
Submitted:
09 July 2023
Posted:
10 July 2023
Read the latest preprint version here
Abstract
Autism Spectrum Disorder (ASD) is characterized by varying degrees of difficulty in social interaction and communication. These deficits are often associated with gastrointestinal symptoms, indicating alterations in both intestinal microbiota composition and metabolic activities. The intestinal microbiota influences the function and development of the nervous system. In individuals with ASD, there is an increase in bacterial genera such as Clostridium, as well as species involved in the synthesis of branched-chain amino acids (BCAA) like Prevotella copri. Conversely, decreased amounts of Akkermansia muciniphila, and Bifidobacterium spp. are observed. Epigallocatechin gallate (EGCG) is one of the polyphenols with the greatest beneficial activity on microbial growth and its consumption is associated with reduced psychological distress. Therefore, the objective of this review is to analyze how EGCG and its metabolites can improve the microbial dysbiosis present in ASD and its impact on the pathology. The analysis reveals that EGCG inhibits the growth of pathogenic bacteria like Clostridium perfringens and Clostridium difficile. Moreover, it increases the abundance of Bifidobacterium spp. and Akkermansia spp. As a result, EGCG demonstrates efficacy in increasing the production of metabolites involved in maintaining epithelial integrity and improving brain function. This identifies EGCG as highly promising for complementary treatment in ASD.
Keywords:
Autism Spectrum Disorder (ASD)
; microbiota
; epigallocatechin-3-gallate (EGCG)
; inflammation
; oxidative stress
1. Introduction
Autism Spectrum Disorder (ASD) is a highly heterogeneous and complex disorder, characterized by two major groups of core symptoms: persistent deficits in social communication and interaction, and restricted and repetitive patterns of behavior, interests, or activities [1,2,3,4,5]. Current epidemiology statistics indicates that the disorder affects approximately 1 in every 160 children worldwide [6].
Characteristic deficits are often associated with a range of gastrointestinal symptoms such as abdominal pain, diarrhea, or constipation [7,8]. Several studies have found that both the composition of the intestinal microbiota and metabolic activities may be altered in individuals with the disorder ASD [9,10]. Therefore, several authors concur that disruptions in the microbiota and intestinal microbiome, i.e., the collection of microorganisms present in the human gastrointestinal tract and their respective genomes [3,11], could trigger many of the gastrointestinal issues experienced by children with ASD, as well as exacerbate some of the core symptoms of the disorder [12,13].
Microbial colonization begins in infancy through the acquisition of maternal microbiota during vaginal delivery [3]. Subsequently, beneficial microorganisms feed on breast milk, which has a high content of oligosaccharides [14]. Similarly, the composition of the microbiota in the early years may be subject to alterations influenced by the delivery method, hygiene habits and feeding practices and routines. Among these factors, formula feeding has a particularly significant impact [3].
It has been shown that the intestinal microbiota affects the function and development of the immune, metabolic and nervous systems. Regarding the immune and metabolic systems, intestinal microbiota and its metabolites impact host physiology by regulating the function of the intestinal barrier, redox and mitochondrial metabolism, and mucosal inflammatory response through the regulation of intestinal lymphocytes that provide resistance against potential pathogens [15,16,17,18,19,20]. On the other hand, intestinal microbiota can influence neurochemistry, function, gene expression and the development of the central nervous system (CNS) through the gut-brain axis, which represents a bidirectional link between the cognitive and emotional functions of the CNS and peripheral intestinal function [15,21,22,23]. It is worth noting that the genus Bifidobacterium can metabolize gamma-aminobutyric acid (GABA), Lactobacillus spp. can metabolize acetylcholine, Bacillus spp. and Serratia spp., dopamine, and Escherichia spp. and Saccharomyces spp., noradrenaline. All these neurotransmitters are essential for the proper functioning of the nervous system as they can enter circulation and directly affect neural processes throughout the body, including the brain [3,24]. Additionally, intestinal proinflammatory cytokines, particularly TNF and IL-6, can have an impact on the brain because they make the blood-brain barrier (BBB) more permeable, allowing peripheral immune cells to enter the brain and stimulating brain cells to produce additional proinflammatory mediators [25,26].
All of this has led to the hypothesis that the development of symptoms related to ASD may be influenced by the disturbance of the gut-brain-microbiota axis caused by changes in the intestinal microbiota.
2. Alterations in the Intestinal Microbiota in ASD
The Firmicutes (40-60%) and Bacteroidetes (20-40%) are the two major phyla of bacteria in the healthy human intestinal flora, followed by Proteobacteria, Actinomycetes, Clostridium spp., and Verruciformis [3,27]. However, compared to healthy population, increased levels of bacterial genera such as Clostridium, Desulfovibrio and Ruminococcus have been observed [15,28], in ASD, along with species that synthesize BCAA such as Bacteroides vulgatus and Prevotella copri. On the other hand, lower quantities of Bacteroides fragilis, Akkermansia muciniphila, Bifidobacterium spp. and Enterococcus spp. have been found [29,30]. Additionally, the ratio of the Escherichia/Shigella genera is altered, as Shigella spp. is present at higher levels, while Escherichia coli is decreased [31].
It should be noted that, despite the consensus regarding these alterations, it is important to consider that not all studies quantifying the microbiota in ASD analyze the same species or genera, leading to some heterogeneity in the results, and various studies emphasize the relationship between the composition of the human intestinal microbiota and the individual's diet, which can lead to variations in composition depending on the region where the analyses were conducted [32]. In fact, a study in the United States highlighted that children with ASD had lower levels of Bifidobacterium spp. and Prevotella spp., but higher levels of Lactobacillus spp., compared to healthy children [33]. Another study in China described that children with ASD had higher levels of the Actinobacteria and Proteobacteria phyla than the control group [34].
Furthermore, the review conducted by Ho et al. (2020) concluded that although the results of different studies are not entirely consistent [35], some articles indicate that individuals with ASD exhibit a decrease in the percentage of the phylum Bacteroidetes [10,35] or no significant difference [31,36,37] compared to the control group of healthy individuals. This could be due to the fact that within the phylum Bacteroidetes there are bacteria of the genus Bacteroides spp. that are increased, such as B. vulgatus, while others are decreased, such as B. fragilis, potentially leading to a compensatory effect that results in an unclear profile [30].
The genus Prevotella presents a more variable pattern. In the aforementioned review, the authors noted a lower relative abundance of Prevotella spp. in children diagnosed with ASD compared to the control group. However, multiple studies indicate an elevated prevalence of Prevotella spp. in children with ASD [15,30,31,38] and this variability could be due to dietary factors. Filippo et al. (2010) compared the composition of Prevotella spp. in the microbiota of children following European and African diets and observed a significant increase in Prevotella spp. in the microbiota of African children, likely due to their consumption of a grain-rich diet [36].
Finally, to establish a correlation between gastrointestinal issues and behavioral problems with the microbiota, the high growth rates of Clostridium histolyticum, Clostridium difficile, Clostridium perfringens and Sutterella spp., the alteration in the Escherichia/Shigella genera ratio, and the decreased Bacteroidetes/Firmicutes phylum ratio have been identified as factors associated with gastrointestinal problems. Furthermore, the relative abundance of Desulfovibrio spp., Clostridium spp. and Bacteroides vulgatus has been linked to behavioral disorders [40].
3. Polyphenols as a Therapeutic Alternative to ASD.
ASD lacks a medical cure, and understanding molecular pathogenic mechanisms is crucial for proposing alternative therapies to current medications used for treating autism-related symptoms [41]. Given disruptions in the microbiota, exploring the microbiota-gut-brain axis is essential to modify disease development. Polyphenols, as bioactive dietary compounds, show great promise in this regard. These compounds act in the intestine, reducing inflammation and modulating the microbiota and its metabolites. They also have neuroprotective effects in the brain by crossing the BBB [42].
Polyphenolic antioxidants capable capable of targeting the intestine and modulating the intestinal microbiota, while reducing intestinal inflammation, have been shown to influence memory, cognition, mood and behavior, thereby contributing to the prevention and treatment of various brain disorders [25]. Polyphenols have garnered attention for their ability to stimulate certain gut bacteria involved in phenolic compound utilization and metabolism, making them a proposed treatment option for neurological disorders [40,41].
Polyphenolic antioxidants capable of targeting the intestine and modulating the intestinal microbiota, while reducing intestinal inflammation, have been shown to influence memory, cognition, mood and behavior, thereby contributing to the prevention and treatment of various brain disorders [25]. Polyphenols have garnered attention for their ability to stimulate certain gut bacteria involved in phenolic compound utilization and metabolism, making them a proposed treatment option for neurological disorders [43,44]. Polyphenols are the most ubiquitous phytochemicals in the human diet, with a total daily intake of approximately 1 g [45]. These phenolic compounds, as secondary metabolites in plants, primarily protect them against various aggressions or infections caused by bacteria, insects or viruses [43,44]. These protective properties in plants are expected to be beneficial for humans as well. Polyphenolic compounds found in green and black tea are the most potent inhibitors of microbial growth.
These compounds include epigallocatechin gallate (EGCG),, epicatechin gallate, epigallocatechin, catechin gallate, epicatechin and catechin [46]. They have shown significant efficacy in inhibiting the growth of pathogens such as Helicobacter pylori [47], Staphylococcus aureus, Escherichia coli O157:H7 [48,49], Salmonella typhimurium DT104, Listeria monocytogenes, methicillin-resistant Staphylococcus aureus [50,51] and Pseudomonas aeruginosa [52]. They have also been effective against viruses like HIV and Epstein-Barr [53,54].
Among all the polyphenols, several studies highlighted by Duda-Chodak et al. (2015) emphasize the significant role of EGCG [55]. EGCG is the primary catechin in green tea, but it is also found in lower concentrations in certain plant-based foods, such as apples, peaches, kiwis, blackberries, pears and nuts like pistachios, hazelnuts and walnuts [56].
Hence, considering the advantageous effects observed on the microbiota from polyphenols, including EGCG, a comprehensive mini-review was proposed. This review was based on a thorough scientific search of published studies in PubMed, without any limitations on publication date. The objective was to examine the potential advantages that EGCG might offer to individuals with ASD. The search was from January to April 2023 and utilized MeSH descriptors like "epigallocatechin gallate," "autism spectrum disorder," "metabolism," "microbiota," and "fatty acids," combined using the boolean operator "AND.".
3.1. Possible Role of EGCG in the Intestinal Microbiota of Patients with ASD
To date, there is a lack of published studies that have used EGCG in ASD. However, there are cellular and animal studies [57,58] that clearly demonstrate the potential of this antioxidant for clinical treatment of ASD. Additionally, in recent years several studies have been published employing other polyphenols that are structurally similar to EGCG, such as luteolin or quercetin, which have achieved evident clinical benefits in the disease [59,60,61,62]. Its consumption is linked to reduced psychological distress [63], possibly by inhibiting the growth of specific pathogenic bacteria such as Clostridium perfringens, Clostridium difficile [64], Bacteroides ovatus, and enterobacteria (Salmonella spp., Escherichia coli, Yersinia pestis, Klebsiella spp., Shigella spp. and Eggerthella spp.) [49,65,66,67]. Furthermore, in the intestinal microbiota, EGCG increases the abundance of Bifidobacterium spp. determined through in vitro studies [65] or in Drosophila models of Parkinson’s disease [66]. EGCG also increases in vitro bacteria of the Bacteroides genus (Bacteroides uniformis, Bacteroides stercoris, Bacteroides thetaiotaomicron and Bacteroides cellulosilyticus), and Lachnoclostridium spp. [65 in Male C57BL/6N mice Akkermansia spp. [67] and in ovariectomized (OVX) mice fed a high fat diet (HFD) Prevotella spp. [68].
A substantial portion of the vital microorganisms in the fecal microbiota of breastfed infants consists of species from the Bifidobacterium genus [69]. Bifidobacterium spp. has psychobiotic effects that help reduce anxiety, stress, and other depressive behaviors, particularly strains B. longum and B. breve[70,71]. This bacterial genus is one of the initial colonizers of the newborn's intestines, and an imbalance in Bifidobacterium spp. can affect infant neurodevelopment [71]. Administering EGCG to increase Bifidobacterium spp. has been found to enhance the intestinal microbiota, as supported by previous animal studies that showed an elevated abundance of Bifidobacterium spp. in response to a diet enriched with EGCG[72].
Bacteria within the Bacteroides genus play a crucial role breaking down complex molecules, particularly carbohydrates, and confer advantages to their hosts by impeding the colonization of potential pathogens in the digestive tract [73,74]. It has been seen that there are bacteria capable of modulating the immune system. For instance, B. fragilis has been shown to restore the balance of T-cell populations in mice affected by ASD [75], while B. uniformis enhances the production of anti-inflammatory cytokines and improve metabolic and immune dysfunction [65]. Moreover, this genus can improve social behaviors and physiological abnormalities in individuals with ASD [76], indicating that an increase in Bacteroides abundance caused by EGCG could enhance the quality of life in patients. Additionally, this polyphenol can reduce B. ovatus, the main intestinal commensal responsible for a systemic antibody response in inflammatory bowel disease [77]. These findings suggest that even within the same genus, EGCG may exert different effects on bacterial species.
Prevotella spp. are bacteria that exist in the human microbiota, and are responsible for degrading plant polysaccharides and participating in the synthesis of vitamin B1 [78]. One specific species within the Prevotella genus is Prevotella copri, which has been linked to enhanced glucose and insulin tolerance. It is commonly found in individuals who consume a diet rich in fiber, indicating a strong association between the effects of this bacterium and dietary habits. Within the Prevotella genus, Prevotella copri can be found. This species has been associated with improved glucose and insulin tolerance and is commonly found in individuals who follow a fiber-rich diet, suggesting a strong connection between the effects of this bacterium and dietary patterns [79,80,81,82]. Consequently, both the bacteria within of the Prevotella genus and their metabolic activity are significantly relevant to ASD: on one hand, children with ASD present higher risk of obesity, which implies insulin resistance [83], an on the other hand, these children exhibit selective eating patterns, with notable preference for high amounts of simple carbohydrates and low intake of fiber [84]. This dietary behavior is associated with lower levels of Prevotella spp. within the microbiota. Additionally, it is worth noting that individuals with ASD have been reported to have low levels of vitamin B1, which plays a significant role in antioxidant activity [85].
In relation to the Lachnoclostridium genus, it has been seen to possess anti-inflammatory properties and contribute to the maintenance of intestinal balance by producing butyric acid [86]. This metabolite aids in the elimination of intestinal gases and its absence is associated with irritable bowel syndrome (IBS) [87]. Therefore, the increase of Lachnoclostridium spp. mediated by EGCG [56] can be seen as a positive impact on the microbiota of ASD patients.
EGCG modulates intestinal microbiota and its metabolites, enriching the population of short-chain fatty acid (SCFA)-producing bacteria such as Akkermansia spp., resulting in an improvement in the production of acetate, propionate and butyrate in a sodium dextran sulfate (SDS)-induced colitis mouse model, where levels of these metabolites were significantly reduced [88,89]; this promotes an anti-inflammatory and antioxidant state in the intestine. [88]. Akkermansia spp. utilizes intestinal epithelial mucin as a source of energy. By degrading mucin, it releases nutrients such as monosaccharides, amino acids and SCFAs, which are used by other bacteria in the microbiota, stimulating their metabolic functions [90]. Akkermansia spp. is involved in the regulation of glucose metabolism and adipose tissue homeostasis, therefore, the fact that EGCG increases the levels of Akkermansia muciniphila improves intestinal dysbiosis and enhances barrier integrity [67,91,92,93].
Moreover, among the bacteria reduced by EGCG, Clostridia stand out as a group of Gram-positive bacilli that, when in excess, can lead to infection in the large intestine [28]. Clostridia produce significant compounds such as butyrate or butyric acid, along with other SCFAs generated from the fermentation of dietary fiber. [94]. Butyrate is an essential metabolite in the human colon as it is the preferred energy source for colonic epithelial cells. It contributes to maintaining intestinal barrier functions and has immunomodulatory and anti-inflammatory properties [95]. However, excessively high levels of this metabolite in the intestinepromote intestinal permeability, which may allow the passage of toxic substances into the bloodstream, potentially leading to inflammation [94]. Thus, EGCG's inhibitory effect on this bacterial group, as observed in rats and resulting in decreased levels of Clostridium spp., balancing their levels, beneficial [72]. This is particularly relevant for Clostridium perfringens, as it significantly impacts disease symptomatology, including gastrointestinal issues. Both the bacterium itself and the gene that produces its toxin (CPB2) have been linked to gastrointestinal complications in ASD and are correlated with disease severity. Moreover, the isolated species from children with ASD show greater antibiotic resistance compared to healthy children [96]. Conversely, the increase of Clostridium difficile has a notably negative impact on ASD, and effective treatments against this bacterium using oral vancomycin have shown improvements in behavior and communication [97,98]. Therefore, the reduction of this strain mediated by EGCG could be beneficial for the clinical presentation of children affected by ASD.
EGCG treatment also leads to a decrease in enterobacteria, which are commonly present in the body but can cause infections when their growth is uncontrolled. Certain species have been linked to specific pathologies. Specifically, Salmonella typhi is responsible for typhoid fever, Shigella dysenteriae is the causative agent of bacillary dysentery and causes infantile gastroenteritis, and Yersinia pestis causes plague [99].
Lastly, it is important to highlight that specific bacteria increased by EGCG, such as Lactobacillus acidophilus, Bifidobacterium longum, Akkermansia muciniphila and Prevotella ruminicola, restore the balance of the intestinal microbiota when included to the diet, reducing oxidative stress in the intestine and the brain [100] (Table 1).
3.2. Outlook on the Anti-Inflammatory and Antioxidant Activity of EGCG in Autism. Neuroprotective Role
Immune system dysregulation, inflammation, and increased oxidative stress are significant factors in ASD [101], which is directly linked to dysbiosis. Comorbid inflammatory conditions associated with immune dysregulation closely contribute to the emergence and progression of clinical features in children with ASD [102,103,104]. Cytokines, small proteins regulating inflammation and neurological development. Increased levels of some of them, particularly, IL-4 o IL-10, have been seen in children with ASD[105]. Notable changes in interferon-α (IFN-α), interleukin-7 (IL-7), IFN-γ-inducible protein-10, and IL-8 are particularly relevant to the disorder's pathogenesis [106].
IL-1β and IL-10 levels produced by blood monocytes, along with the IL-1β/IL-10 ratio and miRNA expression, are relevant to innate immunity [107,108]. All these alterations closely associate with social behavioral impairments and cognitive development in children with ASD [109], which could be due to a brain dysfunction associated with the proper production of growth factors and neurogenesis [110]. Specifically, the brain-derived neurotrophic factor (BDNF) plays important roles in the formation, branching and connectivity of synaptic connections during development [111,112], as well as in synaptic plasticity, as it is involved in learning and memory [113,114].
Oxidative stress is directly linked to the intestinal microbiota, particularly in children with ASD [115]. The fermentation of dietary fibers and resistant starch in the gut generates SCFAs [116], with propionate being associated with gastrointestinal issues and neuroinflammation in ASD [117]. Consequently, oxidative stress, closely tied to dysbiosis, plays a crucial role in the neuroinflammatory response and the development of ASD [118,119,120,121,122]. Children with autism exhibit differences in antioxidant capacity and oxidative stress-related metabolites compared to healthy children [123]. Imbalance in glutathione levels and decreased storage capacity contribute to increased susceptibility to oxidative stress in autistic children [124]. Moreover, increased free radical production, influenced by amyloid precursor proteins (APP), is observed in various human disorders, including autism [125,126,127].
Polyphenols, particularly EGCG, play a role as antioxidants and promote neurogenesis and plasticity in a Down syndrome mouse model [128], showing potential in alleviating autistic symptoms. Other polyphenols such as cyanidin-3-glucosidereduce intestinal inflammation in human intestinal cells, affecting inflammatory cascades like nuclear factor kappa B (NF-kB), activator protein-1 (AP-1) and Janus kinase-signal transducer and activator of transcription (JAK/STAT) [129]. Furthermore, resveratrol in an in vitro modulates inflammatory markers including nitric oxide (NO), tumor necrosis factor-alpha (TNF-α), ionized calcium-binding adapter molecule 1 (Iba1), prostaglandin E2 (PGE2), inducible nitric oxide synthase (iNOS) and cyclooxygenase-(COX-2) [130]. Moreover, a study on rat pups with autistic traits found that a polyphenol-probiotic complex reversed autistic behaviors and modulated biochemical changes in IL-6, TNF-α, BDNF, 5-HT, AchE, and the granular layer, which is supported by all this evidence [131].
Therefore, when analyzing the activity of various polyphenols in inflammation and oxidative stress, these molecules have demonstrated high efficacy in regulating APP levels [132]. . Its ability to cross the BBB (based on the hBMEC model, which utilizes human-derived brain endothelial cells), plays a significant role in protecting cortical neurons against oxidative stress-induced cell death. Comparatively, EGCG demonstrated rapid BBB permeation, while cyanidin-3-glucoside (C3G) showed slower permeation, and quercetin did not cross the BBB. This makes EGCG particularly promising as a neuroprotector among various polyphenols [133]. In relation to ASD, IL-8 stands out among the cytokines involved. The administration of oral EGCG reduces its levels through the inhibition of intracellular Ca2+ levels and the activation of ERK1/2 and NF-kappaB pathways. This is becauseit has been seen in the colon that the treatment of TNF-α-stimulated HT29 cells (human colon carcinoma cell lines) with EGCG inhibits IL-8 production by regulating genes in inflammatory pathways, suggesting its potential for preventing or mitigating colonic disorders in autism [134].
Finally, it should be noted that elevated levels of reactive oxygen species (ROS), specifically hydrogen peroxide (H2 O2) and superoxide (O2-), are reported in the intestine of children with ASD as a cause of damage to the epithelial tissue [135,136,137], and EGCG negatively regulates the inflammatory response in inflamed intestinal epithelial cells, largely through a post-transcriptional regulatory mechanism [138].
3.3. Role of EGCG in the Metabolic Activity Associated with Dysbiosis in ASD
EGCG regulates the production of SCFAs [42], which are metabolites involved in the bidirectional communication between the gut and the brain. Succinate and butyrate, are particularly important and can be altered in individuals with autism due to microbial activity [139]. An imbalanced ratio of succinate production/consumption can cause intestinal disturbances and impact the gut-brain axis. [140]. Cheng et al. determined through a review that altered succinate levels have been linked to disrupted calcium homeostasis and dysregulated metabolic functions in autistic individuals [41].
Regarding butyrate, it has been seen that children with ASD have lower levels of fecal butyrate, as well as a decrease in the abundance of taxa producing this metabolite [141]. Butyrate is particularly relevant in ASD as its production mainly derives from the gut microbiome. Butyrate displays potent anti-inflammatory activity that contributes to gut health, regulates intestinal homeostasis , and modulates the expression of neurotransmitter genes [142,143]. Furthermore, it positively modulates the impaired mitochondrial function in ASD, characterized by reduced activity of complex IV in the electron transport chain (ETC) [144], including improvements in oxidative phosphorylation and beta-oxidation. These observations have underscored the significant neuroprotective role of butyrate in children with ASD [145]. In fact, it has recently been seen that maternal treatment with butyrate in the BTBR mouse model of ASD rescues social and partially repetitive behavior deficits in the offspring [146]. Thus, proper butyrate production at adequate levels is crucial, and this depends on the correct development of the gut bacterial community. Oral EGCG delivery on the DSS-induced murine colitis model has been shown to increase both the abundance of Akkermansia muciniphila and its butyrate production [88], potentially reversing the adverse effects resulting from altered butyrate levels in ASD. On the other hand, among the altered fecal metabolite profiles in children with ASD (not present in neurotypical children), there is an increase in p-cresol, caprate, aspartate, and a reduction in GABA, nicotinate, glutamine, and thymine [147]. Among all these metabolites, p-cresol is a uremic toxin produced by certain strains of Clostridium spp., which has negative biological effects and appears to adversely affect the homeostasis of colonic epithelial cells in children with ASD. When present in excess, p-cresol induces DNA damage in vitro and negatively affects the integrity of colonic epithelial cells [148]. In fact, EGCG may have beneficial effects by reversing the activity of this metabolite. In a recent study, it has been seen that an EGCG-enriched diet reduces plasma and urinary concentrations of p-cresol in mice by suppressing the abundance of Firmicutes at the phylum level and Clostridia at the order level [149].
In conclusion, EGCG has an impact on both the intestinal microbiota and variables directly related to the microbiota such as metabolites, inflammation and oxidative stress, which could provide benefits for patients with ASD (Figure 1).
Another noteworthy aspect is the impact of metabolites generated from the interaction between the polyphenol and the intestinal microbiota. Particularly, in rats, the metabolic pathway of EGCG leads to gallic acid (GA) and epigallocatechin (EGC) [150]. EGC is degraded by colonic bacteria to yield microbial ring-fission metabolites, specifically converting to 5-(3',5'-dihydroxyphenyl)-γ-valerolactone (EGC-M5). Then, the bacteria Adlercreutzia equolifaciens MT4s-5 and Flavonifractor plautii MT42 are capable of degrading EGC. The bacterium Adlercreutzia equolifaciens MT4s-5 catalyzes the conversion of EGC to 1-(3,4,5-trihydroxyphenyl)-3-(2,4,6-trihydroxyphenyl)propan-2-ol (EGC-M1), and subsequently, Flavonifractor plautii MT42 converts the propan-2-ol to 5-(3,4,5-trihydroxyphenyl)-γ-valerolactone (EGC-M5) and 4-hydroxy-5-(3,4,5-trihydroxyphenyl)valeric acid (EGC-M4). Similarly, EGC-M5 is produced from 1-(3,5-dihydroxyphenyl)-3-(2,4,6-trihydroxyphenyl)propan-2-ol (EGC-M3), which is formed from EGC by Adlercreutzia equolifaciens MT4s-5 in the presence of hydrogen [120]. A large portion of the formed EGC-M5 is absorbed and undergoes glucuronidation in the intestinal mucosa and/or liver to form EGC-M5 glucuronide (EGC-M5-GlcUA), which is distributed to various tissues through the bloodstream and ultimately excreted in urine [151] (Figure 2).
The microbial ring-fission metabolites of EGCG are found in plasma in both free and conjugated forms [151], and in vitro studies have shown that they may reach the brain parenchyma through the BBB, promoting neuritogenesis and potentially exerting a relevant activity against the degenerative processes of neurodegenerative diseases [151].
Specifically, when assessing the penetration capacity of these metabolites through the BBB, it has been seen that GA has higher permeability than EGCG and EGC, possibly due to its smaller molecular size [153]. Moreover, comparing only the rif[ng-fission metabolites derived from ECG, EGC-M5 demonstrated greater permeability than its conjugate EGC-M5-GlcUA [150]. Some of these ring-fission metabolites also exhibit anti-inflammatory activity [154]. In particular, EGC-M5 has been found to have immunomodulatory properties, as it enhances the activity of CD4+ T cells and the cytotoxic activity of natural killer cells in BALB/c mice [155]. Therefore, it seems evident that the ring-fission metabolites derived from the intestinal microbiota a of catechins demonstrate a protective capacity against various diseases, including neurodegenerative disorders.
3.4. Liposomal EGCG
However, it is crucial to understand the metabolic process and bioavailability of green tea catechins and EGCG, particularly in assessing their biological activity and comprehending their beneficial effects on human health. EGCG presents significantly lower bioavailability compared to other components of catechins [156,157]. Therefore, when assessing the activity of EGCG and its metabolites, it is essential to emphasize that their applications are greatly limited due to their low solubility, bioavailability and stability. The use of liposomal formulations may serve as an effective strategy for their administration in autism. Liposomal delivery aims to improve the poor stability of polyphenols against temperature, light, pH and oxygen [28,29], as well as their low permeability across intestinal membranes, which results in only a small proportion of these compounds remaining available for absorption in the human body after ingestion [29,30]. Both stability and oral bioavailability are enhanced through liposomal encapsulation, as it provides protection against degradation when passing through the gastrointestinal tract [31]. Moreover, nanotechnology can promote controlled release of the polyphenol and modulating the interaction between polyphenols and the microbiota, which also represents an intriguing approach [31], and it has been seen that EGCG has significantly improved stability when formulated as dual-drug loaded PEGylated PLGA nanoparticles (EGCG/AA NPs). Following oral administration in mice, EGCG accumulated in all major organs, including the brain, leading to increased synapse formation and reduction of neuroinflammation in Alzheimer's disease [158]. Furthermore, concerning its metabolites and their administration using nanotechnology, Abbasalipour H. et al. (2022) studied the neuroprotective effects of gallic acid on oxidative stress-induced cognitive impairment and the expression of the Nrf2/Keap1 gene in an autism model. Gallic acid-loaded nanophytosomes (GNP) were administered, and the results revealed improvements in learning and memory deficits by reducing oxidative stress, enhancing antioxidant enzyme activity and modulating the Keap1/Nrf2 gene expression [159].
3.5. Potential Adverse Effects of EGCG
Green tea, and to a lesser extent black tea, are not only the main sources of EGCG but also contain other molecules such as caffeine. It is noteworthy that caffeine consumption has been specifically linked to an increase in sleep disturbances, characteristic of this population [160], as well as the presence of maladaptive behavior in children with ASD [161]. Furthermore, concerning the microbiota in individuals vulnerable to Clostridium difficile infections, an increased risk in such infections has been associated with tea consumption, as it disrupts the normal microbiome and promotes the overgrowth of facultative pathogens [162]. Tea consumption has also been identified as neo f the risk factors for gastroesophageal reflux disease (GERD) [163], which is relevant considering that GERD is one of the gastrointestinal symptoms of commonly described as ASD [164]. Moreover, excessive consumption of EGCG has been found to be hepatotoxic [165,166,167];
Despite these considerations, tea has been consumed worldwide for many centuries, and the intake of its main catechin, EGCG, has been shown to be particularly safe even at doses up to 800 mg/day [168]. Thus, consuming tea at appropriate doses presents significant health benefits and is highly safe in terms of potential adverse effects or contraindications [169].
4. Conclusions
ASD is a complex disorder characterized by inflammatory processes, high oxidative stress and gastrointestinal disturbances. These issues may be attributed to changes in the intestinal microbiome, resulting in microbial dysbiosis in children with ASD. Polyphenols have demonstrated sufficient antioxidant and anti-inflammatory capacity to counteract ASD symptoms. Their role in dysbiosis has also been studied, revealing their ability to regulate and improve the percentages of beneficial microbial species, thereby enhancing the pathology of ASD and promoting gut-brain axis function, consequently improving brain function. In this regard, EGCG appears particularly effective in improving microbial species and increasing the production of metabolites involved in maintaining epithelial integrity. It also demonstrates effectiveness in reducing inflammation directly linked to ASD by influencing specific pro-inflammatory cytokine levels and reducing oxidative stress. Furthermore, it can modulate the altered metabolic production in ASD by decreasing toxic metabolites such as p-cresol. All of these factors have an impact on brain function, which could provide significant benefits for children with ASD. However, it should be noted that these polyphenols have limitations in terms of intestinal assimilation and bioavailability. Hence, when considering the advantages of nanotechnology in overcoming these limitations, it is advisable to administer them in liposomal form as a complementary treatment for ASD.
Consequently, this review identifies EGCG liposomes as a potential adjunctive therapy for children with ASD, aiming to improve their quality of life and alleviate their symptoms.
Author Contributions
Conceptualization, J.E.d.l.R.O. and A.B.C.-S.; Writing—Original Draft Preparation, J.E.d.l.R.O., A.B.C.-S, M.B., B.P.; and C.E.S.-S.; C.M.; P.S.-B.; G.C.; R.B.-B. and M.M. writing—review and editing, J.E.d.l.R.O., A.B.C.-S and M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding, but has been supported by the Foundation of Catholic University of Valencia San Vicente Mártir.
Data Availability Statement
Not applicable due to data privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Taylor, M.J.; Rosenqvist, M.A.; Larsson, H.; Gillberg, C.; D’Onofrio, B.M.; Lichtenstein, P.; Lundström, S. Etiology of Autism Spectrum Disorders and Autistic Traits Over Time. JAMA Psychiatry 2020, 77, 936–943. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Am. Psychiatr. Assoc. 2013. [Google Scholar]
- Hughes, H.K.; Rose, D.; Ashwood, P. The Gut Microbiota and Dysbiosis in Autism Spectrum Disorders. Curr. Neurol. Neurosci. Reports 2018 1811 2018, 18, 1–15. [Google Scholar] [CrossRef]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Rosenberg, C.R.; White, T.; Durkin, M.S.; Imm, P.; Nikolaou, L.; Yeargin-Allsopp, M.; Lee, L.C.; Harrington, R.; Lopez, M.; Fitzgerald, R.T.; Hewitt, A.; Pettygrove, S.; Constantino, J.N.; Vehorn, A.; Shenouda, J.; Hall-Lande, J.; Naarden Braun, K. Van; Dowling, N.F. Prevalence of Autism Spectrum Disorder among Children Aged 8 Years - Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. MMWR Surveill. Summ. 2018, 67, 1–23. [Google Scholar] [CrossRef]
- Grimaldi, R.; Gibson, G.R.; Vulevic, J.; Giallourou, N.; Castro-Mejía, J.L.; Hansen, L.H.; Leigh Gibson, E.; Nielsen, D.S.; Costabile, A. A Prebiotic Intervention Study in Children with Autism Spectrum Disorders (ASDs). Microbiome 2018, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Autism. https://www.who.int/news-room/fact-sheets/detail/autism-spectrum-disorders (accessed 2023-02-26).
- Li, Q.; Zhou, J.M. The Microbiota–Gut–Brain Axis and Its Potential Therapeutic Role in Autism Spectrum Disorder. Neuroscience 2016, 324, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fu, X.; Liao, X.; Li, Y. Effects of Gut Microbial-Based Treatments on Gut Microbiota, Behavioral Symptoms, and Gastrointestinal Symptoms in Children with Autism Spectrum Disorder: A Systematic Review. Psychiatry Res. 2020, 293. [Google Scholar] [CrossRef] [PubMed]
- de Theije, C.G.M.; Wopereis, H.; Ramadan, M.; van Eijndthoven, T.; Lambert, J.; Knol, J.; Garssen, J.; Kraneveld, A.D.; Oozeer, R. Altered Gut Microbiota and Activity in a Murine Model of Autism Spectrum Disorders. Brain. Behav. Immun. 2014, 37, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; De Filippo, C. New Evidences on the Altered Gut Microbiota in Autism Spectrum Disorders. Microbiome 2017, 5, 1–11. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; Fitzgerald, M.G.; Fulton, R.S.; Giglio, M.G.; Hallsworth-Pepin, K.; Lobos, E.A.; Madupu, R.; Magrini, V.; Martin, J.C.; Mitreva, M.; Muzny, D.M.; Sodergren, E.J.; Versalovic, J.; Wollam, A.M.; Worley, K.C.; Wortman, J.R.; Young, S.K.; Zeng, Q.; Aagaard, K.M.; Abolude, O.O.; Allen-Vercoe, E.; Alm, E.J.; Alvarado, L.; Andersen, G.L.; Anderson, S.; Appelbaum, E.; Arachchi, H.M.; Armitage, G.; Arze, C.A.; Ayvaz, T.; Baker, C.C.; Begg, L.; Belachew, T.; Bhonagiri, V.; Bihan, M.; Blaser, M.J.; Bloom, T.; Bonazzi, V.; Paul Brooks, J.; Buck, G.A.; Buhay, C.J.; Busam, D.A.; Campbell, J.L.; Canon, S.R.; Cantarel, B.L.; Chain, P.S.G.; Chen, I.M.A.; Chen, L.; Chhibba, S.; Chu, K.; Ciulla, D.M.; Clemente, J.C.; Clifton, S.W.; Conlan, S.; Crabtree, J.; Cutting, M.A.; Davidovics, N.J.; Davis, C.C.; Desantis, T.Z.; Deal, C.; Delehaunty, K.D.; Dewhirst, F.E.; Deych, E.; Ding, Y.; Dooling, D.J.; Dugan, S.P.; Michael Dunne, W.; Scott Durkin, A.; Edgar, R.C.; Erlich, R.L.; Farmer, C.N.; Farrell, R.M.; Faust, K.; Feldgarden, M.; Felix, V.M.; Fisher, S.; Fodor, A.A.; Forney, L.J.; Foster, L.; Di Francesco, V.; Friedman, J.; Friedrich, D.C.; Fronick, C.C.; Fulton, L.L.; Gao, H.; Garcia, N.; Giannoukos, G.; Giblin, C.; Giovanni, M.Y.; Goldberg, J.M.; Goll, J.; Gonzalez, A.; Griggs, A.; Gujja, S.; Kinder Haake, S.; Haas, B.J.; Hamilton, H.A.; Harris, E.L.; Hepburn, T.A.; Herter, B.; Hoffmann, D.E.; Holder, M.E.; Howarth, C.; Huang, K.H.; Huse, S.M.; Izard, J.; Jansson, J.K.; Jiang, H.; Jordan, C.; Joshi, V.; Katancik, J.A.; Keitel, W.A.; Kelley, S.T.; Kells, C.; King, N.B.; Knights, D.; Kong, H.H.; Koren, O.; Koren, S.; Kota, K.C.; Kovar, C.L.; Kyrpides, N.C.; La Rosa, P.S.; Lee, S.L.; Lemon, K.P.; Lennon, N.; Lewis, C.M.; Lewis, L.; Ley, R.E.; Li, K.; Liolios, K.; Liu, B.; Liu, Y.; Lo, C.C.; Lozupone, C.A.; Dwayne Lunsford, R.; Madden, T.; Mahurkar, A.A.; Mannon, P.J.; Mardis, E.R.; Markowitz, V.M.; Mavromatis, K.; McCorrison, J.M.; McDonald, D.; McEwen, J.; McGuire, A.L.; McInnes, P.; Mehta, T.; Mihindukulasuriya, K.A.; Miller, J.R.; Minx, P.J.; Newsham, I.; Nusbaum, C.; Oglaughlin, M.; Orvis, J.; Pagani, I.; Palaniappan, K.; Patel, S.M.; Pearson, M.; Peterson, J.; Podar, M.; Pohl, C.; Pollard, K.S.; Pop, M.; Priest, M.E.; Proctor, L.M.; Qin, X.; Raes, J.; Ravel, J.; Reid, J.G.; Rho, M.; Rhodes, R.; Riehle, K.P.; Rivera, M.C.; Rodriguez-Mueller, B.; Rogers, Y.H.; Ross, M.C.; Russ, C.; Sanka, R.K.; Sankar, P.; Fah Sathirapongsasuti, J.; Schloss, J.A.; Schloss, P.D.; Schmidt, T.M.; Scholz, M.; Schriml, L.; Schubert, A.M.; Segata, N.; Segre, J.A.; Shannon, W.D.; Sharp, R.R.; Sharpton, T.J.; Shenoy, N.; Sheth, N.U.; Simone, G.A.; Singh, I.; Smillie, C.S.; Sobel, J.D.; Sommer, D.D.; Spicer, P.; Sutton, G.G.; Sykes, S.M.; Tabbaa, D.G.; Thiagarajan, M.; Tomlinson, C.M.; Torralba, M.; Treangen, T.J.; Truty, R.M.; Vishnivetskaya, T.A.; Walker, J.; Wang, L.; Wang, Z.; Ward, D.V.; Warren, W.; Watson, M.A.; Wellington, C.; Wetterstrand, K.A.; White, J.R.; Wilczek-Boney, K.; Wu, Y.; Wylie, K.M.; Wylie, T.; Yandava, C.; Ye, L.; Ye, Y.; Yooseph, S.; Youmans, B.P.; Zhang, L.; Zhou, Y.; Zhu, Y.; Zoloth, L.; Zucker, J.D.; Birren, B.W.; Gibbs, R.A.; Highlander, S.K.; Methé, B.A.; Nelson, K.E.; Petrosino, J.F.; Weinstock, G.M.; Wilson, R.K.; White, O. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Iovene, M.R.; Bombace, F.; Maresca, R.; Sapone, A.; Iardino, P.; Picardi, A.; Marotta, R.; Schiraldi, C.; Siniscalco, D.; Serra, N.; de Magistris, L.; Bravaccio, C. Intestinal Dysbiosis and Yeast Isolation in Stool of Subjects with Autism Spectrum Disorders. Mycopathologia 2017, 182, (3–4). [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Kapoor, A.; Verma, A.; Ambatipudi, K. Functional Significance of Different Milk Constituents in Modulating the Gut Microbiome and Infant Health. J. Agric. Food Chem. 2022, 70, 3929–3947. [Google Scholar] [CrossRef]
- Ding, H.; Yi, X.; Zhang, X.; Wang, H.; Liu, H.; Mou, W.W. Imbalance in the Gut Microbiota of Children With Autism Spectrum Disorders. Front. Cell. Infect. Microbiol. 2021, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal Gut Microbiota Modulates Brain Development and Behavior. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the Microbiota and the Immune System. Science (80-. ). 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Frye, R.E.; Melnyk, S.; Macfabe, D.F. Unique Acyl-Carnitine Profiles Are Potential Biomarkers for Acquired Mitochondrial Disease in Autism Spectrum Disorder. Transl. Psychiatry 2013, 3. [Google Scholar] [CrossRef]
- Slattery, J.; Macfabe, D.F.; Frye, R.E. The Significance of the Enteric Microbiome on the Development of Childhood Disease: A Review of Prebiotic and Probiotic Therapies in Disorders of Childhood. Clin. Med. Insights Pediatr. 2016, 10, CMPed–S38338. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.; Li, H.; Li, B.; Duan, G.; Zhu, C. The Role of Probiotics in Children with Autism Spectrum Disorders: A Study Protocol for a Randomised Controlled Trial. PLoS One 2022, 17. [Google Scholar] [CrossRef]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-Gut-Microbe Communication in Health and Disease. Front. Physiol. 2011, 2 DEC. [Google Scholar] [CrossRef]
- Jiang, C.; Li, G.; Huang, P.; Liu, Z.; Zhao, B. The Gut Microbiota and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 58, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric Acid Production by Culturable Bacteria from the Human Intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. Polyphenols in the Management of Brain Disorders: Modulation of the Microbiota-Gut-Brain Axis. Adv. Food Nutr. Res. 2020, 91, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Powell, N.; Walker, M.M.; Talley, N.J. The Mucosal Immune System: Master Regulator of Bidirectional Gut–Brain Communications. Nat. Rev. Gastroenterol. Hepatol. 2017 143 2017, 14, 143–159. [Google Scholar] [CrossRef]
- Sun, Q.; Cheng, L.; Zhang, X.; Wu, Z.; Weng, P. The Interaction between Tea Polyphenols and Host Intestinal Microorganisms: An Effective Way to Prevent Psychiatric Disorders. Food Funct. 2021, 12, 952–962. [Google Scholar] [CrossRef]
- Argou-Cardozo, I.; Zeidán-Chuliá, F. Clostridium Bacteria and Autism Spectrum Conditions: A Systematic Review and Hypothetical Contribution of Environmental Glyphosate Levels. Med. Sci. 2018, 6, 29. [Google Scholar] [CrossRef]
- Xu, M.; Xu, X.; Li, J.; Li, F. Association between Gut Microbiota and Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Front. Psychiatry 2019, 10. [Google Scholar] [CrossRef]
- Zou, R.; Xu, F.; Wang, Y.; Duan, M.; Guo, M.; Zhang, Q.; Zhao, H.; Zheng, H. Changes in the Gut Microbiota of Children with Autism Spectrum Disorder. Autism Res. 2020, 13, 1614–1625. [Google Scholar] [CrossRef]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal Microbiota and Metabolome of Children with Autism and Pervasive Developmental Disorder Not Otherwise Specified. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Andreo-Martínez, P.; García-Martínez, N.; Sánchez-Samper, E.P.; Martínez-González, A.E. An Approach to Gut Microbiota Profile in Children with Autism Spectrum Disorder. Environ. Microbiol. Rep. 2020, 12, 115–135. [Google Scholar] [CrossRef]
- Tomova, A.; Husarova, V.; Lakatosova, S.; Bakos, J.; Vlkova, B.; Babinska, K.; Ostatnikova, D. Gastrointestinal Microbiota in Children with Autism in Slovakia. Physiol. Behav. 2015, 138, 179–187. [Google Scholar] [CrossRef]
- Xie, X.; Li, L.; Wu, X.; Hou, F.; Chen, Y.; Shi, L.; Liu, Q.; Zhu, K.; Jiang, Q.; Feng, Y.; Xiao, P.; Zhang, J.; Gong, J.; Song, R. Alteration of the Fecal Microbiota in Chinese Children with Autism Spectrum Disorder. Autism Res. 2022, 15, 996–1007. [Google Scholar] [CrossRef]
- Williams, B.L.; Hornig, M.; Buie, T.; Bauman, M.L.; Cho Paik, M.; Wick, I.; Bennett, A.; Jabado, O.; Hirschberg, D.L.; Lipkin, W.I. Impaired Carbohydrate Digestion and Transport and Mucosal Dysbiosis in the Intestines of Children with Autism and Gastrointestinal Disturbances. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Gondalia, S.V.; Palombo, E.A.; Knowles, S.R.; Cox, S.B.; Meyer, D.; Austin, D.W. Molecular Characterisation of Gastrointestinal Microbiota of Children with Autism (with and without Gastrointestinal Dysfunction) and Their Neurotypical Siblings. Autism Res. 2012, 5, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Son, J.S.; Zheng, L.J.; Rowehl, L.M.; Tian, X.; Zhang, Y.; Zhu, W.; Litcher-Kelly, L.; Gadow, K.D.; Gathungu, G.; Robertson, C.E.; Ir, D.; Frank, D.N.; Li, E. Comparison of Fecal Microbiota in Children with Autism Spectrum Disorders and Neurotypical Siblings in the Simons Simplex Collection. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.K.H.; Tong, V.J.W.; Syn, N.; Nagarajan, N.; Tham, E.H.; Tay, S.K.; Shorey, S.; Tambyah, P.A.; Law, E.C.N. Gut Microbiota Changes in Children with Autism Spectrum Disorder: A Systematic Review. Gut Pathog. 2020, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of Diet in Shaping Gut Microbiota Revealed by a Comparative Study in Children from Europe and Rural Africa. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Nogay, N.H.; Nahikian-Nelms, M. Can We Reduce Autism-Related Gastrointestinal and Behavior Problems by Gut Microbiota Based Dietary Modulation? A Review. Nutr. Neurosci. 2021, 24, 327–338. [Google Scholar] [CrossRef]
- Cheng, N.; Rho, J.M.; Masino, S.A. Metabolic Dysfunction Underlying Autism Spectrum Disorder and Potential Treatment Approaches. Front. Mol. Neurosci. 2017, 10, 34. [Google Scholar] [CrossRef]
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. Polyphenols as Food Bioactive Compounds in the Context of Autism Spectrum Disorders: A Critical Mini-Review. Neurosci. Biobehav. Rev. 2019, 102, 290–298. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Navajas-Porras, B.; López-Maldonado, A.; Hinojosa-Nogueira, D.; Pastoriza, S.; Rufián-Henares, J.Á. Green Tea and Its Relation to Human Gut Microbiome. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Liu, Z.; Bruins, M.E.; Ni, L.; Vincken, J.P. Green and Black Tea Phenolics: Bioavailability, Transformation by Colonic Microbiota, and Modulation of Colonic Microbiota. J. Agric. Food Chem. 2018, 66, 8469–8477. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.; Tomás-Barberán; González-Sarrías, A.; García-Villalba, R. Dietary Polyphenols: Metabolism and Health Effects; Wiley-Blackwell: Murcia, 2020. [Google Scholar]
- Peterson, J.; Dwyer, J.; Bhagwat, S.; Haytowitz, D.; Holden, J.; Eldridge, A.L.; Beecher, G.; Aladesanmi, J. Major Flavonoids in Dry Tea. J. Food Compos. Anal. 2005, 18, 487–501. [Google Scholar] [CrossRef]
- Ankolekar, C.; Johnson, D.; Pinto, M.D.S.; Johnson, K.; Labbe, R.; Shetty, K. Inhibitory Potential of Tea Polyphenolics and Influence of Extraction Time against Helicobacter Pylori and Lack of Inhibition of Beneficial Lactic Acid Bacteria. J. Med. Food 2011, 14, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Chung, H.S. Antibacterial Activities of Phenolic Components from Camellia Sinensis L. on Pathogenic Microorganisms. Prev. Nutr. Food Sci. 2007, 12, 135–140. [Google Scholar] [CrossRef]
- Nakayama, M.; Shigemune, N.; Tsugukuni, T.; Jun, H.; Matsushita, T.; Mekada, Y.; Kurahachi, M.; Miyamoto, T. Mechanism of the Combined Anti-Bacterial Effect of Green Tea Extract and NaCl against Staphylococcus Aureus and Escherichia Coli O157:H7. Food Control 2012, 25, 225–232. [Google Scholar] [CrossRef]
- Kohda, C.; Yanagawa, Y.; Shimamura, T. Epigallocatechin Gallate Inhibits Intracellular Survival of Listeria Monocytogenes in Macrophages. Biochem. Biophys. Res. Commun. 2008, 365, 310–315. [Google Scholar] [CrossRef]
- Si, W.; Gong, J.; Tsao, R.; Kalab, M.; Yang, R.; Yin, Y. Bioassay-Guided Purification and Identification of Antimicrobial Components in Chinese Green Tea Extract. J. Chromatogr. A 2006, 1125, 204–210. [Google Scholar] [CrossRef]
- Bancirova, M. Comparison of the Antioxidant Capacity and the Antimicrobial Activity of Black and Green Tea. Food Res. Int. 2010, 43, 1379–1382. [Google Scholar] [CrossRef]
- Liu, S.; Lu, H.; Zhao, Q.; He, Y.; Niu, J.; Debnath, A.K.; Wu, S.; Jiang, S. Theaflavin Derivatives in Black Tea and Catechin Derivatives in Green Tea Inhibit HIV-1 Entry by Targeting Gp41. Biochim. Biophys. Acta 2005, 1723, 270–281. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tsai, H.L.; Peng, C.W. EGCG Debilitates the Persistence of EBV Latency by Reducing the DNA Binding Potency of Nuclear Antigen 1. Biochem. Biophys. Res. Commun. 2012, 417, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of Dietary Compounds, Especially Polyphenols, with the Intestinal Microbiota: A Review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.D. Epigallocatechin-3-Gallate (EGCG): Chemical and Biomedical Perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Trovò, L.; Fuchs, C.; De Rosa, R.; Barbiero, I.; Tramarin, M.; Ciani, E.; Rusconi, L.; Kilstrup-Nielsen, C. The Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCG) Restores CDKL5-Dependent Synaptic Defects in Vitro and in Vivo. Neurobiol. Dis. 2020, 138. [Google Scholar] [CrossRef]
- Kumaravel, P.; Melchias, G.; Vasanth, N.; Manivasagam, T. DOSE-DEPENDENT AMELIORATION OF EPIGALLOCATECHIN-3-GALLATE AGAINST SODIUM VALPROATE INDUCED AUTISTIC RATS. Int. J. Pharm. Pharm. Sci. 2017, 9, 203–206. [Google Scholar] [CrossRef]
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An Open-Label Pilot Study of a Formulation Containing the Anti-Inflammatory Flavonoid Luteolin and Its Effects on Behavior in Children with Autism Spectrum Disorders. Clin. Ther. 2013, 35, 592–602. [Google Scholar] [CrossRef]
- Tsilioni, I.; Taliou, A.; Francis, K.; Theoharides, T.C. Children with Autism Spectrum Disorders, Who Improved with a Luteolin-Containing Dietary Formulation, Show Reduced Serum Levels of TNF and IL-6. Transl. Psychiatry 2015 59 2015, 5, e647–e647. [Google Scholar] [CrossRef]
- Bertolino, B.; Crupi, R.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Beneficial Effects of Co-Ultramicronized Palmitoylethanolamide/Luteolin in a Mouse Model of Autism and in a Case Report of Autism. CNS Neurosci. Ther. 2017, 23, 87–98. [Google Scholar] [CrossRef]
- Nasiry, D.; Khalatbary, A.R. Natural Polyphenols for the Management of Autism Spectrum Disorder: A Review of Efficacy and Molecular Mechanisms. Nutr. Neurosci. 2023. [Google Scholar] [CrossRef]
- Scholey, A.; Downey, L.A.; Ciorciari, J.; Pipingas, A.; Nolidin, K.; Finn, M.; Wines, M.; Catchlove, S.; Terrens, A.; Barlow, E.; Gordon, L.; Stough, C. Acute Neurocognitive Effects of Epigallocatechin Gallate (EGCG). Appetite 2012, 58, 767–770. [Google Scholar] [CrossRef]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of Tea Phenolics and Their Aromatic Fecal Bacterial Metabolites on Intestinal Microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Liu, Z.; De Bruijn, W.J.C.; Bruins, M.E.; Vincken, J.P. Reciprocal Interactions between Epigallocatechin-3-Gallate (EGCG) and Human Gut Microbiota in Vitro. J. Agric. Food Chem. 2020, 68, 9804–9815. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, M.; Xue, J.; Xiang, L.; Li, Y.; Xiao, J.; Xiao, G.; Wang, H.L. EGCG Ameliorates Neuronal and Behavioral Defects by Remodeling Gut Microbiota and TotM Expression in Drosophila Models of Parkinson’s Disease. FASEB J. 2020, 34, 5931–5950. [Google Scholar] [CrossRef] [PubMed]
- Ushiroda, C.; Naito, Y.; Takagi, T.; Uchiyama, K.; Mizushima, K.; Higashimura, Y.; Yasukawa, Z.; Okubo, T.; Inoue, R.; Honda, A.; Matsuzaki, Y.; Itoh, Y. Green Tea Polyphenol (Epigallocatechin-3-Gallate) Improves Gut Dysbiosis and Serum Bile Acids Dysregulation in High-Fat Diet-Fed Mice. J. Clin. Biochem. Nutr. 2019, 65, 34–46. [Google Scholar] [CrossRef]
- Qu, Y.; Wu, Y.; Cheng, W.; Wang, D.; Zeng, L.; Wang, Y.; Li, T.-T.; Zhang, L.; Yang, J.; Sun, L.; Ai, J. Amelioration of Cognitive Impairment Using Epigallocatechin-3-Gallate in Ovariectomized Mice Fed a High-Fat Diet Involves Remodeling with Prevotella and Bifidobacteriales. Frontiers in Pharmacology 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between Lactoferrin and Beneficial Microbiota in Breast Milk and Infant’s Feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Savignac, H.M.; Kiely, B.; Dinan, T.G.; Cryan, J.F. Bifidobacteria Exert Strain-Specific Effects on Stress-Related Behavior and Physiology in BALB/c Mice. Neurogastroenterol. Motil. 2014, 26, 1615–1627. [Google Scholar] [CrossRef]
- Andreo-Martínez, P.; Martínez-Gonzlez, A.E. Una Propuesta de Probiótico Basada En El Bifidobacterium Para Autismo. Rev. Española Nutr. Humana y Dietética 2022, 26. [Google Scholar] [CrossRef]
- Unno, T.; Sakuma, M.; Mitsuhashi, S. Effect of Dietary Supplementation of (-)-Epigallocatechin Gallate on Gut Microbiota and Biomarkers of Colonic Fermentation in Rats. J. Nutr. Sci. Vitaminol. (Tokyo). 2014, 60, 213–219. [Google Scholar] [CrossRef]
- Tarek El-Banna; El-Aziz, A.A.; EL-Mahdy, N.; Samy, Y. Sherris Medical Microbiology, 4th Editio.; McGraw-Hill, 2004.
- Pérez de Rozas Ruiz de Gauna, A.M.; Badiola Sáiz, I.; Castellà Gómez, G.; Universitat Autònoma de Barcelona. Departament de Sanitat i d’Anatomia Animals.; Centre de Recerca de Sanitat Animal.; Institut de Recerca i Tecnologia Agroalimentàries. Utilización de Cepas de Bacteroides Spp. Como Probiótico En Conejos. TDX (Tesis Dr. en Xarxa) 2014.
- Jiang, C.C.; Lin, L.S.; Long, S.; Ke, X.Y.; Fukunaga, K.; Lu, Y.M.; Han, F. Signalling Pathways in Autism Spectrum Disorder: Mechanisms and Therapeutic Implications. Signal Transduct. Target. Ther. 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Lang, J.D.; Yang, J.; Long, B.; Liu, X.D.; Zeng, X.F.; Tian, G.; You, X. Differences of Gut Microbiota and Behavioral Symptoms between Two Subgroups of Autistic Children Based on ΓδT Cells-Derived IFN-γ Levels: A Preliminary Study. Front. Immunol. 2023, 14, 1100816. [Google Scholar] [CrossRef]
- Saitoh, S.; Noda, S.; Aiba, Y.; Takagi, A.; Sakamoto, M.; Benno, Y.; Koga, Y. Bacteroides Ovatus as the Predominant Commensal Intestinal Microbe Causing a Systemic Antibody Response in Inflammatory Bowel Disease. Clin. Diagn. Lab. Immunol. 2002, 9, 54–59. [Google Scholar] [CrossRef]
- Srikantha, P.; Hasan Mohajeri, M. The Possible Role of the Microbiota-Gut-Brain-Axis in Autism Spectrum Disorder. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Huang, K.D.; Asnicar, F.; Fehlner-Peach, H.; Pasolli, E.; Karcher, N.; Armanini, F.; Manghi, P.; Bonham, K.; Zolfo, M.; De Filippis, F.; Magnabosco, C.; Bonneau, R.; Lusingu, J.; Amuasi, J.; Reinhard, K.; Rattei, T.; Boulund, F.; Engstrand, L.; Zink, A.; Collado, M.C.; Littman, D.R.; Eibach, D.; Ercolini, D.; Rota-Stabelli, O.; Huttenhower, C.; Maixner, F.; Segata, N. The Prevotella Copri Complex Comprises Four Distinct Clades Underrepresented in Westernized Populations. Cell Host Microbe 2019, 26, 666–679. [Google Scholar] [CrossRef]
- O’Grady, J.; O’Connor, E.M.; Shanahan, F. Review Article: Dietary Fibre in the Era of Microbiome Science. Aliment. Pharmacol. Ther. 2019, 49, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Matusheski, N.V.; Caffrey, A.; Christensen, L.; Mezgec, S.; Surendran, S.; Hjorth, M.F.; McNulty, H.; Pentieva, K.; Roager, H.M.; Seljak, B.K.; Vimaleswaran, K.S.; Remmers, M.; Péter, S. Diets, Nutrients, Genes and the Microbiome: Recent Advances in Personalised Nutrition. Br. J. Nutr. 2021, 126, 1489–1497. [Google Scholar] [CrossRef]
- Ortega-Santos, C.P.; Whisner, C.M. The Key to Successful Weight Loss on a High-Fiber Diet May Be in Gut Microbiome Prevotella Abundance. J. Nutr. 2019, 149, 2083–2084. [Google Scholar] [CrossRef]
- Hill, A.P.; Zuckerman, K.E.; Fombonne, E. Obesity and Autism. Pediatrics 2015, 136, 1051–1061. [Google Scholar] [CrossRef]
- Sharp, W.G.; Postorino, V.; McCracken, C.E.; Berry, R.C.; Criado, K.K.; Burrell, T.L.; Scahill, L. Dietary Intake, Nutrient Status, and Growth Parameters in Children with Autism Spectrum Disorder and Severe Food Selectivity: An Electronic Medical Record Review. J. Acad. Nutr. Diet. 2018, 118, 1943–1950. [Google Scholar] [CrossRef]
- Dhir, S.; Tarasenko, M.; Napoli, E.; Giulivi, C. Neurological, Psychiatric, and Biochemical Aspects of Thiamine Deficiency in Children and Adults. Front. psychiatry 2019, 10. [Google Scholar] [CrossRef]
- Dandachi, I.; Anani, H.; Hadjadj, L.; Brahimi, S.; Lagier, J.C.; Daoud, Z.; Rolain, J.M. Genome Analysis of Lachnoclostridium Phocaeense Isolated from a Patient after Kidney Transplantation in Marseille. New Microbes New Infect. 2021, 41, 100863. [Google Scholar] [CrossRef] [PubMed]
- McNabney, S.M.; Henagan, T.M. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.Z.; Zhang, S.; Pang, J.; Wang, S.; Zhang, G.; Zhao, J.; Wang, J. Gut Microbiota from Green Tea Polyphenol-Dosed Mice Improves Intestinal Epithelial Homeostasis and Ameliorates Experimental Colitis. Microbiome 2021, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, S.; Pang, J.; Wang, S.; Zhang, G.; Wang, J. Oral, but Not Rectal Delivery of Epigallocatechin-3-Gallate Alleviates Colitis by Regulating the Gut Microbiota, Oxidative Stress, Inflammation, and Barrier Integrity. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Amaral Montesino, C.; Abrego Sánchez, A.; Díaz Granados, M.A.; González Ponce, R.; Salinas Flores, A.; Rojas García, O.C.; Amaral Montesino, C.; Abrego Sánchez, A.; Díaz Granados, M.A.; González Ponce, R.; Salinas Flores, A.; Rojas García, O.C. Akkermansia Muciniphila, Una Ventana de Investigación Para La Regulación Del Metabolismo y Enfermedades Relacionadas. Nutr. Hosp. 2021, 38, 675–676. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yang, Y.; Xu, N.; Mu, H.; Zhang, H.; Duan, J. Improved Viability of Akkermansia Muciniphila by Encapsulation in Spray Dried Succinate-Grafted Alginate Doped with Epigallocatechin-3-Gallate. Int. J. Biol. Macromol. 2020, 159, 373–382. [Google Scholar] [CrossRef]
- Sheng, L.; Jena, P.K.; Hui-Xin, L.; Hu, Y.; Nagar, N.; Bronner, D.N.; Settles, M.L.; Bäumler, A.J.; Wan, Y.J.Y. Obesity Treatment by Epigallocatechin-3-Gallate−regulated Bile Acid Signaling and Its Enriched Akkermansia Muciniphila. FASEB J. 2018, 32, 6371–6384. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, K.; Jing, N.; Zhao, Y.; Yang, X. EGCG Regulates Fatty Acid Metabolism of High-Fat Diet-Fed Mice in Association with Enrichment of Gut Akkermansia Muciniphila. J. Funct. Foods 2020, 75, 104261. [Google Scholar] [CrossRef]
- Manrique, D.; Elie, V. Ácidos Grasos de Cadena Corta (Ácido Butírico) y Patologías Intestinales. Nutr. Hosp. 2017, 34, 58–61. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Alshammari, M.K.; AlKhulaifi, M.M.; Al Farraj, D.A.; Somily, A.M.; Albarrag, A.M. Incidence of Clostridium Perfringens and Its Toxin Genes in the Gut of Children with Autism Spectrum Disorder. Anaerobe 2020, 61, 102114. [Google Scholar] [CrossRef]
- Liu, J.; Gao, Z.; Liu, C.; Liu, T.; Gao, J.; Cai, Y.; Fan, X. Alteration of Gut Microbiota: New Strategy for Treating Autism Spectrum Disorder. Front. Cell Dev. Biol. 2022, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Sandler, R.H.; Finegold, S.M.; Bolte, E.R.; Buchanan, C.P.; Maxwell, A.P.; Väisänen, M.L.; Nelson, M.N.; Wexler, H.M. Short-Term Benefit From Oral Vancomycin Treatment of Regressive-Onset Autism. J. Child Neurol. 2000, 15, 429–435. [Google Scholar] [CrossRef]
- Enterobacterias - EcuRed. https://www.ecured.cu/Enterobacterias (accessed 2022-10-11).
- Mohammad, F.K.; Palukuri, M.V.; Shivakumar, S.; Rengaswamy, R.; Sahoo, S. A Computational Framework for Studying Gut-Brain Axis in Autism Spectrum Disorder. Front. Physiol. 2022, 13. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. Mitochondrial Dysfunction in Autism Spectrum Disorders: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2012, 17, 290–314. [Google Scholar] [CrossRef] [PubMed]
- Matelski, L.; Van de Water, J. Risk Factors in Autism: Thinking Outside the Brain. J. Autoimmun. 2016, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McAllister, A.K. Immune Contributions to Cause and Effect in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 380–382. [Google Scholar] [CrossRef]
- Meltzer, A.; Van De Water, J. The Role of the Immune System in Autism Spectrum Disorder. Neuropsychopharmacology 2017, 42, 284–298. [Google Scholar] [CrossRef]
- Leviton, A.; Joseph, R.M.; Allred, E.N.; Fichorova, R.N.; O’Shea, T.M.; Kuban, K.K.C.; Dammann, O. The Risk of Neurodevelopmental Disorders at Age 10 Years Associated with Blood Concentrations of Interleukins 4 and 10 during the First Postnatal Month of Children Born Extremely Preterm. Cytokine 2018, 110, 181–188. [Google Scholar] [CrossRef]
- Shen, Y.; Li, Y.; Shi, L.; Liu, M.; Wu, R.; Xia, K.; Zhang, F.; Ou, J.; Zhao, J. Autism Spectrum Disorder and Severe Social Impairment Associated with Elevated Plasma Interleukin-8. Pediatr. Res. 2021, 89, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H.; Geng, L.; Davidow, A.L. Cytokine Profiles by Peripheral Blood Monocytes Are Associated with Changes in Behavioral Symptoms Following Immune Insults in a Subset of ASD Subjects: An Inflammatory Subtype? J. Neuroinflammation 2014, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H.; Geng, L.; Streck, D.L.; Dermody, J.J.; Toruner, G.A. MicroRNA Expression Changes in Association with Changes in Interleukin-1ß/Interleukin10 Ratios Produced by Monocytes in Autism Spectrum Disorders: Their Association with Neuropsychiatric Symptoms and Comorbid Conditions (Observational Study). J. Neuroinflammation 2017, 14, 1–14. [Google Scholar] [CrossRef]
- Masi, A.; Quintana, D.S.; Glozier, N.; Lloyd, A.R.; Hickie, I.B.; Guastella, A.J. Cytokine Aberrations in Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2015, 20, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Ricci, S.; Businaro, R.; Ippoliti, F.; Lo Vasco, V.R.; Massoni, F.; Onofri, E.; Troili, G.M.; Pontecorvi, V.; Morelli, M.; Rapp Ricciardi, M.; Archer, T. Altered Cytokine and BDNF Levels in Autism Spectrum Disorder. Neurotox. Res. 2013, 24, 491–501. [Google Scholar] [CrossRef]
- Binder, D.K.; Scharfman, H.E. Brain-Derived Neurotrophic Factor. Growth Factors 2004, 22, 123–131. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677. [Google Scholar] [CrossRef]
- Bekinschtein, P.; Cammarota, M.; Katche, C.; Slipczuk, L.; Rossato, J.I.; Goldin, A.; Izquierdo, I.; Medina, J.H. BDNF Is Essential to Promote Persistence of Long-Term Memory Storage. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 2711–2716. [Google Scholar] [CrossRef]
- Yamada, K.; Nabeshima, T. Brain-Derived Neurotrophic Factor/TrkB Signaling in Memory Processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef]
- Hu, T.; Dong, Y.; He, C.; Zhao, M.; He, Q. The Gut Microbiota and Oxidative Stress in Autism Spectrum Disorders (ASD). Oxid. Med. Cell. Longev. 2020, 2020. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.; Zheng, B.; Cheung, P.C.K. Short-Chain Fatty Acids in Control of Energy Metabolism. Crit. Rev. Food Sci. Nutr. 2018, 58, 1243–1249. [Google Scholar] [CrossRef]
- Codina-Solà, M.; Rodríguez-Santiago, B.; Homs, A.; Santoyo, J.; Rigau, M.; Aznar-Laín, G.; Del Campo, M.; Gener, B.; Gabau, E.; Botella, M.P.; Gutiérrez-Arumí, A.; Antiñolo, G.; Pérez-Jurado, L.A.; Cuscó, I. Integrated Analysis of Whole-Exome Sequencing and Transcriptome Profiling in Males with Autism Spectrum Disorders. Mol. Autism 2015, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; Osredkar, J.; Urbina, M.A.; Fabjan, T.; El-Houfey, A.A.; Kałużna-Czaplińska, J.; Gątarek, P.; Chirumbolo, S. Oxidative Stress in Autism Spectrum Disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Ahmad, S.F.; Attia, S.M.; AL-Ayadhi, L.Y.; Al-Harbi, N.O.; Bakheet, S.A. Dysregulated Enzymatic Antioxidant Network in Peripheral Neutrophils and Monocytes in Children with Autism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 88, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Messina, A.; Monda, V.; Sessa, F.; Valenzano, A.; Salerno, M.; Bitetti, I.; Precenzano, F.; Marotta, R.; Lavano, F.; Lavano, S.M.; Salerno, M.; Maltese, A.; Roccella, M.; Parisi, L.; Ferrentino, R.I.; Tripi, G.; Gallai, B.; Cibelli, G.; Monda, M.; Messina, G.; Carotenuto, M. Sympathetic, Metabolic Adaptations, and Oxidative Stress in Autism Spectrum Disorders: How Far from Physiology? Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O. Role of Peroxisomes in ROS/RNS-Metabolism: Implications for Human Disease. Biochim. Biophys. Acta 2012, 1822, 1363–1373. [Google Scholar] [CrossRef]
- Cipolla, C.M.; Lodhi, I.J. Peroxisomal Dysfunction in Age-Related Diseases. Trends Endocrinol. Metab. 2017, 28, 297–308. [Google Scholar] [CrossRef]
- James, S.J.; Melnyk, S.; Jernigan, S.; Cleves, M.A.; Halsted, C.H.; Wong, D.H.; Cutler, P.; Bock, K.; Boris, M.; Bradstreet, J.J.; Baker, S.M.; Gaylor, D.W. Metabolic Endophenotype and Related Genotypes Are Associated with Oxidative Stress in Children with Autism. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141B, 947–956. [Google Scholar] [CrossRef]
- Rose, S.; Bennuri, S.C.; Wynne, R.; Melnyk, S.; James, S.J.; Frye, R.E. Mitochondrial and Redox Abnormalities in Autism Lymphoblastoid Cells: A Sibling Control Study. FASEB J. 2017, 31, 904–909. [Google Scholar] [CrossRef]
- Ray, B.; Long, J.M.; Sokol, D.K.; Lahiri, D.K. Increased Secreted Amyloid Precursor Protein-α (SAPPα) in Severe Autism: Proposal of a Specific, Anabolic Pathway and Putative Biomarker. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Sokol, D.K.; Maloney, B.; Long, J.M.; Ray, B.; Lahiri, D.K. Autism, Alzheimer Disease, and Fragile X: APP, FMRP, and MGluR5 Are Molecular Links. Neurology 2011, 76, 1344–1352. [Google Scholar] [CrossRef]
- Lahiri, D.K.; Sokol, D.K.; Erickson, C.; Ray, B.; Ho, C.Y.; Maloney, B. Autism as Early Neurodevelopmental Disorder: Evidence for an SAPPα-Mediated Anabolic Pathway. Front. Cell. Neurosci. 2013, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Valenti, D.; de Bari, L.; de Rasmo, D.; Signorile, A.; Henrion-Caude, A.; Contestabile, A.; Vacca, R.A. The Polyphenols Resveratrol and Epigallocatechin-3-Gallate Restore the Severe Impairment of Mitochondria in Hippocampal Progenitor Cells from a Down Syndrome Mouse Model. Biochim. Biophys. Acta 2016, 1862, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Paixão, J.; Nunes, C.; Dinis, T.C.P.; Almeida, L.M. Cyanidin-3-Glucoside Suppresses Cytokine-Induced Inflammatory Response in Human Intestinal Cells: Comparison with 5-Aminosalicylic Acid. PLoS One 2013, 8, 73001. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Rufino, A.T.; Mendes, A.F.; Almeida, L.M.; Dinis, T.C.P. Resveratrol Modulates Cytokine-Induced JAK/STAT Activation More Efficiently than 5-Aminosalicylic Acid: An In Vitro Approach. PLoS One 2014, 9, e109048. [Google Scholar] [CrossRef]
- Sunand, K.; Mohan, G.K.; Bakshi, V. Synergetic Potential of Combination Probiotic Complex with Phytopharmaceuticals in Valproic Acid Induced Autism: Prenatal Model. Int. J. Appl. Pharm. Sci. Res. 2021, 6, 33–43. [Google Scholar] [CrossRef]
- Avramovich-Tirosh, Y.; Reznichenko, L.; Amit, T.; Zheng, H.; Fridkin, M.; Weinreb, O.; Mandel, S.; Youdim, M. Neurorescue Activity, APP Regulation and Amyloid-Beta Peptide Reduction by Novel Multi-Functional Brain Permeable Iron- Chelating- Antioxidants, M-30 and Green Tea Polyphenol, EGCG. Curr. Alzheimer Res. 2007, 4, 403–411. [Google Scholar] [CrossRef]
- Pogačnik, L.; Pirc, K.; Palmela, I.; Skrt, M.; Kwang, K.S.; Brites, D.; Brito, M.A.; Ulrih, N.P.; Silva, R.F.M. Potential for Brain Accessibility and Analysis of Stability of Selected Flavonoids in Relation to Neuroprotection in Vitro. Brain Res. 2016, 1651, 17–26. [Google Scholar] [CrossRef]
- Porath, D.; Riegger, C.; Drewe, J.; Schwager, J. Epigallocatechin-3-Gallate Impairs Chemokine Production in Human Colon Epithelial Cell Lines. J. Pharmacol. Exp. Ther. 2005, 315, 1172–1180. [Google Scholar] [CrossRef]
- Frye, R.E.; Rossignol, D.A. Mitochondrial Dysfunction Can Connect the Diverse Medical Symptoms Associated with Autism Spectrum Disorders. Pediatr. Res. 2011, 69 Pt 2. [Google Scholar] [CrossRef]
- Ming, X.; Stein, T.P.; Barnes, V.; Rhodes, N.; Guo, L. Metabolic Perturbance in Autism Spectrum Disorders: A Metabolomics Study. J. Proteome Res. 2012, 11, 5856–5862. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.E.; Rose, S.; Slattery, J.; MacFabe, D.F. Gastrointestinal Dysfunction in Autism Spectrum Disorder: The Role of the Mitochondria and the Enteric Microbiome. Microb. Ecol. Health Dis. 2015, 26. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-Inflammatory Effects of Dietary Phenolic Compounds in an in Vitro Model of Inflamed Human Intestinal Epithelium. Chem. Biol. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, S.; Naik, B.; Ramachandra, N.B. Mucosa-Associated Specific Bacterial Species Disrupt the Intestinal Epithelial Barrier in the Autism Phenome. Brain, Behav. Immun. - Heal. 2021, 15, 100269. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S RRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Liu, S.; Li, E.; Sun, Z.; Fu, D.; Duan, G.; Jiang, M.; Yu, Y.; Mei, L.; Yang, P.; Tang, Y.; Zheng, P. Altered Gut Microbiota and Short Chain Fatty Acids in Chinese Children with Autism Spectrum Disorder. Sci. Reports 2019 91 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, W.; Zhang, J.; He, Y.; Wang, J. Analysis of Gut Microbiota Profiles and Microbe-Disease Associations in Children with Autism Spectrum Disorders in China. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Brown, C.T.; Davis-Richardson, A.G.; Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; Hyöty, H.; Veijola, R.; Simell, T.; Simell, O.; Neu, J.; Wasserfall, C.H.; Schatz, D.; Atkinson, M.A.; Triplett, E.W. Gut Microbiome Metagenomics Analysis Suggests a Functional Model for the Development of Autoimmunity for Type 1 Diabetes. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Rose, S.; Bennuri, S.C.; Murray, K.F.; Buie, T.; Winter, H.; Frye, R.E. Mitochondrial Dysfunction in the Gastrointestinal Mucosa of Children with Autism: A Blinded Case-Control Study. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Rose, S.; Bennuri, S.C.; Davis, J.E.; Wynne, R.; Slattery, J.C.; Tippett, M.; Delhey, L.; Melnyk, S.; Kahler, S.G.; MacFabe, D.F.; Frye, R.E. Butyrate Enhances Mitochondrial Function during Oxidative Stress in Cell Lines from Boys with Autism. Transl. Psychiatry 2018, 8. [Google Scholar] [CrossRef]
- Cristiano, C.; Hoxha, E.; Lippiello, P.; Balbo, I.; Russo, R.; Tempia, F.; Miniaci, M.C. Maternal Treatment with Sodium Butyrate Reduces the Development of Autism-like Traits in Mice Offspring. Biomed. Pharmacother. 2022, 156, 113870. [Google Scholar] [CrossRef]
- Kang, D.W.; Ilhan, Z.E.; Isern, N.G.; Hoyt, D.W.; Howsmon, D.P.; Shaffer, M.; Lozupone, C.A.; Hahn, J.; Adams, J.B.; Krajmalnik-Brown, R. Differences in Fecal Microbial Metabolites and Microbiota of Children with Autism Spectrum Disorders. Anaerobe 2018, 49, 121–131. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Lan, A.; Beaumont, M.; Audebert, M.; Wong, X.; Yamada, K.; Yin, Y.; Tomé, D.; Carrasco-Pozo, C.; Gotteland, M.; Kong, X.; Blachier, F. The Deleterious Metabolic and Genotoxic Effects of the Bacterial Metabolite P-Cresol on Colonic Epithelial Cells. Free Radic. Biol. Med. 2015, 85, 219–227. [Google Scholar] [CrossRef]
- Unno, T.; Ichitani, M. Epigallocatechin-3-Gallate Decreases Plasma and Urinary Levels of p-Cresol by Modulating Gut Microbiota in Mice. ACS Omega 2022, 7, 40034–40041. [Google Scholar] [CrossRef]
- Unno, K.; Pervin, M.; Nakagawa, A.; Iguchi, K.; Hara, A.; Takagaki, A.; Nanjo, F.; Minami, A.; Nakamura, Y. Blood–Brain Barrier Permeability of Green Tea Catechin Metabolites and Their Neuritogenic Activity in Human Neuroblastoma SH-SY5Y Cells. Mol. Nutr. Food Res. 2017, 61, 1700294. [Google Scholar] [CrossRef] [PubMed]
- Kohri, T.; Matsumoto, N.; Yamakawa, M.; Suzuki, M.; Nanjo, F.; Hara, Y.; Oku, N. Metabolic Fate of (−)-[4-3H]Epigallocatechin Gallate in Rats after Oral Administration. Am. Chem. Soc. 2001, 49, 4102–4112. [Google Scholar] [CrossRef]
- Pervin, M.; Unno, K.; Nakagawa, A.; Takahashi, Y.; Iguchi, K.; Yamamoto, H.; Hoshino, M.; Hara, A.; Takagaki, A.; Nanjo, F.; Minami, A.; Imai, S.; Nakamura, Y. Blood Brain Barrier Permeability of (-)-Epigallocatechin Gallate, Its Proliferation-Enhancing Activity of Human Neuroblastoma SH-SY5Y Cells, and Its Preventive Effect on Age-Related Cognitive Dysfunction in Mice. Biochem. Biophys. reports 2017, 9, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Pervin, M.; Unno, K.; Takagaki, A.; Isemura, M.; Nakamura, Y. Function of Green Tea Catechins in the Brain: Epigallocatechin Gallate and Its Metabolites. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Lambert, J.D.; Rice, J.E.; Hong, J.; Hou, Z.; Yang, C.S. Synthesis and Biological Activity of the Tea Catechin Metabolites, M4 and M6 and Their Methoxy-Derivatives. Bioorg. Med. Chem. Lett. 2005, 15, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Won, Y.S.; Yang, X.; Kumazoe, M.; Yamashita, S.; Hara, A.; Takagaki, A.; Goto, K.; Nanjo, F.; Tachibana, H. Green Tea Catechin Metabolites Exert Immunoregulatory Effects on CD4(+) T Cell and Natural Killer Cell Activities. J. Agric. Food Chem. 2016, 64, 3591–3597. [Google Scholar] [CrossRef]
- Chen, L.; Lee, M.-J.; Li, H.; Yang, C.S. Absorption, Distribution, and Elimination of Tea Polyphenols in Rats. Drug Metab. Dispos. 1997, 25. [Google Scholar]
- Kim, S.; Lee, M.J.; Hong, J.; Li, C.; Smith, T.J.; Yang, G.Y.; Seril, D.N.; Yang, C.S. Plasma and Tissue Levels of Tea Catechins in Rats and Mice during Chronic Consumption of Green Tea Polyphenols. Nutr. Cancer 2000, 37, 41–48. [Google Scholar] [CrossRef]
- Cano, A.; Ettcheto, M.; Chang, J.H.; Barroso, E.; Espina, M.; Kühne, B.A.; Barenys, M.; Auladell, C.; Folch, J.; Souto, E.B.; Camins, A.; Turowski, P.; García, M.L. Dual-Drug Loaded Nanoparticles of Epigallocatechin-3-Gallate (EGCG)/Ascorbic Acid Enhance Therapeutic Efficacy of EGCG in a APPswe/PS1dE9 Alzheimer’s Disease Mice Model. J. Control. Release 2019, 301, 62–75. [Google Scholar] [CrossRef]
- Abbasalipour, H.; Hajizadeh Moghaddam, A.; Ranjbar, M. Sumac and Gallic Acid-Loaded Nanophytosomes Ameliorate Hippocampal Oxidative Stress via Regulation of Nrf2/Keap1 Pathway in Autistic Rats. J. Biochem. Mol. Toxicol. 2022, 36. [Google Scholar] [CrossRef] [PubMed]
- Mindell, J.A.; Meltzer, L.J.; Carskadon, M.A.; Chervin, R.D. Developmental Aspects of Sleep Hygiene: Findings from the 2004 National Sleep Foundation Sleep in America Poll. Sleep Med. 2009, 10, 771–779. [Google Scholar] [CrossRef]
- Samanta, P.; Panigrahi, A.; Senapati, L.K.; Mishra, D.P.; Ravan, J.R.; Mishra, J. Maladaptive Behavior and Associated Factors among Young Children with Autism. Indian J. Pediatr. 2022, 89, 1134–1136. [Google Scholar] [CrossRef]
- Evans, M.O.; Starley, B.; Galagan, J.C.; Yabes, J.M.; Evans, S.; Salama, J.J. Tea and Recurrent Clostridium Difficile Infection. Gastroenterol. Res. Pract. 2016, 2016. [Google Scholar] [CrossRef]
- Vossoughinia, H.; Salari, M.; Amirmajdi, E.M.; Saadatnia, H.; Abedini, S.; Shariati, A.; Shariati, M.; Khorashad, A.K. An Epidemiological Study of Gastroesophageal Reflux Disease and Related Risk Factors in Urban Population of Mashhad, Iran. Iran. Red Crescent Med. J. 2014, 16. [Google Scholar] [CrossRef]
- Buie, T.; Campbell, D.B.; Fuchs, G.J.; Furuta, G.T.; Levy, J.; Van De Water, J.; Whitaker, A.H.; Atkins, D.; Bauman, M.L.; Beaudet, A.L.; Carr, E.G.; Gershon, M.D.; Hyman, S.L.; Jirapinyo, P.; Jyonouchi, H.; Kooros, K.; Kushak, R.; Levitt, P.; Levy, S.E.; Lewis, J.D.; Murray, K.F.; Natowicz, M.R.; Sabra, A.; Wershil, B.K.; Weston, S.C.; Zeltzer, L.; Winter, H. Evaluation, Diagnosis, and Treatment of Gastrointestinal Disorders in Individuals with ASDs: A Consensus Report. Pediatrics 2010, 125 (SUPPL. 1). [Google Scholar] [CrossRef] [PubMed]
- Kucera, O.; Mezera, V.; Moravcova, A.; Endlicher, R.; Lotkova, H.; Drahota, Z.; Cervinkova, Z. In Vitro Toxicity of Epigallocatechin Gallate in Rat Liver Mitochondria and Hepatocytes. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; Lin, A.; Sultan, A.M.; O’Brien, P.J. Cellular and in Vivo Hepatotoxicity Caused by Green Tea Phenolic Acids and Catechins. Free Radic. Biol. Med. 2006, 40, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Mezera, V.; Kucera, O.; Moravcova, A.; Peterova, E.; Cervinkova, Z. The Effect of Epigallocatechin Gallate on Hepatocytes Isolated from Normal and Partially Hepatectomized Rats. Can. J. Physiol. Pharmacol. 2014, 92, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.-H. S.; Cai, Y.; Hakim, I.A.; Crowell, J.A.; Shahi, F.; Brooks, C.A.; Dorr, R.T.; Hara, Y.; Alberts, D.S. Pharmacokinetics and Safety of Green Tea Polyphenols after Multiple-Dose Administration of Epigallocatechin Gallate and Polyphenon E in Healthy Individuals1. Clin. Cancer Res. 2003, 9, 3312–3319. [Google Scholar]
- Abe, S.K.; Inoue, M. Green Tea and Cancer and Cardiometabolic Diseases: A Review of the Current Epidemiological Evidence. Eur. J. Clin. Nutr. 2021, 75, 865–876. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
EGCG impact in patients with ASD. Alterations in the intestinal microbiota in ASD and its implications on the left; EGCG impact on both the intestinal microbiota and variables directly related to the microbiota such as metabolites, inflammation and oxidative stress on the right.
Figure 1.
EGCG impact in patients with ASD. Alterations in the intestinal microbiota in ASD and its implications on the left; EGCG impact on both the intestinal microbiota and variables directly related to the microbiota such as metabolites, inflammation and oxidative stress on the right.
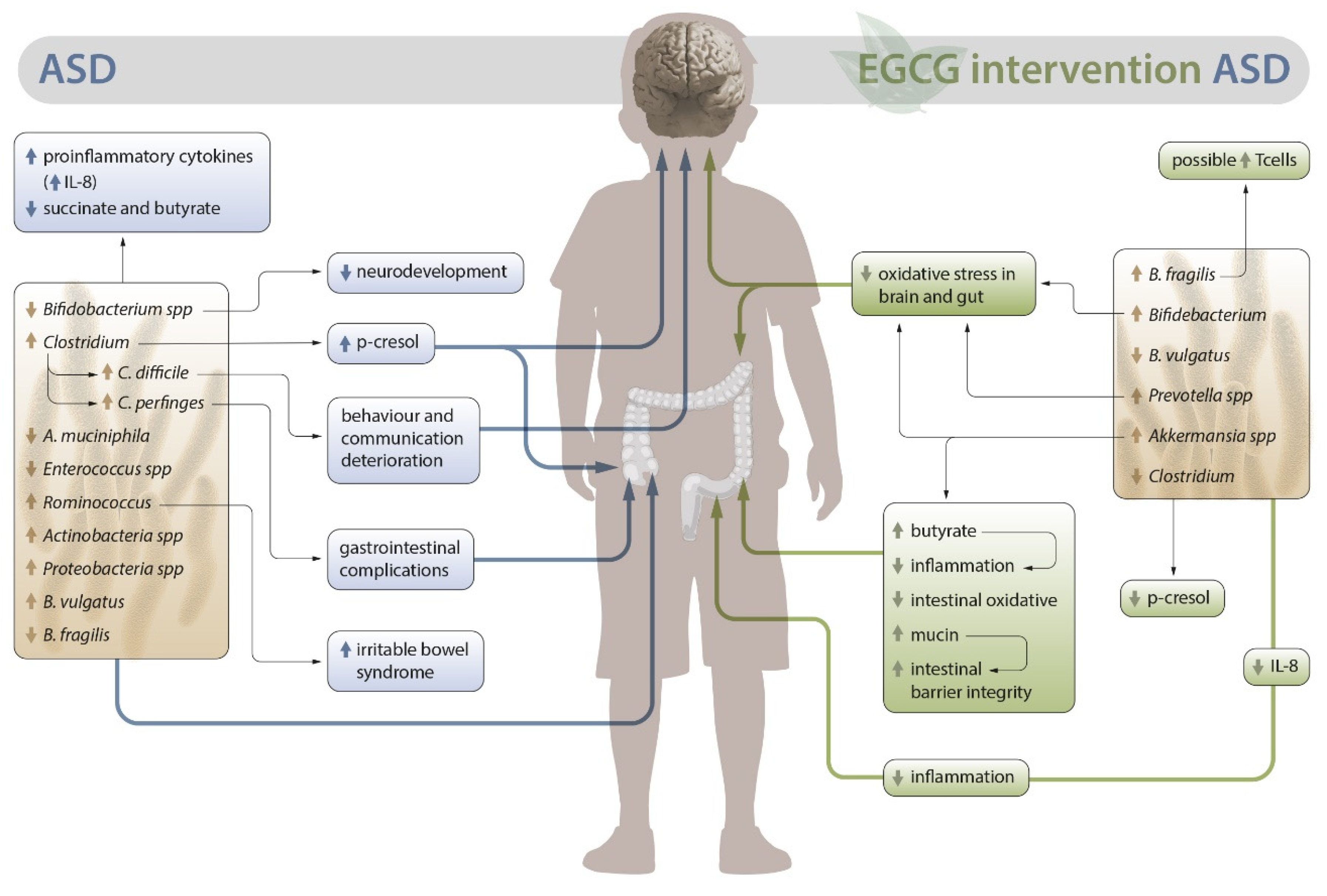
Figure 2.
Mechanism of EGCG metabolite generation in the intestinal microbiota. Reciprocal interactions between EGCG and human gut microbiota in vitro [65].
Figure 2.
Mechanism of EGCG metabolite generation in the intestinal microbiota. Reciprocal interactions between EGCG and human gut microbiota in vitro [65].
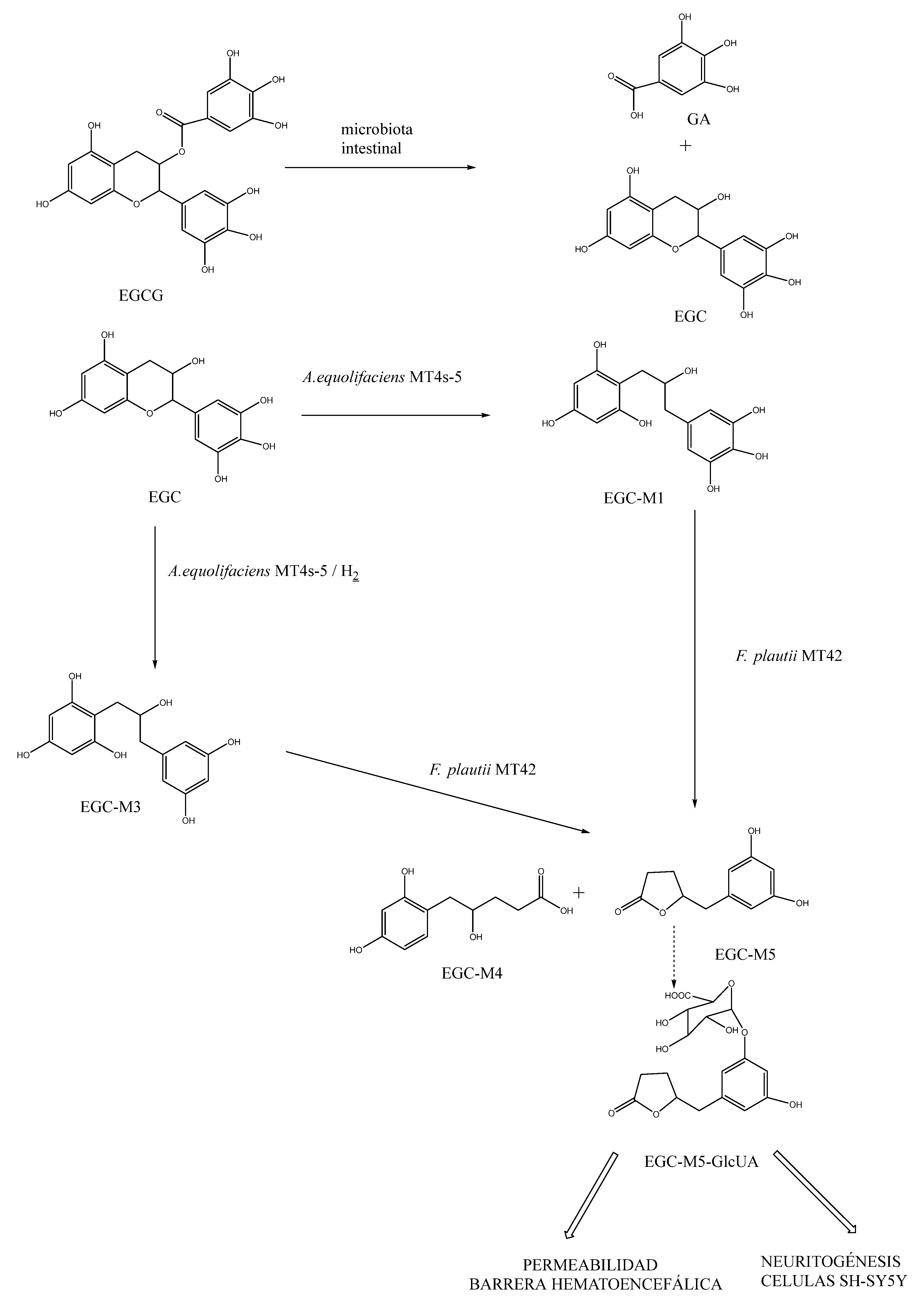

Table 1.
Main results related to EGCG effects on microbiota populations.
| Study | Study Type | Population/sample size | Intervention | Results | Ref. |
|---|---|---|---|---|---|
| Ankolekar C et al. 2011 | Analytical Longitudinal Experimental |
Helicobacter pylori Lactic acid bacteria strains such as B. longum, L. acidophilus, and L. plantarum | They were exposed to tea preparations with rest times of 2 and 5 minutes | The 4 types of tea inhibited H. pylori | 47 |
| Shin JS et al. 2007 | Analytical Cross-sectional Experimental |
The antibacterial activities of the main phenolic components of Camellia sinensis | Minimum Inhibitory Concentration Test (MIC) | (-)-epicatechin is more toxic against S. aureus and E. coli | 48 |
| Nakayama M et al. 2012 | Analytical Longitudinal Experimental |
E. coli O157:H7 producing Stx 1 and Stx 2 S. aureus NBRC 13276 | Green tea extract concentration was 0.0625 mg/ml for S. aureus and 1.0 mg/ml for E. coli O157:H7. | Green tea extract (GTE) inhibits the absorption and secretion of substrates and inhibits enzyme activity in both species | 49 |
| Bancirova M et al. 2010 | Analytical Cross-sectional Experimental |
Standard strains of Enterococcus faecalis 4224, S. aureus 3953 and 4223, P. aeruginosa 3955 and E. coli 3954 and 3988 | Tea infusion (2g/100ml) | E. faecalis was the most resistant bacterial strain and P. aeruginosa was the least resistant bacterial strain. But the predominant antimicrobial activity of non-fermented tea infusions was not confirmed. | 52 |
| Trovò L et al. 2020 | Analytical Longitudinal Experimental |
Cdkl5-KO mice N = 5–6 |
Primary hippocampal neurons were treated daily with EGCG [1 μM, 0.5 μM, 1 μM, and 3 μM] or harmine | Treatment with EGCG efficiently restores defects in dendritic and synaptic development of Cdkl5-KO hippocampal neurons. | 57 |
| Kumaravel P et al. 2017 | Analytical Longitudinal Experimental |
Wistar rats N = 6 per group |
Rats were treated with EGCG in doses of 1, 2 and 5 mg/kg body weight via oral administration. | EGCG ameliorates and reverses autistic attributes possibly due to its neuroprotective activity. | 58 |
| Liu Z et al. 2020 | Analytical Cross-sectional Experimental |
Fecal materia from four volunteers | Catechin concentration of 0.1 mmol/L | EGCG increase the beneficial bacteria Bacteroides, Christensenellaceae, and Bifidobacterium. And inhibited the pathogenic bacteria Fusobacterium varium, Bilophila, and Enterobacteriaceae. | 65 |
| Ushiroda C et al. 2019 |
Analytical Longitudinal Experimental |
Male C57BL/ 6N mice | EGCG at a concentration of 0,32% for 8 weeks | EGCG increase abundance of Adlercreutzia, Akkermansia, Allobaculum and a significantly lower abundance of Desulfovibrionaceae | 67 |
| Qu Y et al. 2022 | Analytical Longitudinal Experimental |
Female C57BL/6 mice (8–13 weeks old) | EGCG [45 mg/kg] were administered for 8 weeks. The mice were then subjected to behavioral and gut microbiota analysis | EGCG effectively increased Prevotella and decreased Bifidobacteriales but had no effect on Alloprevotella or Lactobacillaceae | 68 |
| Unno T et al. 2014 | Analytical Longitudinal Experimental |
Four-week-old male Wistar rats N= 7 per group |
Rats were fed an assigned diet of either a control diet, a 0,3% (w/w) EGCG diet, or a 0,6% (w/w) EGCG diet for 4 weeks | Dietary EGCG affects the growth of certain species of gut microbiota which could be responsible for regulating energy metabolism in the body | 72 |
| Wu Z et al. 2021 | Analytical Longitudinal Experimental |
Seven- to eight-week-old specifc pathogen-free (SPF) female C57BL/6J mice N = 8 per group |
Modulated the gut microbial community of mice by the EGCG [50 mg/kg] pre-supplementation | Oral EGCG suppressed DSS-induced oxidative stress in intestinal mucosa, improved the barrier function and regulated the composition and SCFAs production of gut microbiota | 88 |
| Wu Z et al. 2020 | Analytical Longitudinal Experimental |
Seven- to eight-week-old specific pathogen-free (SPF) female C57L/6J mice | Tested the effect of oral or rectal administration of EGCG [50 mg/kg] | EGCG enriched Akkermansia, Faecalibaculum, and Bifidobacterium and enhanced acetate, propionate and butyrate production. | 89 |
| Sheng L et al. 2018 | Analytical Longitudinal Experimental |
Specific pathogen-free male C57BL/6 wild-type (WT) mice N = 16 |
Mice received vehicle (PBS) or EGCG daily [100 mg/d per gram body weight, orally] for 2 mo. | Increased Verrucomicrobiaceae and Akkermansia muciniphila. | 92 |
| Liu X et al. 2020 | Analytical Longitudinal Experimental |
C57BL/6J male mice N = 8 |
C57BL/6J mice fed with HFD were administrated with 210 mg/kg EGCG for 12 weeks. | Enriched the Verrucomicrobia and decreased the Firmicutes and Saccharibacteria. | 93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



