
Submitted:
15 March 2023
Posted:
21 March 2023
You are already at the latest version
Abstract
;Yams are economic and medicinal crops with a long growth cycle, spanning between 9-11 months due to the prolonged tuber dormancy. Tuber dormancy has constituted a major constraint in yam production and genetic improvement. In this study, we performed non-targeted comparative metabolomic profiling of tubers of two white yam genotypes, (Obiaoturugo and TDr1100873), to identify metabolites and associated pathways that regulate yam tuber dormancy using gas chromatography-mass spectrometry (GC-MS). Yam tubers were sampled between 42 days after physiological maturity (DAPM) till tuber sprouting. The sampling points include 42-DAPM, 56-DAPM, 87DAPM, 101-DAPM, 115-DAPM, and 143-DAPM. A total of 949 metabolites were annotated, 559 in TDr1100873 and 390 in Obiaoturugo. 39 differentially accumulated metabolites (DAMs) were identified across the studied tuber dormancy stages in the two genotypes. 27 DAMs were conserved between the two genotypes, whereas, 5 DAMs were unique in the tubers of TDr1100873 and 7 DAMs were in the tubers of Obiaoturugo. The differentially accumulated metabolites (DAMs) spread across 14 major functional chemical groups. Amines and biogenic polyamines, amino acids and derivatives, Alcohols, flavonoids, alkaloids, phenols, esters, coumarins and phytohormone positively regulated yam tuber dormancy induction and maintenance, whereas, fatty acids, lipids, nucleotides, carboxylic acids, sugars, terpenoids, benzoquinones, and benzene derivatives positively regulated dormancy breaking and sprouting in tubers of both yam genotypes. Metabolite set enrichment analysis (MSEA) revealed that 12 metabolisms were significantly enriched during yam tuber dormancy stages. Metabolic pathway topology analysis further revealed that six metabolic pathways (linoleic acid metabolic pathway, phenylalanine metabolic pathway, galactose metabolic pathway, starch and sucrose metabolic pathway, alanine-aspartate-glutamine metabolic pathways and purine metabolic pathway) exerted significant impact on yam tuber dormancy regulation. This result provides vital insights into molecular mechanisms regulating yam tuber dormancy.
Keywords:
Yam tuber
; metabolomics
; pathways
; molecular-mechanism
; dormancy
; regulation
1. Introduction
Yam is a multi-species crop which has about 613 known species that produce tubers, bulbils or rhizomes [1]. Of these, about 10 are cultivated over larger area and serve as a staple food crop, about 50 other species are also eaten as wild-harvested staples famine food, thus this genus occupies a prominent position in global food insecurity combat, [2,3].Yam is placed fourth position among the utilized root and tuber crops globally after potatoes (Solanum spp.), cassava (Manihot esculenta) and sweet potatoes (Ipomoea spp.) and the second in West Africa after cassava [4,5]. However, the sensorial preference of yam coupled with its better organoleptic properties and storability compared to cassava, potatoes, sweet potatoes and plantain has led to it high demand as a major cash crop in Sub-Sahara Africa [1]. It’s potential as a source of food is attributed to its high levels of carbohydrates including fiber, starch and sugar, contributing about 200 dietary calories per person per day to more than 300 million people in the tropics [6], in addition to other nutritional benefits such as proteins, lipids, vitamins and minerals [2]. Yam tuber also contains abundance of steroidal C27 saponins, and diosgenin. the aglycone portion of the abundant saponin dioscin has been industrially exploited as the starting material for the synthesis of pregnenolone-derived steroid, thus making yam an industrial crop [7].
Yam tuber dormancy is temporary growth arrest of the continuous underlying meristematic cells beneath the dead brown skin of the tuber[8]. It is an inherent mechanism that regulates tuber sprouting. The dormancy phase has been divided into three categories including endodormancy (deep dormancy during which growth arrest is influenced by internal physiological and genetic factors within the meristem), para-dormancy (this occurs when growth is arrested by physiological factors external to the meristem, eco-dormancy (growth is stopped by unfavorable external or environmental factors.
Prolonged tuber dormancy after physiological maturity has constituted a great constraint in yam crop production and genetic improvement [9]. Tuber dormancy is the major cause of ware and seed tubers’ inability to sprout for prolonged period, during which tubers remain dormant; incapable of developing an internal or external shoot buds/sprout for about 150 to 210 days depending on; species, genotype, growing and storage environmental conditions [10,11,12]. Thus, making it impossible to have more than one crop cycle per year, and thereby limiting yam crop productivity, tuber availability for industrial use, and slowing down the rate of genetic improvement [13]. Dormancy induction and release is associated with numerous physiological and biochemical processes that are regulated by gene expression, protein synthesis, hormonal signaling and energy metabolism [14]. In these processes, both the decomposition and synthesis of metabolites and conversion and consumption of energy occur. Thus, generating a genotype and dormancy stage-specific metabolites [15,16]. Some of these metabolites have been found to regulate dormancy in seeds of various plant species. The typical example is phytohormones, whose involvement in seed dormancy regulation has always been a popular topic in seed physiology and biochemistry research [17]. The most reported phytohormone influencing dormancy induction in crops is abscisic acid (ABA), it inhibits the process of seed germination, though complex interactions exist between ABA and other phytohormones [18]. Gibberellic acid (GA) is another important phytohormones known to be involved in dormancy regulation. It acts antagonistically with ABA to promote seed germination (dormancy breaking), while jasmonic acid (JA) shows a synergetic effect with ABA in regulating dormancy [7]. Exogenous treatment with auxin in soybean was found to induce dormancy by decreasing the GA/ABA ratio [19]. Therefore, the roles of various phytohormones in dormancy regulation are complex and subtle.
Polyamine has also been reported to play key role as modulator of plant growth and development, and its involvement in seed dormancy regulation has attracted research attention in the recent. Dormancy breaking led to accumulation of the total polyamine content in the seeds [20]. Polyamine was found to be distributed compartmentally in dormancy breaking seeds, which may suggest its involvement in the process [21]; presoaking with polyamine improved seed germinability[22]. This effect has been suspected to involve starch metabolism, phytohormones interactions and antioxidant defenses [23,24]. In addition, the involvement of amino acids in dormancy regulation has also attracted research attention. For instance, it has been reported that essential amino acids such lysin, methionine, leucine, isoleucine, threonine, phenylalanine, and valine increased significantly during dormancy breaking process in wheat, brown rice, triticale, and Arabidopsis [25,26]. Asparagine, arginine, and γ-aminobutyric (GABA) are the three most common amino compounds in the mobilization of nitrogen reserves in dormancy breaking chestnut seeds [27]. Therefore, as the major transporter of all forms of nitrogen and the key factor of carbon/nitrogen balance in the plants, the role of amino acids in dormancy regulation may not be ignored [18].
Carbohydrates play multiple roles in plant growth and development, besides acting as the primary source of carbon and energy, hence energy metabolism has also been implicated in the dormancy regulation. It has been reported that Embden Meyerhof Parnas (EMP) pathway, tricarboxylic acid (TCA) pathway and pentose phosphate pathway (PPP) are strictly related dormancy in different plants [28]. For instance, the TCA cycle was enhanced, while PPP pathway was slowly decreased during apple bud dormancy breaking [29]. In grape dormancy breaking induced by chemical and low temperature was regulated by PPP, EMP and TCA cycle [30,31]. Also, other pathways have been reported to be involve in dormancy regulation in herbaceous plants, for example, sucrose metabolism, lipid metabolism, amino acid metabolism, were all downregulated in imbibed dormant seeds of herbaceous plants [32]. Thus, demonstrating that the ability of imbibed seeds to synthesize protein and generate adenosine triphosphate (ATP) decreases once dormancy breaking process commences [33,34]. However, two studies have reported that imbibed dormant seeds of Picea glauca and Julans regia exhibited inactivation of sucrose and amino acids due to wet cold induced dormancy [32]. Furthermore, accumulation of galactinol and raffinose family of oligosaccharides (RFOs) during seeds maturation stages in some legume has indicated their roles in desiccation tolerance and longevity [35]. Galactinol content in dry seeds has also been used as a biomarker for seed longevity in Brassiceae and tomato [36]. Sucrose-non-fermenting 1-related kinase1 (SnRK1) transcription family has been reported to play the role of energy metabolism sensors in plant system, they detect when energy is below optimum in the plant system, and inhibit plants growth and developmental processes that are energy consuming such as dormancy breaking (sprouting), in order to maintain energy homeostasis during the low energy economy which characterize dormant state [37,38]. There are also reports that trehalose-6-phospate (T6P) initiates the process of growth from any plant part under growth arrest (dormancy), by inhibiting the action of (SnRK1) [39,40].
Effective reduction of yam tuber dormancy duration through genetic manipulation is considered a reliable solution to the challenge of prolonged tuber dormancy and ensuring more than one yam crop cycle per annual. It is important to note that the paucity of genetic resource information controlling the dormancy trait has been limiting to overcoming the barriers mentioned above. Our study presents an alternative route in understanding implicated pathways at the biochemical and genetic level.
The development of metabolomics technologies has improved our understanding of the complex molecular interaction of biological systems. Due to its high throughput and efficiency, metabolomics has been applied to research on the mechanism of dormancy in several plant species [18]. However, in yam crop only few metabolomics studies exist, and to the best of our knowledge this is the first time, metabolomics approach is being applied to study yam tuber dormancy. In this study, untargeted GC-MS approach was used to profile tubers of two white yam genotypes (Obiaoturugo and TDr1100873) for differentially accumulated metabolites from dormancy induction to dormancy breaking in order to determine the metabolites and their associated pathways that regulate yam tuber dormancy.
2. Experimental Design
2.1. Genetic Material
Two yam genotypes (Obiaoturugo and TDr1100873) of white yam specie (D. rotundata,) were used for the study. Obiaoturugo is a popular landrace while TDr1100873 is an advanced breeding clone. Both individuals were sourced from the Yam breeding program of National Root Crops Research Institute (NRCRI) Umudike, Nigeria.
2.2. Field Study Area
The field was established at the Eastern research farm of the National Root Crops Research Institute (NRCRI) Umudike, Nigeria. Umudike is a rainforest agroecology located at; 5.4729° N, 7.5480° E, 152m ASL with mean annual rainfall of 2093mm, mean annual temperature of 27.3℃ and mean annual relative humidity of 82%, and sandy loam soil with pH range from 4.3 to 5.27.
2.3. Planting and Agronomic Management
Uniform set size (200 g) of only proximal and distal regions of the tubers were planted in order to maintain relatively uniform germination time. A total of 30 plant stands of each genotype were planted in randomized complete block design, replicated three times, with 10 plant stands of each genotype per replicate. The field was adequately maintained under rainfed condition, and all cultural practices including application of NPK fertilizer at the recommended dose of 80:60:100kg/ha [41] were carried out. Tubers were harvested at 50% senescence, which is the tuber physiological maturity stage.

Figure 1.
(a–c); showing a; established field of the yam genotypes, (b) tubers of Obiaoturugo in post-harvest study facility, and c tubers of TDr1100873 in post-harvest study facility.
Figure 1.
(a–c); showing a; established field of the yam genotypes, (b) tubers of Obiaoturugo in post-harvest study facility, and c tubers of TDr1100873 in post-harvest study facility.
2.4. Postharvest Study and Sampling
After harvesting, tubers were freighted to Centre of Plant Molecular Biology and Bioinformatics, Department of Plant Biotechnology Tamil Nadu Agricultural University, Coimbatore, India, where the tubers were stored in green house facility, with natural light condition and sunshine hours per day between 10 to 12 hours, relative humidity range of 38% to 78%, maximum and minimum temperature range from 26 to 36.5℃ and 17.5 to 26.5℃ respectively, during the study. Triplicate samples of tubers were collected at the following days after physiological maturity (DAPM): 42-DAPM, 56-DAPM, 87-DAPM, 101-DAPM, 115-DAPM and sprouted tuber (143-DAPM). The two genotypes (Obiaoturugo and TDr1100873) exhibited different dormant phenotype. Obiaoturugo exhibited shorter dormant phenotype and sprouted at 101days after tuber physiological maturity, and was sampled only four times within this period at (42, 56, 87, 101) days. Whereas, TDr1100873 exhibited longer dormant phenotype and sprouted at 143 days after tuber physiological maturity, and was sampled six times within the period at (42, 56, 87, 101, 115, 143) days after tuber physiological maturity. The samples were frozen in the liquid nitrogen immediately after collection, and stored in -80℃ till when sampling was completed. At the completion of sampling, freeze-dried samples were microwaved for 5 minutes and then oven dried at oven at 72℃ for 48hrs, and grinded into powder using commercial grinder.
2.5. Metabolites Extraction
10g of powdered samples were wrapped in filter papers and dropped in the cylindrical tubes of Soxhlet apparatus and filled with 400mL of 100% methanol solvent. Extraction was done using the at 70℃ in nine (9) extraction cycles for each sample. 50mL of the extract were collected in 50mL centrifuging tubes and air dried to 7.5mL.
2.5.1. Sample Processing
500µL of the extracts were pipetted into 2.0 appendoff tubes and incubated in water-bath for 15mins at 37℃, before centrifuging at 10000rpm and 4℃ for mins. Samples were filtered using 0.2mm filter paper and syringe. After filtering, samples were diluted with methanol at ratio of 1:9 at which samples became colorless. 500mL of the diluted samples were concentrated at 20000rpm for 2hrs. The concentrated solid extracts were derivatized according to [1] with slight modifications by dissolving it in 50mL solution of (20mg methoxylamine hydrochloride + 1mL of pyrimidine), incubated for 1.5hrs at 37℃. 80µL of (N-methy-N-trimethyl-Sisyl)-tri-fluro acetamine) was also added and incubated for 30mins at 37℃.
2.6. GC-MS Analysis
Untargeted metabolomics data was acquired using two auto-injecting GC-MS machines (Turbomass and 8040NX) by injecting 1ml of the sample. The GC oven was held for 2 min at 705°C, ramp at 10°C/min to 150 °C, held for 2 min, ramp at 5°C/min to 220 °C, held 1 min, ramp 10°C/min to 250°C, held 1 min, InjAauto=220°C. Helium was the carrier gas at a flowrate of 1.3 mL/min. The interface with the MS was set at 290 °C and MS performed in full scan mode using 70 eV EI + and scanned from 50 to 500Da. Retention time locking to ribitol was used according to Enfissi, et al., [42]. A mixture of n-alkanes, ranging from 8 to 32 carbons, was used for retention index external calibration. 1ml of the extraction solvent (Methanol) was used as the internal standard. Sample sets were run in batches of three replicates, Solvent Delay=3.00 min, Transfer Temp=250°C, Source Temp=220°C, Scan mass range: 50 to 500Da, Column was set at 30.0m x 250μm.
2.6.1. Data Processing
To identify chromatogram components found in white yam tuber profiles, a mass spectral library was constructed from in-built libraries; NIST ‘20M&R MS lib, Main lib, rep lib, OA_TMS_DB5_67min_V3 lib and FA_ME_SP2560_V3 lib. Component peak identification and spectral deconvolution was performed using the Automated Mass Spectral Deconvolution and Identification System (AMDIS v2.71, NIST); using Kovat’s retention indices (RI) and MS for identification according to the metabolomics reporting guidelines [1]. Each compound was assigned a representative ion and response areas were integrated and expressed relative to internal standard. Linearity was assessed by following the standard extraction procedure on amounts of material (10 mg) and the range further enhanced by taking the quantity of aliquot used (400 μ L). Recovery was assessed by expressing measurements from each extract of sample (10 mg) as a percentage of the total following three sequential extractions. All measurements for the developmental stages were conducted in triplicate and relative to internal standard.
2.6. Data Statistical Analysis and Visualization
Data were further processed in metaboAnalyst 5.0 platform; Row-wise normalization by the sum, logarithmic transformation and scaling by mean centering were performed to normalize the data (Figure S1a,b). All statistical analyses were perform using metaboanalyst software. Venn tool [43], was used to determine the relationship among between differential accumulated metabolites (DAMs) in the tubers of the two genotypes. One-way univariate ANOVA analyses were performed to determine the significant differentially accumulated metabolites across different data points at p-value threshold of (P≤0.05), and (Turkey’s HSD) was used to compare the means. Principal component analysis (PCA) was performed to determine the overall pattern of metabolites distribution in the two genotypes along the data points. In order to filter out the variables in the metabolites that were not related to categorical variables, and to obtain more reliable metabolite information on the structural of different data points, partial least squares – discriminant analysis (PLS-DA) was performed using variable importance in projection (VIP) at threshold of (VIP≤1). Agglomerative hierarchical cluster analysis was performed to classify the differentially accumulated metabolites (DAMs) across the data point using (euclidean and ward.D distance algorithm) and visualized it in clustering heatmap. The DAMs were mapped into PubChem, Kyoto encyclopedia genes and genomes (KEGG) and human metabolome data base (HMDB) to identify their functional groups. Metabolites set enrichment analysis (MSEA) and pathway annotation were performed to determine the significantly induced metabolisms that DAMs are involved in, during the studied yam dormancy stages. Pathway topology analysis was performed to visualize the significantly induced metabolic pathways.
3. Results
3.1. Metabolic Profiling of White Yam Tuber During Dormancy
A total of 949 metabolites were annotated in tubers of the two genotypes during the study period; 390 metabolites were annotated in the tubers of Obiaoturugo and 559 metabolites in annotated in the tubers of TDr1100873. Using Venny interactive tool (https://bioinfogp.cnb.csic.es/tools/venny/index.html) we compared the tuber metabolomes of the two genotypes and it showed that 158 metabolites were common between the genotypes (Figure 2a). Differentially accumulated metabolites (DAMs) in tubers of the two genotypes were also compared, and Figure 2b showed that 27 DAMs were conserved in the tubers of both genotypes, while 7 unique DAMs were observed Obiaoturugo and 5 unique DAMs were observed TDr1100873. It is believed that the phenotypic variation between the two yam genotypes with respect to dormancy duration was driven by these unique DAMs.
Analysis of variance (ANOVA) with Bonferroni correction identified 28 and 30 significantly differentially accumulated metabolites at (P≤0.05 and FDR ≤ 0.05) across the data points in the tubers of Obiaoturugo and TDr1100873 respectively (Figure 3a,b).
3.2. Principal Component Analysis (PCA) and Partial Least Square. Discriminant Analysis (PLS – DA) of Metabolites
Principal component analysis (PCA) was performed to determine the overview of yam tuber metabolome at the six data points of TDr1100873 and four data points of Obiaoturugo. Figure 4a shows the discriminating pattern of TDr1100873 tuber metabolome across the studied dormancy stages. The first principal component (PCI) clearly separated TDr1100873 tuber metabolome at 42-DAPM, 115-DAPM and 143-DAPM separated from the rest of data points and it accounts for 28.4% of total variation, while PC2 accounts for 21.4% of the total variation and it drives the clustering together of the tuber metabolome at 56-DAPM, 87-DAPM and 101-DAPM. This indicates that the later three dormancy stages shared similar metabolome, while, the former three dormancy stages were distinct. Though, 115-DAPM and 143-DAPM together constitute dormancy breaking stage, and were expected to share similarities in their metabolomes, but, PCA did not detect any similarity between them. The tuber metabolome at 115-DAPM which marks the appearance shoot buds (the first physical sign that tuber is getting ready for sprouting), differed substantially from the metabolome at 143-DAPM (sprouted tuber). We used PCA loading to identify the major metabolites contributing to the of tuber metabolome variations at these dormancy stages (Table S1). The metabolites playing key roles in tuber metabolome discrimination at different dormancy stages include; four esters, four amines, two alcohols, an alkaloid, fatty acid, lipid, coumarin, terpenoid, and one flavonoid. Similarly, Figure 4b shows Obiaoturugo tuber metabolome discriminating pattern across its four studied dormancy stages. The tuber metabolomes at 87-DAPM and 101-DAPM were clearly separated from that at 42-DAPM and 56-DAPM by PC1, and it account for 50.5% total variations. While PC2 accounts for 32.4% of total variations and drives the clustering together of 42-DAPM and 56-DAPM together. This is an indicative that Obiaoturugo tuber metabolome at these dormancy stages similar, implying that within the 14days time between the two sampling points no substantive metabolic change occurred. Whereas, the distance between the tuber metabolome at 87-DAPM and 101-DAPM indicates that tuber metabolomes at these two stages are clearly different from each other. 87-DAPM marked the appearance of shoot bud in the tubers of Obiaoturugo, while, 101-DAPM is its sprouted tuber metabolome. PCA loading revealed that the metabolites driving the discriminating of Obiaoturugo tuber metabolome at 87-DAPM and 101-DAPM were; five esters, three amines, three alcohols, lipid, fatty acid, coumarin, flavonoid, and a carbohydrate (Table S2).
We performed partial least squares – discriminant analysis (PLS – DA) to further access the performance of the discrimination of the tuber metabolome under supervised technique across the studied stages, using the very importance in projection (VIP) scores to evaluate the pattern of differential accumulation of important metabolites identified by PLS - DA across the dormancy stages (Figures S2 and S3). Figure 4c shows that the discriminating of TDr110873 tuber metabolome under supervised classification technique across the investigated dormancy stages. Component2 separated tuber metabolome at 115-DAPM and 143-DAPM from the metabolome at the rest of the dormancy stages, and it accounts for 16.8% of the total variations, while component 1 which account for 15.4% variation, separated the tuber metabolome at 101-DAPM from 42-DAPM, 56-DAPM and 87-DAPM. Figure 2e shows the discriminating pattern of Obiaoturugo tuber metabolome across the studied dormancy stages under supervised classification. It indicates that Obiaoturugo metabolome at each stage represents a distinct dormancy progression stage unlike the what was observed with PCA. The separations were driven by components 1 and 2 of the PLS-DA scores plot which account for 32.6% and 36.9% of the total variations respectively. across the studied dormancy stages exhibited consistent pattern in their separation into groups of clusters. The separation into distinct metabolome group or clustering into groups of similar metabolomes were determined mainly by the time duration between dormancy stages, and dormancy phenotype which necessitate change in metabolome composition.
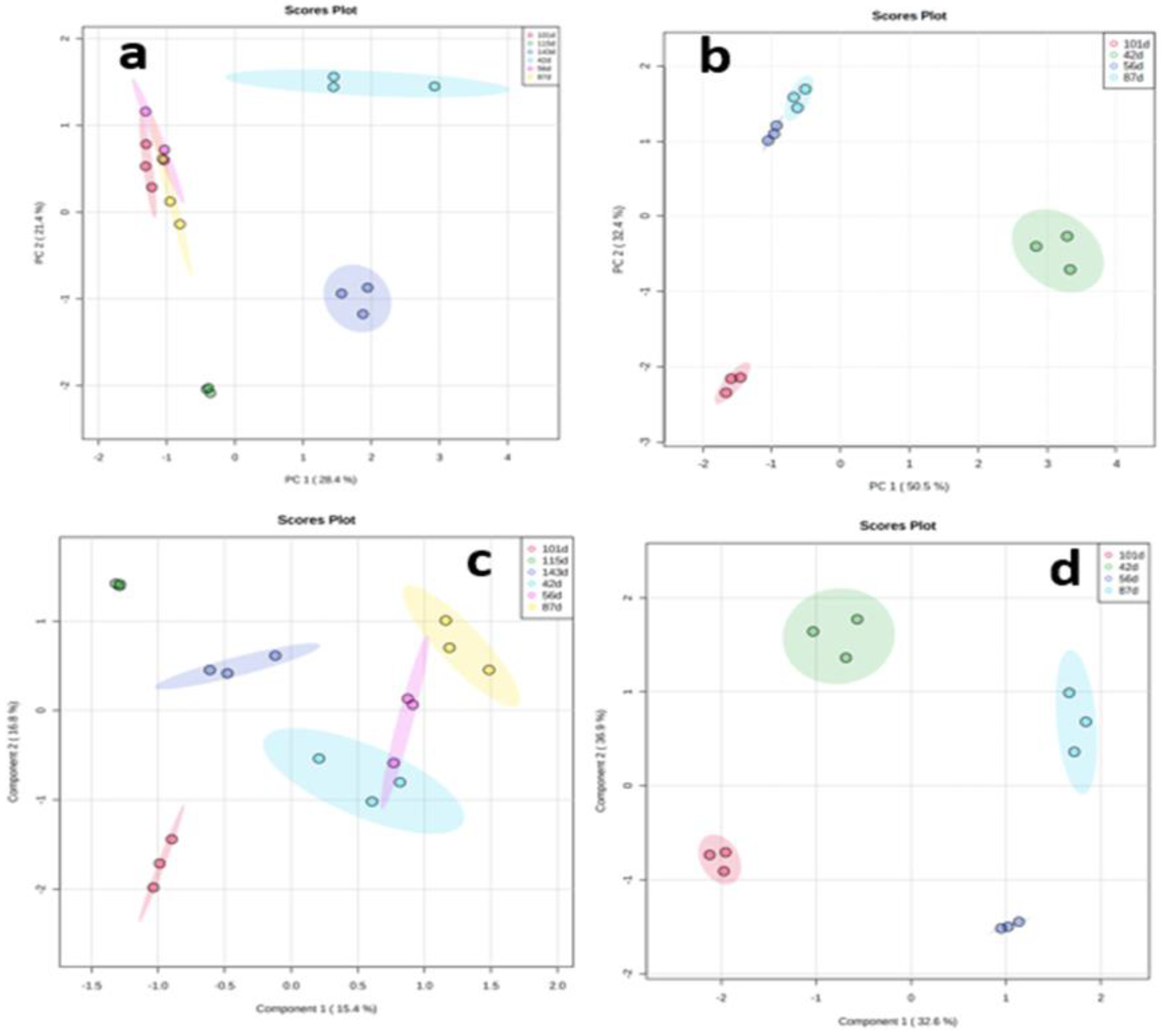
Figure 4.
(a–d) Scores plot between the selected Principal components of PCA and components of PLS-DA showing the discrimination of tuber metabolomes of the two genotypes across the dormancy stages studied, the explained variances are shown in brackets; (a) show the PCA of TDr1100873, (b) shows the PCA of Obiaoturugo, c shows the PLS-DA of TDr110873, while (d) shows the PLS-DA of Obiaoturugo.
Figure 4.
(a–d) Scores plot between the selected Principal components of PCA and components of PLS-DA showing the discrimination of tuber metabolomes of the two genotypes across the dormancy stages studied, the explained variances are shown in brackets; (a) show the PCA of TDr1100873, (b) shows the PCA of Obiaoturugo, c shows the PLS-DA of TDr110873, while (d) shows the PLS-DA of Obiaoturugo.
3.4. Identification of Differentially Accumulated Metabolites Across the Studied Tuber Dormancy Stages
To identify the differential accumulated metabolites (DAMs) across tuber dormancy stages studied, we performed agglomerative hierarchical clustering (AHC) analysis which partitioned DAMs earlier identified by ANOVA and PLS – DA VIP scores into the pattern of the exact change in their concentrations across the different dormancy stages using Euclidean distance measure and ward. D clustering algorithm. If a metabolite is playing a role in biological process, its concentration normally fluctuates during the process to adapt to the physiological changes in the process. First, AHC grouped the studied dormancy stages of TDr1100873 tuber into two main clusters; cluster 1 comprises of 143-DAPM, 115DAPM and 42-DAPM, cluster 2 comprised of 101-DAPM, 87-DAPM, and 56-DAPM (Figure 5a). In Obiaoturugo, the only distinct cluster comprises of 56-DAPM and 87-DAPM, while, 42-DAPM and 101-DAPM were separately linked to it (Figure 5b). This was a deviation from the normal three curves stages in germination process of botanic seed. Although, this was expected because the physical characteristics of dormant botanic seeds that influence its growth curve are existing in yam tuber. while botanical seeds are in desiccation state at maturity necessitating their requirement of imbibition during germination process, yam tubers at maturity are in hydrated state, and do not require imbibition during germination. Instead, as we reported earlier [44], it requires moisture loss to 40% threshold before dormancy breaking process can commence. This might be linked to this sharp deviation of yam tuber germination stages from well-established three curve stages of germination process in botanical seeds.
Agglomerative hierarchical clustering of the DAMs revealed the dynamic changes in the tuber metabolomes of the two genotypes (TDr1100873 and Obiaoturugo) across the studied dormancy stages. It identified 32 DAMs in tubers of TDr1100873, and 34 DAMs in tubers of Obiaoturugo (Figure 5a,b). Figure 5a shows that in TDr1100873, 8 DAMs which comprised of aromatic amine, (2) alcohols and polyols, organosiloxane, amino acid and derivatives, alkaloid, biogenic polyamine, coumarin were up-regulated, and 15 DAMs that comprised of nucleotide (Adenine), (2) alkaloids, amino acid and derivative, galactonolactone (sugar acid), flavonoids, fatty acid, (3) carboxylic acids, coumarin, (2) alcohols and polyols, ester, benzene derivatives and phytohormone, were down-regulated at 42-DAPM. At 56-DAPM, 9 DAMs which comprised of aromatic amine, (2) amino acids and derivatives, (2) alcohols, flavonoid, organosiloxane, biogenic polyamines and ester were up-regulated, and 7 DAMs that comprised of benzene derivatives, phenol, sugar (sucrose), benzoquinone, alcohol and polyol, ester, and carboxylic acid were down-regulated. At 87-DAPM, 10DAMs which comprised of fatty ester, biogenic polyamine, flavonoid, amines, alcohol, phenol, lipid, coumarin, fatty acid, benzene derivatives were up-regulated, and 7 DAMs that comprised of, alkaloids, (2) nucleotides (Adenine and nucleic acid), aromatic amine, alcohol and polyol, amino acid and derivatives, organosiloxane were down-regulated. At 101-DAPM, 6 DAMs which comprised of fatty ester, terpene, alcohol, (2) nucleotides (adenine and nucleotide sugar acid) and alkaloid, were up-regulated, alkaloids, and 7 DAMs that comprised of alcohol, flavonoid, amine, biogenic polyamine, alkaloid, phenol, phytohormone (salicyluric acid) down-regulated. At 115-DAPM, 7 DAMs which comprised of sugar (sucrose), benzoquinone, (2) carboxylic acids, fatty acid, lipid, phytohormone (salicyluric acid) were up-regulated, and 12 DAMs that comprised of fatty ester, biogenic polyamine, terpene, (2) amino acid and derivatives, (3) alcohols and polyols, (2) flavonoids, fatty acid, organosiloxane were down-regulated. At 143-DAPM, 12DAMs which comprised of (2) flavonoids, sugar (sucrose), (2) alkaloids, carboxylic acid, phytohormone (salicyluric acid), biogenic Polyamines, benzene derivatives, amino acid, benzoquinone were up-regulated, whereas, 7 DAMs that comprised of aromatic amine, alcohol, amino acids and derivatives, sugar acid, flavonoid, fatty acid, lipid were down-regulated.
Figure 5b shows that in Obiaoturugo, 15 DAMs which comprised of (3) amino acids and derivatives, (3) alcohols and polyols, (2) alkaloids, (2) carboxylic acids, (2) phytohormones (salicyluric and 3-methylsalicylic acids), sugar acid, amino alcohol, biogenic polyamine were up-regulated, and 14 DAMs that comprised of (2) carboxylic acids, (2) esters, (2) flavonoids, phenol, fatty acid, aromatic amine, organosiloxane, benzoquinone, down-regulated at 42-DAPM. At 56-DAPM, 7 DAMs which comprised of amine, (2) flavonoids, benzene derivatives, (2) nucleotides (adenine and nucleic acid), sugar (sucrose) were up-regulated, and 5 DAMs that comprised of ester, nucleotide (nucleotide sugar), alcohol, biogenic polyamine, and alkaloid were down-regulated. At 87-DAPM, 11DAMs which comprised of (2) nucleotides (nucleic acid and nucleotide sugar), benzoquinone, (2) esters, aromatic amine, fatty acid, carboxylic acid, flavonoid, alcohol, organosiloxane were up-regulated, and 4 DAMs that comprised of flavonoids, phytohormone (methylsalicylic acid), alkaloids, amino alcohol were down-regulated. At 101-DAPM, 8 DAMs which comprised of (2) flavonoids, benzoquinone, ester, carboxylic acid, alcohol organosiloxane and biogenic polyamines were up-regulated, and 9DAMs that comprised of phytohormone (methysalicylic acid), amino acid and derivatives, alcohols and polyols, sugar acid, inorganic oxide, (2) carboxylic acids were down-regulated. This result demonstrates that amines, biogenic polyamines, amino acids and derivatives, Alcohols, flavonoids, alkaloids, organosiloxane, phenols, esters, coumarins and phytohormone dominated the metabolic activities during dormancy period, whereas, fatty acids, lipids, nucleotides, carboxylic acids, sugars, amino acid (glutamine), terpenoids, benzoquinones, and benzene derivatives dominated the metabolic activities during tuber dormancy breaking and sprouted tubers of both yam genotypes. However, polyamine accumulated in the tubers of both genotype during the deep dormant stages and in sprouted tubers, indicating that it might be playing role in both dormancy induction and yam vine development.

Figure 5.
(a–b) Agglomerative Hierarchical Cluster heatmap of DAMs across the studied dormancy stages in each of the genotype. Metabolites and dormancy stages were separated into clusters using distance measure Euclidean distance measure, and clustering algorithms ward. linkage. The relative concentration of each metabolite at different studied dormancy stages was standardized, ranked using Spearman's rank correlation, and shown in the box according to the color criterion indicating their log fold change in concentration at each dormancy stage. The log fold change values according to the color criterion and dormancy stages are shown on the legends at right of the figures. (a) show the hierarchical cluster heatmap of DAMs in the tubers of in TDr1100873 across the studied dormancy stages, while (b) shows the hierarchical cluster heatmap of DAMs in the tubers of Obiaoturugo across the studied dormancy stages.
Figure 5.
(a–b) Agglomerative Hierarchical Cluster heatmap of DAMs across the studied dormancy stages in each of the genotype. Metabolites and dormancy stages were separated into clusters using distance measure Euclidean distance measure, and clustering algorithms ward. linkage. The relative concentration of each metabolite at different studied dormancy stages was standardized, ranked using Spearman's rank correlation, and shown in the box according to the color criterion indicating their log fold change in concentration at each dormancy stage. The log fold change values according to the color criterion and dormancy stages are shown on the legends at right of the figures. (a) show the hierarchical cluster heatmap of DAMs in the tubers of in TDr1100873 across the studied dormancy stages, while (b) shows the hierarchical cluster heatmap of DAMs in the tubers of Obiaoturugo across the studied dormancy stages.
3.3. Metabolites Mapping and Chemical Functional Groups Identification
To perform further analysis with the identified DAMs the compound labels or identity need to be standardized. The raw metabolites labels were used as queries in mapping DAMs into HMDB, PubChem, and KEGG data bases to identify their standardized compound names, chemical IDs and functional groups. It was observed that the DAMs were distributed across sixteen (14) major chemical functional groups, comprising of primary and secondary metabolisms. Alcohols and polyols were the most abundance DAMs, and it constitutes 12.82% proportion of the total DAMs, followed by fatty acid and esters, and amines and polyamines as each of them constitute 10.26% (Figure 6). Table 1 shows the conversion of the raw DAMs labels to standard compound names, their chemical functional groups and IDs in HMDB, PubChem and KEGG.
3.5. Metabolites Sets Enrichment Analysis and Pathways Annotation
We performed metabolite set enrichment analysis using hypergeometric test to determine whether metabolism of any set of DAMs is represented more than expected by chance during the studied yam tuber dormancy stages. It was found that 12 metabolisms were significantly enriched at (P≤0.05) during the studied yam tuber dormancy stages (Figure 7). The enrichment ratio of each of the induced metabolism denoted by the its bar length indicates the fold change in a particular metabolism from normal, while, the p-value indicates determines whether a particular metabolism has significant impact on the biological process under consideration. Linoleic acid metabolism recording the highest enrichment ratio of over 20 at (p = 0.02), an indicative that linoleic acid metabolism plays the most important role in yam tuber dormancy regulation. Whereas other metabolisms such as; galactose metabolism, biosynthesis of unsaturated fatty acids, pyrimidine metabolism, nitrogen metabolism, purine metabolism, D-glutamine and D-glutamate metabolism, Phenylalanine metabolism, alpha-linoleic acid metabolism, Arginine biosynthesis, Glycerolipid metabolism, and starch and sucrose metabolism playing some roles in yam tuber dormancy regulation.
3.6. Pathway Topology Analysis
Since the metabolic network is directed, we used the relative betweenness centrality of the metabolites in a node to determine the impact of pathway on yam tuber dormancy regulation. The result indicates that 7 metabolic pathways (linoleic acid metabolic pathway, Phenylalanine metabolic pathway, galactose metabolic pathway, starch and sucrose metabolic, Alanine, aspartate and glutamate metabolic pathway, purine metabolic pathway and lysine biosynthesis) had significant impact on yam tuber dormancy regulation, with linoleic acid metabolic pathway exacting maximum impact of 1 (Figure 8). Two pathways (Aminoacy-tRNA biosynthesis, and porphyrin and chlorophyll metabolism) were also observed to be suppressed during tuber endodormancy dormancy stages. Generally, the 7 significantly induced pathways can be grouped into three major functional categories, which include; lipids metabolism, energy and nucleotides metabolism, and amino acids metabolism. Lipids and energy nucleotides metabolisms were majorly involved yam tuber dormancy breaking activities and their metabolites were preponderance during tuber dormancy breaking stages. While amino acids metabolisms dominated the activities during yam tuber endodormancy dormancy stages. Table 2 presents the summary of all induced pathways, their log-fold change and impacts on yam tuber dormancy regulation.
Total is the number of compounds in the pathway, expected is the normal ratio value, hits is the matched number from the uploaded data, Raw p is the p-value before adjustment, Holm p is the P-value adjusted using Holm-Bonferroni method. FDR is p value adjusted using false discovering rate, Impact is the effect of the pathway on a biological process (yam tuber dormancy regulation).
4. Discussions
Dormancy in yam tuber is complex process that is synergistically modulated by molecular processes and environmental signals. Physiologically, during the period of dormancy to sprouting, yam tuber transit into three modes of nutrition; from heterotrophic to autotrophic and back to heterotrophic. Metabolites undoubtably play essential roles during these processes, since they contribute the raw materials for cellular structure composition, energy supply, biochemical reactions and signaling which are required for the completion of the physiological process of dormancy induction and breaking [18,28]. Usually, upon dormancy induction some specific metabolites are accumulated and some signals activated to maintain low energy condition in yam tuber that characterize dormancy period, whereas, during dormancy breaking, varieties of metabolites and metabolisms are activated to provide nutrients and energy required for tuber dormancy breaking [7,45]. However, elucidating the key metabolites and metabolic pathways that regulate yam tuber dormancy, to the best of our knowledge has not been done previously, although the functions of some metabolites in botanical seeds have been established [32,46,47,48]. Our study used untargeted metabolomics profiling to determine some key metabolites and their associated metabolic pathways that regulate yam tuber dormancy.
4.1. Efficiency of Metabolomics in Deciphering Molecular Mechanisms in a Biological System
In recent years, metabolomics has been frequently used in the study of seed dormancy [28,46,49]. In Davidia involucrate: a dove tree that is common in China, which has very recalcitrant seeds that stay in dormant state for about four years, metabolome profiling identified 48 differentially accumulated metabolites (DAMs) which were enriched in purine metabolism, pyrimidine metabolism, arginine and proline metabolism, flavone and flavonol biosynthesis, phenylpropanoid biosynthesis, arginine biosynthesis pathways, as well as key phytohormones, abscisic acid, indole-3 acetic acid and sinapic acid that were implicated in the seed dormancy regulation [32]. In rice, comparative metabolic profiling identified 74 candidate metabolites, among which 29 belong to ornithine-asparagine-polyamine module and the shikimic acid-tyrosine-tryptamine-phenylalamine-flavonoid module. The result revealed that shikimic acid promoted seed dormancy breaking [18]. In Arabidopsis, metabolomic and transcriptomic profiling identified two distinctive profiles involved in the metabolic regulation of seed dormancy breaking and seedling establishment [35]. The distribution of metabolites in two inbred lines (B73 and Mo17) of maize seeds revealed that the two inbred lines were highly differentiated in their metabolite profiles during dormancy breaking, with regard to amino acids, sugar alcohols, and organic acids accumulation, which determined their dormancy phenotype [50]. In the present study, we performed a comparative metabolomics profiling of tubers of two white yam genotypes at different time points during dormancy to dormancy breaking. We observed that a total of 39 metabolites were differentially accumulated in tubers of the two yam genotypes during the study period. The 39 DAMs were dominated by amino acids and derivatives, alkaloids and alcohols, sugars, nucleotides, amines and polyamines, fatty acids and esters, phenols and benzene derivatives, and organic acids. Therefore, this study demonstrated the efficiency of our metabolomic profiling approach in identifying differential accumulated metabolites, that revealed the dynamic physiological processes in white yam tubers from dormancy to sprouting.
4.2. Differentially Accumulated Metabolites Determined the Phenotypic Variation in Dormancy Duration of the Two White Yam Genotypes
Our data showed genotype-specific variation in differentially accumulated metabolites during tuber dormancy and dormancy break of the two white yam genotypes (TDr1100873 and Obiaoturugo). The tubers of TDr1100873 had 5 unique DAMs that spread between dormancy and dormancy break periods, and exhibited long dormancy phenotype. The 5 unique DAMs observed in the tubers of TDr1100873 comprised of terpenoids, amines, phenol and 2 fatty acids. Among the 5 unique DAMs observed in tubers of TDr110873, terpenoid was accumulated at 101-DAPM, amine accumulated at 56-DAPM, while phenol and one of the fatty acids accumulated at 87-DAPM, all these time points, except 101-DAPM constitute dormant period in TDr1100873. Obiaoturugo had 7 unique DAMs (nucleotide sugar, L-malic acid, amino alcohol, amine, phytohormone, carboxylic acid and amino acid and derivatives) that also spread between dormancy and dormancy break periods and exhibited short dormant phenotype. Among the 7 unique DAMs observed in the tubers of Obiaoturugo, nucleotide sugar, amino acid and carboxylic acid were all upregulated at 87-DAPM which is the dormancy breaking point in Obiaoturugo. The role of amines in dormancy regulation has been shown to be specie-specific. In some plant they promote dormancy breaking (germination), and in others, they induce dormancy, [46,51,52]. Its accumulation in the tuber of long dormant phenotype is indicative that it induced or at least function to prolong yam tuber dormancy. Study has shown phenol as an antioxidant that participate in the scavenging of reactive oxygen species (ROS) thus catalyzing oxygenation reactions through formation of metallic complexes, and inhibiting the activities of oxidizing enzymes [53]. It’s accumulation at the point when the tubers of short dormant genotype-initiated dormancy breaking this suggest it plays a vital role in the process. Conversely, the presence of unique nucleotide sugar and amino acid (glutamine) that accumulated at 87-DAPM which marked the endodormancy breaking point suggest that these metabolites play some roles in determining early dormancy breaking. It has been demonstrated that nucleotide sugar play regulatory role in cell division. A strong correlation was observed between the supply of Glc and the expression of cyclins (cycD2;1, D3;2, A3;2, and B1;2) [54,55], that the accumulation of nucleotide sugar in the tubers of short dormant phenotype might have triggered the early reactivation of cell division and consequently dormancy break. Similarly, it has been reported that L-Glutamine is direct precursor of Glutamate, and Glutamate occupies a central position in amino acid metabolism in plants and as well plays key role initiation of dormancy breaking process in seeds [56]. We infer that 2 (amine and phenol) out of the 5 unique DAMs in TDr1100873 tubers might be playing some roles in maintaining prolonged dormancy duration, whereas, 2 (nucleotide sugar and amino acid) out of the 7 unique DAMs in Obiaoturugo tubers might be playing some roles in early dormancy break. Our results agreed with the finding of [57] who reported that composition and changes in concentration of metabolites in the seeds of two wild barley. Our study buttresses the fact that variation in duration of tuber dormancy could be driven by the absence of nucleotide sugar and nucleic amino acid (L-Glutamine) in the tubers of long dormancy duration genotype (TDr1100873).
4.3. Metabolic Regulation of Yam Tuber Dormancy
Using three typical methods; ANOVA, PLS-DA and AHC, for comparative metabolome analysis at different time points during tuber dormancy of the two white yam genotypes, we identified 39 candidates metabolites associated with tuber dormancy regulation, distributed across 14 well-known dormancy regulating chemical functional groups such as: amines and biogenic polyamines, amino acids and derivatives, carboxylic acids, sugars, Alkaloids, alcohols and polyols, flavonoids, phenols and benzene derivatives, nucleotides, fatty acids and esters, lipids, coumarins and terpenoids, and phytohormones [7,26,58,59]. We did not identify any of the two established dormancy regulating phytohormones; Abscisic acid (ABA) and Gibberellic acids (GA) directly, although, their precursors were observed in some of the metabolic pathways enriched. Our data showed the dynamic changes in the concentrations of the identified DAMs across the studied dormancy stages of the tubers of the two white yam genotypes in response to different physiological processes that characterized each of the studied dormancy progression stage. Metabolites set enrichment analysis revealed that these DAMs were involved in 12 significantly induced metabolisms during white yam tuber dormancy-to-dormancy break period. These observations can be further grouped into amines and amino acids metabolisms, secondary metabolites metabolisms, energy metabolisms and nucleotides metabolisms.
4.4. Amines and Amino Acids Metabolisms
Generally, most of amines and amino acids were accumulated in the tubers of both yam genotypes during dormant stages, except for L-Glutamine that accumulated during dormancy breaking stage in the tubers of short dormant genotype. The preponderance of amines and amino acids during yam tuber dormancy stage is an indicative that the cells of yam tubers during dormancy were in post-replication phase, implying that replication which is the hallmark of cell cycle and consequently growth process was halted during this period. It has been reported that active dormancy induction is characterized by the arrest of cell cycle process at G1 phase [54]. Studies have also revealed that amino acids have prominent functions in plant growth and development, besides their role as protein building block, they as well play pivotal roles in biosynthesis of secondary metabolites, and during signaling processes in plant stress response [60,61]. These functions of amino acids, amines and polyamines have been shown to be specie-specific, in some plant they promote dormancy breaking (germination), and in others, they induce dormancy [52,62]. The molecular mechanism of exerting any of the dormancy regulatory action (either induction or inhibition) differs too. In dormancy induction it seems that amino acids and amines utilize the same molecular module that they use when they act as osmo-protectant during plant abiotic and biotic stress response and biosynthesis secondary metabolites. It has been demonstrated that amino acids have antioxidant property and act as reactive oxygen species (ROS) scavenger during plant response to stressful conditions [63]. ROS also serve as signaling molecules during plant growth and in the development and responses to biotic and abiotic stresses [64]. ROS had been shown to directly act on plant cell-wall polysaccharides by breaking their glycosidic bonds, which cause the cell wall to relax, thereby promoting cell division and elongation, which marks the breaking of endodormancy, [65]. During seed germination of dicotyledonous plants, such as cress and lettuce, ROS accumulation in their micropylar endosperm and radicle, led to endosperm weakening and radicle elongation, thereby promoting seed germination [66]. Certain cell-wall hydrolases and non-enzymatic substances, such as ROS and expansin, are needed for these processes [67]. Li et al., [68] have shown that ROS accumulated in the radicle and coleorhiza of rice seeds during germination and that ROS production and the number of germinated seeds decreased in the presence of ROS inhibitors, i.e., diphenyleneiodonium chloride and guazatine. We therefore infer that the ROS inhibition action of amino acids in yam tuber led to dormancy induction and maintenance, and hence they accumulated during tuber dormancy in the two-genotype studied. Previous studies have also made similar observations, in soybean (Glaycin max), high seed-specific expression of feedback-insensitive genes encoding dihydrodipicolinate synthase (DHPS) and aspartate kinase (AK) resulted in wrinkled seeds and low germination rates [69], while grain yield reduction was observed in high lysine crops [70]. Studies suggest that lysine also effect starch synthesis and endosperm development in maize (Zea mays Linn) and rice (Orayze sative) [70,71]. It mobilizes sucrose and other energy sources in a cell thereby leading to state of low-energy condition that characterize dormancy period. Also, seed mobilization of minor proteins such as seed maturation and dormancy-associated proteins that are degraded during imbibition to provide primary amino acids during germination have been reported [72,73]. In red rice, pyruvate in arginine-proline metabolism was induced in dormant seed, and implicated in seed dormancy induction and maintenance [74]. In Dove tree (D. involucrata Baill), the seeds that do not germinated after stratification treatment had the strongest pyruvate – arginine - proline metabolism induction, suggesting that this metabolic pathway might be a key contributor to dormancy induction [32]. In strawberry, polyamine in ABA-dominated, IAA and ethylene-participating crosstalk inhibited seed ripening and germination [75]. In the present study, our pathway topology analysis also revealed that arginine-proline was significantly induced during yam tuber dormancy (Table 2). Our pathway topology analysis also revealed that lysine metabolism leads to biosynthesis of secondary metabolites such as; tropane, piperidine and pyridine alkaloids, which are known for dormancy induction and maintenance in seed.
Polyamine (putrescine) has also been implicated in other plant physiological processes such as senescence induction, and stress tolerance [76,77]. It has been reported that senescence induction in yam crop is the physiological marker of dormancy onset [7], hence, it is reasonable to hypothesize that putrescine can induce tuber dormancy through the same molecular module that it uses to induce senescence. In the current study, putrescine accumulated in the tubers of both yam genotypes during dormancy stage, and was suppressed as dormancy progresses towards breaking (germination). Phenylalanine is another important amine which accumulated during dormancy in the present study. Although, the aromatic amine acts as a nitrogen source participate in activation of cellular processes that induce germination, but its metabolic pathway is also a hub for biosynthesis of diverse secondary metabolites, most of which counter its own growth inducing action [78,79]. Study has also demonstrated that upregulation of alanine aminotransferase (AlaAT) isoenzymes encoded by long and short dormancy alleles differ in barley seed embryo led to exhibition of different dormant phenotypes [80] The reduced dormancy allele Qsd1 mutant enhanced early and uniform germination, whereas long dormancy allele qsd1 mutant exhibited prolonged dormancy. In the current study, pathway topology analysis revealed that phenylalanine accumulated during tuber dormancy stages, suggesting that the metabolite plays role in tuber dormancy induction. Our pathway topology analysis also revealed that phenylalanine was significant induced during yam tuber dormancy, and it exerted significant impact on yam tuber dormancy regulation.
Our result contrasted several studies that have demonstrated that majority of amino acids and amines promote dormancy break (germination), and inhibit dormancy induction and maintenance in botanical seeds [52,81,82]. Ornithine and arginine are both nitrogenous amino acids that can form polyamines through decarboxylation [18]. In Canola (Brassica napus), it was demonstrated that tyramine differentially accumulated in osmoprimed seeds under saline stress, and led to early seed germination [83]. Llebres, et al., [84] identified the genes involved in arginine metabolism in Pinus pinaster Ait and found that arginine plays important role in seed germination in Pinus pinaster. In cotton, nitrogen flow occurs in seed during seed formation and germination through asparagine —> arginine —> storage protein —> arginine —> asparagine sequence [85]. The shikimic acid–tyrosine–tryptamine–phenylalanine–flavonoid module is closely related to the pathway of phenylalanine, tyrosine and tryptophan biosynthesis along with biosynthesis of phenylpropanoids. This module is also well known for its involvement in plants defense system, especially shikimic acid (shikimate). Some of the members of this pathway have been reported to be involved in seed germination. In legume seeds, tryptamine was found to be the main biogenic amine detected, and its concentration considerably increased during the germination process [86], suggesting that it might be playing role in dormancy breaking. These aforementioned findings contradict the present result, because tyramine accumulated during yam tuber deep dormancy stage in both genotypes, suggesting that it might be playing role in yam tuber dormancy induction. We also identified one amino acid (sarcosine) that accumulated during yam tuber dormancy, but has not been reported to have any role in plant growth regulation.
Our result demonstrated that amines and amino acids’ role in dormancy induction and maintenance in yam tuber is due to their antioxidant property, and action of secondary metabolites synthesized in their biosynthetic pathways (Figure 5). The contrast between our result and some of the previous reports of amines and amino acids actions in botanical seeds dormancy regulation could possibly be explained by two reasons namely; (1) The presence of seed coat and requirement of imbibition for dormancy break in botanical seeds, and non-requirement of imbibition for yam tuber dormancy break. In addition to developmental pathways in zygotic tissues, the seed coat plays a critical role in the control of dormancy in botanical seeds of many species. In some seeds, dormancy is known as coat imposed, and dormancy can be removed or reduced by simple removal of or damage to the seed coat. In various species the seed coat has been shown to restrict germination by regulating permeability to either water [87,88], oxygen [89] or germination inhibitors that leach from the seed coat into the embryo [90]. It has been reported that aromatic amines, and amino acids reduced the thickness and increased seed tegument porosity which helped in water imbibition and boost seed germination rate [91]. whereas in yam tuber, the reverse is the case, as we earlier reported that yam tuber required moisture reduction to critical threshold of 40% before dormancy breaking process could be initiated [44]. Study has also demonstrated that phenolics, aromatic amines and amino acids in exerting their antioxidant property inhibit enzymes that generate ROS in a cell system [68], in doing this, they might also inhibit enzymes that catalyze the removal of cell cycle arrest at G1 phase, thereby preventing the resumption of mitotic cell division which mark endodormancy break. (2) The botanical seed has a well-organized embryonic system in which dormancy regulation is processed, whereas, yam tuber has a continuous layer of meristematic cells beneath the dried tuber skin in which tuber dormancy regulation is processed, and the basis of yam tuber dormancy on this structure has been explained by Nwogha, et al. [7]. These postulations however need to be further validated through more studies.
4.5. Energy Metabolism
It is believed that yam tuber dormancy induction is an adaptive response to low energy condition due to stoppage of supply of sugar from photosynthesis as result of vine senescence at tuber physiological maturity [44]. Since photosynthetic sugar supply and mineral uptake systems are not active during yam tuber dormancy, the energy needs for low and high physiological activities during tuber dormancy is supplied from reserved energy-source metabolites in the tuber [45]. In the precent study, several energy source metabolites such as; (sucrose, galactose, carboxylic acids, malic acid, fatty acids, esters, and lipids) accumulated at different stages of tuber dormancy. The role of sugar signaling in dormancy regulation has been reported [92]. Based on changes in carbohydrate metabolism, it was suggested that sugar signaling is involved in regulating bud dormancy in non-woody perennial specie leafy spurge [93]. In woody perennials, bud dormancy is associated with gene expression pattern in typical manner of carbon starvation, while bud burst in spring depends on sucrose availability, and carbohydrate import from a distant wood parenchyma [94,95]. Expression of an Arabidopsis sucrose-phosphate synthase gene resulted in earlier bud dormancy break in spring transgenic and hybrid poplar (Populus alba x Populus grandidentata) [96]. Wengler [92], reported that carbohydrate mobilization for bud dormancy break may depend on interactions between the clock and sugar signaling. While in rice seeds, study demonstrated that the amount of total storable sugar does not affect the seed dormancy duration, but a mixture of sucrose and raffinose, and thus hypothesized that the ratio of oligosaccharides to sucrose may be the determinant of seed dormancy duration [49,97]. In our study, sucrose accumulated in the tubers of short dormant genotype (Obiaoturugo) at 56-DAPM, whereas, it accumulated in the tubers of long dormancy duration genotype (TDr1100873) at 115-DAP. The 56-DAPM represents 28days before appearance of shoot bud in the short dormant genotype (Obiaoturugo), whereas, 115-DAPM was the point of appearance of shoot bud in long dormant genotype (TDr1100873). This is an indicative that the stored starch might have been mobilized into sucrose at the onset of dormancy breaking process to provide the required energy to reactivated cellular activities towards tuber dormancy break. our result is an agreement with the findings of [94,95] and the report of Wengler; [92]. Also, we earlier reported that biochemical profiling of non-structural sugar from yam tuber dormancy to dormancy break revealed that quantity of nonreducing sugar which include sucrose increased up to 9 and 10 folds in the tubers of TDr110073 and Obiaoturugo respectively and that this increase started from 56-DAPM in tubers of Obiaoturugo and 101-DAPM in tubers of TDr1100873 [44]. This Indicates that sucrose play role in the early cellular activities culminating in dormancy break in the tubers of both genotypes.
The tricarboxylic acid (TCA) cycle intermediates, L-malic acid was observed in the tubers of Obiaoturugo (the short dormancy duration genotype), but not in the tubers of TDr110873, and surprisingly, it accumulated at 42-DAPM only. As dormancy progressed towards breaking L-malic acid was downregulated to almost non-existence level. However, TCA cycles precursors and intermediates such as, carboxylic acids, esters, lipid and fatty acids accumulated from 28days before appearance of shoot bud to dormancy break stage in the tubers of both genotypes. This indicates that TCA cycle plays essential role in energy metabolism during yam tuber dormancy breaking. Our pathway topology analysis also revealed that TCA cycle pathway was induced during yam tuber dormancy progression stage (Table 2). TCA cycle is one of central energy metabolic pathways in biological system, as well as being the biosynthetic pathway of many metabolites [98]. The plant TCA cycle has a set of eight enzymes primarily linking to the oxidation of pyruvate and malate (generated in the cytosol) to CO2 with the generation of nicotinamide adenine dinucleotide reduced form (NADH) for oxidation by mitochondrial respiratory chain [99], and subsequently releases the energy required to reactivate cell cycle, which had been arrested at G1 phase during dormancy induction. It has been demonstrated that accumulation of organic acids, particularly TCA cycle intermediates in plants supports many cellular processes, which are species, tissue and developmental stage specific [100]. In plants, lipids are the major component of cell membranes, and are used as a compact energy source for seed germination [101]. This was further supported by our enrichment and pathway topology analyses, which revealed that linoleic acid (lipid) metabolism was significantly induced with fold change ratio above 20 (Figure 7), and its pathway exerted maximum impact of 1 on yam tuber dormancy regulation (Table 2). We therefore, infer that lipids, carboxylic acids, fatty acids, and esters were metabolized through Embden-Meyerhof -Parnas (EMP), glycolysis, pyruvate, and TCA cycle pathways to generate the required energy pool for yam tuber dormancy break. It is noteworthy to mention that linoleic acid metabolism is very essential for white yam tuber dormancy break, and therefore can be a target metabolite in genetic manipulation of white yam tuber dormancy duration. Previous studies have also demonstrated that pathways in carbohydrate metabolism and ATP production such as; glycolysis, TCA cycle, EMP pathway, and oxidative phosphorylation were accumulated at the end of endodormancy to produce energy for bud sprouting and as well as precursors for amino acids biosynthesis [46,102]. Put together our results demonstrated that sucrose, carboxylic acids, esters, lipids fatty acids with exception of (octadecadienoic acid) provided the energy required for yam tuber dormancy breaking, with lipid metabolism and its pathway exerting strongest regulatory impact on the yam dormancy.
4.6. Nucleotides Metabolism and Cellular Processes
Four nucleotides comprised of L-Glutamine (nucleic amino acid), Adenine, nucleotide-sugar, and sugar acid were identified as part of DAMs in our study. Two of these nucleotides (L-Glutamine and nucleotide sugar) were found only in the short dormancy duration genotype (Obiaoturugo), whereas, Adenine and sugar acid were found in both genotypes. Adenine accumulated in the tubers of Obiaoturugo at 56-DAPM and in the tubers of TDr1100873 at 101-DAPM. Note; that due to short dormancy duration of the tubers of Obiaoturugo, 56-DAPM in the tubers of Obiaoturugo is physiological equivalent to 101-DAPM in the tubers of TDr1100873. Hence, in physiological timing of Adenine accumulation in the tubers of both genotypes is actually the same time, and it marked the point of commencement of molecular activities towards tuber dormancy break in tubers of both genotypes. Hence, the accumulation of adenine in the tubers of both genotypes at their respective stages suggests that adenine plays key role in reactivation of cellular processes that led to dormancy break irrespective of genotypic variation. Nucleotide sugar and nucleic amino acid (L-Glutamine) all accumulated at 87-DAPM which marked the appearance of shoot bud in the short dormancy duration genotype, suggesting that these two metabolites play key role in a later cellular processes that hastened dormancy breaking process in the tubers of Obiaoturugo. Previous study has demonstrated that L-Glutamine is direct precursor of Glutamate, and Glutamate occupies a central position in amino acid metabolism in plants and as well plays key role initiation of dormancy breaking process in seeds [56]. Glutamate, glutamine, aspartate and asparagine are the central regulators of carbon/nitrogen metabolism and they interact with multiple metabolic networks to reactivate the cell cycle process [103,104]. Glutamine and glutamate are precursors of nitrogenous compounds in plants, and the production of glutamate form glutamine is a key point in the synthesis of a variety of organic molecules, such as nucleic acids, amino acids and secondary metabolites [103]. We infer that L-glutamine could be a biomarker for early yam tuber dormancy breaking, and can be a target metabolic candidate in genetic manipulation of yam tuber for short dormancy duration.
Two nucleotide sugars (nucleotide sugar and sugar acid) were identified in this study and both of them accumulated at onset of dormancy breaking stage. Studies have demonstrated regulatory role of sugar in cell division. A close correlation was observed between the supply of Glc and the expression of cyclins (cycD2;1, D3;2, A3;2, and B1;2) [54,55]. The D-type cyclins have been reported to act as a sensor of external conditions within the cellular environment, and are associated with cyclin-dependent kinase (CDKA) to regulated cell cycles [105]. It has also been reported that the regulatory effect of Glc on the rate of cell division is primarily due to signaling rather than nutrient availability and energy status, as cell proliferating activity positively correlated with endogenous hexose levels, but not their uptake rate [54,55]. Implying that nucleotide sugar is not functioning as energy source, but rather as signaling to reactivate and mobilize other molecular machineries towards dormancy breaking process. These reports are in agreement with our data which shows that nucleotide sugar accumulated in the tubers of short dormancy duration genotype (Obiaoturugo) at onset of dormancy breaking activities. Sugar acid (galactonolactone) is a precursor of a nucleotide amino acid (ascorbate). Plant has evolved an efficient multi-components antioxidant defense system to protect vital molecules from the damage of mitochondrial generated ROS, as a consequence of activity respiratory chains [106]. This antioxidant defense system includes both enzymatic and non-enzymatic element [107]. One of the most important non- enzymatic members of this system is the water-soluble antioxidant, ascorbate [106]. Paradiso, et al. [108] demonstrated that Ascorbate biosynthesis improved wheat seed kernel development and delayed programmed cell death (PCD), thereby delaying senescing by feeding the plant with its immediate precursor L-galactone-γ-lactone (GL). We infer that galactose accumulation at early stage of cellular activities towards dormancy break in the current study suggest that it plays double roles during yam tuber dormancy breaking process. 1) it acted as precursor in biosynthesis of ascorbate that scavenged ROS generated during the reactivation of cell cycle. 2) Galactonolactone also promoted growth and protected the immature cells generated during cell division from being oxidized by ROS, which culminated in dormancy break. Our result showed that adenine accumulated in the tubers of both genotypes at the onset of dormancy break, 42days and 45days before physical germination of tubers of TD1100873 and Obiaoturugo respectively (Figure 5a,b). This indicates that non de novo synthesis of purine from adenine might have taken place, and synthesis of purine positively regulated the release of G1 phase of cell cycle that was arrested during yam tuber dormancy. The release of the arrested G1 phase of cell cycle marks the release the release of yam tuber endodormancy, and this also coincides with the 40days plus that has been reported to be the duration between yam tuber endodormancy release and eco-dormancy release [7]. We postulate that adenine is the metabolic marker for yam tuber endo-dormancy break, whereas, L-glutamine and nucleotide sugar are the potential candidates’ metabolites for early tuber dormancy breaking. In yam, these three candidate metabolites hold promise of metabolic manipulation of yam crop for a desirable tuber dormancy duration. Therefore, we hypothesize that nucleotide sugar (glucose) might have acted as a signal that triggered the accumulation of L-glutamine, which function together with the accumulated adenine to increase the nitrogen/carbon ratio (a critical factor in energy metabolism and DNA synthesis) in short dormancy duration genotype (Obiaoturugo), thereby leading to its tuber earlier dormancy break.
4.7. Secondary Metabolites and Phytohormones Metabolism
Secondary metabolites play key roles in plant growth and developmental processes such as; cell division, hormonal regulation, photosynthetic activity, radicals’ homeostasis, modulation of seeds transition to quiescent state at maturity, environmental signals transduction, nutrient mobilization and mineralization, stress tolerance, plant-soil, and environmental interactions [109,110,111]. In the current study, diverse secondary metabolites that accumulated at different yam tuber dormancy progression stages include; flavonoids, coumarins, terpenoids, alkaloids and alcohols, benzoquinone and benzoic derivatives, phenols, and salicylic acid (hormone). In the tubers of long dormant genotype (TDr1100873) the 3 flavonoids that were detected accumulated during tuber dormancy at (42-DAPM, 56-DAPM) and on sprouted tuber (143-DAPM), whereas, in the tubers of short dormant genotype (Obiaoturugo), they accumulated from onset of dormancy break (56-DAPM, 87-DAPM), and on sprouted tuber (101-DAPM). This suggests that in the tubers of long dormant genotype flavonoids play role in dormancy induction and maintenance, whereas in short dormant genotype they promote dormancy break (germination). This also explained the variation between the two genotypes with respect to tuber dormancy duration. The flavonoids accumulation in the sprouted tubers indicates that, it might be playing role also in vine elongation. It has been reported that flavonoids play different roles in different plant species, at different plant growth stages, organs and tissues with regards to growth regulation [112]. Studies have demonstrated that in botanical seeds, flavonoids induce and prolong seed dormancy [28,58,110,113]. It has been shown that flavonoids induce dormancy by inhibiting reactive oxygen species (ROS), through; suppression of singlet oxygen, inhibition of enzymes that generate ROS, such as (cyclooxygenase, lipoxygenase, monooxygenase and xanthine oxidase), chelation of ions transition which catalyze ROS production, suppressing the cascade of free-radicals in lipid peroxidation and recycling of other antioxidants such as amino acids and polyamines [114,115,116]. This agreed with our data on the tubers of long dormant genotype, but varies with data on the tubers of short dormant genotype. The opposing role of flavonoids in the tubers of short dormant genotype compared to long dormant genotype and botanical seeds presents a frontier for exploration of new role of flavonoids in dormancy regulation.
Similarly, coumarins also accumulated in the tubers of long dormant genotype during dormant stage (87-DAPM), and in the tubers of short dormant during the onset of dormancy breaking stages (56-DAPM and 87-DAPM). This also indicates that coumarins prolonged dormancy in the tubers of long dormant genotype, but induced dormancy breaking process in the tubers of short dormant genotype. Studies have demonstrated that coumarins induced both primary and secondary seed dormancy [117,118,119]. It has been reported that coumarins induce dormancy due to its ability to antagonize gibberellins and auxin’s function, while synergizing with abscisic acid (ABA) [118,120,121]. Our result on the tubers of long dormant phenotype with regard to coumarins metabolism is supported by this model. Conversely, it has been reported that plants show different response depending on the species, size, growth stage, and process of cellular respiration and associated-enormous changes in the mitochondria [122]. This may explain the dormancy breaking effect of coumarin observed on the tubers of short dormant genotype in the current study. It has been shown that secondary metabolite can also serve as energy source for cellular processes, when degraded [113]. In the light of this, we hypothesize that there could be a mechanism in the tubers of short dormant genotype that inhibits dormancy induction action of secondary metabolites, and subsequently degrade them at onset of dormancy breaking to provide additional energy source required to fuel the cell cycle reactivation process, thus leading to their accumulation during this period. It also could be as a result of differential accumulation quantity, because it has been reported that extreme high quantity of phenolic compounds including coumarins such as; free (1042%), bound (120%) and total phenolic acid content of (741%) in canary grass seed promoted germination [123].
Terpenoid was detected only in the tubers of long dormant phenotype, and it accumulated at the onset of dormancy breaking activities (101-DAPM). The accumulation of terpenes at 101-DAPM is indicative that it might be playing some role in cellular primary metabolism that is involve in cell cycle reactivation. Although, direct involvement of terpene in dormancy or plant growth regulation is sparsely reported in the literature. However, terpenes play vital roles in many other plants physiological processes especially in protection against external invaders, and response to environmental signals [111]. In the recent, it has been reported that terpenes also participate in plant primary metabolism [124]. There is also report that most dormancy regulating phytohormones belong to one terpenoid group or the other, for instance, gibberellins belong diterpenoids, brassinolide belongs to triterpenoids, abscisic acid and strigolactones belong to tetraterpenoids (carotenoids), cytokinin belongs to isoprene and is also known as primary metabolism terpene [111]. In the light of this, it is reasonable to postulate that the terpene that accumulated in the tuber of long dormant genotype at the onset of dormancy break stage in this study might be precursor of one of these hormones especially the gibberellins and cytokinin which are known for their role in dormancy breaking and promotion of activities towards germination. Benzoquinone and benzene derivative also accumulated from the onset dormancy breaking activities to sprouted tubers in both genotypes, an indication that these metabolites are implicated in yam tuber dormancy breaking process. Phenol accumulation was recorded only in the tubers of long dormant genotype, at its mid dormancy stage (87-DAPM). Study has shown that phenol as an antioxidant, also participate in the scavenging of reactive oxygen species (ROS), catalyzing oxygenation reactions through formation of metallic complexes, and inhibiting the activities of oxidizing enzymes [53], thereby inducing prolong dormancy. The absence of phenol in the tubers of short dormant genotype, and its cumulation in the tubers of long dormant genotype at 87-DAPM, when endodormancy was broken in the short dormant genotype is indicative of the phenol role in yam tuber dormancy elongation.
In the tubers of short dormant genotype, the two alkaloids detected in this study accumulated during dormancy stage at (42-DAPM), whereas, in the tubers of long dormant genotype one (galactosylglycerol) accumulated at dormant stage (42-DAPM) and in sprouted tuber. Isopyridoxal accumulated at dormancy break stage and in sprouted tuber. This result demonstrates alkaloids play diverse roles in plants growth and developmental process that depend on specie, plant variety, and growth stage. Hence, in the tubers of short dormant genotype it induced dormancy, and did not play any role in vine development. In the tubers long dormant genotype, galactosylglycerol played role in dormancy induction, and also show to be playing some role vine development. 5 Alcohols detected in this study all accumulated during early dormancy stage in the tubers of both genotypes, although, 2 of the alcohols accumulated in dormant and sprouted tubers of both genotypes. Our result shows that alcohols like other osmo-protectants are also involve in yam tuber dormancy induction, whereas the 2 that accumulated in sprouted yam tubers might have been mobilized to serve as energy additional source during the early stage of vine elongation. Studies have demonstrated that alcohols improved crops tolerance against abiotic stresses, possess antioxidant property, and promote germination and seedling development of botanical seeds [125,126,127]. The two hormones (salicylic acid and 3-Cresotinic acid) that were identified as DAMs in this study, all accumulated in the tubers of short dormant genotype during dormant stage at (42-DAPM), whereas, 3-Cresotinic acid was not found in the tubers of long dormant genotype. Salicylic acid accumulated in its tubers at dormancy break stage and in sprouted tuber. Salicylic acid has been classified as one of the phenolics compounds among the secondary metabolites, and has been reported to act in synergy with ABA to induce dormancy, while it functions alone to promote dormancy break [7,123].
4.8. Pathways Elucidations
Our pathway topology analysis revealed that linoleic acid has the strongest influence on yam tuber dormancy regulation process (Table 2). The pathway view shows that it is linoleic acid biosynthetic process regulated by Cytochrome P450 (CYP) genes family. CYPs have been implicated in Gibberellic acids (GAs) biosynthesis and response to environmental signal (light and photoperiod) during dormancy breaking in many botanical seeds [7], and GAs is one of the major plant growth regulatory hormones that promotes seed dormancy break, by inhibiting the action abscisic acid (ABA). ABA and GAs are the two major phytohormones that act in antagonistic manner to each other to induce and break dormancy respectively, in all crops. The ratio of ABA quantity to GAs quantity in seeds or any other plant tissue that has capability of being dormant has been reported to be the determinant factor of the seed or tissue dormant status [7], and the effect of this factor (ABA/GAs ratio) surpasses the effect of any other factors and molecular processes that regulate dormancy in plant system. In maize, CYP01A26 was reported to exhibit ent-kaurene oxidase activity which led to increased accumulation bioactive GAs and consequently result in early seed dormancy break [128,129] Similarly, in rice, 13-hydroxylation pathway that involves the activity of 13-hydroxlases which were coded by CYP714b1 and CYP714B2 genes led to early seed germination [130]. We therefore infer that the activities of CYP1A2, CYP2C, CYP2J, CYP2EI, CYP3A4, found in linoleic acid metabolic pathway might have led to increase biosynthesis of GAs and activities of GAs in the tubers, led to dormancy break. Although, we did not detect GAs in our study, probably because of the difference between metabolomic profiling and hormonal profiling protocols, however, some of its intermediates were detected which indicates its presence. The Cytochrome P450 (CYP) genes upregulation of linoleic acid biosynthesis can lead to inhibition of abscisic acids (ABA) dormancy inducing action, and increased the accumulation gibberellic acids (GAs) which consequently led to yam tuber dormancy break. Hence, we further infer that linoleic acid pathway is a putative metabolic pathway that can be exploited towards genetic manipulation of yam tuber dormancy. Our pathway topology analysis also revealed that phenylalanine metabolic pathway has significant impact on yam tuber dormancy regulation process. Phenylalanine pathway is a phenylalanine, tyrosine and tryptophan biosynthetic pathway, and was shown to be significantly induced during yam tuber dormancy (Table 2).
The pathway view shows that it is a hub for biosynthesis of secondary metabolite, amino acid derivatives and degradation of one ester that accumulated during yam tuber dormancy breaking stage (Figure 9). The secondary metabolites synthesized in this pathway accumulated during tuber dormancy stage, thus establishing a crosstalk link between secondary metabolites and amino acids and how their synergetic action leads to induction and prolonging of dormancy in yam tuber. The ester (Octanoic acid, 2-phenylethyl ester) that was degraded in this pathway eventually accumulated during the tuber dormancy breaking, implying that in addition to antioxidant property effects of secondary metabolites and amino acids, they might be antagonizing some tuber dormancy break promoting metabolites such as Octanoic acid, 2-phenylethyl ester, whose action promotes dormancy break. This further supports our earlier postulation that dormancy inducing effect of most of amino acids in yam tuber was partly as a result of secondary metabolites synthesis along their biosynthetic pathways. In plants, β-alanine is important for the synthesis of pantothenate and subsequently coenzyme A, which is an essential coenzyme in lipid and carbohydrate metabolism [131]. Also, glycolysis and TCA cycle have been reported to be linked to alanine aminotransferase during hypoxia induced waterlogging [132], confirming the role alanine metabolic pathway in stress tolerance, which is been linked to dormancy inducing-action. Moreover, in rice, alanine aminotransferase 1 encoded by flo12 gene was found to simultaneously regulate carbon and nitrogen metabolism, while the flo12 mutant exhibit a floury white-core endosperm [133]. Previous study has demonstrated that phenylalanine, tyrosine and tryptophan are not only essential components of protein synthesis, but are also located upstream of a number of growth hormones and secondary metabolites with multiple biological functions and health-promoting properties, such as protection against abiotic and biotic stress [134]. It has been further shown that phenylalanine is required for protein biosynthesis and cell survival during desiccation in botanical seed, and act as a precursor of a large number of multi-functional secondary metabolites, among which is a principal structural component in the supporting tissues of vascular plants and some algae [135]. Tyrosine has been reported to be the central hub to various specialized metabolic pathways, including vitamin E and plastoquinone which are essential metabolites of plant nutrition and antioxidant synthesis, as well as a precursor of specialized metabolites with diverse physiological functions such as protein, amino acids, attractants and defense compounds [136]. It has been demonstrated that tryptophan is an essential amino acid in synthesis of large number of bioactive molecules, such as auxin, tryptamine derivatives, phytoalexins, indole glucosinolates and terpenoid indole alkaloids, as well as playing pivotal role in the regulation of plant growth and development [61,62].
Two energy metabolic pathways (starch and sucrose metabolism and galactose metabolism) were significantly induced during this study (Figure 9b,c). These pathways are biosynthetic pathways of three important metabolites (sucrose, sugar acid and nucleotide sugar) that were identified as DAMs in this study. Interestingly, these sugars all accumulated at the onset of dormancy breaking stage in the tubers of both genotypes, except for nucleotide sugar which was detected only in the tubers of short dormant genotype, and has been earlier highlighted as the possible putative candidate metabolite driving the early dormancy break in that genotype. The significant induction and high impact of these pathways on yam tuber dormancy regulation process supports our earlier postulation that accumulation of these sugars at onset of tuber dormancy breaking is an indicative that they serve as an energy source for reactivation of cell cycle which marks the breaking of endodormancy in yam tuber. In addition to biosynthesis of these sugars, these pathways are also involve the pentose and glucuronate interconversions through UDP-glucose, pentose phosphate pathway, starch mobilization to amylose, galactosamine, D-glucose-6p, treholse-6p, all of which have been reported to play pivotal role in cell cycle reactivation, including working in synergy with auxin during dormancy breaking process, and as well serve as signaling molecules [37,108,137]. Alanine Aspartate and Glutamate metabolism and purine metabolism are another two pathways that were induced during our study period in yam tubers. They are mainly involved in biosynthesis of nucleotides such as adenine, L-glutamine, DNA precursor, DNA, RNA, histidine proteins and pentose phosphate. Adenine and L-glutamine are the two major metabolites in these pathways that were identified as DAMs in our study, and both accumulated at onset of dormancy break stage in both genotypes, which indicate that they play pivotal role in the array of early cellular events towards dormancy break such as; histidine unpacking, DNA synthesis and replication, synthesis of replicated cells components, including house-keeping RNA during cell division, these required central carbon and nitrogen metabolism. L-glutamine was detected only in the tubers of short dormant genotype, therefore, the induction and impact of its biosynthesis as revealed by pathway topology analysis supports our earlier inference that L-glutamine could be a putative metabolic marker determining the early yam tuber dormancy break phenotype. Put together, our results demonstrate that adenine and galactose (sugar acid) might be conserved putative metabolic for yam tuber dormancy break irrespective of dormancy duration phenotype. While, L-glutamine and nucleotide sugar are the two putative metabolic markers for yam tuber short dormant phenotype. Hence, alanine-aspartate-glutamine and purine metabolic pathways are some of the pathways that hold promise around metabolic manipulation of yam tuber dormancy.
5. Conclusions
This result demonstrates that yam tuber dormancy is a variable trait, and the two genotypes used in this study varied in the duration of their tuber dormancy. Tubers of TDr110873 exhibited long dormant phenotype, whereas, tubers of Obiaoturugo exhibited short dormant phenotype. This variation was driven by differential accumulation of terpenoid, phenol, amine and one fatty acid in the tubers of TDr110873, and differential accumulation of nucleotide sugar, amino acid (L-glutamine) and one carboxylic acid in the tubers of Obiaoturugo. The metabolomics changes from dormancy to sprouting in the tubers of both genotypes were systematically revealed, and a total of 39 differentially accumulated metabolites were identified across the investigated yam tuber dormancy stages of both yam genotypes. Amines and biogenic polyamines, amino acids and derivatives, Alcohols, flavonoids, alkaloids, phenols, esters, coumarins and phytohormone regulated yam tuber dormancy induction and maintenance, whereas, fatty acids, lipids, nucleotides, carboxylic acids, sugars, terpenoids, benzoquinones, and benzene derivatives regulated dormancy breaking and sprouting process in tubers of both yam genotypes. 12 metabolisms were significantly enriched during yam tuber dormancy and dormancy break process, and these were involved majorly in six metabolic (linoleic acid, phenylalanine, galactose, starch and sucrose, purine and alanine-aspartate-glutamate) pathways that exerted significant impacts on yam tuber dormancy regulation. Among these metabolic pathways, only phenylalanine metabolic regulated tuber dormancy induction, whereas, the rest five (linoleic acid, galactose, starch and sucrose, purine and alanine-aspartate-glutamate) pathways regulated dormancy break process, with linoleic acid metabolic pathway exerting strongest regulatory control. Therefore, linoleic acid pathway holds a promise and potential of being a putative metabolic pathway to be targeted in genetic manipulation of yam tuber dormancy. In addition, metabolites such as; adenine, sucrose and galactose (sugar acid) exhibited potentials of being conserved putative metabolic markers for yam tuber dormancy break irrespective of genotypic variation with respect to dormancy duration. Our results also demonstrated that nucleotide sugar and glutamine are the two potential metabolic markers for early tuber dormancy break, and therefore determined the dormancy duration phenotypic variation between the tubers of two yam genotypes studied. Comparative to the literature information on botanical seeds dormancy metabolic regulation mechanisms, our result demonstrated that yam tubers exhibit deviation, especially with regards to the roles of amino acids, amine and polyamines, and some secondary metabolites in dormancy regulations. Our result provides insights into molecular mechanisms regulating yam tuber dormancy, basis for further metabolomics investigation of yam tuber dormancy regulation mechanisms and future genetic manipulation of yam tuber dormancy.
Supplementary Materials
The following supporting information have also been submitted and can be downloaded at: Preprints.org, Figure S1a, Figure S1b, Figure S2, Figure S3: title; Table S1, Table S2.
Author Contributions
Conceptualization, Jeremiah S. Nwogha, and Jude E. Obidiegwu, and Cynthia A. Chilaka.; methodology, Muthurajan Raveendran., Jeremiah S. Nwogha and Cynthia A. Chilaka.; software, Jeremiah S. Nwogha; validation, Muthurajan Raveendran., Jude E. Obidiegwu. and Rohit Kambale.; formal analysis, Jeremiah S. Nwogha.; investigation, Jeremiah S, Nwogha.; Rohit Kambale and Cynthia A. Chilaka.; resources, Jude E. Obidiegwu and Muthurajan Raveendran; data curation, Jeremiah S. Nwogha, Rohit Kambale and Veera R. Rajagopalan; writing—original draft preparation, Jeremiah S. Nwogha.; writing—review and editing, Jude E. Obidiegwu and Abtew G. Wosene.; visualization, Jeremiah S. Nwogha.; supervision Abtew G. Wosene, Muthurajan Raveendran and Jude E. Obidiegwu.; project administration, Happiness O. Oselebe and Abtew G. Wosene.; funding acquisition, Happiness O. Oselebe and Abtew G. Wosene. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by GENES Intra-Africa Academic Mobility Project; “Mobility for plant genomics scholars to accelerate climate-smart adaptation options and food security in Africa (GENES)” funded by European Union. GENES Intra-Africa Academic Mobility Project. grant number EACEA/2017/2552. The APC will be funded by the authors.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the Supplementary Material here.
Acknowledgments
The authors acknowledge the field staff of yam research programme of NRCRI, Umudike Nigeria, especially Anthony Udeagbara, and staff of department Plant biotech- nology Tamil Nadu Agricultural University, Coimbatore, India for their technical support in field and laboratory experimentations respectively.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Price, E.J.; Wilkin, P.; Sarasan, V.; Fraser, P.D. Metabolite profiling of Dioscorea (yam) species reveals underutilised biodiversity and renewable sources for high-value compounds. Scientific reports 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam)—An appraisal of nutritional and therapeutic potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef]
- Mignouna, H.D.; Abang, M.M.; Asiedu, R. Harnessing modern biotechnology for tropical tuber crop improvement: Yam (Dioscorea spp.) molecular breeding. African Journal of Biotechnology 2003, 2, 478–485. [Google Scholar]
- Lev, L.S.; Shriver, A.L. A trend analysis of yam production, area, yield, and trade (1961-1996). Colloques-CIRAD 1998, 11–20. [Google Scholar]
- Dansi, A.; Dantsey-Barry, H.; Dossou-Aminon, I.; N’kpenu, E.; Agré, A.; Sunu, Y.; Kombaté, K.; Loko, Y.; Dansi, M.; Assogba, P. Varietal diversity and genetic erosion of cultivated yams (Dioscorea cayenensis Poir-D. rotundata Lam complex and D. alata L.) in Togo. Int. J. Biodivers. Conserv 2013, 5, 223–239. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://wwwfaoorg/ faostat/en/#data/QC. 2020.
- Nwogha, J.S.; Wosene, A.G.; Raveendran, M.; Oselebe, H.O.; Obidiegwu, J.E.; Amirtham, D. Physiological and Molecular basis of dormancy in yam tuber: A way forward towards genetic manipultion of dormancy in yam tubers. Global Journal of science frontier research 2022b, XXll, 47–73. [Google Scholar]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohy- drates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Yolou; Elie Idossou1 ADECHOKAN Hibath Audrey, A.; Didier, M.Z.J. Evaluation of yam (Dioscorea cayenensis–Dioscorea rotundata) seed germination grown in Centre Benin. International Journal 2015, 3, 277–284.
- Ile, E.; Craufurd, P.; Battey, N.; Asiedu, R. Phases of dormancy in yam tubers (Dioscorea rotundata). Annals of Botany 2006, 97, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Passam, H. Dormancy of yams in relation to storage. Yams. Ignames. 1982, 285–293. [Google Scholar]
- Hamadina, E.I.; Craufurd, P.Q. Changes in Free Phenolics Contents during Tuber Development, Dormancy and Sprouting in White Yam (Dioscorea rotundata Poir.). International Journal of Plant Research 2015, 5, 34–41. [Google Scholar]
- Shangguan, L.; Chen, M.; Fang, X.; Xie, Z.; Gong, P.; Huang, Y.; Wang, Z.; Fang, J. Comparative transcriptome analysis provides insight into regulation pathways and temporal and spatial expression characteristics of grapevine (Vitis vinifera) dormant buds in different nodes. BMC Plant Biology 2020, 20, 1–20. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Zabaras, D.; Frank, D.; Arcot, J. Effect of germination and fermentation on carbohydrate composition of Australian sweet lupin and soybean seeds and flours. Journal of Agricultural and Food Chemistry 2017, 65, 10064–10073. [Google Scholar] [CrossRef] [PubMed]
- González-Orenga, S.; Ferrer-Gallego, P.P.; Laguna, E.; López-Gresa, M.P.; Donat-Torres, M.P.; Verdeguer, M.; Vicente, O.; Boscaiu, M. Insights on salt tolerance of two endemic Limonium species from Spain. Metabolites 2019, 9, 294. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.-d.; Xie, Q.; He, Z.-h. Two faces of one seed: Hormonal regulation of dormancy and germination. Molecular plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Lyv, Y.; Zheng, W.; Yang, C.; Li, Y.; Wang, X.; Chen, R.; Wang, C.; Luo, J.; Qu, L. Comparative metabolomics reveals two metabolic modules affecting seed germination in rice (Oryza sativa). Metabolites 2021, 11, 880. [Google Scholar] [CrossRef]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Scientific Reports 2017, 7, 12620. [Google Scholar] [CrossRef]
- Kralj Cigić, I.; Rupnik, S.; Rijavec, T.; Poklar Ulrih, N.; Cigić, B. Accumulation of agmatine, spermidine, and spermine in sprouts and microgreens of alfalfa, fenugreek, lentil, and daikon radish. Foods 2020, 9, 547. [Google Scholar] [CrossRef]
- Sepúlveda, G.; de Jiménez, E.S. Polyamine distribution among maize embryonic tissues and its relation to seed germination. Biochemical and biophysical research communications 1988, 153, 881–887. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.; Rehman, H.u.; Hussain, M. Seed priming with polyamines improves the germination and early seedling growth in fine rice. Journal of New Seeds 2008, 9, 145–155. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Zhang, X.-Q.; Ma, X.; Huang, L.-K.; Yan, Y.-H. Exogenous spermidine improves seed germination of white clover under water stress via involvement in starch metabolism, antioxidant defenses and relevant gene expression. Molecules 2014, 19, 18003–18024. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, C.; He, F.; Li, Z.; Guan, Y.; Hu, Q.; Hu, J. Exogenous spermidine improves seed germination of sweet corn via involvement in phytohormone interactions, H2O2 and relevant gene expression. BMC Plant Biology 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Sibian, M.S.; Saxena, D.C.; Riar, C.S. Effect of germination on chemical, functional and nutritional characteristics of wheat, brown rice and triticale: A comparative study. Journal of the Science of Food and Agriculture 2017, 97, 4643–4651. [Google Scholar] [CrossRef]
- Gipson, A.B.; Morton, K.J.; Rhee, R.J.; Simo, S.; Clayton, J.A.; Perrett, M.E.; Binkley, C.G.; Jensen, E.L.; Oakes, D.L.; Rouhier, M.F. Disruptions in valine degradation affect seed development and germination in Arabidopsis. The Plant Journal 2017, 90, 1029–1039. [Google Scholar] [CrossRef]
- Desmaison, A.M.; Tixier, M. Amino acids content in germinating seeds and seedlings from Castanea sativa L. Plant physiology 1986, 81, 692–695. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, Y.; Zhan, Y.; Cao, X.; Liu, C.; Zhang, Y.; Gai, S. Metabolomics analysis reveals Embden Meyerhof Parnas pathway activation and flavonoids accumulation during dormancy transition in tree peony. BMC Plant Biology 2020, 20, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Jiao, H.J.; Faust, M. Changes in metabolic enzyme activities during thidiazuron-induced lateral budbreak of apple. HortScience 1991, 26, 171–173. [Google Scholar] [CrossRef]
- Halaly, T.; Pang, X.; Batikoff, T.; Crane, O.; Keren, A.; Venkateswari, J.; Ogrodovitch, A.; Sadka, A.; Lavee, S.; Or, E. Similar mechanisms might be triggered by alternative external stimuli that induce dormancy release in grape buds. Planta 2008, 228, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, H.; Wang, X.; Wang, B.; Zheng, X.; Shi, X.; Liu, W.; Liu, F. Respiratory changes during dormancy of grape buds. Sci Agric Sin 2013, 46, 1201–1207. [Google Scholar]
- Deng, S.; Xiao, Q.; Xu, C.; Hong, J.; Deng, Z.; Jiang, D.; Luo, S. Metabolome profiling of stratified seeds provides insight into the regulation of dormancy in Davidia involucrata. Plant Diversity 2022, 44, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Arc, E.; Chibani, K.; Grappin, P.; Jullien, M.; Godin, B.a.; Cueff, G.; Valot, B.; Balliau, T.; Job, D.; Rajjou, L. Cold stratification and exogenous nitrates entail similar functional proteome adjustments during Arabidopsis seed dormancy release. Journal of proteome research 2012, 11, 5418–5432. [Google Scholar] [CrossRef] [PubMed]
- Szczotka, Z.; Pawłowski, T.; Krawiarz, K. Proteins and polyamines during dormancy breaking of European beech (Fagus sylvatica L.) seeds. Acta Physiologiae Plantarum 2003, 25, 423–435. [Google Scholar] [CrossRef]
- Silva, A.C.d.; Suassuna, J.F.; Melo, A.S.d.; Costa, R.R.; Andrade, W.L.d.; Silva, D.C.d. Salicylic acid as attenuator of drought stress on germination and initial development of sesame. Revista Brasileira de Engenharia Agrícola e Ambiental 2017, 21, 156–162. [Google Scholar] [CrossRef]
- de Souza Vidigal, D.; Willems, L.; van Arkel, J.; Dekkers, B.J.; Hilhorst, H.W.; Bentsink, L. Galactinol as marker for seed longevity. Plant Science 2016, 246, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Baena-González, E.; Lunn, J.E. SnRK1 and trehalose 6-phosphate–two ancient pathways converge to regulate plant metabolism and growth. Current Opinion in Plant Biology 2020, 55, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR signaling promotes accumulation of BZR1 to balance growth with carbon availability in Arabidopsis. Current Biology 2016, 26, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Bledsoe, S.W.; Henry, C.; Griffiths, C.A.; Paul, M.J.; Feil, R.; Lunn, J.E.; Stitt, M.; Lagrimini, L.M. The role of Tre6P and SnRK1 in maize early kernel development and events leading to stress-induced kernel abortion. BMC Plant Biology 2017, 17, 1–17. [Google Scholar] [CrossRef]
- Figueroa, C.M.; Lunn, J.E. A tale of two sugars: Trehalose 6-phosphate and sucrose. Plant physiology 2016, 172, 7–27. [Google Scholar] [CrossRef]
- D. TIAMA1, N.S.; R. E. TRAORE1, M.Y.; P. BATIONO-KANDO; J. ZOUNDJIHEKPON2; SAWADOGO, M.; ZONGO1, J.D. Effect of chemical fertilizers on production of yams (nyù) of passore in farmers’ environment. Agronomie Africaine 2018, 30, 99–105. [Google Scholar]
- Enfissi, E.M.; Barneche, F.; Ahmed, I.; Lichtlé, C.; Gerrish, C.; McQuinn, R.P.; Giovannoni, J.J.; Lopez-Juez, E.; Bowler, C.; Bramley, P.M. Integrative transcript and metabolite analysis of nutritionally enhanced DE-ETIOLATED1 downregulated tomato fruit. The Plant Cell 2010, 22, 1190–1215. [Google Scholar] [CrossRef]
- Oliveros, J.C. VENNY. An interactive tool for comparing lists with Venn Diagrams. http://bioinfogp. cnb. csic. es/tools/venny/index. html 2007.
- S., N.J.; Abtew, W.G.; Raveendran, M.; Oselebe, H.O.; Obidiegwu, J.E.; Chilaka, C.A.; Amirtham, D.D. Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata). Agriculture (Switzerland) 2023, 13. https://doi.org/.
- Han, C.; Zhen, S.; Zhu, G.; Bian, Y.; Yan, Y. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination. Plant Physiology and Biochemistry 2017, 115, 320–327. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Dong, S.; Liu, Y.-L.; Li, Z.-H. The Opposite Roles of White Light in Regulating Germination of Fresh and Aged Seed in Tobacco. Plants 2021, 10, 2457. [Google Scholar] [CrossRef]
- Yang, M.; Yang, J.; Su, L.; Sun, K.; Li, D.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. Metabolic profile analysis and identification of key metabolites during rice seed germination under low-temperature stress. Plant Science 2019, 289, 110282. [Google Scholar] [CrossRef] [PubMed]
- Mazlan, O.; Aizat, W.M.; Baharum, S.N.; Azizan, K.A.; Noor, N.M. Metabolomics analysis of developing Garcinia mangostana seed reveals modulated levels of sugars, organic acids and phenylpropanoid compounds. Scientia Horticulturae 2018, 233, 323–330. [Google Scholar] [CrossRef]
- Yan, S.; Huang, W.; Gao, J.; Fu, H.; Liu, J. Comparative metabolomic analysis of seed metabolites associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiology and Biochemistry 2018, 127, 590–598. [Google Scholar] [CrossRef]
- Feenstra, A.D.; Alexander, L.E.; Song, Z.; Korte, A.R.; Yandeau-Nelson, M.D.; Nikolau, B.J.; Lee, Y.J. Spatial mapping and profiling of metabolite distributions during germination. Plant physiology 2017, 174, 2532–2548. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. Journal of Integrative Plant Biology 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Horikoshi, H.M.; Sekozawa, Y.; Kobayashi, M.; Saito, K.; Kusano, M.; Sugaya, S. Metabolomics analysis of'Housui'Japanese pear flower buds during endodormancy reveals metabolic suppression by thermal fluctuation. Plant Physiology and Biochemistry 2018, 126, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Weidner, S. Biological activity of grapevine phenolic compounds. Grapevine molecular physiology & biotechnology 2009, 389–405. [Google Scholar]
- Wang, L.; Ruan, Y.-L. Regulation of cell division and expansion by sugar and auxin signaling. Frontiers in plant science 2013, 4, 163. [Google Scholar] [CrossRef]
- Hartig, K.; Beck, E. Crosstalk between auxin, cytokinins, and sugars in the plant cell cycle. Plant Biology 2006, 8, 389–396. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. Journal of experimental botany 2007, 58, 2339–2358. [Google Scholar] [CrossRef]
- Huang, Y.; Cai, S.; Ye, L.; Hu, H.; Li, C.; Zhang, G. The effects of GA and ABA treatments on metabolite profile of germinating barley. Food Chemistry 2016, 192, 928–933. [Google Scholar] [CrossRef]
- Chen, C.; Zeng, L.; Zhao, H.; Ye, Q. Proteomic analysis of the early development of the Phalaenopsis amabilis flower bud under low temperature induction using the iTRAQ/MRM approach. Molecules 2020, 25, 1244. [Google Scholar] [CrossRef]
- Tiburcio, A.F.; Alcazar, R. Potential applications of polyamines in agriculture and plant biotechnology. Polyamines: Methods and Protocols 2018, 489–508. [Google Scholar]
- Ma, Z.; Bykova, N.V.; Igamberdiev, A.U. Cell signaling mechanisms and metabolic regulation of germination and dormancy in barley seeds. The Crop Journal 2017, 5, 459–477. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino acid catabolism in plants. Molecular plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, D.; Liu, Q. Connections between amino acid metabolisms in plants: Lysine as an example. Frontiers in Plant Science 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. International Journal of Molecular Sciences 2022, 23, 5186. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. The Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.P.; Ivakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.-M.; Stitt, M. The sucrose–trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. Journal of experimental botany 2014, 65, 1051–1068. [Google Scholar] [CrossRef]
- Muller, K.; Linkies, A.; Vreeburg, R.A.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant physiology 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Calle, V.; Barrero-Sicilia, C.; Carbonero, P.; Iglesias-Fernandez, R. R. Mannans and endo-β-mannanases (MAN) in Brachypodium distachyon: Expression profiling and possible role of the BdMAN genes during coleorhiza-limited seed germination. J. Exp. Bot. 2015, 66, 3753–3764. [Google Scholar] [CrossRef]
- Li, W.-Y.; Chen, B.-X.; Chen, Z.-J.; Gao, Y.-T.; Chen, Z.; Liu, J. Reactive oxygen species generated by NADPH oxidases promote radicle protrusion and root elongation during rice seed germination. International Journal of Molecular Sciences 2017, 18, 110. [Google Scholar] [CrossRef] [PubMed]
- Falco, S.; Guida, T.; Locke, M.; Mauvais, J.; Sanders, C.; Ward, R.; Webber, P. Transgenic canola and soybean seeds with increased lysine. Bio/technology 1995, 13, 577–582. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, C.; Wang, X.; Liu, Q.; Yuan, D.; Pan, G.; Sun, S.S.; Tu, J. Development of high-lysine rice via endosperm-specific expression of a foreign LYSINE RICH PROTEIN gene. BMC Plant Biology 2016, 16, 1–13. [Google Scholar] [CrossRef]
- Jia, M.; Wu, H.; Clay, K.L.; Jung, R.; Larkins, B.A.; Gibbon, B.C. Identification and characterization of lysine-rich proteins and starch biosynthesis genes in the opaque2mutant by transcriptional and proteomic analysis. BMC plant biology 2013, 13, 1–14. [Google Scholar] [CrossRef]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Okamoto, M.; Koshiba, T.; Kamiya, Y.; Nambara, E. Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: Epigenetic and genetic regulation of transcription in seed. The plant journal 2005, 41, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Gianinetti, A.; Finocchiaro, F.; Bagnaresi, P.; Zechini, A.; Faccioli, P.; Cattivelli, L.; Valè, G.; Biselli, C. Seed dormancy involves a transcriptional program that supports early plastid functionality during imbibition. Plants 2018, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Liu, X.; Sun, F.; Cao, J.; Huo, N.; Wuda, B.; Xin, M.; Hu, Z.; Du, J.; Xia, R. Wheat miR9678 affects seed germination by generating phased siRNAs and modulating abscisic acid/gibberellin signaling. The Plant Cell 2018, 30, 796–814. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Honda, C.; Moriguchi, T. Involvement of polyamine in floral and fruit development. Japan Agricultural Research Quarterly: JARQ 2006, 40, 51–58. [Google Scholar] [CrossRef]
- Nambeesan, S.; Avtar K., Handa; Mattoo, A.K. Polyamines and Regulation of Ripening and Senescence. In Postharvest Biology and Technology of Fruit, Vegetables and Flowers; Paliyath, G., Murr, D.P., Handa, A.K., Lurie, S., Eds.; Wiley-Blackwell, 2016. [Google Scholar]
- Feduraev, P.; Skrypnik, L.; Riabova, A.; Pungin, A.; Tokupova, E.; Maslennikov, P.; Chupakhina, G. Phenylalanine and tyrosine as exogenous precursors of wheat (Triticum aestivum L.) secondary metabolism through PAL-associated pathways. Plants 2020, 9, 476. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Y.; Ma, C.; Qin, J.; Nguyen, T.H.N.; Liu, D.; Gan, H.; Ding, S.; Luo, Z.-B. Phenylalanine as a nitrogen source induces root growth and nitrogen-use efficiency in Populus× canescens. Tree physiology 2018, 38, 66–82. [Google Scholar] [CrossRef]
- Sato, K.; Yamane, M.; Yamaji, N.; Kanamori, H.; Tagiri, A.; Schwerdt, J.G.; Fincher, G.B.; Matsumoto, T.; Takeda, K.; Komatsuda, T. Alanine aminotransferase controls seed dormancy in barley. Nature communications 2016, 7, 11625. [Google Scholar] [CrossRef]
- Das, A.; Kim, D.-W.; Khadka, P.; Rakwal, R.; Rohila, J.S. Unraveling key metabolomic alterations in wheat embryos derived from freshly harvested and water-imbibed seeds of two wheat cultivars with contrasting dormancy status. Frontiers in Plant Science 2017, 8, 1203. [Google Scholar] [CrossRef]
- Verslues , P.E., and Sharma , Sandeep. Proline Metabolism and Its Implications for Plant- Environment Interaction Authors : Published By : The American Society of Plant Biologists Proline Metabolism and Its Implications for Plant-Environment Interactio. In The Arabidopsis Book, 2010(8), https://doi.org/10.1199/tab.0140, Ed.; The American Society of Plant Biologists: 2010. [CrossRef]
- Lechowska, K.; Wojtyla, Ł.; Quinet, M.; Kubala, S.; Lutts, S.; Garnczarska, M. Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress Katarzyna. International Journal of Molecular Sciences 2022, 23. [Google Scholar]
- Llebrés, M.-T.; Pascual, M.-B.; Debille, S.; Trontin, J.-F.; Harvengt, L.; Avila, C.; Cánovas, F.M. The role of arginine metabolic pathway during embryogenesis and germination in maritime pine (Pinus pinaster Ait.). Tree physiology 2018, 38, 471–484. [Google Scholar] [CrossRef]
- Dilworth, M.F.; Dure III, L. Developmental biochemistry of cotton seed embryogenesis and germination: X. Nitrogen flow from arginine to asparagine in germination. Plant Physiology 1978, 61, 698–702. [Google Scholar] [CrossRef]
- Shalaby, A. Changes in biogenic amines in mature and germinating legume seeds and their behavior during cooking. Food/Nahrung 2000, 44, 23–27. [Google Scholar] [CrossRef]
- MacGregor, R.D.; Sarah, L. Kendall; Hannah Florance; Fabio Fedi; Karen Moore; Konrad Paszkiewicz; Smirnoff, N.; Penfield, S. Seed production temperature regulation of primary dormancy occurs through control of seed coat phenylpropanoid metabolism. New Phytol. 2014, 205, 642–652. [Google Scholar] [CrossRef]
- E., W.J. Seed coat and water absorption properties of seed of near-isogenic snap bean lines differing in seed coat color. Journal of the American Society for Horticultural Science 1977, 102, 478–480. [CrossRef]
- Corbineau, F.; M.A. Picard; Côme, D. Germinability of leek seeds and its improvement by osmopriming. Acta Horticulture 1993. [CrossRef]
- MM., E. Dormancy in seeds of Charlock: III. Occurrence and mode ofaction ofan inhibitor associated with dormancy. Journal of Experimental Botany 1968, 19, 601–610. [CrossRef]
- Tobe, K.; Zhang, L.; Yu Qiu, G.; Shimizu, H.; Omasa, K. Characteristics of seed germination in five non-halophytic Chinese desert shrub species. Journal of Arid Environments 2001, 47, 191–201. [Google Scholar] [CrossRef]
- Wingler, A.; Henriques, R. Sugars and the speed of life—Metabolic signals that determine plant growth, development and death. Physiologia Plantarum 2022, 174, e13656. [Google Scholar] [CrossRef]
- Anderson, G.H.; Veit, B.; MR, H. The Arabidopsis AtRaptor genes are essential for post-embryonic plant growth. BMC Biol 2005, 3. [Google Scholar] [CrossRef]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.; Orozco, J.; Zwieniecki, M.A. Non-structural carbohydrates in dormant woody perennials; the tale of winter survival and spring arrival. Frontiers in Forests and Global Change 2019, 2. [Google Scholar] [CrossRef]
- Tarancon, C.; González-Grando, E.; Oliveros, J.C.; Nicolas, M.; Cubas, P. A conserved carbon starvation response underlies bud dormancy in woody and herbaceous species. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Canam, T.; Kang, K.Y.; Unda, F.; Mansfield, S.D. Sucrose phosphate synthase expression influences poplar phenology. Tree Physiology 2009, 29, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, C.; Teulat-Merah, B.; MorÈRe-Le Paven, M.C.; Leprince, O.; Ly Vu, B.; Viau, L.; Ledroit, L.; Pelletier, S.; Payet, N.; Satour, P.; Lebras, C.; Gallardo, K.; Huguet, T.; Limami, A.M.; et al. Quantitative trait loci analysis reveals a correlation between the ratio of sucrose/raffinose family oligosaccharides and seed vigour in Medicago truncatula. Plant Cell Environ. 2011, 34, 1473–1487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fernie, A.R. On the role of the tricarboxylic cid cycle in plant productivity. Journal of Integrative Plant Biology 2018, 60, 1199–1216. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; LJ, S. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr Opin Plant Biol 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Fernie AR; E, M. Malate. Jack of all trades or master of a few? Phytochemistry 2009, 70, 828–883. [CrossRef]
- Kim, H.U. Lipid metabolism in plants. Plants 2020, 9, 1–4. [Google Scholar] [CrossRef]
- Mitio, H.; Sekozawa, Y.; Kobayashi, M.; Saito, K.; Kusano, M.; Sugaya, S. Metabolomics analysis of ' Housui ' Japanese pear fl ower buds during endodormancy reveals metabolic suppression by thermal fl uctuation. Plant Physiology and Biochemistry 2018, 126, 134–141. [Google Scholar]
- Qiu, X.M.; Sun, Y.Y.; Ye, X.Y.; Li, Z.G. Signaling Role of Glutamate in Plants. Frontiers in Plant Science 2020, 10, 1–11. [Google Scholar] [CrossRef]
- M, K.; Araújo, W.L.; AR, F.; Galili, G. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning 2012, 63, 695–709. [Google Scholar]
- Nieuwland, J., Menges, M., and Murray, J. A. H. . ''The plant cyclins,” in Cell Cycle Control and Plant Development, . In Plant J, Inze, D., Ed.; Oxford: Blackwell Publishing: Oxford, 2007; Volume 41. Oxford: Blackwell Publishing: Oxford.
- Zsigmond, L.; Tomasskovics, B.; Deák, V.; Rigó, G.; Szabados, L.; Bánhegyi, G.; Szarka, A. Plant Physiology and Biochemistry Enhanced activity of galactono-1, 4-lactone dehydrogenase and ascorbate e glutathione cycle in mitochondria from complex III de fi cient Arabidopsis. Plant Physiology et Biochemistry 2011, 49, 809–815. [Google Scholar] [CrossRef]
- Blokhina, O.; Fagerstedt, K.V. Reactive oxygen species and nitric oxide in plant mitochondria: Origin and redundant regulatory systems. Physiologia Plantarum 2010, 138, 447–462. [Google Scholar] [CrossRef]
- Paradiso, A.; Pinto, M.C.D.; Locato, V.; Gara, L.D. Galactone- c -lactone-dependent ascorbate biosynthesis alters wheat kernel maturation. Plant Biology 2012, 1435–8603. [Google Scholar] [CrossRef] [PubMed]
- Xool-tamayo, J.; Tamayo-ordoñez, Y.; Monforte-gonz, M.; Muñoz-s, A.; Felipe, V. Alkaloid Biosynthesis in the Early Stages of the Germination of Argemone mexicana L. (Papaveraceae). Plants https://doi.org/. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Smith, D.L. Flavonoids in Agriculture : Chemistry and Roles in, Biotic and Abiotic Stress Responses, and Microbial Associations. Agronomy 2020, 10. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds ? New Phytologist 2018, 220, 692–702. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as Important Molecules of Plant Interactions with the Environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Corso, M.; Perreau, F.; Mouille, G.; Lepiniec, L. Specialized phenolic compounds in seeds: Structures, functions, and regulations. Plant Science 2020, 296, 110–471. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Byrem, T.M.; Nair, M.G.; Strasburg, G.M. Modulation of liposomal membrane fluidity by flavonoids and isoflavonoids. Arch. Biochem. Biophys. 2000, 373. [Google Scholar] [CrossRef]
- Cotelle, N.; Bernier, J.L.; Catteau, J.P.; Pommery, J.; Wallet, J.C.; Gaydou, E.M. Antioxidant properties of hydroxy-flavones. Free Radic. Biol. Med. 1996, 20, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Prinsi, B. Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism. Plants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Chattha, F.A.; Munawar, M.A.; Nisa, M.; Kousar, S. Plant growth regulating activity of coumarins. Plant growth regulating activity of coumarins. In Coumarin-Based Heteroaromatics as Plant Growth Coumarin-Based Heteroaromatics as Plant Growth Regulators Regulators; University of the Punjab: Lahore, Pakistan, 2016; pp. 91–104. [Google Scholar]
- Chen, Z.; Ma, Y.; Yang, R.; Gu, Z.; Wang, P. Effects of exogenous Ca2+ on phenolic accumulation and physiological changes in germinated wheat (Triticum aestivum L.) under UV-B radiation. Food Chem. 2019, 288, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Pergo, E.M., D. ; Abrahim, P.C.; Soares, K.A.; Kern, L.J.D.; Silva, E.V.; Iwamoto., E.L.I. Bidens pilosa L. exhibits high sensitivity to coumarin in comparison with three other weed species. J. Chem. Ecol. 2008, 34, 499–507. [CrossRef] [PubMed]
- Berrie, A.M.M.; Parker, W.; Knights, B.A.; Hendrie, M.R. Studies on lettuce seed germination-I. Coumarin induced dormancy. Phytochemistry 1968, 7, 567–573. [Google Scholar] [CrossRef]
- Goren, R.; Tomer, E. Effect of seselin and coumarin on growth, indoleacetic acid oxidase and peroxidase with special reference to cucumber radicles. Plant Physiol. 1971, 47, 312–316. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of Terpenes and Recent Advances in Plant Protection. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Kumar, D.A.; Touhidur Rahman Anik; Md. Mezanur Rahman; Keya, S.S.; Islam, M.R.; Rahman, M.A.; Sultana, S.; Ghosh, P.K.; Khan, S.; Ahamed, T.; et al. Ethanol Treatment Enhances Physiological and Biochemical Responses to Mitigate Saline Toxicity in Soybean. Plants 2022, 11.
- Ventura, I.; Brunello, L.; Iacopino, S.; Valeri, M.C.; Novi, G.; Dornbusch, T.; Perata, P.; Loreti, E. Arabidopsis phenotyping reveals the importance of alcohol dehydrogenase and pyruvate decarboxylase for aerobic plant growth. Scientific Reports 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Su, W.; Ren, Y.; Wang, D.; Su, Y.; Feng, J.; Zhang, C.; Tang, H.; Xu, L.; Muhammad, K.; Que, Y. The alcohol dehydrogenase gene family in sugarcane and its involvement in cold stress regulation. BMC Genomics 2020, 21, 1–17. [Google Scholar] [CrossRef]
- Skalák, J.; Vercruyssen, L.; Claeys, H.; Hradilová, J.; Černý, M.; Novák, O.; Plačková, L.; Saiz-Fernández, I.; Skaláková, P.; Coppens, F.; et al. Multifaceted activity of cytokinin in leaf development shapes its size and structure in Arabidopsis. Plant Journal 2019, 97, 805–824. [Google Scholar] [CrossRef]
- Mao, J.; Li, W.; Mi, B.; Mujitaba, M.; Ma, Z.; Zhang, Y.; Chen, B.; Caldero, A. Different exogenous sugars affect the hormone signal pathway and sugar metabolism in ‘‘ Red Globe ’’ (Vitis vinifera L.) plantlets grown in vitro as shown by transcriptomic analysis. Planta 2017, 246, 537–552. [Google Scholar] [CrossRef]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. PNAS www.pnas.org/cgi/doi/10.1073/pnas.1215788110 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Savka, M.A.; Hudson, A.O. The synthesis and role of β-alanine in plants. Frontiers in plant science 2019, 10, 921. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.; Sodek, L.; Licausi, F.; Hameed, M.W.; Dornelas, M.C.; Van Dongen, J.T. nalysis of alanine aminotransferase in various organs of soybean (Glycine max) and in dependence of different nitrogen fertilisers during hypoxic stress. Amino Acids 2010, 39, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Liu, X.; Liu, F.; Ren, Y.; Wang, Y.; Zhu, J.; Teng, X.; Duan, E.; Wang, F.; Zhang, H. FLOURY ENDOSPERM12 encoding alanine aminotransferase 1 regulates carbon and nitrogen metabolism in rice. Journal of Plant Biology 2019, 62, 61–73. [Google Scholar] [CrossRef]
- Tzin, V. Malitsky S.; Galili G, A.A. Expression of a bacterial bi-functional chorismate mutase/prephenate dehydratase modulates primary and secondary metabolism associated with aromatic amino acids in Arabidopsis. Plant J. 2009, 60, 156–167. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Current opinion in biotechnology 2019, 56, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Schenck, C.A.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [CrossRef]
- Dennis, R. The role of primary carbohydrate metabolism in wheat grain dormancy and germination. 2019.
Figure 2.
(a,b); (a) Comparison of total annotated metabolites in the tubers of the two yam genotypes, (b) comparison of differentially accumulated metabolites in the tubers of the two genotypes.
Figure 2.
(a,b); (a) Comparison of total annotated metabolites in the tubers of the two yam genotypes, (b) comparison of differentially accumulated metabolites in the tubers of the two genotypes.

Figure 3.
(a–b) Differential accumulated metabolites identified by ANOVA plot with p value threshold 0.05, FDR ≤ 0.05 and logFC 1.5. means were compared using Turkey’s Post-hoc test. a shows DAMs in the tubers of TDr1100873 across the studied tuber dormancy stages. (b) shows DAMs in the tubers of Obiaoturugo across the studied tuber dormancy stages. Red dots are significant differential accumulated metabolites, while the green dots are the metabolites that were differentially accumulated, but are not significant. The vertical axis indicates the metabolites log-fold-change, while the horizontal axis shows the mass peak of the metabolites.
Figure 3.
(a–b) Differential accumulated metabolites identified by ANOVA plot with p value threshold 0.05, FDR ≤ 0.05 and logFC 1.5. means were compared using Turkey’s Post-hoc test. a shows DAMs in the tubers of TDr1100873 across the studied tuber dormancy stages. (b) shows DAMs in the tubers of Obiaoturugo across the studied tuber dormancy stages. Red dots are significant differential accumulated metabolites, while the green dots are the metabolites that were differentially accumulated, but are not significant. The vertical axis indicates the metabolites log-fold-change, while the horizontal axis shows the mass peak of the metabolites.
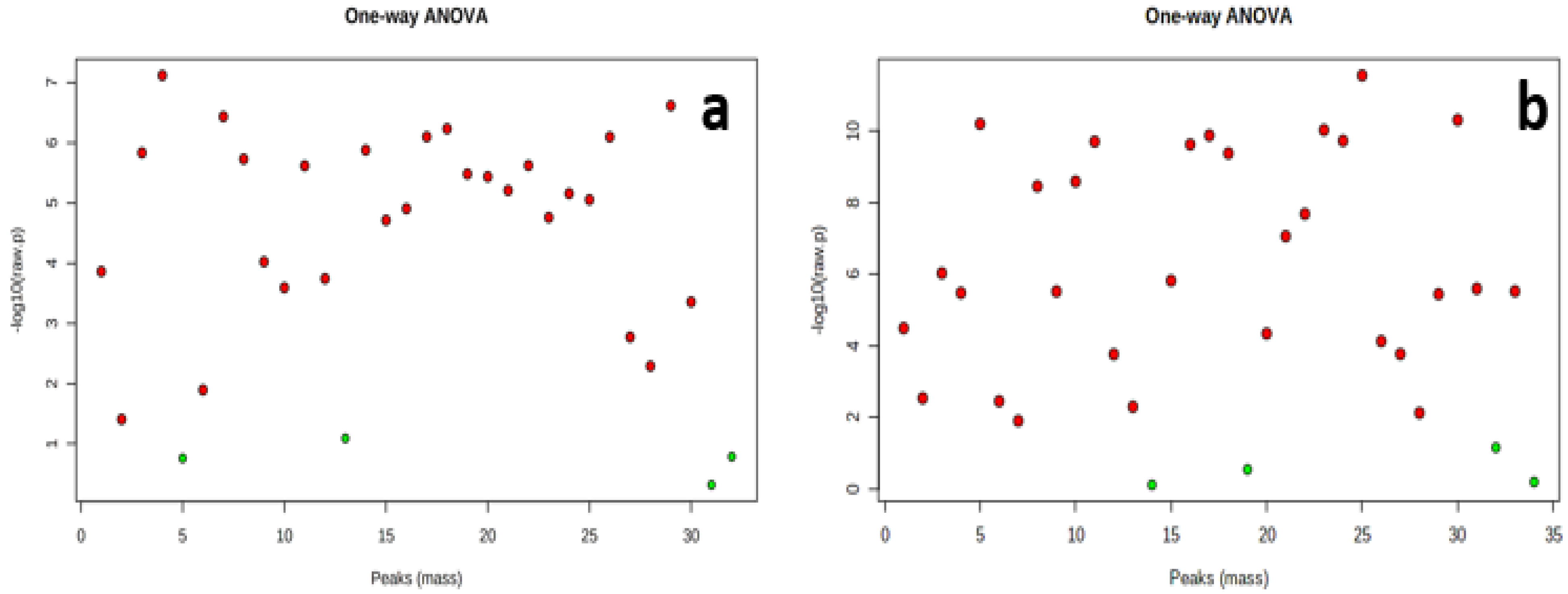
Figure 6.
Proportional composition of chemical functional groups of the DAMs. The color and area of each component of the pie chart shows the percentage composition of a particular chemical functional group.
Figure 6.
Proportional composition of chemical functional groups of the DAMs. The color and area of each component of the pie chart shows the percentage composition of a particular chemical functional group.
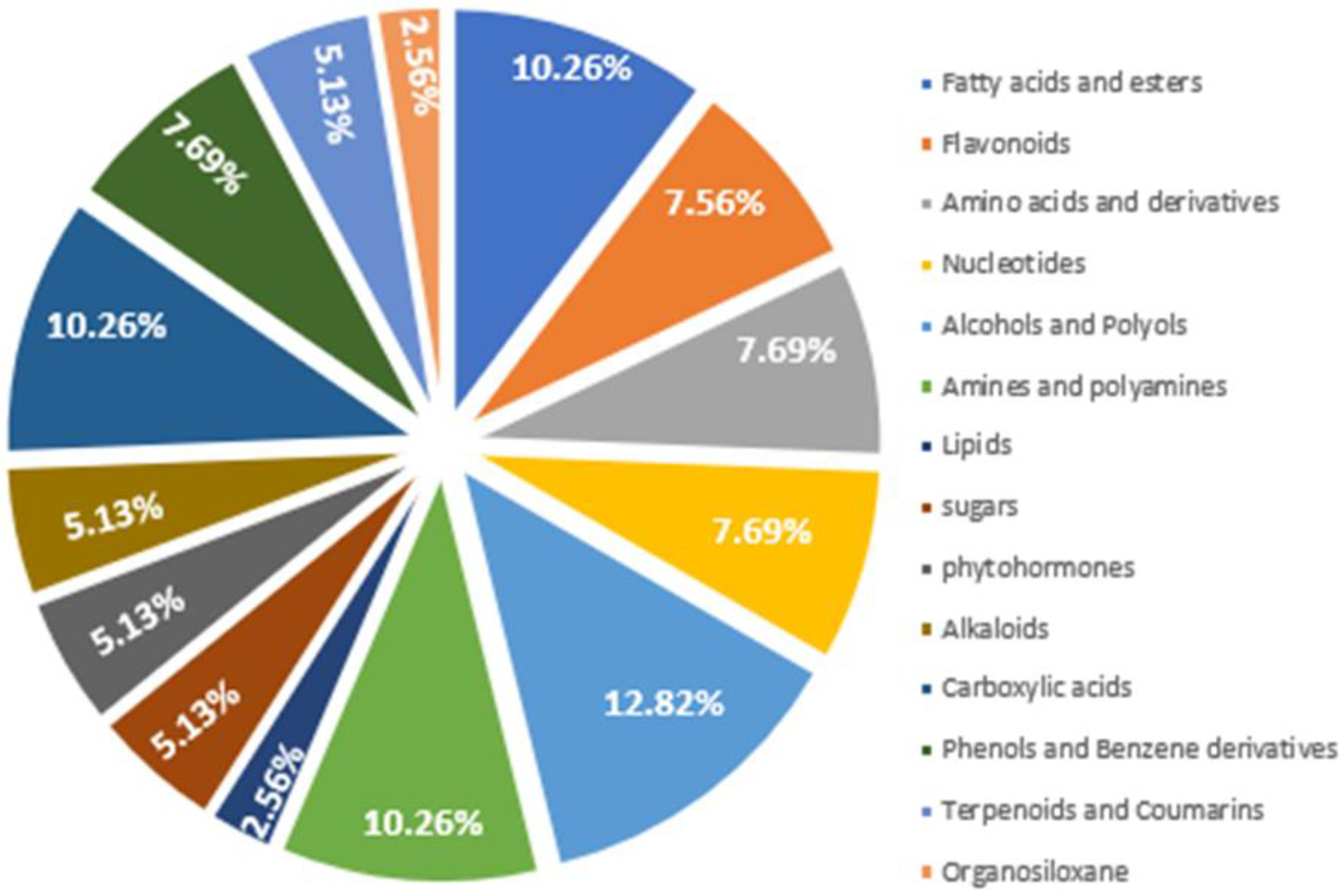
Figure 7.
Bar plots for metabolites set enrichment analysis showing top 23 metabolisms of enriched metabolites. The enrichment ratio values are the ratio between the fold change in the metabolisms across the studied dormancy stages and their expected values under normal condition. The color criterion indicates the p-value at which each metabolism was induced to its enrichment ratio, and it determines whether enrichment is significant or not.
Figure 7.
Bar plots for metabolites set enrichment analysis showing top 23 metabolisms of enriched metabolites. The enrichment ratio values are the ratio between the fold change in the metabolisms across the studied dormancy stages and their expected values under normal condition. The color criterion indicates the p-value at which each metabolism was induced to its enrichment ratio, and it determines whether enrichment is significant or not.
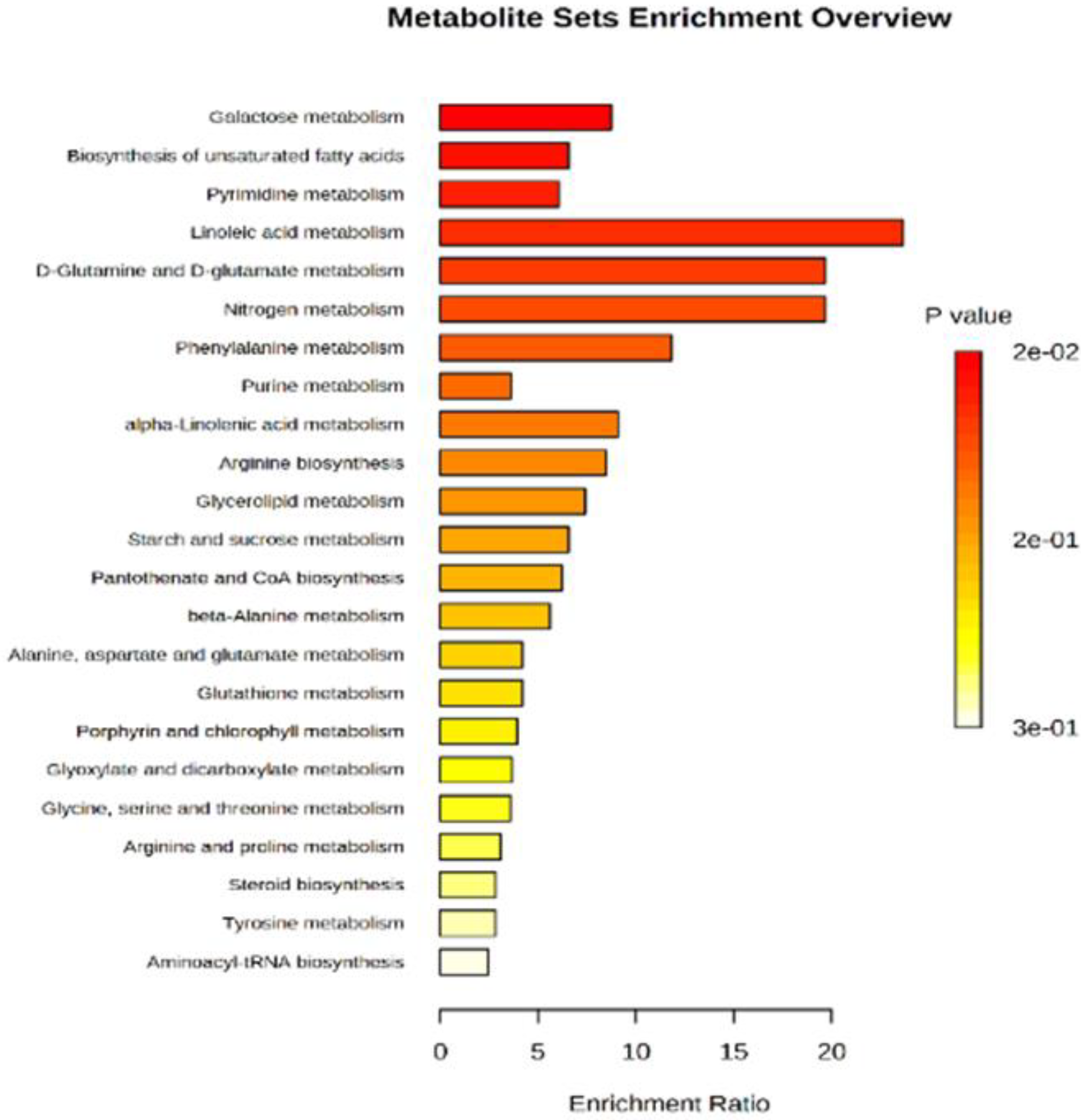
Figure 8.
Metabolome view of pathway topology showing the pathways that significantly impacted on yam tuber dormancy regulation. The dots denote metabolic pathways, size of the dots indicates the value of the impact of the pathway on yam tuber dormancy regulation on the scale of 0 to 1, whereas, the color intensity of the dots indicates the fold change of the pathway.
Figure 8.
Metabolome view of pathway topology showing the pathways that significantly impacted on yam tuber dormancy regulation. The dots denote metabolic pathways, size of the dots indicates the value of the impact of the pathway on yam tuber dormancy regulation on the scale of 0 to 1, whereas, the color intensity of the dots indicates the fold change of the pathway.
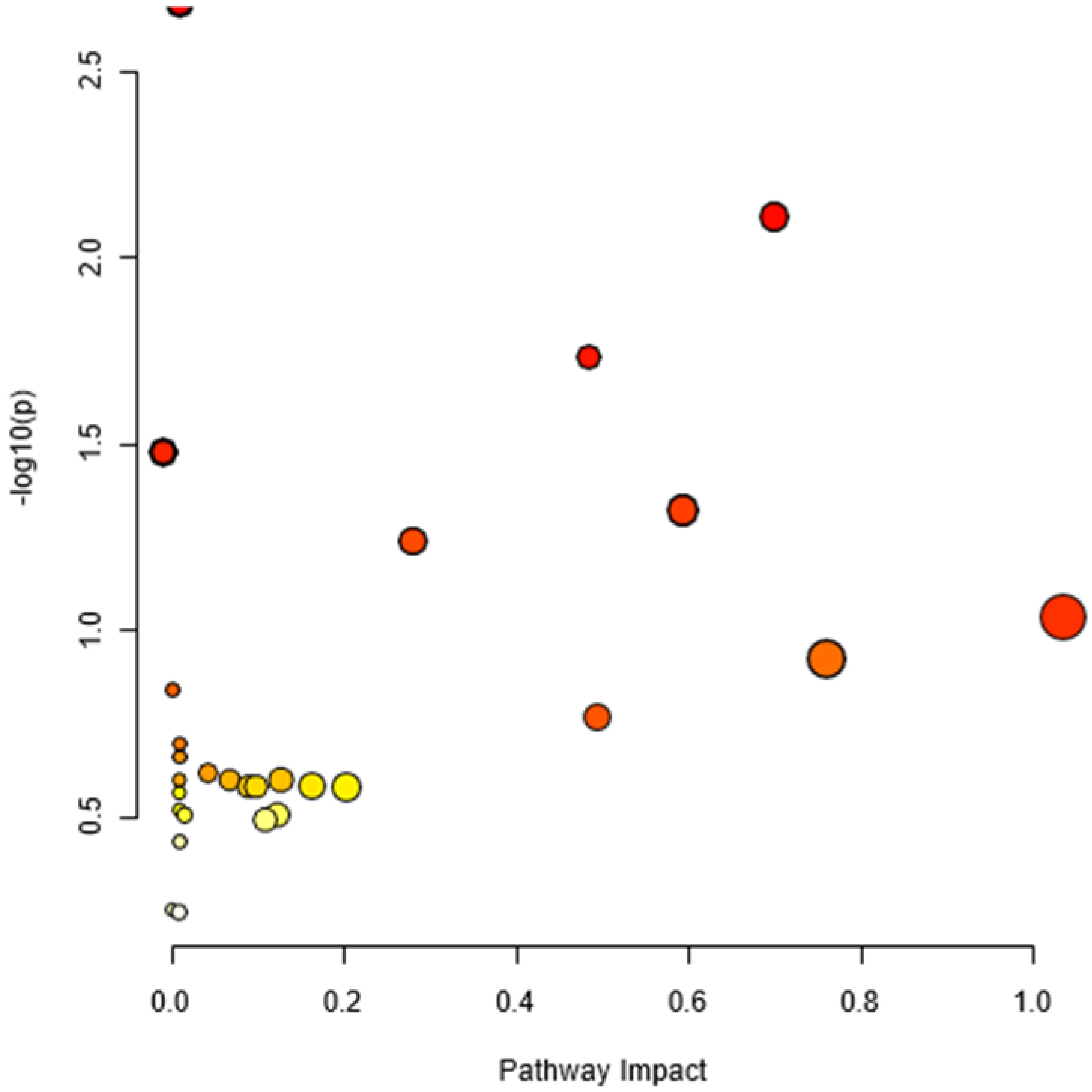
Figure 9.
(a–f) Six candidate pathways identified by pathway topology analysis that have significant impact on yam tuber dormancy regulation. (a) is linoleic acid metabolic pathway, it exerted maximum impact of 1, on a scale of 0 to 1. (b) is phenylalanine metabolic pathway, it exerted impact of 0.79. (c) is galactose metabolic pathway with impact value of 0.76. (d) is starch and sucrose metabolic pathway with impact value of 0.59. (e) is Alanine-aspartate-glutamine metabolic pathway with impact value of 0.57. (f) is purine metabolic pathway with impact value of 0.54.
Figure 9.
(a–f) Six candidate pathways identified by pathway topology analysis that have significant impact on yam tuber dormancy regulation. (a) is linoleic acid metabolic pathway, it exerted maximum impact of 1, on a scale of 0 to 1. (b) is phenylalanine metabolic pathway, it exerted impact of 0.79. (c) is galactose metabolic pathway with impact value of 0.76. (d) is starch and sucrose metabolic pathway with impact value of 0.59. (e) is Alanine-aspartate-glutamine metabolic pathway with impact value of 0.57. (f) is purine metabolic pathway with impact value of 0.54.
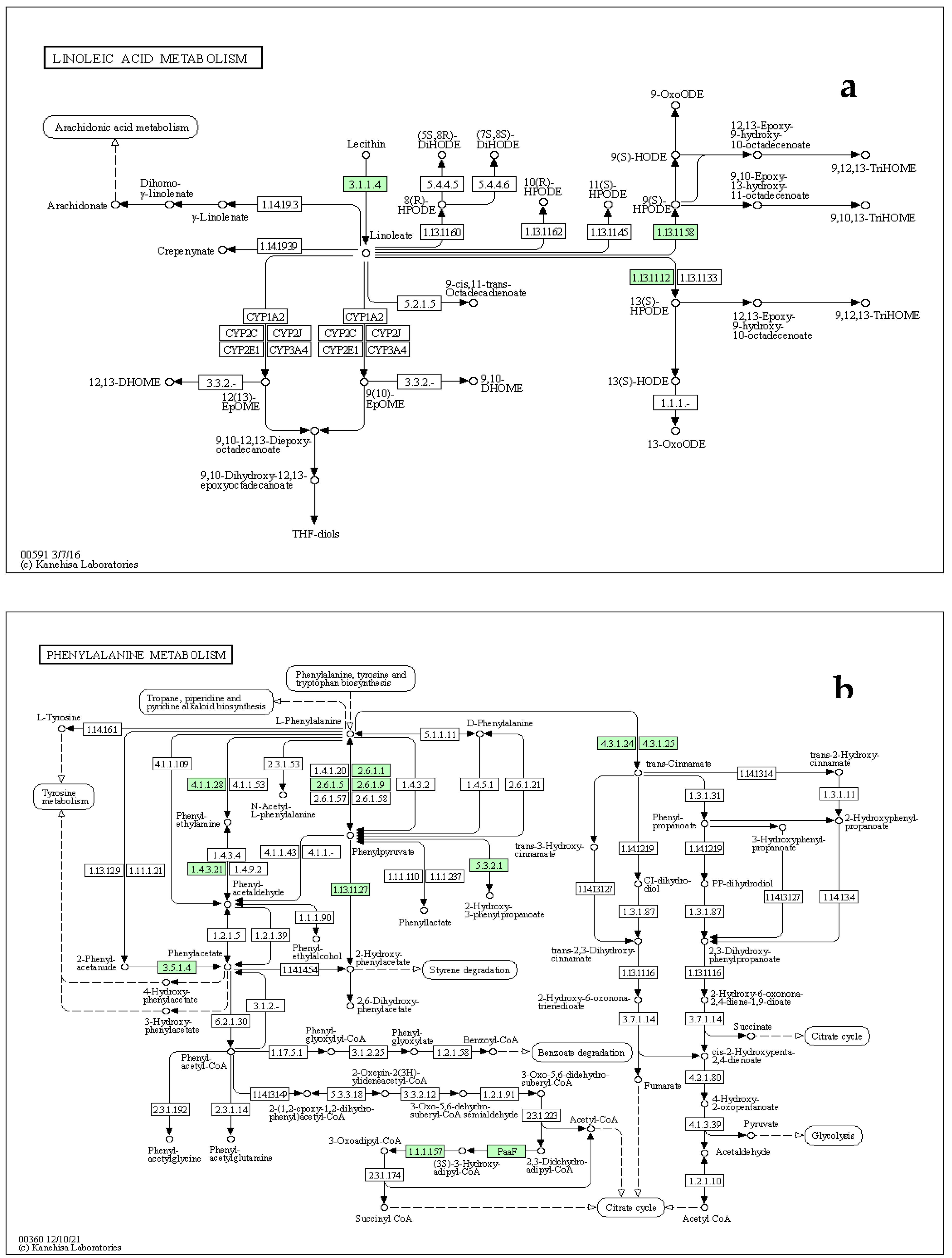
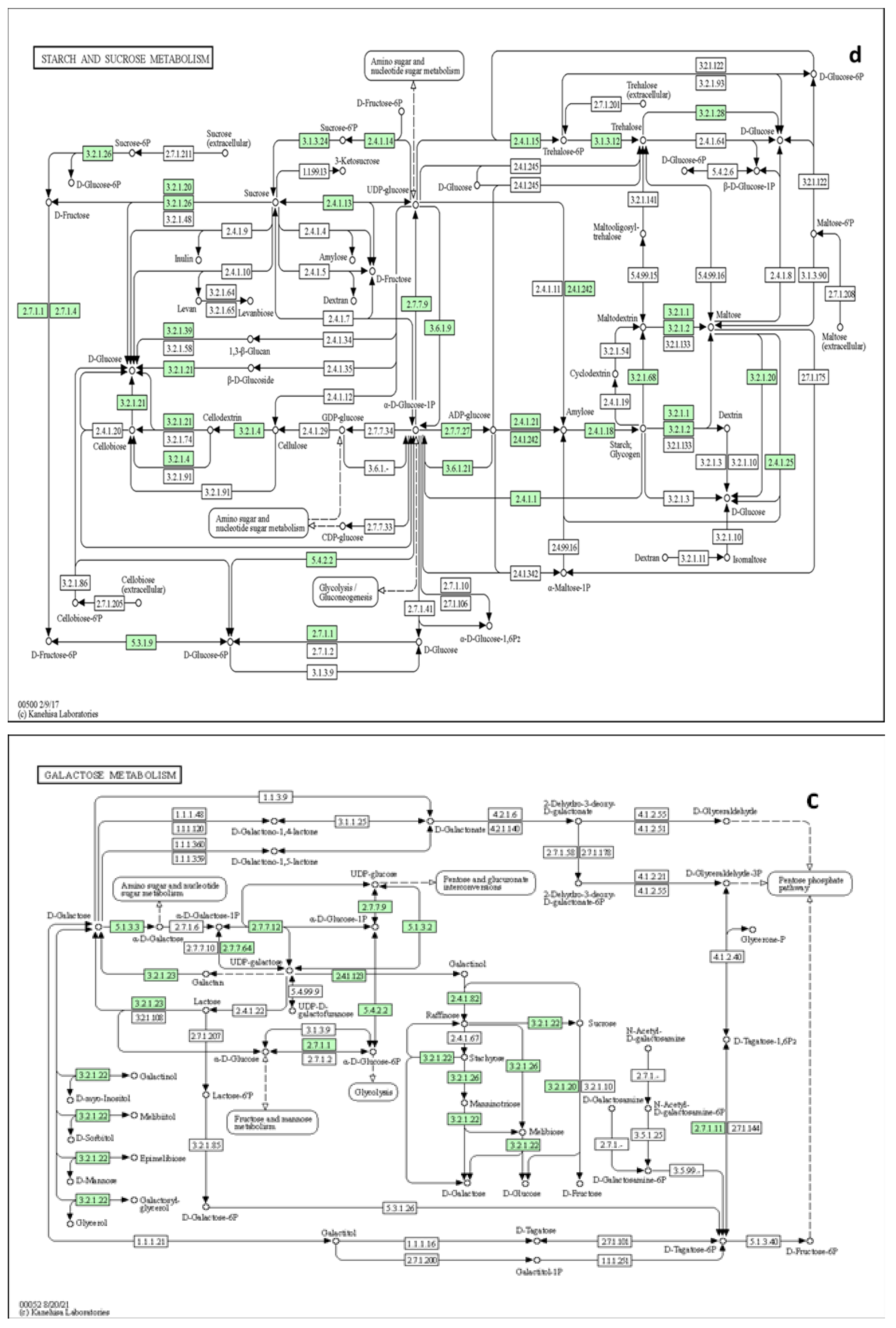
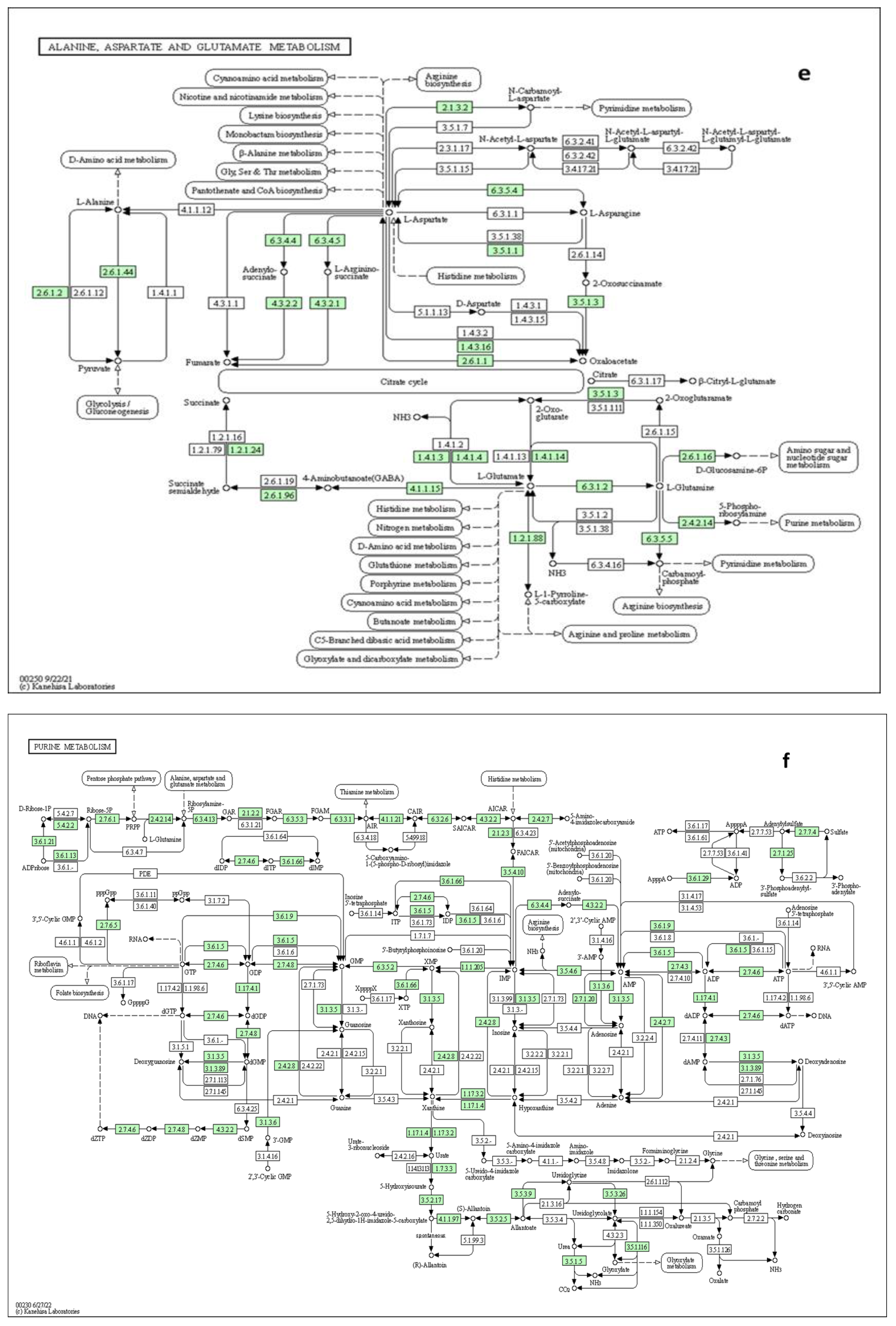

Table 1.
Result of mapping of DAMs in HMDB, PubChem and KEGG for standard name, chemical functional group and IDs determination.
Table 1.
Result of mapping of DAMs in HMDB, PubChem and KEGG for standard name, chemical functional group and IDs determination.
| Query | Match | Chem-Fun Group | HMDB | PubChem | KEGG |
|---|---|---|---|---|---|
| 9,12,15-Octadecatrienoic acid, (Z,Z,Z)- | Alpha-Linolenic acid | Fatty acids | HMDB0001388 | 5280934 | C06427 |
| Ethylenediamine-N,N'-dipropionic acid | Edetic Acid | Flavonoids | HMDB0015109 | 6049 | C00284 |
| 1-Butanol, 2-methyl-, acetate | Methyl methacrylate | Esters | HMDB0032385 | 6658 | C19504 |
| Tetraacetyl-d-xylonic nitrile | Sarcosine | Amino Acids and Derivatives | HMDB0000271 | 1088 | C00213 |
| (+)-N-Acetylmuramic acid | UDP-N-acetylmuraminate | nucleotide-sugar | HMDB0011720 | 24755495 | C01050 |
| 1-Butanol, 3-methyl-, formate | 3-Methylbutyl formate | Fatty esters | HMDB0034163 | 8052 | C12293 |
| 1,3-Propanediol, 2-ethyl-2-(hydroxymethyl)- | 1,3-Butanediol | Alcohols and Polyols | HMDB0031320 | 7896 | C20335 |
| 1,8-Di(4-nitrophenylmethyl)-3,6-diazahomoadamantan-9-one | Tyramine | Amino Acids and Derivatives | HMDB0000306 | 5610 | C00483 |
| Benzeneethanamine, 2-fluoro-á,3,4-trihydroxy-N-isopropyl- | Phenylethylamine | Amines, Aromatic | HMDB0012275 | 1001 | C05332 |
| Desulphosinigrin | Delphinidin 3-O-sophoroside | Flavonoids | 47205615 | C16307 | |
| Glycerin | Glycerol | Alcohols and Polyols | HMDB0000131 | 753 | C00116 |
| 9,12-Octadecadienoic acid (Z,Z)- | Linoleic acid | Lipids | HMDB0000673 | 5280450 | C01595 |
| Dasycarpidan-1-methanol, acetate (ester) | Acetaminophen | Flavonoids | HMDB0001859 | 1983 | C06804 |
| 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | 3-Phenylpropyl 2-methylpropanoate | Alcohols | HMDB0034472 | 7662 | C02008 |
| Sucrose | Sucrose | Disaccharides | HMDB0000258 | 5988 | C00089 |
| 5-O-Methyl-d-gluconic acid dimethylamide | 3-Cresotinic acid | Phytohormone | HMDB0002390 | 6738 | C14088 |
| 2,6-Dioxa-tricyclo[3.3.2.0(3,7)]decan-9-one | Ibuprofen | Amines | HMDB0001925 | 3672 | C01588 |
| 1,3-Cyclohexanedione, 2,5,5-trimethyl- | 1,4-Cyclohexanedione | Alcohols | 10263 | C08063 | |
| 5-Methoxypyrrolidin-2-one | Galactosylglycerol | Alkaloids | HMDB0006790 | 656504 | C05401 |
| 2(3H)-Furanone, 5-heptyldihydro | Galactonolactone | Sugar Acids | HMDB0002541 | 5640 | C03383 |
| 2-Propanol, 1,1'-oxybis- | Propranolol | Amino Alcohols | HMDB0001849 | 4946 | C07407 |
| 10-Hydroxydecanoic acid | 12-Hydroxydodecanoic acid | Fatty Acids | HMDB0002059 | 79034 | C08317 |
| Succinamide | Putrescine | Biogenic Polyamines | HMDB0001414 | 1045 | C00134 |
| 2-Octenoic acid, 4,5,7-trhydroxy | Methyl acrylate | Carboxylic Acids | HMDB0033977 | 7294 | C19443 |
| 2-Hydroxy-3-methoxy-succinic acid, dimethyl ester | L-Malic acid | carboxylic acid | HMDB0000156 | 222656 | C00149 |
| Octanoic acid, 2-phenylethyl ester | Styrene | Benzene Derivatives | HMDB0034240 | 7501 | C19506 |
| 2-Cyclohexen-1-one, 4-(3-hydroxy-1-butenyl)-3,5,5-trimethyl- | Amoxycillin | Terpenoids | HMDB0030500 | 2171 | C06827 |
| 8-Hydroxy-7-methoxycoumarin | Oxybenzone | Coumarins | HMDB0015497 | 4632 | C14285 |
| Pyrimidine-4,6-diol, 5-methyl- | Uracil | Nucleic Acids and Derivatives | HMDB0000300 | 1174 | C00106 |
| Oxime-, methoxy-phenyl-_ | 3,4-Dihydroxyphenylacetaldehyde | phenols | HMDB0003791 | 119219 | C04043 |
| Adenine | Adenine | Nucleotides | HMDB0000034 | 190 | C00147 |
| 2,5-Cyclohexadiene-1,4-dione, dioxime | Quinone | Benzoquinones | HMDB0003364 | 4650 | C00472 |
| 2,5-Pyrrolidinedione, 3-(1-aminoethylidene)-4-methyl- | Isopyridoxal | Alkaloids | HMDB0004290 | 440899 | C06051 |
| Phthalic acid, hept-4-yl isobutyl ester | Thalidomide | Carboxylic Acids | HMDB0015175 | 92142 | C07910 |
| L-Glutamine | L-Glutamine | Amino Acids and Derivatives | HMDB0000641 | 5961 | C00064 |
| Methyl cis-13,16-Docosadienate | Methylitaconate | carboxylic acid | METPA0268 | C02295 | |
| 2-Hydroxyhippuric acid-3TMS | Salicyluric acid | Phytohormone | HMDB0000840 | 10253 | C07588 |
| 2,4-Di-tert-butylphenoxytrimethylsilane | Butenafine | Amines | HMDB0015223 | 2484 | C08067 |
| Pentasiloxane, dodecamethyl- | Dodecamethylpentasiloxane | organosiloxane | HMDB0062731 | 8853 |
Table 2.
Result of pathway topology analysis, showing the significantly induced pathways and their impact on yam tuber dormancy regulation.
Table 2.
Result of pathway topology analysis, showing the significantly induced pathways and their impact on yam tuber dormancy regulation.
| Total | Expected | Hits | Raw p | -log10(p) | Holm adjust | FDR | Impact | |
|---|---|---|---|---|---|---|---|---|
| Isoquinoline alkaloid biosynthesis | 6 | 0.08 | 2 | 2.26E-03 | 2.65E+00 | 2.15E-01 | 2.03E-01 | 0.00 |
| Galactose metabolism | 27 | 0.35 | 3 | 4.28E-03 | 2.37E+00 | 4.02E-01 | 2.03E-01 | 0.76 |
| Tyrosine metabolism | 18 | 0.23 | 2 | 2.11E-02 | 1.68E+00 | 1.00E+00 | 6.67E-01 | 0.04 |
| Biosynthesis of unsaturated fatty acids | 22 | 0.28 | 2 | 3.09E-02 | 1.51E+00 | 1.00E+00 | 7.33E-01 | 0.00 |
| Linoleic acid metabolism | 4 | 0.05 | 1 | 5.04E-02 | 1.30E+00 | 1.00E+00 | 8.15E-01 | 1.00 |
| Glyoxylate and dicarboxylate | 29 | 0.37 | 2 | 5.14E-02 | 1.29E+00 | 1.00E+00 | 8.15E-01 | 0.06 |
| metabolism | ||||||||
| Pyrimidine metabolism | 38 | 0.49 | 2 | 8.32E-02 | 1.08E+00 | 1.00E+00 | 1.00E+00 | 0.06 |
| Lysine biosynthesis | 9 | 0.12 | 1 | 1.10E-01 | 9.59E-01 | 1.00E+00 | 1.00E+00 | 0.47 |
| Nitrogen metabolism | 12 | 0.15 | 1 | 1.44E-01 | 8.41E-01 | 1.00E+00 | 1.00E+00 | 0.54 |
| Phenylalanine metabolism | 12 | 0.15 | 1 | 1.44E-01 | 8.41E-01 | 1.00E+00 | 1.00E+00 | 0.79 |
| Purine metabolism | 63 | 0.81 | 2 | 1.92E-01 | 7.17E-01 | 1.00E+00 | 1.00E+00 | 0.26 |
| Arginine biosynthesis | 18 | 0.23 | 1 | 2.09E-01 | 6.81E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| beta-Alanine metabolism | 18 | 0.23 | 1 | 2.09E-01 | 6.81E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Citrate cycle (TCA cycle) | 20 | 0.26 | 1 | 2.29E-01 | 6.40E-01 | 1.00E+00 | 1.00E+00 | 0.33 |
| Zeatin biosynthesis | 21 | 0.27 | 1 | 2.39E-01 | 6.22E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Carbon fixation in photosynthetic organ- | 21 | 0.27 | 1 | 2.39E-01 | 6.22E-01 | 1.00E+00 | 1.00E+00 | 0.06 |
| isms | ||||||||
| Glycerolipid metabolism | 21 | 0.27 | 1 | 2.39E-01 | 6.22E-01 | 1.00E+00 | 1.00E+00 | 0.26 |
| Phenylalanine, tyrosine and tryptophan | 22 | 0.28 | 1 | 2.49E-01 | 6.04E-01 | 1.00E+00 | 1.00E+00 | 0.08 |
| biosynthesis | ||||||||
| Starch and sucrose metabolism | 22 | 0.28 | 1 | 2.49E-01 | 6.04E-01 | 1.00E+00 | 1.00E+00 | 0.59 |
| Pyruvate metabolism | 22 | 0.28 | 1 | 2.49E-01 | 6.04E-01 | 1.00E+00 | 1.00E+00 | 0.24 |
| Alanine, aspartate and glutamate | 22 | 0.28 | 1 | 2.49E-01 | 6.04E-01 | 1.00E+00 | 1.00E+00 | 0.57 |
| metabolism | ||||||||
| Pantothenate and CoA biosynthesis | 23 | 0.30 | 1 | 2.59E-01 | 5.87E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Cyanoamino acid metabolism | 26 | 0.33 | 1 | 2.87E-01 | 5.42E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Glutathione metabolism | 27 | 0.35 | 1 | 2.97E-01 | 5.28E-01 | 1.00E+00 | 1.00E+00 | 0.01 |
| alpha-Linolenic acid metabolism | 27 | 0.35 | 1 | 2.97E-01 | 5.28E-01 | 1.00E+00 | 1.00E+00 | 0.11 |
| Arginine and proline metabolism | 28 | 0.36 | 1 | 3.06E-01 | 5.14E-01 | 1.00E+00 | 1.00E+00 | 0.16 |
| Glycine, serine and threonine metabolism | 33 | 0.42 | 1 | 3.50E-01 | 4.56E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Aminoacyl-tRNA biosynthesis | 46 | 0.59 | 1 | 4.53E-01 | 3.44E-01 | 1.00E+00 | 1.00E+00 | 0.00 |
| Porphyrin and chlorophyll metabolism | 47 | 0.60 | 1 | 4.61E-01 | 3.37E-01 | 1.00E+00 | 1.00E+00 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



