
Submitted:
09 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
The use of several light-related variables, such as the Secchi disc depth, the euphotic depth, and in particular, the diffuse attenuation coefficient (Kd), is deeply rooted in phytoplankton research, but these are not the most appropriate indicators of the amount of light available for photosynthesis. We argue that the variable of interest for phytoplankton is the mean light intensity in the mixed layer (Im), which represents the mean light to which phytoplankton cells are exposed throughout their life cycle, while being continuously mixed in the mixed layer. We use empirical data collected in different coastal ecosystems in southern Portugal to demonstrate why Im should be the preferred metric instead of the deeply rooted Kd. We show that, although the relationship between Im and Kd is inversely proportional, it is not always strong or even significant. Different Im values can be associated to the same Kd, but distinct Im have different physiological effects of phytoplankton. Therefore, Kd does not capture the amount of light available for photosynthesis, given that, unlike Im, Kd calculation does not consider the depth of the mixed layer. Therefore, we urge phytoplankton researchers to consider the measurement and calculation of Im when evaluating light-related processes in phytoplankton ecology.
Keywords:
phytoplankton
; light measurements
; light limitation
; coastal ecosystems
1. Introduction
Light availability is of paramount importance for phytoplankton, especially in turbid ecosystems [1,2,3,4,5], but it has not yet received the same attention as nutrients as an environmental driver of phytoplankton dynamics. Indeed, studies dealing with the effects of nutrients on phytoplankton are abundant (e.g., [6,7]), whilst papers focused on the effects of light are rather scarce (e.g., [8,9]). The main reason behind this discrepancy is probably methodological. Dissolved inorganic macronutrients can be easily and directly analysed in the lab, using simple colorimetric methods (e.g., [10]). In addition, water samples can be collected and preserved for a considerable amount of time before the actual nutrient analysis is performed.
Measurements of underwater light are not as straightforward. Solar radiation reaching the Earth’s surface is composed by a large spectrum of radiation with different energies and wavelengths, including ultraviolet, infrared, and visible radiation. Photoautotrophic organisms, such as plants and algae, can only use a fraction of the total solar radiation for the process of photosynthesis; this photosynthetically active radiation (PAR) corresponds roughly to the visible light of the electromagnetic spectrum, with wavelengths between 400 and 700 nm. PAR constitutes approximately 50% of the total solar radiation that reaches the Earth’s surface, considering both direct and diffuse sources [11]. The amount of light that penetrates the water surface depends on several factors, such as solar elevation and its daily and annual variation [12], but a reflectance of 6.6% for a flat surface and irradiance coming equally from all directions is generally assumed [11]. The radiation that penetrates the water is scattered and eventually all photons are absorbed by water molecules and by dissolved and particulate matter, resulting in an exponential decrease of light intensity with depth, according to the Beer-Lambert law. The rate at which light disappears in the water column with depth can be expressed by the light extinction coefficient or diffuse attenuation coefficient (Kd). Kd is classified as an apparent optical property of water, depending on the composition of the medium and on the directional structure of the ambient light field [11].
The diffuse attenuation coefficient is typically estimated using the depth of disappearance of the Secchi disc (SD), as k/SD, where k is a constant; k values of 1.7 [13] and 1.44 [14], for non-turbid and turbid (euphotic depths <5 m) waters, respectively, are commonly used. However, the constant k can vary widely, between 1.27 and 2 [15], and this uncertainty may lead to errors in the estimation of light availability for phytoplankton. In addition, this equation does not account for all of Kd variability, which is affected by any optically active component and represents the sum of water, phytoplankton, seston and chromophoric dissolved organic matter (CDOM) variability [16]. Indeed, the relationship between SD and Kd may vary seven-fold in waters with high turbidity and CDOM variability [17].
The best estimate of Kd (m-1) is given by an exponential fit of light measurements in the water column as a function of depth, according to equation 1:
where Iz (µmol photons m-2 s-1) is the light intensity at depth level Z (m) and I0 is the light intensity at the water surface. Kd may also be expressed as (equation 2):
where I1 and I2 are the light intensities at depth levels Z1 and Z2, respectively. Z1 is the depth immediately below water surface. When light intensity in the water column is not available, Kd is generally estimated using empirical coefficients. Yet, a hyperbolic fit of Secchi disk measurements and Kd (obtained from vertical light attenuation measurements) data will result in a better function to estimate Kd from SD in a given ecosystem, than these constants (e.g., [16]).
The use of Kd is deeply rooted in phytoplankton research and many efforts have been made to develop better empirical relationships between SD and Kd (e.g. [18,19,20]). However, the diffuse attenuation coefficient per se does not provide any information on the quantity of PAR available for phytoplankton and, therefore, does not allow the evaluation of the underwater light environment and its role as a limiting factor of phytoplankton growth. To determine potential light limitation, light intensity values are needed, just like nutrient concentrations are required to assess nutrient limitation [21,22]. Thus, the mean light intensity in the mixed layer (Im) is a more relevant metric to evaluate the underwater light environment.
Im represents the mean PAR to which cells are exposed throughout most of their life cycle, while being continuously mixed in the mixed layer. Im is therefore a useful indicator of the underwater light environment, and it can be used to evaluate the occurrence of potential light limitation of phytoplankton growth. Im determination (equation 3) considers the incident light at the water surface (I0), the diffuse attenuation coefficient in the water column (Kd), and the depth of the mixed layer (Zm):
However, given the easiness of Secchi depth determination, the high cost of quanta sensors [20], and the common lack of daily integrated I0 measurements, necessary for a realistic determination of Im, Kd is frequently the only light-related variable determined in phytoplankton studies. In our opinion, Kd is used in a rather excessive and forced fashion to draw askew conclusions about the amount of light available for phytoplankton photosynthesis. It is commonly assumed that the relationship between Kd and light availability is inversely proportional, but light availability for phototrophs depends on the depth of the mixed layer, whereas Kd is unrelated with depth. For instance, a turbid (higher Kd) and shallow ecosystem (e.g., a coastal lagoon) may present higher light availability than a non-turbid system (lower Kd) with a deeper mixing layer (e.g., coastal zone, continental shelf, oceanic zone).
The assessment of underwater light availability is essential to understand phytoplankton dynamics and thus to assess the impacts of natural and human-induced perturbations to ecosystems. In this article we aim to expose constraints that phytoplankton researchers are faced with when characterizing the underwater light environment and to clarify the light-related variables of interest that, in our opinion, should be used in phytoplankton research and environmental monitoring. To accomplish this goal, we present underwater light data collected in distinct ecosystems in southern Portugal and analyse relationships between the diffuse attenuation coefficient and mean light intensity in the mixed layer.
2. Materials and Methods
Several light variables were measured in three distinct ecosystems in southern Portugal (SW Iberia): the Guadiana estuary (divided into freshwater, brackish, and marine zones), the Ria Formosa coastal lagoon, and the coastal zone adjacent to the Ria Formosa coastal lagoon. Measurements were integrated in different research projects aimed at phytoplankton dynamics and results were published elsewhere [5,23,24]. Data presented here was collected from 2001 to 2009 in the Guadiana estuary, and between 2012 and 2014 in the Ria Formosa coastal lagoon and adjacent coastal zone.
Light penetration in the water column was determined using two methods: a) calculation of Kd based on the disappearance of the Secchi disc and b) calculation of Im based on vertical profiles of PAR intensity. Using the disappearance of the Secchi disc, i.e., the Secchi depth (SD), the diffuse attenuation coefficient (kd, m-1) was calculated as 1.4/SD for highly turbid ecosystems [14], namely the Guadiana estuary, or as 1.7/SD for non-turbid ecosystems [13], namely the Ria Formosa coastal lagoon and the adjacent coastal zone. Vertical profiles of PAR intensity in the mixed layer (Im, µmol photons m-2 s-1) were measured using a LI-COR 4π quantum sensor and Kd was according to equation 1 (see Introduction). The mean light intensity in the mixed layer (Im, µmol photons m-2 s- 1) was calculated according to equation 3 (Jumars, 1993).
Data analysis was carried out by means of descriptive statistics; Pearson’s correlation was conducted to assess the relationships between Im and Kd. All data analysis were performed using IBM SPSS Statistics 28.
3. Results and Discussion
The relationship between the diffuse attenuation coefficient and the amount of light available for photosynthesis, expressed as the mean light intensity in the mixed layer, is typically inversely proportional, but not always significant. The highest Kd and lower Im values were found in the freshwater (Kd = 0.90 - 8.47 m-1, Im = 0.99 – 440.86 µmol photons m-2 s-1) and brackish (Kd = 0.85 - 14.40 m-1; Im = 0.42 – 104.97 µmol photons m-2 s-1) estuarine zones of the Guadiana estuary (Figure 1), whereas for coastal locations, Kd values were lower and Im was higher (Kd = 0.08 – 4.26 m-1, Im = 12.82 – 1397.64 µmol photons m-2 s-1) (Figure 2). The strength of the relationship between the two variables in the different coastal ecosystems was always weak, with determination coefficients ranging between 0.00 to 0.33.
It was also clear that similar Kd values were associated with different Im (Table 1), reflecting the importance of considering the depth of the mixed layer to properly evaluate PAR availability for photosynthesis. For instance, a Kd of 0.6 m-1 corresponds to Im values ranging between 100 and 427 µmol photons m-2 s-1 in the Ria Formosa coastal lagoon, and between 50 and 332 µmol photons m-2 s-1 in the Guadiana estuary (See Figure 1 and Figure 2). For each type of ecosystem, each Kd value presents a wide range of corresponding Im values; for instance, in coastal zone, a Kd of 0.09 m-1 was associated with Im values of 89 and 907 µmol photons m-2 s-1 (Table 1). These Im values may have different physiological effects on phytoplankton. For instance, light enrichment experiments conducted in the Guadiana estuary demonstrated that exposure of natural phytoplankton communities to Im values of 50, 70, 120, and 225 µmol photons m-2 s-1 had significantly different effects, from an enhancement to a decline of phytoplankton growth, leading to changes in phytoplankton biomass and community structure (Domingues et al. 2011b). Therefore, a Kd value of 0.6 m-1 may correspond not only to different mean light intensities in the mixed layer, but also to different degrees of light-limited growth and photoinhibition of phytoplankton photosynthesis. To avoid erroneous conclusions about the availability of underwater light for phytoplankton based solely on light attenuation coefficients, we urge phytoplankton researchers to include estimates of both Kd and Im in phytoplankton studies and monitoring programs. But how can Im me estimated in a timely and inexpensive manner?
Firstly, the depth of the mixed layer (Zm) should be established. The mixed layer is the top, unstratified layer in the water column where no significant density gradients are found, and phytoplankton is thus continuously mixed. Empirically, the mixed layer is where temperature or density variability with depth is less than 0.5ºC or 0.125 sigma-t, respectively [25]. In shallow, well mixed ecosystems, Zmix usually corresponds to the whole water column depth [26]. In an estuarine ecosystem, for instance, Zmix will vary along the channel’s cross-section and with tidal phase. A mean mixing depth should therefore be established for coastal, shallow waters, taking into consideration bathymetry and tidal amplitude. For deeper waters, the mixing depth should be determined in each sampling campaign, using vertical profiles of T and S.
A common lapse in phytoplankton studies is the characterization of light availability for phytoplankton using, besides Kd, the compensation depth, that corresponds to the bottom of the euphotic zone and is defined as the depth where PAR is 1% of I0. Given that this amount of PAR is sufficient to sustain photosynthesis, the compensation depth is another metric widely used in phytoplankton research (e.g., [27]). However, the euphotic layer and the mixed layer are usually neither coincident nor proportional, and whilst the euphotic depth depends on inherent optical properties of the water, the mixing depth is mainly affected by physical-meteorological forcing, bathymetry and even tides. For phytoplankton, it really does not matter how deep the euphotic zone is; what matters is the mean light intensity to which cells are exposed while being continuously mixed in the mixed layer. A useful approach, given by [28], combines both Zeu and Zmix to characterize light availability for phytoplankton growth, in the form of a mixing depth to euphotic depth ratio (Zmix:Zeu). It is usually considered that when the mixing depth is more than 5 times the euphotic depth (Zmix:Zeu > 5), no net phytoplankton growth will occur.
Secondly, the light attenuation coefficient must be determined to calculate Im. Two methods can be used, as referred above: vertical profiles of PAR in the water column or a function that relates Kd with Secchi depth. PAR measurements with a spherical quantum sensor are more reliable than the Secchi disk, which relies on the sensitivity of the human eye. But even vertical profiles of PAR are not exempt of problems. Several measurements of PAR intensity should be taken in the water column at specified depths (usually every meter), to adjust the exponential function. All measurements must made in the same conditions, considering intermittent cloud cover and making sure that the cable supporting the light sensor is always vertical. Kd is usually considered constant with depth, so only a few data points are necessary for an accurate estimate of Kd, especially in turbid systems, where light attenuation closely follows an exponential function and thus can be characterized by a single Kd [11]. In clear waters light attenuation with depth may present a biphasic behaviour, characterized by two Kd values [11].
It should be noted, however, that light attenuation in the water column may show significant daily variability. In shallow turbid ecosystems, significant and positive correlations between SPM and Kd are common, indicating that light attenuation is mainly controlled by suspended sediments [29,30], which in turn may show significant short-term variability associated to tidal cycles and river flow [2]. For instance, SPM in the Guadiana estuary is usually higher during spring tides, due to stronger tidal currents and higher resuspension of bottom sediments [30], and during flood, due to the resuspension of sediments deposited during the preceding long low tide slack [31]. However, light attenuation coefficients are typically considered constant throughout the day, which may lead to erroneous estimates of Im. In the Guadiana estuary, Kd varies along the semidiurnal cycle, with higher values during ebb and flood and lower at slack water. During an autumn 2008 spring tide, Kd ranged between 7.3 m-1 during flood and 3.9 m-1 three hours later, at high tide [30]. Considering Zm = 9.4 m and I0 = 1,000 µmol photons m-2 s-1, Im calculation based on Kd = 3.9 m-1 would be 27 µmol photons m-2 s-1, whilst using kd = 7.3 m-1, Im is 15 µmol photons m-2 s-1, roughly half of the first value.
Thirdly, incident light at the water surface (I0) shows a significant daily variability. If sampling is conducted in the early morning when incident solar radiation is lower, Im will be lower than if measurements were taken in the afternoon; likewise, sampling around noon will result in higher Im values. When these isolated estimates are taken as a proxy for the whole day, the mean light availability in the mixed layer over the light period may be severely under- or overestimated. Therefore, light measurements should consider the significant daily variability of solar radiation, to avoid inaccurate assessments of light limitation of phytoplankton growth. Ideally, Im estimates should use the mean radiation for the whole light period. Data on daily solar radiation should thus be obtained, either from a public database or by making continuous measurements of PAR at the water surface throughout the light period.
The mean light intensity in the mixed layer is the variable of interest to evaluate the underwater light environment for phytoplankton. However, to assess the occurrence of light limitation of phytoplankton growth using Im, a previous knowledge on how a phytoplankton community responds to light is necessary. The effects of light on phytoplankton growth and community structure can be determined using bioassays where micro- or mesocosms of natural phytoplankton assemblages are exposed to different light intensities. The outcomes of these experiments can be extrapolated to the field and used to assess the occurrence of light limitation of phytoplankton growth. Light addition experiments also provide threshold Im values that can be used to evaluate the occurrence of light limitation in natural phytoplankton communities (e.g., [4,8]). In addition, the measurement of carbon incorporation by phytoplankton (e.g., using the 14C method: [32]) under different light intensities allows the determination of a light-response curve of photosynthesis, known as a photosynthesis-irradiance (P-E) curve, which describes the variability of photosynthetic characteristics of phytoplankton over a range of light conditions. Again, experimental PAR values can be compared to Im values in the field, providing an array of information concerning the effects of light on phytoplankton, such as the saturating irradiance, photosynthetic efficiency, and the occurrence of photoinhibition.
4. Conclusions
The diffuse attenuation coefficient (Kd) is the most common indicator of light availability, used not only in phytoplankton research, but also in water quality studies. We argued that Kd is not the most appropriate metric to evaluate the underwater light environment, given that the amount of light to which phytoplankton is exposed to in the water column depends not only on the rate of attenuation, but also on the depth of the mixing layer. Therefore, we propose the measurement and estimation of the mean light intensity in the mixed layer (Im) instead of just Kd. We showed, using data collected in different coastal ecosystems in southern Portugal, that the same Kd value was associated with hugely different values of Im, and different Im values have distinct physiological consequences, namely in terms of light availability for photosynthesis. For instance, for the same incident light (I0) and Kd, Im will be higher for a shallower mixing depth, and lower for a deeper mixing depth. Therefore, Kd cannot properly characterize the underwater light environment for phototrophs. We urge phytoplankton and water quality researchers to consider the measurement of Im instead, as this indicator is more relevant from an ecophysiological standpoint.
Author Contributions
Conceptualization, R.D. and A.B.; formal analysis, R.D.; investigation, R.D.; writing – original draft, R.D; writing – review & editing, R.D. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
The Portuguese Foundation for Science and Technology (FCT) supported this work through projects UID/00350/2020 CIMA and LA/P/0069/2020 (Associate Laboratory ARNET), and a researcher contract (DL57/2016/CP1361/CT0017).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cole, B.E.; Cloern, J.E. Significance of Biomass and Light Availability to Phytoplankton Productivity in San Francisco Bay. Mar Ecol Prog Ser 1984, 17, 15–24. [CrossRef]
- Kromkamp, J.; Peene, J.; Rijswijk, P.; Sandee, A.; Goosen, N. Nutrients, Light and Primary Production by Phytoplankton and Microphytobenthos in the Eutrophic, Turbid Westerschelde Estuary (The Netherlands). Hydrobiologia 1995, 311, 9–19. [CrossRef]
- Kocum, E.; Underwood, G.J.C.; Nedwell, D.B. Simultaneous Measurement of Phytoplanktonic Primary Production, Nutrient and Light Availability along a Turbid, Eutrophic UK East Coast Estuary (the Colne Estuary). Mar Ecol Prog Ser 2002, 231, 1–12. [CrossRef]
- Domingues, R.B.; Anselmo, T.P.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Light as a Driver of Phytoplankton Growth and Production in the Freshwater Tidal Zone of a Turbid Estuary. Estuar Coast Shelf Sci 2011, 91, 526–535. [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Will Nutrient and Light Limitation Prevent Eutrophication in an Anthropogenically-Impacted Coastal Lagoon? Cont Shelf Res 2017, 141, 11–25. [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global Analysis of Nitrogen and Phosphorus Limitation of Primary Producers in Freshwater, Marine and Terrestrial Ecosystems. Ecol Lett 2007, 10, 1135–1142. [CrossRef]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient Co-Limitation of Primary Producer Communities. Ecol Lett 2011, 14, 852–862. [CrossRef]
- Carter, C.M.; Ross, A.H.; Schiel, D.R.; Howard-Williams, C.; Hayden, B. In Situ Microcosm Experiments on the Influence of Nitrate and Light on Phytoplankton Community Composition. J Exp Mar Biol Ecol 2005, 326, 1–13. [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Are Nutrients and Light Limiting Summer Phytoplankton in a Temperate Coastal Lagoon? Aquat Ecol 2015, 49, 127–146. [CrossRef]
- Hansen, H.P.; Koroleff, F. Determination of Nutrients. In Methods of Seawater Analysis; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; WILEY-VCH Verlag GmbH, 1999; pp. 159–228 ISBN 9783527613984.
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems; 3rd ed.; Cambridge University Press, 2011;
- Reynolds, C.S. Vegetation Processes in the Pelagic: A Model for Ecosystem Theory; Ecology Institute: Oldendorf/Luhe, 1997;
- Poole, H.H.; Atkins, W.R.G. Photo-Electric Measurements of Submarine Illumination throughout the Year. Journal of the Marine Biological Association of the United Kingdom 1929, 16, 297–324. [CrossRef]
- Holmes, R.W. The Secchi Disk in Turbid Coastal Waters. Limnol Oceanogr 1970, 15, 688–694. [CrossRef]
- Lee, Z.; Shang, S.; Du, K.; Wei, J. Resolving the Long-Standing Puzzles about the Observed Secchi Depth Relationships. Limnol Oceanogr 2018, 63, 2321–2336. [CrossRef]
- Xu, J.; Hood, R.R.; Chao, S.-Y. A Simple Empirical Optical Model for Simulating Light Attenuation Variability in a Partially Mixed Estuary. Estuaries 2005, 28, 572–580. [CrossRef]
- Koenings, J.P.; Edmundson, J.A. Secchi Disk and Photometer Estimates of Light Regimes in Alaskan Lakes: Effects of Yellow Color and Turbidity. Limnol Oceanogr 1991, 36, 91–105. [CrossRef]
- Mayora, G.; Devercelli, M. Spatio-temporal Variability in Underwater Light Climate in a Turbid River-floodplain System: Driving Factors and Estimation Using Secchi Disc. River Res Appl 2019, rra.3429. [CrossRef]
- Zhang, Y.; Liu, X.; Yin, Y.; Wang, M.; Qin, B. Predicting the Light Attenuation Coefficient through Secchi Disk Depth and Beam Attenuation Coefficient in a Large, Shallow, Freshwater Lake. Hydrobiologia 2012, 693, 29–37. [CrossRef]
- Padial, A.A.; Thomaz, S.M. Prediction of the Light Attenuation Coefficient through the Secchi Disk Depth: Empirical Modeling in Two Large Neotropical Ecosystems. Limnology (Tokyo) 2008, 9, 143–151. [CrossRef]
- Dortch, Q.; Whitledge, T.E. Does Nitrogen or Silicon Limit Phytoplankton Production in the Mississippi River Plume and Nearby Regions? Cont Shelf Res 1992, 12, 1293–1309. [CrossRef]
- Domingues, R.B.; Anselmo, T.P.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Nutrient Limitation of Phytoplankton Growth in the Freshwater Tidal Zone of a Turbid, Mediterranean Estuary. Estuar Coast Shelf Sci 2011, 91, 282–297. [CrossRef]
- Domingues, R.B.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Phytoplankton Composition, Growth and Production in the Guadiana Estuary (SW Iberia): Unraveling Changes Induced after Dam Construction. Science of the Total Environment 2012, 416, 300–313. [CrossRef]
- Krug, L.A.; Platt, T.; Barbosa, A.B. Delineation of Ocean Surface Provinces over a Complex Marine Domain (off SW Iberia): An Objective Abiotic-Based Approach. Reg Stud Mar Sci 2018, 18, 80–96. [CrossRef]
- Levitus, S. Climatological Atlas of the World Ocean. Eos, Transactions American Geophysical Union 1983, 64, 962. [CrossRef]
- Domingues, R.B.; Galvão, H.M. Phytoplankton and Environmental Variability in a Dam Regulated Temperate Estuary. Hydrobiologia 2007, 586, 117–134. [CrossRef]
- Golubkov, M.; Golubkov, S. Photosynthetically Active Radiation, Attenuation Coefficient, Depth of the Euphotic Zone, and Water Turbidity in the Neva Estuary: Relationship with Environmental Factors. Estuaries and Coasts 2023. [CrossRef]
- Cloern, J.E. Turbidity as a Control on Phytoplankton Biomass and Productivity in Estuaries. Cont Shelf Res 1987, 7, 1367–1381. [CrossRef]
- May, C.; Koseff, J.; Lucas, L.; Cloern, J.; Schoellhamer, D. Effects of Spatial and Temporal Variability of Turbidity on Phytoplankton Blooms. Mar Ecol Prog Ser 2003, 254, 111–128. [CrossRef]
- Domingues, R.B.; Anselmo, T.P.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Tidal Variability of Phytoplankton and Environmental Drivers in the Freshwater Reaches of the Guadiana Estuary (SW Iberia). Int Rev Hydrobiol 2010, 95, 352–369. [CrossRef]
- Garel, E.; Pinto, L.; Santos, A.; Ferreira, Ó. Tidal and River Discharge Forcing upon Water and Sediment Circulation at a Rock-Bound Estuary (Guadiana Estuary, Portugal). Estuar Coast Shelf Sci 2009, 84, 269–281. [CrossRef]
- Steeman-Nielsen, E. The Use of Radio-Active Carbon (C14) for Measuring Organic Production in the Sea. Journal du Counseil International pour l’Exploration de la Mer 1952, 18, 117–140. [CrossRef]
Figure 1.
Relationship between the light attenuation coefficient (determined through Secchi disc depth) and the mean light intensity in the mixed layer (determined through vertical profiles of PAR). Data points in red represent the Kd x Im pairs presented in Table 1. Data collected at different locations in the Guadiana estuary: A) freshwater zone (n = 97), B) brackish zone (n = 121), and C) marine zone (n = 46).
Figure 1.
Relationship between the light attenuation coefficient (determined through Secchi disc depth) and the mean light intensity in the mixed layer (determined through vertical profiles of PAR). Data points in red represent the Kd x Im pairs presented in Table 1. Data collected at different locations in the Guadiana estuary: A) freshwater zone (n = 97), B) brackish zone (n = 121), and C) marine zone (n = 46).
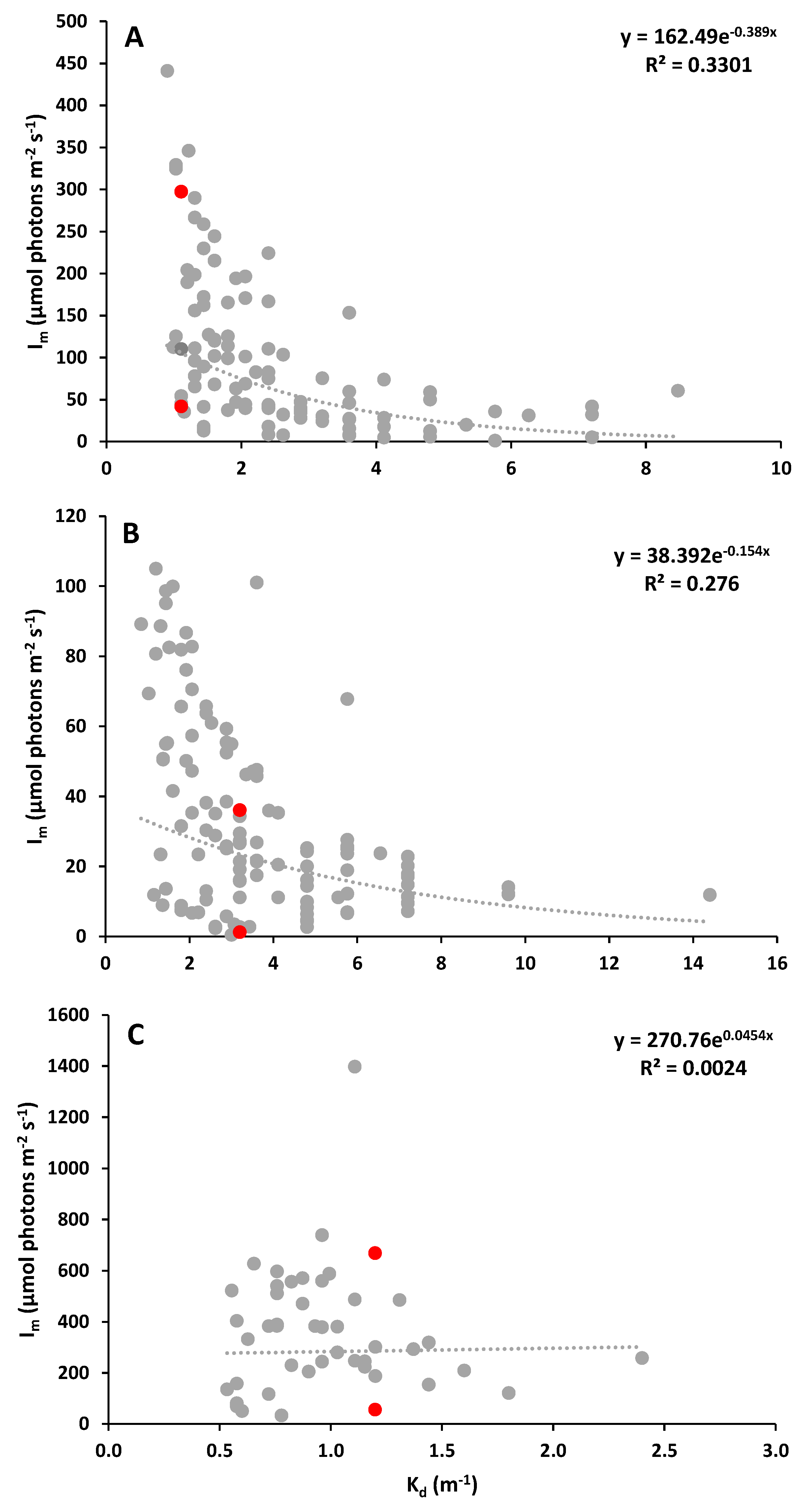
Figure 2.
Relationship between the light attenuation coefficient (determined through Secchi disc depth) and the mean light intensity in the mixed layer (determined through vertical profiles of PAR). Data points in red represent the Kd x Im pairs presented in Table 1. Data collected in A) Ria Formosa coastal lagoon (n = 71), and B) adjacent coastal zone (n = 15).
Figure 2.
Relationship between the light attenuation coefficient (determined through Secchi disc depth) and the mean light intensity in the mixed layer (determined through vertical profiles of PAR). Data points in red represent the Kd x Im pairs presented in Table 1. Data collected in A) Ria Formosa coastal lagoon (n = 71), and B) adjacent coastal zone (n = 15).
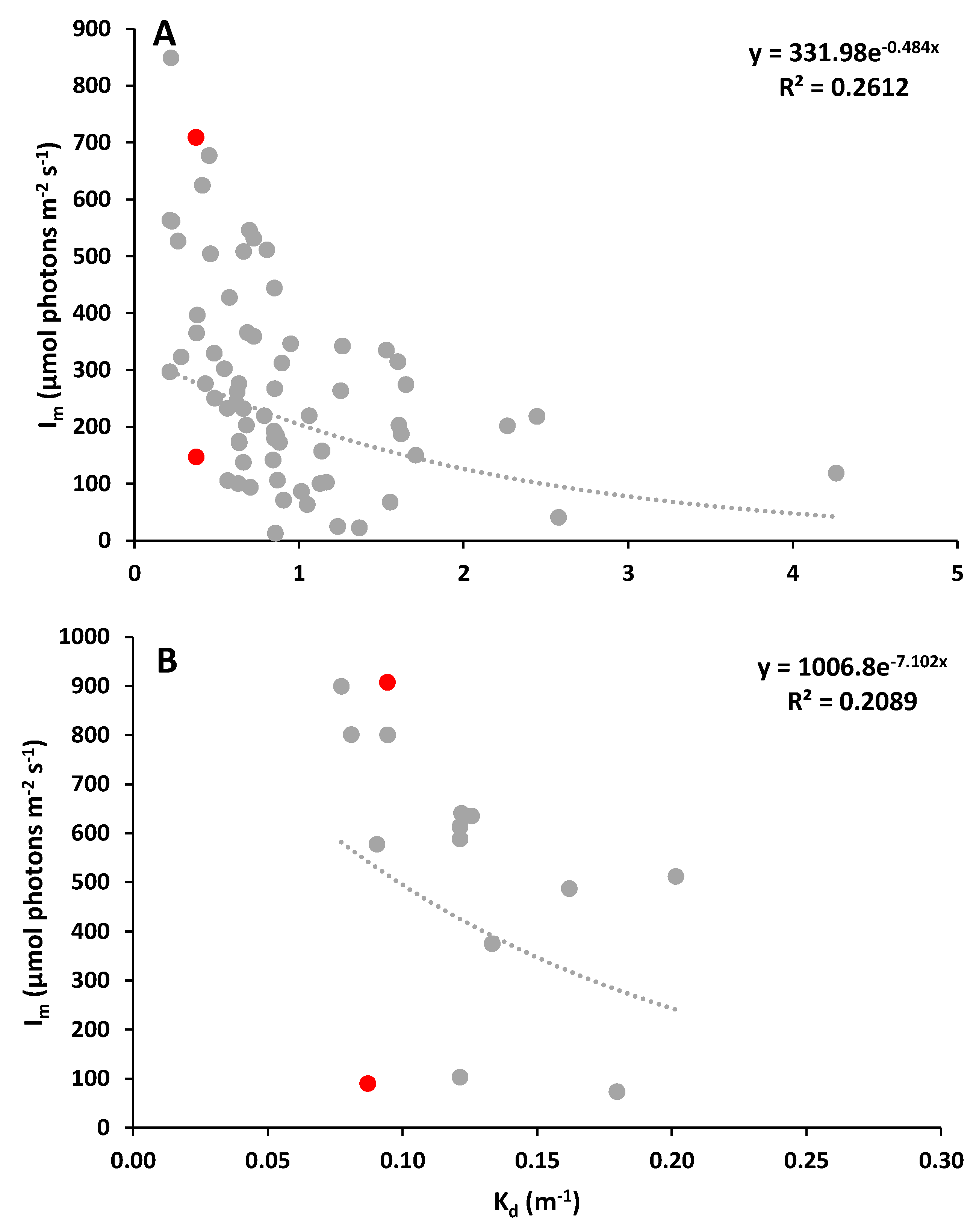

Table 1.
Examples of Kd (m-1, obtained using Secchi disc depth) and corresponding Im values (µmol photons m-2 s-1) for different types of ecosystems. The ten Kd x Im values are represented as red dots in Figure 1 and Figure 2.
| Ecosystem | Kd | Associated Im values |
|---|---|---|
| Estuary – freshwater | 1.1 | 42, 297 |
| Estuary – brackish | 3.2 | 1, 36 |
| Estuary – marine | 1.2 | 56, 669 |
| Coastal lagoon | 0.4 | 146, 708 |
| Adjacent coastal waters | 0.1 | 89, 907 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



