
Submitted:
14 March 2023
Posted:
23 March 2023
You are already at the latest version
Abstract
The more than thirty spacetime theorems developed over the past five decades establish that the universe and its spacetime dimensions have emerged from a cause/Causal Agent beyond the cosmos. Thus, to infer that this cause/Causal Agent may have intervened in the origin and history of Earth and Earth’s life resides well within the bounds of reason. Meanwhile, proponents of each of the three prevailing naturalistic models for the origin and history of Earth’s life have marshalled arguments and evidence that effectively undermine and refute the other two models. A biblical perspective and approach to Earth’s life can help resolve this impasse. While a superficial and pervasive appeal to divine intervention thwarts scientific advance, so does a rigid adherence to naturalism. A productive way forward is to identify which models (or parts of models), whether naturalistic, theistic, or a combination, most effectively narrow, rather than widen, knowledge gaps, minimize anomalies, offer the most comprehensive and detailed explanation of the data, and prove most successful in predicting scientific discoveries.
Keywords:
spacetime theorems
; naturalism
; abiogenesis
; panspermia
; process structuralism
; Cambrian explosion
; amino acids
; homochirality
; hand of God dilemma
Introduction
Naturalism is the belief that the laws of physics governing the universe and natural processes are fully sufficient to explain all that exists in the universe. Its proponents claim that it can explain the origin, dynamics, operations, and evolution of all the universe’s components, including all forms of life. In other words, physics and chemistry alone explain everything we can detect and measure in the universe.
For the past century, naturalism has dominated scientific research on the origin and history of Earth’s life. Such near complete dominance is evident in the leading scientific journals on the origin and history of life. Examples include Origin of Life and Evolution of the Biosphere, Current Biology, Astrobiology, Proceedings of the National Academy of Sciences USA, Science, and Nature.
The success of a naturalistic approach to scientific research is undeniable. Much of what scientists observe in the natural realm can, indeed, be explained from a strictly naturalistic perspective. However, does it follow from this success that naturalism explains everything we observe in nature? In light of the laws of thermodynamics, gravity, and electromagnetism, which imply that the universe and everything in it is proceeding toward ever-increasing disorder and decay, is it reasonable to assert that life comes from non-life and that simple, primitive life inexorably progresses to more complex and advanced life? Is it reasonable to conclude that organisms possessing mind, will, and emotions naturalistically arise from life forms that lack mind, will, and emotions? Is it realistic that intelligent life capable of launching and sustaining high-technology civilization arose through strictly natural means from species of life that possessed no innate intelligence?
Spacetime Theorems
In 1970, physicists Stephen Hawking and Roger Penrose published the first of the cosmic space-time theorems (Hawking and Penrose 1970). Based on only two primary assumptions, Hawking and Penrose demonstrated that the cosmic beginning is not just a beginning of matter and energy but also the beginning of space and time as well. That is, space and time began to exist when the universe began to exist.
The first assumption, that the universe contains mass, is undisputed. The second assumption, that general relativity reliably describes the dynamics, or movements, of massive bodies in the universe, has since been confirmed by astronomical observations to better than a trillionth of a percent precision (Penrose 1994; Weisberg and Huang 2016; Hector 2021). Today, general relativity ranks as the most exhaustively tested and firmly verified principle in all of physics (Ross 2018; Do et al. 2019; Kramer et al. 2021).
In 1994–2003, theoretical physicists Arvind Borde and Alexander Vilenkin published five extensions of the space-time theorems, culminating in what is now known as the Borde-Guth-Vilenkin theorem (Borde, Guth, and Vilenkin 2003). These extensions demonstrate that, regardless of the homogeneity, isotropy, and energy conditions of the universe, the universe must be subject to the relentless grip of an initial space-time singularity. Any cosmic model in which the universe obeys an average past expansion (a necessary requirement, as pointed by Borde and Vilenkin, for physical life to possibly exist in the universe) must be traceable back within finite time to an actual beginning of space and time.
According to physicist Sean Carroll (Carroll 2008; Carroll 2010), previous to 10-43 seconds after the cosmic origin event (the quantum gravity era where quantum mechanics competes with gravity in determining the dynamics of the universe), a possibility exists that quantum spacetime fluctuations could have been large enough to permit an escape from of an initial space-time singularity. However, quantum spacetime fluctuations during this first tiny split second of the universe’s 13.8-billion-year existence would accumulate, that is become “frothier,” over long pathways through space. Such an accumulation would blur the images of distantly observed quasars and blazars, especially at short wavelengths. The conclusion from observations on distant quasars is that “all the main QG [quantum gravity] scenarios [proposed to escape a spacetime beginning] are excluded” (Tamburini et al 2011). Without exception, all observations relevant to the quantum gravity era sustain a spacetime beginning to the universe (Perlman 2015; Romoli 2017; Acciari et al 2020; Bartlett 2021; Bolmont et al 2022).
Given that space and time came into existence at the cosmic origin event, we know something about where the universe has come from. It must have arisen not from within spacetime but, rather, from an “entity” or “reality” beyond space and time. In coming to this conclusion, astrophysical researchers have echoed the biblical explanation for the origin of the universe and of spacetime. As Hebrews 11:3 declares, “We understand that the universe was formed at God’s command, so that what is seen [detected] was not made out of what was visible [detectable].” According to 2 Timothy 1:9 and Titus 1:2, time has a beginning and God was active “before the beginning of time.” Physicists likely would have produced the spacetime theorems sooner had they started with the biblical assertion that time has a beginning.
The spacetime theorems establish that the Bible cannot be dismissed as scientifically irrelevant. Nor can scientists honestly assert that supernatural events never happen. Demonstrating that the universe—all that scientists can detect and measure—came into existence through the agency of a Cause beyond space and time is the greatest supernatural event that any scientist can hope to uncover.
The Bible on the Origin and History of Life
In addition to crediting God with the origin of the universe, the Bible also points to God as the creator of Earth’s life. Psalm 104:24 declares, “How countless are your works, Lord! In wisdom you have made them all; the earth is full of your creatures.” Genesis 1 uses the Hebrew verbs bara (create) and asa (make) to describe God’s action in bringing into existence different life forms. Genesis 1 also uses the Hebrew verbs haya (let there be) and sharas (teem) with respect to other life forms.
These biblical texts and others imply that God is responsible for all life on Earth. He brings different life forms on Earth throughout the history of life on Earth via supernatural acts that transcend the laws of physics (bara), as well as supernatural acts performed within the laws of physics (asa) and actions occurring through guided or unguided natural processes and/or through supernatural interventions (haya, sharas).
From a biblical perspective, therefore, research on the origin and history of Earth’s life should not be closed to consideration of only natural causation or only supernatural causation. It should be open to both. It should also develop tests to determine which causation sources are most likely and/or best able to explain all the observational and experimental data. The Bible predicts that strictly naturalistic models for the origin and history of Earth’s life will fail to explain all the data. The Bible implies that the science of the origin and history of life will proceed more efficiently, generate more successful outcomes, and demonstrate greater explanatory and predictive power if both natural and supernatural causations are taken into account.
Origin of Life
The simplest known non-parasitic life form is the bacterium Pelagibacter ubique. Its genome consists of about 1,100 gene products (Giovannoni et al. 2005). (A gene product refers to proteins and functional RNAs, such as ribosomal and transfer RNA.) This bacterium’s genome is at the theoretical minimum size for an independently existing life form capable of long-term survival and reproduction (Itaya 1995; Maniloff 1996; Leslie 2021;).
Proteins and functional RNAs are complex. Life-essential proteins range in size from 200–2,000 amino acids. Transfer RNAs, messenger RNAs, and ribosomal RNAs are all life-essential. Transfer RNAs (about 15% of the total RNA in cells) contain between 75–95 nucleotides. Messenger RNAs (about 5% of the total RNA in cells) contain 400–12,000 nucleotides. Ribosomal RNAs (about 80% of the total RNA in cells) contain 1,500–5,000 nucleotides. Single cells contain between 50,000 and 3 billion proteins and RNAs (Milo 2013; Dolgalev et al. 2023).
Life, even in its simplest form, therefore, is intricate and complex to a mind-boggling degree. This intricacy and complexity pose a profound problem for naturalistic origin-of-life models. The chance of a living cell forming through random, repeated shufflings of amino acids, nucleotides, lipids, and sugars is exceedingly remote, to a point that is indistinguishable from zero. The chance that all the required amino acids, nucleotides, lipids, and sugars would be available in one place, with no contaminants, and where random shufflings can occur appears just as remote and indistinguishable from zero.
These improbabilities have reduced the number of naturalistic origin-of-life models still seriously considered viable by origin-of-life researchers to just three. These three include (1) abiogenesis, (2) panspermia, and (3) directed panspermia. Hundreds of scientists, tens of millions of research dollars, and several decades of concerted research effort have been dedicated to demonstrating the scientific feasibility of each. All these dedicated resources have yielded progressively more evidence to suggest that science is best and most successfully done if both natural and supernatural causes are considered and tested.
Abiogenesis Model
The central dogma of origin-of-life research is abiogenesis, the view that sometime in the past, favorable natural conditions occurred on Earth for the formation of complex organic molecules—the twenty bioactive amino acids, the five bioactive nucleobases, the two ribose sugars, and three categories of lipids—and that, in turn, these gave rise to the self-organization of proteins, DNA, RNA, and lipid membranes. Abiogenesis goes on to propose a naturalistic pathway whereby the proteins, DNA, RNA, and lipid membranes self-assemble into primitive living cells. The central dogma of biological evolution asserts that all life has naturalistically evolved from the first primitive living cells.
However, the abiogenesis model appears fraught with intractable problems. A sampling of just six is described, briefly, here:
Missing amino acids: The twenty bioactive amino acids fall into four categories: nonpolar, polar uncharged, acidic (negatively charged), and basic (positively charged). The three basic (positively charged) bioactive amino acids are arginine, histidine, and lysine. Laboratory prebiotic synthesis experiments at both normal temperatures (0–200° Celsius) and high temperatures (200–700° Celsius) failed to produce any arginine or lysine. Reanalysis of products from normal temperature Miller-Urey-type spark discharge experiments detected neither arginine nor lysine (Johnson 2008). Prebiotic hydrothermal synthesis experiments yielded no detectible level of amino acid production, where it would have been clearly discernible (Hennet, Holm, and Engel 1992; Aubrey, Cleaves, and Bada 2009).
The failure of any prebiotic synthesis experiment to produce either lysine or arginine under any remotely conceivable early Earth conditions led some astronomers to propose that comets, asteroids, and/or meteoroids and impact-shock synthesis brought these life-essential amino acids to Earth’s surface (Chyba and Sagan 1992). However, the only basic amino acid reportedly detected in any meteorite is lysine, and it was identified only tentatively in just one chondrite (Kotra et al. 1979). The chondrites with the highest levels of amino acids, EET92042, GRA95229, and GRO 95577, contained no measurable lysine or arginine (Martins 2007). In the words of two origin-of-life chemists, “a general consensus seems to have emerged that lysine and arginine are ‘prebiotically implausible’” (McDonald and Storrie-Lombardi 2010).
Given the absence of lysine and arginine on the early Earth, biochemists sought to determine what biochemical functions can and cannot be achieved without these amino acids (McDonald and Storrie-Lombardi 2010). They discovered that while the lack of lysine and arginine did not halt the formation of proteins, it did prevent the formation of proteins that take any role in protein-nucleic acid biochemistry. Given that arginine-and-lysine-containing proteins are essential for crucial protein-DNA interactions, naturalistic explanations for life’s origin, on this basis alone, appear ruled out.
Homochirality: Amino acids cannot be linked together to make proteins in living organisms (in virtually cases) without catastrophic consequences unless all the amino acids have the same “handed” configuration, in the case of known life, left-handed (Banreti et al. 2022). Neither can nucleosides be linked together in living organisms (in all cases) to make DNA and RNA unless they are connected by pentose sugars all with the same right-handed configurations. Outside of organisms, amino acids and pentose sugars exist in a racemic mixture, that is, random mixtures of left-handed and right-handed configuration.
No natural source exists on Earth for producing the homochiral molecules required for life’s emergence. Organic chemist William Bonner writes, “I spent 25 years looking for terrestrial mechanisms for homochirality and trying to investigate them and didn’t find any supporting evidence. Terrestrial explanations are impotent or nonviable” (Bonner 1995).
The only possible astronomical sources that could possibly drive a racemic mixture of amino acids toward a slight left-handed inclination are intense circularly polarized ultraviolet radiation (CPUR) emitted by either neutron stars or black holes and dense neutrino fluxes blasted from supernovae or neutron stars or Wolf-Rayet stars (high-mass helium-burning stars with surface temperatures ranging from 20,000 K to 210,000 K) in close orbits about massive stars. For both these radiation sources, the departure from a racemic mixture takes place through right-handed amino acids being destroyed at a faster rate than the destruction of left-handed amino acids.
Laboratory simulations show that, at best, CPUR from neutron stars or black holes would generate only a 1.34% excess of left-handed (relative to right-handed) amino acids (Takano et al. 2007; de Marcellus et al. 2011). Other laboratory simulation experiments demonstrate that, at best, dense neutrino fluxes from supernovae or neutron stars and Wolf-Rayet stars closely orbiting massive stars would produce only a 1% excess of left-handed (Boyd et al. 2018).
According to the Kuhn-Condon rule of quantum mechanics (Kuhn 1930; Condon 1937), CPUR destroys more left-handed than right-handed amino acids only if the CPUR is strictly monochromatic. Astronomers know of no astrophysical source of monochromatic CPUR.
In the Milky Way Galaxy, Wolf-Rayet stars and neutron stars in close orbits about massive stars are rare (Boyd et al. 2018). Thus, they would be unable to populate our galaxy with a sufficient number of meteoroids containing amino acids with a slight preference for left-handed configurations.
Chemists in sophisticated laboratory experiments have been successful in amplifying an original tiny excess of left-handed chiral molecules that are closely analogous to bioactive amino acids (Shibata et al. 1998). However, any higher excess thereby achieved becomes unstable over time (Frank 1953). Furthermore, the greater the amplification of excess, the lower the remaining quantity of the original amino acid sample (Flores, Bonner, and Massey 1977). All laboratory experiments are consistent with the conclusion that long before a sample of amino acids becomes 100 percent left-handed, the entire original sample is destroyed.
Insufficient time: In a short graduate astronomy course at the University of Toronto in 1970, astronomer Carl Sagan set forth three minimal requirements for abiogenesis:
- A vast “soup” of prebiotic molecules, at least as vast as all Earth’s oceans;
- A rich abundance of exclusively homochiral (100% left-handed or 100% right-handed) prebiotic molecules; and
- A very long time, at least a billion years.
Researchers now shows that the time window for life’s origin on Earth was much briefer than a billion years. Until 4.0 billion years ago, the Sun showered Earth in deadly radiation. Earth’s surface was, at least episodically, hellishly hot until 3.84 billion years ago. Stable liquid water oceans and stable rocky land masses did not appear on Earth’s surface until 3.84–3.83 billion years ago.
When living organisms extract inorganic material from their environment, they preferentially choose the lighter isotopes of that material. For example, they choose carbon-12 at the expense of carbon-13, nitrogen-14 at the expense of nitrogen-15, and sulfur-32 at the expense of sulfur-34. The earliest undisputed multiple isotope evidence for life on Earth dates back to 3.825 ± 0.006 billion years ago (Manning, Mojzsis, and Harrison 2006). Ancient zircons discovered in Western Australia (Bell et al. 2015) and northern Canada (Dodd et al. 2017) reveal brief episodes 4.1–4.0 billion years ago when liquid water, solid rocks, and the possible isotope signatures of life existed. Evolutionary biologist Niles Eldredge, comments on this evidence: “One of the most arresting facts I have ever learned is that life goes back as far in Earth history as we can possibly trace it. …[I]n the very oldest rocks that stand a chance of showing signs of life, we find those signs” (Eldredge 2000).
The moment physical and chemical conditions permit life’s existence on Earth, life appears. It appears not in just one form, as naturalistic models would predict. Isotope evidence reveals the existence of a diversity of microbial species at the time of life’s origin (Schidlowski 2001; Garcia, Cavanaugh, and Kacar 2021).
Missing soup: The significance of evidence for multiple isotopes indicative of life in Earth’s oldest rocks is this: despite the absence of a primordial prebiotic soup, life existed abundantly on Earth previous to 3.8 billion years ago. For many decades, scientists have scoured Earth’s crust and oceans in a quest for evidence of prebiotics. Extensive research reveals none. Isotope ratios of carbonaceous molecules in Earth’s oldest rocks show evidence of postbiotics but not of prebiotics (Schidlowski 1998; Rosing 1999; Grassineau et al. 2005). If the latter ever existed, they must have been in such low abundance as to be of no use for abiogenesis.
Scientists now understand why Earth never had a prebiotic soup. The explanation comes from the “oxygen-ultraviolet paradox.” If oxygen were present in the early Earth’s atmosphere or oceans, even at a very low abundance level, that oxygen would have prevented any prebiotic chemistry from functioning. However, if there were no oxygen at all in Earth’s atmosphere and ocean, ultraviolet radiation from the early Sun would have flowed to Earth’s surface unimpeded and, in this case, too, halted prebiotic chemistry. For the early Earth, radiolysis of water by uranium, thorium, and plutonium isotopes produced sufficient oxygen to prevent what we know as prebiotic chemistry from occurring (Draganic 2005).
With Earth essentially ruled out as a site for prebiotic chemistry, researchers committed to considering only naturalistic answers began to look to outer space. In several meteorites, they found eight of the twenty bioactive amino acids, though at abundance levels of only a few parts per million (Cronin and Pizzarello 1983; Burton et al. 2012; Lymer et al. 2021). In one comet, researchers discovered the simplest amino acid, glycine, but at less than one part per billion (Elsila, Glavin, and Dworkin 2009).
The original sources of these amino acids are dense interstellar molecular clouds. To quote Carl Sagan, “We are made of star-stuff” (Sagan 1973). Molecular clouds in the Milky Way Galaxy’s spiral arms and core possess a much higher abundance and diversity of carbonaceous molecules than any other known astrophysical sources. Astronomers have discovered over 140 carbonaceous molecules in interstellar molecular clouds so far. They have yet to find, however, any amino acids, nucleobases, or ribose sugars—critical building block molecules for proteins, DNA, and RNA (Kuan et al. 2003; Snyder et al. 2005). While chemical conditions within the densest and largest interstellar molecular clouds permit limited production of nucleobases and simple amino acids, these same chemical conditions operate to destroy most of the nucleobases and amino acids produced. The remaining amounts likely fall below a few parts per billion, a quantity insufficient to support any conceivable naturalistic origin-of-life scenario.
Life as we know it? Recognizing the many intractable barriers confronting naturalistic abiogenesis models, some origin-of-life researchers began to speculate about the existence of alternatives to life as we know it, that is, life that is not carbon-based in its chemistry. The only elements in the periodic table other than carbon on which complex molecules might conceivably be based would be arsenic, boron, and silicon.
Arsenic and boron are rare, however. Compared to carbon both are more than 5,000 times less abundant in the solar system (Arnett 1996a). When concentrated, both elements prove poisonous, deadly to life. Silicon is only 89 times less abundant than carbon in the solar system (Arnett 1996b). However, while carbon easily forms double and triple bonds, silicon rarely does. Because of silicon’s very high affinity to oxygen, polymers of silicon will be built on Si–O chains rather than on Si–Si chains. Furthermore, as a MIT research team demonstrated, “The vast potential theoretical space of silicon chemistry is almost entirely unstable in water, and hence not available to a biochemistry based on water as a solvent” (Petkowski, Bains, and Seager 2020). Of all the elements in the periodic table, only carbon allows for the complexity and stability of chemical bonding that life molecules require. Any physical life in the universe must be virtually the same as “life as we know it.” It must be carbon-based.
Hand of God dilemma:
Origin-of-life research chemists have achieved amazing outcomes in laboratory experiments that attempt to demonstrate how the components of life molecules potentially could be assembled. Their greatest success is the joining together of bioactive amino acids to construct short protein segments. However, for this success to be possible, the experimenters had to repeatedly intervene. They discovered that each chemical step needed a specific chemical environment and set of physical conditions. Nearly always, a subtraction reaction must occur simultaneously with an addition reaction, and both reactions must occur at specified rates.
In living cells, biochemical synthesis occurs through catalyzed reactions among different enzymes, each enzyme requiring a distinct, specified microenvironment at its active site for the reaction to run. In simulating an enzyme-free prebiotic scenario, experimenters have found that they must employ multiple, specifically ordered chemical steps that involve precipitation, crystallization, purification, and drastic changes in the chemical conditions to go from one synthesis step to the next. Even then, success is rarely achieved.
Origin-of-life chemist Clemens Richert, in a Nature Communications article, explains that the reputed goal of experimental origin-of-life biochemists is to re-enact what may have occurred when life arose from abiotic matter on the early Earth (Richert 2018a). Richert noted, however, that the most successful origin-of-life experiments in the laboratory required numerous cycles of hydration and dehydration and/or cooling and heating. To be productive, these experiments necessitated repeated transitions from arctic to volcanic conditions in a single location then back to arctic again within only a few hours or days—an unrealistic natural scenario to say the least.
Richert has referred to these required experimenter interventions “the hand of God dilemma” (Richert 2018b). He recommended that in their publications, origin-of-life research chemists should state as accurately as possible how many times and exactly when and where in their experiments they commit the Hand of God dilemma. His recommendation would help researchers in other disciplines, and especially the lay public, understand what has and has not been determined by origin-of-life laboratory experiments.
Experimenter intervention, Richert claims, is equivalent to asserting that God did it. In his paper Richert acknowledges that, “yes, most of us [origin-of-life research chemists] are not comfortable with the idea of divine intervention in this context” (Richert 2018c). He and his peers may be uncomfortable with their finding, but what they have demonstrated in their experiments, nonetheless, is that an Agent much more knowledgeable, intelligent, and capable than they must have assembled life on the early Earth.
Panspermia Models
The manifold failures of the abiogenesis model partly explain why since the beginning of the twenty-first century origin-of-life conferences and research endeavors now seem dominated by astronomers and astrobiologists. The current thinking of scientists committed to naturalism is that life’s origin must have occurred on some extra-terrestrial site and later been transported to Earth’s surface. Here are the two main panspermia scenarios put forward, interplanetary and interstellar:
Interplanetary panspermia: In the 1990s and first decade of the twenty-first century, astrobiologists speculated that life may have originated on Mars and piggybacked on a meteorite traveling from Mars to Earth. In 1996, U.S. president Bill Clinton famously danced around a Martian meteorite reputed to contain unmistakable signatures of microbial life. Later, however, that bold claim was refuted (Maniloff et al. 1997; Bada 1998; Kazmierczak and Kempe 2003; Weiss 2004).
The piggybacking of life on meteorites certainly occurs, but the other way around. Meteorites have sent the fossil remains of Earth’s microbial life to nearly all the solar system’s planets and moons. While Earth’s geologic activity has made the fossils of Earth’s first life unrecognizable, millions of pristine fossils of Earth’s first life litter the landscape of the Moon, just waiting to be discovered (Armstrong, Wells, and Gonzalez 2002; Armstrong; Ross 2007; Armstrong 2010). Once collected and closely examined these fossilized remains of Earth’s earliest life could go a long way to determining how life originated and whether its origin most closely aligns with a naturalistic or supernatural origin. One would hope astrobiologists would be motivated to send spacecraft to the Moon to recover these fossils, given their potential to answer crucial questions of life’s origin, whichever direction the evidence may point.
All the conditions that stymie a naturalistic origin of life on Earth also exist on Mars. What is more, several other characteristics of Mars also serve to thwart a naturalistic origin of life. Mars’ atmosphere always has been thinner than Earth’s. Therefore, Mars’ surface has been exposed to more ultraviolet radiation. Mars’ soil/crust is more oxidizing and contains sixty times the concentration of sulfur and sulfur compounds. Mars’ rotation axis tilt varies chaotically. Carbonate formation when Mars was warm and wet would have rapidly and permanently removed carbon dioxide from the Martian atmosphere, which quickly made Mars cold and dry (Carr 2000).
Interstellar panspermia: Conditions for a naturalistic origin of life on solar system bodies other than on Earth or Mars prove even more intractable. This recognition has turned naturalists’ attention toward consideration of interstellar panspermia. Adherents to this scenario have proposed several modes for transportation of the seeds of life from far-off regions of the cosmos to Earth:
- 1.
- radiation pressure
Nobel Prize-winning chemist Svante Arrhenius developed the first scientific model for interstellar panspermia in 1907 (Arrhenius 1908). He reasoned that Earth was too young for life to have arisen here naturalistically. Therefore, he proposed that life arose over much more time on planets orbiting stars outside the solar system. He presumed that “spores” escaped from these planets and that interstellar radiation pressure wafted them to Earth. Astronomers later discovered that the required radiation pressure would have destroyed any such spores (Zagorski 2007).
- 2.
- dust and meteoroids
The most popular candidates offered for transporting the seeds of life to Earth across the vast reaches of space have been interstellar dust and meteoroids. However, interstellar dust and meteoroids cannot protect microbes, genes, proteins, or life-essential amino acids and nucleotides from the deadly dangers of interstellar space. A dust grain massive enough to safely carry a microbe or spore across interstellar space would require propulsion by starlight at an intensity sufficient to destroy the microbe or spore and all its proteins, DNA, and RNA. The probability of an interstellar meteoroid of planetary origin large enough to protect a dormant microbe or spore during its trip to and landing upon Earth would be no greater than one chance in a hundred thousand over the entire history of Earth (Melosh 2003). For both dust grains and meteoroids, interstellar travel times range from the millions to tens of millions of years, a time scale much longer than the half-lives of the proteins, DNA, and RNA crucial for maintaining the viability of any such microbe or spore (Levy and Miller 1998; Larralde, Robertson, and Miller. 1995).
- 3.
- comets
In 1981, astronomers Fred Hoyle and Chandra Wickramasinghe updated Arrhenius’ model in their book Evolution from Space (Hoyle and Wickramasinghe 1981). They proposed that Earth was seeded with life by comets from other planetary systems.
In 2018, thirty-three astrobiologists extended the Hoyle-Wickramasinghe model to propose that comets 0.54 billion years ago seeded Earth with diverse genes, explaining the Cambrian explosion event, when 50+ phyla of life suddenly appeared (Steele et al. 2018a). As recently as 2020, three astrobiologists further developed the Hoyle-Wickramasinghe model, claiming that interstellar comets seeded Earth with a community of diverse microbes 3.8 billion years ago (Wickramasinghe, Wickramasinghe, and Steele 2020).
Astronomers estimate that about 20% of all stars in the Milky Way Galaxy possess comet belts. When stars experience close encounters with one another, they occasionally exchange comets. Such exchanges could potentially cut interstellar travel times from millions of years down to tens of thousands, conceivably even just thousands of years.
Nevertheless, for multiple reasons, comets make poor candidates for the transport of life or life’s genes across interstellar space. First, the comets that are exchanged during stellar encounters are those orbiting distantly from their host stars. Such comets have an extremely remote probability of ever being close enough to a presumably “inhabited” planet to capture any of that planet’s life or genetic material.
Second, any comet captured from another star has an extremely remote probability of making a close enough flyby (not a collision) with Earth, in less than a hundred thousand years, to safely deposit any microbes or genes on Earth. Third, even with a planet as heavily populated with microbes and viruses as Earth, the density of such microbes and viruses at distances where any comet could conceivably capture these microbes or viruses is very low. The density is far too low for a sufficiently large comet to realistically capture, encase, and protect a microbe or virus so that it can survive an interstellar trip, even a relatively short one (Wainwright 2013).
Additional challenges to interstellar panspermia
An obvious problem with appeals to interstellar panspermia is that it simply shifts the burden of explaining the naturalistic origin of life from Earth to some other planet orbiting some other star. The thirty-three astrobiologists who attempted to get around this problem did so by depending on two speculations. First, they asserted that the spacetime theorems and the big bang origin model must be incorrect. Both, they declare, must be replaced by some kind of quasi-steady-state cosmic (QSSC) model (Hoyle, Burbidge, and Narlikar 2000; Narlikar, Vishwakarma, and Burbidge 2003; Narlikar, Burbidge, and Vishwakarma 2007). Such models would allow infinite time for life to evolve from prebiotic chemicals. Second, they assert as fact the claim that “hundreds of billions of habitable planets exist in the [Milky Way] galaxy alone” (Steele et al. 2018b).
These astrobiologists are to be commended for acknowledging that only through a denial of decades of research findings about the origin and history of the universe—as well as long-standing biblical statements (Ross 2023)—can a naturalistic model for the origin and history of Earth’s life be defended. However, as much as these astrobiologists dislike the big bang creation model and the spacetime theorems, the experimental and observational evidence sustaining them still stands strong in the face of thorough testing (Ross 2018). Likewise, the observational case against the QSSC model in all its modifications remains both overwhelming and pervasive.
According to the QSSC models, the spectral redshifts of quasars fail to establish their great distances. Until recently, this negative assertion could not be refuted by the direct trigonometric-distance-measuring methods used by surveyors to determine the distances and heights of buildings and mountains or by astronomers to determine the distances to nearby stars. However, by exploiting an intercontinental array of radio telescopes that yield the resolving power of an 8,000-kilometer diameter telescope, astronomers were able to achieve a direct distance measurement showing that the quasar 3C 279 must be at least 5.9 billion light years away (Homan and Wardle 2000). By use of this same technique, astronomers now confirm that the spectral redshifts of galaxies as far as 460 million light years away, do, indeed, accurately indicate their distances (Kuo et al. 2013).
If the QSSC models were accurately describing cosmic reality, at least some of the spectra of quasars should be blue-shifted. Astronomers observe only redshifts, and the quasar redshifts always are proportional to their distances. The QSSC models also contradict the cosmic microwave background radiation (CMBR) evidence showing that the big bang creation event occurred only 13.8 billion years ago. By contrast, the QSSC models assert that there is enough dust in the universe to explain CMBR maps without a relatively recent big bang. However, observations now establish that the quantity of intergalactic dust amounts to 100–125 times less than the minimum required by QSSC models (Thacker 2013).
Number of habitable planets in the Milky Way Galaxy
Two teams of astrobiologists have calculated that there could as many as 40 or 45.5 billion habitable planets in our galaxy (Guo et al. 2009; Petigura, Howard, and Marcy 2013), a number significantly shy of the QSSC’s hundreds of billions. These numbers shrink still further if various habitability criteria are applied. The estimate of 40 or 45.5 billion habitable planets took into account nothing more than the possibility of a planet’s orbiting a star in the zone where liquid water could conceivably exist on some part of the planet’s surface for a limited time— “the liquid water habitable zone.” It also included all host stars as candidates, regardless of their mass, age, and composition.
The liquid water planetary habitable zone was the first planetary habitable zone to be described by astronomers. Today, astronomers are aware of at least thirteen distinct planetary habitable zones (Ross 2016a, Ross 2019a, Ross 2019b, Green et al. 2020; Ross 2022a). For a planet to be truly habitable, it must simultaneously reside in all the known planetary habitable zones. Of the 5,342 planets that astronomers have discovered thus far and for which orbital features have been determined (exoplanet TEAM 2023), only one resides simultaneously in even three of the thirteen known planetary habitable zones—the same one that resides in all thirteen such zones—not to mention in all the known galactic and intergalactic habitable zones, as well (Lineweaver, Fenner, and Gibson. 2004; Spitoni, Gioannini, and Matteucci. 2017; Ross 2022b).
Directed Panspermia Model
In 1999, the conference of the International Society for the Study of the Origin of Life (ISSOL), a triennial gathering of 300+ researchers, ended with a dramatic summation, but not by one of the official ISSOL speakers. A conference participant stepped to the open microphone to offer his overview of the week’s presentations. He noted that during the first day of the conference, Earth had been eliminated as the location where life originated. During the second day, Mars was eliminated. During the third day, the remainder of solar system bodies were ruled out. During the fourth day, interstellar panspermia was tossed out. The only possible explanation remaining, he said, was that an intelligent civilization from another planetary system must have sent a spaceship to Earth 3.8 billion years ago for the purpose of planting life on Earth.
This research scholar explained that his “directed panspermia” proposal was not a new one, nor was it original to him. None other than Nobel laureate chemist Francis Crick and famed origin-of-life researcher Leslie Orgel (present at the conference) had made such a proposal in 1973 (Crick and Orgel. 1973). The directed panspermia model, said the speaker, held the advantage of explaining other events in the history of Earth’s life that have proven intractable for all naturalistic models, events such as the Avalon and Cambrian explosions and other mass speciation events that occurred startingly soon after mass extinction events. He went on to propose that intelligent aliens visited Earth not only once but, rather, must have returned to Earth multiple times to rescue Earth from total extinction and to ensure life’s steady progression from primitive to more advanced forms. With that comment, the man left the microphone and the conference was over.
The directed panspermia model he summarized clearly acknowledges the physical and chemical impossibility that life could have arisen on Earth apart from intelligent input. It frankly admits that the existence of life on Earth requires capable, intentional—thus personal—agency.
The model, however, suffers from several devastating flaws. First, it is physically impossible for intelligent aliens with advanced technology to have existed in the universe 3.8 billion years ago, or even 0.54 billion years ago. For such beings to exist and develop advanced technology, they would have had to be living on a planet with strong, enduring plate tectonics and a strong, enduring magnetosphere. They also would have needed to live on a planet orbiting a star virtually identical to the Sun, and that star would need to be 4.57 billion years old.
For these conditions to have been met, the host planet would have required an exceptionally high accretional heat. That accretional heat would need to have been as great as what Earth gained from its collision (during its infancy) with the solar system’s fifth rocky planet, Theia (Canup 2012; Wang and Jacobsen 2016; Steenstra et al. 2020). This interior planetary accretional heat by itself would not have been sufficient. It would have required precise augmentation by heat from the radioactive decay of a superabundant supply of thorium and uranium. To obtain this superabundance, the host planet would need to have formed when the cosmic supply of uranium and thorium peaked. That peak occurred when the universe was about 9 billion years old (see Figure 1).
Uranium and thorium are exclusively manufactured during supernova eruption events and merger events of neutron stars. As the universe expands and stars disperse, these events become progressively less frequent. Meanwhile, thorium-232, uranium-235, and uranium-238 decay with half-lives of 14.05, 0.704, and 4.468 billion years respectively. Consequently, the concentration of uranium and thorium builds up during the first several billion years of cosmic history when star formation rates are high. Later, when star formation rates decline, the radioactive decay of uranium and thorium supersedes the production of uranium and thorium from supernova eruptions and mergers of neutron stars. One reason why Earth has such an enormous concentration of uranium and thorium is that it formed when the cosmic abundance of uranium and thorium had peaked (Yungelson and Livio 2000; Kobayashi and Nomoto 2009; Spina et al. 2016; Tsujimoto 2023). diagram credit: Hugh Ross
Our star, the Sun, has the lowest flaring activity level and the greatest luminosity stability among all known stars (Maehara et al. 2012; Maehara et al. 2015; Reinhold et al. 2020; Zhang et al. 2020). However, during the Sun’s history this low flaring activity level and luminosity stability were lacking and will be lacking again in the future. Any star hosting a planet on which intelligent life has been able to launch and sustain high-technology civilization must be just like the Sun—with extreme luminosity stability and a very low level of flaring activity.
Figure 2 shows the Sun’s past and present flaring activity levels and its future flaring activity levels, as determined from observations of the most Sun-like stars spanning ages from 0.1 to 9.0 billion years. Only when the Sun is 4.57 billion years old is its flaring activity sufficiently low to permit the emergence of high-technology civilization.
The date for the peak cosmic abundance of uranium and thorium (plus 4.57 billion years) adds up to about 13.8 billion years. Therefore, while it is theoretically conceivable that other physical, extraterrestrial intelligent life could exist in the universe, that life could not predate us to any significant degree. It is wholly infeasible to suggest that physical intelligent aliens from another planetary system might have seeded life on Earth 3.8 billion years ago or seeded the Cambrian explosion 0.54 billion years ago.
As a second consideration, the laws of physics and the radiation, particles, gas, and dust in interstellar space present a huge challenge to the successful flight of spaceships across interstellar space. The damage to interstellar spacecraft from interstellar radiation, particles, gas, and dust increases with the square of the spacecraft’s velocity. It also increases with the square of spacecraft’s cross section. For these reasons, researchers intending to send spacecraft to the nearest planet outside our solar system, a planet only 4.25 light years away, recognize that spacecraft larger than 10 centimeters across and traveling at a fifth the velocity of light cannot survive the journey (Hoang et al. 2017). Thus, the latest plan calls for sending a thousand spacecraft, all much smaller than 10 centimeters across, at a tenth light’s velocity (Clery 2016). Even so, more than half the spacecraft will most likely be inoperable on arrival and the other half, only partly operable. Clearly, no living organism, seed, or spore could survive the trip.
A third fatal flaw for directed panspermia is that the home planet for extraterrestrial physical intelligent aliens is either impossibly distant or non-existent. Astronomers have yet to discover a star that is sufficiently identical to the Sun that it could be a candidate to host a planet on which intelligent physical beings can exist and develop a technologically advanced civilization. As noted already, the habitability requirements for a planet capable of hosting intelligent life are numerous. So far, everywhere astronomers have searched in our galaxy and the universe they see only conditions hostile to intelligent physical life. At best, a planet capable of hosting extraterrestrial intelligent beings who have successfully launched high-technology civilization must be so distant—more than 10,000 light years away—as to make a life-seeding mission to Earth exceedingly impractical.
A remaining major problem with the directed panspermia model is the origin of the extraterrestrial intelligent beings. Explaining the origin of an intelligent species bound by the laws of physics on some distant planet presents a challenge as big, if not bigger than, explaining life’s origin on Earth.
It appears, therefore, that the origin of life on Earth must be sought within Earth’s confines. It appears, too, that the cause or originator must be Someone who is not constrained by the laws of physics governing the universe or by the universe’s spacetime dimensions.
Process Structuralism
There is a growing acknowledgment among evolutionary biologists that natural selection, mutations, gene exchange, and epigenetics are inadequate to explains the fossil record and the history of Earth’s life. This recognition has given rise to a school of biological theory known as process, or biological, structuralism.
Process structuralists argue that there must exist one or more hidden natural physical or chemical forces in addition to natural selection, mutations, gene exchange, and epigenetics that shape and direct the development and the appearance of new species of plants and animals (Goodwin 2001; Morris 2003; Denton 2013). They also assert that these hidden forces play a more dominant role in the history of Earth’s life than natural selection, mutations, gene exchange, and epigenetics. These forces, they claim, embody self-organizing principles that impact biological development in a major way.
Process structuralists cite the Avalon and Cambrian explosions of life and the mass speciation events that quickly follow mass extinction events as evidence for their hypothesis. They point out that the four known naturalistic mechanisms for generating change in life forms all make relatively small stepwise changes. Consequently, these four mechanisms all require long time periods and relatively stable or very gradually altering environments to produce significant change.
The known natural mechanisms predict a bottom-up development of taxonomic hierarchy. That is, the four known natural processes over time will first produce a proliferation of species, which, in much more time, will produce a proliferation of genera. The proliferation of genera will, eventually, produce new families. The proliferation of families over still more time will yield new orders. New orders will produce new classes, and, last of all, new phyla may be expected to appear.
The problem with the predicted bottom-up taxonomic development is that the fossil record of both the Avalon and Cambrian explosions reveals the opposite. As paleontologists have observed with respect to these explosions, “The major pulse of diversification of phyla occurs before that of classes, classes before that of orders, and orders before that of families” (Erwin, Valentine, and Sepkoski 1987).
The Cambrian explosion refers to the sudden appearance some 538.79 ± 0.21 million years ago (Linnemann et al. 2019), of animals with digestive tracts, circulatory systems, and skeletons, as well as internal, and external organs. These animals require a minimum atmospheric oxygen level of 10%, and they appear in the fossil record at the geological moment this essential level is reached (Tatzel et al. 2017; Ye et al. 2020; Ross 2023).
The Cambrian phyla do not appear in a time-separated sequence of increasing complexity. The most advanced phylum, chordata, the phylum to which humans and all vertebrates belong, appears at the same time as the most primitive Cambrian phyla. That epoch? It is the early, not the middle or the end portion of the Cambrian era. Furthermore, it is not just the non-vertebrate chordates that appear at the beginning of the Cambrian. Vertebrate fish also appear at that time (Xiang-Guang et al. 2002; Shu et al. 2003; Morris and Caron. 2014).
The problem with process structuralists’ appeals to hidden natural self-organizing forces to explain the Avalon and Cambrian explosions is that the changes being attributed to such forces are so far from trivial. They are, in fact, predominant. These proposed forces would be playing such dominant roles that it seems inconceivable that today’s community of research scientists would have missed uncovering and identifying them, if indeed such forces are strictly natural. As paleontologists Kevin Peterson, Michael Dietrich, and Mark McPeek state in a review paper on the Cambrian explosion, “Elucidating the materialistic basis for the Cambrian explosion has become more elusive, not less, the more we know about the event itself” (Peterson, Dietrich, and McPeek 2009). They are not alone among paleontologists in drawing such a conclusion (Wray 1992; Levinton 2008).
Biblical Approach to Researching the Origin and History of Life
The Bible offers a straightforward explanation for the “process structuralist forces” hidden from scientists. The process structuralists are correct that natural selection, mutations, gene exchange, and epigenetics remain inadequate to explain the history of Earth’s life. They are correct, also, in noting that these four mechanisms play only partial, relatively minor, roles in explaining Earth’s life history. However, the hidden self-organizing principles responsible for most of the changes in Earth’s life over time cannot be natural processes or they would not be hidden from direct scientific investigation More likely, they are not merely natural processes.
On the Bible’s first page we see a straightforward account of how life on Earth changed from primitive to progressively more advanced. For six creation “days,” God supernaturally intervened to alter Earth’s environment and fill it with progressively more advanced life forms. On the seventh day, God ceased from his work of physical creation activity.
The description for each of the six creation days ends with the statement that reads in the literal Hebrew: “evening was and morning was, day [X].” This phrase indicates that each of the six creation days had a start time and an end time within Earth’s history. For the seventh day, the “evening and morning” phrase is absent. Its omission suggests that while the seventh day began, it has not yet ended. Rather, it continues. Both Psalm 95:8–11 and Hebrews 4:1–11 affirm this inference.
The cessation of God’s creation work during the seventh day answers the fossil record enigma. It explains why naturalistic processes explain everything we see occurring in the life and physical sciences during the human era. but something more had to be going on during the pre-human eras. God’s supernatural interventions to fill Earth with an abundance and diversity of life forms explain why scientists have been unable to identify current natural processes capable, by themselves, of explaining the origin of life, the increasing complexity of life, or such fossil record events as the Avalon and Cambrian explosions and other mass speciation following mass extinction events. God’s supernatural interventions in the history of Earth’s life leading up to the creation of the first humans—the re-creations mentioned in Psalm 104:29–30—also explains why new life forms that replace extinct life forms continually and perfectly compensate for the Sun’s ongoing brightening by 19–23% over the past 3.8 billion years (Goldblatt and Zahnle 2011; Ross 2016b).
Taking into account both natural and supernatural causes for the origin and history of Earth’s life, rather than limiting science to natural causes alone, avoids conflict with the laws of thermodynamics. It also avoids conflict with the principle that lesser causes do not produce substantially greater effects. The exceptionally high entropy measure of the universe (Pavón and Radicella 2013; Valetim and Jesus 2020) poses an additional problematic barrier to the emergence of life from non-life.
It is philosophically unrealistic and unnecessary to demand that all scientists performing research on the origin and history of life adopt a biblical worldview in conducting their research. However, scientists of all worldviews can and should participate in the endeavor to establish the boundaries between natural causation and supernatural causation, with the vitally important caveat that scientists can no longer pretend that no such boundaries exist. Some scientists in interpreting the data may tend to push the boundaries more firmly toward supernatural causation while others will tend to pull the boundaries more toward natural causation.
Through this give-and-take endeavor along with careful, pervasive testing of competing models through ongoing experiments and observations, the boundaries between supernatural and natural causes are most likely to become more clearly defined. Through this approach science and theology can most efficiently and rapidly advance. Once again, as was the case during the first two centuries of the Reformation, scientists and theologians can become allies rather than enemies in the quest for fuller understanding of reality.
References
- Acciari, V. A. et al. 2020. Bounds on Lorentz Invariance Violation from MAGIC Observation of GRB 190114C. Physical Review Letters 125: id. 021301, DOI:10.1103/PhysRevLett.125.021301. [CrossRef]
- Armstrong, John C., Wells, Llyd E., and Gonzalez, Guillermo. 2002. Rummaging Through Earth’s Attic for the Remains of Ancient Life. Icarus 160: 183–96, DOI:10.1006/icar.2002.6957. [CrossRef]
- Armstrong, John C. 2010. Distribution of Impact Locations and Velocities of Earth Meteorites on the Moon. Earth, Moon, and Planets 107: 43–54, DOI:10.1007/s11038-010-9355-2. [CrossRef]
- Arnett, David. 1996a. Supernovae and Nucleosynthesis (Princeton, NJ: Princeton University Press): 14–15.
- Arnett, David. 1996b. Supernovae and Nucleosynthesis (Princeton, NJ: Princeton University Press), 11.
- Arrhenius, Svante. 1908. Worlds in the Making: The Evolution of the Universe (New York: Harper & Row).
- Aubrey, A. D., Cleaves, H. J., and Bada, Jeffrey L. 2009. The Role of Submarine Hydrothermal Systems in the Synthesis of Amino Acids. Origins of Life and Evolution of Biospheres 39: 91–108, DOI:10.1007/s11084-008-9153-2. [CrossRef]
- Bada, J. L. et al. 1998. A Search for Endogenous Amino Acids in Martian Meteorite ALH84001. Science 279: 362–65, DOI:10.1126/science.279.5349.362. [CrossRef]
- Banreti, Agnes et al. 2022. Biological Effects of the Loss of Homochirality in a Multicellular Organism. Nature Communications 13: id. 7059, DOI:10.1038/s41467-022-34516-x. [CrossRef]
- Bartlett, D. J. et al. 2021. Constraints on Quantum Gravity and the Photon Mass from Gamma Ray Bursts. Physical Review D 104: id. 103516, DOI:10.1103/PhysRevD.104.103516. [CrossRef]
- Bell, Elizabeth A. et al. 2015. Potentially Biogenic Carbon Preserved in a 4.1 Billion-Year-Old Zircon. Proceedings of the National Academy of Sciences USA 112: 14518–21, DOI:10.1073/pnas.1517557112. [CrossRef]
- Bolmont, Julien et al. 2022. First Combined Study of Lorentz Invariance Violation from Observations of Energy-Dependent Time Delays from Multiple-Type Gamma Ray Sources. I. Motivation, Method Description, and Validation Through Simulations of H.E.S.S., MAGIC, and VERITAS Data Sets. Astrophysical Journal 930: id. 75, DOI:10.3847/1538-4357/ac5048. [CrossRef]
- Bonner, William A. 1995. Quoted in Cohen, Jon. Getting All Turned Around Over the Origins of Life on Earth. Science 267: 1265, DOI:10.1126/science.7871419. [CrossRef]
- Borde, Arvind, Guth, Alan H., and Vilenkin, Alexander. 2003. Inflationary Spacetimes Are Incomplete in Past Directions. Physical Review Letters 90: id. 151301, DOI:1103/PhysRevLett.90.151301. [CrossRef]
- Boyd, Richard N. et al. 2018. Sites That Can Produce Left-Handed Amino Acids in the Supernova Neutrino Amino Acid Processing Model. Astrophysical Journal 856: id. 26, DOI:10.3847/1538-4357/aaad5f. [CrossRef]
- Burton, Aaron S. et al. 2012. Understanding Prebiotic Chemistry Through the Analysis of Extraterrestrial Amino Acids and Nucleobases in Meteorites. Chemical Society Reviews 41: 5459–72, DOI:10.1039/c2cs.35109a. [CrossRef]
- Canup, Robin M. 2012. Forming a Moon with an Earth-Like Composition Via a Giant Impact. Science 338: 1052–55, DOI:10.1126/science.1226073. [CrossRef]
- Carr, Michael H. 2000. The Habitability of Mars. Journal of the International Astrobiology Society 30: 392, DOI:10.1023/A:1017300204711. [CrossRef]
- Carroll, Sean M. 2008. What If Time Really Exists? eprint: arXiv:0811.3722.
- Carroll, Sean. 2010. From Eternity to Here: The Quest for the Ultimate Theory of Time (New York: Dutton).
- Chyba, Christopher and Sagan, Carl. 1992. Endogenous Production, Exogenous Delivery and Impact-Shock Synthesis of Organic Molecules: An Inventory for the Origins of Life. Nature 355: 125–32, DOI:10.1038/355125a0. [CrossRef]
- Clery, Daniel. 2016. Interstellar Spacecraft: Breakthrough Starshot Dreams of Using Lasers to Power Trip to Alpha Centauri. Science, News, Scienceinsider, DOI:10.1126/science.aaf9905. [CrossRef]
- Condon, E. U. 1937. Theories of Optical Rotary Power. Reviews of Modern Physics 9: 432–57, DOI:10.1103/RevModPhys.9.432. [CrossRef]
- Crick, F. H. and Orgel, L. E. 1973. Directed Panspermia. Icarus 19: 341–346, DOI:10.1016/0019-1035(73)90110-3. [CrossRef]
- Cronin, J. R. and Pizzarello, S. 1983. Amino Acids in Meteorites. Advances in Space Research 3: 5–18, DOI:10.1016/0273-1177(83)90036-4. [CrossRef]
- de Marcellus, Pierre et al. 2011. Non-Racemic Amino Acid Production by Ultraviolet Irradiation of Achiral Interstellar Ice Analogs with Circularly Polarized Light. Astrophysical Journal Letters 727: id. L27, DOI:10.1088/2041-8205/727/2/L27. [CrossRef]
- Denton, Michael J. 2013. The Types: A Persistent Structuralist Challenge to Darwinian Pan-Selectionism. BIO-Complexity 2013: id. 3, DOI:10.5048/BIO-C.2013.3. [CrossRef]
- Do, Tuan et al. 2019. Relativistic Redshift of the Star S0-2 Orbiting the Galactic Center Supermassive Black Hole. Science 365: 664–68, DOI:10.1126/science.aav8137. [CrossRef]
- Dodd, Matthew S. et al. 2017. Evidence for Early Life in Earth’s Oldest Hydrothermal Vent Precipitates. Nature 543: 60–64, DOI:10.1038/nature21377. [CrossRef]
- Dolgalev, Georgii V. et al. 2023. Estimating Total Quantitative Protein Content in Escherichia coli, Saccharomyces cerevisiae, and HeLa Cells. International Journal of Molecular Sciences 24: id. 2081, DOI:10.3390/ijms24032081. [CrossRef]
- Draganic, Ivan G. 2005. Radiolysis of Water: A Look at Its Origin and Occurrence in Nature. Radiation Physics and Chemistry 72: 181–86, DOI:10.1016/j.radphyschem.2004.09.012. [CrossRef]
- Eldridge, Niles. 2000. The Triumph of Evolution and the Failure of Creationism (New York: W. H. Freeman): 35–36.
- Elsila, Jamie, Glavin, Daniel, and Dworkin, Jason. 2009. Cometary Glycine Detected in Samples Returned by Stardust Meteoritics & Planetary Science 44: 1323–30, DOI:10.1111/j.1945-5100.2009.tb01224.x. [CrossRef]
- Erwin, Douglas H., Valentine, James W., and Sepkoski, J. John Jr. 1987. A Comparative Study of Diversification Events: The Early Paleozoic versus the Mesozoic. Evolution 41: 1183, doi:10.2307/2409086. [CrossRef]
- Exoplanet TEAM. 2023. The Extrasolar Planets Encyclopaedia, Exoplanet Catalog. available online: http://exoplanet.eu/catalog/.
- Flores, Jose J., Bonner, William A., and Massey, Gail A. 1977. Asymmetric Photolysis of (RS)-Leucine with Circularly Polarized Ultraviolet Light. Journal of the American Chemical Society 99: 3622–25, DOI:10.1021/ja00453a018. [CrossRef]
- Frank, Frederic C. 1953. On Spontaneous Asymmetric Synthesis. Biochimica et Biophysica Acta 11: 459–63, DOI:10.1016/0006-3002(53)90082-1. [CrossRef]
- Garcia, Amanda K., Cavanaugh, Colleen M., and Kacar, Betul. 2021. The Curious Consistency of Carbon Biosignatures over Billions of Years of Earth-Life Coevolution. ISME Journal 15: 2183–2194, DOI:10.1038/s41396-021-00971-5. [CrossRef]
- Giovannoni, Stephen J. et al. 2005. Genome Streamlining in a Cosmopolitan Oceanic Bacterium. Science 309: 1242–45, DOI:10.1126/science.1114057. [CrossRef]
- Goldblatt, Colin and Zahnle, Kevin. 2011. Faint Young Sun Paradox Remains. Nature 474: id. E1, DOI:10.1038/nature09961. [CrossRef]
- Goodwin, Brian. 2001. How the Leopard Changed Its Spots: The Evolution of Complexity (Princeton, NJ: Princeton University Press).
- Grassineau, N. V. et al. 2005. Distinguishing Biological from Hydrothermal Signatures via Sulphur and Carbon Isotopes in Archaean Mineralizations at 3.8 and 2.7 Ga. Geological Society London, Special Publications 248: 195–212, DOI:10.1144/GSL.SP.2005.248.01.11. [CrossRef]
- Green, James et al. 2020. When the Moon Had a Magnetosphere. Science Advances 6: id. eabc0865, DOI:10.1126/sciadv.abc0865. [CrossRef]
- Guo, Jianpo et al. 2009. Probability Distribution of Terrestrial Planets in the Habitable Zones around Host Stars. Astrophysics and Space Science 323: 367–73, DOI:10.1007/s10509-009-0081-z. [CrossRef]
- Hawking, Stephen and Penrose, Roger. 1970. The Singularities of Gravitational Collapse and Cosmology. Proceedings of the Royal Society of London A 314: 529–48, DOI:10.1098/rspa.1970.0021. [CrossRef]
- Hector, Silva O. et al. 2021. Astrophysical and Theoretical Physics Implications from Multimessenger Neutron Star Observations. Physical Review Letters 126: id. 181101, DOI:10.1103/PhysRevLett.126.181101. [CrossRef]
- Hennet, R. J. -C., Holm, N. G., and Engel, M. H. 1992. Abiotic Synthesis of Amino Acids under Hydrothermal Conditions and the Origin of Life: A Perpetual Phenomenon? Naturwissenschaften 79: 361–65, DOI:10.1007/BF01140180. [CrossRef]
- Hoang, Thiem et al. 2017. The Interaction of Relativistic Spacecrafts with the Interstellar Medium. Astrophysical Journal 837: id. 5, Doi:10.3847/1538-4357/aa5da6. [CrossRef]
- Homan, D. C. and Wardle, J. F. C. 2000. Direct Distance Measurements to Superluminal Radio Sources. Astrophysical Journal 535: 575–85, DOI:10.1086/308884. [CrossRef]
- Hoyle, Fred and Wickramasinghe, N. C. 1981. Evolution from Space: A Theory of Cosmic Creationism (New York: Simon and Schuster).
- Hoyle, Fred, Burbidge, Geoffrey, and Narlikar, Jayant V. 2000. A Different Approach to Cosmology: From a Static Universe through the Big Bang towards Reality (Cambridge, UK: Cambridge University Press).
- Itaya, Mitsuhiro. 1995. An Estimation of Minimal Genome Size Required for Life. FEBS Letters 362: 257–60, DOI:10.1016/0014-5793(95)00233-y. [CrossRef]
- Johnson, Adam P. et al. 2008. The Miller Volcanic Spark Discharge Experiment. Science 322: 404, DOI:10.1126/science.1161527. [CrossRef]
- Kazmierczak, Jósef and Kempe, Stephan. 2003. Modern Terrestrial Analogues for the Carbonate Globules in Martian Meteorite ALH84001. Naturwissenschaften 90: 167–72, Doi:10.1007/s00114-003-0411-x. [CrossRef]
- Kobayashi, Chiaki and Nomoto, Ken’ichi. 2009. The Role of Type Ia Supernovae in Chemical Evolution. I. Lifetime of Type Ia Supernovae and Metallicity Effect. Astrophysical Journal 707: 1466–84, DOI:10.1088/0004-637X/707/2/1466. [CrossRef]
- Kotra, R. K. et al. 1979. Amino Acids in a Carbonaceous Chondrite from Antarctica. Journal of Molecular Evolution 13: 179–83, DOI:10.1007/BF01739477. [CrossRef]
- Kramer, Michael et al. 2021. Strong-Field Gravity Tests with the Double Pulsar. Physical Review X 11: id. 041050, DOI:10.1103/PhysRevX.11.041050. [CrossRef]
- Kuan, Yi-Jehng et al. 2003. A Search for Interstellar Pyrimidine. Monthly Notices of the Royal Astronomical Society 345: 650–56, DOI:10.1046/j.1365-8711.2003.06975.x. [CrossRef]
- Kuhn, Werner. 1930. The Physical Significance of Optical Rotary Power. Transactions of the Faraday Society 26: 293–308, DOI:10.1039/TF9302600293. [CrossRef]
- Kuo, C. Y. et al. 2013. The Megamaser Cosmology Project. V. An Angular Diameter Distance to NGC 6264 at 140 Mpc. Astrophysical Journal 767: id. 155, DOI:10.1088/0004-637X/767/2/155. [CrossRef]
- Larralde, Rosa, Robertson, M. P., and Miller, Stanley L. 1995. Rates of Decomposition of Ribose and Other Sugars: Implications for Chemical Evolution. Proceedings of the National Academy of Sciences USA 92: 8158–60, DOI:10.1073/pnas.92.18.8158. [CrossRef]
- Leslie, Mitch. 2021. Scientists Coax Cells with the World’s Smallest Genomes to Reproduce Normally. Science: News March 29, DOI:10.1126/science.abi7601. [CrossRef]
- Levinton, Jeffrey S. 2008. The Cambrian Explosion: How Do We Use the Evidence? BioScience 58: 855, DOI:10.1641/B580912. [CrossRef]
- Levy, Matthew and Miller, Stanley L. 1998. The Stability of the RNA Bases: Implications for the Origin of Life. Proceedings of the National Academy of Sciences USA 95: 7933–38, DOI:10.1073/pnas.95.13.7933. [CrossRef]
- Lineweaver, Charles H., Fenner, Yeshe, and Gibson Brad K. 2004. The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way. Science 303: 59–62, DOI:10.1126/science.1092322. [CrossRef]
- Linnemann et al. 2019. New High-Resolution Age Data from the Ediacaran-Cambrian Boundary Indicate Rapid, Ecologically Driven Onset of the Cambrian Explosion. Terra Nova 31: 49–58, doi:10.1111/ter.12368. [CrossRef]
- Lymer, Elizabeth A. et al. 2021. UV Time-Resolved Laser-Induced Fluorescence Spectroscopy of Amino Acids Found in Meteorites: Implications for Space Science and Exploration. Astrobiology 21: 1350–1362, DOI:10.1089/ast.2021.0006. [CrossRef]
- Maehara, Hiroyuki et al. 2012. Superflares on Solar-Type Stars. Nature 485: 478–81, DOI:10.1038/nature11063. [CrossRef]
- Maehara, Hiroyuki et al. 2015. Statistical Properties of Superflares on Solar-Type Stars Based on 1-Minute Cadence Data. Earth, Planets, and Space 67: id. 59, DOI:10.1186/s40623-015-0217-z. [CrossRef]
- Maniloff, Jack. 1996. The Minimal Cell Genome: “On Being the Right Size”. Proceedings of the National Academy of Sciences USA 93: 10004–6, DOI:10.1073/pnas.93.19.10004. [CrossRef]
- Manniloff, Jack et al. 1997. Nannobacteria: Size Limits and Evidence. Science 276: 1773–76, DOI:10.1126/science.276.5320.1773e. [CrossRef]
- Manning, Craig E., Mojzsis, Stephen J., and Harrison, T. Mark. 2006. Geology, Age and Origin of Supracrustal Rocks at Akilia, West Greenland. American Journal of Science 306: 303–366, DOI:10.2475/05.2006.02. [CrossRef]
- McDonald, Gene D. and Storrie-Lombardi, Michael C. 2010. Biochemical Constraints in a Protobiotic Earth Devoid of Basic Amino Acids: The ‘BAA(-) World’. Astrobiology 10: 989–1000, DOI:10.1089/ast.2010.0484. [CrossRef]
- Melosh, H. J. 2003. Exchange of Meteorites (and Life?) Between Stellar Systems. Astrobiology, 3: 207–215, DOI:10.1089/153110703321632525. [CrossRef]
- Milo, Ron. 2013. What Is the Total Number of Protein Molecules per Cell Volume? A Call to Rethink Some Published Values. Bioessays 35: 1050–55, DOI:10.1002/bies.201300066. [CrossRef]
- Morris, Simon Conway. 2003. Life’s Solution: Inevitable Humans in a Lonely Universe (Cambridge, UK: University of Cambridge Press), DOI:10.1017/CBO97805111535499. [CrossRef]
- Morris, Simon Conway and Caron, Jean-Bernard. 2014. A Primitive Fish from the Cambrian of North America. Nature 512: 419–22, DOI:10.1038/nature13414. [CrossRef]
- Narlikar, J. V., Vishwakarma, R. G., and Burbidge, G. 2003. Inhomogeneities in the Microwave Background Radiation Interpreted within the Framework of the Quasi-Steady State Cosmology. Astrophysical Journal 585: 1–11, DOI:10.1086/345928. [CrossRef]
- Narlikar, Jayant V., Burbidge, Geoffrey, and Vishwakarma, R. G. 2007. Cosmology and Cosmogony in a Cyclic Universe. Journal of Astrophysics and Astronomy 28: 67–99, DOI:10.1007/s12036-007-0007-5. [CrossRef]
- Pavón, Diego and Radicella, Ninfa. 2013. Does the Entropy of the Universe Tend to a Maximum? General Relativity and Gravitation 45: 63–68, DOI:10.1007/s10714-012-1457-x. [CrossRef]
- Penrose, Roger. 1994. Shadows of the Mind: A Search for the Missing Science of Consciousness (New York: Oxford University Press): 230.
- Perlman, E. S. et al. 2015. New Constraints on Quantum Gravity from X-Ray and Gamma-Ray Observations. Astrophysical Journal 805: id. 10, DOI:10.1088/0004-637X/805/1/10. [CrossRef]
- Peterson, Kevin J., Dietrich, Michael R., and McPeek, Mark A. 2009. MicroRNAs and Metazoan Macroevolution: Insights into Canalization, Complexity, and the Cambrian Explosion. BioEssays 31: 737, doi:10.1002/bies.200900033. [CrossRef]
- Petigura, Erik A., Howard, Andrew W., and Marcy, Geoffrey W. 2013. Prevalence of Earth-Size Planets Orbiting Sun-Like Stars. Proceedings of the National Academy of Sciences USA 110: 19273–78, DOI:10.1073/pnas.1319909110. [CrossRef]
- Petlowski, Janusz Jurand, Bains, William, and Seager, Sara. 2020. On the Potential of Silicon as a Building Block for Life. Life 10: id. 84, DOI:10.3390/life10060084. [CrossRef]
- Reinhold, Timo et al. 2020. The Sun Is Less Active Than Other Solar-Like Stars. Science 368: 518–21, DOI:10.1126/science.aay3821. [CrossRef]
- Richert, Clemens. 2018a. Prebiotic Chemistry and Human Intervention. Nature Communications 9: id. 5177, DOI:10.1038/241467-018-07219-5. [CrossRef]
- Richert Clemens. 2018b. Prebiotic Chemistry and Human Intervention. Nature Communications 9: id. 5177, page 2, DOI:10.1038/241467-018-07219-5. [CrossRef]
- Richert Clemens. 2018c. Prebiotic Chemistry and Human Intervention. Nature Communications 9: id. 5177, page 2, DOI:10.1038/241467-018-07219-5. [CrossRef]
- Romoli, C. et al. 2017. Observation of the Extremely Bright Flare of the FSRQ 3C279 with H.E.S.S. II. Proceedings of Science 301: id. 649, DOI:10.22323/1.301.0649. [CrossRef]
- Rosing, Minik T. 1999. 13C-Depleted Carbon Microparticles in >3700-Ma Sea–Floor Sedimentary Rocks from West Greenland. Science 283: 674–76, DOI:10.1126/science.283.5402.674. [CrossRef]
- Ross, Hugh. 2007. Why We Need to Return to the Moon. Today’s New Reason to Believe. available online: https://reasons.org/explore/publications/articles/why-we-need-to-return-to-the-moon.
- Ross, Hugh. 2016a. Improbable Planet (Grand Rapids, MI: Baker Books): 81–93.
- Ross, Hugh. 2016b. Improbable Planet (Grand Rapids, MI: Baker Books): 143–164.
- Ross, Hugh. 2018. The Creator and the Cosmos, 4th ed’n (Covina, CA: RTB Press): 45–120.
- Ross, Hugh. 2019a. Tiny Habitable Zones for Complex Life. Today’s New Reason To Believe, available online: https://reasons.org/explore/blogs/todays-new-reason-to-believe/read/todays-new-reason-to-believe/2019/03/04/tiny-habitable-zones-for-complex-life.
- Ross, Hugh. 2019b. Complex Life’s Narrow Requirements for Atmospheric Gases. Today’s New Reason To Believe, available online: https://reasons.org/explore/blogs/todays-new-reason-to-believe/read/todays-new-reason-to-believe/2019/07/01/complex-life-s-narrow-requirements-for-atmospheric-gases.
- Ross, Hugh. 2022a. Designed to the Core (Covina, CA: RTB Press): 132–181.
- Ross, Hugh. 2022b. Designed to the Core (Covina, CA: RTB Press): 55–90.
- Ross, Hugh. 2023. What Does the Bible Say About the Big Bang? Today’s New Reason to Believe. available online: https://reasons.org/explore/blogs/todays-new-reason-to-believe/what-does-the-bible-say-about-the-big-bang.
- Sagan, Carl. 1973. The Cosmic Connection: An Extraterrestrial Perspective, (produced by Jerome Agel, Garden City, NY: Anchor Press/Doubleday): 189–190.
- Schidlowski, Manfred. 1988. A 3,800-Million-Year Isotopic Record of Life from Carbon in Sedimentary Rocks. Nature 333: 313–18, DOI:10.1038/333313a0. [CrossRef]
- Schidlowski, Manfred. 2001. Carbon Isotopes as Biogeochemical Recorders of Life Over 3.8 Ga of Earth History: Evolution of a Concept. Precambrian Research 106: 117–134, DOI:10.1016/S0301-9268(00)00128-5. [CrossRef]
- Shibata, Takanori et al. 1998. Amplification of a Slight Enantiomeric Imbalance in Molecules Based on Asymmetric Autocatalysis: The First Correlation between High Enantiomeric Enrichment in a Chiral Molecule and Circularly Polarized Light. Journal of the American Chemical Society 120: 12157–58, DOI:10.1021/ja980815w. [CrossRef]
- Shu, D-G et al. 2003. Head and Backbone of the Early Cambrian Vertebrate Haikouichthys. Nature 421: 526-29, DOI:10.1038/nature01264. [CrossRef]
- Snyder, L. E. et al. 2005. A Rigorous Attempt to Verify Interstellar Glycine. Astrophysical Journal 619: 914–30, DOI:10.1086/426677. [CrossRef]
- Spina, L. et al. 2016. Nucleosynthesis History of Elements in the Galactic Disk. [X/Fe]-Age Relations from High-Precision Spectroscopy. Astronomy & Astrophysics 593: id. A125, DOI:10.0004-6361/2016285577. [CrossRef]
- Spitoni, E., Gioannini, L., and Matteucci, F. 2017. Galactic Habitable Zone Around M and FGK Stars with Chemical Evolution Models that Include Dust. Astronomy & Astrophysics 605: id. A38, DOI:10.1051/0004-6361/201730545. [CrossRef]
- Steele, Edward J. 2018a. Cause of Cambrian Explosion – Terrestrial or Cosmic? Progress in Biophysics and Molecular Biology 136: 3–23, DOI:10.1016/j.pbiomolbio.2018.03.004. [CrossRef]
- Steele, Edward J. 2018b. Cause of Cambrian Explosion – Terrestrial or Cosmic? Progress in Biophysics and Molecular Biology 136: 5, DOI:10.1016/j.pbiomolbio.2018.03.004. [CrossRef]
- Steenstra, E. S. et al. 2020. A Possible High-Temperature Origin of the Moon and Its Geochemical Consequences. Earth and Planetary Science Letters 538: id. 116222, DOI:10.1016/j.epsl.2020.116222. [CrossRef]
- Tamburini, F. et al. 2011. No Quantum Gravity Signature from the Farthest Quasars. Astronomy & Astrophysics 533: id. A71, DOI:10.1051/0004-6361/201015808. [CrossRef]
- Thacker, Cameron et al. 2013. H-ATLAS: The Cosmic Abundance of Dust from the Far-Infrared Background Power Spectrum. Astrophysical Journal 768: id. 58, DOI:10.1088/0004-637X/768/1/58. [CrossRef]
- Tokano, Yoshinori et al. 2007. Asymmetric Synthesis of Amino Acid Precursors in Interstellar Complex Organics by Circularly Polarized Light. Earth and Planetary Science Letters 254: 106–114, DOI:10.1016/j.epsl.2006.11.030. [CrossRef]
- Tsujimoto, T. 2023. From Galactic chemical evolution to cosmic supernova rates synchronized with core-collapse supernovae limited to the narrow progenitor mass range. Monthly Notices of the Royal Astronomical Society 518: 3475–81, DOI:10.1093/mnras/stac3351. [CrossRef]
- Valentin, R. and Jesus, J. F. 2020. Thermodynamic Constraints on Matter Creation Models. European Physical Journal C 80: id. 1035, DOI:10.1140/epjc/s10052-020-08592-6. [CrossRef]
- Wainwright, Milton et al. 2013. Isolation of Biological Entities from the Stratosphere (22–27 Km). Journal of Cosmology 22: 10189–97, DOI:10.5334/jc.sg. [CrossRef]
- Wang, Kun and Jacobsen, Stein B. 2016. Potassium Isotope Evidence for a High-Energy Giant Impact Origin of the Moon. Nature 538: 487–90, DOI:10.1038/nature19341. [CrossRef]
- Weisberg, J. M. and Huang, Y. 2016. Relativistic Measurements from Timing the Binary Pulsar PSR B1913+16. Astrophysical Journal 829: id. 55, DOI:10.3847/0004-637X/829/1/55. [CrossRef]
- Weiss, Benjamin P. et al. 2004. Magnetic Tests for Magnetosome Chains in Martian Meteorite ALH84001. Proceedings of the National Academy of Sciences USA 101: 8281–84, DOI:10.1073/pnas.0402292101. [CrossRef]
- Wickramasinghe, N. Chandra, Wickramasinghe, Dayal T., and Steele, Edward J. 2020. Cometary Panspermia and Origin of Life? Advances in Genetics 106: id. 5, DOI:10.1016/bs.adgen.2020.03.003. [CrossRef]
- Wray, Gregory A. 1992. Rates of Evolution in Developmental Processes. American Zoologist 32: 131, DOI:10.1093/icb/32.1.123. [CrossRef]
- Xiang-Guang, Hou et al. 2002. New Evidence on the Anatomy and Phylogeny of the Earliest Vertebrates. Proceedings of the Royal Society B: Biological Sciences 269: 1865–69, DOI:10.1098/rspb.2002.2104. [CrossRef]
- Yungelson, Lev R. and Livio, Mario. 2000. Supernova Rates: A Cosmic History. Astrophysical Journal 528: 108–117, DOI:10.1086/308174. [CrossRef]
- Zagorski, Zbigniew Pavel. 2007. Question 2: Relation of Panspermia-Hypothesis to Astrobiology. Origins of Life and Evolution of Biospheres 37: 351–355, DOI:10.1007/s11084-007-9074-5. [CrossRef]
- Zhang, Jinghua et al. 2020. Solar-Type Stars Observed by LAMOST and Kepler. Astrophysical Journal Letters 894: id. L11, DOI:10.3847/2041-8213/ab8795. [CrossRef]
Figure 1.
Abundance of Uranium and Thorium Throughout Cosmic History.
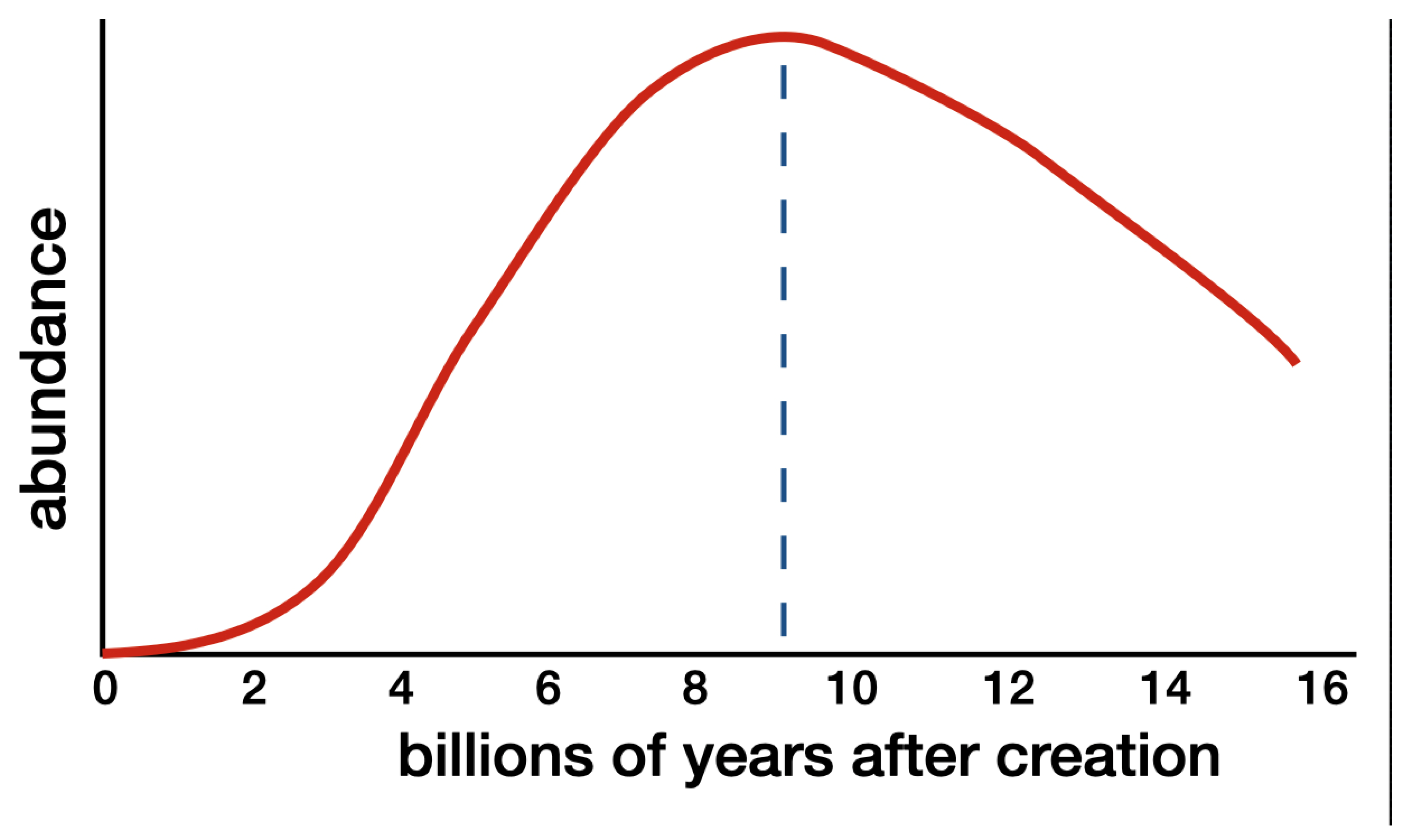
Figure 2.
Sun’s Flaring Activity and X-Ray Radiation Levels Throughout Its History. The y-axis is logarithmic. The dotted line indicates the present. During its first half-billion years, the Sun’s flaring activity levels were more than 10,000 times greater than its present level. The Sun’s intensity of particle radiation, gamma-ray, x-ray, and ultraviolet emission strongly correlate with its flaring activity level. Only for stars that are virtually identical to the Sun and 4.57 billion years old is global high-technology civilization possible on one of its planets. diagram credit: Hugh Ross
Figure 2.
Sun’s Flaring Activity and X-Ray Radiation Levels Throughout Its History. The y-axis is logarithmic. The dotted line indicates the present. During its first half-billion years, the Sun’s flaring activity levels were more than 10,000 times greater than its present level. The Sun’s intensity of particle radiation, gamma-ray, x-ray, and ultraviolet emission strongly correlate with its flaring activity level. Only for stars that are virtually identical to the Sun and 4.57 billion years old is global high-technology civilization possible on one of its planets. diagram credit: Hugh Ross
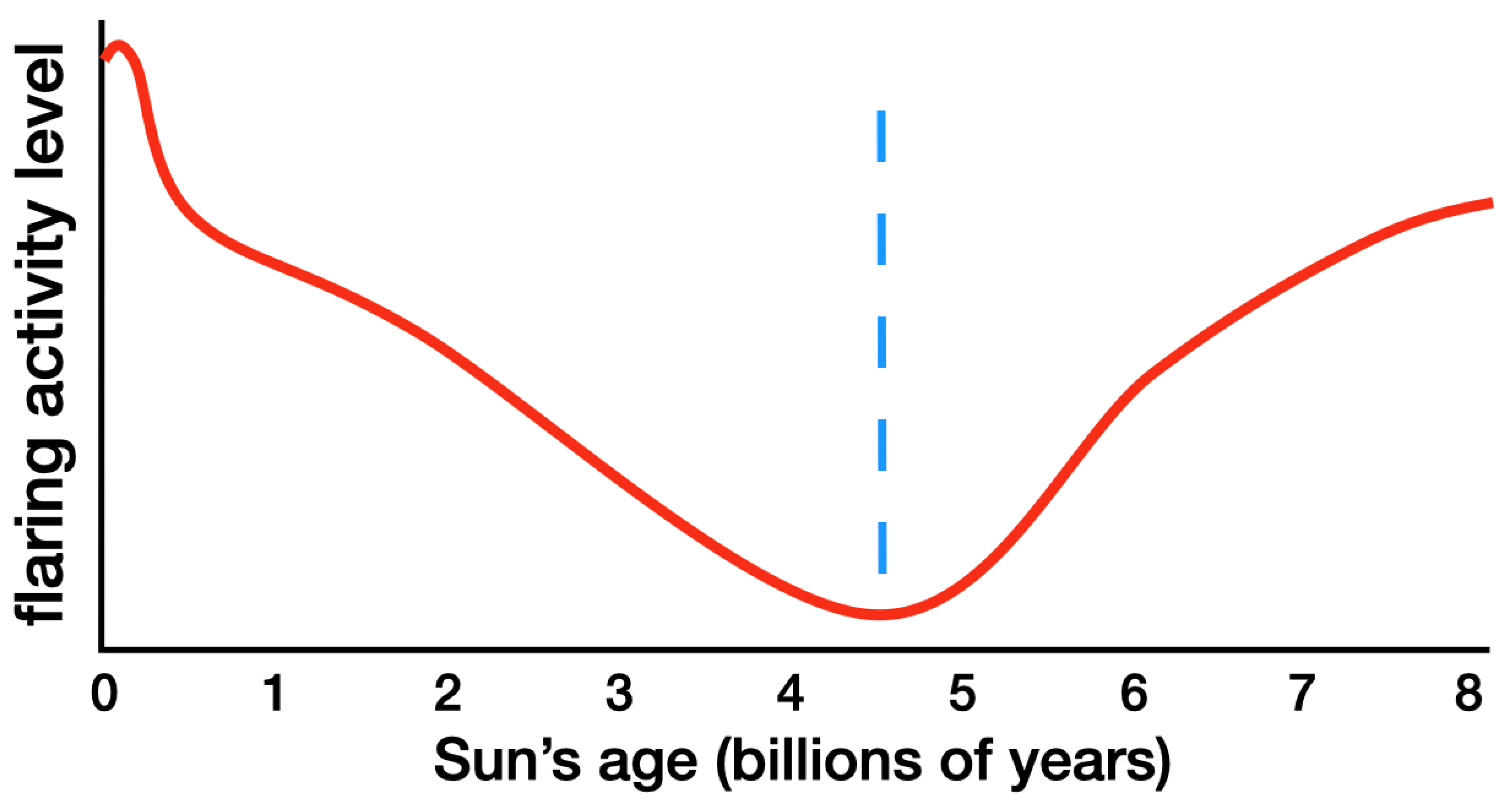
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



