36 µM, respectively. Against Trypanosoma brucei farnesyl pyrophosphate synthase, compounds 41-45 afforded IC50 values of 3.12, 0.66, 3.57 and 4.54 µM, respectively [66]. Nonnitrogen-containing BPs are known to induce apoptosis, as these compounds metabolize into cytotoxic, nonhydrolyzable analogs of ATP, which interfere with mitochondrial function [67,68,69,70]. Other nitrogen-containing BPs (compounds 51-60) were also reported as inhibitors of farnesyl pyrophosphate synthase. Complete growth arrest of the extracellular epimastigote form of T. cruzi (IC50: 150 µM) by the bisphosphonate risedronate (51) was attributed to the depletion of the parasite’s endogenous sterols (inhibition of ergosterol synthesis with IC50 value 65.4 µM) [71]. Likewise, two alkyl 1-amino-1,1-bisphosphonates (compounds 52 and 53) inhibited T. cruzi farnesyl pyrophosphate synthase with IC50 values of 0.382 and 3.57 µM, respectively. Further in vitro screening of compounds 52 and 53 against the amastigote forms of T. cruzi showed inhibitory effects with IC50 values of 77.0 and 72.0 µM, respectively [72]. Szajnman et al. [73] described the inhibitory effect of a series of 2-alkylaminoethyl-1,1-bisphosphonic acids (compounds 54-60) [1-[(n-But-1-ylamino)ethyl] 1,1-bisphosphonic acid (29); 1-[(n-Pent-1-ylamino)ethyl] 1,1-bisphosphonic acid (30); 1-[(n-Hex-1-ylamino)ethyl] 1,1-bisphosphonic acid (31); 1-[(n-Hept-1-ylamino)ethyl] 1,1-bisphosphonic acid (32); 1-[(n-Oct-1-ylamino)ethyl] 1,1-bisphosphonic acid (54); 1-[(3-Methyl-but-1-ylamino)ethyl] 1,1-bisphosphonic acid (33); and 1-[(tert-Butylamino)ethyl] 1,1-bisphosphonic acid (55)] against the intracellular form of T. cruzi, exhibiting IC50 values of 2.28, 1.84, 0.49, 0.058, 1.014, 0.42 and 1.21 µM, for compounds 29-32, 54, 33 and 55, respectively. These compounds were further assayed for inhibitory activity on T. cruzi farnesyl diphosphate synthase, and the results showed IC50 values of 2.28, 1.84, 0.49, 0.058, 1.014, 0.42, and 1.21 µM for compounds 29-32, 54, 33 and 55, respectively [73]. Additionally, 1-[(n-dodec-1-ylamino)ethyl] 1,1-bisphosphonic acid (56) inhibited the growth of a more clinically relevant form of Trypanosoma cruzi with an IC50 value of 0.67 μM compared to benznidazole, the positive control considered (IC50: 2.77 μM). This compound was also effective against amastigote forms of T. cruzi, exhibiting an IC50 value of 0.67 µM. The target enzyme T. cruzi farnesyl diphosphate synthase (TcFPPS) was also inhibited by this compound (IC50 value: 0.81 µM) [74]. Furthermore, Rodrígues-Poveda et al. [9] synthesized a series of 2-alkyl aminoethyl-1,1-bisphosphonic acids (compounds 29-32, 55, 57 and 34) as potent inhibitors of T. cruzi farnesyl pyrophosphate synthase with IC50 values of 2.28, 1.84, 0.49, 0.058, 1.21, 69.8, and 57.3 nM, respectively. Further in vitro studies demonstrated that compounds 29-32, 55, 57 and 34 inhibit the growth of T. cruzi amastigotes with IC50 values of 4.8, 0.54, 0.84, 10.0, 10.0, 1.39 and >10 nM, respectively [9].
Additionally, 2-(n-propylamino) ethane-1,1-diyl]bisphosphonic acid (58) and [2-(n-heptylamino)ethane-1,1-diyl]bisphosphonic acid (32) afforded IC50 values of 38.0 and 58.0 nM, respectively, when tested on T. cruzi farnesyl diphosphate synthase [75]. In a paper published by Recher et al. [76], compounds 59, 60 and 61 exhibited ED50 values of 15.8, 12.8, and 22.4 µM, respectively, when tested against amastigotes of T. cruzi. These cellular activities matched the inhibition of the enzymatic activity of the target enzyme (TcFPPS) with IC50 values of 6.4, 1.7 and 0.097 µM, respectively [76]. Moreover, Lindert et al. [77] described the inhibitory effect of a series of compounds (62-71) on farnesyl diphosphate synthase with IC50 values of 1.8, 1.9, 2.5, 7.0, 10.7, 13.7, 20.3, 21.0, 22.3 and 35.0 µM for compounds 62-71, respectively [77]. In a paper published by Liu et al. [12], the quinone methide celastrol (72) showed an inhibitory effect on Trypanosoma brucei farnesyl diphosphate synthase (FPPS) with an IC50 value ∼20 µM [12].
In 2014, Aripirala et al. [78] reported the inhibitory potential of two nitrogen-containing bisphosphonates [1-(2-hydroxy-2,2-diphosphonoethyl)-3-phenylpyridinium (73) and 3-fluoro-1-(2-hydroxy-2,2-diphosphonoethyl)-pyridinium (74) in inhibiting L. major farnesyl diphosphate synthase with Ki values of 9 and 50 µM, respectively, vs zoledronate (11 µM) [78].
Recently, Galaka et al. [79] reported the inhibitory effect of three 1-alkylaminomethyl-1,1-bisphosphonic acids (compounds 31, 32 and 56), with IC50 values of 0.49, 0.058 and 0.81 µM, respectively, against Trypanosoma brucei farnesyl pyrophosphate synthase. After incubation of compounds 31, 32 and 56 with T. cruzi, IC50 values of 0.84, 10.0 and 0.67 µM were obtained, respectively, vs benznidazole (IC50: 2.58 µM) [79].
Table 2.
Potential inhibitors of farnesyl diphosphate synthase.
| Serial number | Compound name | Model | Significant results | Reference |
|---|---|---|---|---|
| 41-50 |
Compounds 41-45, 46, 47, 48, 49 and 50 |
-Trypanosoma cruzi farnesyl pyrophosphate synthase -Trypanosoma brucei farnesyl pyrophosphate synthase |
IC50 values: 42.83, 1.94, 2.37, 9.36, 8.45, 5.71, 5.67, 4.54, 19.73 and 4.25 µM for compounds 41-45, 46, 47, 48, 49 and 50, respectively. IC50 values: 3.12, 0.66, 3.57 and 4.54 µM, for compounds 41-45, respectively |
[66] |
| 51 | Risedronate (51) | NS | IC50: 65.4 µM | [71] |
| 52-53 | Alkyl 1-amino-1,1-bisphosphonates (compounds 52 and 53) | T. cruzi farnesyl pyrophosphate synthase | IC50 values 0.382 and 3.57 µM, for compounds 10 and 11, respectively; compounds 52 and 53, respectively. | [72] |
| 54-55 | [1-[(n-But-1-ylamino)ethyl] 1,1-bisphosphonic acid (29); 1-[(n-Pent-1-ylamino)ethyl] 1,1-bisphosphonic acid (30); 1-[(n-Hex-1-ylamino)ethyl] 1,1-bisphosphonic acid (31); 1-[(n-Hept-1-ylamino)ethyl] 1,1-bisphosphonic acid (32); 1-[(n-Oct-1-ylamino)ethyl] 1,1-bisphosphonic acid (54); 1-[(3-Methyl-but-1-ylamino)ethyl] 1,1-bisphosphonic acid (33); and 1-[(tert-Butylamino)ethyl] 1,1-bisphosphonic acid (55)] | T. cruzi farnesyl diphosphate synthase | IC50s: 2.28, 1.84, 0.49, 0.058, 1.014, 0.42 and 1.21 µM, for compounds 29-32, 54, 33 and 55, respectively; |
[73] |
| 56 | 1-[(n-dodec-1-ylamino)ethyl] 1,1-bisphosphonic acid (56) | T. cruzi farnesyl diphosphate synthase | IC50: 0.67 μM, vs benznidazole (IC50: 2.77 μM) | [74] |
| 57 | 2-alkylaminoethyl-1,1-bisphosphonic acids (compounds 29-32, 55, 57 and 34) |
T. cruzi farnesyl pyrophosphate synthase | IC50 values: 4.8, 0.54, 0.84, 10.0, 10.0, 1.39 and >10 nM, for compounds 29-32, 55, 57 and 34, respectively | [9] |
| 58 | 2-(n-propylamino) ethane-1,1-diyl]bisphosphonic acid (58) and [2-(n-heptylamino)ethane-1,1-diyl]bisphosphonic acid (32) | T. cruzi farnesyl diphosphate synthase | IC50 values: 38.0 and 58.0 nM, for 58 and 32, respectively. | [75] |
| 59-61 | Compounds 59, 60 and 61 | Target enzyme TcFPPS | IC50 values: 6.4, 1.7, and 0.097 µM, for compounds 59, 60 and 61, respectively | [76] |
| 73-74 | [1-(2-hydroxy-2,2-diphosphonoethyl)-3-phenylpyridinium (73) and 3-fluoro-1-(2-hydroxy-2,2-diphosphonoethyl)-pyridinium (74) | L. major farnesyl diphosphate synthase | Ki values: 9 and 50 µM for compounds 73 and 74, respectively, vs zoledronate (11 µM) | [78] |
| 3 | N4,N4’-bis(3-(4,5-dihydro-1H-imidazol-2-yl)phenyl)-[1,1’-biphenyl]-4,4’-dicarboxamide (62); N1,N3-bis(4-(4,5-dihydro-1H-imidazol-2-yl)phenyl)isophthalamide (63); 2-amino-N1-(4-(4-methyl-4,5-dihydro-1l4,3l2-imidazol-2-yl)phenyl)-N4-(4-(4-methyl-4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (64); N1,N4-bis(2-(4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (65); N1,N4-bis(3-carbamoyl-5-(4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (66); N1,N4-bis(3-(4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (67); 2-amino-N1,N4-bis(4-(4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (68); 2-amino-N1,N4-bis(3-(4,5-dihydro-1H-imidazol-2-yl)phenyl)terephthalamide (69); 2-amino-N1,N3-bis(4-(4,5-dihydro-1H-imidazol-2-yl)phenyl)isophthalamide (70); N1,N4-bis(4-(4,5-dihydro-1H-imidazol-2-yl)-3-hydroxyphenyl)terephthalamide (71). | Human FPPS | IC50: 1.8, 1.9, 2.5, 7.0, 10.7, 13.7, 20.3, 21.0, 22.3 and 35.0 µM, for compounds 62-71, respectively | [77] |
| 5 | Quinone methide celastrol (72) | Trypanosoma brucei farnesyl diphosphate synthase (FPPS) | IC50 value ∼20 µM | [12] |
| 11 | Compounds 31, 32 and 56 | Trypanosoma brucei farnesyl pyrophosphate synthase | IC50 values: 0.49, 0.058 and 0.81 µM for compounds 31, 32 and 56, respectively | [79] |
NS: Not Specified.
As already discussed, farnesyl diphosphate synthase is one of the crucial enzymes involved in the isoprenoid pathway, which is required for sterol biosynthesis. In the present study, forty (40) inhibitors of farnesyl diphosphate synthase were recorded across the literature. Among these compounds, nineteen (19) were reported to be active against Trypanosoma cruzi and included compounds 41, 43, 45 and 49 (IC50 values: 21.4, 18.1, 65.8 and 22.36 µM, respectively; amastigote forms of T. cruzi, [66]); compounds 52 and 53 (IC50 values of 77.0 and 72.0 µM, respectively, [72]; compounds 29-32, 54, 33, and 55, IC50 values: 2.28, 1.84, 0.49, 0.058, 1.014, 0.42 and 1.21 µM, respectively, [73]; 1-[(n-dodec-1-ylamino)ethyl] 1,1-bisphosphonic acid (56); IC50: 0.67 μM; clinically relevant form of T. cruzi, [74]; compounds 29-32, 55, 57 and 34 (IC50s: 4.8, 0.54, 0.84, 10.0, 10.0, 1.39 and >10 nM, respectively; T. cruzi amastigotes, [9]; compounds 59, 60 and 61 (ED50s : 15.8, 12.8, and 22.4 µM, respectively; T. cruzi amastigotes, [76]; and compounds 31, 32 and 56 (IC50s: 0.84, 10.0 and 0.67 µM, respectively; T. cruzi, [79]). This observation clearly verifies the target-based antiparasitic drug discovery approach, in which the activity of a compound can be screened by targeting enzymes that are crucial for the survival of the parasite.
9. Critical assessment and discussion
The present study aimed to emphasize the gaps in our knowledge on the following: (i) the use of squalene synthase and farnesyl diphosphate synthase as potential target proteins for the discovery of new anti-Trypanosomatidae treatments and (ii) the issues thus far not suitably explored.
As already discussed, a total of forty compounds were reported to inhibit squalene synthase. Additionally, forty compounds (40) were found to inhibit the activity of farnesyl diphosphate synthase. Notably, among the inhibitors of squalene synthase, eight compounds (3, 22, 23, and 29-33) were found to be active against Leishmania spp., whereas five compounds (22, 23, 24, 25 and 26) exhibited anti-T. cruzi activity. For the inhibitors of farnesyl diphosphate synthase, nineteen compounds [41, 43, 45 and 49; 52 and 53; 29-32, 54, 55, 57, 33, 34, 59, 60, 61 and 56] displayed anti-Trypanosoma cruzi activity.
However, the following research gaps were noted: (i) lack of appropriate controls (negative and positive) in experiments; (ii) in most of the papers, the enzymatic assays were not followed by phenotypic screening of the compounds for anti-Trypanosomatid activity; (iii) there was a lack of data regarding the cytotoxicity of bioactive compounds against human mammalian cells; and (iv) noteworthy, data regarding enzymatic assays and phenotypic screening in vivo are scarce.
As already discussed, reverse pharmacology or target-based drug discovery has been used to identify lead compounds against the parasites responsible for leishmaniasis and trypanosomiasis [62]. However, there is room to improve anti-Trypanosomatidae drug discovery and development: validation of molecular targets, widening of the chemical space explored, increase in the predictive value of surrogate models, characterization of the effect of leishmaniasis on pharmacological properties of drugs and combinations, sustainability of the drug discovery and development process, an increase in funding and public‒private partnerships, among others [63]. Although rational design of multitargeting agents is extremely complex in polypharmacology modeling, this concept could be useful in drug discovery as it involves interaction of drug molecules with multiple targets, which may interfere with a single or multiple disease pathways [80]. In addition to these activities, toxicity studies and pharmacokinetics of the most promising compounds should be investigated. Phenotypic screening of inhibitors of squalene synthase and farnesyl diphosphate synthase should be performed for anti-trypanosomatid activity against Leishmania and Trypanosoma species to verify the activity on the parasite. In vitro and in vivo toxicity studies of the bioactive compounds should be carried out to evaluate their selectivity.
10. Authors’ opinion on the topic
Neglected tropical diseases, such as leishmaniases and trypanosomiases, are prevalent in several sub-Saharan African and South American countries [2]. These diseases are caused by trypanosomatid parasites that interact with a wide range of insects and mammals to complete their life cycles. In general, Trypanosoma and Leishmania species are the parasites causing trypanosomiases and leishmaniases, respectively. A number of approaches, including phenotypic drug discovery and target-based drug discovery, have been used to identify anti-Trypanosomatidae compounds. We have summarized and discussed inhibitors of squalene synthase and farnesyl diphosphate synthase in this manuscript. Notably, a number of inhibitors of these enzymes exhibited moderate to high anti-trypanosomatid activity upon in vitro phenotypic screening, attesting to the involvement of squalene synthase and farnesyl diphosphate synthase in the pathogenesis of trypanosomatids. Indeed, squalene synthase and farnesyl diphosphate synthase are the main enzymes that intervene in the isoprenoid pathway and biosynthesis of ergosterol and other sterol compounds in trypanosomatids. The five best inhibitors of squalene synthase that displayed superior anti-trypanosomatid activity included compounds 30, 31 and 33 (IC50 values: 0.54, 0.84 and 0.94 nM, respectively; [9]) and compounds 22 and 23 (IC50s: 1.7 and 14.7 nM, respectively; [59]). The five best inhibitors of farnesyl diphosphate synthase that exhibited higher anti-trypanosomatid activity included compound 56; IC50: 0.67 μM [74]; 30 and 31 (IC50s: 0.54 and 0.84 nM, respectively [9]; and 31 and 56 (IC50s: 0.84 and 0.67 µM, respectively) [79]. More importantly, three compounds that showed anti-Trypanosomatidae activity were identified as inhibitors of both farnesyl diphosphate synthase and squalene synthase and included compounds 30, 31 and 33. Based on these observations, squalene synthase and farnesyl diphosphate synthase are potential targets for the identification of hit compounds that can serve as scaffolds for the discovery of anti-Trypanosomatid drugs. However, more studies are needed to potentially verify this target-based drug discovery to traditional phenotypic drug screening, as almost half of the reported squalene synthase and farnesyl diphosphate synthase inhibitors have not been tested on trypanosomatids. Thus, doors are opened for researchers who work on anti-trypanosomatid drug discovery to screen such inhibitors. More in vitro and in vivo toxicity studies, pharmacokinetics, and structural medications of the most prominent scaffolds are desired to improve and select the best potential hit candidates that can be pursued further for anti-trypanosomatid drug discovery. Nevertheless, three broad approaches are used for drug discovery against trypanosomatids: (i) target-based approaches that involve screening for inhibitors against a purified protein, for example, an enzyme. Compounds identified through the screening (or structure-based) process are subsequently optimized to show efficacy in a cellular model; ii) phenotypic approaches involve screening for growth inhibitors directly against an intact parasite, usually in an in vitro culture; (iii) compound repositioning, which is the redeployment of compounds previously developed for an alternative indication as anti-trypanosomatid therapies [81, 82].
11. Conclusions and future perspectives
Addressing tropical diseases, such as leishmaniasis and trypanosomiases, requires cross-sectoral approaches for drug discovery that span from phenotypic-based screening to target-based screening. Moreover, the pathogenic mechanisms used by trypanosomatids are incompletely understood, and a better understanding could speed both vaccine development and new drug discovery. We hypothesized that dynamic changes in sterol composition during parasite development are crucial for the virulence of the Leishmania and Trypanosoma parasites that are the most infectious for mammalian hosts, such as humans. Two key enzymes that are involved in the biosynthesis of cholesterol in humans and ergosterol in trypanosomatids include farnesyl diphosphate synthase and squalene synthase. In fact, these enzymes are involved in the isoprenoid pathway and sterol biosynthesis, respectively, to generate cholesterol in humans and ergosterol in trypanosomatids. Notably, the latter compound (ergosterol) is also essential for parasite survival. Thus, inhibitors of these enzymes (farnesyl diphosphate synthase and squalene synthase), which contribute to ergosterol biosynthesis, can be used as potential targets for the development of anti-trypanosomatid drugs.
In fact, the present study aimed to summarize recent developments in anti-Trypanosomatidae hit compounds from a target-based approach using farnesyl diphosphate synthase and squalene synthase as the target enzymes. Validation of the anti-trypanosomatid activity of farnesyl diphosphate synthase and squalene synthase inhibitors using phenotypic screening against trypanosomatids is also highlighted. Thus, there is no denying that inhibitors of farnesyl diphosphate and squalene synthases can afford potentially active compounds against Leishmania and Trypanosoma species. However, more in vitro and in vivo phenotypic screenings are needed to verify the potential activity of these inhibitors against Leishmania and Trypanosoma species. More toxicity studies and pharmacokinetics and structural modifications are recommended to identify the most active inhibitors as starting points for the discovery of new anti-trypanosomatid drugs.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
No.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization (WHO). The neglected tropical diseases. FactSheets. 2022. Available online: https://www.who.int/health-topics/neglected-tropical-diseases#tab=tab_1 (accessed on 26 April 2023).
- Ochola, E.A.; Karanja, D.M.S.; Elliott, S.J. The impact of Neglected Tropical Diseases (NTDs) on health and wellbeing in sub-Saharan Africa (SSA): A case study of Kenya. PLoS Negl. Trop. Dis. 2021, 15, 1–19. [Google Scholar] [CrossRef]
- Hotez, P.J.; Kamath, A. Neglected tropical diseases in subsaharan Africa: review of their prevalence, distribution, and disease burden. PLoS Negl. Trop. Dis. 2009, 3, 1–10. [Google Scholar] [CrossRef]
- World Health Organization (WHO). 2023a. The fact sheets. Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 22 March 2023).
- World Health Organization (WHO). 2023b. The Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 22 March 2023).
- World Health Organization (WHO), 2023c. The Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 22 March 2023).
- Aronson, N.; Herwaldt, B.L.; Libman, M.; Pearson, R.; Lopez-Velez, R.; Weina, P.; Carvalho, E.; Ephros, M.; Jeronimo, S.; Magill, A. Diagnosis and treatment of leishmaniasis: Clinical practice guidelines by the infectious diseases society of America (IDSA) and the American Society of Tropical Medicine and Hygiene (ASTMH). Am. J. Trop. Med. Hyg. 2017, 96, 24–45. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Basu, S.; Zhang, K. Farnesyl pyrophosphate synthase is essential for the promastigote and amastigote stages in Leishmania major. Mol. Biochem. Parasitol. 2019, 230, 8–15. [Google Scholar] [CrossRef]
- Rodrígues-Poveda, C.A.; González-Pacanowska, D.; Szajnman, S.H.; Rodríguez, J.B. 2-alkylaminoethyl-1,1-bisphosphonic acids are potent inhibitors of the enzymatic activity of Trypanosoma cruzi squalene synthase. Antimicrob. Agents Chemother. 2012, 56, 4483–4486. [Google Scholar] [CrossRef]
- Urbina, J.A.; Concepcion, J.L.; Rangel, S.; Visbal, G.; Lira, R. Squalene synthase as a chemotherapeutic target in Trypanosoma cruzi and Leishmania mexicana. Mol. Biochem. Parasitol. 2002, 125, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Urbina, J.A.; Concepcion, J.L.; Caldera, A.; Payares, G.; Sanoja, C.; Otomo, T.; Hiyoshi, H. In vitro and in vivo activities of E5700 and ER-119884, two novel orally active squalene synthase inhibitors, against Trypanosoma cruzi. Antimicrob. Agents Chemother. 2004, 48, 2379–2387. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lindert, S.; Zhu, W.; Wang, K.; McCammon, J.A.; Oldfield, E. Taxodione and arenarone inhibit farnesyl diphosphate synthase by binding to the isopentenyl diphosphate site. Proc. Natl. Acad. Sci. U S A. 2014, 111, E2530–E2539. [Google Scholar] [CrossRef]
- Shang, N.; Li, Q.; Ko, T.P.; Chan, H.-C.; Li, J.; Zheng, Y.; Huang, C.-H.; Ren, F.; Chen, C.-C.; Zhu, Z.; Galizzi, M.; Li, Z.-H.; Rodrigues-Poveda, C.A.; Gonzalez-Pacanowska, D.; Veiga-Santos, P.; Ulisses de Carvalho, T.M.; de Souza, W.; Urbina, J.A.; Wang, A.H.-J.; Docampo, R.; Li, K.; Liu, Y.-L.; Oldfield, E.; Guo, R.-T. Squalene synthase as a target for Chagas disease therapeutics. PLoS Pathog. 2014, 10, 1–16. [Google Scholar] [CrossRef]
- Sun, S.; McKenna, C.E. Farnesyl pyrophosphate synthase modulators: a patent review (2006 - 2010). Expert Opin. Ther. Pat. 2011, 21, 1433–1451. [Google Scholar] [CrossRef]
- Elenga, V.A.; Lissom, A.; Elion, D.O.A.; Vouvoungui, J.C.; Djontu, J.C.; Boumpoutou, R.K.; Gabriel Ahombo, G.; Ntoumi, F. Risk factors and prevalence of human African trypanosomiasis in individuals living in remote areas of the republic of Congo. BMC Public Health. 2022, 22, 1–9. [Google Scholar] [CrossRef]
- Álvarez-Rodríguez, A.; Jin, B.K.; Radwanska, M.; Magez, S. Recent progress in diagnosis and treatment of Human African Trypanosomiasis has made the elimination of this disease a realistic target by 2030. Front Med (Lausanne). Front. Med. 2022, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bouteille, B.; Buguet, A. The detection and treatment of human African trypanosomiasis. Res. Rep. Trop. Med. 2012, 3, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.R. The developmental cell biology of Trypanosoma brucei. J. Cell Sci. 2005, 118 Pt 2 Pt 2, 283–290. [Google Scholar] [CrossRef]
- Martinez, S.J.; Romano, P.S.; Engman, D.M. Precision health for Chagas disease: Integrating parasite and host factors to predict outcome of infection and response to therapy. Front. Cell Infect. Microbiol. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Balouz, V.; Agüero, F.; Buscaglia, C.A. Chagas disease diagnostic applications: present knowledge and future steps. Adv Parasitol. 2017, 97, 1–45. [Google Scholar]
- Bivona, A.E.; Alberti, A.S.; Cerny, N.; Trinitario, S.N.; Malchiodi, E.L. Chagas disease vaccine design: the search for an efficient Trypanosoma cruzi immune-mediated control. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Estani, S.; Colantonio, L.; Segura, E.L. Therapy of Chagas disease: implications for levels of prevention. J. Trop. Med. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhou, X.N. Preventing the transmission of American trypanosomiasis and its spread into nonendemic countries. Infect. Dis. Poverty 2015, 4, 1–11. [Google Scholar] [CrossRef]
- Madusanka, R.K.; Silva, H.; Karunaweera, N.D. Treatment of cutaneous leishmaniasis and insights into species-specific responses: A narrative review. Infect. Dis. Ther. 2022, 11, 695–711. [Google Scholar] [CrossRef]
- Arevalo, J.; Ramirez, L.; Adaui, V.; Zimic, M.; Tulliano, G.; Miranda-Verástegui, C.; Lazo, M.; Loayza-Muro, R.; De Doncker, S.; Maurer, A.; Chappuis, F.; Dujardin, J.-C.; Llanos-Cuentas, A. Influence of Leishmania (Viannia) species on the response to antimonial treatment in patients with American tegumentary leishmaniasis. J. Infect. Dis. 2007, 195, 1846–1851. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Jaya, J. Liposomal amphotericin B and leishmaniasis: dose and response. J. Glob. Infect. Dis. 2010, 2, 159–166. [Google Scholar] [CrossRef]
- Tahir, M.; Bashir, U.; Hafeez, J.; Ghafoor, R. Safety and efficacy of miltefosine in cutaneous leishmaniasis: an open label, noncomparative study from Balochistan. Pak. J. Med. Sci. 2019, 35, 495–499. [Google Scholar] [CrossRef]
- Asilian, A.; Davami, M. Comparison between the efficacy of photodynamic therapy and topical paromomycin in the treatment of Old World cutaneous leishmaniasis: a placebo-controlled, randomized clinical trial. Clin. Exp. Dermatol. 2006, 31, 634–637. [Google Scholar] [CrossRef] [PubMed]
- Galvão, E.L.; Rabello, A.; Cota, G.F. Efficacy of azole therapy for tegumentary leishmaniasis: a systematic review and meta-analysis. PLoS One 2017, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, D.E.; Benchimol, M.; Rodrigues, J.C.; Crepaldi, P.H.; Pimenta, P.F.; de Souza, W. The cell biology of Leishmania: how to teach using animations. PLoS Pathog. 2013, 9, 1–4. [Google Scholar] [CrossRef]
- Tansey, T.R.; Shechter, I. Squalene synthase: structure and regulation. Prog. Nucleic Acid Res. Mol. Biol. 2001, 65, 157–195. [Google Scholar]
- Wadanambi, P.M.; Mannapperuma, U. Computational study to discover potent phytochemical inhibitors against drug target, squalene synthase from Leishmania donovani. Heliyon 2021, 7, 1–14. [Google Scholar] [CrossRef]
- de Souza, W.; Rodrigues, J.C. Sterol biosynthesis pathway as target for anti-trypanosomatid drugs. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 1–19. [Google Scholar] [CrossRef]
- Carrero-Lérida, J.; Pérez-Moreno, G.; Castillo-Acosta, V.M.; Ruiz-Pérez, L.M.; González-Pacanowskda, D. Intracellular location of the early steps of the isoprenoid biosynthetic pathway in the trypanosomatids Leishmania major and Trypanosoma brucei. Int. J. Parasitol. 2009, 39, 307–314. [Google Scholar] [CrossRef]
- Ortiz-Gómez, A.; Jiménez, C.; Estévez, A.M.; Carrero-Lérida, J.; Ruiz-Pérez, L.M.; González-Pacanowska, D. Farnesyl diphosphate synthase is a cytosolic enzyme in Leishmania major promastigotes and its overexpression confers resistance to risedronate. Eukaryotic Cell. 2006, 5, 1057–1064. [Google Scholar] [CrossRef]
- Ishihara, T.; Kakuta, H.; Moritani, H.; Ugawa, T.; Yanagisawa, I. Synthesis and biological evaluation of novel propylamine derivatives as orally active squalene synthase inhibitors. Bioorg. Med. Chem. 2004, 12, 5899–5908. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, M.; Ohtsuka, M.; Ohki, H.; Ota, M.; Haginoya, N.; Itoh, M.; Shibata, Y.; Sugita, K.; Ishigai, Y.; Terayama, K.; Kanda, A.; Usui, H. Discovery of DF-461, a potent squalene synthase inhibitor. ACS Med. Chem. Lett. 2013, 4, 932–936. [Google Scholar]
- Pospiech, M.; Owens, S.E.; Miller, D.J.; Austin-Muttitt, K.; Mullins, J.G.L.; Cronin, J.G.; Allemann, R.K.; Sheldon, I.M. Bisphosphonate inhibitors of squalene synthase protect cells against cholesterol-dependent cytolysins. FASEB J. 2021, 35, e21640. [Google Scholar] [CrossRef]
- Baxter, A.; Fitzgerald, B.J.; Hutson, J.L.; McCarthy, A.D.; Motteram, J.M.; Ross, B.C.; Sapra, M.; Snowden, M.A.; Watson, N.S.; Williams, R.J.; Wright, C. Squalestatin 1, a potent inhibitor of squalene synthase, which lowers serum cholesterol in vivo. J. Biol. Chem. 1992, 17, 11705–11708. [Google Scholar] [CrossRef]
- Wentzinger, L.F.; Bach, T.J.; Hartmann, M.A. Inhibition of squalene synthase and squalene epoxidase in tobacco cells triggers an upregulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase. Plant Physiol. 2002, 130, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.H.; Holdgate, G.A.; Freeman, S.; McTaggart, F.; Girdwood, P.A.; Davidson, R.G.; Mallion, K.B.; Brown, G.R.; Eakin, M.A. Inhibition of squalene synthase in vitro by 3-(biphenyl-4-yl)-quinuclidine. Biochem. Pharmacol. 1996, 51, 1489–1501. [Google Scholar]
- McTaggart, F.; Brown, G.R.; Davidson, R.G.; Freeman, S.; Holdgate, G.A.; Mallion, K.B.; Mirrlees, D.J.; Smith, G.J.; Ward, W.H. Inhibition of squalene synthase of rat liver by novel 3’ substituted quinuclidines. Biochem. Pharmacol. 1996, 51, 1477–1487. [Google Scholar] [CrossRef]
- Bergstrom, J.D.; Dufresne, C.; Bills, G.F.; Nallin-Omstead, M.; Byrne, K. Discovery, biosynthesis, and mechanism of action of the zaragozic acids: potent inhibitors of squalene synthase. Annu. Rev. Microbiol. 1995, 49, 607–639. [Google Scholar] [CrossRef]
- Shechter, I.; Guan, G.; Boettcher, B.R. 2.09 – Squalene synthase. Reference Module in Chemistry, Molecular Sciences and Chemical Engineering. Comprehensive Natural Products Chemistry. 1999, 2, 245–266. [Google Scholar]
- Bedi, M.; Niesen, M.; Lopez, D. Inhibition of squalene synthase upregulates PCSK9 expression in rat liver. Arch. Biochem. Biophys. 2008, 470, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Amin, D.; Rutledge, R.Z.; Needle, S.N.; Galczenski, H.F.; Neuenschwander, K.; Scotese, A.C.; Maguire, M.P.; Bush, R.C.; Hele, D.J.; Bilder, G.E.; Perrone, M.H. RPR 107393, a Potent Squalene Synthase Inhibitor and Orally Effective Cholesterol-Lowering Agent: Comparison with Inhibitors of HMG-CoA Reductase. J. Pharmacol. Exp. Ther. 1997, 281, 746–752. [Google Scholar] [PubMed]
- Ugawa, T.; Kakuta, H.; Moritani, H.; Matsuda, K.; Ishihara, T.; Yamaguchi, M.; Naganuma, S.; Lizumi, Y.; Shikama, H. A novel squalene synthase inhibitor, reduces plasma cholesterol and triglyceride levels in several animal species. Br. J. Pharmacol. 2000, 131, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Hiyoshi, H.; Yanagimachi, M.; Ito, M.; Ohtsuka, I.; Yoshida, I.; Saeki, T.; Tanaka, H. Effect of ER-27856, a novel squalene synthase inhibitor, on plasma cholesterol in rhesus monkeys: comparison with 3-hydroxy-3-methylglutaryl-coa reductase inhibitors. J. Lipid Res. 2000, 41, 1136–1144. [Google Scholar] [CrossRef]
- Gong, L.; Altman, R.B.; Klein, T.E. Bisphosphonates pathway. Pharmacogenet. Genomics 2011, 21, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Kori, M.; Mabuchi, H.; Tozawa, R.; Nishimoto, T.; Sugiyama, Y.; Teshima, K.; Yukimasa, H. Synthesis of novel 4,1-benzoxazepine derivatives as squalene synthase inhibitors and their inhibition of cholesterol synthesis. J. Med. Chem. 2002, 45, 4571–4580. [Google Scholar] [CrossRef]
- Bailey, S.J.; Toth, M. Variability in the benzodiazepine response of serotonin 5-HT1A receptor null mice displaying anxiety-like phenotype: evidence for genetic modifiers in the 5-HT-mediated regulation of GABAA receptors. J. Neurosci. 2004, 24, 6343–6351. [Google Scholar] [CrossRef]
- Jaka, O.; Iturria, I.; van der Toorn, M.; de Mendoza, J.H.; Latino, D.A.R.S.; Alzualde, A.; Peitsch, M.C.; Hoeng, J.; Koshibu, K. Effects of natural monoamine oxidase inhibitors on anxiety-like behavior in zebrafish. Front. Pharmacol. 2012, 12, 1–13. [Google Scholar] [CrossRef]
- Scala, R.; Maqoud, F.; Antonacci, M.; Dibenedetto, J.R.; Perrone, M.G.; Scilimati, A.; Castillo, K.; Latorre, R.; Conte, D.; Bendahhou, S.; Tricarico, D. Bisphosphonates Targeting Ion Channels and Musculoskeletal Effects. Front. Pharmacol. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Ishihara, T.; Kakuta, H.; Moritani, H.; Ugawa, T.; Sakamoto, S.; Tsukamoto, S.; Yanagisawa, I. Syntheses and biological evaluation of novel quinuclidine derivatives as squalene synthase inhibitors. Bioorg. Med. Chem. 2003, 11, 2403–2414. [Google Scholar] [CrossRef]
- Ishihara, T.; Kakuta, H.; Moritani, H.; Ugawa, T.; Sakamoto, S.; Tsukamoto, Si.; Yanagisawa, I. Syntheses of 3-ethylidenequinuclidine derivatives as squalene synthase inhibitors. Part 2: enzyme inhibition and effects on plasma lipid levels. Bioorg. Med. Chem. 2003, 11, 3735–3745. [Google Scholar] [CrossRef]
- Sealey-Cardona, M.; Cammerer, S.; Jones, S.; Ruiz-Pérez, L.M.; Brun, R.; Gilbert, I.H.; Urbina, J.A.; González-Pacanowska, D. Kinetic characterization of squalene synthase from Trypanosoma cruzi: selective inhibition by quinuclidine derivatives. Antimicrob. Agents Chemother. 2007, 51, 2123–2129. [Google Scholar] [CrossRef] [PubMed]
- Orenes Lorente, S.; Gómez, R.; Jiménez, C.; Cammerer, S.; Yardley, V.; de Luca-Fradley, K.; Croft, S,L. ; Ruiz Perez, L.M.; Urbina, J.; Gonzalez Pacanowska, D.; Gilbert, I.H. Biphenylquinuclidines as inhibitors of squalene synthase and growth of parasitic protozoa. Bioorg. Med. Chem. 2005, 13, 3519–3529. [Google Scholar] [CrossRef] [PubMed]
- Tavridou, A.; Kaklamanis, L.; Megaritis, G.; Kourounakis, A.P.; Papalois, A.; Roukounas, D.; Rekka, E.A.; Kourounakis, P.N.; Charalambous, A.; Manolopoulos, V.G. Pharmacological characterization in vitro of EP2306 and EP2302, potent inhibitors of squalene synthase and lipid biosynthesis. Eur. J. Pharmacol. 2006, 535, 34–42. [Google Scholar] [CrossRef]
- Fernandes Rodrigues, J.C.; Concepcion, J.L.; Rodrigues, C.; Caldera, A.; Urbina, J.A.; de Souza, W. In vitro activities of ER-119884 and E5700, two potent squalene synthase inhibitors, against Leishmania amazonensis: antiproliferative, biochemical, and ultrastructural effects. Antimicrob. Agents Chemother. 2008, 52, 4098–4114. [Google Scholar] [CrossRef]
- Huang, H.; Chu. C.L.; Chen, L.; Shui, D. Evaluation of potential inhibitors of squalene synthase based on virtual screening and in vitro studies. Comput. Biol. Chem. 2019, 80, 390–397. [Google Scholar] [CrossRef]
- Macías-Alonso, M.; Andrés, L.S.; Córdova-Guerrero, I.; Estolano-Cobián, A.; Díaz-Rubio, L.; Marrero, J.G. Inhibition of squalene synthase of rat liver by abietane diterpenes derivatives. Nat. Prod. Res. 2021, 35, 2972–2976. [Google Scholar] [CrossRef]
- Lage, O.M.; Ramos, M.C.; Calisto, R.; Almeida, E.; Vasconcelos, V.; Vicente, F. Current screening methodologies in drug discovery for selected human diseases. Mar. Drugs 2018, 16, 1–31. [Google Scholar] [CrossRef]
- Olías-Molero, A.I.; de la Fuente, C.; Cuquerella, M.; Torrado, J.J.; Alunda, J.M. Antileishmanial drug discovery and development: Time to reset the model? Microorganisms 2021, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Wilson, M.E. Dynamics of sterol synthesis during development of Leishmania spp. parasites to their virulent form. Parasit. Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef]
- Picard, M.È.; Nisole, A.; Béliveau, C.; Sen, S.; Barbar, A.; Shi, R.; Cusson, M. Structural characterization of a lepidopteran type-II farnesyl diphosphate synthase from the spruce budworm, Choristoneura fumiferana: Implications for inhibitor design. Insect. Biochem. Mol. Biol. 2018, 92, 84–92. [Google Scholar] [CrossRef]
- Szajnman, S.H.; Montalvetti, A.; Wang, Y.; Docampo, R.; Rodriguez, J.B. Bisphosphonates derived from fatty acids are potent inhibitors of Trypanosoma cruzi farnesyl pyrophosphate synthase. Bioorg. Med. Chem. Lett. 2003, 13, 3231–3235. [Google Scholar] [CrossRef]
- McCloskey, E.; Paterson, A.H.; Powles, T.; Kanis, J.A. 2021. Clodronate. Bone. 2021, 143, 1–23. [Google Scholar]
- Panagiotakou, A.; Yavropoulou, M.; Nasiri-Ansari, N.; Makras, P.; Basdra, E.K.; Papavassiliou, A.G.; Kassi, E.N. Extraskeletal effects of bisphosphonates. Metabolism 2020, 110, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Lehenkari, P.P.; Kellinsalmi, M.; Napankangas, J.P.; Ylitalo, K.V.; Monkkonen, J.; Rogers, M.J.; Azhayev, A.; Vaananen, K.; Hassinen, I.E. Further insight into mechanism of action of clodronate: inhibition of mitochondrial ADP/ATP translocase by a nonhydrolyzable, adenine-containing metabolite. Mol. Pharmacol. 2002, 61, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Malwal, S.R.; O’Dowd, B.; Feng, X.; Turhanen, P.; Shin, C.; Yao, J.; Kim, B.K.; Baig, N.; Zhou, T.; Bansal, S.; Khade, R.L.; Zhang, Y.; Oldfield, E. Bisphosphonate-generated ATP-analogs inhibit cell signaling pathways. J. Am. Chem. Soc. 2018, 140, 7568–7578. [Google Scholar] [CrossRef]
- Garzoni, L.R.; Caldera, A.; Meirelles, M.de.N.; de Castro, S.L.; Docampo, R.; Meints, G.A.; Oldfield, E.; Urbina, J.A. Selective in vitro effects of the farnesyl pyrophosphate synthase inhibitor risedronate on Trypanosoma cruzi. Int. J. Antimicrob. Agents 2004, 23, 273–285. [Google Scholar] [CrossRef]
- Szajnman, S.H.; Ravaschino, E.L.; Docampo, R.; Rodriguez, J.B. Synthesis and biological evaluation of 1-amino-1,1-bisphosphonates derived from fatty acids against Trypanosoma cruzi targeting farnesyl pyrophosphate synthase. Bioorg. Med. Chem. Lett. 2005, 15, 4685–4690. [Google Scholar] [CrossRef]
- Szajnman, S.H.; García Liñares, G.E.; Li, Z.H.; Jiang, C.; Galizzi, M.; Bontempi, E.J.; Ferella, M.; Moreno, S.N.; Docampo, R.; Rodriguez, J.B. Synthesis and biological evaluation of 2-alkylaminoethyl-1,1-bisphosphonic acids against Trypanosoma cruzi and Toxoplasma gondii targeting farnesyl diphosphate synthase. Bioorg. Med. Chem. 2008, 16, 3283–3290. [Google Scholar] [CrossRef]
- Rosso, V.S.; Szajnman, S.H.; Malayil, L.; Galizzi, M.; Moreno, S.N.J.; Docampo, R.; Rodriguez, J.B. Synthesis and biological evaluation of new 2-alkylaminoethyl-1,1-bisphosphonic acids against Trypanosoma cruzi and Toxoplasma gondii targeting farnesyl diphosphate synthase. Bioorg. Med. Chem. 2011, 19, 2211–2217. [Google Scholar] [CrossRef]
- Aripirala, S.; Szajnman, S.H.; Jakoncic, J.; Rodriguez, J.B.; Docampo, R.; Gabelli, S.B.; Amzel, L.M. Design, synthesis, calorimetry, and crystallographic analysis of 2-alkylaminoethyl-1,1-bisphosphonates as inhibitors of Trypanosoma cruzi farnesyl diphosphate synthase. J. Med. Chem. 2012, 55, 6445–6454. [Google Scholar] [CrossRef] [PubMed]
- Recher, M.; Barboza, A.P.; Li, Z.H.; Galizzi, M.; Ferrer-Casal, M.; Szajnman, S.H.; Docampo, R.; Moreno, S.N.; Rodriguez, J.B. Design, synthesis and biological evaluation of sulfur-containing 1,1-bisphosphonic acids as antiparasitic agents. Eur. J. Med. Chem. 2013, 60, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Lindert, S.; Zhu, W.; Liu, Y.L.; Pang, R.; Oldfield, E.; McCammon, J.A. Farnesyl diphosphate synthase inhibitors from in silico screening. Chem. Biol. Drug Des. 2013, 81, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Aripirala, S.; Gonzalez-Pacanowska, D.; Oldfield, E.; Kaiser, M.; Amzel, L.M.; Gabelli, S.B. Structural and thermodynamic basis of the inhibition of Leishmania major farnesyl diphosphate synthase by nitrogen-containing bisphosphonates. Acta Crystallogr. D Biol. Crystallogr. 2014, 70 (Pt 3), 802–810. [Google Scholar] [CrossRef]
- Galaka, T.; Falcone, B.N.; Li, C.; Szajnman, S.H.; Moreno, S.N.J.; Docampo, R.; Rodriguez, J.B. Synthesis and biological evaluation of 1-alkylaminomethyl-1,1-bisphosphonic acids against Trypanosoma cruzi and Toxoplasma gondii. Bioorg. Med. Chem. 2019, 27, 3663–3673. [Google Scholar] [CrossRef]
- Reddy, A.S.; Zhang, S. Polypharmacology: drug discovery for the future. Expert Rev. Clin. Pharmacol. 2013, 6, 41–47. [Google Scholar] [CrossRef]
- Boran, A.D.; Iyengar, R. Systems approaches to polypharmacology and drug discovery. Curr. Opin. Drug Discov. Devel. 2010, 13, 297–309. [Google Scholar]
- Field, M.C.; Horn, D.; Fairlamb, A.H.; Ferguson, M.A.J.; Gray, D.W.; Read, K.D.; Rycker, M.D.; Torrie, L.S.; Wyatt, P.G.; Wyllie, S.; Gilbert, I.H. Antitrypanosomatid drug discovery: an ongoing challenge and a continuing need. Nat. Rev. Microbiol. 2017, 15, 217–231. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The mevalonate pathway to obtain mevalonic acid from acetyl-SCoA.
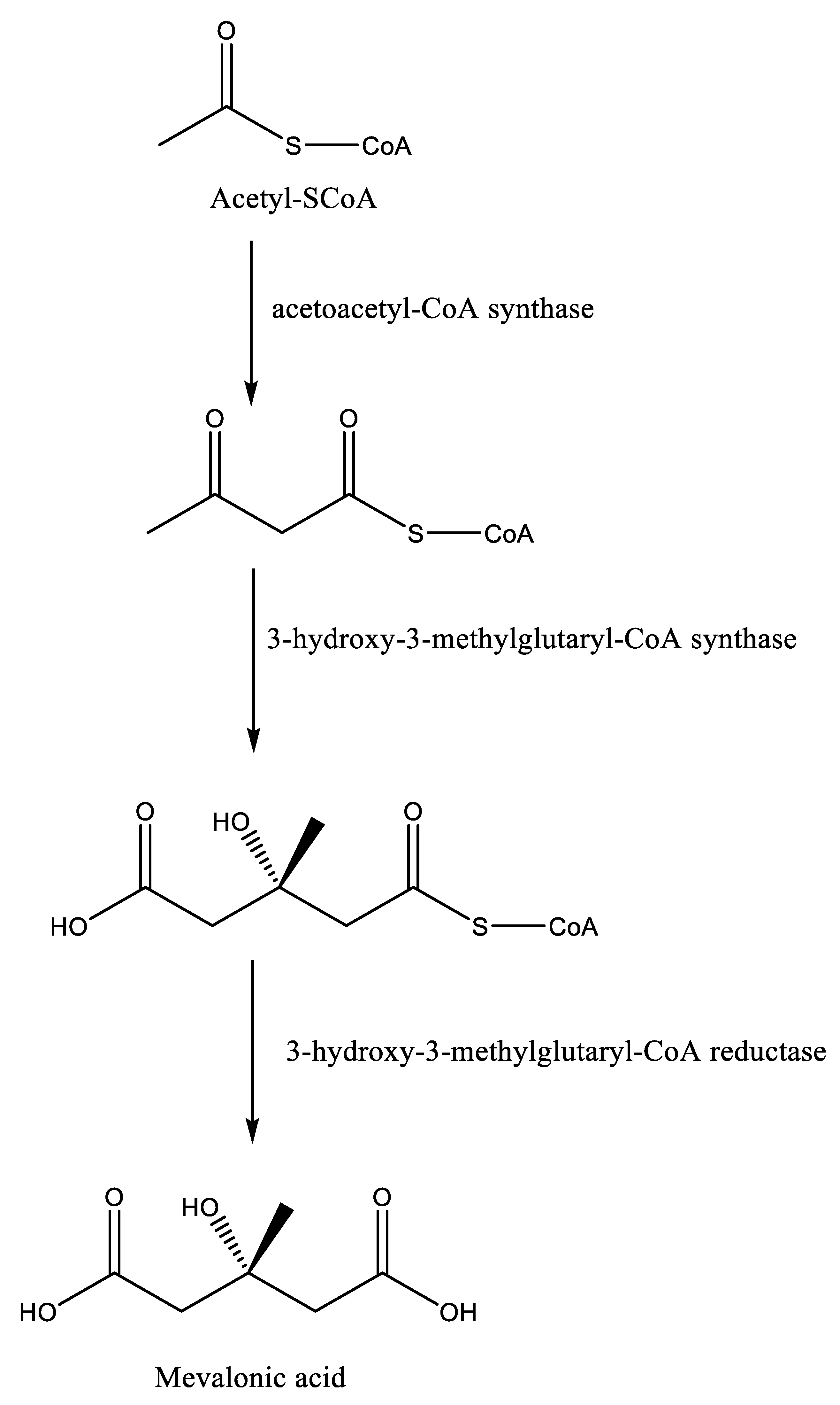
Figure 2.
Biosynthesis of cholesterol/ergosterol from mevalonic acid (isoprenoid and squalene pathways).
Figure 2.
Biosynthesis of cholesterol/ergosterol from mevalonic acid (isoprenoid and squalene pathways).
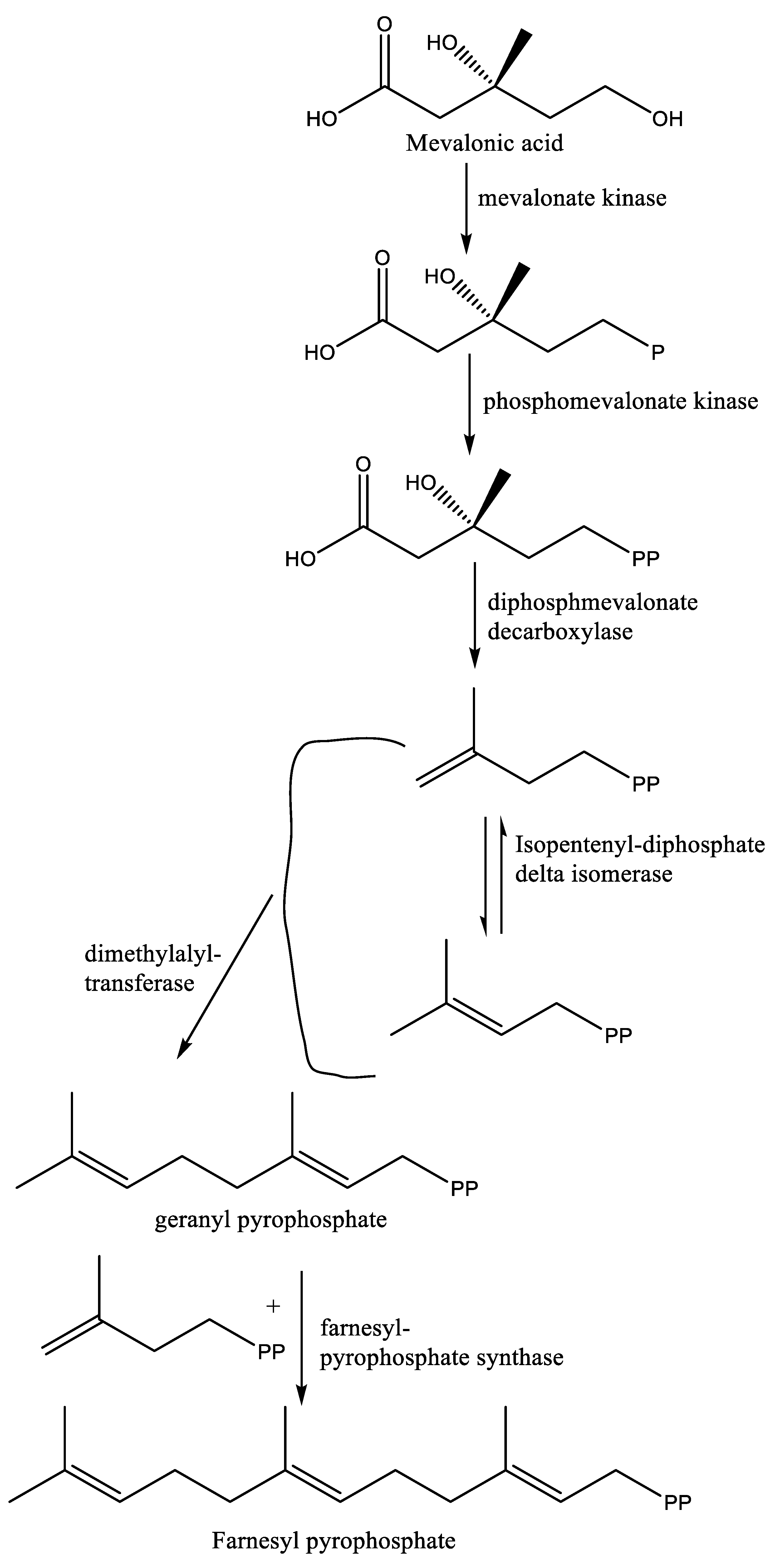
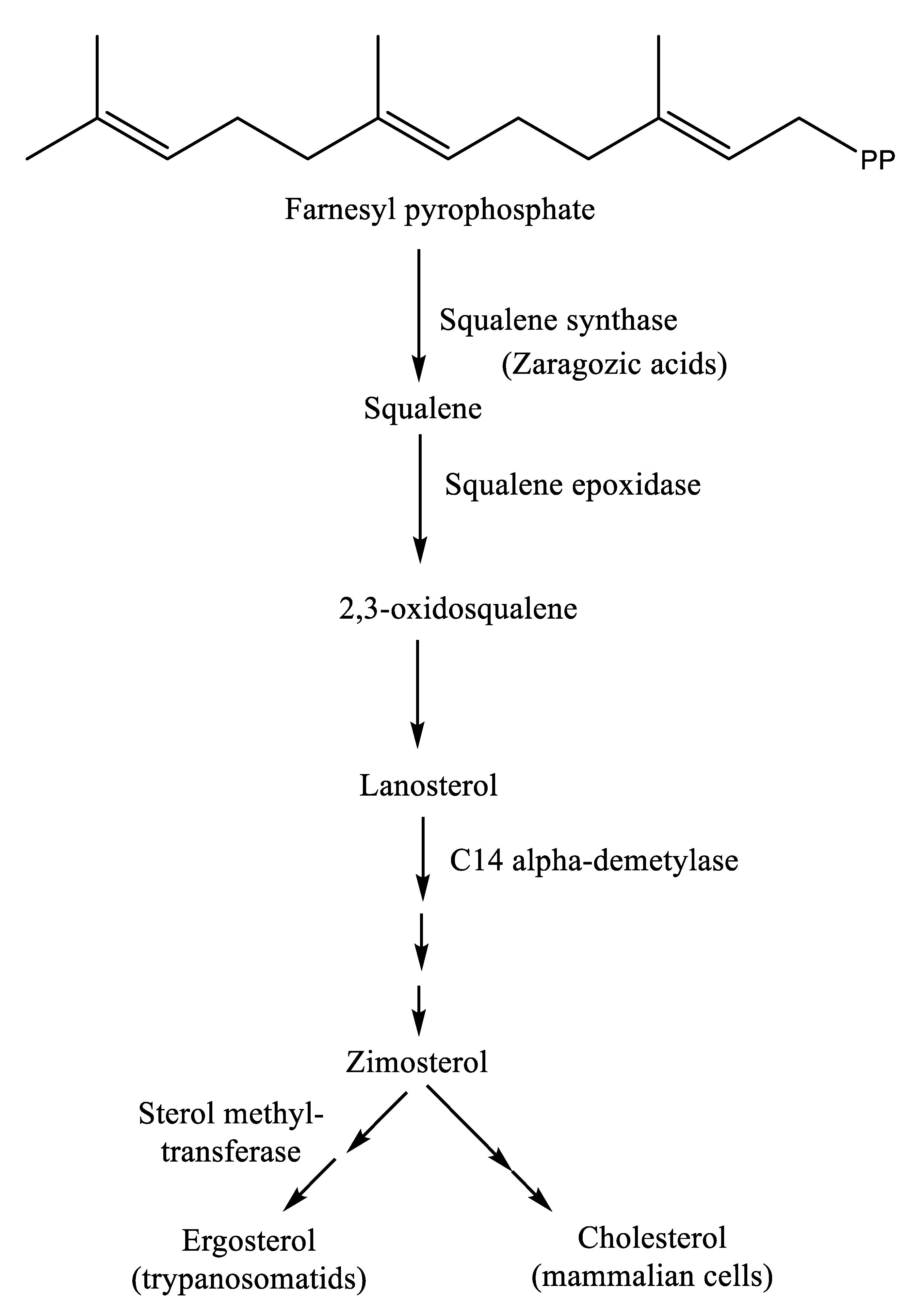
Figure 3.
Potential inhibitors of squalene synthase.
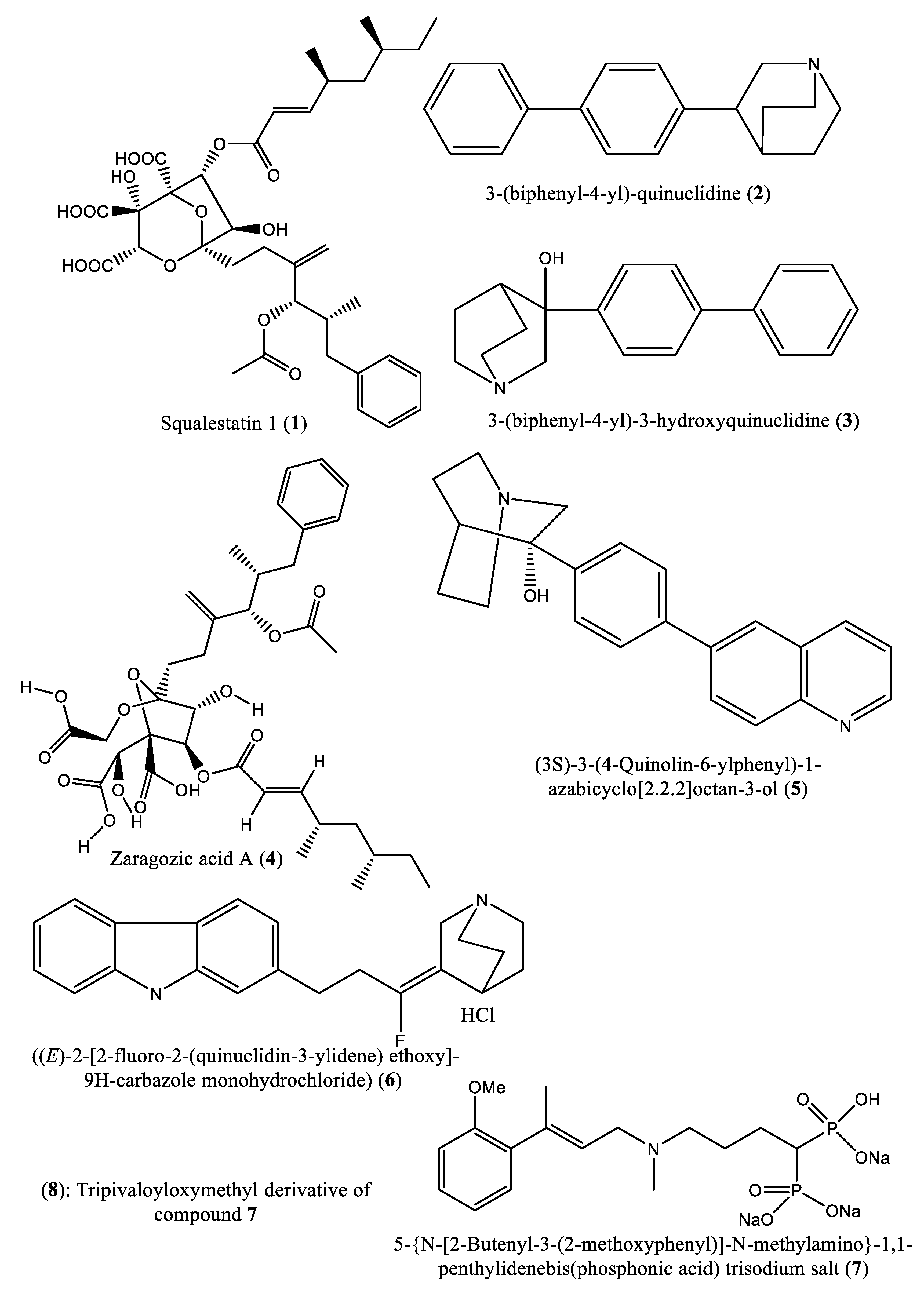
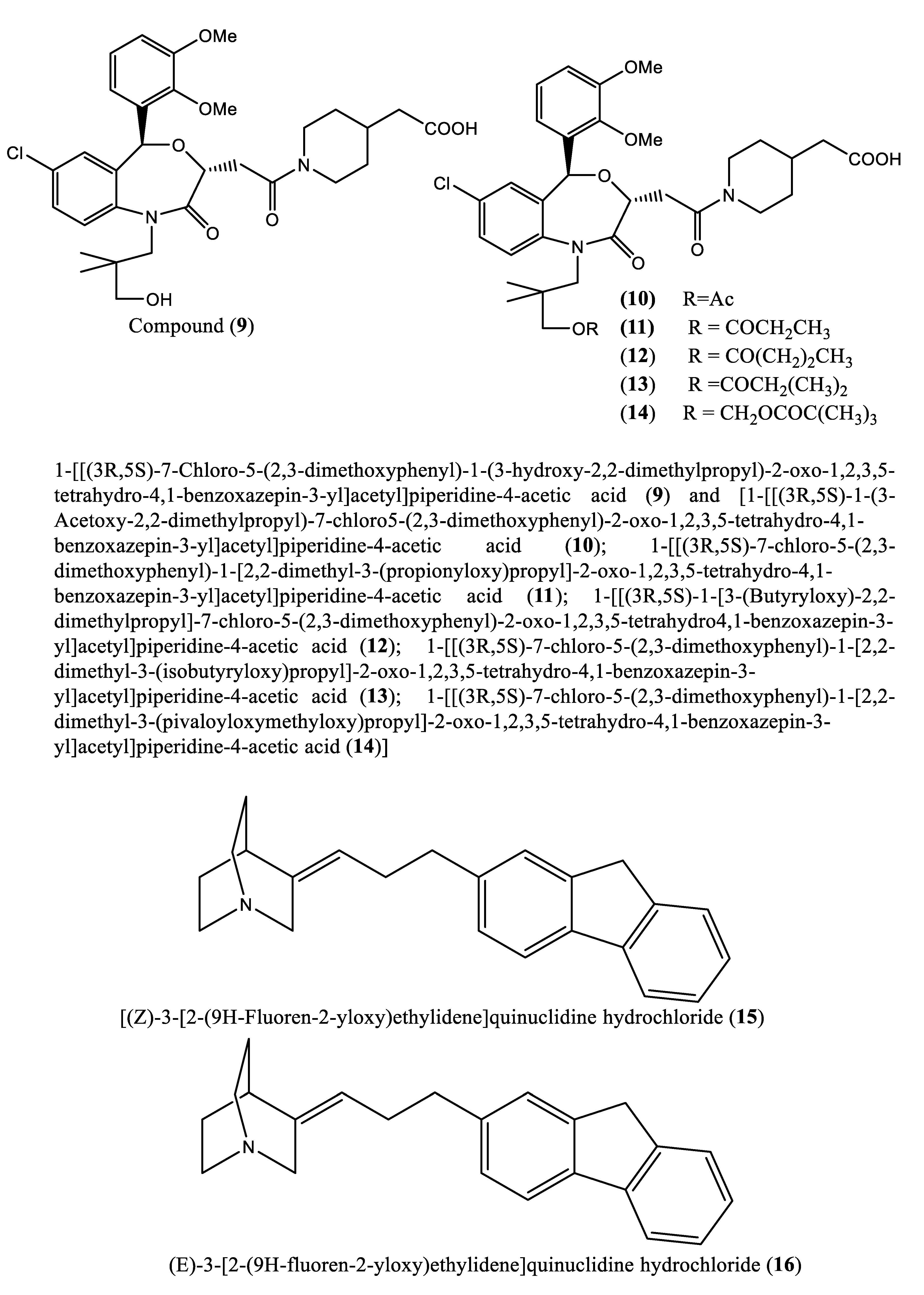
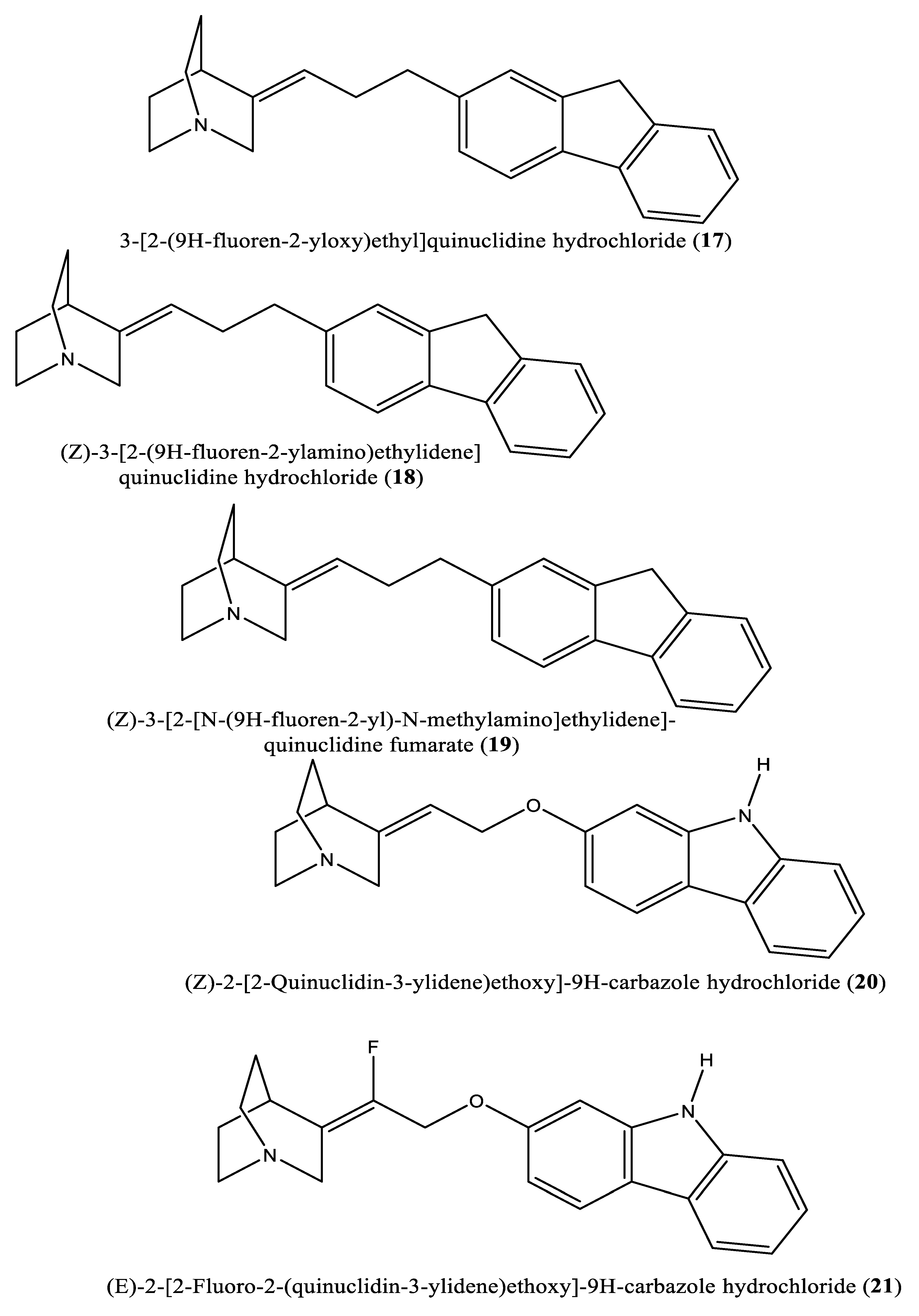
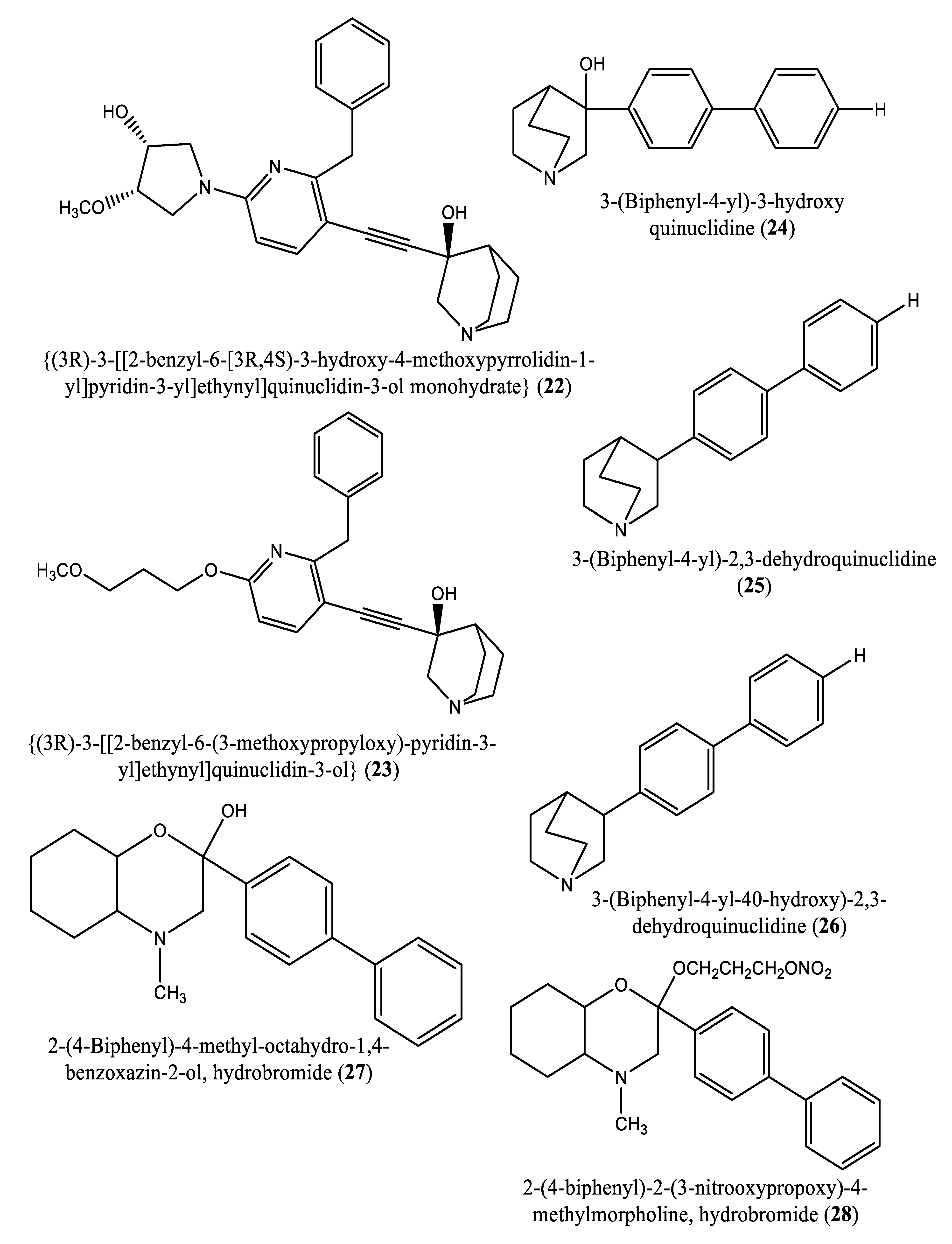
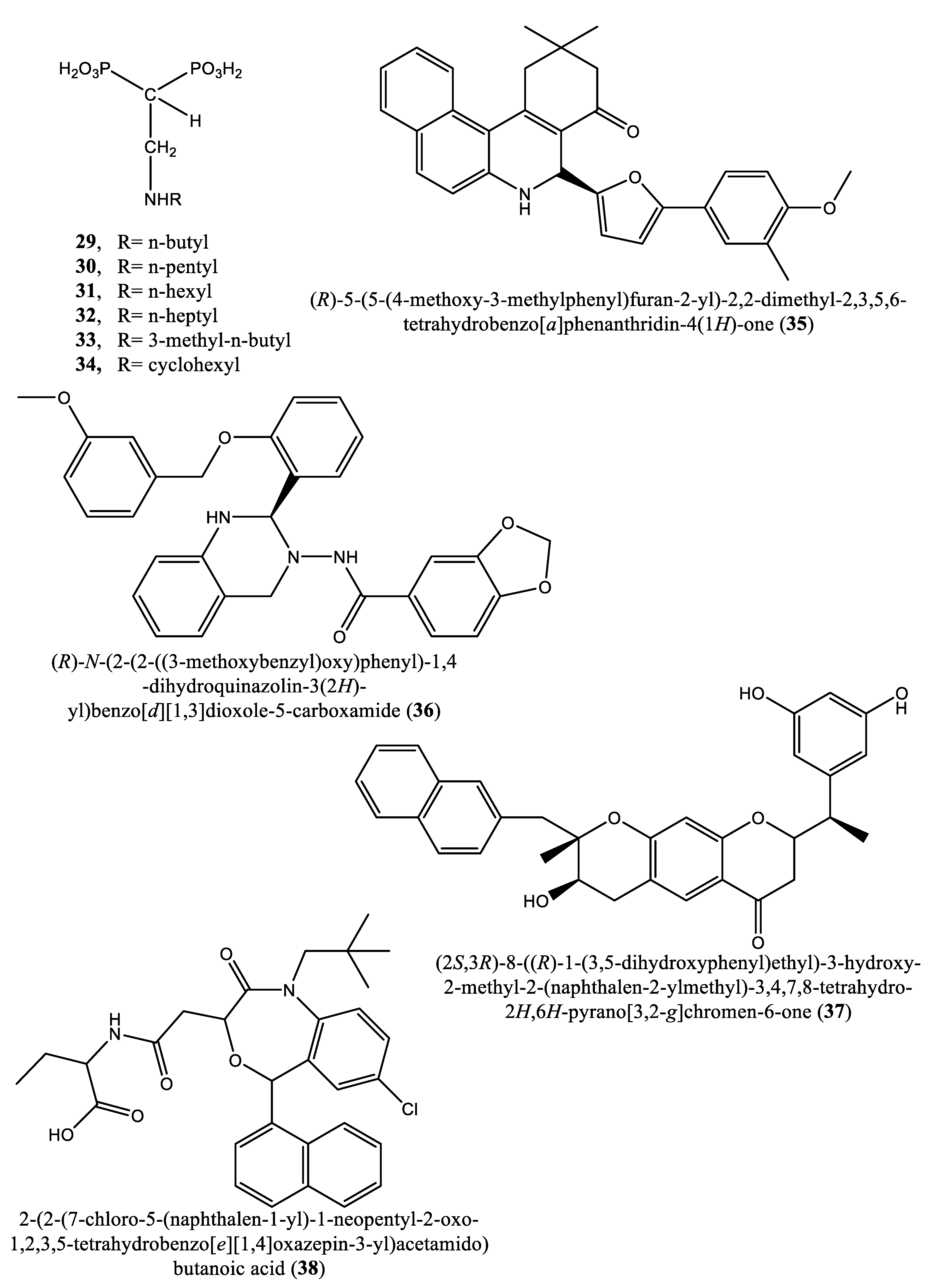
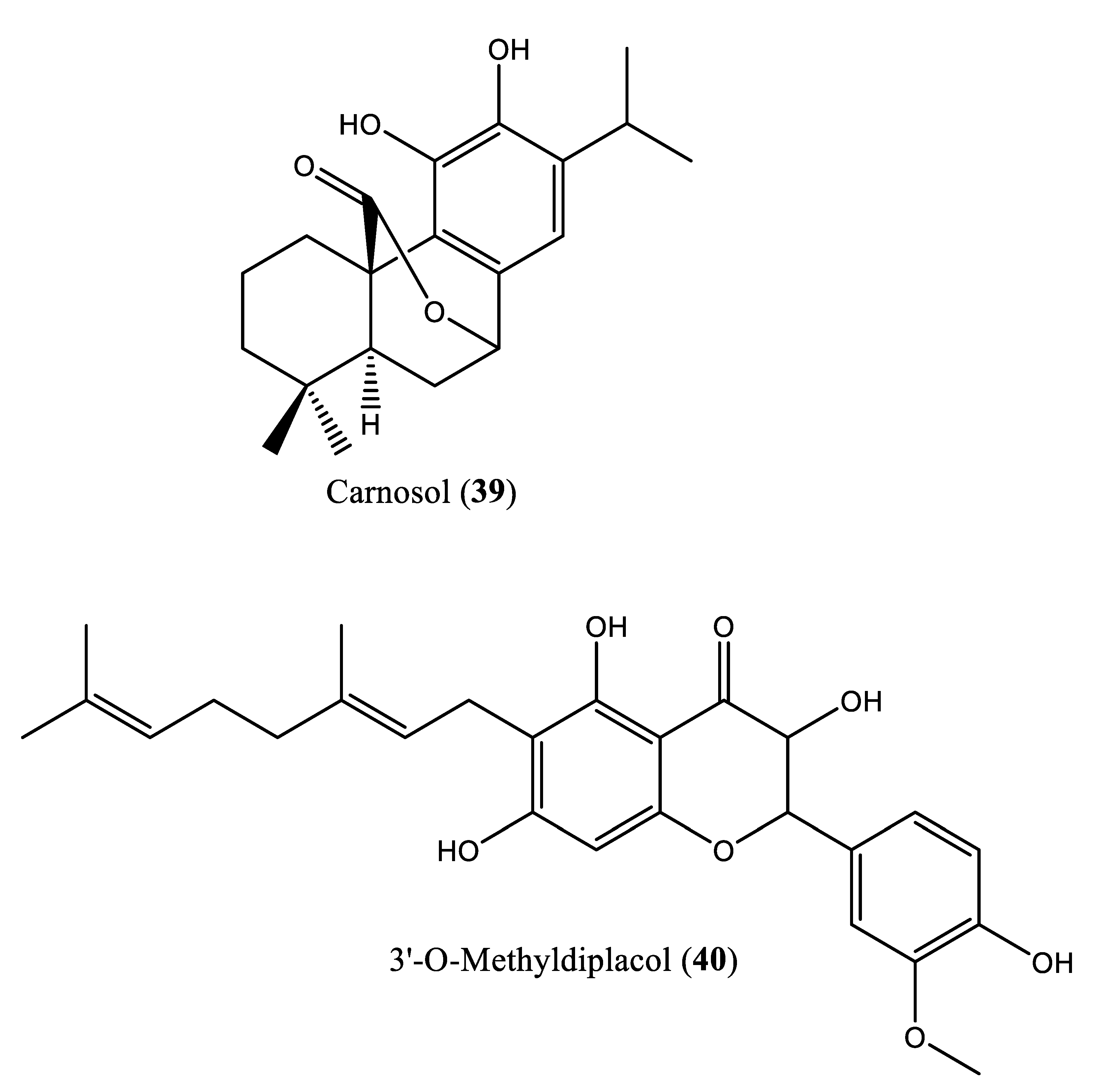
Figure 4.
Potential inhibitors of farnesyl diphosphate synthase.
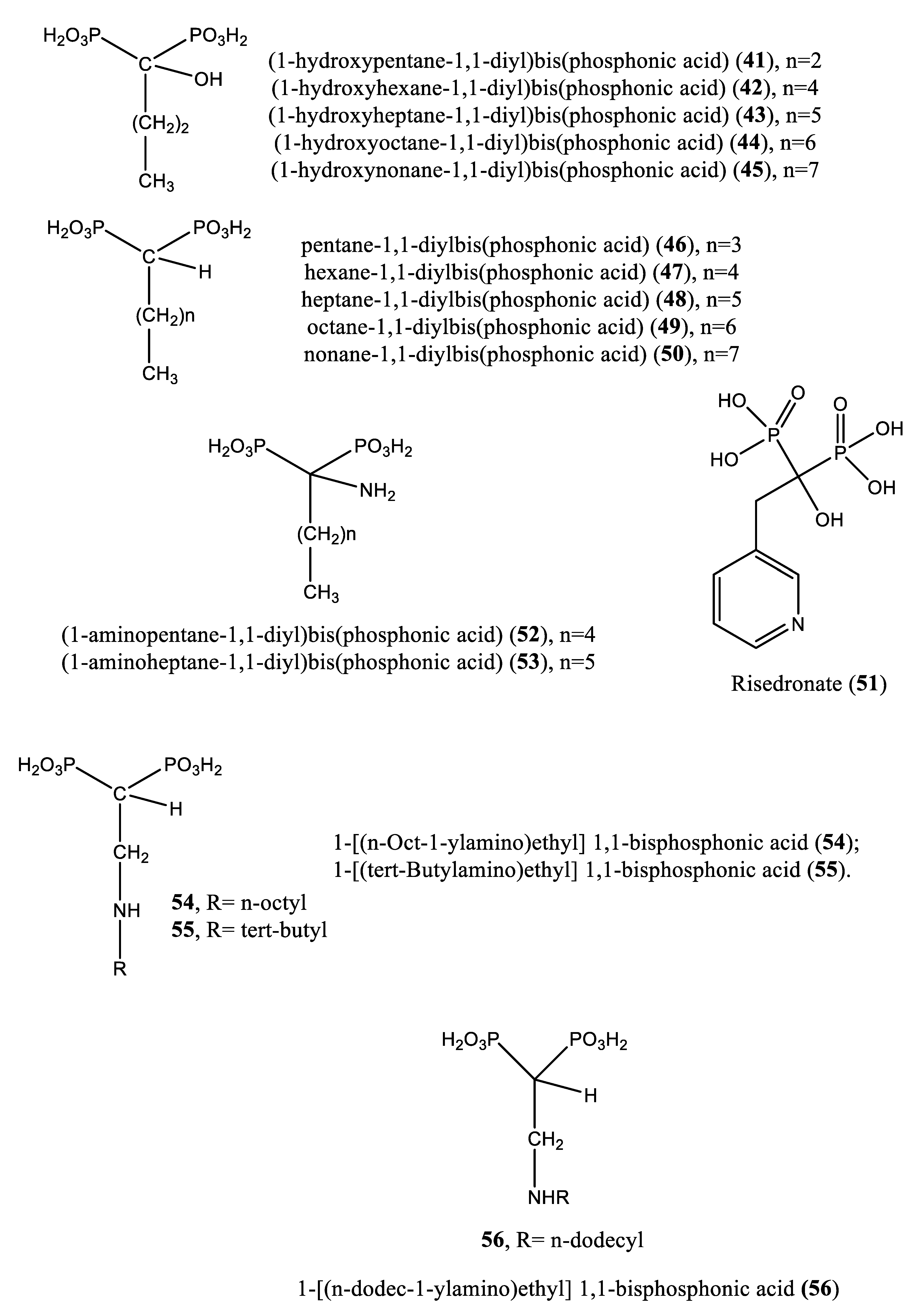
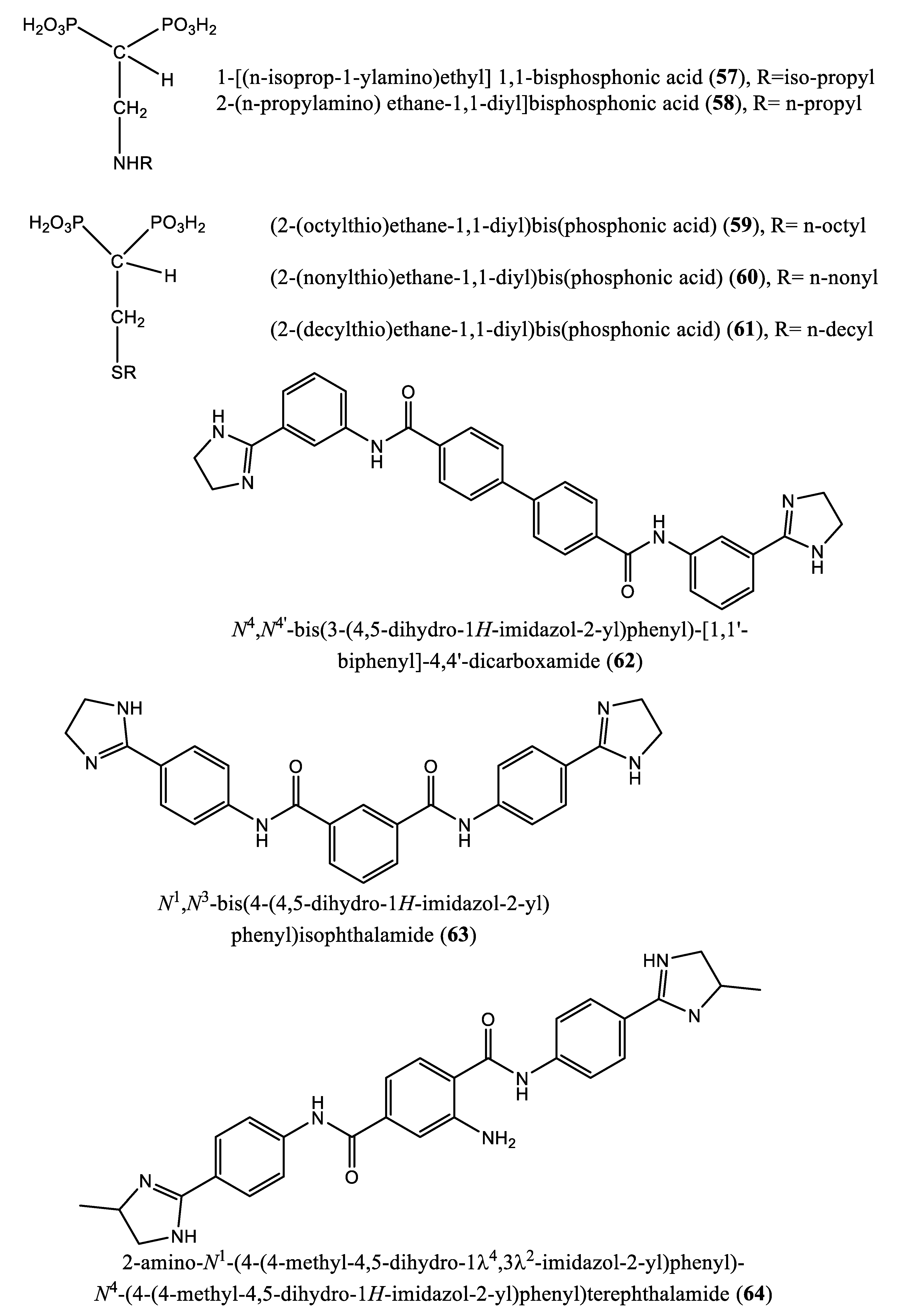
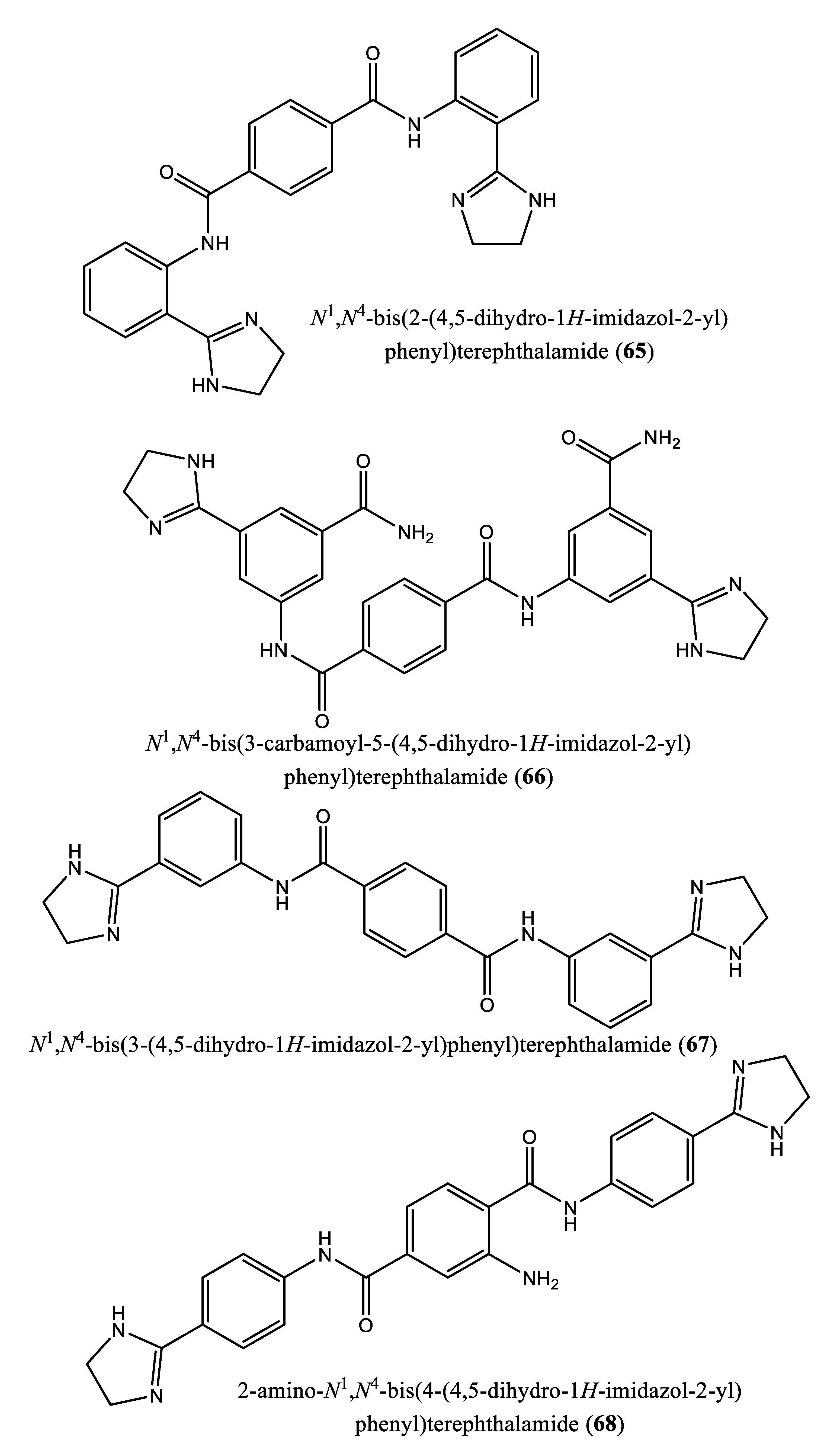
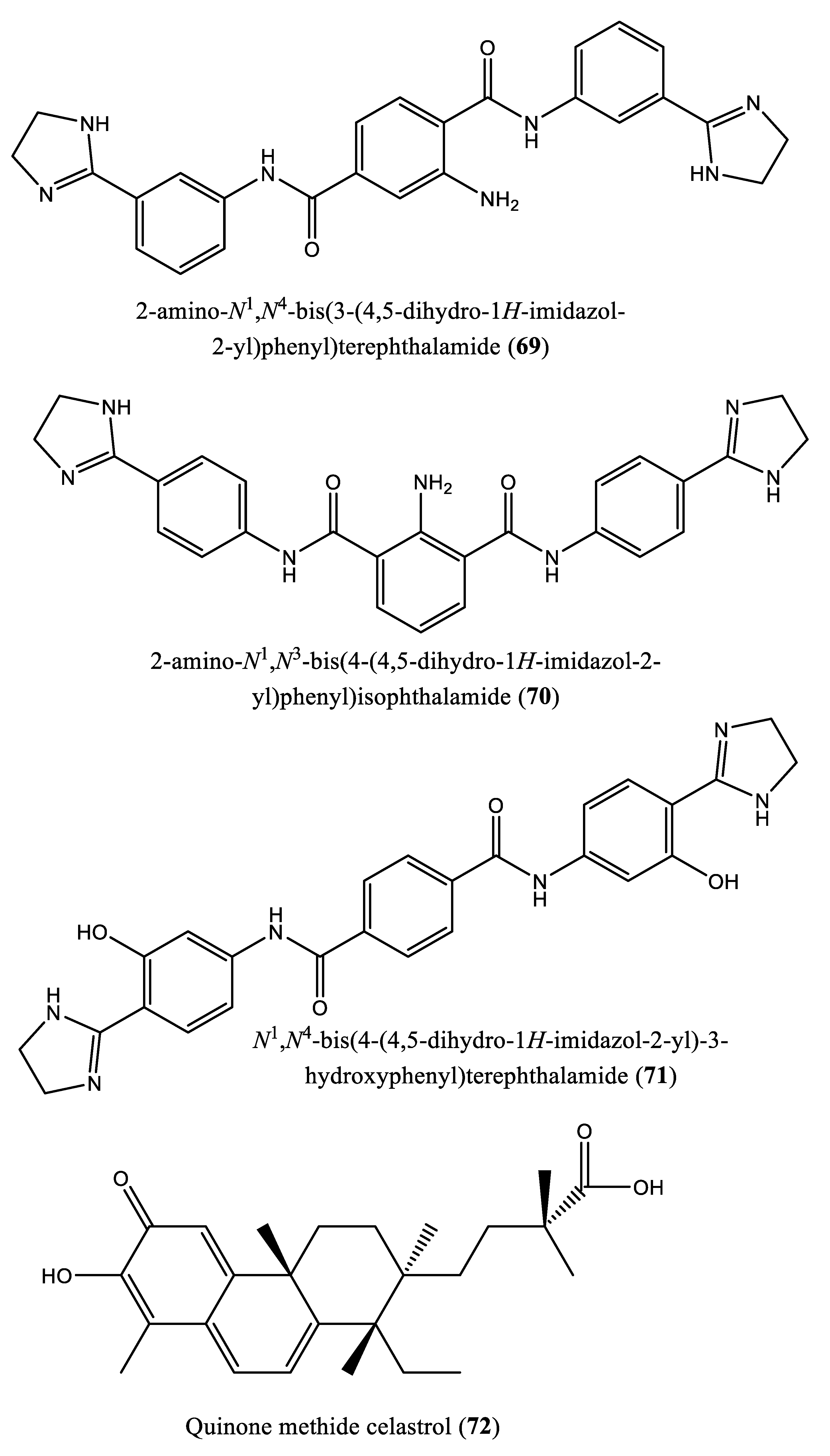
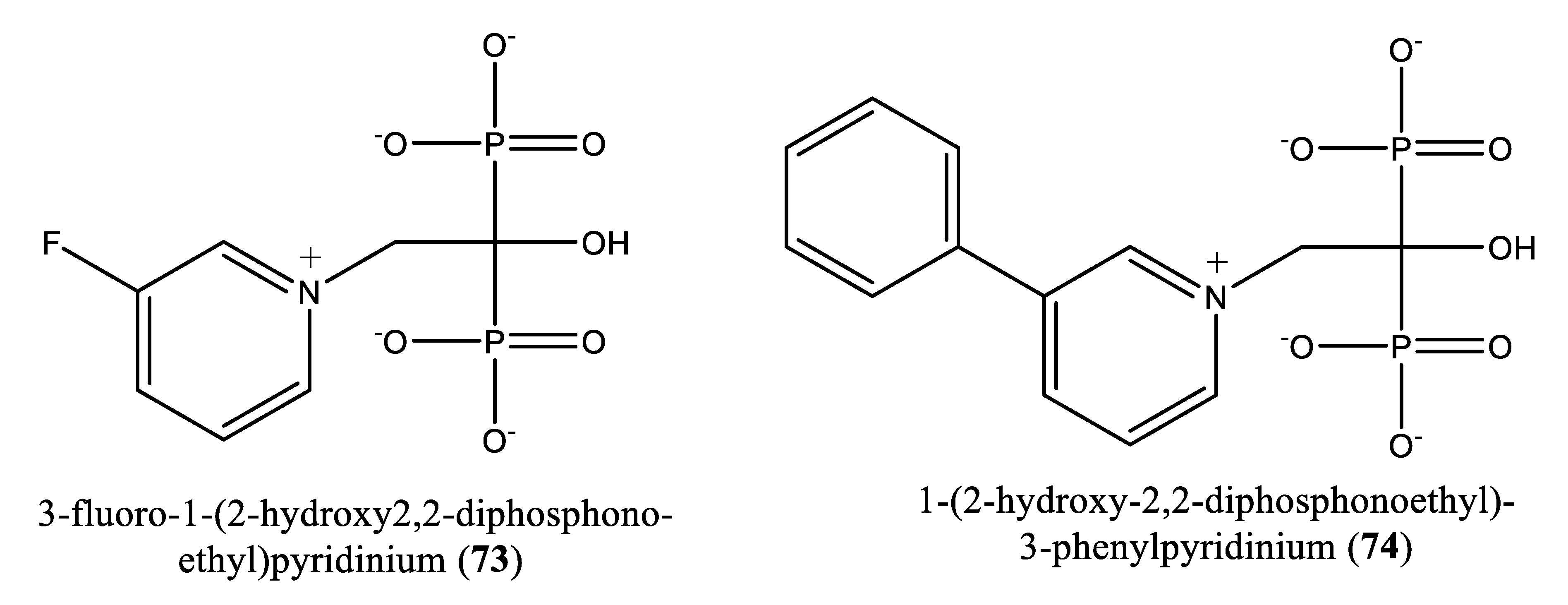
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



