Submitted:
19 June 2023
Posted:
19 June 2023
You are already at the latest version
Abstract
No major breakthroughs have entered mainstream clinical fertility practice since egg donation and intracytoplasmic sperm injection decades ago, and oocyte deficits secondary to advanced age continue as the main manifestation of diminished ovarian reserve. In the meantime, several unproven IVF ‘accessories’ have emerged including so-called ovarian rejuvenation which entails placing fresh autologous platelet-rich plasma (PRP) directly into ovarian tissue. Among cellular responses attributed to this intervention are reduced oxidative stress, slowed apoptosis, and improved metabolism. Besides impacting the existing follicle pool, platelet growth factors might also facilitate de novo oocyte recruitment by specified gene upregulation targeting uncommitted ovarian stem cells. Because disordered activity at mechanistic target of rapamycin (mTOR) has been shown to exacerbate or accelerate ovarian aging, PRP-discharged plasma cytokines combined with mTOR suppression by pulsed/cyclic rapamycin represents a novel fusion technique to enhance ovarian function. While beneficial effects have already been observed experimentally in oocytes and embryos with mTOR inhibition alone, this is the first discussion of intraovarian platelet cytokines followed by low-dose, phased rapamycin. For refractory cases, this investigational, tailored approach could amplify or sustain ovarian capacity sufficient to permit retrieval of competent oocytes via distinct but complementary pathways—thus reducing dependency on oocyte donation.
Keywords:
mTOR
; rapamycin
; PRP
; platelet cytokines
; ovarian reserve
; IVF
1. Introduction
Ovarian reserve constrains human fertility potential as female reproductive capacity deteriorates with increasing age. The primordial follicle pool and its rate of activation are compromised by chronic low-grade inflammation over time, culminating in eventual loss of both egg quality and quantity. Once considered nonrenewable, primordial follicles supply the cells which move gradually into the active (growing) follicle group [1,2]. Auditing this process has practical relevance, since few fertility clinics will embark on an IVF cycle without estimating ovarian reserve first. The failure to respond to gonadotropins, irrespective of dose or duration, becomes the final common pathway for any ‘low reserve’ problem.
While choice of IVF stimulation protocols can sometimes be empiric [3], a better understanding of how phosphatidylinositol 3-kinase (PI3K)/mammalian-mechanistic target of rapamycin (mTOR) can be driven by specific gonadotropins has been helpful. Some IVF medications, for example, preferentially boost insulin-like growth factor 1 (IGF-1) with a view to improve oocyte quality [4,5]. Because IGF-1, epidermal growth factor (EGF), platelet derived growth factor (PDGF), and vascular endothelial growth factor (VEGF) are among cytokines sourced from activated platelets (PLTs), this led to a hypothesis that reproductive outcome might be shaped by intraovarian platelet-rich plasma (PRP) [6-8]. Insulin/IGF-1 signaling can also regulate the PI3K/mTOR cascade to advance follicle stimulating hormone (FSH)-mediated granulosa cell development [9]. In mammalian ovarian function, the PI3K/mTOR pathway imbricates with other signaling motifs to calibrate steroidogenesis, granulosa proliferation, corpus luteum survival, and oocyte maturation [10,11]. Yet satisfactory IVF stimulations (with or without intraovarian PRP) can still unfortunately yield cycle cancellation or failure, so an alternative therapeutic option—rapamycin—is explored here for selected refractory, poor-prognosis IVF patients. While rapamycin is known for its general anti-aging effects [12-14], what features would be most salient if this were to be repurposed for an ovarian application, especially if used in tandem with intraovarian PRP?
2. PRP and its cytokine constituents
Aging in the ovary unfortunately traces a different trajectory compared to somatic cells, with anti- and pro-longevity genes in oocytes tending to change in opposite directions over time [15]. Ovarian reserve begins to decline measurably often by about age 35yrs and despite advancements of oocyte donation and ICSI [16,17], the need to enlarge the oocyte reservoir remains acute. In this regard, PRP includes components able to upregulate pluripotency genes (e.g., c-Myc, Klf4, Oct3/4, Sox2) associated with reprogramming somatic cells for a pluripotent lineage [18]. Here Oct4 has special relevance for ovarian remodeling, as human testicular cells express this marker after PRP culture [19] and a parallel response in ovarian tissue is plausible following intraovarian activated platelet-derived cytokines.
Perhaps most crucially, stem cells near ovarian surface epithelium [20,21] are well placed for research and clinical access. These cells display features which permit cellular transformations into different functional lineages (e.g., epithelium to mesenchyme) [22-24]. Local macrophages assert some role in this, although how this influence occurs is not known exactly. Xiao et al recently (2022) reported that inflammation associated with ovulation in mice drives selective activation of primordial follicles at each estrous cycle [25] depending on follicular macrophages having either M1 or M2 polarization. Interestingly, newborn ovaries cocultured with these macrophage subtypes evince stimulatory features with M1 macrophages, but dormancy characteristics with M2 macrophages [25].
This discovery aligned with earlier data which found M2 macrophages more often in older murine retinal tissue [26]. Importantly, this M1/M2 switching is controlled by the PI3K/mTOR signaling pathway [25]. PI3K/mTOR also orchestrates complex intracellular signaling systems which direct proliferation, cellular quiescence, and longevity. With relevance to ovarian rejuvenation practice, PI3K/mTOR is enhanced or regulated by specific PRP cytokine components including EGF, fibroblast growth factor 2 (FGF-2), and IGF-1 [27-29]. Such research is consistent with metabolic cross-talk among platelet- derived cytokines and pluripotency networks, perhaps explaining more fully what has become known as ‘ovarian rejuvenation’ [30].
3. Rapamycin, mTOR, and reproductive biology
First described in 1972, rapamycin was isolated from Streptomyces hygroscopicus found in soil and plant samples collected on Rapa Nui (Easter Island). First developed as an antifungal and immunosuppressant (see Figure 1), the substance was later found to have potent anti-tumor properties [31]. Further work on mechanism of action showed rapamycin complexes with the 12 kDa peptidyl-prolyl cis-trans isomerase FK506-binding protein-12 (FKBP12), to block proliferation and cell growth [32].
In 1991, the protein target of rapamycin (TOR) was discovered in Saccharomyces cerevisiae, where TOR gene mutations were noted to cause rapamycin resistance [33]. Subsequent research confirmed mTOR as the allosteric binding site for the rapamycin-FKBP12 complex in mammalian cells [34] where its organizing role in autophagy and cellular senescence was later characterized [35]. It is now agreed mTOR is a serine/ threonine protein kinase in the Class IV PI3K superfamily, which regulates proliferation, growth, and cell survival [36].
Integral to many complex signaling networks, mTOR drives adult stem cell proliferation and dictates the differentiation programs of stem cells [37]. Full deletion of mTOR is lethal shortly after embryo implantation [38], yet mTOR null blastocysts can have near-normal early morphology. Nevertheless, trophoblast formation is impaired such that cells taken from the inner cell mass will not proliferate when cultured in vitro. Hence, proper mTOR activity is mandatory for normal embryo development past the blastocyst stage [39]. Rapamycin slows proliferative decay via p16 and butyrate-induced p21 [12], and partial mTOR inhibition enhances maturation of selected populations of human stem cell–derived cardiomyocytes [40].
The consequences of mTOR overactivation are evident in animal progeria models. For example, in the Ercc1-/Δ accelerated aging (mouse) model, rapamycin improved muscle-derived stem cell function via autophagy [41]. Interrupting mTOR signaling by blocking its downstream target (S6K) resulted in longer lifespan and preserved cell function [42]. Specifically, rapamycin was able to recover differentiation and proliferation, reduce senescence, and enhance autophagy in a murine progeria model [43]. Hyperactive mTOR with aging thus seems to have serious and harmful consequences for somatic stem cells [13], and mice given rapamycin for the first 45d of life attain longer lifespans [14]. In human umbilical vein endothelial cells, rapamycin suppresses migration, reverses TGF-β1 stimulated endothelial-to-mesenchymal transitions, and downregulates the mesenchymal marker SMA-α [44].
In clinical medicine, a severe derangement of aging is confronted in Hutchinson- Gilford syndrome, an ultrarare progeria [45]. In this condition, progerin deposits occur due to a single point mutation c.1824C→T in exon 11 of the LMNA gene [46]. Abnormal primary transcript splicing while forming the lamin A mRNA generates progerin as an intracellular accretion; children with this mutation experience accelerated aging with death often before age 15yrs. In this disease, one promising therapy is to stimulate autophagy for clearance of toxic progerin by rapamycin [46]. Of note, rapamycin was recently used for successful treatment of an unrelated cardiomyopathy which involved a LMNA gene variant and dysregulated mTOR [47].
Altered mitochondrial status is another hallmark of aging, and key quality control checks have evolved to prevent vertical transmission of any ovarian mtDNA error. Palozzi & Hurd (2023) recently completed an RNAi screen in Drosophila to find mtDNA integrity surveilled via mTOR complex 1 (mTORC1), implicating the mitophagy adaptor BNIP3 and RNA-binding protein Atx2 as major elements [48]. This extended earlier research which showed that BNIP3 mediates inhibition of mTOR in response to hypoxia [49]. Specifically, Atx2 (C. elegans homolog of human ATXN2L and ATXN2) regulates mTOR and the ‘dietary restriction’ phenotype [50]. Indeed, local nutrient availability (e.g., folate [51]) is sensed by mTOR which then coordinates metabolism, growth, and autophagy functions. More recently in early mammalian embryos, the impact of rapamycin on mitochondrial fission and mitophagy [52] was studied under varied rapamycin concentrations, with significant improvements noted in blastocyst development, autophagy formation, and mitochondrial activation with rapamycin compared to no treatment [52]. In addition, among >200 immature human oocytes submitted for in vitro maturation then fertilized by ICSI, more high-quality embryos were obtained with rapamycin culture vs. untreated controls [53], and histone γH2AX levels (indicating double-strand DNA breaks) in oocytes cultured with rapamycin were also markedly reduced vs. controls [53]. While one-way (permanent) loss of growth potential was previously observed as blocked in arrested cells, rapamycin does not push the arrested cells into proliferation. Instead, rapamycin enables a permissively reversible aging condition [12].
Notwithstanding its use in organ transplantation which succeed best with minimal inflammation, rapamycin also paradoxically increases some pro-inflammatory cytokine outputs (i.e., IL-6, IL-12, IL-23) while lowering production of the anti-inflammatory IL-10 [54]. Such actions would not be welcome if rapamycin were planned for simultaneous use with conventional intraovarian PRP, which by design places PLT boluses beneath surface epithelium [55]. That PLTs exposed to rapamycin undergo functional change is uncontested—a property exploited in vascular stents coated with rapamycin to prevent restenosis [56]. Higher-dose rapamycin interferes with aspirin’s ability to block PLT aggregation [57,58] and more research is needed to know its impact on PLT morphology, membrane phosphatidylserine, and thrombin formation [36]. A biphasic disruption in PLT calcium homeostasis does occur with rapamycin, mediated by slowed activation and granule release [59]. Thus, if PLT function were disturbed by rapamycin, then pairing it with conventional PRP would make an odd therapeutic combination.
4. PLT cytokine augmentation by rapamycin?
The puzzle of a treatment plan interlocking ovarian PRP with rapamycin, given the latter’s interference with PLT features, may at first seem intractable. But using condensed PLT cytokines isolated as a cell-free product would bypass the rapamycin tampering problem, and this PRP variant has already been used successfully—without rapamycin—for human ovaries [60]. In other words, if PLT cytokines are separated first and then inserted into ovarian tissue as a filtered condensate by PLT subtraction, the microclimate affected by rapamycin would, in aggregate, end up supporting the potential cytokine commitment of ovarian stem precursors to an oocyte lineage. In this way, rapamycin sustains or possibly extends the ovarian response as entrained by fresh, autologous PLT cytokines.
Investigating how lowering mTOR action affects ovarian biology is not original here, as measurements by NYU researchers found that a two- to four-fold dampening of mTOR activity preserves ovarian function and parity [61]. The current model assumes that limited local inflammation is not injurious but essential, and from this perspective, intraovarian PLT growth factors followed by rapamycin would be an adaptation of the latter’s recognized role in tissue repair. If introduced at a low dose following ovarian treatment with condensed PLT cytokines, pulsed rapamycin might provide metabolic gains similar to fasting [62] for a nascent follicle pool undergoing induction by platelet-derived growth factors.
5. Rapamycin—Scheduling & toxicity issues
Rapamycin at relatively low dose is more likely to confer a beneficial response with preferential mTORC1 inhibition, while blunting any undesirable effects on mTORC2 [63]. In USA, Rapamycin (Sirolimus) is available as 0.5, 1, and 2mg tablets as well as 1mg/mL oral solution; it is classified as FDA Pregnancy category C. An improved immune response was reported from one short-term clinical study using the ‘rapalog’ everolimus [64], but a rapamycin protocol specific to the adult human ovary has not been standardized. Unlike high-dose rapamycin used for immunosuppression in transplant medicine which has received close monitoring, establishing a toxicity threshold for rapamycin use in ovarian (fertility) applications awaits additional study. Poison control records include a failed suicide attempt where a female age 18yrs consumed more than 100 tablets of 1mg rapamycin each, and high serum cholesterol was the only documented abnormality [65]. In rats, the LD50 for rapamycin could not be calculated because it exceeded 2500mg/kg [66].
Inconsistency in rapamycin dosing required for adequate mTOR suppression has also complicated its therapeutic use. Differing rapamycin sensitivities are likely due to varied functional characteristics in mTOR complex 1 and 2 (mTORC1 & mTORC2). For example, mTORC1 is inhibited at low nM levels of rapamycin while mTORC2 suppression usually requires chronic use at higher concentrations [67]. PI3K/Akt works with Hippo signaling to accelerate recruitment of primordial follicles, while mTORC1 suppression suppresses uncoordinated ‘flash’ discharges resulting in mass follicular activation [9,68]. Platelet-rich plasma (or its condensed cytokine derivatives) plus rapamycin has not been previously studied, probably because both treatments are somewhat novel and available research on each is unsettled.
The combined method favored here for ovarian use is a variation on a prior rapamycin dosing calendar with short, pulsed exposure at low doses [64,69]. Specifically, this means phased/cyclic monthly rapamycin with the first day’s oral dose at 3mg, taken one week after the office PRP procedure. For subsequent days, 1mg/d is taken for six days and then no rapamycin for the next three weeks. Next, a 3mg loading dose is repeated on the first day of Cycle #2 again with 1mg/d taken x 6d. Cycles #3 and #4 follow the same pattern, so a total of four pulses is completed before planned IVF (see Figure 2).
By comparison, oral rapamycin tablets given for prevention of organ transplant rejection can entail a larger initial first-day 6mg loading dose, followed by 2mg/d [70]. Baseline and follow-up laboratory measurements appropriate for rapamycin use include serum AMH, complete blood count (w/PLT), C-reactive protein, comprehensive metabolic panel, serum E2, ferritin, FSH/LH, with fasting insulin and lipid panel. Additional testing for specific gene regulators may not be available in all centers.
6. Conclusions
Sponsorship for rapamycin investigation significantly outpaces ovarian PRP research. At present >1000 rapamycin anti-aging clinical trials are formally registered [71], although none focus specifically on premature ovarian insufficiency or low reserve. Because the total number of registered ovarian PRP clinical trials remains less than ten [72], it is unsurprising that research joining both topics is lacking. Nearly a decade ago, the failure to define general dosing guidelines for rapamycin was acknowledged as a major impediment to its use [73], and consensus on rapamycin use in ovarian biology is likewise absent.
While pulsed or phased oral rapamycin dosing has been discussed in the setting of wound healing [74], this is the first attempt to outline dosing before IVF. Oocyte capacity sets downstream reproductive fidelity through meiosis, fertilization, nidation, and eventual development to term, and treatments to affect these have been reviewed for the adult human ovary [75,76]. The experimental nature of intraovarian PRP [7] and condensed plasma cytokines [60] notwithstanding, these interventions alone may still be insufficient for some patients. Because intraovarian injection of PLT growth factors can increase serum AMH (indicating expansion of the follicle/oocyte unit) [7,77] and mTOR inhibition has been suggested to boost ovarian reserve [78], a protocol incorporating both as described here might provide a useful synergy where single-agent refractoriness is encountered.
Author Contributions
E.S.S. developed the protocol and organized initial drafts; E.S.S., C.H., S.H.W. and S.L.T. reviewed the literature and revised drafts. All authors read and approved the final manuscript.
Funding
This project received no external funding.
Conflicts of Interest
ESS has been awarded U.S. Trademark #88505430 for process & method using autologous platelet cytokines for ovarian therapy. CH, SHW and SLT have no disclosures.
References
- Kallen A, Polotsky AJ, Johnson J. Untapped reserves: Controlling primordial follicle growth activation. Trends Mol Med 2018;24(3):319-31. [CrossRef]
- Ford EA, Beckett EL, Roman SD, McLaughlin EA, Sutherland JM. Advances in human primordial follicle activation and premature ovarian insufficiency. Reproduction 2020;159(1):R15-29. [CrossRef]
- Shrestha D, La X, Feng HL. Comparison of different stimulation protocols used in in vitro fertilization: A review. Ann Transl Med 2015;3(10):137. [CrossRef]
- Mehta BN, Chimote NM, Chimote MN, Chimote NN, Nath NM. Follicular fluid insulin like growth factor-1 (FF IGF-1) is a biochemical marker of embryo quality and implantation rates in in vitro fertilization cycles. J Hum Reprod Sci 2013;6(2):140-6. [CrossRef]
- Ji Z, Quan X, Lan Y, Zhao M, Tian X, Yang X. Gonadotropin vs. follicle-stimulating hormone for ovarian response in patients undergoing in vitro fertilization: A retrospective cohort comparison. Curr Ther Res Clin Exp 2019;92:100572. [CrossRef]
- Banu J, Tarique M, Jahan N, Lasker N, Sultana N, Alamgir CF, et al. Efficacy of autologous platelet rich plasma for ovarian rejuvenation in infertile women having poor ovarian reserve. Int J Reprod Contracept Obstet Gynecol 2022;11:2948-53. [CrossRef]
- Cakiroglu Y, Yuceturk A, Karaosmanoglu O, Kopuk SY, Korun ZEU, Herlihy N, et al. Ovarian reserve parameters and IVF outcomes in 510 women with poor ovarian response (POR) treated with intraovarian injection of autologous platelet rich plasma. Aging (Albany NY) 2022;14(6):2513-23. [CrossRef]
- Herlihy NS, Seli E. The use of intraovarian injection of autologous platelet rich plasma (PRP) in patients with poor ovarian response and premature ovarian insufficiency. Curr Opin Obstet Gynecol 2022;34(3):133-7. [CrossRef]
- Grosbois J, Demeestere I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum Reprod 2018;33(9):1705-14. [CrossRef]
- Gareis NC, Huber E, Hein GJ, Rodríguez FM, Salvetti NR, Angeli E, et al. Impaired insulin signaling pathways affect ovarian steroidogenesis in cows with COD. Anim Reprod Sci 2020;192:298-312. [CrossRef]
- Papageorgiou K, Mastora E, Zikopoulos A, Grigoriou ME, Georgiou I, Michaelidis TM. Interplay between mTOR and Hippo signaling in the ovary: Clinical choice guidance between different gonadotropin preparations for better IVF. Front Endocrinol (Lausanne) 2021;12:702446. [CrossRef]
- Demidenko ZN, Zubova SG, Bukreeva EI, Pospelov VA, Pospelova TV, Blagosklonny MV. Rapamycin decelerates cellular senescence. Cell Cycle 2009;8(12):1888-95. [CrossRef]
- Hambright WS, Philippon MJ, Huard J. Rapamycin for aging stem cells. Aging (Albany NY) 2020;12(15):15184-5. [CrossRef]
- Shindyapina AV, Cho Y, Kaya A, Tyshkovskiy A, Castro JP, Deik A, et al. Rapamycin treatment during development extends life span and health span of male mice and Daphnia magna. Sci Adv 2022;8(37):eabo5482. [CrossRef]
- Smits MAJ, Janssens GE, Goddijn M, Hamer G, Houtkooper RH, Mastenbroek S. Longevity pathways are associated with human ovarian ageing. Hum Reprod Open 2021;2021(2):hoab020. [CrossRef]
- Rosenwaks Z, Veeck LL, Liu HC. Pregnancy following transfer of in vitro fertilized donated oocytes. Fertil Steril 1986;45(3):417-20. [CrossRef]
- Palermo G, Joris H, Devroey P, Van Steirteghem AC. Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet 1992;340(8810):17-8. [CrossRef]
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006;126(4):663-76. [CrossRef]
- Khadivi F, Koruji M, Akbari M, Jabari A, Talebi A, Ashouri Movassagh S, et al. Application of platelet-rich plasma (PRP) improves self-renewal of human spermatogonial stem cells in two-dimensional and three-dimensional culture systems. Acta Histochem 2020;122(8):151627. [CrossRef]
- Parte S, Bhartiya D, Telang J, Daithankar V, Salvi V, Zaveri K, et al. Detection, characterization, and spontaneous differentiation in vitro of very small embryonic-like putative stem cells in adult mammalian ovary. Stem Cells Dev 2011;20(8):1451-64. [CrossRef]
- Bhartiya D, Singh P, Sharma D, Kaushik A. Very small embryonic-like stem cells (VSELs) regenerate whereas mesenchymal stromal cells (MSCs) rejuvenate diseased reproductive tissues. Stem Cell Rev Rep 2022;18(5):1718-27. [CrossRef]
- Virant-Klun I, Skutella T. Stem cells in aged mammalian ovaries. Aging (Albany NY) 2010;2(1):3-6. [CrossRef]
- Wu M, Lu Z, Zhu Q, Ma L, Xue L, Li Y, et al. DDX04+ stem cells in the ovaries of postmenopausal women: Existence and differentiation potential. Stem Cells 2022;40(1):88-101. [CrossRef]
- Woods DC, Tilly JL. Revisiting claims of the continued absence of functional germline stem cells in adult ovaries. Stem Cells 2023;41(2):200-4. [CrossRef]
- Xiao Y, Peng X, Peng Y, Zhang C, Liu W, Yang W, et al. Macrophage-derived extracellular vesicles regulate follicular activation and improve ovarian function in old mice by modulating local environment. Clin Transl Med 2022;12(10):e1071. [CrossRef]
- Cruz-Guilloty F, Saeed AM, Echegaray JJ, Duffort S, Ballmick A, Tan Y, et al. Infiltration of proinflammatory M1 macrophages into the outer retina precedes damage in a mouse model of age-related macular degeneration. Int J Inflam 2013;2013:503725. [CrossRef]
- Peltier J, O'Neill A, Schaffer DV. PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation. Dev Neurobiol 2007;67(10):1348-61. [CrossRef]
- Ojeda L, Gao J, Hooten KG, Wang E, Thonhoff JR, Dunn TJ, et al. Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF. PLoS One 2011;6(8):e23414. [CrossRef]
- Marck RE, Gardien KLM, Vlig M, Breederveld RS, Middelkoop E. Growth factor quantification of platelet-rich plasma in burn patients compared to matched healthy volunteers. Int J Mol Sci 2019;20(2):288. [CrossRef]
- Pantos K, Nitsos N, Kokkali G, Vaxevanoglou T, Markomichali C, Pantou A, et al. Ovarian rejuvenation and folliculogenesis reactivation in peri-menopausal women after autologous platelet-rich plasma treatment (abstract) ESHRE 32nd Annual Meeting [Helsinki] 3-6 July, 2016. Hum Reprod 2016:i301. https://sa1s3.patientpop.com/assets/docs/111052.pdf.
- Vézina C, Kudelski A, Sehgal SN. Rapamycin (AY-22,989), a new antifungal antibiotic. I. Taxonomy of the producing streptomycete and isolation of the active principle. J Antibiot (Tokyo) 1975;28:721-6. [CrossRef]
- Chung J, Kuo CJ, Crabtree GR, Blenis J. Rapamycin-FKBP specifically blocks growth-dependent activation of and signaling by the 70kd S6 protein kinases. Cell 1992;69:1227-36. [CrossRef]
- Heitman J, Movva NR, Hall MN. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991;253(5022):905-9. [CrossRef]
- Sabatini DM, Erdjument-Bromage H, Lui M, Tempst P, Snyder SH. RAFT1: A mammalian protein that binds to FKBP12 in a rapamycin-dependent fashion and is homologous to yeast TORs. Cell 1994;78: 35-43. [CrossRef]
- Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and disease. Cell 2017;168(6):960-76. [CrossRef]
- Śledź KM, Moore SF, Durrant TN, Blair TA, Hunter RW, Hers I. Rapamycin restrains platelet procoagulant responses via FKBP-mediated protection of mitochondrial integrity. Biochem Pharmacol 2020;177:113975. [CrossRef]
- Meng D, Frank AR, Jewell JL. mTOR signaling in stem and progenitor cells. Development 2018;145(1):dev152595. [CrossRef]
- Murakami M, Ichisaka T, Maeda M, Oshiro N, Hara K, Edenhofer F, et al. mTOR is essential for growth and proliferation in early mouse embryos and embryonic stem cells. Mol Cell Biol 2004;24(15):6710-8. [CrossRef]
- Alhasan BA, Gordeev SA, Knyazeva AR, Aleksandrova KV, Margulis BA, Guzhova IV, et al. The mTOR pathway in pluripotent stem cells: Lessons for understanding cancer cell dormancy. Membranes (Basel) 2021;11(11):858. [CrossRef]
- Garbern JC, Helman A, Sereda R, Sarikhani M, Ahmed A, Escalante GO, et al. Inhibition of mTOR signaling enhances maturation of cardiomyocytes derived from human-induced pluripotent stem cells via p53-induced quiescence. Circulation 2020;141(4):285-300. [CrossRef]
- Takayama K, Kawakami Y, Lavasani M, Mu X, Cummins JH, Yurube T, et al. mTOR signaling plays a critical role in the defects observed in muscle-derived stem/progenitor cells isolated from a murine model of accelerated aging. J Orthop Res 2017;35(7):1375-82. [CrossRef]
- Papadopoli D, Boulay K, Kazak L, Pollak M, Mallette F, Topisirovic I, et al. mTOR as a central regulator of lifespan and aging. F1000Res 2019;8:F1000 Faculty Rev-998. [CrossRef]
- Kawakami Y, Hambright WS, Takayama K, Mu X, Lu A, Cummins JH, et al. Rapamycin rescues age-related changes in muscle-derived stem/progenitor cells from progeroid mice. Mol Ther Methods Clin Dev 2019;14:64-76. [CrossRef]
- Tian D, Zeng X, Wang W, Wang Z, Zhang Y, Wang Y. Protective effect of rapamycin on endothelial-to-mesenchymal transition in HUVECs through the Notch signaling pathway. Vascul Pharmacol 2019;113:20-6. [CrossRef]
- Gordon LB, Norris W, Hamren S, Goodson R, LeClair J, Massaro J, et al. Plasma progerin in patients with Hutchinson-Gilford progeria syndrome: Immunoassay development and clinical evaluation. Circulation 2023: in press. [CrossRef]
- Mosevitsky, MI. Progerin and Its role in accelerated and natural aging. Mol Biol (Mosk) 2022;56(2):181-205. [CrossRef]
- Neupane B, Pradhan K, Ortega-Ramirez AM, Aidery P, Kucikas V, Marks M, et al. Personalized medicine approach in a DCM patient with LMNA mutation reveals dysregulation of mTOR signaling. J Pers Med 2022;12(7):1149. [CrossRef]
- Palozzi JM, Hurd TR. The role of programmed mitophagy in germline mitochondrial DNA quality control. Autophagy 2023:1-2. [CrossRef]
- Li Y, Wang Y, Kim E, Beemiller P, Wang CY, Swanson J, et al. Bnip3 mediates the hypoxia-induced inhibition on mammalian target of rapamycin by interacting with Rheb. J Biol Chem 2007;282(49):35803-13. [CrossRef]
- Bar DZ, Charar C, Dorfman J, Yadid T, Tafforeau L, Lafontaine DL, et al. Cell size and fat content of dietary-restricted Caenorhabditis elegans are regulated by ATX-2, an mTOR repressor. Proc Natl Acad Sci USA 2016;113(32):E4620-9. [CrossRef]
- Silva E, Rosario FJ, Powell TL, Jansson T. Mechanistic target of rapamycin is a novel molecular mechanism linking folate availability and cell function. J Nutr 2017;147(7):1237-42. [CrossRef]
- Park HJ, Heo GD, Yang SG, Koo DB. Rapamycin encourages the maintenance of mitochondrial dynamic balance and mitophagy activity for improving developmental competence of blastocysts in porcine embryos in vitro. Mol Reprod Dev 2023: in press. [CrossRef]
- Yang Q, Xi Q, Wang M, Liu J, Li Z, Hu J, Jin L, Zhu L. Rapamycin improves the developmental competence of human oocytes by alleviating DNA damage during IVM. Hum Reprod Open 2022;2022(4):hoac050. [CrossRef]
- Allan, S. Seeing mTOR in a new light. Nat Rev Immunol 2008;8:904. [CrossRef]
- Sills ES, Wood SH. Epigenetics, ovarian cell plasticity, and platelet-rich plasma: Mechanistic theories. Reprod Fertil 2022;3(4):C44-C51. [CrossRef]
- Bae IH, Lim KS, Park DS, Shim JW, Lee SY, Jang EJ, et al. Sirolimus coating on heparinized stents prevents restenosis and thrombosis. J Biomater Appl 2017;31(10):1337-45. [CrossRef]
- Babinska A, Markell MS, Salifu MO, Akoad M, Ehrlich YH, Kornecki E. Enhancement of human platelet aggregation and secretion induced by rapamycin. Nephrol Dial Transplant 1998;13(12):3153-9. [CrossRef]
- Wu Q, Huang KS, Chen M, Huang DJ. Rapamycin enhances platelet aggregation induced by adenosine diphosphate in vitro. Platelets 2009;20(6):428-31. [CrossRef]
- López E, Berna-Erro A, Bermejo N, Brull JM, Martinez R, Garcia Pino G, et al. Long-term mTOR inhibitors administration evokes altered calcium homeostasis and platelet dysfunction in kidney transplant patients. J Cell Mol Med 2013;17(5):636-47. [CrossRef]
- Sills ES, Rickers NS, Svid CS, Rickers JM, Wood SH. Normalized ploidy following 20 consecutive blastocysts with chromosomal error: Healthy 46, XY pregnancy with IVF after intraovarian injection of autologous enriched platelet-derived growth factors. Int J Mol Cell Med 2019;8(1):84-90. [CrossRef]
- Goldman KN, Chenette D, Arju R, Duncan FE, Keefe DL, Grifo JA, et al. mTORC1/2 inhibition preserves ovarian function and fertility during genotoxic chemotherapy. Proc Natl Acad Sci USA 2017;114(12):3186-91. [CrossRef]
- Escobar KA, Cole NH, Mermier CM, VanDusseldorp TA. Autophagy and aging: Maintaining the proteome through exercise and caloric restriction. Aging Cell 2019;18(1):e12876. [CrossRef]
- Lamming DW, Ye L, Sabatini DM, Baur JA. Rapalogs and mTOR inhibitors as anti-aging therapeutics. J Clin Invest 2013;123(3):980-9. [CrossRef]
- Mannick JB, Del Giudice G, Lattanzi M, Valiante NM, Praestgaard J, Huang B, et al. mTOR inhibition improves immune function in the elderly. Sci Transl Med 2014;6(268):268ra179. [CrossRef]
- Ceschi A, Heistermann E, Gros S, Reichert C, Kupferschmidt H, Banner NR, et al. Acute sirolimus overdose: A multicenter case series. PLoS One 2015;10(5):e0128033. [CrossRef]
- Blagosklonny, MV. Rapamycin for longevity: Opinion article. Aging (Albany NY) 2019;11(19):8048-67. [CrossRef]
- Foster DA, Toschi A. Targeting mTOR with rapamycin: One dose does not fit all. Cell Cycle 2009;8(7):1026-9. [CrossRef]
- Sills ES, Wood SH, Walsh APH. Intraovarian condensed platelet cytokines for infertility and menopause-Mirage or miracle? Biochimie 2023;204:41-7. [CrossRef]
- Kennedy BK, Lamming DW. The mechanistic target of rapamycin: The grand conducTOR of metabolism and aging. Cell Metab 2016;23(6):990-1003. [CrossRef]
- Mayo Clinic Health Information (Drugs & Supplements) – Sirolimus DRG-20068199. Mayo Foundation for Medical Education & Research, 1 March 2023 https://www.mayoclinic.org/drugs-supplements/sirolimus-oral-route/proper-use/drg-20068199 [accessed 15 May 2023].
- U.S. National Library of Medicine, https://clinicaltrials.gov [accessed ]. 15 May.
- Sills ES, Wood SH. Progress in human ovarian rejuvenation: Current platelet-rich plasma and condensed cytokine research activity by scope and international origin. Clin Exp Reprod Med 2021;48(4):311-5. [CrossRef]
- Kaeberlein, M. Rapamycin and ageing: When, for how long, and how much? J Genet Genomics 2014;41(9):459-63. [CrossRef]
- Blagosklonny, MV. Fasting and rapamycin: Diabetes versus benevolent glucose intolerance. Cell Death Dis 2019;10(8):607. [CrossRef]
- Atkinson L, Martin F, Sturmey RG. Intraovarian injection of platelet-rich plasma in assisted reproduction: Too much too soon? Hum Reprod 2021;36(7):1737-50. [CrossRef]
- Sills, ES. Ovarian recovery via autologous platelet-rich plasma: New benchmarks for condensed cytokine applications to reverse reproductive aging. Aging Med (Milton) 2022;5(1):63-7. [CrossRef]
- Garavelas A, Mallis P, Michalopoulos E, Nikitos E. Clinical benefit of autologous platelet-rich plasma infusion in ovarian function rejuvenation: Evidence from a before-after prospective pilot study. Medicines (Basel) 2023;10(3):19. [CrossRef]
- Papapanou M, Syristatidi K, Gazouli M, Eleftheriades M, Vlahos N, Siristatidis C. The effect of stimulation protocols (GnRH agonist vs. antagonist) on the activity of mTOR and Hippo pathways of ovarian granulosa cells and its potential correlation with the outcomes of in vitro fertilization: A hypothesis. J Clin Med 2022;11(20):6131. [CrossRef]
Figure 1.
Rapamycin (Sirolimus), a macrolide lactone with potent immunosuppressant, antiproliferative, and antifungal properties, received U.S. FDA approval in 1999. While widely used in organ transplant surgery, lower-dose applications of this mTOR inhibitor have successfully decelerated cellular aging to extend lifespan. Any emergent oocyte precursors available after intraovarian injection of PLT cytokines may benefit from reduced mTOR activity, as described here.
Figure 1.
Rapamycin (Sirolimus), a macrolide lactone with potent immunosuppressant, antiproliferative, and antifungal properties, received U.S. FDA approval in 1999. While widely used in organ transplant surgery, lower-dose applications of this mTOR inhibitor have successfully decelerated cellular aging to extend lifespan. Any emergent oocyte precursors available after intraovarian injection of PLT cytokines may benefit from reduced mTOR activity, as described here.
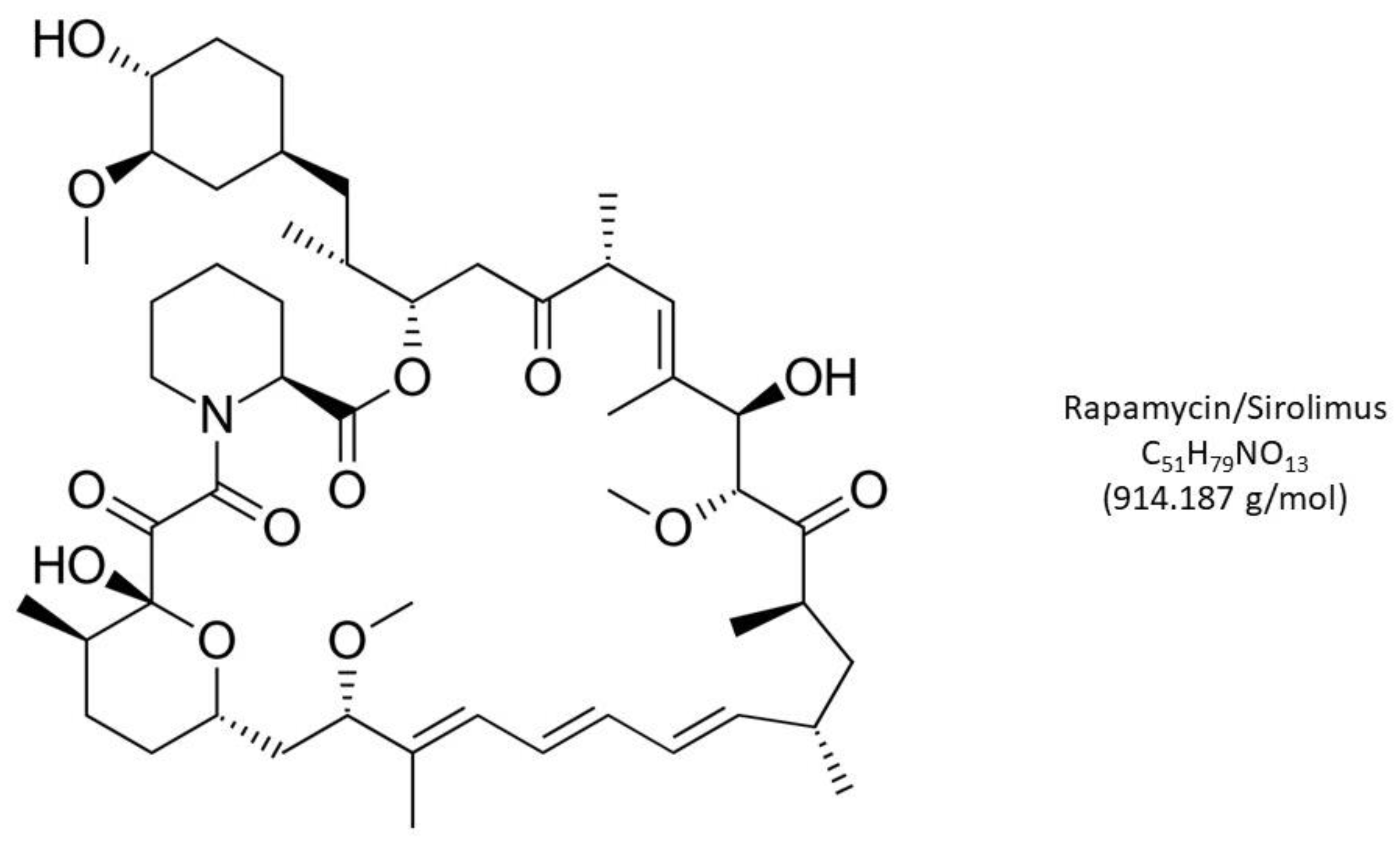
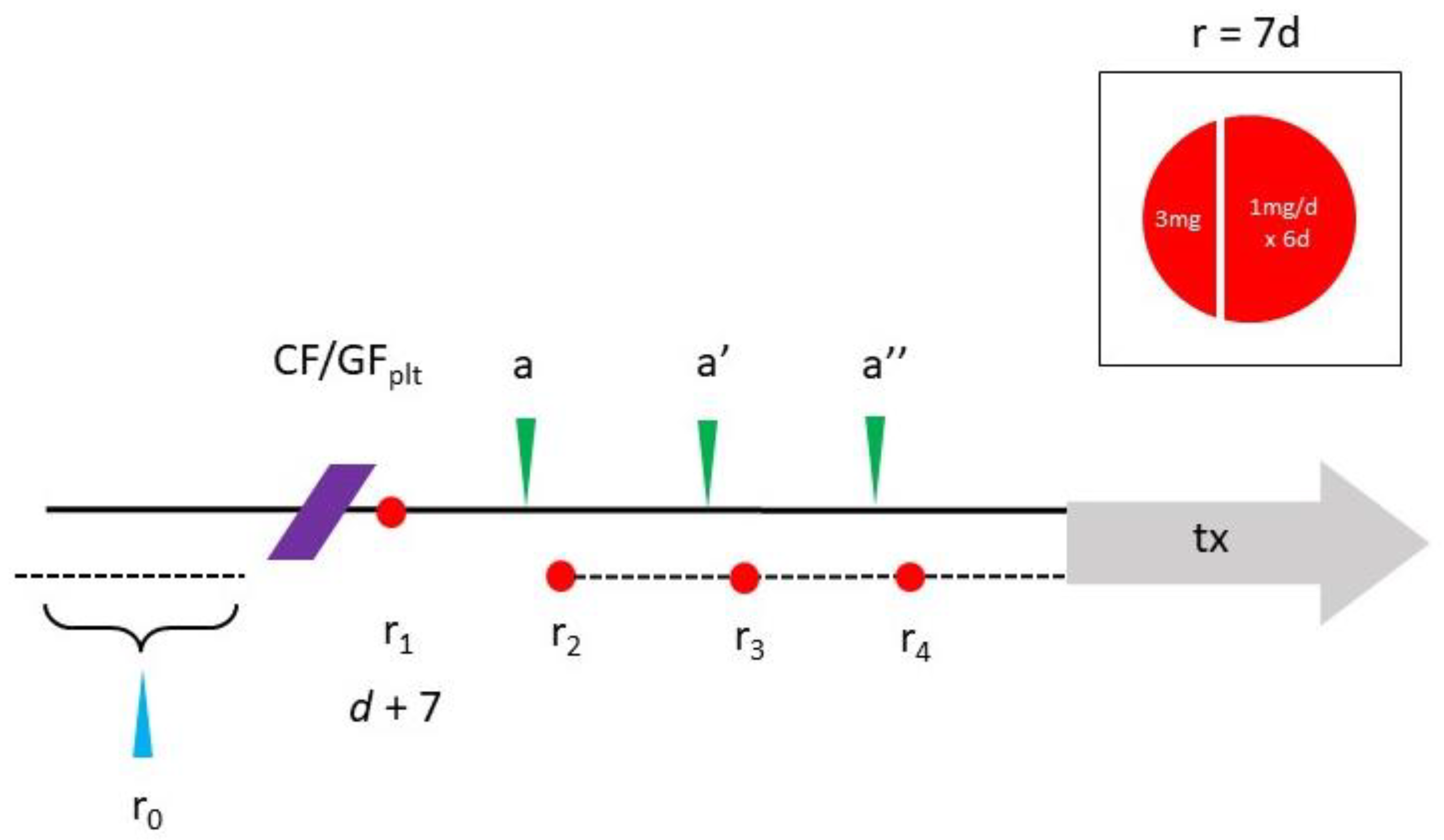
Figure 2.
Schematic for combined intraovarian condensed PLT growth factors (CF/GF) and oral cyclic rapamycin (r x4). Following IRB approval, the study protocol includes enrollment labs (blue arrow) reviewed within 1mo of ovarian injection (purple). Weekly oral rapamycin phased one-week-per-month begins 7d after ovarian injection, where a 3mg loading dose is followed 1g/d for the next 6d (inset). Subsequent testing scheduled monthly (a-a'') allows for close monitoring, intended to improve ovarian reserve sufficient for fertility treatment later (tx).
Figure 2.
Schematic for combined intraovarian condensed PLT growth factors (CF/GF) and oral cyclic rapamycin (r x4). Following IRB approval, the study protocol includes enrollment labs (blue arrow) reviewed within 1mo of ovarian injection (purple). Weekly oral rapamycin phased one-week-per-month begins 7d after ovarian injection, where a 3mg loading dose is followed 1g/d for the next 6d (inset). Subsequent testing scheduled monthly (a-a'') allows for close monitoring, intended to improve ovarian reserve sufficient for fertility treatment later (tx).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



