
Submitted:
02 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
This review systematically examines the multifaceted relationship between the tumor microenvironment, stromal interactions, and colorectal cancer (CRC) progression and metastasis. The analysis begins by assessing the gut microbiome's influence on CRC development, emphasizing its association with gut-associated lymphoid tissue (GALT). The role of the Wnt signaling pathway in CRC tumor stroma is scrutinized, elucidating its impact on disease progression. Tumor budding, its effect on tumor stroma, and implications for patient prognosis are investigated. The review also identifies conserved oncogenic signatures (COS) within CRC stroma and explores their potential as therapeutic targets. Lastly, the seed and soil hypothesis is employed to contextualize metastasis, accentuating the significance of both tumor cells and the surrounding stroma in metastatic propensity. In summary, this review highlights the intricate interdependence between CRC cells and their microenvironment, providing valuable insights into prospective therapeutic approaches targeting tumor-stroma interactions.
Keywords:
Colorectal cancer
; tumor stroma
; gut microbiome
; gut-associated lymphoid tissue
; Wnt signaling pathway
; tumor budding
; Conserved Oncogenic Signatures
; metastasis
1. Introduction
Colorectal cancer (CRC) represents a significant global health burden, ranking as the third most common cancer and the second leading cause of cancer-related deaths worldwide. According to the World Health Organization, approximately 1.8 million new cases of CRC were diagnosed, and nearly 900,000 deaths were reported in 2020 [1]. The high morbidity and mortality associated with CRC can be attributed to several factors, including late-stage diagnosis, limited treatment options, and therapy resistance.
Early detection and diagnosis of CRC are crucial for improving patient outcomes, as the 5-year survival rate for patients diagnosed at an early stage is significantly higher than for those diagnosed at advanced stages. Current screening methods for CRC include fecal occult blood tests (FOBT), fecal immunochemical tests (FIT), flexible sigmoidoscopy, and colonoscopy [2]. While these methods have been effective in reducing CRC incidence and mortality, they are not without limitations. Barriers to CRC screening include patient discomfort, invasiveness, financial constraints, and low adherence to screening guidelines.
Treatment options for CRC primarily depend on the stage of the disease and may include surgery, chemotherapy, radiotherapy, targeted therapy, and immunotherapy. Despite advances in surgical techniques and the development of novel therapeutic agents, the prognosis for patients with advanced or metastatic CRC remains poor, with a 5-year survival rate of less than 20% [3]. Furthermore, the emergence of therapy resistance and the occurrence of tumor recurrence after initial treatment contribute to the challenges associated with CRC management.
The complexity and heterogeneity of CRC, along with the dynamic interplay between cancer cells and the tumor microenvironment (TME), underscore the need for a deeper understanding of the underlying disease mechanisms. The tumor stroma in CRC shares many similarities with the stroma of other solid tumors, such as the presence of cancer-associated fibroblasts (CAFs), tumor-associated macrophages (TAMs), extracellular matrix (ECM) components, and immune cells. However, there are some specific features of the colorectal tumor stroma that distinguish it from other types of cancer (Figure 1):
1) Microbiome influence - bacteria can modulate the tumor stroma by influencing immune cell recruitment and activation, as well as by promoting a pro-inflammatory environment;
2) Mucosal immune system: the colorectal mucosa has a unique immune system, known as the gut-associated lymphoid tissue (GALT), the balance between immune tolerance and anti-tumor immune responses can be disrupted, leading to alterations in the composition and function of immune cells within the tumor stroma;
3) Wnt signaling pathway: Aberrant activation of the Wnt signaling pathway is a hallmark of colorectal cancer, particularly in the case of tumors with mutations in the adenomatous polyposis coli (APC) gene. This pathway can also influence the tumor stroma by promoting the activation of CAFs and the secretion of factors that support cancer cell growth and invasion;
4) Tumor budding: Tumor budding, characterized by the presence of small clusters or single cancer cells at the invasive front of the tumor, is a distinctive feature of colorectal cancer and has been associated with a more aggressive phenotype and worse prognosis. These budding cancer cells can interact with the stromal cells, such as CAFs and immune cells, to promote invasion and metastasis;
5) Conserved Oncogenic Signatures (COS): Colorectal cancer stroma displays unique gene expression signatures known as Conserved Oncogenic Signatures, which are specific to the stromal compartment of colorectal tumors.
This review will focus on the specific features of the colorectal tumor stroma, discussing the cellular and molecular mechanisms that govern its role in CRC progression and therapy resistance. A better understanding of these mechanisms may pave the way for the development of novel diagnostic tools and targeted therapeutic strategies, ultimately improving the prognosis for CRC patients.
2. The Gut Microbiome and Colorectal Cancer: Dysbiosis, Tumor Stroma Modulation, and Emerging Therapeutic Strategies
The gut microbiome, composed of trillions of commensal microorganisms, plays a vital role in maintaining homeostasis and overall health. Mounting evidence suggests that alterations in the gut microbiome, referred to as dysbiosis, may contribute to the initiation and progression of CRC by modulating the tumor microenvironment (TME), including the tumor stroma [4].
One crucial aspect of this modulation is the direct interaction of specific bacterial species with the tumor stroma. For instance, Fusobacterium nucleatum, Bacteroides fragilis, and Escherichia coli have been closely associated with CRC development, while some species exhibit antitumor activity (Table 1) [5,6,7,8,9]. These bacteria directly interact with stromal cells, including CAFs and immune cells, influencing their activation and function. F. nucleatum, for example, adheres to and invades CAFs, leading to the production of pro-inflammatory cytokines like IL-6 and IL-8, which in turn promote cancer cell proliferation, survival, and migration [10,11,12].
Once in the nucleus, β-catenin interacts with T-cell factor/lymphoid enhancer factor (TCF/LEF) transcription factors, activating the transcription of target genes such as Transforming growth factor-beta (TGF-β), Connective tissue growth factor (CTGF), and Matrix metalloproteinases (MMPs). These factors influence the tumor microenvironment through the activation of cancer-associated fibroblasts (CAFs), promotion of extracellular matrix (ECM) remodeling, enhancement of cell adhesion, migration, and proliferation, and facilitation of tumor invasion and metastasis.
Another aspect of gut microbiome influence on the CRC stroma is through the production of bacterial metabolites, such as short-chain fatty acids (SCFAs), secondary bile acids, and polyamines [24,25,26,27,28]. SCFAs, like butyrate, exhibit anti-inflammatory and anti-tumorigenic properties by modulating the activation of immune cells and CAFs [24,25,26]. In contrast, secondary bile acids and polyamines promote a pro-inflammatory environment and stimulate reactive oxygen species (ROS) production, leading to DNA damage and activation of oncogenic pathways in both cancer and stromal cells [27,28,29,30,31].
Dysbiosis can also result in chronic inflammation, a major risk factor for CRC. Pro-inflammatory bacteria stimulate the production of cytokines and chemokines, such as IL-6, IL-8, IL-1β, and TNF-α, which recruit and activate various immune cells, including tumor-associated macrophages (TAMs), T cells, and myeloid-derived suppressor cells (MDSCs) [11,32,33,34]. The complex interplay among bacteria, immune cells, and the tumor stroma creates a self-perpetuating pro-inflammatory and pro-tumorigenic environment, facilitating CRC development and progression.
The gut microbiome can also impact the composition and remodeling of the extracellular matrix (ECM) in the CRC stroma. Bacteria and their metabolites modulate the expression and activity of matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs), which are crucial for ECM remodeling [35,36]. Changes in ECM composition and stiffness influence cancer cell invasion, metastasis, angiogenesis, and immune cell infiltration.
Given the role of the gut microbiome in colorectal tumor stroma development and progression, researchers are actively exploring strategies to manipulate it. Promising approaches include probiotics and prebiotics, fecal microbiota transplantation (FMT), dietary interventions, targeted antimicrobial therapy, and combination therapies.
Probiotics, live microorganisms conferring health benefits when administered in adequate amounts, may help restore immune homeostasis and reduce pro-inflammatory and pro-tumorigenic stimuli contributing to CRC progression [37,38,39]. Prebiotics, non-digestible food components that selectively stimulate the growth and activity of beneficial gut bacteria, promote the production of beneficial bacterial metabolites, such as SCFAs, potentially counteracting dysbiosis' adverse effects on colorectal tumor stroma development [40,41].
FMT, involving the transfer of fecal material containing a healthy donor's gut microbiota into a recipient's gastrointestinal tract, aims to restore the recipient's gut microbial balance. While FMT has been primarily used for treating recurrent Clostridioides difficile infection, emerging evidence suggests potential for modulating the gut microbiome in colorectal cancer patients, thereby affecting tumor stroma development and disease progression [42,43,44].
Dietary interventions offer another means to influence gut microbial composition and function. Adopting a diet rich in fruits, vegetables, whole grains, and lean proteins, while limiting the intake of processed and red meats, high-fat dairy products, and added sugars, can promote a healthy gut microbiome [45,46,47]. Such dietary changes may potentially reduce inflammation and the risk of colorectal cancer by modulating the gut microbiome and its interactions with the tumor stroma.
Targeted antimicrobial therapy, selectively targeting specific pathogenic bacteria implicated in colorectal cancer progression such as Fusobacterium nucleatum and Bacteroides fragilis, could be a potential approach to mitigate their influence on the tumor stroma [48,49,50]. However, developing targeted antimicrobial therapies requires a thorough understanding of the complex interactions between these bacteria and the colorectal tumor stroma, as well as the identification of specific molecular targets.
Combination therapies, which involve combining microbiome-targeting interventions with conventional cancer therapies such as chemotherapy, radiotherapy, or immunotherapy, may enhance treatment efficacy by modulating the tumor stroma and improving the overall tumor microenvironment [51]. These combination strategies could help overcome therapy resistance and improve patient outcomes.
In summary, the gut microbiome plays a crucial role in colorectal cancer development and progression by modulating the tumor stroma through direct bacterial interactions, production of bacterial metabolites, and bacteria-induced inflammation. Dysbiosis can lead to a pro-inflammatory and pro-tumorigenic environment, further promoting colorectal cancer. Strategies such as probiotics, prebiotics, fecal microbiota transplantation, dietary interventions, targeted antimicrobial therapy, and combination therapies hold promise for mitigating the gut microbiome's influence on colorectal tumor stroma development and progression. Further research is needed to optimize these approaches and improve patient outcomes.
3. Gut-Associated Lymphoid Tissue (GALT) and its Influence on Colorectal Tumor Stroma Development and Stability
The gut-associated lymphoid tissue (GALT) is a critical component of the mucosal immune system, responsible for maintaining intestinal homeostasis and protecting the host from pathogens. However, under certain conditions, GALT can contribute to the development and stability of the colorectal tumor stroma, ultimately supporting tumor survival.
One way GALT can influence tumor survival is through immune tolerance and immunosuppression. Tumors can exploit GALT's tolerogenic environment to evade immune surveillance. Regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) are essential for maintaining immune tolerance within GALT. Tumor cells can recruit Tregs and MDSCs to the tumor stroma, where they suppress anti-tumor immune responses by inhibiting the function of cytotoxic T cells and natural killer (NK) cells. This immunosuppressive microenvironment enables the tumor to survive and progress [52].
Additionally, GALT plays a central role in regulating inflammatory responses. In the context of colorectal cancer, GALT-driven chronic inflammation can contribute to tumor stroma development and progression. Pro-inflammatory cytokines and chemokines, such as IL-6, IL-8, IL-1β, and TNF-α, can activate stromal cells, including CAFs and tumor-associated macrophages (TAMs). These cells, in turn, support tumor growth, angiogenesis, and metastasis, creating a pro-tumorigenic environment in the colorectal tumor stroma [53,54].
Furthermore, antigen presentation and immune cell infiltration should be considered. GALT contains a high concentration of antigen-presenting cells (APCs), such as dendritic cells and macrophages, which play a pivotal role in shaping the immune response. In the context of colorectal cancer, dysfunctional APCs may inefficiently present tumor antigens, leading to suboptimal activation of cytotoxic T cells and a weakened anti-tumor immune response. Additionally, the complex cellular composition of the tumor stroma, including immune cells like TAMs, MDSCs, and Tregs, can impede the infiltration and function of effector T cells, further supporting tumor immune escape [54,55,56].
Lastly, lymphangiogenesis is a crucial factor. GALT is rich in lymphatic vessels, which are essential for the transport of immune cells and antigens. Tumor cells can exploit the lymphatic network within GALT to facilitate metastasis to regional lymph nodes and distant organs. Moreover, tumor-induced lymphangiogenesis can modulate the tumor stroma, allowing for increased infiltration of immunosuppressive cells and further supporting tumor survival and progression [52].
Given the fundamental mechanisms of GALT's influence on tumor stroma development, several potential targets within GALT associated with immune regulation and inflammation can be explored as intervention methods. Regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) play a central role in immune suppression within the tumor microenvironment. Inhibiting their recruitment or function could potentially enhance anti-tumor immunity. Current investigations focus on strategies to deplete or block the function of Tregs and MDSCs, including the use of monoclonal antibodies, small molecules, and immune checkpoint inhibitors [57,58]. Immune checkpoint blockade is a promising strategy for enhancing anti-tumor immunity in various cancers, including colorectal cancer. Immune checkpoints are inhibitory pathways that regulate immune responses and maintain self-tolerance. Tumors often exploit these pathways to evade immune surveillance. Blocking immune checkpoints, such as CTLA-4, PD-1, and PD-L1, with monoclonal antibodies has shown promise in multiple cancer types [59].
Another potential target is pro-inflammatory cytokines and chemokines, such as IL-6, IL-8, IL-1β, and TNF-α, which contribute to the promotion of tumor growth, angiogenesis, and metastasis. Targeting these key pro-inflammatory mediators could reduce inflammation within the tumor microenvironment. Inhibitors of these cytokines or their receptors are being explored as potential therapeutic agents for various cancers, including colorectal cancer [60]. In addition to inhibition, enhancing the function of APCs, including dendritic cells and macrophages, could lead to more efficient activation of cytotoxic T cells and a stronger anti-tumor immune response. Immunotherapeutic approaches, such as dendritic cell vaccines or adoptive cell transfer, aim to improve the antigen-presenting capacity of these cells [61].
Targeting lymphangiogenesis could potentially limit the spread of tumor cells to regional lymph nodes and distant organs and is an attractive approach for treatment. Therapeutic agents targeting the vascular endothelial growth factor (VEGF) family members or their receptors, which play a crucial role in lymphangiogenesis, are being explored as potential treatment options [62,63].
In conclusion, understanding the complex role of GALT in colorectal cancer progression offers valuable insights for developing novel therapeutic strategies. By targeting key components of the immune response, inflammation, and lymphangiogenesis within GALT, it may be possible to interfere with the pathological processes that support tumor survival and progression. This approach holds promise for the development of more effective therapies for colorectal cancer and may ultimately improve patient outcomes.
4. The Role of the Wnt Signaling Pathway in Colorectal Cancer Tumor Stroma Development and Maintenance
The Wnt signaling pathway is a crucial signaling cascade involved in various physiological processes such as embryonic development, cell proliferation, differentiation, and tissue homeostasis. Aberrant activation of the Wnt pathway has been implicated in numerous human cancers, including CRC [64]. In CRC, the Wnt signaling pathway plays a central role in driving tumorigenesis and maintaining the tumor microenvironment. It is also essential for tumor stroma development, remodeling, and stability, making it a critical contributor to tumor progression. The Wnt signaling pathway's specificity for CRC provides potential molecular targets for regulating this pathway to halt tumor support.
The Wnt signaling pathway can be broadly divided into two categories: canonical (β-catenin-dependent) and non-canonical (β-catenin-independent). In CRC, the canonical Wnt/β-catenin signaling is the primary pathway involved in tumorigenesis. Under normal physiological conditions, β-catenin levels are tightly regulated through phosphorylation and subsequent degradation. However, in CRC, mutations in genes encoding components of the Wnt pathway, such as APC, CTNNB1 (encoding β-catenin), and AXIN2, lead to stabilization and accumulation of β-catenin [65]. Consequently, β-catenin translocates to the nucleus, where it interacts with the TCF/LEF family of transcription factors, leading to the activation of target genes involved in cell proliferation, survival, and stemness [66].
The specificity of Wnt signaling in CRC can be attributed to the high frequency of mutations in the APC gene, occurring in approximately 80% of sporadic CRC cases [67]. The APC protein, a key component of the β-catenin destruction complex, plays a crucial role in maintaining intestinal epithelial homeostasis. Loss of APC function due to mutations results in aberrant activation of Wnt/β-catenin signaling, leading to uncontrolled cell proliferation and the initiation of CRC. Furthermore, the Wnt signaling pathway is intimately involved in the maintenance and regulation of intestinal stem cells, which are essential for tissue regeneration and repair. Dysregulation of Wnt signaling in CRC disrupts the balance between stem cell proliferation and differentiation, contributing to tumor growth and progression [68].
Wnt signaling contributes to the formation and maintenance of the tumor stroma in CRC through various mechanisms. One such mechanism is the crosstalk between tumor cells and stromal cells, where Wnt signaling can mediate communication between tumor cells and stromal cells like CAFs and tumor-associated macrophages (TAMs). In turn, these stromal cells secrete Wnt ligands like Wnt3a, Wnt5a, and Wnt7b, and other factors that further activate Wnt signaling in tumor cells, creating a positive feedback loop that promotes tumor growth and progression [69,70].
Another mechanism involves the regulation of extracellular matrix (ECM) remodeling. Wnt signaling can modulate the expression of matrix metalloproteinases (MMPs) and other ECM remodeling enzymes, which are involved in the degradation and reorganization of the ECM. This remodeling process creates a permissive environment for tumor cell invasion and metastasis [71].
Wnt signaling also plays a role in angiogenesis, promoting the formation of new blood vessels by regulating the expression of pro-angiogenic factors, such as vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) [72,73]. Enhanced angiogenesis within the tumor stroma supports tumor growth by providing nutrients and oxygen while also facilitating the metastatic spread of tumor cells through the Additionally, Wnt signaling can influence the tumor immune microenvironment by affecting the recruitment and function of immune cells within the tumor stroma. Activation of Wnt signaling can lead to the recruitment of immunosuppressive cells, such as myeloid-derived suppressor cells (MDSCs) and regulatory T cells (Tregs), which inhibit anti-tumor immune responses and promote tumor progression [74,75].
Given the critical role of the Wnt signaling pathway in colorectal cancer and its impact on the tumor stroma, several molecular targets have been proposed for therapeutic intervention. These targets aim to inhibit Wnt signaling and disrupt the tumor-supporting functions of the tumor stroma:
- Porcupine inhibitors: Porcupine is an enzyme required for the palmitoylation and secretion of Wnt ligands. Inhibition of Porcupine prevents the secretion of Wnt ligands, thereby blocking Wnt signaling activation [76]. Several Porcupine inhibitors, such as LGK974 and ETC-159, are currently being evaluated in clinical trials for the treatment of Wnt-driven cancers, including CRC [77,78].
- Frizzled receptor antagonists: Frizzled receptors are cell surface receptors that bind to Wnt ligands and activate Wnt signaling [77]. Targeting Frizzled receptors with antagonistic antibodies or small molecules can prevent Wnt ligand binding and subsequent pathway activation. OMP-18R5 (vantictumab) and OMP-54F28 (ipafricept) are two examples of Frizzled receptor antagonists under clinical investigation [79,80].
- Tankyrase inhibitors: Tankyrases are enzymes that regulate the stability of the scaffold protein AXIN, a component of the β-catenin destruction complex. Tankyrase inhibitors stabilize AXIN, promoting the degradation of β-catenin and thus inhibiting Wnt signaling [81,82]. G007-LK and NVP-TNKS656 are examples of Tankyrase inhibitors in preclinical development [83].
- β-catenin inhibitors: Directly targeting β-catenin can inhibit its interaction with TCF/LEF transcription factors, preventing the activation of Wnt target genes [82]. Small molecules, such as PRI-724 and BC2059, have been developed to target the β-catenin/TCF interaction and are currently in clinical trials [84,85].
In conclusion, the Wnt signaling pathway plays a pivotal role in colorectal cancer by influencing the tumor stroma and promoting tumor growth and progression. The specificity of Wnt signaling in CRC, primarily due to the high frequency of APC mutations, makes it an attractive target for therapeutic intervention. Several molecular targets within the Wnt pathway have been identified, and their modulation holds promise for disrupting the tumor-supporting functions of the tumor stroma in CRC. Continued research and clinical trials are needed to fully understand the potential of these therapeutic strategies in treating colorectal cancer patients.
5. Tumor Budding and Its Influence on the Tumor Stroma in Colorectal Cancer
Tumor budding is a histological feature characterized by small clusters or single cancer cells at the invasive front of the tumor [86]. These budding tumor cells possess features of epithelial-mesenchymal transition (EMT) and display increased migratory and invasive properties. Tumor budding is considered an important and unique prognostic factor in CRC, as it is associated with aggressive tumor behavior, lymph node metastasis, and poor clinical outcomes. The interaction between tumor budding cells and the tumor stroma is crucial for the progression of CRC.
Tumor budding cells exhibit EMT, a process in which epithelial cells lose their polarity and cell-cell adhesion properties and acquire mesenchymal features. This process enhances their migratory and invasive capabilities, ultimately leading to tumor invasion and metastasis. EMT in CRC is driven by various signaling pathways, such as TGF-β, Wnt, and Notch, which are also involved in modulating the tumor stroma [87].
The tumor stroma, composed of CAFs, immune cells, and extracellular matrix (ECM), plays a crucial role in supporting tumor growth, angiogenesis, and metastasis. Tumor budding cells secrete various growth factors, cytokines, and chemokines that influence the stroma and create a tumor-promoting microenvironment, such as TGF-β, VEGF, IL-6, IL-8, Chemokine (C-X-C motif) ligand 12 (CXCL12) and its receptor C-X-C chemokine receptor type 4 (CXCR4), and Matrix Metalloproteinases (MMPs). In turn, the stromal cells produce factors that enhance the invasive and metastatic potential of the budding cells, such as Fibroblast Growth Factors (FGFs), Hepatocyte Growth Factor (HGF), Platelet-Derived Growth Factor (PDGF), Stromal Cell-Derived Factor-1 (SDF-1, also known as CXCL12), Matrix Metalloproteinases (MMPs), Tenascin-C, and Lysyl Oxidase (LOX). This establishes a reciprocal feedback loop [88,89,90,91].
Tumor budding is strongly associated with tumor invasion and metastasis in CRC. Budding cells invade the stroma and degrade the ECM through the secretion of MMPs. They can also enter the lymphatic or blood vessels, leading to lymph node and distant organ metastasis. The stromal cells, particularly CAFs, support this process by secreting factors that promote invasion and angiogenesis [88,92].
Targeting tumor budding in CRC is a promising therapeutic strategy, as it may disrupt the tumor-stroma crosstalk, inhibit invasion and metastasis, and improve clinical outcomes. Several cellular and molecular targets are being investigated for their potential to stop tumor budding and its supportive role in cancer progression:
- EMT Inhibitors: Targeting the EMT process can inhibit tumor budding and potentially prevent invasion and metastasis. Inhibition of EMT-driving pathways, such as TGF-β, Wnt, and Notch, can restore epithelial properties and reduce the invasive potential of tumor cells [93,94,95]. Small molecules, monoclonal antibodies, and other therapeutic agents targeting these pathways are under investigation.
- Targeting CAFs: CAFs play a critical role in supporting tumor budding by secreting factors that promote EMT, invasion, and angiogenesis. Inhibiting CAF activation, proliferation, or function may disrupt the tumor-stroma crosstalk and reduce the supportive role of CAFs in tumor budding. Several strategies, including targeting CAF-derived factors, such as fibroblast activation protein (FAP) [96] or transforming growth factor-beta (TGF-β), are being explored [97].
- Inhibition of MMPs: MMPs are enzymes that degrade the ECM, facilitating tumor invasion and metastasis [98]. Budding tumor cells produce MMPs to invade the stroma and promote tumor progression. Inhibiting MMP activity may prevent the degradation of the ECM and reduce tumor budding-associated invasion and metastasis. Several MMP inhibitors have been developed, and some are currently being tested in preclinical and clinical studies for their potential in CRC treatment [99,100].
- Targeting Immune Cells: The immune cells within the tumor stroma, such as tumor-associated macrophages (TAMs) and myeloid-derived suppressor cells (MDSCs), contribute to the supportive tumor microenvironment and facilitate tumor budding. Modulating the function of these immune cells or reprogramming them to adopt an anti-tumor phenotype may help inhibit tumor budding and improve CRC outcomes. Immunotherapies, such as immune checkpoint inhibitors and adoptive cell transfer, are being investigated for their potential to target these immune cells [101,102].
- Targeting Tumor-Stroma Interactions: The reciprocal feedback loop between tumor budding cells and the stroma is crucial for CRC progression. Disrupting the signaling pathways involved in this crosstalk, such as the chemokine ligand-receptor axis (e.g., CXCL12-CXCR4), may inhibit tumor budding and its associated invasion and metastasis. Therapeutic agents targeting these signaling pathways are under investigation [103,104].
- Therapeutic Approaches Targeting Tumor-Initiating Cells: Tumor-initiating cells (TICs) or cancer stem cells are a subpopulation of tumor cells that possess self-renewal and differentiation capabilities, driving tumor heterogeneity and resistance to therapy. Tumor budding has been linked to the presence of TICs, which are capable of initiating new tumor growth at the invasive front. Targeting TICs using specific surface markers, such as CD133, CD44, and Lgr5, or cellular processes like self-renewal and differentiation, can help inhibit TICs and tumor budding [105,106,107]. Targeting metabolic reprogramming in TICs, such as glucose metabolism, glutamine metabolism, or fatty acid synthesis, may selectively affect TICs and impair their survival and function [108,109].
In conclusion, tumor budding is an essential and unique feature of colorectal cancer that significantly influences the tumor stroma and contributes to aggressive tumor behavior, invasion, and metastasis. Several potential cellular and molecular targets are being investigated to stop tumor budding and its supportive role in CRC progression. Targeting tumor budding may disrupt the tumor-stroma crosstalk, inhibit invasion and metastasis, and ultimately improve the clinical outcomes of CRC patients. Further research is required to identify and validate effective therapeutic strategies that specifically target tumor budding and its underlying molecular mechanisms in colorectal cancer.
6. Conserved Oncogenic Signatures in Colorectal Cancer Stroma
Colorectal cancer stroma displays unique gene expression signatures known as Conserved Oncogenic Signatures (COS), which are specific to the stromal compartment of colorectal tumors. These signatures represent consistent and reproducible patterns of gene expression that have been identified across various colorectal cancer patients and studies [110,111,112]. The presence of COS suggests that there are specific interactions between colorectal cancer cells and their stromal components, which may drive tumor progression and influence treatment outcomes. Key players in this signature include Fibronectin (FN1), Matrix metalloproteinases (MMPs), Vascular endothelial growth factor (VEGF), Transforming growth factor-beta (TGF-β), and Interleukins (e.g., IL-6, IL-8) [113,114,115,116].
Although COS represent consistent and reproducible patterns of gene expression, there may still be some degree of inter-patient variability due to factors such as genetic background, lifestyle, and tumor stage. These inter-patient differences in COS may influence treatment response and outcomes, highlighting the importance of personalized medicine approaches. By analyzing the unique molecular characteristics of each patient's tumor, including their specific COS, clinicians can develop tailored treatment plans that maximize therapeutic efficacy and minimize side effects.
For example, some of the genes identified in the COS are involved in extracellular matrix remodeling, which is known to play a critical role in facilitating tumor invasion and metastasis [117]. These genes include matrix metalloproteinases, which degrade various components of the extracellular matrix; lysyl oxidase, which contributes to collagen cross-linking and stabilization; fibronectin, a major component of the extracellular matrix that mediates cell adhesion and migration; and collagens, the primary structural proteins in the extracellular matrix (Table 2). By targeting the specific genes or pathways involved in this process, it may be possible to inhibit tumor progression and improve treatment outcomes.
Moreover, the study of COS in colorectal cancer stroma has revealed the potential role of the immune system in colorectal cancer development and progression. Some of the gene expression signatures identified are associated with immune cell infiltration, immune cell activation, and immune checkpoint pathways, indicating that the immune system may play a significant role in shaping the tumor microenvironment (Table 2). Targeting these immune-related genes or pathways may help modulate the immune response and enhance the effectiveness of immunotherapies in colorectal cancer patients.
Another important aspect of COS in colorectal cancer stroma is the identification of genes and pathways involved in angiogenesis. By identifying specific angiogenesis-related genes in the COS, researchers can better understand the mechanisms driving this process and develop targeted therapies to inhibit angiogenesis in colorectal cancer patients (Table 2).
In conclusion, COS in colorectal cancer stroma is a distinct concept from the previous chapters on the Gut Microbiome, the Gut-Associated Lymphoid Tissue (GALT), the Wnt Signaling, and the Tumor Budding. While the previous chapters focus on specific components or processes within the tumor microenvironment, COS represents a unique gene expression signature specific to the stromal compartment of colorectal tumors. Although there may be some overlap or interplay among these topics, COS is an individual mechanism that could potentially serve as a therapeutic target.
7. Culmination of Colorectal Cancer Tumor-Stroma Interactions in Metastasis: The Seed and Soil Hypothesis
The development of metastatic CRC significantly impacts patient prognosis and treatment outcomes, as metastatic disease is often more challenging to treat and associated with a poorer prognosis. A deeper understanding of the mechanisms underlying metastasis in CRC is essential for developing novel therapeutic strategies that can improve patient survival and quality of life. In this context, the concept of "the seed" and "the soil" plays a crucial role in understanding the intricate interplay between cancer cells and their supportive stroma during the metastatic process.
The "seed" represents the colon cancer cell, which undergoes a series of genetic and epigenetic alterations during carcinogenesis. As the cancer cell accumulates mutations, it acquires immunogenic properties that enable it to evade the host's immune system [150]. Simultaneously, the tumor stroma, or "the soil," undergoes changes that promote immunosuppression and create a nurturing environment for the growing cancer cell [151,152]. The dynamic relationship between the cancer cell and its stroma is critical for primary tumor progression and the eventual development of metastasis.
In the metastatic process, it is unlikely that a single, isolated cancer cell from the primary tumor can form a metastatic lesion in a distant organ. Intact organs represent a "desert" that lacks the supportive stromal environment necessary for a highly immunogenic cancer cell to survive and thrive. Instead, the hypothesis suggests that metastasis may arise from conglomerates of cells, consisting of cancer cells and associated stromal elements, that together create a suitable microenvironment for the survival and growth of metastatic cancer cells.
One potential example of such a metastatic unit is the presence of Cancer-Associated Macrophage-like Cells (CAMLs) in circulation [153]. CAMLs are a unique cell population that consists of both cancer cells and stromal macrophages. These cells have been detected in the bloodstream of cancer patients and have been suggested to play a role in metastasis [154,155]. In this scenario, the cancer cell and the macrophage together form a primary metastatic unit that is capable of colonizing distant organs and establishing metastatic lesions. The presence of the stromal macrophage in this unit may provide the necessary support and immunosuppressive environment for the highly immunogenic cancer cell to survive and grow in the "desert" of an intact organ (Figure 2).
This hypothesis of metastasis involving cancer cell-stromal cell conglomerates underscores the importance of considering both cancer cells and their surrounding stroma in the development of therapeutic strategies for metastatic CRC. By targeting both the "seed" and the "soil," researchers can potentially disrupt the metastatic process and improve patient outcomes.
8. Conclusions
In conclusion, the primary colorectal tumor stroma plays a vital role in the development of metastasis. As cancer cells acquire immunogenic properties, the tumor stroma adapts to create an immunosuppressive and trophic environment that supports cancer cell growth. The metastatic process may involve cancer cell-stromal cell conglomerates that together establish metastatic lesions in distant organs. Understanding this complex interplay between cancer cells and their stroma is crucial for developing novel therapeutic strategies that can effectively target both the "seed" and the "soil" to prevent or treat metastatic colorectal cancer.
Author Contributions
N.S., L.M., A.B. and Z.G. – conceptualization, investigation, visualization, writing—original draft preparation, writing—review and editing, and supervision; N.S., A.M., D.A., I.B., M.T. and A.O. – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Russian Science Foundation, grant No 23-15-00140.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data and materials are available upon reasonable request. Address to I.B. (email: buchwalow@pathologie-hh.de) or M.T. (email: mtiemann@hp-hamburg.de) Institute for Hematopathology, Hamburg, Germany.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
APC, Adenomatous Polyposis Coli; APCs, Antigen-presenting Cells; CAMLs, Cancer-Associated Macrophage-like Cells; CAFs, Cancer-Associated Fibroblasts; CD133, Cluster of Differentiation 133; CD44, Cluster of Differentiation 44; COS, Conserved Oncogenic Signatures; CRC, Colorectal Cancer; CTGF, Connective tissue growth factor; CTLA-4, Cytotoxic T-lymphocyte-associated protein 4; CXCL12, Chemokine (C-X-C motif) ligand 12; CXCR4, C-X-C chemokine receptor type 4; EMT, Epithelial-Mesenchymal Transition; ECM, Extracellular Matrix; FAP, Fibroblast Activation Protein; FGF, Fibroblast Growth Factor; FIT, Fecal Immunochemical Tests; FOBT, Fecal Occult Blood Tests; GALT, Gut-Associated Lymphoid Tissue; HGF, Hepatocyte Growth Factor; IL, Interleukin; LOX, Lysyl Oxidase; Lgr5, Leucine-rich repeat-containing G-protein coupled receptor 5; MMPs, Matrix Metalloproteinases; MDSCs, Myeloid-Derived Suppressor Cells; Notch, Signaling pathway; PD-1, Programmed cell death protein 1; PDGF, Platelet-Derived Growth Factor; PD-L1, Programmed death-ligand 1; SDF-1, Stromal Cell-Derived Factor-1; TAMs, Tumor-Associated Macrophages; TGF-β, Transforming Growth Factor-beta; TICs, Tumor-Initiating Cells; TIMPs: Tissue Inhibitors of Metalloproteinases; TNF-α, Tumor Necrosis Factor Alpha; TME, Tumor Microenvironment; Tregs, Regulatory T cells; VEGF, Vascular Endothelial Growth Factor; WHO, World Health Organization.
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int J Cancer 2021. https://doi.org/10.1002/ijc.33588. [CrossRef]
- Ladabaum, U.; Dominitz, J.A.; Kahi, C.; Schoen, R.E. Strategies for Colorectal Cancer Screening. Gastroenterology 2020, 158, 418-432. https://doi.org/10.1053/j.gastro.2019.06.043. [CrossRef]
- Biller, L.H.; Schrag, D. Diagnosis and Treatment of Metastatic Colorectal Cancer: A Review. JAMA 2021, 325, 669-685. https://doi.org/10.1001/jama.2021.0106. [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322-340. https://doi.org/10.1053/j.gastro.2019.06.048. [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/beta-catenin modulator Annexin A1. EMBO Rep 2019, 20. https://doi.org/10.15252/embr.201847638. [CrossRef]
- Brennan, C.A.; Clay, S.L.; Lavoie, S.L.; Bae, S.; Lang, J.K.; Fonseca-Pereira, D.; Rosinski, K.G.; Ou, N.; Glickman, J.N.; Garrett, W.S. Fusobacterium nucleatum drives a pro-inflammatory intestinal microenvironment through metabolite receptor-dependent modulation of IL-17 expression. Gut Microbes 2021, 13, 1987780. https://doi.org/10.1080/19490976.2021.1987780. [CrossRef]
- Sui, H.; Zhang, L.; Gu, K.; Chai, N.; Ji, Q.; Zhou, L.; Wang, Y.; Ren, J.; Yang, L.; Zhang, B.; et al. YYFZBJS ameliorates colorectal cancer progression in Apc(Min/+) mice by remodeling gut microbiota and inhibiting regulatory T-cell generation. Cell Commun Signal 2020, 18, 113. https://doi.org/10.1186/s12964-020-00596-9. [CrossRef]
- Knippel, R.J.; Drewes, J.L.; Sears, C.L. The Cancer Microbiome: Recent Highlights and Knowledge Gaps. Cancer Discov 2021, 11, 2378-2395. https://doi.org/10.1158/2159-8290.CD-21-0324. [CrossRef]
- Chattopadhyay, I.; Dhar, R.; Pethusamy, K.; Seethy, A.; Srivastava, T.; Sah, R.; Sharma, J.; Karmakar, S. Exploring the Role of Gut Microbiome in Colon Cancer. Appl Biochem Biotechnol 2021, 193, 1780-1799. https://doi.org/10.1007/s12010-021-03498-9. [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443-1448. https://doi.org/10.1126/science.aal5240. [CrossRef]
- Engevik, M.A.; Danhof, H.A.; Ruan, W.; Engevik, A.C.; Chang-Graham, A.L.; Engevik, K.A.; Shi, Z.; Zhao, Y.; Brand, C.K.; Krystofiak, E.S.; et al. Fusobacterium nucleatum Secretes Outer Membrane Vesicles and Promotes Intestinal Inflammation. mBio 2021, 12. https://doi.org/10.1128/mBio.02706-20. [CrossRef]
- Casasanta, M.A.; Yoo, C.C.; Udayasuryan, B.; Sanders, B.E.; Umana, A.; Zhang, Y.; Peng, H.; Duncan, A.J.; Wang, Y.; Li, L.; et al. Fusobacterium nucleatum host-cell binding and invasion induces IL-8 and CXCL1 secretion that drives colorectal cancer cell migration. Sci Signal 2020, 13. https://doi.org/10.1126/scisignal.aba9157. [CrossRef]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11. https://doi.org/10.3390/nu11071613. [CrossRef]
- Cheng, W.T.; Kantilal, H.K.; Davamani, F. The Mechanism of Bacteroides fragilis Toxin Contributes to Colon Cancer Formation. Malays J Med Sci 2020, 27, 9-21. https://doi.org/10.21315/mjms2020.27.4.2. [CrossRef]
- Lee, C.W.; Chen, H.J.; Chien, Y.H.; Hsia, S.M.; Chen, J.H.; Shih, C.K. Synbiotic Combination of Djulis (Chenopodium formosanum) and Lactobacillus acidophilus Inhibits Colon Carcinogenesis in Rats. Nutrients 2019, 12. https://doi.org/10.3390/nu12010103. [CrossRef]
- Faghfoori, Z.; Pourghassem Gargari, B.; Saber, A.; Seyyedi, M.; Fazelian, S.; Khosroushahi, A.Y. Prophylactic effects of secretion metabolites of dairy lactobacilli through downregulation of ErbB-2 and ErbB-3 genes on colon cancer cells. Eur J Cancer Prev 2020, 29, 201-209. https://doi.org/10.1097/CEJ.0000000000000393. [CrossRef]
- Williamson, A.J.; Jacobson, R.; van Praagh, J.B.; Gaines, S.; Koo, H.Y.; Lee, B.; Chan, W.C.; Weichselbaum, R.; Alverdy, J.C.; Zaborina, O.; et al. Enterococcus faecalis promotes a migratory and invasive phenotype in colon cancer cells. Neoplasia 2022, 27, 100787. https://doi.org/10.1016/j.neo.2022.100787. [CrossRef]
- Banna, G.L.; Torino, F.; Marletta, F.; Santagati, M.; Salemi, R.; Cannarozzo, E.; Falzone, L.; Ferrau, F.; Libra, M. Lactobacillus rhamnosus GG: An Overview to Explore the Rationale of Its Use in Cancer. Front Pharmacol 2017, 8, 603. https://doi.org/10.3389/fphar.2017.00603. [CrossRef]
- Nouri, R.; Hasani, A.; Shirazi, K.M.; Alivand, M.R.; Sepehri, B.; Sotoodeh, S.; Hemmati, F.; Rezaee, M.A. Escherichia coli and Colorectal Cancer: Unfolding the Enigmatic Relationship. Curr Pharm Biotechnol 2022, 23, 1257-1268. https://doi.org/10.2174/1389201022666210910094827. [CrossRef]
- Dikeocha, I.J.; Al-Kabsi, A.M.; Chiu, H.T.; Alshawsh, M.A. Faecalibacterium prausnitzii Ameliorates Colorectal Tumorigenesis and Suppresses Proliferation of HCT116 Colorectal Cancer Cells. Biomedicines 2022, 10. https://doi.org/10.3390/biomedicines10051128. [CrossRef]
- Long, X.; Wong, C.C.; Tong, L.; Chu, E.S.H.; Ho Szeto, C.; Go, M.Y.Y.; Coker, O.O.; Chan, A.W.H.; Chan, F.K.L.; Sung, J.J.Y.; et al. Peptostreptococcus anaerobius promotes colorectal carcinogenesis and modulates tumour immunity. Nat Microbiol 2019, 4, 2319-2330. https://doi.org/10.1038/s41564-019-0541-3. [CrossRef]
- Pasquereau-Kotula, E.; Martins, M.; Aymeric, L.; Dramsi, S. Significance of Streptococcus gallolyticus subsp. gallolyticus Association With Colorectal Cancer. Front Microbiol 2018, 9, 614. https://doi.org/10.3389/fmicb.2018.00614. [CrossRef]
- Slezak, M.; Smolar, M.; Drobna Saniova, B.; Hosala, M.; Miklusica, J. Clostridium septicum foot gangrene associated with colorectal cancer. Neuro Endocrinol Lett 2022, 43, 57-64.
- Ebert, M.N.; Klinder, A.; Peters, W.H.; Schaferhenrich, A.; Sendt, W.; Scheele, J.; Pool-Zobel, B.L. Expression of glutathione S-transferases (GSTs) in human colon cells and inducibility of GSTM2 by butyrate. Carcinogenesis 2003, 24, 1637-1644. https://doi.org/10.1093/carcin/bgg122. [CrossRef]
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O.; Eid, J. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes 2021, 13, 1-28. https://doi.org/10.1080/19490976.2021.1907272. [CrossRef]
- Park, C.H.; Eun, C.S.; Han, D.S. Intestinal microbiota, chronic inflammation, and colorectal cancer. Intest Res 2018, 16, 338-345. https://doi.org/10.5217/ir.2018.16.3.338. [CrossRef]
- Ryu, T.Y.; Kim, K.; Han, T.S.; Lee, M.O.; Lee, J.; Choi, J.; Jung, K.B.; Jeong, E.J.; An, D.M.; Jung, C.R.; et al. Human gut-microbiome-derived propionate coordinates proteasomal degradation via HECTD2 upregulation to target EHMT2 in colorectal cancer. ISME J 2022, 16, 1205-1221. https://doi.org/10.1038/s41396-021-01119-1. [CrossRef]
- Liu, Y.; Zhang, S.; Zhou, W.; Hu, D.; Xu, H.; Ji, G. Secondary Bile Acids and Tumorigenesis in Colorectal Cancer. Front Oncol 2022, 12, 813745. https://doi.org/10.3389/fonc.2022.813745. [CrossRef]
- Fang, Y.; Yan, C.; Zhao, Q.; Xu, J.; Liu, Z.; Gao, J.; Zhu, H.; Dai, Z.; Wang, D.; Tang, D. The roles of microbial products in the development of colorectal cancer: a review. Bioengineered 2021, 12, 720-735. https://doi.org/10.1080/21655979.2021.1889109. [CrossRef]
- Caliceti, C.; Punzo, A.; Silla, A.; Simoni, P.; Roda, G.; Hrelia, S. New Insights into Bile Acids Related Signaling Pathways in the Onset of Colorectal Cancer. Nutrients 2022, 14. https://doi.org/10.3390/nu14142964. [CrossRef]
- Huang, C.Y.; Fang, Y.J.; Abulimiti, A.; Yang, X.; Li, L.; Liu, K.Y.; Zhang, X.; Feng, X.L.; Chen, Y.M.; Zhang, C.X. Dietary Polyamines Intake and Risk of Colorectal Cancer: A Case-Control Study. Nutrients 2020, 12. https://doi.org/10.3390/nu12113575. [CrossRef]
- Bolte, L.A.; Vich Vila, A.; Imhann, F.; Collij, V.; Gacesa, R.; Peters, V.; Wijmenga, C.; Kurilshikov, A.; Campmans-Kuijpers, M.J.E.; Fu, J.; et al. Long-term dietary patterns are associated with pro-inflammatory and anti-inflammatory features of the gut microbiome. Gut 2021, 70, 1287-1298. https://doi.org/10.1136/gutjnl-2020-322670. [CrossRef]
- Yang, Y.; Li, L.; Xu, C.; Wang, Y.; Wang, Z.; Chen, M.; Jiang, Z.; Pan, J.; Yang, C.; Li, X.; et al. Cross-talk between the gut microbiota and monocyte-like macrophages mediates an inflammatory response to promote colitis-associated tumourigenesis. Gut 2020, 70, 1495-1506. https://doi.org/10.1136/gutjnl-2020-320777. [CrossRef]
- Stephens, M.; von der Weid, P.Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes 2020, 11, 421-432. https://doi.org/10.1080/19490976.2019.1629235. [CrossRef]
- Rodrigues, D.M.; Sousa, A.J.; Hawley, S.P.; Vong, L.; Gareau, M.G.; Kumar, S.A.; Johnson-Henry, K.C.; Sherman, P.M. Matrix metalloproteinase 9 contributes to gut microbe homeostasis in a model of infectious colitis. BMC Microbiol 2012, 12, 105. https://doi.org/10.1186/1471-2180-12-105. [CrossRef]
- Wei, Z.; Cao, S.; Liu, S.; Yao, Z.; Sun, T.; Li, Y.; Li, J.; Zhang, D.; Zhou, Y. Could gut microbiota serve as prognostic biomarker associated with colorectal cancer patients' survival? A pilot study on relevant mechanism. Oncotarget 2016, 7, 46158-46172. https://doi.org/10.18632/oncotarget.10064. [CrossRef]
- Bell, H.N.; Rebernick, R.J.; Goyert, J.; Singhal, R.; Kuljanin, M.; Kerk, S.A.; Huang, W.; Das, N.K.; Andren, A.; Solanki, S.; et al. Reuterin in the healthy gut microbiome suppresses colorectal cancer growth through altering redox balance. Cancer Cell 2022, 40, 185-200 e186. https://doi.org/10.1016/j.ccell.2021.12.001. [CrossRef]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. Biomed Res Int 2018, 2018, 9478630. https://doi.org/10.1155/2018/9478630. [CrossRef]
- Legesse Bedada, T.; Feto, T.K.; Awoke, K.S.; Garedew, A.D.; Yifat, F.T.; Birri, D.J. Probiotics for cancer alternative prevention and treatment. Biomed Pharmacother 2020, 129, 110409. https://doi.org/10.1016/j.biopha.2020.110409. [CrossRef]
- Ciecierska, A.; Drywien, M.E.; Hamulka, J.; Sadkowski, T. Nutraceutical functions of beta-glucans in human nutrition. Rocz Panstw Zakl Hig 2019, 70, 315-324. https://doi.org/10.32394/rpzh.2019.0082. [CrossRef]
- Wang, H.; Chen, K.; Ning, M.; Wang, X.; Wang, Z.; Yue, Y.; Yuan, Y.; Yue, T. Intake of Pro- and/or Prebiotics as a Promising Approach for Prevention and Treatment of Colorectal Cancer. Mol Nutr Food Res 2023, 67, e2200474. https://doi.org/10.1002/mnfr.202200474. [CrossRef]
- Zhang, J.; Wu, K.; Shi, C.; Li, G. Cancer Immunotherapy: Fecal Microbiota Transplantation Brings Light. Curr Treat Options Oncol 2022, 23, 1777-1792. https://doi.org/10.1007/s11864-022-01027-2. [CrossRef]
- Xu, H.; Cao, C.; Ren, Y.; Weng, S.; Liu, L.; Guo, C.; Wang, L.; Han, X.; Ren, J.; Liu, Z. Antitumor effects of fecal microbiota transplantation: Implications for microbiome modulation in cancer treatment. Front Immunol 2022, 13, 949490. https://doi.org/10.3389/fimmu.2022.949490. [CrossRef]
- Ting, N.L.; Lau, H.C.; Yu, J. Cancer pharmacomicrobiomics: targeting microbiota to optimise cancer therapy outcomes. Gut 2022, 71, 1412-1425. https://doi.org/10.1136/gutjnl-2021-326264. [CrossRef]
- Zhao, Y.; Zhan, J.; Wang, Y.; Wang, D. The Relationship Between Plant-Based Diet and Risk of Digestive System Cancers: A Meta-Analysis Based on 3,059,009 Subjects. Front Public Health 2022, 10, 892153. https://doi.org/10.3389/fpubh.2022.892153. [CrossRef]
- Kato, I.; Sun, J. Microbiome and Diet in Colon Cancer Development and Treatment. Cancer J 2023, 29, 89-97. https://doi.org/10.1097/PPO.0000000000000649. [CrossRef]
- Gomes, S.; Teixeira-Guedes, C.; Silva, E.; Baltazar, F.; Preto, A. Colon microbiota modulation by dairy-derived diet: new strategy for prevention and treatment of colorectal cancer. Food Funct 2022, 13, 9183-9194. https://doi.org/10.1039/d2fo01720b. [CrossRef]
- Jia, F.; Yu, Q.; Wang, R.; Zhao, L.; Yuan, F.; Guo, H.; Shen, Y.; He, F. Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression. Int J Mol Sci 2023, 24. https://doi.org/10.3390/ijms24021469. [CrossRef]
- Jang, H.I.; Rhee, K.J.; Eom, Y.B. Antibacterial and antibiofilm effects of alpha-humulene against Bacteroides fragilis. Can J Microbiol 2020, 66, 389-399. https://doi.org/10.1139/cjm-2020-0004. [CrossRef]
- Li, S.; Liu, J.; Zheng, X.; Ren, L.; Yang, Y.; Li, W.; Fu, W.; Wang, J.; Du, G. Tumorigenic bacteria in colorectal cancer: mechanisms and treatments. Cancer Biol Med 2021, 19, 147-162. https://doi.org/10.20892/j.issn.2095-3941.2020.0651. [CrossRef]
- Alam, Z.; Shang, X.; Effat, K.; Kanwal, F.; He, X.; Li, Y.; Xu, C.; Niu, W.; War, A.R.; Zhang, Y. The potential role of prebiotics, probiotics, and synbiotics in adjuvant cancer therapy especially colorectal cancer. J Food Biochem 2022, 46, e14302. https://doi.org/10.1111/jfbc.14302. [CrossRef]
- Yaseen, A.; Ladenheim, A.; Olson, K.A.; Libertini, S.J.; McPherson, J.D.; Matsukuma, K. Whole exome sequencing of a gut-associated lymphoid tissue neoplasm points to precursor or early form of sporadic colon carcinoma. Pathol Res Pract 2021, 220, 153406. https://doi.org/10.1016/j.prp.2021.153406. [CrossRef]
- Muzes, G.; Bohusne Barta, B.; Sipos, F. Colitis and Colorectal Carcinogenesis: The Focus on Isolated Lymphoid Follicles. Biomedicines 2022, 10. https://doi.org/10.3390/biomedicines10020226. [CrossRef]
- Morbe, U.M.; Jorgensen, P.B.; Fenton, T.M.; von Burg, N.; Riis, L.B.; Spencer, J.; Agace, W.W. Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunol 2021, 14, 793-802. https://doi.org/10.1038/s41385-021-00389-4. [CrossRef]
- Barone, F.; Patel, P.; Sanderson, J.D.; Spencer, J. Gut-associated lymphoid tissue contains the molecular machinery to support T-cell-dependent and T-cell-independent class switch recombination. Mucosal Immunol 2009, 2, 495-503. https://doi.org/10.1038/mi.2009.106. [CrossRef]
- Cosola, C.; Rocchetti, M.T.; Gesualdo, L. Gut Microbiota, the Immune System, and Cytotoxic T Lymphocytes. Methods Mol Biol 2021, 2325, 229-241. https://doi.org/10.1007/978-1-0716-1507-2_16. [CrossRef]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci 2019, 110, 2080-2089. https://doi.org/10.1111/cas.14069. [CrossRef]
- Law, A.M.K.; Valdes-Mora, F.; Gallego-Ortega, D. Myeloid-Derived Suppressor Cells as a Therapeutic Target for Cancer. Cells 2020, 9. https://doi.org/10.3390/cells9030561. [CrossRef]
- Pandey, P.; Khan, F.; Qari, H.A.; Upadhyay, T.K.; Alkhateeb, A.F.; Oves, M. Revolutionization in Cancer Therapeutics via Targeting Major Immune Checkpoints PD-1, PD-L1 and CTLA-4. Pharmaceuticals (Basel) 2022, 15. https://doi.org/10.3390/ph15030335. [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal Transduct Target Ther 2021, 6, 263. https://doi.org/10.1038/s41392-021-00658-5. [CrossRef]
- Gardner, A.; de Mingo Pulido, A.; Ruffell, B. Dendritic Cells and Their Role in Immunotherapy. Front Immunol 2020, 11, 924. https://doi.org/10.3389/fimmu.2020.00924. [CrossRef]
- Korhonen, E.A.; Murtomaki, A.; Jha, S.K.; Anisimov, A.; Pink, A.; Zhang, Y.; Stritt, S.; Liaqat, I.; Stanczuk, L.; Alderfer, L.; et al. Lymphangiogenesis requires Ang2/Tie/PI3K signaling for VEGFR3 cell-surface expression. J Clin Invest 2022, 132. https://doi.org/10.1172/JCI155478. [CrossRef]
- Roy, S.; Banerjee, P.; Ekser, B.; Bayless, K.; Zawieja, D.; Alpini, G.; Glaser, S.S.; Chakraborty, S. Targeting Lymphangiogenesis and Lymph Node Metastasis in Liver Cancer. Am J Pathol 2021, 191, 2052-2063. https://doi.org/10.1016/j.ajpath.2021.08.011. [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/beta-catenin signaling pathway in cancer. J Hematol Oncol 2020, 13, 165. https://doi.org/10.1186/s13045-020-00990-3. [CrossRef]
- Malki, A.; ElRuz, R.A.; Gupta, I.; Allouch, A.; Vranic, S.; Al Moustafa, A.E. Molecular Mechanisms of Colon Cancer Progression and Metastasis: Recent Insights and Advancements. Int J Mol Sci 2020, 22. https://doi.org/10.3390/ijms22010130. [CrossRef]
- Boonekamp, K.E.; Heo, I.; Artegiani, B.; Asra, P.; van Son, G.; de Ligt, J.; Clevers, H. Identification of novel human Wnt target genes using adult endodermal tissue-derived organoids. Dev Biol 2021, 474, 37-47. https://doi.org/10.1016/j.ydbio.2021.01.009. [CrossRef]
- Aghabozorgi, A.S.; Bahreyni, A.; Soleimani, A.; Bahrami, A.; Khazaei, M.; Ferns, G.A.; Avan, A.; Hassanian, S.M. Role of adenomatous polyposis coli (APC) gene mutations in the pathogenesis of colorectal cancer; current status and perspectives. Biochimie 2019, 157, 64-71. https://doi.org/10.1016/j.biochi.2018.11.003. [CrossRef]
- Steinhart, Z.; Angers, S. Wnt signaling in development and tissue homeostasis. Development 2018, 145. https://doi.org/10.1242/dev.146589. [CrossRef]
- Lecarpentier, Y.; Schussler, O.; Hebert, J.L.; Vallee, A. Multiple Targets of the Canonical WNT/beta-Catenin Signaling in Cancers. Front Oncol 2019, 9, 1248. https://doi.org/10.3389/fonc.2019.01248. [CrossRef]
- Yoshida, G.J. Regulation of heterogeneous cancer-associated fibroblasts: the molecular pathology of activated signaling pathways. J Exp Clin Cancer Res 2020, 39, 112. https://doi.org/10.1186/s13046-020-01611-0. [CrossRef]
- Wang, J.; Cai, H.; Liu, Q.; Xia, Y.; Xing, L.; Zuo, Q.; Zhang, Y.; Chen, C.; Xu, K.; Yin, P.; et al. Cinobufacini Inhibits Colon Cancer Invasion and Metastasis via Suppressing Wnt/beta-Catenin Signaling Pathway and EMT. Am J Chin Med 2020, 48, 703-718. https://doi.org/10.1142/S0192415X20500354. [CrossRef]
- Kasprzak, A. Angiogenesis-Related Functions of Wnt Signaling in Colorectal Carcinogenesis. Cancers (Basel) 2020, 12. https://doi.org/10.3390/cancers12123601. [CrossRef]
- Yang, P.; Yu, D.; Zhou, J.; Zhuang, S.; Jiang, T. TGM2 interference regulates the angiogenesis and apoptosis of colorectal cancer via Wnt/beta-catenin pathway. Cell Cycle 2019, 18, 1122-1134. https://doi.org/10.1080/15384101.2019.1609831. [CrossRef]
- Zhang, J.; Fan, J.; Zeng, X.; Nie, M.; Luan, J.; Wang, Y.; Ju, D.; Yin, K. Hedgehog signaling in gastrointestinal carcinogenesis and the gastrointestinal tumor microenvironment. Acta Pharm Sin B 2021, 11, 609-620. https://doi.org/10.1016/j.apsb.2020.10.022. [CrossRef]
- Li, X.; Xiang, Y.; Li, F.; Yin, C.; Li, B.; Ke, X. WNT/beta-Catenin Signaling Pathway Regulating T Cell-Inflammation in the Tumor Microenvironment. Front Immunol 2019, 10, 2293. https://doi.org/10.3389/fimmu.2019.02293. [CrossRef]
- Flanagan, D.J.; Woodcock, S.A.; Phillips, C.; Eagle, C.; Sansom, O.J. Targeting ligand-dependent wnt pathway dysregulation in gastrointestinal cancers through porcupine inhibition. Pharmacol Ther 2022, 238, 108179. https://doi.org/10.1016/j.pharmthera.2022.108179. [CrossRef]
- Liu, Y.; Qi, X.; Donnelly, L.; Elghobashi-Meinhardt, N.; Long, T.; Zhou, R.W.; Sun, Y.; Wang, B.; Li, X. Mechanisms and inhibition of Porcupine-mediated Wnt acylation. Nature 2022, 607, 816-822. https://doi.org/10.1038/s41586-022-04952-2. [CrossRef]
- Chua, K.; Sim, A.Y.L.; Yeo, E.Y.M.; Bin Masroni, M.S.; Naw, W.W.; Leong, S.M.; Lee, K.W.; Lim, H.J.; Virshup, D.M.; Lee, V.K.M. ETC-159, an Upstream Wnt inhibitor, Induces Tumour Necrosis via Modulation of Angiogenesis in Osteosarcoma. Int J Mol Sci 2023, 24. https://doi.org/10.3390/ijms24054759. [CrossRef]
- Diamond, J.R.; Becerra, C.; Richards, D.; Mita, A.; Osborne, C.; O'Shaughnessy, J.; Zhang, C.; Henner, R.; Kapoun, A.M.; Xu, L.; et al. Phase Ib clinical trial of the anti-frizzled antibody vantictumab (OMP-18R5) plus paclitaxel in patients with locally advanced or metastatic HER2-negative breast cancer. Breast Cancer Res Treat 2020, 184, 53-62. https://doi.org/10.1007/s10549-020-05817-w. [CrossRef]
- Sun, Y.; Wang, W.; Zhao, C. Frizzled Receptors in Tumors, Focusing on Signaling, Roles, Modulation Mechanisms, and Targeted Therapies. Oncol Res 2021, 28, 661-674. https://doi.org/10.3727/096504020X16014648664459. [CrossRef]
- Shirai, F.; Mizutani, A.; Yashiroda, Y.; Tsumura, T.; Kano, Y.; Muramatsu, Y.; Chikada, T.; Yuki, H.; Niwa, H.; Sato, S.; et al. Design and Discovery of an Orally Efficacious Spiroindolinone-Based Tankyrase Inhibitor for the Treatment of Colon Cancer. J Med Chem 2020, 63, 4183-4204. https://doi.org/10.1021/acs.jmedchem.0c00045. [CrossRef]
- Katoh, M. Multi-layered prevention and treatment of chronic inflammation, organ fibrosis and cancer associated with canonical WNT/beta-catenin signaling activation (Review). Int J Mol Med 2018, 42, 713-725. https://doi.org/10.3892/ijmm.2018.3689. [CrossRef]
- Moon, J.H.; Hong, S.W.; Kim, J.E.; Shin, J.S.; Kim, J.S.; Jung, S.A.; Ha, S.H.; Lee, S.; Kim, J.; Lee, D.H.; et al. Targeting beta-catenin overcomes MEK inhibition resistance in colon cancer with KRAS and PIK3CA mutations. Br J Cancer 2019, 120, 941-951. https://doi.org/10.1038/s41416-019-0434-5. [CrossRef]
- He, G.; Nie, J.J.; Liu, X.; Ding, Z.; Luo, P.; Liu, Y.; Zhang, B.W.; Wang, R.; Liu, X.; Hai, Y.; et al. Zinc oxide nanoparticles inhibit osteosarcoma metastasis by downregulating beta-catenin via HIF-1alpha/BNIP3/LC3B-mediated mitophagy pathway. Bioact Mater 2023, 19, 690-702. https://doi.org/10.1016/j.bioactmat.2022.05.006. [CrossRef]
- Braggio, D.A.; Costas, C.d.F.F.; Koller, D.; Jin, F.; Zewdu, A.; Lopez, G.; Batte, K.; Casadei, L.; Welliver, M.; Horrigan, S.K.; et al. Preclinical efficacy of the Wnt/beta-catenin pathway inhibitor BC2059 for the treatment of desmoid tumors. PLoS One 2022, 17, e0276047. https://doi.org/10.1371/journal.pone.0276047. [CrossRef]
- Shah, A.H.; Gami, A.J.; Desai, N.H.; Gandhi, J.S.; Trivedi, P.P. Tumor budding as a prognostic indicator in colorectal carcinoma: a retrospective study of primary colorectal carcinoma cases in a tertiary care center. Indian J Surg Oncol 2022, 13, 459-467. https://doi.org/10.1007/s13193-022-01498-7. [CrossRef]
- Okuyama, K.; Suzuki, K.; Yanamoto, S. Relationship between Tumor Budding and Partial Epithelial-Mesenchymal Transition in Head and Neck Cancer. Cancers (Basel) 2023, 15. https://doi.org/10.3390/cancers15041111. [CrossRef]
- Gao, L.F.; Zhong, Y.; Long, T.; Wang, X.; Zhu, J.X.; Wang, X.Y.; Hu, Z.Y.; Li, Z.G. Tumor bud-derived CCL5 recruits fibroblasts and promotes colorectal cancer progression via CCR5-SLC25A24 signaling. J Exp Clin Cancer Res 2022, 41, 81. https://doi.org/10.1186/s13046-022-02300-w. [CrossRef]
- Moller, T.; James, J.P.; Holmstrom, K.; Sorensen, F.B.; Lindebjerg, J.; Nielsen, B.S. Co-Detection of miR-21 and TNF-alpha mRNA in Budding Cancer Cells in Colorectal Cancer. Int J Mol Sci 2019, 20. https://doi.org/10.3390/ijms20081907. [CrossRef]
- Bradley, C.A.; Dunne, P.D.; Bingham, V.; McQuaid, S.; Khawaja, H.; Craig, S.; James, J.; Moore, W.L.; McArt, D.G.; Lawler, M.; et al. Transcriptional upregulation of c-MET is associated with invasion and tumor budding in colorectal cancer. Oncotarget 2016, 7, 78932-78945. https://doi.org/10.18632/oncotarget.12933. [CrossRef]
- Mielcarska, S.; Kula, A.; Dawidowicz, M.; Kiczmer, P.; Chrabanska, M.; Rynkiewicz, M.; Wziatek-Kuczmik, D.; Swietochowska, E.; Waniczek, D. Assessment of the RANTES Level Correlation and Selected Inflammatory and Pro-Angiogenic Molecules Evaluation of Their Influence on CRC Clinical Features: A Preliminary Observational Study. Medicina (Kaunas) 2022, 58. https://doi.org/10.3390/medicina58020203. [CrossRef]
- Gonzalez, L.O.; Eiro, N.; Fraile, M.; Sanchez, R.; Andicoechea, A.; Fernandez-Francos, S.; Schneider, J.; Vizoso, F.J. Joint Tumor Bud-MMP/TIMP Count at the Invasive Front Improves the Prognostic Evaluation of Invasive Breast Carcinoma. Biomedicines 2021, 9. https://doi.org/10.3390/biomedicines9020196. [CrossRef]
- Zhao, H.; Ming, T.; Tang, S.; Ren, S.; Yang, H.; Liu, M.; Tao, Q.; Xu, H. Wnt signaling in colorectal cancer: pathogenic role and therapeutic target. Mol Cancer 2022, 21, 144. https://doi.org/10.1186/s12943-022-01616-7. [CrossRef]
- Gulley, J.L.; Schlom, J.; Barcellos-Hoff, M.H.; Wang, X.J.; Seoane, J.; Audhuy, F.; Lan, Y.; Dussault, I.; Moustakas, A. Dual inhibition of TGF-beta and PD-L1: a novel approach to cancer treatment. Mol Oncol 2022, 16, 2117-2134. https://doi.org/10.1002/1878-0261.13146. [CrossRef]
- Fendler, A.; Bauer, D.; Busch, J.; Jung, K.; Wulf-Goldenberg, A.; Kunz, S.; Song, K.; Myszczyszyn, A.; Elezkurtaj, S.; Erguen, B.; et al. Inhibiting WNT and NOTCH in renal cancer stem cells and the implications for human patients. Nat Commun 2020, 11, 929. https://doi.org/10.1038/s41467-020-14700-7. [CrossRef]
- Li, M.; Younis, M.H.; Zhang, Y.; Cai, W.; Lan, X. Clinical summary of fibroblast activation protein inhibitor-based radiopharmaceuticals: cancer and beyond. Eur J Nucl Med Mol Imaging 2022, 49, 2844-2868. https://doi.org/10.1007/s00259-022-05706-y. [CrossRef]
- Huang, C.Y.; Chung, C.L.; Hu, T.H.; Chen, J.J.; Liu, P.F.; Chen, C.L. Recent progress in TGF-beta inhibitors for cancer therapy. Biomed Pharmacother 2021, 134, 111046. https://doi.org/10.1016/j.biopha.2020.111046. [CrossRef]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 (MMP-9) and its inhibitors in cancer: A minireview. Eur J Med Chem 2020, 194, 112260. https://doi.org/10.1016/j.ejmech.2020.112260. [CrossRef]
- Fuerst, R.; Choi, J.Y.; Knapinska, A.M.; Cameron, M.D.; Ruiz, C.; Delmas, A.; Sundrud, M.S.; Fields, G.B.; Roush, W.R. Development of a putative Zn(2+)-chelating but highly selective MMP-13 inhibitor. Bioorg Med Chem Lett 2022, 76, 129014. https://doi.org/10.1016/j.bmcl.2022.129014. [CrossRef]
- Sanyal, S.; Amin, S.A.; Banerjee, P.; Gayen, S.; Jha, T. A review of MMP-2 structures and binding mode analysis of its inhibitors to strategize structure-based drug design. Bioorg Med Chem 2022, 74, 117044. https://doi.org/10.1016/j.bmc.2022.117044. [CrossRef]
- Secinti, I.E.; Ozgur, T.; Dede, I. PD-L1 Expression in Colorectal Adenocarcinoma Is Associated With the Tumor Immune Microenvironment and Epithelial-Mesenchymal Transition. Am J Clin Pathol 2022, 158, 506-515. https://doi.org/10.1093/ajcp/aqac077. [CrossRef]
- Zhang, L.; Li, Z.; Skrzypczynska, K.M.; Fang, Q.; Zhang, W.; O'Brien, S.A.; He, Y.; Wang, L.; Zhang, Q.; Kim, A.; et al. Single-Cell Analyses Inform Mechanisms of Myeloid-Targeted Therapies in Colon Cancer. Cell 2020, 181, 442-459 e429. https://doi.org/10.1016/j.cell.2020.03.048. [CrossRef]
- Zhou, W.; Guo, S.; Liu, M.; Burow, M.E.; Wang, G. Targeting CXCL12/CXCR4 Axis in Tumor Immunotherapy. Curr Med Chem 2019, 26, 3026-3041. https://doi.org/10.2174/0929867324666170830111531. [CrossRef]
- Alghamri, M.S.; Banerjee, K.; Mujeeb, A.A.; Mauser, A.; Taher, A.; Thalla, R.; McClellan, B.L.; Varela, M.L.; Stamatovic, S.M.; Martinez-Revollar, G.; et al. Systemic Delivery of an Adjuvant CXCR4-CXCL12 Signaling Inhibitor Encapsulated in Synthetic Protein Nanoparticles for Glioma Immunotherapy. ACS Nano 2022, 16, 8729-8750. https://doi.org/10.1021/acsnano.1c07492. [CrossRef]
- Kobayashi, H.; Choyke, P.L. Near-Infrared Photoimmunotherapy of Cancer. Acc Chem Res 2019, 52, 2332-2339. https://doi.org/10.1021/acs.accounts.9b00273. [CrossRef]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: therapeutic implications. J Hematol Oncol 2018, 11, 64. https://doi.org/10.1186/s13045-018-0605-5. [CrossRef]
- Ma, Z.; Guo, D.; Wang, Q.; Liu, P.; Xiao, Y.; Wu, P.; Wang, Y.; Chen, B.; Liu, Z.; Liu, Q. Lgr5-mediated p53 Repression through PDCD5 leads to doxorubicin resistance in Hepatocellular Carcinoma. Theranostics 2019, 9, 2967-2983. https://doi.org/10.7150/thno.30562. [CrossRef]
- Xia, L.; Oyang, L.; Lin, J.; Tan, S.; Han, Y.; Wu, N.; Yi, P.; Tang, L.; Pan, Q.; Rao, S.; et al. The cancer metabolic reprogramming and immune response. Mol Cancer 2021, 20, 28. https://doi.org/10.1186/s12943-021-01316-8. [CrossRef]
- Li, Z.; Sun, C.; Qin, Z. Metabolic reprogramming of cancer-associated fibroblasts and its effect on cancer cell reprogramming. Theranostics 2021, 11, 8322-8336. https://doi.org/10.7150/thno.62378. [CrossRef]
- Chifman, J.; Pullikuth, A.; Chou, J.W.; Bedognetti, D.; Miller, L.D. Conservation of immune gene signatures in solid tumors and prognostic implications. BMC Cancer 2016, 16, 911. https://doi.org/10.1186/s12885-016-2948-z. [CrossRef]
- Uddin, M.N.; Li, M.; Wang, X. Identification of Transcriptional Signatures of Colon Tumor Stroma by a Meta-Analysis. J Oncol 2019, 2019, 8752862. https://doi.org/10.1155/2019/8752862. [CrossRef]
- Jasso, G.J.; Jaiswal, A.; Varma, M.; Laszewski, T.; Grauel, A.; Omar, A.; Silva, N.; Dranoff, G.; Porter, J.A.; Mansfield, K.; et al. Colon stroma mediates an inflammation-driven fibroblastic response controlling matrix remodeling and healing. PLoS Biol 2022, 20, e3001532. https://doi.org/10.1371/journal.pbio.3001532. [CrossRef]
- Kim, M.S.; Ha, S.E.; Wu, M.; Zogg, H.; Ronkon, C.F.; Lee, M.Y.; Ro, S. Extracellular Matrix Biomarkers in Colorectal Cancer. Int J Mol Sci 2021, 22. https://doi.org/10.3390/ijms22179185. [CrossRef]
- Sandberg, T.P.; Stuart, M.; Oosting, J.; Tollenaar, R.; Sier, C.F.M.; Mesker, W.E. Increased expression of cancer-associated fibroblast markers at the invasive front and its association with tumor-stroma ratio in colorectal cancer. BMC Cancer 2019, 19, 284. https://doi.org/10.1186/s12885-019-5462-2. [CrossRef]
- Stefani, C.; Miricescu, D.; Stanescu, S., II; Nica, R.I.; Greabu, M.; Totan, A.R.; Jinga, M. Growth Factors, PI3K/AKT/mTOR and MAPK Signaling Pathways in Colorectal Cancer Pathogenesis: Where Are We Now? Int J Mol Sci 2021, 22. https://doi.org/10.3390/ijms221910260. [CrossRef]
- Kobayashi, H.; Gieniec, K.A.; Lannagan, T.R.M.; Wang, T.; Asai, N.; Mizutani, Y.; Iida, T.; Ando, R.; Thomas, E.M.; Sakai, A.; et al. The Origin and Contribution of Cancer-Associated Fibroblasts in Colorectal Carcinogenesis. Gastroenterology 2022, 162, 890-906. https://doi.org/10.1053/j.gastro.2021.11.037. [CrossRef]
- Liang, Y.; Lv, Z.; Huang, G.; Qin, J.; Li, H.; Nong, F.; Wen, B. Prognostic significance of abnormal matrix collagen remodeling in colorectal cancer based on histologic and bioinformatics analysis. Oncol Rep 2020, 44, 1671-1685. https://doi.org/10.3892/or.2020.7729. [CrossRef]
- Lindgren, M.; Rask, G.; Jonsson, J.; Berglund, A.; Lundin, C.; Jonsson, P.; Ljuslinder, I.; Nystrom, H. Type IV Collagen in Human Colorectal Liver Metastases-Cellular Origin and a Circulating Biomarker. Cancers (Basel) 2022, 14. https://doi.org/10.3390/cancers14143396. [CrossRef]
- Buttacavoli, M.; Di Cara, G.; Roz, E.; Pucci-Minafra, I.; Feo, S.; Cancemi, P. Integrated Multi-Omics Investigations of Metalloproteinases in Colon Cancer: Focus on MMP2 and MMP9. Int J Mol Sci 2021, 22. https://doi.org/10.3390/ijms222212389. [CrossRef]
- Cui, G.; Cai, F.; Ding, Z.; Gao, L. MMP14 predicts a poor prognosis in patients with colorectal cancer. Hum Pathol 2019, 83, 36-42. https://doi.org/10.1016/j.humpath.2018.03.030. [CrossRef]
- Tian, L.; Zhao, Z.F.; Xie, L.; Zhu, J.P. Taurine up-regulated 1 accelerates tumorigenesis of colon cancer by regulating miR-26a-5p/MMP14/p38 MAPK/Hsp27 axis in vitro and in vivo. Life Sci 2019, 239, 117035. https://doi.org/10.1016/j.lfs.2019.117035. [CrossRef]
- Xuefeng, X.; Hou, M.X.; Yang, Z.W.; Agudamu, A.; Wang, F.; Su, X.L.; Li, X.; Shi, L.; Terigele, T.; Bao, L.L.; et al. Epithelial-mesenchymal transition and metastasis of colon cancer cells induced by the FAK pathway in cancer-associated fibroblasts. J Int Med Res 2020, 48, 300060520931242. https://doi.org/10.1177/0300060520931242. [CrossRef]
- Hu, L.; Wang, J.; Wang, Y.; Wu, L.; Wu, C.; Mao, B.; Maruthi Prasad, E.; Wang, Y.; Chin, Y.E. LOXL1 modulates the malignant progression of colorectal cancer by inhibiting the transcriptional activity of YAP. Cell Commun Signal 2020, 18, 148. https://doi.org/10.1186/s12964-020-00639-1. [CrossRef]
- Wang, J.; Li, R.; Li, M.; Wang, C. Fibronectin and colorectal cancer: signaling pathways and clinical implications. J Recept Signal Transduct Res 2021, 41, 313-320. https://doi.org/10.1080/10799893.2020.1817074. [CrossRef]
- Zhang, Z.; Fang, C.; Wang, Y.; Zhang, J.; Yu, J.; Zhang, Y.; Wang, X.; Zhong, J. COL1A1: A potential therapeutic target for colorectal cancer expressing wild-type or mutant KRAS. Int J Oncol 2018, 53, 1869-1880. https://doi.org/10.3892/ijo.2018.4536. [CrossRef]
- Kamali Zonouzi, S.; Pezeshki, P.S.; Razi, S.; Rezaei, N. Cancer-associated fibroblasts in colorectal cancer. Clin Transl Oncol 2022, 24, 757-769. https://doi.org/10.1007/s12094-021-02734-2. [CrossRef]
- Mortezapour, M.; Tapak, L.; Bahreini, F.; Najafi, R.; Afshar, S. Identification of key genes in colorectal cancer diagnosis by co-expression analysis weighted gene co-expression network analysis. Comput Biol Med 2023, 157, 106779. https://doi.org/10.1016/j.compbiomed.2023.106779. [CrossRef]
- Thibaudin, M.; Limagne, E.; Hampe, L.; Ballot, E.; Truntzer, C.; Ghiringhelli, F. Targeting PD-L1 and TIGIT could restore intratumoral CD8 T cell function in human colorectal cancer. Cancer Immunol Immunother 2022, 71, 2549-2563. https://doi.org/10.1007/s00262-022-03182-9. [CrossRef]
- Yang, Z.; Wu, G.; Zhang, X.; Gao, J.; Meng, C.; Liu, Y.; Wei, Q.; Sun, L.; Wei, P.; Bai, Z.; et al. Current progress and future perspectives of neoadjuvant anti-PD-1/PD-L1 therapy for colorectal cancer. Front Immunol 2022, 13, 1001444. https://doi.org/10.3389/fimmu.2022.1001444. [CrossRef]
- Zeynep, O.; Funda, C.; Evrim, Y.; Deniz, A.; Bulent, Y.; Fatih, Y.N. PD-L1 and PD-L2 expression in colorectal cancer. Indian J Pathol Microbiol 2023, 66, 31-37. https://doi.org/10.4103/ijpm.ijpm_814_21. [CrossRef]
- Cheruku, S.; Rao, V.; Pandey, R.; Rao Chamallamudi, M.; Velayutham, R.; Kumar, N. Tumor-associated macrophages employ immunoediting mechanisms in colorectal tumor progression: Current research in Macrophage repolarization immunotherapy. Int Immunopharmacol 2023, 116, 109569. https://doi.org/10.1016/j.intimp.2022.109569. [CrossRef]
- Liu, J.N.; Kong, X.S.; Huang, T.; Wang, R.; Li, W.; Chen, Q.F. Clinical Implications of Aberrant PD-1 and CTLA4 Expression for Cancer Immunity and Prognosis: A Pan-Cancer Study. Front Immunol 2020, 11, 2048. https://doi.org/10.3389/fimmu.2020.02048. [CrossRef]
- Chen, J.; Ye, X.; Pitmon, E.; Lu, M.; Wan, J.; Jellison, E.R.; Adler, A.J.; Vella, A.T.; Wang, K. IL-17 inhibits CXCL9/10-mediated recruitment of CD8(+) cytotoxic T cells and regulatory T cells to colorectal tumors. J Immunother Cancer 2019, 7, 324. https://doi.org/10.1186/s40425-019-0757-z. [CrossRef]
- Shang, S.; Yang, Y.W.; Chen, F.; Yu, L.; Shen, S.H.; Li, K.; Cui, B.; Lv, X.X.; Zhang, C.; Yang, C.; et al. TRIB3 reduces CD8(+) T cell infiltration and induces immune evasion by repressing the STAT1-CXCL10 axis in colorectal cancer. Sci Transl Med 2022, 14, eabf0992. https://doi.org/10.1126/scitranslmed.abf0992. [CrossRef]
- Bess, S.N.; Greening, G.J.; Rajaram, N.; Muldoon, T.J. Macrophage-targeted anti-CCL2 immunotherapy enhances tumor sensitivity to 5-fluorouracil in a Balb/c-CT26 murine colon carcinoma model measured using diffuse reflectance spectroscopy. BMC Immunol 2022, 23, 20. https://doi.org/10.1186/s12865-022-00493-5. [CrossRef]
- Li, J.; Yang, J.; Xing, R.; Wang, Y. A novel inflammation-related signature for predicting prognosis and characterizing the tumor microenvironment in colorectal cancer. Aging (Albany NY) 2023, 15, 2554-2581. https://doi.org/10.18632/aging.204630. [CrossRef]
- Soylemez, Z.; Arikan, E.S.; Solak, M.; Arikan, Y.; Tokyol, C.; Seker, H. Investigation of the expression levels of CPEB4, APC, TRIP13, EIF2S3, EIF4A1, IFNg, PIK3CA and CTNNB1 genes in different stage colorectal tumors. Turk J Med Sci 2021, 51, 661-674. https://doi.org/10.3906/sag-2010-18. [CrossRef]
- Wang, R.; Ma, Y.; Zhan, S.; Zhang, G.; Cao, L.; Zhang, X.; Shi, T.; Chen, W. B7-H3 promotes colorectal cancer angiogenesis through activating the NF-kappaB pathway to induce VEGFA expression. Cell Death Dis 2020, 11, 55. https://doi.org/10.1038/s41419-020-2252-3. [CrossRef]
- Chen, C.; Huang, Z.; Mo, X.; Song, Y.; Li, X.; Li, X.; Zhang, M. The circular RNA 001971/miR-29c-3p axis modulates colorectal cancer growth, metastasis, and angiogenesis through VEGFA. J Exp Clin Cancer Res 2020, 39, 91. https://doi.org/10.1186/s13046-020-01594-y. [CrossRef]
- Chen, C.; Huang, Z.; Mo, X.; Song, Y.; Li, X.; Li, X.; Zhang, M. Retraction Note: The circular RNA 001971/miR-29c-3p axis modulates colorectal cancer growth, metastasis, and angiogenesis through VEGFA. J Exp Clin Cancer Res 2022, 41, 114. https://doi.org/10.1186/s13046-022-02342-0. [CrossRef]
- Zhong, M.; Li, N.; Qiu, X.; Ye, Y.; Chen, H.; Hua, J.; Yin, P.; Zhuang, G. TIPE regulates VEGFR2 expression and promotes angiogenesis in colorectal cancer. Int J Biol Sci 2020, 16, 272-283. https://doi.org/10.7150/ijbs.37906. [CrossRef]
- Cai, X.; Wei, B.; Li, L.; Chen, X.; Yang, J.; Li, X.; Jiang, X.; Lv, M.; Li, M.; Lin, Y.; et al. Therapeutic Potential of Apatinib Against Colorectal Cancer by Inhibiting VEGFR2-Mediated Angiogenesis and beta-Catenin Signaling. Onco Targets Ther 2020, 13, 11031-11044. https://doi.org/10.2147/OTT.S266549. [CrossRef]
- Gaibar, M.; Galan, M.; Romero-Lorca, A.; Anton, B.; Malon, D.; Moreno, A.; Fernandez-Santander, A.; Novillo, A. Genetic Variants of ANGPT1, CD39, FGF2 and MMP9 Linked to Clinical Outcome of Bevacizumab Plus Chemotherapy for Metastatic Colorectal Cancer. Int J Mol Sci 2021, 22. https://doi.org/10.3390/ijms22031381. [CrossRef]
- Munakata, S.; Ueyama, T.; Ishihara, H.; Komiyama, H.; Tsukamoto, R.; Kawai, M.; Takahashi, M.; Kojima, Y.; Tomiki, Y.; Sakamoto, K. Angiopoietin-2 as a Prognostic Factor in Patients with Incurable Stage IV Colorectal Cancer. J Gastrointest Cancer 2021, 52, 237-242. https://doi.org/10.1007/s12029-020-00392-1. [CrossRef]
- Jary, M.; Hasanova, R.; Vienot, A.; Asgarov, K.; Loyon, R.; Tirole, C.; Bouard, A.; Orillard, E.; Klajer, E.; Kim, S.; et al. Molecular description of ANGPT2 associated colorectal carcinoma. Int J Cancer 2020, 147, 2007-2018. https://doi.org/10.1002/ijc.32993. [CrossRef]
- Wang, J.; Yu, S.; Chen, G.; Kang, M.; Jin, X.; Huang, Y.; Lin, L.; Wu, D.; Wang, L.; Chen, J. A novel prognostic signature of immune-related genes for patients with colorectal cancer. J Cell Mol Med 2020, 24, 8491-8504. https://doi.org/10.1111/jcmm.15443. [CrossRef]
- Caiado, H.; Conceicao, N.; Tiago, D.; Marreiros, A.; Vicente, S.; Enriquez, J.L.; Vaz, A.M.; Antunes, A.; Guerreiro, H.; Caldeira, P.; et al. Data on the evaluation of FGF2 gene expression in Colorectal Cancer. Data Brief 2020, 31, 105765. https://doi.org/10.1016/j.dib.2020.105765. [CrossRef]
- Cheng, X.; Jin, Z.; Ji, X.; Shen, X.; Feng, H.; Morgenlander, W.; Ou, B.; Wu, H.; Gao, H.; Ye, F.; et al. ETS variant 5 promotes colorectal cancer angiogenesis by targeting platelet-derived growth factor BB. Int J Cancer 2019, 145, 179-191. https://doi.org/10.1002/ijc.32071. [CrossRef]
- Sae-Lim, S.; Soontornworajit, B.; Rotkrua, P. Inhibition of Colorectal Cancer Cell Proliferation by Regulating Platelet-Derived Growth Factor B Signaling with a DNA Aptamer. Asian Pac J Cancer Prev 2019, 20, 487-494. https://doi.org/10.31557/APJCP.2019.20.2.487. [CrossRef]
- Zheng, Y.; Fu, Y.; Wang, P.P.; Ding, Z.Y. Neoantigen: A Promising Target for the Immunotherapy of Colorectal Cancer. Dis Markers 2022, 2022, 8270305. https://doi.org/10.1155/2022/8270305. [CrossRef]
- Gilazieva, Z.; Ponomarev, A.; Rizvanov, A.; Solovyeva, V. The Dual Role of Mesenchymal Stromal Cells and Their Extracellular Vesicles in Carcinogenesis. Biology (Basel) 2022, 11. https://doi.org/10.3390/biology11060813. [CrossRef]
- Currais, P.; Rosa, I.; Claro, I. Colorectal cancer carcinogenesis: From bench to bedside. World J Gastrointest Oncol 2022, 14, 654-663. https://doi.org/10.4251/wjgo.v14.i3.654. [CrossRef]
- Tang, C.M.; Adams, D.L. Clinical Applications of Cancer-Associated Cells Present in the Blood of Cancer Patients. Biomedicines 2022, 10. https://doi.org/10.3390/biomedicines10030587. [CrossRef]
- Kasabwala, D.M.; Bergan, R.C.; Gardner, K.P.; Lapidus, R.; Tsai, S.; Aldakkak, M.; Adams, D.L. Micronuclei in Circulating Tumor Associated Macrophages Predicts Progression in Advanced Colorectal Cancer. Biomedicines 2022, 10. https://doi.org/10.3390/biomedicines10112898. [CrossRef]
- Sutton, T.L.; Patel, R.K.; Anderson, A.N.; Bowden, S.G.; Whalen, R.; Giske, N.R.; Wong, M.H. Circulating Cells with Macrophage-like Characteristics in Cancer: The Importance of Circulating Neoplastic-Immune Hybrid Cells in Cancer. Cancers (Basel) 2022, 14. https://doi.org/10.3390/cancers14163871. [CrossRef]
Figure 1.
Unique Features and Complexity of the Colorectal Tumor Microenvironment (TME). (A): In this illustration, a comprehensive overview of the complex colorectal tumor microenvironment (TME) is provided. The image demonstrates the interaction between bacteria and their metabolites with the TME, emphasizing the influence of the microbiome on the tumor stroma. Colorectal cancer (CRC) cells are depicted within the tumor and its stroma, surrounded by key immune cells, such as tumor-associated macrophages (TAMs) and myeloid-derived suppressor cells (MDSCs). Cancer-associated fibroblasts (CAFs), which support tumor growth and invasion, are also displayed in the TME. The illustration features regulatory T cells (Tregs) that modulate immune responses, as well as the unique mucosal immune system in the colorectal mucosa, represented by the gut-associated lymphoid tissue (GALT) and its interactions with immune cells. Tumor budding, characterized by the presence of small clusters or single cancer cells at the invasive front of the tumor, is also depicted. This visual representation effectively captures the distinctive features and interactions within the colorectal TME. (B): Colorectal cancer frequently exhibits driver mutations in Wnt pathway genes, such as APC and β-catenin (CTNNB1). The canonical Wnt/β-catenin pathway is depicted, beginning with extracellular Wnt proteins binding to cell surface receptors Frizzled (Fz) and low-density lipoprotein receptor-related protein 5/6 (LRP5/6). Upon Wnt binding, the Fz receptor recruits and activates the intracellular protein Dishevelled (Dvl), leading to the inhibition of the β-catenin destruction complex, which consists of Axin, Adenomatous Polyposis Coli (APC), Glycogen Synthase Kinase-3β (GSK-3β), and Casein Kinase 1α (CK1α). This inhibition prevents the phosphorylation and subsequent degradation of β-catenin, allowing it to accumulate in the cytoplasm and translocate into the nucleus.
Figure 1.
Unique Features and Complexity of the Colorectal Tumor Microenvironment (TME). (A): In this illustration, a comprehensive overview of the complex colorectal tumor microenvironment (TME) is provided. The image demonstrates the interaction between bacteria and their metabolites with the TME, emphasizing the influence of the microbiome on the tumor stroma. Colorectal cancer (CRC) cells are depicted within the tumor and its stroma, surrounded by key immune cells, such as tumor-associated macrophages (TAMs) and myeloid-derived suppressor cells (MDSCs). Cancer-associated fibroblasts (CAFs), which support tumor growth and invasion, are also displayed in the TME. The illustration features regulatory T cells (Tregs) that modulate immune responses, as well as the unique mucosal immune system in the colorectal mucosa, represented by the gut-associated lymphoid tissue (GALT) and its interactions with immune cells. Tumor budding, characterized by the presence of small clusters or single cancer cells at the invasive front of the tumor, is also depicted. This visual representation effectively captures the distinctive features and interactions within the colorectal TME. (B): Colorectal cancer frequently exhibits driver mutations in Wnt pathway genes, such as APC and β-catenin (CTNNB1). The canonical Wnt/β-catenin pathway is depicted, beginning with extracellular Wnt proteins binding to cell surface receptors Frizzled (Fz) and low-density lipoprotein receptor-related protein 5/6 (LRP5/6). Upon Wnt binding, the Fz receptor recruits and activates the intracellular protein Dishevelled (Dvl), leading to the inhibition of the β-catenin destruction complex, which consists of Axin, Adenomatous Polyposis Coli (APC), Glycogen Synthase Kinase-3β (GSK-3β), and Casein Kinase 1α (CK1α). This inhibition prevents the phosphorylation and subsequent degradation of β-catenin, allowing it to accumulate in the cytoplasm and translocate into the nucleus.
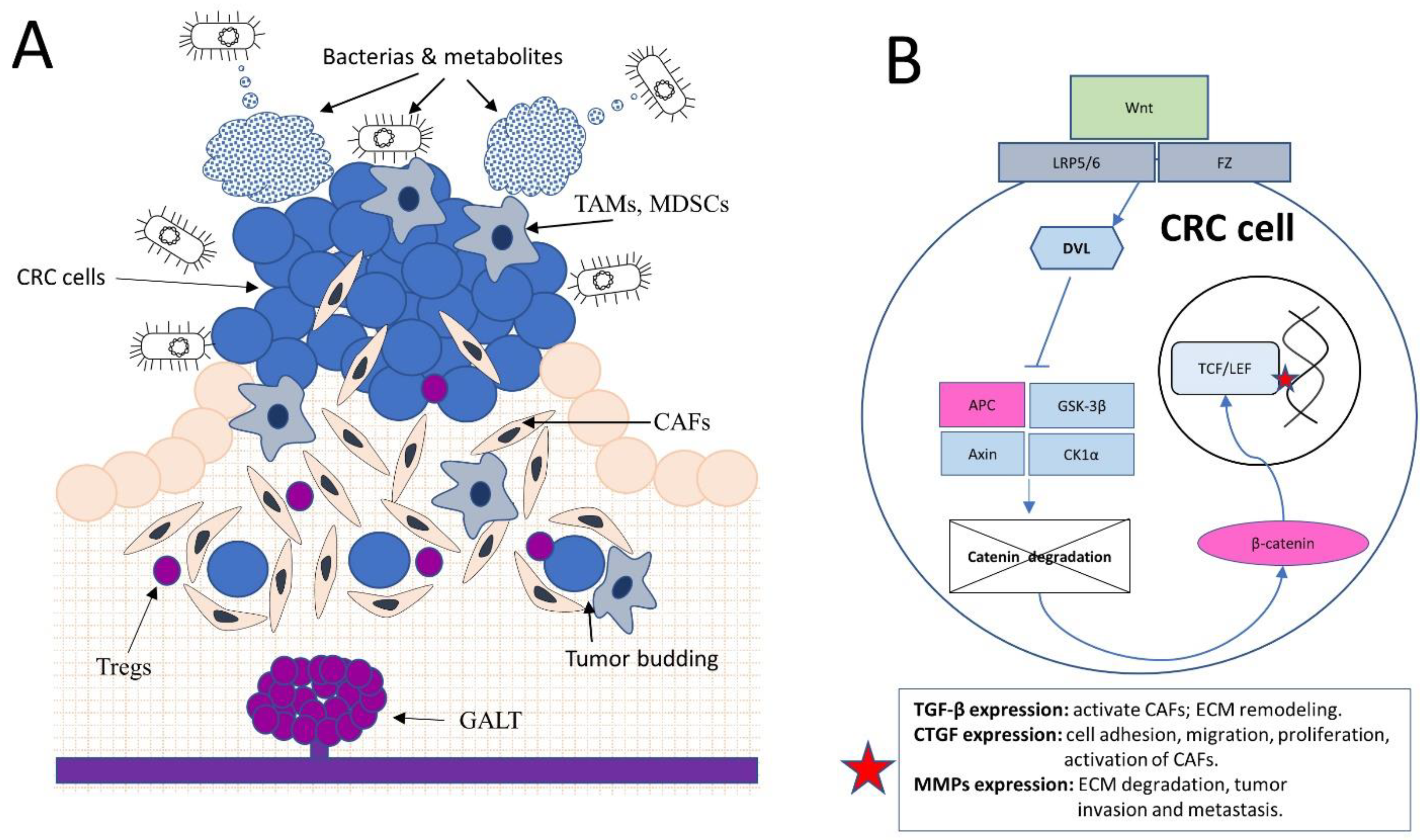
Figure 2.
Colorectal Cancer Metastasis: Tumor-Stroma Interactions and the Seed and Soil Hypothesis. (A): During tumor progression, a colorectal cancer (CRC) cell develops through mutational and epigenetic events, exhibiting uncontrolled proliferation and evasion from apoptosis. The CRC cell, or "seed," thrives within the "soil" provided by cancer-associated fibroblasts (CAFs) and tumor-associated macrophages (TAMs). These stromal cells create a humoral (“bubbles” in the picture) and cellular shield and facilitate the supply of nutrients and exogenous growth stimuli to tumor cells. Key factors involved in this process include matrix metalloproteinases (MMP2, MMP9, MMP14), chemokines (CCL2, CXCL9, CXCL10), growth factors (VEGFA, FGF2, PDGFB, TGF, TNF-a), and cytokines (IL-1b, IL-10), which contribute to extracellular matrix remodeling, immune cell recruitment, and angiogenesis. Surface molecules such as CD274 (PD-L1), PDCD1 (PD-1), and CTLA4 suppress T-cell activation, enabling tumor cells to evade immune surveillance and fostering a supportive microenvironment for the CRC cell. (B): The illustration demonstrates the interaction and invasion of Cancer-Associated Macrophage-like Cells (CAMLs) into a blood vessel. As a hybrid of macrophages and CRC cells, CAMLs utilize specific molecules, such as matrix metalloproteinases (MMPs), integrins, and selectins, to infiltrate the vessel wall, emphasizing their invasive capabilities. Moreover, CAMLs can attach to cancer-associated fibroblasts (CAFs) or facilitate tumor cells undergoing epithelial-mesenchymal transition (EMT) to ease invasion into the blood vessel. This visual representation illustrates the complex interactions within the tumor microenvironment that contribute to tumor progression and metastasis. (C): Formation of metastasis: tumor and stromal cell associates (such as CAMLs) carry the necessary "seeds" and an initial supply of "soil" to protect CRC cells during the early stages and prepare fertile "soil" for the protection and further proliferation of tumor cells. These cells utilize various molecular factors, such as integrins, matrix metalloproteinases (MMPs), and chemokines, to facilitate invasion and adaption to new tissues, promoting metastatic growth and colonization.
Figure 2.
Colorectal Cancer Metastasis: Tumor-Stroma Interactions and the Seed and Soil Hypothesis. (A): During tumor progression, a colorectal cancer (CRC) cell develops through mutational and epigenetic events, exhibiting uncontrolled proliferation and evasion from apoptosis. The CRC cell, or "seed," thrives within the "soil" provided by cancer-associated fibroblasts (CAFs) and tumor-associated macrophages (TAMs). These stromal cells create a humoral (“bubbles” in the picture) and cellular shield and facilitate the supply of nutrients and exogenous growth stimuli to tumor cells. Key factors involved in this process include matrix metalloproteinases (MMP2, MMP9, MMP14), chemokines (CCL2, CXCL9, CXCL10), growth factors (VEGFA, FGF2, PDGFB, TGF, TNF-a), and cytokines (IL-1b, IL-10), which contribute to extracellular matrix remodeling, immune cell recruitment, and angiogenesis. Surface molecules such as CD274 (PD-L1), PDCD1 (PD-1), and CTLA4 suppress T-cell activation, enabling tumor cells to evade immune surveillance and fostering a supportive microenvironment for the CRC cell. (B): The illustration demonstrates the interaction and invasion of Cancer-Associated Macrophage-like Cells (CAMLs) into a blood vessel. As a hybrid of macrophages and CRC cells, CAMLs utilize specific molecules, such as matrix metalloproteinases (MMPs), integrins, and selectins, to infiltrate the vessel wall, emphasizing their invasive capabilities. Moreover, CAMLs can attach to cancer-associated fibroblasts (CAFs) or facilitate tumor cells undergoing epithelial-mesenchymal transition (EMT) to ease invasion into the blood vessel. This visual representation illustrates the complex interactions within the tumor microenvironment that contribute to tumor progression and metastasis. (C): Formation of metastasis: tumor and stromal cell associates (such as CAMLs) carry the necessary "seeds" and an initial supply of "soil" to protect CRC cells during the early stages and prepare fertile "soil" for the protection and further proliferation of tumor cells. These cells utilize various molecular factors, such as integrins, matrix metalloproteinases (MMPs), and chemokines, to facilitate invasion and adaption to new tissues, promoting metastatic growth and colonization.
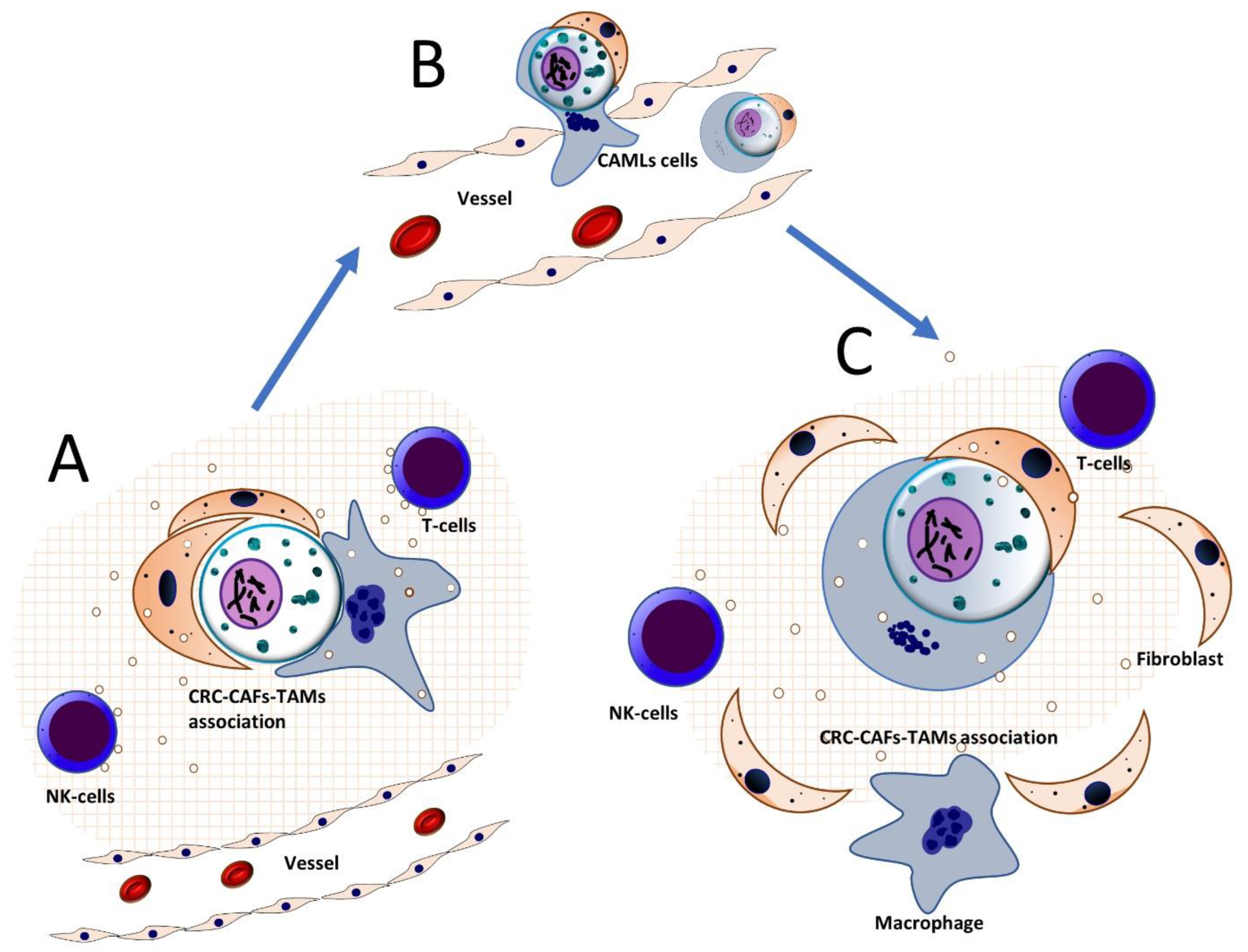

Table 1.
Bacterias of gut microbiome with pro- and anticancer activities.
| Pro-Cancer Bacterias | Anti-Cancer Bacterial Species |
|---|---|
| Fusobacterium nucleatum [5,6] | Bifidobacterium longum [13] |
| Bacteroides fragilis [14] | Lactobacillus acidophilus [15,16] |
| Enterococcus faecalis [17] | Lactobacillus rhamnosus [16,18] |
| Escherichia coli [19] | Faecalibacterium prausnitzii [20] |
| Peptostreptococcus anaerobius [21] | Bifidobacterium breve [13] |
| Streptococcus gallolyticus [22] | Lactobacillus reuteri [16] |
| Clostridium septicum [23] | Bifidobacterium adolescentis [13] |
| Ruminococcus gnavus [13] | Lactobacillus plantarum [16] |
Table 2.
Cytokines and Growth Factors Involved in Colorectal Cancer Stroma.
| Gene | Full Name | Role in Colorectal Cancer Stroma | References |
|---|---|---|---|
| MMP2 | Matrix Metalloproteinase 2 | ECM remodeling, degrades various ECM components, facilitates tumor cell invasion and metastasis. | [118,119] |
| MMP9 | Matrix Metalloproteinase 9 | ECM remodeling, degrades collagen and other ECM components, promotes tumor cell invasion, supports angiogenesis. | [118,119] |
| MMP14 | Matrix Metalloproteinase 14 | ECM remodeling, involved in the cleavage of cell surface proteins and the breakdown of ECM components, promotes tumor invasion and angiogenesis. | [120,121] |
| LOX | Lysyl Oxidase | ECM remodeling, catalyzes the cross-linking of collagens and elastin, contributes to the stiffening of the tumor microenvironment and promotes tumor progression. | [122,123] |
| FN1 | Fibronectin | ECM remodeling, involved in cell adhesion, migration, and proliferation; its increased expression is associated with tumor progression and poor prognosis in colorectal cancer. | [124] |
| COL1A1 | Collagen Type I Alpha 1 Chain | ECM remodeling, major structural component of the ECM, its increased expression is associated with tumor progression and poor prognosis in colorectal cancer. | [125] |
| COL3A1 | Collagen Type III Alpha 1 Chain | ECM remodeling, another structural component of the ECM, its increased expression is associated with tumor progression and poor prognosis in colorectal cancer. | [126] |
| COL5A1 | Collagen Type V Alpha 1 Chain | ECM remodeling, another structural component of the ECM, its increased expression is associated with tumor progression and poor prognosis in colorectal cancer. | [127] |
| CD274 | Programmed Death-Ligand 1 (PD-L1) | Immune checkpoint molecule, inhibits T cell activation, promotes immune evasion by tumor cells. | [128-130] |
| PDCD1 | Programmed Cell Death Protein 1 (PD-1) | Immune checkpoint receptor, dampens immune response, allows tumor cells to escape immune surveillance. | [128-130] |
| CTLA4 | Cytotoxic T-Lymphocyte-Associated Protein 4 | Immune checkpoint receptor, inhibits T cell activation, contributes to immune evasion by tumor cells. | [131,132] |
| CXCL9 | Chemokine (C-X-C motif) Ligand 9 | Recruits immune cells, such as T cells and natural killer cells, to the tumor microenvironment; enhanced anti-tumor immunity. | [133] |
| CXCL10 | Chemokine (C-X-C motif) Ligand 10 | Recruits immune cells, such as T cells and natural killer cells, to the tumor microenvironment; enhanced anti-tumor immunity. | [133,134] |
| CCL2 | Chemokine (C-C motif) Ligand 2 | Recruitment of monocytes, macrophages, and other immune cells to the tumor site; altered expression associated with immune cell infiltration and tumor progression. | [135,136] |
| IFNG | Interferon Gamma | Activates and modulates immune response against tumor cells, affects expression of immune checkpoint molecules and other immune-related genes. | [137] |
| VEGFA | Vascular Endothelial Growth Factor A | Promotes growth of new blood vessels from existing vasculature, stimulates endothelial cell proliferation, migration, and survival. | [138-140] |
| VEGFR2 | Vascular Endothelial Growth Factor Receptor 2 (KDR) | Primary receptor for VEGFA on endothelial cells, activation by VEGFA leads to a signaling cascade promoting angiogenesis and vascular permeability. | [141,142] |
| ANGPT1 | Angiopoietin-1 | Regulates angiogenesis by binding to the endothelial cell receptor tyrosine kinase, Tie2, promotes vessel maturation and stability. | [143,144] |
| ANGPT2 | Angiopoietin-2 | Acts as an antagonist of ANGPT1, binds to Tie2, promotes vessel destabilization and sprouting angiogenesis. | [144,145] |
| FGF2 | Fibroblast Growth Factor 2 (bFGF) | Regulates angiogenesis, stimulates endothelial cell proliferation, migration, and differentiation, acts synergistically with VEGFA to promote blood vessel formation. | [146,147] |
| PDGFB | Platelet-Derived Growth Factor B | Promotes recruitment of pericytes to newly formed blood vessels, essential for blood vessel maturation and stabilization. | [148,149] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



