
Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
Background: The intrauterine and early extrauterine development represents a “window of opportunity” in the immunological development. The underlying mechanisms are still poorly understood. The aim of this study was to provide reference values B cell subpopulations in cord blood of term newborns, juveniles and in adults to find the spectrum of their physiological age-related variation.
Methods: In this study, we used flow cytometry to evaluate human B lymphocytes and subpopulations in cord blood (n = 10), in peripheral blood from healthy juveniles aged 1 to 17 years (n=20) and from donors aged 24 to 62 years (n = 10).
Results: Our findings showed increasing frequencies of IgM memory B cells, class-switched memory B cells, marginal zone B cells and plasmablasts, from cord blood to peripheral blood of juveniles and adults. In con-trast, the percentage of naïve B cells was higher in newborns than in juveniles and adults. The frequencies of were similar in cord blood and peripheral blood of adults. Interestingly, transitional B cells frequencies were similar in cord blood and adults but significantly lower in juveniles.
Conclusions: The frequencies of circulating B cell subpopulation are subject to considerable changes during on-togeny, reflecting overlying effects of maturation and of the acquisition of an adaptive immune memory.
Keywords:
B cells
; B1 cells
; human
; leukocytes
; cord blood
; development
; ontogeny
1. Introduction
B cells play a crucial role in the adaptive immune response, especially in the defense against extracellular infectious pathogens [1]. The development of the human immune system starts in the fetal stage and is not yet complete at birth [2]. Directly after birth, newborns possess a sufficient number of immune cells, which have to be activated by antigen exposure. During puberty, this functional learning process is completed in many aspects, meaning that the individual has the immunological abilities of an adult. However, the immune system of an adult is also subject to a continuous learning process throughout senescense [3].
During the first weeks after birth, B lymphocytes proliferate excessively, probably due to the massive stimulation with previously unknown environmental antigens. Such stimulation triggers the proliferation and differentiation of antigen-specific cells [4,5].
The maturation of B lymphocytes is a complex, highly regulated, multi-stage process that starts in the bone marrow.
Transitional B cells migrate into secondary lymphoid structures, e.g. follicles and germinal centres of spleen and lymph nodes where they undergo further maturational steps. After class switch and affinity-maturation the memory B cells circulate continuously between blood and lymphatic organs, whereas the plasma cells settle in the bone marrow and secrete high-affinity antibodies [6,7]. B1 cells constitutively and spontaneously secrete "natural" antibodies which convey immunity to an array of antigens even in the neonate [8]. While B1 cells are well characterized in murine experimental models [8,9], their analysis in humans is difficult. The quantity and even the existence of B1 cells are under debate in human, and various definitions had been suggested. While in mice B1 cells are defined as CD5+ and/or CD11b+ cells, a substantial number of B1 like cells do not express CD5 in humans [10,11,12]. Griffin et al. [13] identified a CD20+CD27+CD43+ B cell subpopulation in human adult peripheral blood and cord blood with functional characteristics that they describe as typical B1 cell attributes: spontaneous IgM secretion, constitutive BCR signaling and the ability to control allogeneic T cell proliferation. Cord blood contains a greater number of B cells than adult venous blood. At birth, CD27+IgM+IgD+ memory B cells represent the largest B cell subpopulation, but the frequency of this subpopulation decreases during childhood and then stabilizes in young adults. In contrast, CD27+ class-switched B cells gradually enrich in the peripheral blood with age [14,15]. Nevertheless, the data available to date are still patchy. Hence, this study aims to gain a deeper insight into the composition of B cell subpopulations as a function of age using up-to-date cell definitions.
2. Results
2.1. Phenotypic characterization of memory B cell subsets in healthy newborns, juveniles and adults
To compare the age-related influences on the distribution of B lymphocyte subpopulations in neonates, juveniles and adults, we analyzed cord blood samples from 10 term neonates, and venous blood samples from 20 juveniles and 10 adults.
The B cell subpopulations were quantified by flow cytometry. To study the diversity in the human B cell compartment, we defined eight B cell subpopulations (Supplemental Table S2). The surface markers are listed in Supplemental Table S1 and the gating strategies are depicted in Figure 1. Immature B cells were defined as CD19+CD27-IgM-IgD- (Figure 1C). The frequency of immature B cells (Figure 3A) did not differ significantly between neonates (1.4 ± 0.3 %), juveniles (1.7± 0.3 %) and adults (2.3 ± 0.3 %) (neonates vs juveniles p > 0.99, neonates vs adults p = 0.15, juveniles vs adults p = 0.27). Our flow cytometry analysis showed significantly higher naïve B cell (CD19+CD27-IgD+IgM+) frequencies in cord blood of neonates (93.2 ± 1.2 %) than in peripheral blood of juveniles (80.8 ± 1.8 %; p = 0.002) and adults (69.1 ± 3.4 %; p < 0.001) (Figure 3B). The results show that the frequency of the naïve B cell decreases with increasing age.
The transitional B cells are reliably identified as CD19+CD27+CD38+IgM+ lymphocytes (Figure 1D) [16,17]. The proportions of these transitional B cells were nearly equal in adults 30.9 ± 3.7 % and neonates (27.9 ± 5.7 %; p = 0.3527) (Figure 3C). In juveniles the frequencies of transitional B cells were significantly lower (9.0 ± 1.0 %; neonates vs juveniles p < 0.001; juveniles vs adults p < 0.001). Memory cells can be subdivided in class-switched memory B cells, defined as CD19+CD27+IgM-IgD- B cells, IgM memory B cells, described as CD19+CD27+IgM+IgD- B cells and CD19+CD27+IgM+IgD+ marginal zone B cells (Figure 1E). We found significantly higher class-switched memory B cells, IgM memory B cells and marginal zone B cells in peripheral blood of juveniles (6.4 ± 1.0 %; 1.4 ± 0.2 %; 7.1 ± 0.8 %) and adults (14.1 ± 2.0 %; 1.7 ± 0.3 %; 10.7 ± 1.4 %) than in cord blood of neonates (2.5 ± 0.4 %; 0.2 ± 0.1 %; 0.5 ± 0.1 %) (each p = 0.002 or lower) (Figure 3D-F). These results show that the frequency of the class-switched memory B cells, IgM memory B cells and marginal zone B cells increases with age. Further population that develops from activated B cells are plasmablasts (CD19+CD27+IgM-CD38++) (Figure 1F). The proportions of CD19+CD27+IgM-CD38++B cells are significantly larger in peripheral blood of adults (1.4 ± 0.3 %) compared to cord blood of neonates (0.5 ± 0.2 %; p = 0.02) (Figure 3G).
2.2. B1 cells in cord blood and peripheral blood of adults
B1 cells, also known as pre-naïve B cells, represent an intermediate stage between the transitional and naïve B cells. In humans, B1 cells are CD20+, CD27+, CD43+ and CD69- [13] and are given as a percentage of the number of CD20+ cells (Figure 2). The frequencies of CD20+CD27+CD43+CD69- B1 cells did not significantly differ between both groups analyzed (neonates 1.8 ± 0.3 %; adults 2.7 ± 0.4 %, p = 0.12) (Figure 3H).
Figure 2.
Gating strategy and B1 cell frequency. (A) SSC vs FSC density plot. (B) Lymphocytes were gated to CD43 vs CD69 plot, (C) histogram of CD43+CD69- cells vs CD27, (D) histogram of CD43+CD69-CD27+ vs CD20 to identify B1 cells (CD20+CD27+CD43+CD69-).
Figure 2.
Gating strategy and B1 cell frequency. (A) SSC vs FSC density plot. (B) Lymphocytes were gated to CD43 vs CD69 plot, (C) histogram of CD43+CD69- cells vs CD27, (D) histogram of CD43+CD69-CD27+ vs CD20 to identify B1 cells (CD20+CD27+CD43+CD69-).
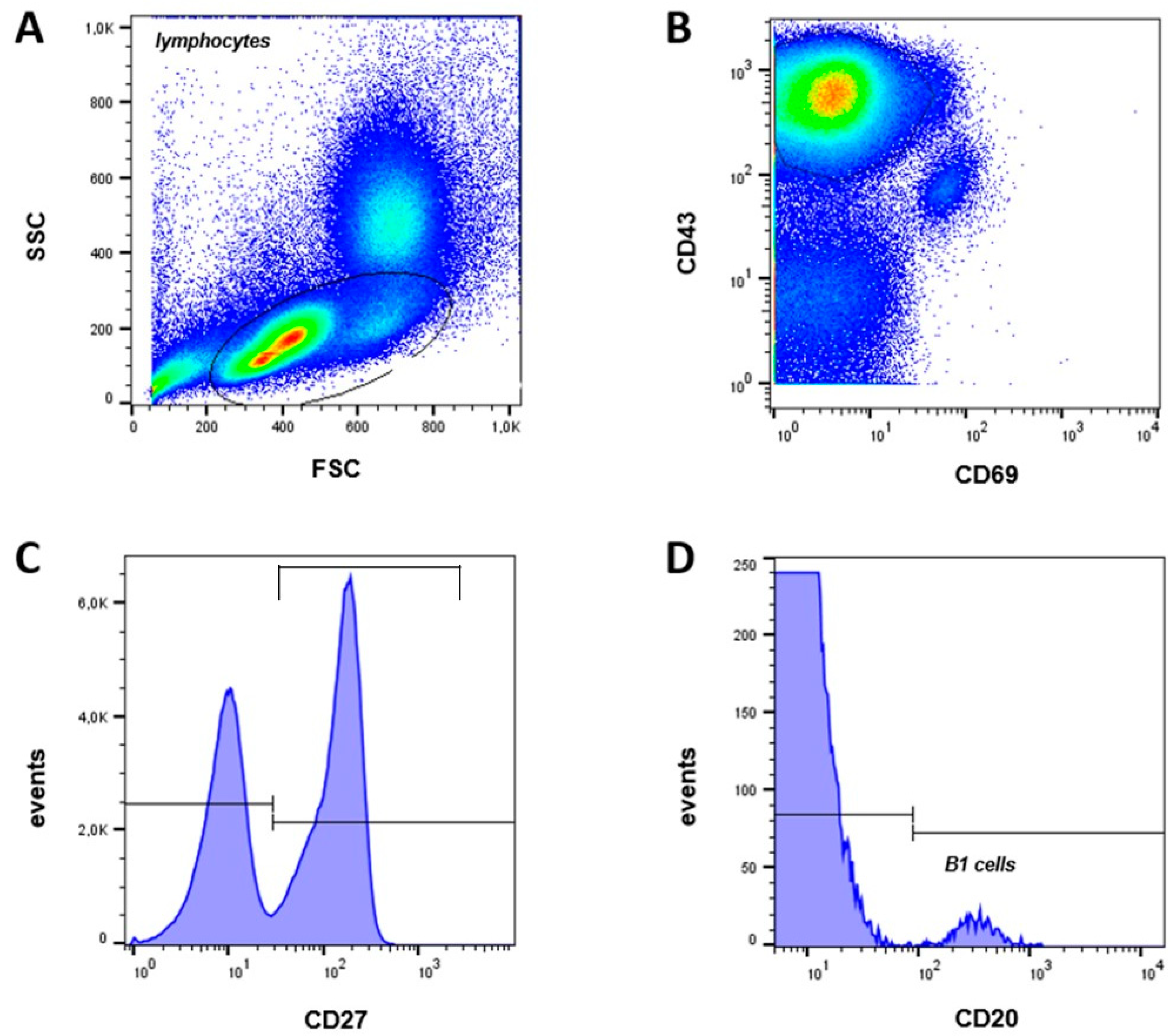
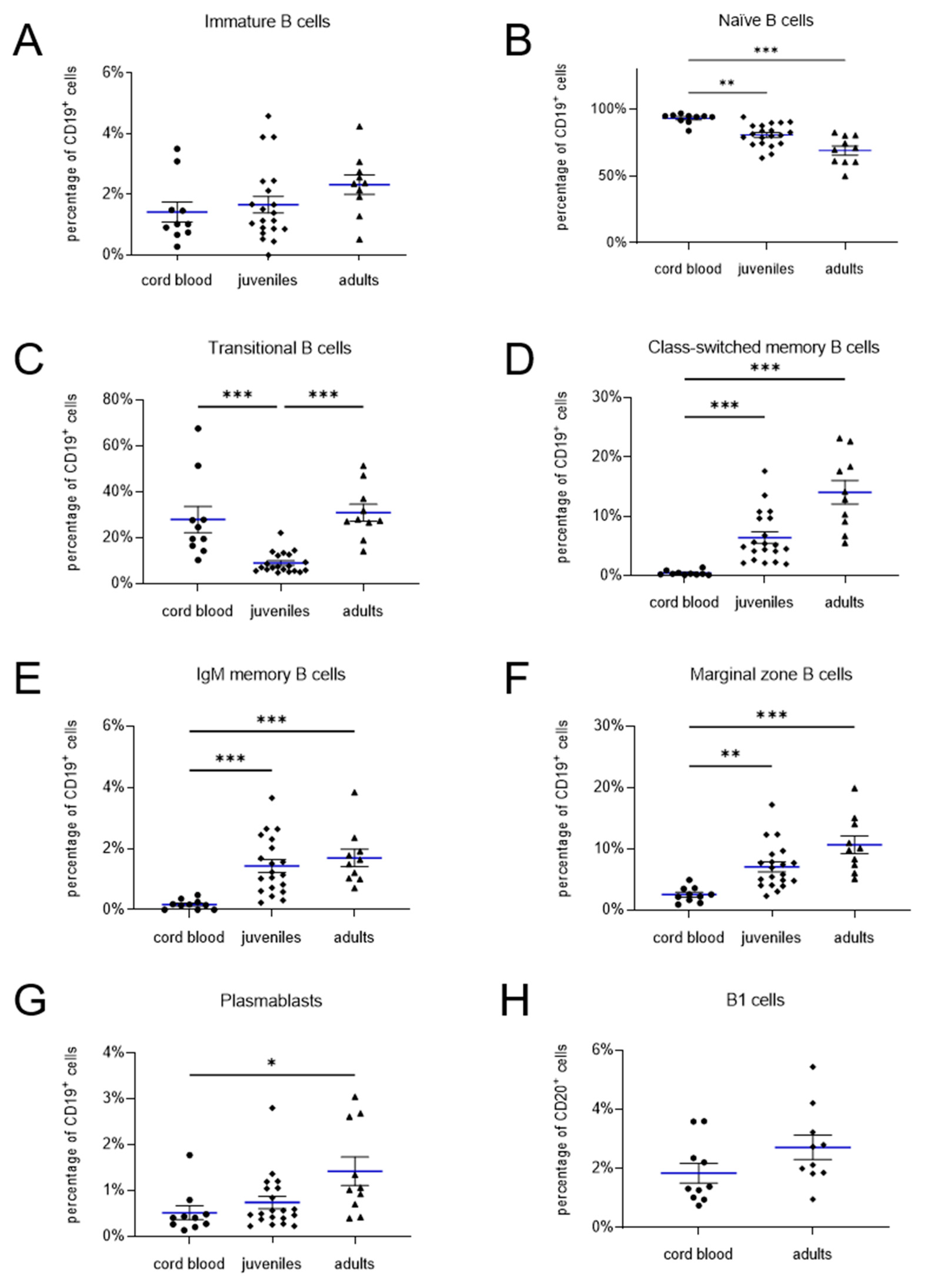
Figure 3.
Quantification of lymphocyte subpopulations in cord blood, juveniles and adults. Immature B cells (A), naïve B cells (B), transitional B cells (C), marginal zone B cells (D), IgM memory B cells (E), class switched memory B cells (F) and plasmablasts (G) values refer to percentage of CD19+ cells, B1 cells (H) values refer to percentage of CD20+ cells. The frequency of immature B cells (A) as well as B1 cells (H) does not differ between neonatal cord blood and non-neonatal peripheral blood. Naïve B cells (B) are more prominent in cord blood and decreases with age. The frequency of the transitional B cells (C) decreases from the cord blood to the children and increases again in the adults. However, the percentage of class-switched and IgM memory B cells (D, E), marginal zone B cells (F), and plasmablasts (G) increases with age. (mean shown as blue lines, SEM shown as black bars).
Figure 3.
Quantification of lymphocyte subpopulations in cord blood, juveniles and adults. Immature B cells (A), naïve B cells (B), transitional B cells (C), marginal zone B cells (D), IgM memory B cells (E), class switched memory B cells (F) and plasmablasts (G) values refer to percentage of CD19+ cells, B1 cells (H) values refer to percentage of CD20+ cells. The frequency of immature B cells (A) as well as B1 cells (H) does not differ between neonatal cord blood and non-neonatal peripheral blood. Naïve B cells (B) are more prominent in cord blood and decreases with age. The frequency of the transitional B cells (C) decreases from the cord blood to the children and increases again in the adults. However, the percentage of class-switched and IgM memory B cells (D, E), marginal zone B cells (F), and plasmablasts (G) increases with age. (mean shown as blue lines, SEM shown as black bars).
3. Discussion
We provide reference values for eight B lymphocyte subpopulations, including B1 cells, in cord blood of healthy mature newborns. The human B1 cell population, which appears to be subject to changes during ontogeny, has been re-defined by flow cytometric and functional analyses [18,19,20,21,22].
Our results show significant changes in relative counts of B cell subpopulations in cord blood compared to juveniles and adult peripheral blood. Age-dependent accumulation of immune experience resulting from contact with foreign antigens leads to a gradual shift from naïve to mature B cells. Meanwhile the total number of B lymphocytes gradually decreases with age along with their increasing ability to respond to foreign antigens [15]. B cell subpopulations are defined by characteristic expression patterns of surface antigens, cytokines and master transcription factors. We found that the proportion of immature B cells, defined as CD19+CD27-IgM-IgD-, is similar in newborns, juveniles and adults. The immature cells enter the spleen, lymph nodes, or other lymphoid tissues. There they mature into long-lived, adult naïve B cells [16,23,24]. Naïve B cells continue to express IgD at the surface, but they do not yet display CD27 expression, which is induced by antigen-receptor activation in B cells. In umbilical cord blood of newborns, naïve B cells (CD19+CD27-IgM+IgD+) represent 93 % of the total population of CD19+ cells and thus the frequency is 25 % higher than in the peripheral blood of adults. This means the population of naïve B cells decreases with age, with the relative counts for the population under study higher than described previously. Piątosa et al. showed that naïve CD19+CD27-IgD+ cells increase during the first year of life and gradually decrease during the following few years, reaching plateau in children aged between 5 and 10 years [15]. In order to propose a convenient flow cytometric approach, we decided to use CD19, CD38 and IgM expression to delineation of B cells. The population of transitional B cells defined by CD19+CD27-IgM+CD38+ phenotype [19,20,21,25] represent an intermediate stage between the immature B cells of bone marrow and mature B cells of the periphery [23]. We found similar frequencies of transitional B cells in neonates and adults, whereas a decline with age has been reported previously [15,43]. Our data show that cells with CD19+CD27-IgM+CD38+ phenotype decreases from cord blood to infants but increases again in adulthood. The mature naïve B cells differentiate into memory and plasma cells [29]. CD19+CD27+IgM+IgD- memory B cells were virtually absent in cord blood, most likely reflecting the lack of exposure to extrauterine antigens in comparison to adults where this cell population represents about 15 % of all CD19+ B lymphocytes [10,15,26,27]. The population of CD19+CD27+IgM+IgG+ marginal zone B lymphocytes is small in cord blood accumulates in peripheral blood once the marginal zone develops during the first two years of life [28]. The proportion of CD27+CD38++ plasmablasts did significantly differ between cord blood, juveniles and adult blood and almost exceeded 5 % of total B cells [14].
B1 cells stand apart from the conventional B cell subpopulations and can be classified as elements of the innate immune system since they constitutively and spontaneously secrete „natural“ IgM antibodies [8,29]. B1 cells differ from conventional B cells by their surface phenotype, function, localization and ontogenesis [30,31]. The identity and even existence of human B1 cells was under debate because of the absence of known surface markers for this population. B1 cells were described first in mice as CD5+ B cells, now known as B1a cells. Human B1 cells were initially defined as CD19+CD5+ [8,32,33]. However, in humans CD5+ is expressed on several other human B cell subpopulations, including pre-naïve, transitional, and activated B cells [20]. Inspired by Griffin et al., who re-defined human B1 cells according to morphological and functional studies, we defined B1 cells as CD20+CD27+CD43+CD69- [13]. CD20 was used as B cell marker because it is lost during mature B cell differentiation [34], unlike CD19, and this assured that immunoglobulin-secreting plasmablasts and plasma cells are excluded. So far, it appears that no other B cells beyond B1 cells constitutively express CD43, even though naïve and memory B cells do so after activation. However, CD43-expressing activated naïve and memory B cells express CD69 and CD70, whereas CD43−expressing B1 cells do not [13]. Previous studies have shown, that the frequency of human B1 cells decreases with age [13,35,36] while the frequency of murine B1 cells increases with age [37]. In our study, we found a slight increase in B1 cells from neonatal cord blood in comparison to adults, but this difference did not reach statistical significance. Neonates as well as adults have a population size of about 1 – 3 % of CD20+ B cells. Griffin et al. described a variation in the frequency of circulating B1 cells with a range from less than 1 % to more than 9 % and an age-dependent trend of the B1 cells as well [38]. Recent studies revealed that CD20+CD27+CD43+ D70- B1 cells are closely related to plasmablasts and pre-plasmablasts according to their gene expression profiles and functional characteristics [39,40]. These CD20+CD27+CD43+CD70- cells were later found to include CD20+CD38hi precursors for plasmablasts and pre-plasmablasts [41]. Consequently, CD20+CD27+CD43+CD38lo/int is currently regarded as a more rigorous and refined phenotype of human B1 cells. Prabhu et al. defined B1 cells as CD19+CD20brightCD38dullCD27+CD43+ and found, in line with our data, an increase in B1 cells from neonatal cord blood in comparison to adults [42]. For these reasons, it is necessary in the future to compare the effects of all these definitions currently available for B1 cells on their quantity in the same samples in order to be able to evaluate the results in a disease-associated or immunologically regulated context.
In conclusion, we found that B cell subpopulations differ considerably between cord blood, juveniles and adult blood. These changes most likely represent an overlap between developmental maturation and the establishment of an adaptive immunological memory. A limitation of the study is the low sample size and the scarce number of markers used to phenotype the B cells. In addition, we must consider the peripheral compartment might differ from the real quantity of B cells, e.g. plasmablasts tend to localize in the bone marrow. There are only few studies about different B cell subsets in cord blood of newborn. Hence, the strength of our study is the consideration of cord blood as a link between intrauterine and extrauterine development.
4. Materials and Methods
4.1. Patient samples
Blood samples were collected from umbilical cord blood of term neonates (range 37-42 weeks gestation), from peripheral blood of healthy juveniles (mean 4 years, range 1-17 years) and adult donors (mean 37.5 years, range 24-62 years) without infectious, immunologic, hematologic, or other chronical disease or treatment of any kind, which could affect the immune system. Umbilical cord blood was collected in vacutainer tubes containing ethylenediaminetetraacetic acid (EDTA) by venipuncture from healthy neonate cords immediately following uncomplicated delivery. Adult and juveniles peripheral blood samples were obtained in EDTA. All subjects were Caucasians. The study was conducted in accordance with the guidelines of the World Medical Association's Declaration of Helsinki. The Institutional Review Boards of the Philipps-University Marburg approved the study protocol. The written informed consent of parents and adult donors was obtained.
4.2. Cell isolation and B cell enrichment
For cell isolation, we used 8-10 ml of blood. Erythrocytes were lysed and peripheral blood mononuclear cells (PBMCs) or cord blood mononuclear cells (CBMC) were isolated by Ficoll-Hypaque gradient (PAA, Linz, Austria). 1 x 107 untouched B cells were enriched by negative selection using Magnetic Bead-Activated Cell Sorting (MACS) with B cell (B-CLL) isolation kit (MiltenyiBiotec, Bergisch Gladbach, Germany) according to the manufacturer`s instructions.
4.3. Flow cytometry
To quantify the B lymphocyte subsets, they were labelled according to Supplemental Table S1 with anti-CD19 (PE/APC), anti-CD20-FITC, anti-CD27 (FITC/PE/PerCP), anti-CD38 (APC), anti-CD43 (APC), anti-CD69 (PE), anti-IgD (FITC) (all from BD Biosciences, Heidelberg, Germany) and anti-IgM (PerCP) (BioLegend, Fell, Germany). All staining steps were performed in CellWASH buffer (BD Biosciences, Heidelberg, Germany). Cells were stained using a master mix of antibodies to maintain consistency among the samples.
Flow cytometry was directly performed on a four-colour FACS Calibur flow cytometer (BD Biosciences, Heidelberg, Germany) using Summit 4.3 software (Beckman Coulter, Krefeld, Germany). Cells were gated into a physical lymphocyte gate in the forward scatter/side scatter plot. Gates were preset and the measurements were performed blinded for sample identity.
4.4. Statistical Analysis
Statistical analyses were performed using Prism 9.5 (GraphPad Software, La Jolla, California) and SPSS (Chicago, USA). Normal distribution was analyzed using Shapiro-Wilk-test. Data with normal distribution were tested for group differences using either the two-tailed unpaired t-test or the ordinary one-way ANOVA test followed by Holm-Šídák's multiple comparisons post-test. For non-normally distributed data, Kruskal-Wallis test followed by Dunn’s multiple comparisons post-test was used. Differences with p-values of p < 0.05 were deemed significant. Means are given with standard error (SEM, Table 1).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: title; Table S2: title.
Author Contributions
SGF: data analysis and interpretation, manuscript writing, final approval of manuscript. MB: data interpretation, manuscript writing, final approval of manuscript. RW, TS: manuscript writing, final approval of manuscript. EK: statistical analysis, manuscript writing. SM: collection and assembly of data, data analysis and interpretation, manuscript writing, final approval of manuscript. TR, SK, RFM, MZ: conception and design, manuscript writing, final approval of manuscript.
Funding
Funded by a grant from the “von Behring-Roentgen Foundation” and by a grant from the “Rhoen Klinikum AG”.
Institutional Review Board Statement
The Ethics Committee of the Philipps-University Marburg (AZ 20/07) approved this study.
Informed Consent Statement
The written informed consent of parents and adult donors was obtained
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article or are available from the authors upon request.
Acknowledgments
We thank Sabine Jennemann and Regina Stoehr for excellent technical help.
Conflicts of Interest
Tobias Rogosch is an employee of CSL Behring. The remaining authors declare no conflict of interest.
References
- LeBien, T.W.; Tedder, T.F. B lymphocytes: how they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Comans-Bitter, W.M.; de Groot, R.; van den Beemd, R.; Neijens, H.J.; Hop, W.C.; Groeneveld, K.; Hooijkaas, H.; van Dongen, J.J. Immunophenotyping of blood lymphocytes in childhood. Reference values for lymphocyte subpopulations. J Pediatr 1997, 130, 388–393. [Google Scholar] [CrossRef]
- Bufe, A.; Peters, M. Unterschiede zwischen kindlichem und erwachsenem Immunsystem. Haut (2013).
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: virtues of innate-like antibody-producing lymphocytes. Nat Rev Immunol 2013, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Berrón-Ruíz, L.; López-Herrera, G.; Ávalos-Martínez, C.E.; Valenzuela-Ponce, C.; Ramírez-SanJuan, E.; Santoyo-Sánchez, G.; Mújica Guzmán, F.; Espinosa-Rosales, F.J.; Santos-Argumedo, L. Variations of B cell subpopulations in peripheral blood of healthy Mexican population according to age: Relevance for diagnosis of primary immunodeficiencies. Allergol Immunopathol (Madr). 2016, 44, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mei, H.; Dörner, T.; Hiepe, F.; Radbruch, A.; Fillatreau, S.; Hoyer, B.F. Memory B and memory plasma cells. Immunol Rev 2010, 237, 117–139. [Google Scholar] [CrossRef]
- Tarlinton, D.; Good-Jacobson, K. Diversity among memory B cells: origin, consequences, and utility. Science 2013, 341, 1205–1211. [Google Scholar] [CrossRef]
- Baumgarth, N. The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol 2011, 11, 34–46. [Google Scholar] [CrossRef]
- Hardy, R.R. B-1 B Cell Development. The Journal of Immunology 2006, 177, 2749–2754. [Google Scholar] [CrossRef]
- Klein, U.; Rajewsky, K.; Küppers, R. Human immunoglobulin (Ig)M+IgD+ peripheral blood B cells expressing the CD27 cell surface antigen carry somatically mutated variable region genes: CD27 as a general marker for somatically mutated (memory) B cells. J Exp Med 1998, 188, 1679–1689. [Google Scholar] [CrossRef]
- Tangye, S.G.; Liu, Y.J.; Aversa, G.; Phillips, J.H.; de Vries, J.E. Identification of functional human splenic memory B cells by expression of CD148 and CD27. J Exp Med 1998, 188, 1691–1703. [Google Scholar] [CrossRef]
- Jazayeri, M.H.; Pourfathollah, A.A.; Jafari, M.E.; Rasaee, M.J.; Dargahi, Z.V. The association between human B-1 cell frequency and aging: From cord blood to the elderly. Biomedicine & Aging Pathology 2013, 3, 20–22. [Google Scholar] [CrossRef]
- Griffin, D.O.; Holodick, N.E.; Rothstein, T.L. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+ CD27+ CD43+ CD70-. J Exp Med 2011, 208, 67–80. [Google Scholar] [CrossRef]
- Morbach, H.; Eichhorn, E.M.; Liese, J.G.; Girschick, H.J. Reference values for B cell subpopulations from infancy to adulthood. Clin Exp Immunol 2010, 162, 271–279. [Google Scholar] [CrossRef]
- Piątosa, B.; Wolska-Kuśnierz, B.; Pac, M.; Siewiera, K.; Gałkowska, E.; Bernatowska, E. B cell subsets in healthy children: reference values for evaluation of B cell maturation process in peripheral blood. Cytometry B Clin Cytom 2010, 78, 372–381. [Google Scholar] [CrossRef]
- Dehne, J. Charakterisierung und Quantifizierung von Lymphozyten-Subpopulationen bei Kindern und Jugendlichen mit allergischen Erkrankungen. Marburg: Fachbereich Medizin der Philipps-Universität Marburg (2016).
- Marie-Cardine, A.; Divay, F.; Dutot, I.; Green, A.; Perdrix, A.; Boyer, O.; Contentin, N.; Tilly, H.; Tron, F.; Vannier, J.-P.; et al. Transitional B cells in humans: characterization and insight from B lymphocyte reconstitution after hematopoietic stem cell transplantation. Clin Immunol 2008, 127, 14–25. [Google Scholar] [CrossRef]
- Weller, S.; Braun, M.C.; Tan, B.K.; Rosenwald, A.; Cordier, C.; Conley, M.E.; Plebani, A.; Kumararatne, D.S.; Bonnet, D.; Tournilhac, O.; et al. Human blood IgM “memory” B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood 2004, 104, 3647–3654. [Google Scholar] [CrossRef]
- Lee, J.; Kuchen, S.; Fischer, R.; Chang, S.; Lipsky, P.E. Identification and characterization of a human CD5+ pre-naive B cell population. J Immunol 2009, 182, 4116–4126. [Google Scholar] [CrossRef] [PubMed]
- Sims, G.P.; Ettinger, R.; Shirota, Y.; Yarboro, C.H.; Illei, G.G.; Lipsky, P.E. Identification and characterization of circulating human transitional B cells. Blood 2005, 105, 4390–4398. [Google Scholar] [CrossRef] [PubMed]
- Palanichamy, A.; Barnard, J.; Zheng, B.; Owen, T.; Quach, T.; Wei, C.; Looney, R.J.; Sanz, I.; Anolik, J.H. Novel human transitional B cell populations revealed by B cell depletion therapy. J Immunol 2009, 182, 5982–5993. [Google Scholar] [CrossRef] [PubMed]
- Huck, K.; Feyen, O.; Ghosh, S.; Beltz, K.; Bellert, S.; Niehues, T. Memory B-cells in healthy and antibody-deficient children. Clin Immunol 2009, 131, 50–59. [Google Scholar] [CrossRef]
- Viau, M.; Zouali, M. B-lymphocytes, innate immunity, and autoimmunity. Clinical Immunology 2005, 114, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Warnatz, K.; Schlesier, M. Flowcytometric phenotyping of common variable immunodeficiency. Cytometry Part B: Clinical Cytometry 2008, 74B, 261–271. [Google Scholar] [CrossRef]
- Wehr, C.; Kivioja, T.; Schmitt, C.; Ferry, B.; Witte, T.; Eren, E.; Vlkova, M.; Hernandez, M.; Detkova, D.; Bos, P.R.; et al. The EUROclass trial: defining subgroups in common variable immunodeficiency. Blood 2008, 111, 77–85. [Google Scholar] [CrossRef]
- Banchereau, J.; Rousset, F. Human B lymphocytes: phenotype, proliferation, and differentiation. Adv Immunol 1992, 52, 125–262. [Google Scholar] [CrossRef] [PubMed]
- Klein, U.; Goossens, T.; Fischer, M.; Kanzler, H.; Braeuninger, A.; Rajewsky, K.; Küppers, R. Somatic hypermutation in normal and transformed human B cells. Immunol Rev 1998, 162, 261–280. [Google Scholar] [CrossRef]
- Sanz, I.; Wei, C.; Lee, F.E.-H.; Anolik, J. Phenotypic and functional heterogeneity of human memory B cells. Semin Immunol 2008, 20, 67–82. [Google Scholar] [CrossRef]
- Hayakawa, K.; Hardy, R.R.; Parks, D.R.; Herzenberg, L.A. The “Ly-1 B” cell subpopulation in normal immunodefective, and autoimmune mice. J Exp Med 1983, 157, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Hardy, R.R.; Stall, A.M.; Herzenberg, L.A.; Herzenberg, L.A. Immunoglobulin-bearing B cells reconstitute and maintain the murine Ly-1 B cell lineage. European Journal of Immunology 1986, 16, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Sidman, C.L.; Shultz, L.D.; Hardy, R.R.; Hayakawa, K.; Herzenberg, L.A. Production of immunoglobulin isotypes by Ly-1+ B cells in viable motheaten and normal mice. Science 1986, 232, 1423–1425. [Google Scholar] [CrossRef]
- Gupta, R.; Jain, P.; Deo, S.V.S.; Sharma, A. Flow cytometric analysis of CD5+ B cells: a frame of reference for minimal residual disease analysis in chronic lymphocytic leukemia. Am J Clin Pathol 2004, 121, 368–372. [Google Scholar] [CrossRef]
- Wuttke, N.J.; Macardle, P.J.; Zola, H. Blood group antibodies are made by CD5+ and by CD5- B cells. Immunol Cell Biol 1997, 75, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Jego, G.; Bataille, R.; Pellat-Deceunynck, C. Interleukin-6 is a growth factor for nonmalignant human plasmablasts. Blood 2001, 97, 1817–1822. [Google Scholar] [CrossRef] [PubMed]
- Descatoire, M.; Weill, J.-C.; Reynaud, C.-A.; Weller, S. A human equivalent of mouse B-1 cells? J Exp Med 2011, 208, 2563–2564. [Google Scholar] [CrossRef]
- Perez-Andres, M.; Grosserichter-Wagener, C.; Teodosio, C.; van Dongen, J.J.M.; Orfao, A.; van Zelm, M.C. The nature of circulating CD27+CD43+ B cells. J Exp Med 2011, 208, 2565–2566. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.M.; Blevins, M.W.; High, K.P.; Pang, B.; Swords, W.E.; Yammani, R.D. Aging promotes B-1b cell responses to native, but not protein-conjugated, pneumococcal polysaccharides: implications for vaccine protection in older adults. J Infect Dis 2014, 209, 87–97. [Google Scholar] [CrossRef]
- Griffin, D.O.; Rothstein, T.L. Human B1 Cell Frequency: Isolation and Analysis of Human B1 Cells. Front Immunol 2012, 3. [Google Scholar] [CrossRef]
- Covens, K.; Verbinnen, B.; Geukens, N.; Meyts, I.; Schuit, F.; Van Lommel, L.; Jacquemin, M.; Bossuyt, X. Characterization of proposed human B-1 cells reveals pre-plasmablast phenotype. Blood 2013, 121, 5176–5183. [Google Scholar] [CrossRef]
- Inui, M.; Hirota, S.; Hirano, K.; Fujii, H.; Sugahara-Tobinai, A.; Ishii, T.; Harigae, H.; Takai, T. Human CD43+ B cells are closely related not only to memory B cells phenotypically but also to plasmablasts developmentally in healthy individuals. Int Immunol 2015, 27, 345–355. [Google Scholar] [CrossRef]
- Quách, T.D.; Rodríguez-Zhurbenko, N.; Hopkins, T.J.; Guo, X.; Hernández, A.M.; Li, W.; Rothstein, T.L. Distinctions among Circulating Antibody-Secreting Cell Populations, Including B-1 Cells, in Human Adult Peripheral Blood. J Immunol 2016, 196, 1060–1069. [Google Scholar] [CrossRef]
- Prabhu, S.B.; Rathore, D.K.; Nair, D.; Chaudhary, A.; Raza, S.; Kanodia, P.; Sopory, S.; George, A.; Rath, S.; Bal, V.; et al. Comparison of Human Neonatal and Adult Blood Leukocyte Subset Composition Phenotypes. PLoS ONE 2016, 11, e0162242. [Google Scholar] [CrossRef]
- Budeus, B.; Kibler, A.; Brauser, M.; Homp, E.; Bronischewski, K.; Ross, J.A.; Görgens, A.; Weniger, M.A.; Dunst, J.; Kreslavsky, T.; Vitoriano da Conceição Castro, S.; Murke, F.; Oakes, C.C.; Rusch, P.; Andrikos, D.; Kern, P.; Könin-ger, A.; Lindemann, M.; Johansson, P.; Hansen, W.; Lundell, A.C.; Rudin, A.; Dürig, J.; Giebel, B.; Hoffmann, D.; Küppers, R.; Seifert, M. Human Cord Blood B Cells Differ from the Adult Counterpart by Conserved Ig Re-pertoires and Accelerated Response Dynamics. J Immunol. 2021, 206, 2839–2851, Epub 2021 Jun 11. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gating strategy and B cells subpopulations frequency. (A) SSC vs FSC density plot. (B) B cells: CD19+ (C), immature B cells (CD19+CD27-IgM-IgD-), naive B cells (CD19+CD27-IgM+IgD+), (D) transitional B cells (CD19+CD27-IgM+CD38+), (E) marginal zone B cells (CD19+CD27+IgM+IgD+), IgM memory B cells (CD19+CD27+IgM+IgD-), class-switched memory B cells (CD19+CD27+IgM-IgD-), (F) plasmablasts (CD19+CD27+IgM-CD38++).
Figure 1.
Gating strategy and B cells subpopulations frequency. (A) SSC vs FSC density plot. (B) B cells: CD19+ (C), immature B cells (CD19+CD27-IgM-IgD-), naive B cells (CD19+CD27-IgM+IgD+), (D) transitional B cells (CD19+CD27-IgM+CD38+), (E) marginal zone B cells (CD19+CD27+IgM+IgD+), IgM memory B cells (CD19+CD27+IgM+IgD-), class-switched memory B cells (CD19+CD27+IgM-IgD-), (F) plasmablasts (CD19+CD27+IgM-CD38++).
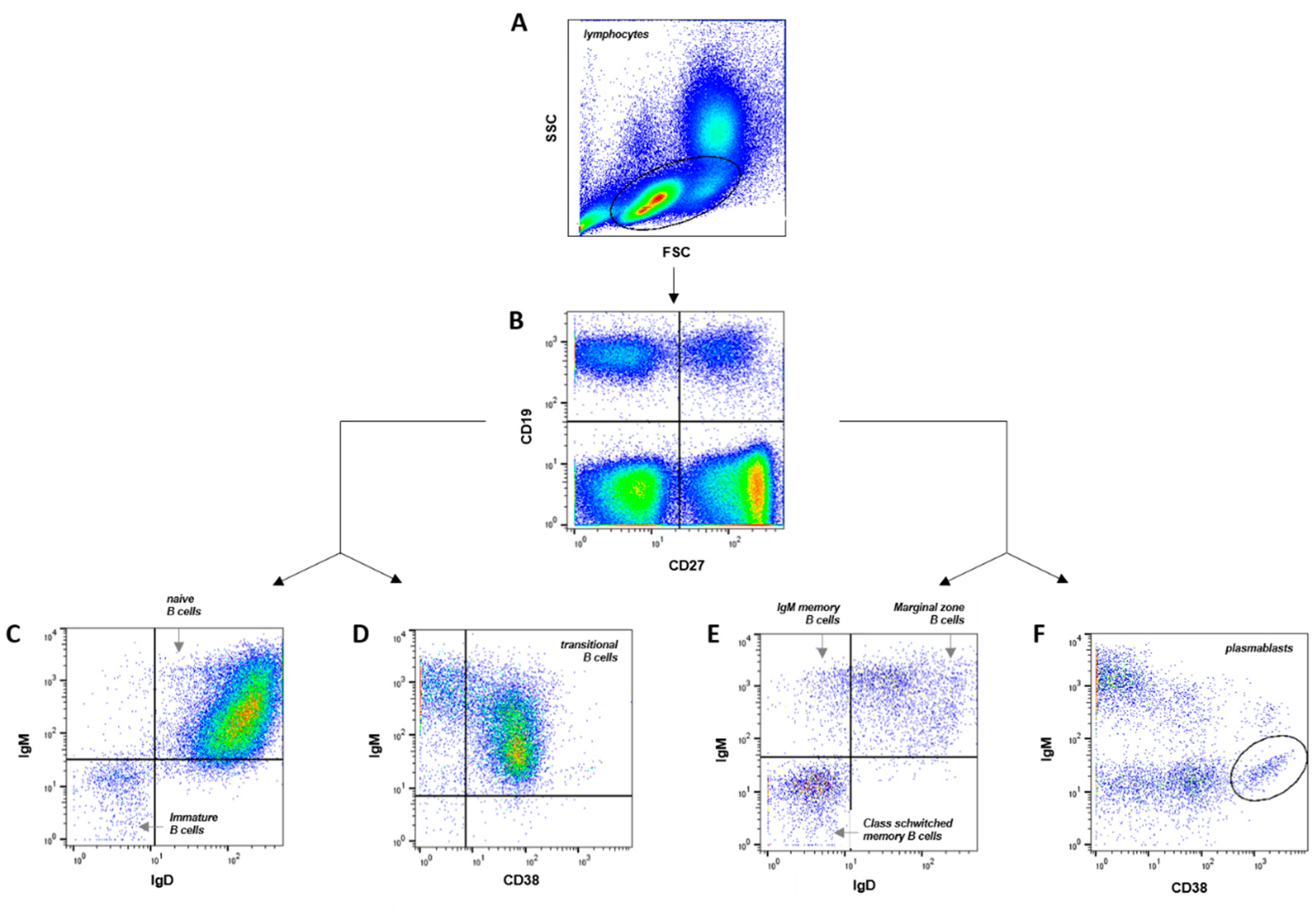

Table 1.
B cell populations in cord blood, juveniles and adult venous blood. Immature B cells, naïve B cells, transitional B cells, marginal zone B cells, IgM memory B cells, class switched memory B cells and plasmablasts values refer to percentage of CD19+ cells, B1 cells values refer to percentage of CD20+ cells.
Table 1.
B cell populations in cord blood, juveniles and adult venous blood. Immature B cells, naïve B cells, transitional B cells, marginal zone B cells, IgM memory B cells, class switched memory B cells and plasmablasts values refer to percentage of CD19+ cells, B1 cells values refer to percentage of CD20+ cells.
| Lymphocyte Subset | Neonates (Cord Blood) |
Juveniles (Peripheral Blood) |
Adults (Peripheral Blood) |
|---|---|---|---|
| immature B cells | 1.4 ± 0.3 % | 1.7 ± 0.3 % | 2.3 ± 0.3 % |
| naïve B cells | 93.2 ± 1.2 % | 80.8 ± 1.8 % | 69.1 ± 3.4 % |
| transitional B cells | 27.9 ± 5.7 % | 9.0 ± 1.0 % | 30.9 ± 3.7 % |
| marginal zone B cells | 2.5 ± 0.4 % | 7.1 ± 0.8 % | 10.7 ± 1.4 % |
| IgM memory B cells | 0.2 ± 0.1 % | 1.4 ± 0.2 % | 1.7 ± 0.3 % |
| class switched memory B cells | 0.5 ± 0.1 % | 6.4 ± 1.0 % | 14.1 ± 2.0 % |
| plasmablasts | 0.5 ± 0.2 % | 0.7 ± 0.1 % | 1.4 ± 0.3 % |
| B1 cells (% of CD20+ cells) | 1.8 ± 0.3 % | - | 2.7 ± 0.4 % |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



