
Submitted:
10 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
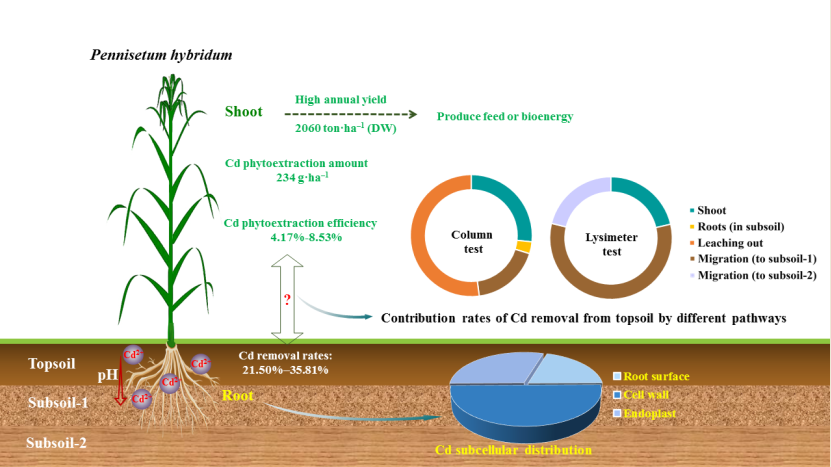
This study was conducted to identify soil cadmium (Cd) removal pathways and their contribution rates during phytoremediation by Pennisetum hybridum, as well as to comprehensively assess its phytoremediation potential. Multilayered soil column tests and farmland simulating lysimeter tests were conducted to investigate the Cd phytoextraction and migration patterns in topsoil and subsoil simultaneously. The aboveground annual yield of P. hybridum grown in the lysimeter was 206 ton·ha–1. The total amount of Cd extracted in P. hybridum shoots was 234 g·ha–1, which was similar to that of other typical phytoremediation plants. After the test, the topsoil Cd removal rate was 21.50%–35.81%, whereas the extraction efficiency in P. hybridum shoots was only 4.17%–8.53%. These findings indicate that extraction by plant shoots was not the most important contributor to the decrease of Cd in the topsoil. The proportions of Cd retained by root cell wall was approximately 50% of the total Cd in root. Based on column test results, P. hybridum treatment led to a significant decrease in soil pH and considerably enhanced Cd migration to subsoil and groundwater. P. hybridum decreases Cd in the topsoil through multiple pathways and provides a relatively ideal material for phytoremediation of Cd-contaminated soils.
Keywords:
Pennisetum hybridum
; Cadmium
; Phytoextration
; Migration
; Removal pathways
1. Introduction
Cadmium (Cd) contamination has emerged as one of the world’s most concerning environmental issues. In China, the rate of sites containing Cd in excess of regulatory limits has reached more than 7% [1]. Rice (Oryza sativa L.), which is a major grain crop grown worldwide, usually faces a high risk of Cd exposure [2]. Approximately 1/10 of the rice produced in China has a Cd content above the limit of the national food safety standard (0.2 mg·kg–1) [3]. As a result, there is an urgent need to control and remediate Cd-contaminated farmland soils.
Phytoextraction is a cost-effective approach that is suitable for the remediation of heavy metal-contaminated soils over large areas and provides high comprehensive benefits [4]. Tolerant plants with high biomass have recently been developed and used for phytoextraction of heavy metals. The hybrid giant napier (Pennisetum hydridum) is an energy plant with strong stress resistance, high productivity, and well-developed roots [5]. Based on its tolerance to heavy metals [6], P. hybridum has been proposed for phytostabilization of heavy metals in soils [7]. It has also been suggested that P. hybridum could be used for phytoextraction of Cd from contaminated soils [8]. Subsequent research showed that ammonium chloride enhanced the removal of soil Cd and Zn by P. hybridum, whereas digestate increased its shoot biomass and local Cd accumulation [9]. Additionally, intercropping of Solanum photeinocarpum (a hyperaccumulator) with P. hybridum led to considerable improvement of the phytoremediation potential for Cd-contaminated soil [10]. However, researchers have often explored the mechanisms by which P. hybridum remediates soil through pot experiments with a single soil layer. Moreover, the majority of previous studies have focused on Cd phytoextraction and removal by the shoots of P. hybridum, while the indirect effects and phytoremediation potential of the roots have received little attention. There have also been few comprehensive and in-depth analyses of the pathways of Cd removal from soil systems.
The developed roots of P. hydridum have a larger bioconcentration factor (BCF) than the shoots, which calls for further investigation of the effects and mechanisms of rhizoremediation. Therefore, multilayered soil column and farmland simulating lysimeter tests were conducted to investigate the phytoextraction and migration patterns of Cd in different soil layers. The various pathways of Cd removal from soil and their contribution rates during the remediation process were clarified to assess the phytoremediation potential and effects of P. hydridum.
2. Results
2.1. Plant growth of P. hybridum
Table 1 and Table 2 present the plant growth observed during this study. There was no remarkable stress on plant growth or development of P. hybridum in soil under moderate Cd pollution, and normal growth without inhibition at any stages was maintained. The root length of P. hybridum reached more than 50 cm under column test conditions, indicating that the developed roots of P. hybridum can pass through the topsoil layer (0–20 cm) to enter the subsoil layer. Under the lysimeter test conditions, the plant height of P. hybridum was greater than 250 cm in all cases, and the tallest plants reached 312.5 cm. The yield per hectare in each harvest of P. hybridum was greater than 4.4×104 kg dry weight (DW), which was markedly higher than that of rice grown under the same conditions. These findings verified that P. hybridum is a high-biomass energy plant.
2.2. Cd distribution in various parts of P. hybridum
As shown in Figure 1, the distribution of Cd concentrations in various parts of P. hybridum followed the order topsoil roots > stems > leaves > subsoil roots (based on column tests; Figure 1a) and topsoil roots > leaves > stems > subsoil roots (based on lysimeter test results; Figure 1c). When compared with other parts, topsoil roots had significantly higher Cd concentrations (α=0.05), and the weighted mean Cd concentration of roots was higher than that of shoots. The cumulative Cd concentration of P. hybridum roots was 2 mg·kg–1 under mild pollution and close to 4 mg·kg–1 under moderate pollution. These results demonstrate that roots are involved in P. hybridum having the highest cumulative concentration of Cd.
In the lysimeter test, the Cd concentrations of P. hybridum shoots were significantly higher than those of common Simiao rice and lower than those of upland rice (Figure 1b/c). The shoot Cd concentrations of P. hybridum were all less than 2 mg·kg–1 dry weight, which corresponded to less than 0.5 mg·kg–1 fresh weight based on a 75% water content. These results were below the standard limit (1 mg·kg–1) specified in the hygienic standard for feeds (GB 13078-2017), indicating that P. hybridum shoots can be used for feed production as well as biofuel.
2.3. Cd uptake and accumulation patterns of P. hybridum
2.3.1. Bioconcentration and translocation factors of Cd in various parts of P. hybridum
Since BCF and TF are key reference factors for evaluating a plant’s capacity to take up and translocate heavy metals, they can be used to evaluate the phytoextraction capacity of plants [11]. The BCF and TF values of Cd in various parts of P. hybridum are listed in Table 3.
The topsoil roots of P. hybridum achieved the highest BCFs for soil Cd (BCF > 2), with larger BCF values being associated with longer planting times. The BCFs varied between 0.65 and 1.25 in the stems and leaves, suggesting that P. hybridum is not an accumulator plant. The BCFs in subsoil roots of P. hybridum were comparable to those in the leaves. Additionally, higher BCFs were observed for P. hybridum in the lysimeter test (close to the field environment), which was most likely a result of the longer time for root growth and Cd accumulation. The TFs in P. hybridum shoots were all less than 1, with no significant differences between the stems and leaves.
The above results suggest that the capacity of P. hybridum to translocate Cd from roots into shoots is not high, and that P. hybridum roots have relatively strong Cd retention.
2.3.2. Location of Cd storage in P. hybridum roots
We analyzed the Cd subcellular distribution to determine the location and form of Cd stored in P. hybridum roots (Figure 2). Of the Cd contained in P. hybridum roots, 20.13% was intercepted at the root surface and 49.95% was immobilized in the root cell wall, whereas only 29.92% entered the endoplast. These results indicate that root cell wall binding is the primary mechanism that allows P. hybridum to retain heavy metals in its roots.
2.4. Phytoextraction capacity of P. hybridum for Cd
Table 4 presents the amount and efficiency of soil Cd extraction by P. hybridum determined in the column test. In test 1, the amount of Cd extracted was highest (0.567 mg·plant–1) in topsoil roots of P. hybridum. In test 2, the highest amount of Cd extracted (0.1136 mg·plant–1) was observed in stems of P. hybridum. The Cd extraction efficiency in P. hybridum shoots was 4.17% and 5.74% for the two tests, respectively. In test 1, the extraction efficiency was ranked as roots > shoots, whereas in test 2 it was shoots > roots. This may have occurred because the duration of test 2 was longer than that of test 1, allowing the shoot biomass of P. hybridum to increase and enhance heavy metals translocation to the shoots. Overall, these results indicate P. hybridum can accumulate a larger amount of Cd in its shoots than roots under certain conditions.
We simulated the field environment using percolating filters to further explore the phytoextraction and phytoremediation potential of P. hybridum by shoots for soil Cd (Table 5). Four harvests of P. hybridum were obtained over one year under the simulated field conditions, with a total Cd extraction amount of 23.40 mg·m–2 and a Cd extraction efficiency of 8.53% in the shoots. When upland rice and Simiao rice were grown for one cropping season under the same soil conditions, the total Cd extraction amount in their shoots was only 3.02 mg·m–2; however, the shoot Cd extraction of P. hybridum was ~7.5 times that of rice. Despite having no prominent ability to translocate Cd to the shoots, P. hybridum has a relatively high extraction efficiency for soil Cd because of its high biomass.
2.5. Variation of soil pH and Cd concentration
2.5.1. Variation of soil pH
Table 6 and Table 7 show the soil pH under different treatments before and after the test. In both column tests, a significant decrease in the pH of topsoil and subsoil treated with P. hybridum occurred. In the lysimeter test, the topsoil and subsoil pH also decreased with increasing harvest under P. hybridum treatment. In addition, the pH differed significantly between treatments.
2.5.2. Variation of soil Cd concentration
The concentrations of Cd in topsoil treated with P. hybridum decreased significantly (α = 0.05) in both the column and lysimeter tests (Figure 3). After 4 months, 6 months, and 1 year of phytoremediation, the topsoil Cd concentrations decreased by 23.62%, 21.50%, and 35.81% in column test 1, column test 2, and the lysimeter test, respectively (Table 8). In the lysimeter test, which was closer to the field environment, the phytoremediation effect on topsoil manifested in the fourth harvest (1 year). The resulting decrease in topsoil Cd concentrations was greater than that observed after planting rice. These findings indicate that P. hybridum has excellent phytoremediation effects on Cd-contaminated soils.
Following P. hybridum treatment, the Cd concentration of the second soil layer also increased significantly relative to the initial concentration; however, there was no noticeable difference between plants. In the lysimeter test, there was an increase in the Cd concentration of the bottom (third) soil layer, and this increase was more pronounced in the treatment with continuous planting of P. hybridum. These results indicate that the roots of P. hybridum have the capacity to transfer soil Cd towards lower layers.
2.6. Soil Cd migration and removal pathways
We calculated soil Cd migration and removal pathways based on the column test. The distribution patterns of Cd in the soil systems of various treatments after testing are shown in Table 9. The contribution of different pathways to soil Cd removal occurred in the following order: leaching from soil > extraction by shoots > migration to subsoil > translocation in subsoil roots. These results showed that leaching from soil was the largest contributor to topsoil Cd removal, accounting for ~50%. The second largest contributor was extraction by aboveground shoots, which accounted for only 20%–30% (Figure 4a).
In the lysimeter test, it was not possible to collect the roots completely. In the topsoil of the lysimeter test, the different removal pathways of Cd were: extraction by shoots and migration to subsoil layer 1 (20–40 cm), and migration to subsoil layer 2 (40–90 cm) (sealed at bottom). Overall, the contribution rates of these three removal pathways were 21.37%, 57.49%, and 21.14%, respectively (Figure 4b). Additionally, the rate of extraction by shoots in the lysimeter test was similar to that in the column test. However, migration to subsoil layer 1 made the greatest contribution to soil Cd removal in the lysimeter test, accounting for ~60%. Accordingly, the extent of downward Cd leaching in lysimeters was lower than that in soil columns. Overall, these results indicate that the primary pathway by which P. hybridum decreases topsoil Cd is not extraction by shoots, but rather promotion of downward Cd migration.
3. Discussion
3.1. Advantages of P. hybridum for phytoremediation of Cd-contaminated soils
It has been suggested that P. hybridum is highly tolerant to Cd contamination [12]. In the present study, soils contaminated with low concentrations of Cd had little impact on growth of P. hybridum, and its annual yield in lysimeters reached 2.06×105 kg·ha–1 DW. Given the high biomass of P. hybridum, it has great potential for removal of Cd from soil [13]. Generally, the effects of plant roots on soil heavy metals is limited to a few millimeters to 1–2 cm from the root surface in the rhizosphere zone [14]. Our observations indicate that the fibrous roots of P. hybridum are highly developed and can form fibrous root networks in a relatively short period. Therefore, it is likely that P. hybridum roots can extract high levels of heavy metals by contacting more soil particles. Moreover, the developed roots of P. hybridum are likely to affect the subsoil below the plow layer. In summary, P. hybridum is easy to grow and manage, has a well-developed root system and has high biomass, which is advantageous for phytoremediation of contaminated soils.
The results of this study showed that the total amount of Cd extracted by P. hybridum shoots annually was 234 g·ha–1 under simulated field environmental conditions. Among hyperaccumulators grown under field conditions, Sedum alfredii Hance can extract 184 g·ha–1 of Cd per year, whereas Sauropus androgynus (L.) Merr. can only extract 64 g·ha-1 per year [15]. Overall, the results of the present study indicate that the amount of Cd extracted by P. hybridum shoots can exceed that extracted by S. alfredii and S. androgynus. Some studies have suggested that the high biomass and rapid growth of P. hybridum contribute to a relatively large total extraction amount of heavy metals, despite its poor ability to transfer heavy metals to shoots; accordingly, P. hybridum may also be effective at phytoremediation of soils contaminated with heavy metals [16]. Based on a field test, Xie et al. [17] demonstrated that the amount of Cd extracted by P. hybridum and the extraction efficiency were 119.9 g·ha–1 and 6.98%, respectively, indicating its phytoremediation effect was superior to that of Solanum nigrum L., a Cd hyperaccumulator.
Based on the decrease in total soil Cd concentration, the rate of Cd removal from the topsoil in column test 1, column test 2, and the lysimeter test was 23.62%, 21.50%, and 35.81%, respectively (Table 8). However, the efficiency of P. hybridum extraction by shoots was only 4.17%, 5.74%, and 8.53%, respectively (Table 4 and Table 5), which was significantly different from the soil Cd removal rate. This indicates that extraction by shoots is not the most prominent contributor to phytoremediation of Cd-contaminated soils.
Evaluation of the removal pathways indicated that P. hybridum removes more Cd by facilitating its movement from the topsoil to deeper soil layers and groundwater than via extraction by shoots. However, the specific mechanisms by which this occurs were not elucidated. It is possible that these mechanisms are related to leaching being enhanced by the decreased soil pH and root exudates, as the amount of Cd brought directly to the subsoil by plant roots was insignificant as shown in this study.
Ma et al. [18] found that planting with P. hybridum considerably neutralized the alkalinity of a contaminated soil and strongly absorbed trace heavy metals such as Zn, Mn, Fe and Cu, causing a significant decrease in the heavy metals concentrations in the contaminated soil. In the present study, a remarkable decrease in soil pH occurred after planting P. hybridum (Table 6 and Table 7). The increase in H+ concentration that occurred after planting could lead to increased Cd ion exchange at the surface of soil particles, increasing the Cd concentration in soil solution [19]. However, organic acids in root exudates could chelate heavy metal ions and mobilize heavy metals near plant roots [20]. Additional research is required to further decipher the possible dynamic effects of the developed roots and root exudates of P. hybridum on the speciation and migration of soil heavy metals.
In situ chemical flushing can cause secondary pollution and damage soil structure. The ‘bioleaching’ of P. hybridum roots can be considered an alternative to conventional chemical leaching, which allows Cd removal from the plow layer through downward migration. Previous studies using P. hybridum for soil phytoremediation have mainly looked at extraction by shoots, resulting in the phytoremediation efficiency calculated based on the extraction amount in shoots being too conservative and the phytoremediation capacity of P. hybridum being underestimated. Improving the phytoremediation capacity of P. hybridum necessitates in-depth research of various removal pathways and strengthening the migration-promoting ability of plant roots.
3.2. Forms of Cd stored in roots of P. hybridum
Root surface interception is a self-protective behavior [21]. The surfaces of rice roots have been found to intercept ~30% of Cd through iron plaques without the addition of exogenous Fe [22]. Zhao et al. [23] reported that early Cd stress response induces transcriptional changes in cell wall remodeling, which may be involved in Cd stress tolerance and Cd ion accumulation in P. hybridum. Other similar herbs can effectively detoxify Cr, As, and Pb through cell wall precipitation and vesicular compartmentalization [24]. The results of the present study demonstrate that root cell wall retention is the principal mechanism of Cd storage and detoxification in P. hybridum roots, but that root surface interception also plays an important role in this process.
4. Materials and Methods
4.1. Experimental materials
Seed stems of P. hydridum were provided by the ecological farm of South China Agricultural University (Guangzhou, Guangdong Province, China). All Cd-contaminated soils (topsoils) were collected from actual paddy fields under moderate to mild pollution. The contaminated soil used in column test 1 was obtained from a town in A City, Guangdong Province, while that used in column test 2 and the lysimeter test came from a town in B City, Guangdong Province. Clean soils (subsoils) were collected from the experimental farm of South China Agricultural University (column tests 1 and 2) and a town in B City, Guangdong Province (lysimeter test). The chemical properties and Cd contents of the tested soils are summarized in Table 10.
4.2. Experimental design
4.2.1. Column test
Time of test 1: September 8, 2020–January 8, 2021
Time of test 2: April 25, 2021–October 25, 2021
The column test was conducted outdoors with ventilation and light at the experimental farm of South China Agricultural University. In each test, six plexiglass columns with a 10 cm inner diameter and 50 cm height were prepared. Three columns were used as blank controls (no plant), while the other three were planted with one P. hydridum each. All columns were wrapped with thin black film and filled with the following materials from bottom to top: 5 cm of quartz sand, 20 cm of uncontaminated soil (subsoil), and 20 cm of Cd-contaminated soil (topsoil). Before starting the test, soils were air-dried and passed through a 1 cm sieve. After the column was filled, it was saturated with tap water. During the test period, the plants in columns were managed with routine practices.
4.2.2. Lysimeter test (July 12, 2021–July 13, 2022)
Six lysimeters (square percolating filters) with 1 m long sides were filled with 50 cm of uncontaminated local soil in the bottom layer (the same subsoil used in column test 1 (Table 10)), 20 cm of uncontaminated subsoil in the second layer, and 20 cm of Cd-contaminated topsoil in the surface layer. A black nylon net was spread between each soil layer to separate them. Before beginning the test, the soil was saturated by tap water irrigation. There were two treatments (P. hydridum and rice) and three replications, which were randomly assigned to six filters. Rice was grown at a density of 16 plants·m–2, with upland rice (cv. Hanyou-73) planted as late rice in 2021 and Simiao rice (cv. Zengcheng) as early rice in 2022. P. hydridum was grown at a density of 4 plants·m–2. P. hydridum was mowed once every three months, after which the stubble was left to re-grow.
4.3. Sample preparation and analysis
Soil samples were air-dried and ground with an agate mortar, passed through 20- and 100-mesh sieves, and then stored in sealed plastic bags at 0°C–4°C prior to analysis. Samples of different plant parts were deactivated in a 70°C oven for 30 min and then dried at 55°C until constant weight. Dry plant samples were pulverized using a pulverizer, passed through a 100-mesh sieve, and then stored in sealed plastic bags at 0°C–4°C before analysis.
Soil total Cd and plant total Cd were analyzed by microwave digestion-graphite furnace atomic absorption photometry based on the environmental standard method (HJ 832–2017) and the national standard method (GB 5009.15–2014), respectively. Test results were verified using national standard materials for soil (GBW07405a) and plants (GBW(E)100348a). Soil pH was measured with a pH meter at a water:soil ratio of 2.5:1 (v/w).
One group of fresh root samples of P. hydridum was digested with a mixture of concentrated HNO3–HClO4 (4:1, v/v) and then used to determine the total Cd content (GB 5009.15-2014). Another group of fresh root samples was soaked in 20 mmol·L–1 Na2-EDTA to remove the Cd adsorbed at the root surface [25]. After filtration, the Cd concentration in the solution was determined and used to calculate the Cd content at the root surface. Finally, the remaining Cd content in the soaked root samples was determined using the same method for the analysis of root total Cd (GB 5009.15-2014).
The Cd adsorbed by the root surface was removed as previously described [25], after which a methanol-chloroform mixture (2:1, v/v) was used to remove cell inclusions based on the method described by Hart et al. [26]. This left morphologically intact root cell walls, which were then analyzed for Cd content using the same method that was used for analysis of root total Cd (GB 5009.15-2014).
4.4. Data analysis
Data processing and graphing were accomplished using Microsoft Excel 365 (Microsoft Corp., Redmond, WA, USA). One-way analysis of variance (ANOVA) and Duncan’s multiple range tests were used to identify statistically significant differences between treatments (α = 0.05) using SPSS 21.0 (IBM Corp., Armonk, NY, USA).
The following methods were used to calculate the factors related to heavy metal migration in the soil–plant system:
- Bioconcentration factor (BCF) = Cd concentration in a plant part/initial soil Cd total concentration
- Translocation factor (TF) = Cd concentration in an aerial part/root Cd concentration
- Extraction efficiency = Cd extraction amount by plants/initial total Cd content in soil
- Removal rate = decrease in soil Cd content after planting/initial total Cd content in soil
5. Conclusions
P. hybridum with stubble growth produced high biomass under environmental conditions in lysimeters that simulated field conditions. Specifically, its annual yield was greater than 206 ton·ha–1 DW, and the total amount of Cd extracted was 234 g·ha–1 per year. Moreover, the amount of Cd removed by P. hybridum shoots was similar to the amount removed by other typical phytoremediation plants.
In all cases, the total Cd in topsoil decreased dramatically following P. hybridum treatment, with removal of 21.50%–35.81%. However, the efficiency of Cd extraction by P. hybridum shoots was only 4.17%–8.53%, indicating that extraction by plant shoots was not the most important contributor to the decrease of Cd in topsoil.
P. hybridum treatment resulted in a significant decrease in soil pH. Additionally, P. hybridum roots could penetrate the subsoil and enable topsoil Cd to transfer to subsoil layers. Therefore, P. hybridum is an ideal material for phytoremediation of Cd-contaminated soils and further investigation of the dynamic effects of its roots on heavy metal speciation and migration are warranted.
Author Contributions
Conceptualization, Q.T. Wu; methodology, C. Chen; investigation, C. Chen and K.Hu.; resources, Q.T.W. and Z. Wei.; writing—original draft preparation, C. Chen; writing—review and editing, Q.T. Wu; supervision, Q.T. Wu. and Z. Wei; funding acquisition, Q.T. Wu. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research & Developmental Program of China, grant no. 2022YFC3701304 and the Local Innovation and Entrepreneurship Team Project of Guangdong Special Support Program, grant no. 2019BT02L218.
Data Availability Statement
The data that support the findings are presented in this paper. Other data are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Liwen Bianji, Edanz Group China for editing the English text of a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, X.; Zhong, T.; Liu, L.; Ouyang, X. Impact of soil heavy metal pollution on food safety in China. PLoS One 2015, 10, e0135182. [Google Scholar] [CrossRef] [PubMed]
- Chaney, R.L. How does contamination of rice soils with Cd and Zn cause high incidence of human Cd disease in subsistence rice farmers. Current Pollution Reports 2015, 1, 13–22. [Google Scholar] [CrossRef]
- Li, J.; Xu, Y. WITHDRAWN: Immobilization of Cd in a paddy soil using moisture management and amendment. Chemosphere 2015, 122, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Zia Ur Rehman, M.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci Total Environ 2018, 631-632, 1175–1191. [Google Scholar] [CrossRef]
- Hei, L.; Lee, C.C.; Wang, H.; Lin, X.Y.; Chen, X.H.; Wu, Q.T. Using a high biomass plant Pennisetum hydridum to phyto-treat fresh municipal sewage sludge. Bioresour Technol 2016, 217, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.C.; He, J.B.; Cheng, H.; Luo, S.M.; He, H.Z.; Zhang, W.Q.; Zhang, Z.M.; Li, H.S. Effects of Cd polluted soil on the modular growth and physiological characteristics of Pennisetum hydridum. Journal of Agro-Environment Science 2014, 33, 276–282. [Google Scholar]
- Wiangkham, N.; Prapagdee, B. Potential of Napier grass with cadmium-resistant bacterial inoculation on cadmium phytoremediation and its possibility to use as biomass fuel. Chemosphere 2018, 201, 511–518. [Google Scholar] [CrossRef]
- Hu, L.; Wang, R.; Liu, X.; Xu, B.; Xie, T.; Li, Y.; Wang, M.; Wang, G.; Chen, Y. Cadmium phytoextraction potential of king grass (Pennisetum sinese Roxb.) and responses of rhizosphere bacterial communities to a cadmium pollution gradient. Environ Sci Pollut Res Int 2018, 25, 21671–21681. [Google Scholar] [CrossRef]
- He, L.; Zhu, Q.; Wang, Y.; Chen, C.; He, M.; Tan, F. Irrigating digestate to improve cadmium phytoremediation potential of Pennisetum hybridum. Chemosphere 2021, 279, 130592. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wu, P.; Feng, J.F.; Guo, Y.H.; Gao, B. Effects of intercropping on Cd, Pb, and Zn accumulation using hyperaccumulators and energy plants. Journal of Agro⁃Environment Science 2021, 40, 1481–1491. [Google Scholar]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals--concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Hu, L.; Liu, X.G.; Deng, Y.W.; Liu, M.J.; Xu, B.; Wang, M.K.; Wang, G. Influences of king grass (Pennisetum sinese Roxb)-enhanced approaches for phytoextraction and microbial communities in multi-metal contaminated soil. Geoderma 2017, 307, 253–266. [Google Scholar] [CrossRef]
- Cui, H.; Fan, Y.; Yang, J.; Xu, L.; Zhou, J.; Zhu, Z. In situ phytoextraction of copper and cadmium and its biological impacts in acidic soil. Chemosphere 2016, 161, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Wang, W.; Jiang, R.f.; Li, H.f. Comparison of phytoextraction efficiency of Cd with the hyperaccumulator Thlaspi caerulescens and three high biomass species. Act Ecologica Sinica 2009, 29, 2732–2737. [Google Scholar]
- Shen, S.L.; Li, H.S.; Xia, B.C.; Yang, C.L. A field experiment on phytoextraction of heavy metals from highly contaminated soil using big biomass plants of sauropus androgynus and manihot sp. Journal of Agro-Environment Science 2013, 32, 572–578. [Google Scholar]
- Ma, C.J.; Ming, H.; Lin, C.H.; Naidu, R.; Bolan, N. Phytoextraction of heavy metal from tailing waste using Napier grass. Catena 2016, 136, 74–83. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, X.M.; Xie, Z.; Wu, K.Q.; Li, X.L.; Yang, R.G.; Peng, B.; Yu, M.H.; He, J.H. Phytoremediation efficiency of Pennisetum hydridum for acid- and cadmium-polluted soil and its safe utilization. Journal of Agro-Environment Science 2016, 35, 478–484. [Google Scholar]
- Ma, C.J.; Liu, F.G.; Lin, C.H. Study on improving the degraded soil with Napier grass (Pennisetum Hydridum). Journal of Shaoguan University ·Natural Science 2012, 33, 44–47. [Google Scholar]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Factors affecting phytoextraction: A review. Pedosphere 2016, 26, 148–166. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Hao, X.W.; Lei, M.; Tie, B.Q. The remediation technology and remediation practice of heavy metals-contaminated soil. Journal of Agro-Environment Science 2013, 32, 409–417. [Google Scholar]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Lannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environmental and Experimental Botany 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Sun, X.X.; Zhu, J.; Tao, R.P.; Xiong, H.X.; Xu, Y.Q. Effect of exogenous iron on soil Cd accumulation of rice. Journal of Yangzhou University (Natural Science Edition) 2022, 25, 74–78. [Google Scholar]
- Zhao, J.; Xia, B.; Meng, Y.; Yang, Z.; Pan, L.; Zhou, M.; Zhang, X. Transcriptome analysis to shed light on the molecular mechanisms of early responses to cadmium in roots and leaves of king grass (Pennisetum americanum x P. purpureum). Int J Mol Sci 2019, 20, 2532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Tian, C.; Gao, B. Heavy metal tolerance and phytoremediation potential of energy crop, king grass. Chinese Journal of Environmental Engineering 2017, 11, 3204–3213. [Google Scholar]
- Du, R.J.; He, E.K.; Tang, Y.T.; Hu, P.J.; Ying, R.R.; Morel, J.L.; Qiu, R.L. How phytohormone IAA and chelator EDTA affect lead uptake by Zn/Cd hyperaccumulator Picris divaricata. Int J Phytoremediation 2011, 13, 1024–1036. [Google Scholar] [CrossRef]
- Hart, J.J.; Di Tomaso, J.M.; Linscott, D.L.; Kochian, L.V. Characterization of the transport and cellular compartmentation of paraquat in roots of intact maize seedlings. Pesticide Biochemistry and Physiology 1992, 43, 212–222. [Google Scholar] [CrossRef]
Figure 1.
Concentration of Cd in different parts of Pennisetum hybridum and rice (dry weight basis). (a): Column test; (b/c) : Lysimeter test.
Figure 1.
Concentration of Cd in different parts of Pennisetum hybridum and rice (dry weight basis). (a): Column test; (b/c) : Lysimeter test.
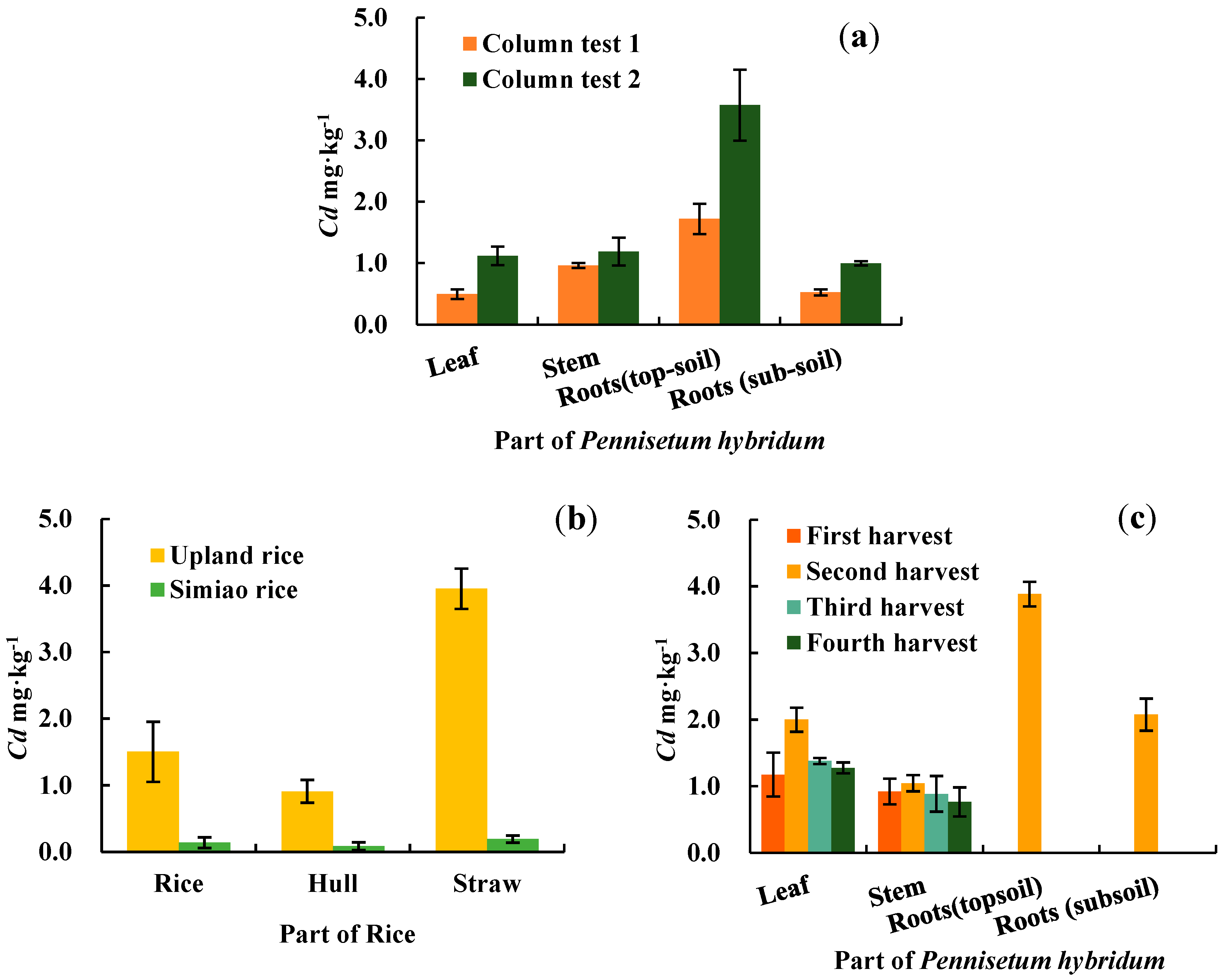
Figure 2.
Cd subcellular distribution in roots of Pennisetum hybridum.
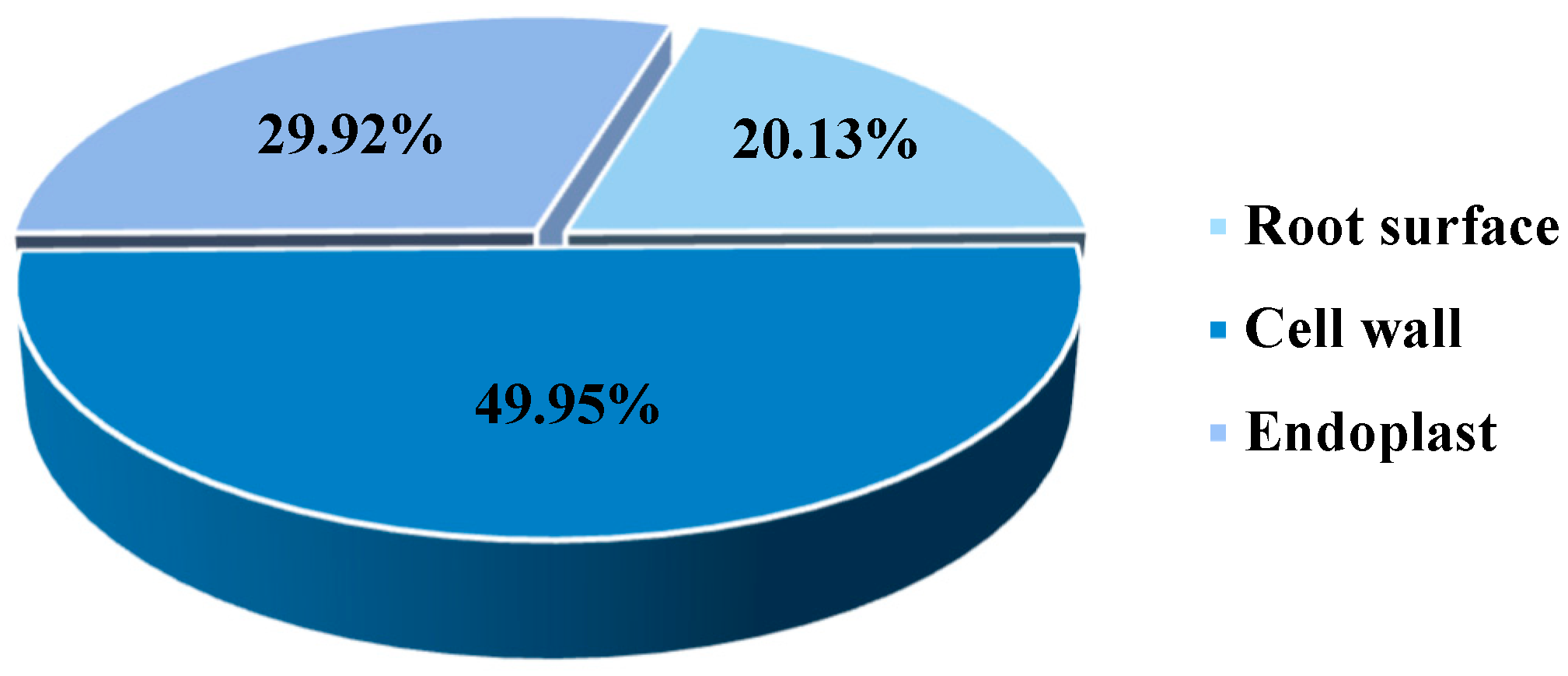
Figure 3.
Changes in Cd content in soil before and after the test. (a): column tests; (b): lysimeter test; HGN: Hybrid giant napier (Pennisetum hybridum). Values for the same test and soil affected by the same letter were not significantly different according to Duncan’s multiple range tests (α=0.05).
Figure 3.
Changes in Cd content in soil before and after the test. (a): column tests; (b): lysimeter test; HGN: Hybrid giant napier (Pennisetum hybridum). Values for the same test and soil affected by the same letter were not significantly different according to Duncan’s multiple range tests (α=0.05).
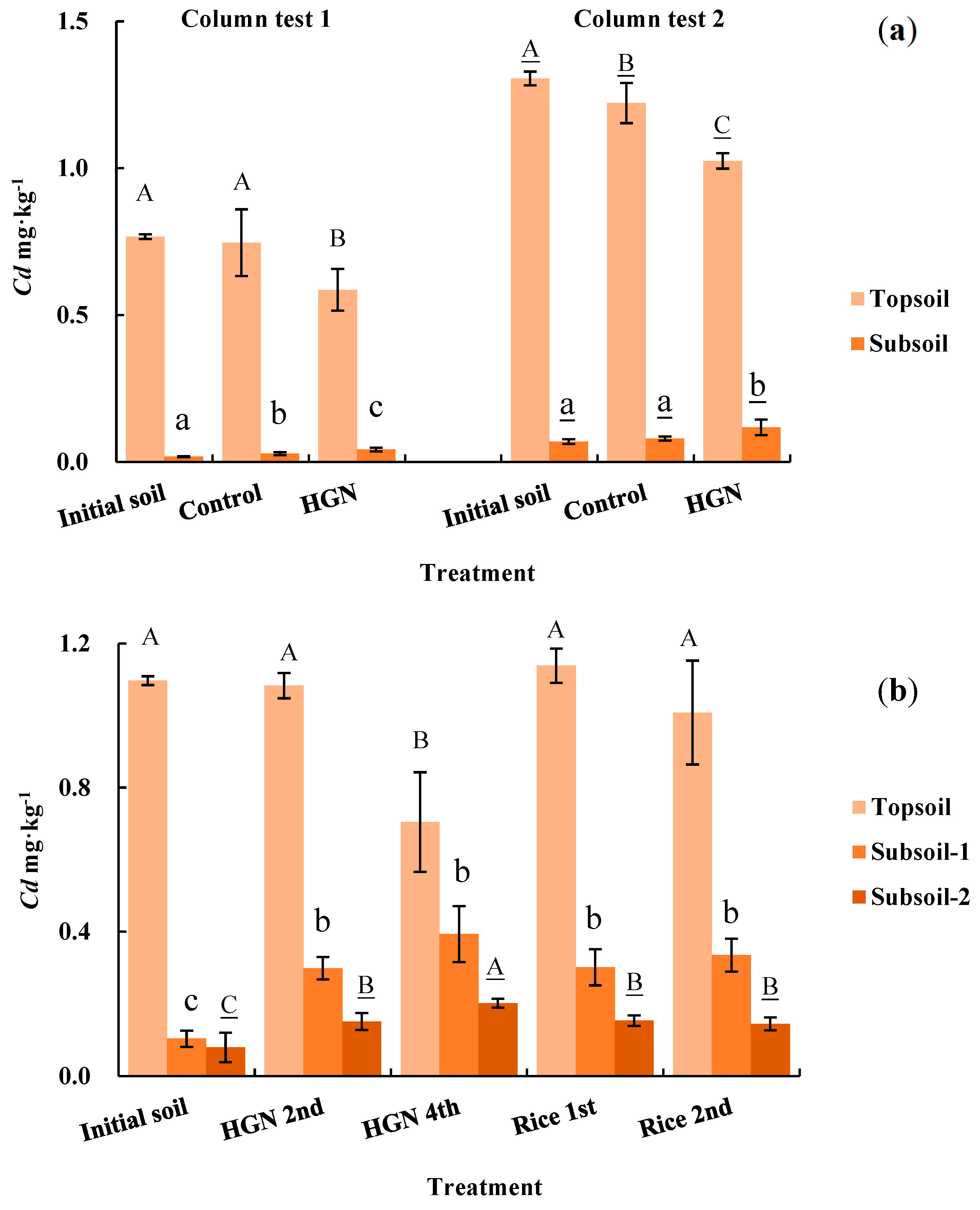
Figure 4.
Contribution of different pathways to Cd removal from the topsoil. (a): column test (inner ring: test 1, outer ring: test 2); (b): lysimeter test.
Figure 4.
Contribution of different pathways to Cd removal from the topsoil. (a): column test (inner ring: test 1, outer ring: test 2); (b): lysimeter test.
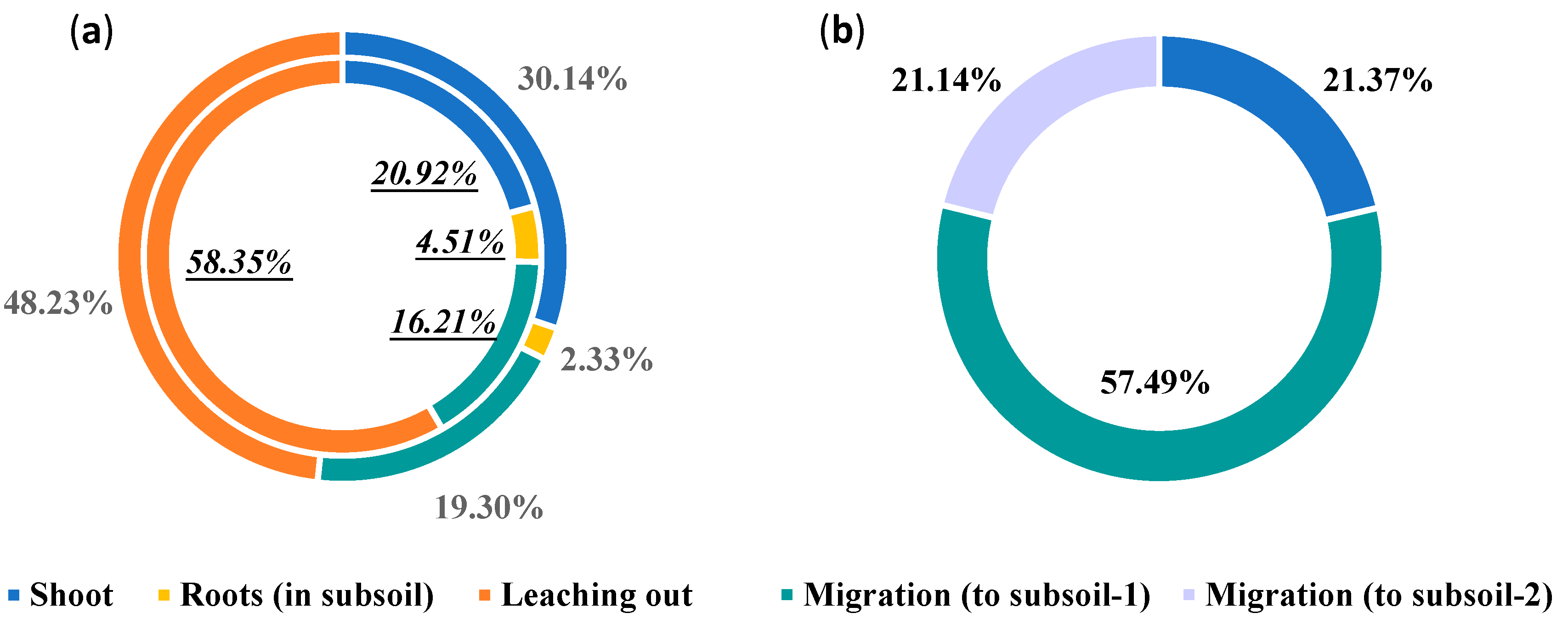

Table 1.
Plant growth and biomass of Pennisetum hybridum in column test.
| Root length (cm) | Height (cm) | Biomass (DW, /g·plant–1) | ||||
|---|---|---|---|---|---|---|
| Root (in topsoil) | Root (in supsoil-1) | Stem | Leaf | |||
| Test 1 | 57.67±2.08 | 174.00±4.58 | 16.84±1.09 | 9.60±1.11 | 44.36±2.52 | 42.27±7.10 |
| Test 2 | 51.67±4.51 | 190.00±8.66 | 17.91±1.94 | 11.59±0.90 | 95.51±2.25 | 25.96±3.00 |
Table 2.
Plant growth and biomass in lysimeter test.
| Height (cm) | Biomass (DW, g·plant –1) | Yield (DW, 104 kg·ha–1) | |||
|---|---|---|---|---|---|
| Pennisetum hybridum | Stem | Leaf | Stem | Leaf | |
| First harvest | 312.50±5.45 | 856.95±92.15 | 332.86±36.98 | 3.430 | 1.332 |
| Second harvest | 267.08±12.14 | 919.83±102.2 | 555.17±62.00 | 3.681 | 2.222 |
| Third harvest | 259.17±15.28 | 625.15±47.32 | 478.89±36.37 | 2.502 | 1.917 |
| Fourth harvest | 265.00±10.00 | 721.52±59.47 | 653.39±54.68 | 2.888 | 2.615 |
| Rice | Straw | Grain | Straw | Grain | |
| Upland rice | 130.00±6.00 | 31.73±1.69 | 43.29±1.95 | 0.5080 | 0.6930 |
| Simiao rice | 120.00±5.50 | 22.96±3.97 | 15.03±1.48 | 0.3675 | 0.2406 |
Table 3.
Bioconcentration factors (BCF) and translocation factors (TF) of Cd for different parts of Pennisetum hybridum.
Table 3.
Bioconcentration factors (BCF) and translocation factors (TF) of Cd for different parts of Pennisetum hybridum.
| BCF | TF | ||||||
| Root (in topsoil) | Root (in subsoil-1) | Stem | Leaf | Stem | Leaf | ||
| Column test 1 | 2.24 | 0.68 | 1.25 | 0.65 | 0.56 | 0.29 | |
| Column test 2 | 2.74 | 0.76 | 0.91 | 0.86 | 0.33 | 0.31 | |
| Lysimeter test | 3.54 | 1.89 | 0.95 | 1.82 | 0.27 | 0.51 | |
Table 4.
Extraction amount and efficiency of soil Cd by Pennisetum hybridum in column test.
| Extraction amount (mg·plant–1) | Extraction efficiency(%) | |||||||
| Root (in topsoil) | Root (in subsoil-1) | Stem | Leaf | Total | Roots | Shoots | ||
| Test 1 | 0.0567 ±0.0044 | 0.0138 ±0.0021 | 0.0428±0.0043 | 0.0212±0.0060 | 0.1345 | 4.60 | 4.17 | |
| Test 2 | 0.0642±0.0086 | 0.0116±0.0008 | 0.1136±0.0022 | 0.0363±0.0034 | 0.2256 | 2.90 | 5.74 | |
Table 5.
Amount and efficiency of Cd extraction by Pennisetum hybridum or rice in lysimeter test.
| Extraction amount (mg·m–2) | Extraction efficiency (%) | |||||
|---|---|---|---|---|---|---|
| Pennisetum hybridum | Stem | Leaf | Each harvest | Annual | ||
| First harvest | 3.153±0.085 | 1.564±0.042 | 4.717 | 23.40 | 8.53 | |
| Second harvest | 3.850±0.107 | 4.438±0.123 | 8.288 | |||
| Third harvest | 2.216±0.041 | 2.641±0.049 | 4.857 | |||
| Fourth harvest | 2.207±0.046 | 3.334±0.069 | 5.541 | |||
| Rice | Straw | Grain | ||||
| Upland rice | 2.006±0.107 | 0.913±0.041 | 2.919 | 3.02 | 1.10 | |
| Simiao rice | 0.071±0.012 | 0.029±0.003 | 0.100 | |||
Table 6.
Variation of soil pH after planting Pennisetum hybridum in column test.
| Test 1 | Test 2 | ||||||
|---|---|---|---|---|---|---|---|
| Initial value | Control | P. hybridum | Initial value | Control | P. hybridum | ||
| Topsoil | 5.57±0.03 A | 5.20±0.27 B | 4.87±0.05 C | 5.27±0.01 a | 5.14±0.01 b | 4.81±0.01 b | |
| Subsoil-1 | 4.47±0.03 A | 5.20±0.28 A | 3.76±0.12 B | 6.27±0.04 a | 6.42±0.02 a | 5.86±0.19 b | |
Means in the same row followed by the same letter were not significantly different according to Duncan’s multiple range tests (α=0.05).
Table 7.
Variation of soil pH after planting in lysimeter test.
| Initial value | Pennisetum hybridum | Rice | ||||
|---|---|---|---|---|---|---|
| Second harvest | Fourth harvest | Upland rice | Simiao rice | |||
| Topsoil | 5.58±0.11 B | 5.06±0.14 C | 4.37±0.23 D | 5.47±0.10 B | 5.95±0.31 A | |
| Subsoil-1 | 5.59±0.25 A | 5.10±0.03 C | 4.76±0.17 D | 5.45±0.03 AB | 5.30±0.14 BC | |
Means in the same row followed by the same letter were not significantly different according to Duncan’s multiple range tests (α=0.05).
Table 8.
Topsoil Cd concentration and removal rate.
| Cd concentration (mg·kg–1) | Cd removal rate (%) | ||
|---|---|---|---|
| Initial | After planting | ||
| Column test 1 | 0.7676±0.0077 | 0.5863±0.0429 | 23.62 |
| Column test 2 | 1.3058±0.0232 | 1.0250±0.0265 | 21.50 |
| Lysimeter test | 1.0970±0.0123 | 0.7041±0.1385 | 35.81 |
Table 9.
Cd changes in different parts of column test with and without plants (mg).
| Shoots | Roots (in topsoil) |
Roots (in subsoil) |
Topsoil reduction | Subsoil increase | Leaching out | |
|---|---|---|---|---|---|---|
| Planted test 1 | 0.0640 | 0.0567 | 0.0138 | 0.3626 | 0.0496 | 0.1785 |
| Planted test 2 | 0.1499 | 0.0642 | 0.0116 | 0.5616 | 0.0960 | 0.2399 |
| No plant test 1 | - | - | - | 0.0418 | 0.0209 | 0.0209 |
| No plant test 2 | - | - | - | 0.1677 | 0.0207 | 0.1470 |
Table 10.
Chemical properties and Cd contents of the tested soil.
| Column test 1 | Column test 2 | Lysimeter test | ||||||
|---|---|---|---|---|---|---|---|---|
| Topsoil | Subsoil | Topsoil | Subsoil | Topsoil | Subsoil | |||
| pH | 5.57±0.09 | 4.47±0.04 | 5.27±0.01 | 6.27±0.04 | 4.95±0.05 | 4.86±0.11 | ||
| OM g·kg–1 | 34.42±0.08 | 5.60±0.02 | 39.94±0.17 | 10.51±0.30 | 39.53±4.43 | 16.10±2.46 | ||
| TN g·kg–1 | 1.96±0.10 | 0.29±0.02 | 1.27±0.12 | 0.62±0.06 | 2.24±0.03 | 0.89±0.11 | ||
| TP g·kg–1 | 0.75±0.08 | 0.26±0.02 | 0.44±0.02 | 0.16±0.02 | 0.47±0.04 | 0.24±0.02 | ||
| TK g·kg–1 | 7.59±0.19 | 3.29±0.62 | 9.56±0.15 | 8.34±0.30 | 10.43±0.13 | 12.63±0.20 | ||
| Cd mg·kg–1 | 0.7676±0.0077 | 0.0181±0.0019 | 1.3058±0.0232 | 0.0698±0.0078 | 1.0236±0.0294 | 0.1472±0.0298 | ||
Note: OM, organic matter; TN, total nitrogen; TP, total phosphorus; TK, total potassium.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



