
Submitted:
16 March 2023
Posted:
16 March 2023
You are already at the latest version
Abstract
CC chemokine receptor 6 (CCR6) is one of the members of G protein-coupled receptor (GPCR) family that is upregulated in many immune-related cells, including B lymphocytes, effector and memory T cells, regulatory T cells, and immature dendritic cells. Coordination between CCR6 and its ligand CC motif chemokine ligand 20 (CCL20) is deeply involved in the pathogenesis of various diseases, such as cancer, autoimmune diseases, and psoriasis. Therefore, CCR6 is an attractive target for therapy and is being investigated as a diagnostic marker for patients. In a previous study, we developed an anti-mouse CCR6 (mCCR6) monoclonal antibody (mAb), C6Mab-13 (rat IgG1, kappa), applicable for flow cytometry by immunizing a rat with N-terminal peptide of mCCR6. This study investigated the binding epitope of C6Mab-13 using enzyme-linked immunosorbent assay (ELISA) and surface plasmon resonance (SPR) methods with the synthesized point mutated-peptides within 1-20 amino acids region of mCCR6. In ELISA, C6Mab-13 lost the reaction to the alanine-substituted peptide of D11A. The epitope of C6Mab-13 was identified to be Asp11 in ELISA. Furthermore, in SPR analysis, the dissociation constants (KD) could not be calculated for G9A and D11A mutants due to lack of binding. The SPR analysis demonstrated that the C6Mab-13 epitope comprises Gly9 and Asp11. Taken together, the key binding epitope of C6Mab-13 was determined to be around Asp11 on mCCR6. Based on the epitope information, C6Mab-13 could be useful for further functional analysis of mCCR6 in future studies.
Keywords:
mouse CCR6
; monoclonal antibody
; epitope
; ELISA
; SPR
1. Introduction
The CC chemokine receptor 6 (CCR6) is a seven transmembrane chemokine receptor belonging to the member of G-protein-coupled receptors (GPCRs) family [1]. CCR6 is identified as a specific receptor for CC motif chemokine ligand 20 (CCL20) in 1997 [2]. CCR6 has been reported to be related to various diseases, such as cancer [3], autoimmune diseases [4], psoriasis [5], and inflammatory bowel disease (IBD) [6]. Since the expression of CCR6 is found in B cells or T cells, such as effector T cells, memory T cells, immature dendritic cells, Th17 cells, and regulatory T (Treg) cells, it affects the activity and directionality of immune cells [7,8,9]. In CCR6-deficient mice, the intestinal lymphoid tissue and Peyer's patch are not developed normally, and immune responses are also abnormal in contact hypersensitivity and delayed-type hypersensitivity models, indicating that CCR6 is found to be a key factor in the formation of the immune system [10].
The chemokine ligand CCL20, also known as macrophage inflammatory protein-3α (MIP-3α), liver and activation-regulated chemokine (LARC), or Exodus-1, is crucial ligand of CCR6 [1]. CCL20 also binds to scavenging receptor atypical chemokine receptor 4 (ACKR4), but G-protein canonical signaling has been occurred upon binding to CCR6 [11,12]. CCL20 is secreted by various immune-related cells, such as B cells, Th17 cells, dendritic cells, and natural killer cells [13]. Although there are various CC chemokine receptor-ligand pairs, the CCR6/CCL20-regulated immune response currently become a hotspot for immunological research, including disease development. The expression of CCR6 and CCL20 are dysregulated in the colonic mucosa and serum from IBD patients [14,15]. CCR6+ T cells are involved in exacerbation in an imiquimod-induced model of psoriasis [16]. Furthermore, tumor promoting effects of CCR6/CCL20 within the tumor microenvironment have been reported in many cancer types, such as renal cell carcinoma [17], gastric cancer [18], cervical cancer [19], and lung cancer [20,21]. Treg cells in peripheral blood (~60%) express CCR6 with stronger suppressive activity and higher FOXP3 expression in human oral squamous cell carcinoma patients [22]. These findings make CCL20/CCR6 axis an attractive therapeutic target for a wide variety of diseases. Development of inhibitors targeting CCR6/CCL20 axis has also been actively carried out [13].
We have previously developed various monoclonal antibodies (mAbs) against chemokine receptors, including mouse CCR2 [23], mouse CCR3 [24,25,26,27], mouse CCR4 [28], mouse CCR6 (mCCR6) [29], mouse CCR8 [30], mouse CCR9 [31], mouse CXCR6 [32], human CCR2 [33], and human CCR9 [34,35]. We have also investigated the epitope of those mAbs in previous studies [36,37,38,39,40,41]. The N-terminus of GPCRs, including CCR6, CCR9, and CXCR6, is identified as the ligand-binding domain [42,43,44,45]. Interestingly, the structure of binding between CCL20 and CCR6 has been elucidated. CCR6 and CCL20 have a shallow binding mode on the receptor surface, which induces allosteric conformational changes and is considered to trigger binding to intracellular G-proteins [46]. As well as analysis of the ligand-binding mode, the characterization of antibody epitopes is important for predicting neutralizing activity and assessment of efficacy against antigens, which regulate cellular conditions.
In this study, we performed the epitope identification of a rat anti-mCCR6 mAb (C6Mab-13; IgG1, kappa) using enzyme-linked immnosorbent assay (ELISA) and surface plasmon resonance (SPR) analysis against alanine-substituted N-terminal peptides of mCCR6.
2. Materials and Methods
2.1. Antibodies
A rat anti-mCCR6 mAb (clone C6Mab-13) was previously established by immunizing a rat with a keyhole limpet hemocyanin (KLH)-conjugated N-terminal peptide of mCCR6 (1-19 amino acids [aa] + C-terminal cysteine) [29]. We purchased secondary peroxidase-conjugated anti-rat immunoglobulins from Sigma-Aldrich Corp. (St. Louis, MO)
2.2. Peptides
The mCCR6 (Accession No.: NM_001190333.1) peptide (1-MNSTESYFGTDDYDNTEYYS-20) and 1×alanine residue-substituted peptides (Table 1) were synthesized by utilizing PEPscreen (Sigma-Aldrich Corp.).
2.3. ELISA
Synthesized mCCR6 peptides were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific Inc., Waltham, MA) at a concentration of 10 µg/mL for 30 min at 37℃. After washing with phosphate-buffered saline (PBS) containing 0.05% Tween20 (PBST; Nacalai Tesque, Inc., Kyoto, Japan), wells were blocked with 1% bovine serum albumin (BSA)-containing PBST for 30 min at 37℃. The plates were incubated with 10 µg/mL of C6Mab-13 for 30 min at 37℃, followed by a peroxidase-conjugated anti-rat immunoglobulins (1:20000 diluted; Sigma-Aldrich Corp.) for 30 min at 37℃. Enzymatic reactions were performed at room temperature using the ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc.). Optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA).
2.4. Measurements of KD via Surface Plasmon Resonance (SPR)
Measurements of KD between C6Mab-13 and the epitope region peptides were performed using SPR. C6Mab-13 was immobilized on the sensor chip CM5 according to the manufacture’s protocol by Cytiva (Marlborough, MA). In brief, C6Mab-13 was diluted to 10 μg/mL by the acetate buffer (pH 4.0: Cytiva) and immobilized using amine coupling reaction. The surface of the flow cell 2 of the sensor chip CM5 was treated with 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide and N-hydroxysuccinimide (NHS), followed by the injection of C6Mab-13. Unreacted NHS-ester was blocked with ethanolamine after C6Mab-13 immobilization. The KD between C6Mab-13 and alanine-substituted peptides described in Table 2 were measured using Biacore X100 (Cytiva) at 25˚C. The buffer was filtrated-PBS containing 0.05% (v/v) of Tween 20 and 0.24% (v/v) of dimethyl sulfoxide (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). A single cycle kinetics method was used to measure the binding signals. The data were analyzed by 1:1 binding kinetics to determine the association rate constant (ka), dissociation rate constant (kd), and dissociation constants (KD) using BIAevaluation software (Cytiva).
3. Results
3.1. Epitope Identification of C6Mab-13 by ELISA using 1×Alanine-Substituted mCCR6 Peptides
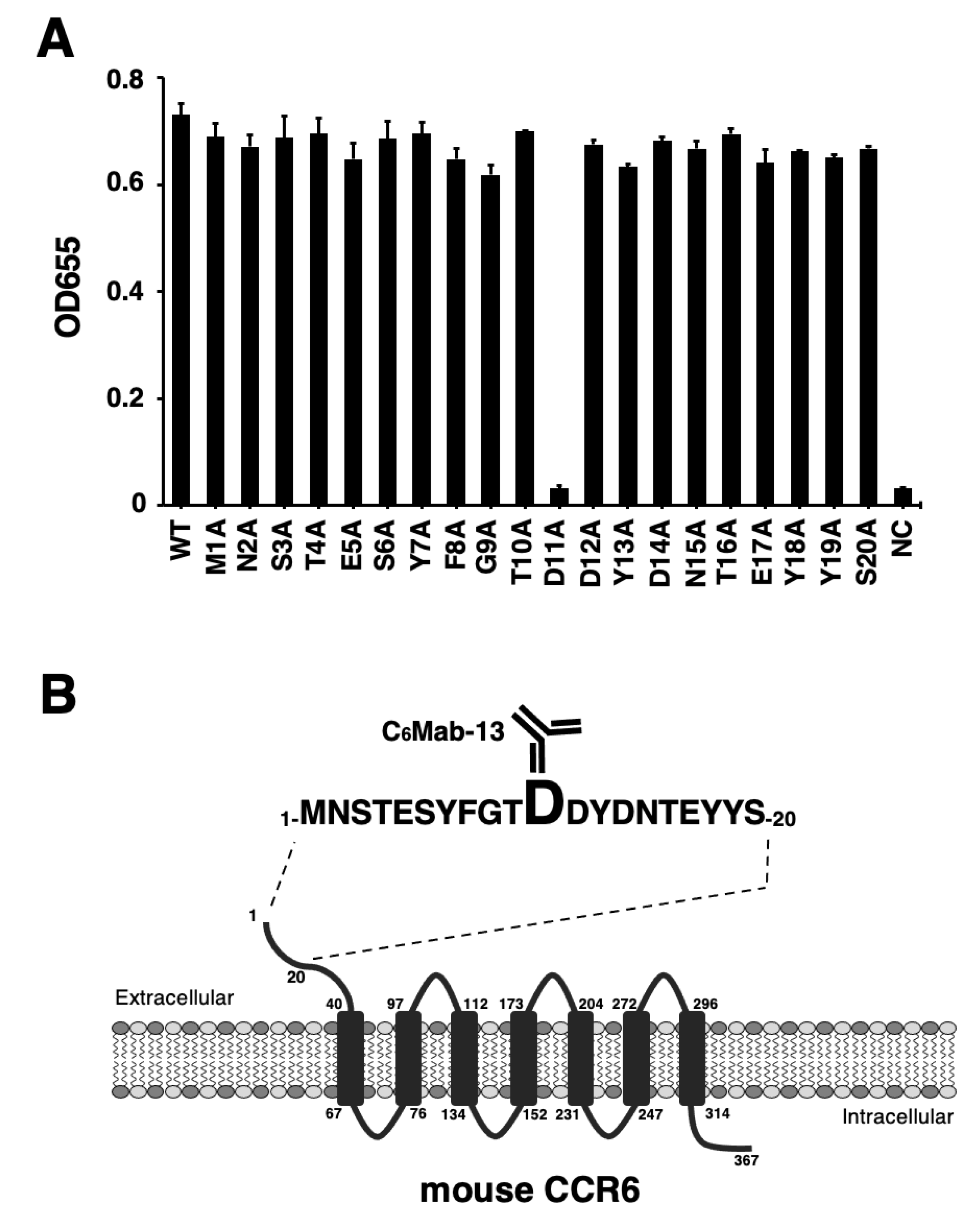
We developed an anti-mCCR6 mAb, C6Mab-13 (rat IgG1, kappa) by immunizing a rat with the KLH-conjugated mCCR6 N-terminal peptide [29]. To characterize the binding epitope of C6Mab-13, we synthesized 20 different 1×alanine-substituted mCCR6 peptides between Met1 to Ser20. The sequences are listed in Table 1. The results of ELISA using alanine-substituted peptides and C6Mab-13 demonstrated that C6Mab-13 bound to point mutants, such as M1A, N2A, S3A, T4A, E5A, S6A, Y7A, F8A, G9A, T10A, D12A, Y13A, D14A, N15A, T16A, E17A, Y18A, Y19A, and S20A as well as the 1-20 aa wild type (WT) sequence (positive control) (Fig. 1A). In contrast, C6Mab-13 did not react with D11A peptide (Fig. 1A). These results indicated that Asp11 was determined to be the critical aa, which is included in the C6Mab-13 epitope. The results are summarized schematically in Figure 1B.

Table 1.
Identification of C6Mab-13 epitope using point mutant peptides of mCCR6 by enzyme-linked immunosorbent assay.
Table 1.
Identification of C6Mab-13 epitope using point mutant peptides of mCCR6 by enzyme-linked immunosorbent assay.
| Peptides | Sequences | C6Mab-13 reactivity |
| p1_20 (WT) | MNSTESYFGTDDYDNTEYYS | +++ |
| M1A | ANSTESYFGTDDYDNTEYYS | +++ |
| N2A | MASTESYFGTDDYDNTEYYS | +++ |
| S3A | MNATESYFGTDDYDNTEYYS | +++ |
| T4A | MNSAESYFGTDDYDNTEYYS | +++ |
| E5A | MNSTASYFGTDDYDNTEYYS | +++ |
| S6A | MNSTEAYFGTDDYDNTEYYS | +++ |
| Y7A | MNSTESAFGTDDYDNTEYYS | +++ |
| F8A | MNSTESYAGTDDYDNTEYYS | +++ |
| G9A | MNSTESYFATDDYDNTEYYS | +++ |
| T10A | MNSTESYFGADDYDNTEYYS | +++ |
| D11A | MNSTESYFGTADYDNTEYYS | - |
| D12A | MNSTESYFGTDAYDNTEYYS | +++ |
| Y13A | MNSTESYFGTDDADNTEYYS | +++ |
| D14A | MNSTESYFGTDDYANTEYYS | +++ |
| N15A | MNSTESYFGTDDYDATEYYS | +++ |
| T16A | MNSTESYFGTDDYDNAEYYS | +++ |
| E17A | MNSTESYFGTDDYDNTAYYS | +++ |
| Y18A | MNSTESYFGTDDYDNTEAYS | +++ |
| Y19A | MNSTESYFGTDDYDNTEYAS | +++ |
| S20A | MNSTESYFGTDDYDNTEYYA | +++ |
+++, OD655≧0.3; -, OD655<0.1
3.2. Epitope Identification of C6Mab-13 by SPR using 1×Alanine-Substituted mCCR6 Peptides
To confirm the epitope of C6Mab-13, we next measured the binding affinity between C6Mab-13 and synthesized peptides, including 20 point mutants and WT of mCCR6, using Biacore X100. The sequences of peptides are described in Table 1. The measured values are summarized in Table 2. The association rate constant (ka), dissociation rate constant (kd), and dissociation constants (KD) of G9A and D11A were not determined. These results demonstrated that Gly9 and Asp11 were determined to be the critical aa of the C6Mab-13 epitope.
Mutant peptides of F8A, T10A, Y13A, and D14A increased the KD values by 15.5-, 4.4-, 16.5-, and 2.8-fold, respectively (Table 2), indicating that Phe8, Thr10, Tyr13, and Asp14 might contribute to C6Mab-13-binding to mCCR6.
Table 2.
The KD between C6Mab-13 and 1×Alanine-substituted peptides by surface plasmon resonance.
| Peptides | ka (/ms) | kd (/s) | KD (M) |
| p1_20 (WT) | 6.84 × 103 | 3.77 × 10-3 | 5.52 × 10-7 |
| M1A | 6.94 × 103 | 4.15 × 10-3 | 5.99 × 10-7 |
| N2A | 7.86 × 103 | 4.23 × 10-3 | 5.38 × 10-7 |
| S3A | 7.62 × 103 | 4.53 × 10-3 | 5.94 × 10-7 |
| T4A | 7.92 × 103 | 4.55 × 10-3 | 5.75 × 10-7 |
| E5A | 8.20 × 103 | 4.64 × 10-3 | 5.65 × 10-7 |
| S6A | 9.05 × 103 | 5.25 × 10-3 | 5.81 × 10-7 |
| Y7A | 8.16 × 103 | 3.45 × 10-3 | 4.23 × 10-7 |
| F8A | 1.43 × 103 | 1.23 × 10-2 | 8.55 × 10-6 |
| G9A | ND | ND | ND |
| T10A | 1.31 × 104 | 3.15 × 10-2 | 2.40 × 10-6 |
| D11A | ND | ND | ND |
| D12A | 7.43 × 103 | 7.09 × 10-3 | 9.55 × 10-7 |
| Y13A | 1.43 × 103 | 1.30 × 10-2 | 9.12 × 10-6 |
| D14A | 6.87 × 103 | 1.05 × 10-2 | 1.53 × 10-6 |
| N15A | 6.19 × 103 | 5.61 × 10-3 | 9.06 × 10-7 |
| T16A | 6.23 × 103 | 5.17 × 10-3 | 8.30 × 10-7 |
| E17A | 6.38 × 103 | 6.67 × 10-3 | 1.05 × 10-6 |
| Y18A | 5.23 × 103 | 5.56 × 10-3 | 1.06 × 10-6 |
| Y19A | 5.75 × 103 | 6.02 × 10-3 | 1.05 × 10-6 |
| S20A | 4.68 × 103 | 5.96 × 10-3 | 1.27 × 10-6 |
ND, not determined.
4. Discussion
This study examines the binding epitope of C6Mab-13 by 1×alanine-substituted peptide scanning method using ELISA and SPR. We concluded the pivotal epitope as Asp11 using ELISA, and as Gly9 and Asp11 using SPR. They are located outside the region of all three extracellular domains of CCR6 and N-terminal residues from Tyr27 to Leu38, to which the chemokine ligand CCL20 binds [46]. Therefore, C6Mab-13 might not show the neutralizing activity for CCL20. In contrast, structural changes might occur when C6Mab-13 binds to CCR6. Also, it has been reported that different affinities of antibodies dramatically alter the functional activities [47]. Therefore, we plan to investigate the neutralizing activity of C6Mab-13 between CCL20 and CCR6 in the future study.
The epitope mapping results using ELISA (Figure 1) and SPR (Table 2) indicated the similar region of mCCR6 as the binding epitope. However, Gly9 is shown as the critical aa by only SPR analysis (Table 2). The experimental system is different between two experiments such as; i) the synthesized peptides were immobilized on immunoplates in ELISA, whereas C6Mab-13 was immobilized on a sensor chip CM5 in SPR analysis; ii) the reaction time between the antigen and the antibody is different; iii) the secondary antibody is used only in ELISA. These different conditions may lead to the inconsistent results of two experiments in this study.
In SPR analysis, mutant peptides of F8A, T10A, Y13A, and D14A increased the KD values by 15.5-, 4.4-, 16.5-, and 2.8-fold, respectively (Table 2). These results indicate that Phe8, Thr10, Tyr13, and Asp14 might contribute to C6Mab-13-binding to mCCR6. In the future, we will perform the cell-based alanine-scanning or 2×alanine-scanning methods for more detailed epitope analysis for C6Mab-13 as we previously clarified the epitopes of mAbs [36,38,40]. We further plan to apply a RIEDL insertion for epitope mapping (REMAP) method for the epitope mapping of C6Mab-13 [48,49]. The REMAP method is useful for determining the conformational epitope, which could not be identified by alanine scanning. The REMAP method will be effective for more detailed epitope analysis of C6Mab-13.
When CCL20 is secreted in tumor tissues, it attracts CCR6-expressing Treg cells [50], which are involved in tumor progression and poor prognosis in patients with hepatocellular carcinoma [51]. Using this biological response of CCR6, novel cancer treatment strategies using CCR6-expressing chimeric antigen receptor-T (CAR-T) cells were designed [52,53]. Also, removal of immunosuppressive cells, such as CCR6+ Treg cells, may enhance antitumor efficacy [54]. We demonstrated that C6Mab-13 possesses high binding affinity against mCCR6, which is expressed in Chinese hamster ovary-K1 cells (KD : 2.8×10-9 M by flow cytometric analysis) in the previous study [29]. Therefore, C6Mab-13 is expected to be useful for antitumor evaluation by depletion of CCR6-expressing Treg cells in mouse models.
Author Contributions
T.T. and M.T. performed the experiments. M.K.K. and Y.K. designed the experiments. T.T. and H.S. analyzed the data. T.T., H.S. and Y.K. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP22ama121008 (to Y.K.), JP22am0401013 (to Y.K.), JP22bm1004001 (to Y.K.), JP22ck0106730 (to Y.K.), and JP21am0101078 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 21K20789 (to T.T.), 22K06995 (to H.S.), 21K07168 (to M.K.K.), and 22K07224 (to Y.K.).
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest involving this article.
References
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev 2003, 14, 409–426. [Google Scholar] [CrossRef]
- Baba, M.; Imai, T.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Hieshima, K.; Nomiyama, H.; Yoshie, O. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J Biol Chem 1997, 272, 14893–14898. [Google Scholar] [CrossRef]
- Kadomoto, S.; Izumi, K.; Mizokami, A. The CCL20-CCR6 Axis in Cancer Progression. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun Rev 2021, 20, 102846. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, S.; Xing, M.; Hong, S.; Liu, L.; Ding, X.J.; Sun, X.Y.; Luo, Y.; Wang, C.X.; Zhang, M.; et al. Current evidence on the role of lipid lowering drugs in the treatment of psoriasis. Front Med (Lausanne) 2022, 9, 900916. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.J.; Adams, D.H. Chemokines and Chemokine Receptors as Therapeutic Targets in Inflammatory Bowel Disease; Pitfalls and Promise. J Crohns Colitis 2018, 12, S641–s652. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Takata, H.; Takiguchi, M. Functional expression of chemokine receptor CCR6 on human effector memory CD8+ T cells. Eur J Immunol 2007, 37, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, F.; Cosmi, L.; Santarlasci, V.; Maggi, L.; Liotta, F.; Mazzinghi, B.; Parente, E.; Filì, L.; Ferri, S.; Frosali, F.; et al. Phenotypic and functional features of human Th17 cells. J Exp Med 2007, 204, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.S.; Körner, H. The CCR6-CCL20 axis in humoral immunity and T-B cell immunobiology. Immunobiology 2019, 224, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Varona, R.; Villares, R.; Carramolino, L.; Goya, I.; Zaballos, A.; Gutiérrez, J.; Torres, M.; Martínez, A.C.; Márquez, G. CCR6-deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed-type hypersensitivity responses. J Clin Invest 2001, 107, R37–45. [Google Scholar] [CrossRef]
- Matti, C.; D'Uonnolo, G.; Artinger, M.; Melgrati, S.; Salnikov, A.; Thelen, S.; Purvanov, V.; Strobel, T.D.; Spannagel, L.; Thelen, M.; et al. CCL20 is a novel ligand for the scavenging atypical chemokine receptor 4. J Leukoc Biol 2020, 107, 1137–1154. [Google Scholar] [CrossRef]
- Meyrath, M.; Reynders, N.; Uchański, T.; Chevigné, A.; Szpakowska, M. Systematic reassessment of chemokine-receptor pairings confirms CCL20 but not CXCL13 and extends the spectrum of ACKR4 agonists to CCL22. J Leukoc Biol 2021, 109, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, R.; Eri, R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina (Kaunas) 2018, 54. [Google Scholar] [CrossRef] [PubMed]
- Skovdahl, H.K.; Granlund, A.; Østvik, A.E.; Bruland, T.; Bakke, I.; Torp, S.H.; Damås, J.K.; Sandvik, A.K. Expression of CCL20 and Its Corresponding Receptor CCR6 Is Enhanced in Active Inflammatory Bowel Disease, and TLR3 Mediates CCL20 Expression in Colonic Epithelial Cells. PLoS One 2015, 10, e0141710. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Ludwiczek, O.; Holzmann, S.; Moschen, A.R.; Weiss, G.; Enrich, B.; Graziadei, I.; Dunzendorfer, S.; Wiedermann, C.J.; Mürzl, E.; et al. Increased expression of CCL20 in human inflammatory bowel disease. J Clin Immunol 2004, 24, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Ichimura, Y.; Kubota, N.; Konishi, R.; Nakamura, Y.; Mizuno, S.; Takahashi, S.; Fujimoto, M.; Nomura, T.; Okiyama, N. The Role of PD-L1 on Langerhans Cells in the Regulation of Psoriasis. J Invest Dermatol 2022, 142, 3167–3174e3169. [Google Scholar] [CrossRef]
- Kadomoto, S.; Izumi, K.; Hiratsuka, K.; Nakano, T.; Naito, R.; Makino, T.; Iwamoto, H.; Yaegashi, H.; Shigehara, K.; Kadono, Y.; et al. Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis. Cancers (Basel) 2019, 12. [Google Scholar] [CrossRef]
- Han, G.; Wu, D.; Yang, Y.; Li, Z.; Zhang, J.; Li, C. CrkL meditates CCL20/CCR6-induced EMT in gastric cancer. Cytokine 2015, 76, 163–169. [Google Scholar] [CrossRef]
- Yu, Q.; Lou, X.M.; He, Y. Preferential recruitment of Th17 cells to cervical cancer via CCR6-CCL20 pathway. PLoS One 2015, 10, e0120855. [Google Scholar] [CrossRef]
- Zhang, X.P.; Hu, Z.J.; Meng, A.H.; Duan, G.C.; Zhao, Q.T.; Yang, J. Role of CCL20/CCR6 and the ERK signaling pathway in lung adenocarcinoma. Oncol Lett 2017, 14, 8183–8189. [Google Scholar] [CrossRef]
- Wei, W.; Zhao, X.; Zhu, J.; Zhang, L.; Chen, Y.; Zhang, B.; Li, Y.; Wang, M.; Zhang, Z.; Wang, C. lncRNA-u50535 promotes the progression of lung cancer by activating CCL20/ERK signaling. Oncol Rep 2019, 42, 1946–1956. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kao, K.C.; Chiu, Y.L.; Jung, C.J.; Liu, C.J.; Cheng, S.J.; Chang, Y.L.; Ko, J.Y.; Chia, J.S. Enrichment of Human CCR6(+) Regulatory T Cells with Superior Suppressive Activity in Oral Cancer. J Immunol 2017, 199, 467–476. [Google Scholar] [CrossRef]
- Tanaka, T.; Li, G.; Asano, T.; Saito, M.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a Novel Anti-Mouse CCR2 Monoclonal Antibody (C(2)Mab-6) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 80–86. [Google Scholar] [CrossRef]
- Asano, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 3 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 107–112. [Google Scholar] [CrossRef]
- Saito, M.; Harigae, Y.; Li, G.; Asano, T.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. C(3)Mab-2: An Anti-Mouse CCR3 Monoclonal Antibody for Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 45–49. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Tanaka, T.; Saito, M.; Li, G.; Goto, N.; Nanamiya, R.; Kaneko, M.K.; Kato, Y. C(3)Mab-3: A Monoclonal Antibody for Mouse CC Chemokine Receptor 3 for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2022, 41, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Suzuki, H.; Goto, N.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Establishment of Novel Anti-Mouse CCR3 Monoclonal Antibodies (C(3)Mab-6 and C(3)Mab-7) by N-terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Suzuki, H.; Asano, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-Mouse CCR4 Monoclonal Antibody (C(4)Mab-1) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 87–93. [Google Scholar] [CrossRef]
- Asano, T.; Tanaka, T.; Suzuki, H.; Li, G.; Nanamiya, R.; Tateyama, N.; Isoda, Y.; Okada, Y.; Kobayashi, H.; Yoshikawa, T.; et al. Development of a Novel Anti-Mouse CCR6 Monoclonal Antibody (C(6)Mab-13) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 343–349. [Google Scholar] [CrossRef]
- Tanaka, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Sano, M.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 65–70. [Google Scholar] [CrossRef]
- Kobayashi, H.; Asano, T.; Suzuki, H.; Tanaka, T.; Yoshikawa, T.; Kaneko, M.K.; Kato, Y. Establishment of a Sensitive Monoclonal Antibody Against Mouse CCR9 (C(9)Mab-24) for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2022. [CrossRef]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx(6)Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Li, G.; Saito, M.; Suzuki, H.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-human CCR2 Monoclonal Antibody (C(2)Mab-9) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 188–193. [Google Scholar] [CrossRef]
- Nanamiya, R.; Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Hosono, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Suzuki, H.; Isoda, Y.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Takahashi, N.; Okuno, S.; Yoshikawa, T.; et al. Development of a Sensitive Anti-Human CCR9 Monoclonal Antibody (C(9)Mab-11) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 303–310. [Google Scholar] [CrossRef]
- Kobayashi, H.; Asano, T.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C(9)Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method. Antibodies (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Tateyama, N.; Asano, T.; Suzuki, H.; Li, G.; Yoshikawa, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry. Antibodies (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Isoda, Y.; Tanaka, T.; Suzuki, H.; Asano, T.; Yoshikawa, T.; Kitamura, K.; Kudo, Y.; Ejima, R.; Ozawa, K.; Kaneko, M.K.; et al. Epitope Mapping Using the Cell-Based 2 × Alanine Substitution Method About the Anti-mouse CXCR6 Monoclonal Antibody, Cx(6)Mab-1. Monoclon Antib Immunodiagn Immunother 2023, 42, 22–26. [Google Scholar] [CrossRef]
- Tanaka, T.; Suzuki, H.; Asano, T.; Li, G.; Nanamiya, R.; Tateyama, N.; Isoda, Y.; Okada, Y.; Kobayashi, H.; Yoshikawa, T.; et al. Epitope Mapping of an Anti-Mouse CCR2 Monoclonal Antibody (C(2)Mab-6) Using Enzyme-Linked Immunosorbent Assay. Monoclon Antib Immunodiagn Immunother 2022, 41, 339–342. [Google Scholar] [CrossRef]
- Isoda, Y.; Tanaka, T.; Suzuki, H.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Komatsu, Y.; Okuno, S.; Takahashi, N.; et al. Epitope Mapping of an Anti-Mouse CXCR6 Monoclonal Antibody (Cx(6)Mab-1) Using the 2 × Alanine Scanning Method. Monoclon Antib Immunodiagn Immunother 2022, 41, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Li, G.; Asano, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Epitope Mapping of the Anti-Human CCR2 Monoclonal Antibody C(2)Mab-9. Monoclon Antib Immunodiagn Immunother 2022, 41, 150–156. [Google Scholar] [CrossRef]
- Chamorro, S.; Vela, M.; Franco-Villanueva, A.; Carramolino, L.; Gutiérrez, J.; Gómez, L.; Lozano, M.; Salvador, B.; García-Gallo, M.; Martínez, A.C.; et al. Antitumor effects of a monoclonal antibody to human CCR9 in leukemia cell xenografts. MAbs 2014, 6, 1000–1012. [Google Scholar] [CrossRef]
- Chain, B.; Arnold, J.; Akthar, S.; Brandt, M.; Davis, D.; Noursadeghi, M.; Lapp, T.; Ji, C.; Sankuratri, S.; Zhang, Y.; et al. A Linear Epitope in the N-Terminal Domain of CCR5 and Its Interaction with Antibody. PLoS One 2015, 10, e0128381. [Google Scholar] [CrossRef]
- Gómez-Melero, S.; García-Maceira, F.I.; García-Maceira, T.; Luna-Guerrero, V.; Montero-Peñalvo, G.; Túnez-Fiñana, I.; Paz-Rojas, E. Amino terminal recognition by a CCR6 chemokine receptor antibody blocks CCL20 signaling and IL-17 expression via β-arrestin. BMC Biotechnol 2021, 21, 41. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Durán, G.; Romo-Mancillas, A. Behavior of Chemokine Receptor 6 (CXCR6) in Complex with CXCL16 Soluble form Chemokine by Molecular Dynamic Simulations: General Protein‒Ligand Interaction Model and 3D-QSAR Studies of Synthetic Antagonists. Life (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Wasilko, D.J.; Johnson, Z.L.; Ammirati, M.; Che, Y.; Griffor, M.C.; Han, S.; Wu, H. Structural basis for chemokine receptor CCR6 activation by the endogenous protein ligand CCL20. Nat Commun 2020, 11, 3031. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Orr, C.M.; Chan, H.T.C.; James, S.; Penfold, C.A.; Kim, J.; Inzhelevskaya, T.; Mockridge, C.I.; Cox, K.L.; Essex, J.W.; et al. Reducing affinity as a strategy to boost immunomodulatory antibody agonism. Nature 2023, 614, 539–547. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. RIEDL tag: A novel pentapeptide tagging system for transmembrane protein purification. Biochem Biophys Rep 2020, 23, 100780. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 162–167. [Google Scholar] [CrossRef]
- Rutihinda, C.; Haroun, R.; Saidi, N.E.; Ordoñez, J.P.; Naasri, S.; Lévesque, D.; Boisvert, F.M.; Fortier, P.H.; Belzile, M.; Fradet, L.; et al. Inhibition of the CCR6-CCL20 axis prevents regulatory T cell recruitment and sensitizes head and neck squamous cell carcinoma to radiation therapy. Cancer Immunol Immunother 2022. [CrossRef]
- Chen, K.J.; Lin, S.Z.; Zhou, L.; Xie, H.Y.; Zhou, W.H.; Taki-Eldin, A.; Zheng, S.S. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS One 2011, 6, e24671. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Cao, L.; Zhu, Y.; Cao, J.; Li, X.; Zhou, J.; Liu, B.; Zhao, T. Enhance anti-lung tumor efficacy of chimeric antigen receptor-T cells by ectopic expression of C-C motif chemokine receptor 6. Sci Bull (Beijing) 2021, 66, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Chen, J. CAR-T cell engineering with CCR6 exhibits superior anti-solid tumor efficacy. Sci Bull (Beijing) 2021, 66, 755–756. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.H.; Kang, M.; Jeon, M.; Chung, Y.; Kim, A.R.; Lee, Y.J.; Kim, E.S.; Nam, H.; Park, J.; Lee, J.Y.; et al. CEACAM1 marks highly suppressive intratumoral regulatory T cells for targeted depletion therapy. Clin Cancer Res 2023. [CrossRef]
Figure 1.
Determination of C6Mab-13 epitope by ELISA using alanine-substituted peptides of mCCR6. (A) Synthesized peptides of mCCR6 (10 µg/mL respectively) were immobilized on immunoplates for 30 min at 37℃. The plates were incubated with 10 µg/mL of C6Mab-13, followed by the treatment of peroxidase-conjugated anti-rat immunoglobulins. Optical density was measured at 655 nm (OD655) using a microplate reader. (B) mCCR6 and the epitope of C6Mab-13 were schematically illustrated. The C6Mab-13 epitope of mCCR6 comprises Asp11 from ELISA experiments.
Figure 1.
Determination of C6Mab-13 epitope by ELISA using alanine-substituted peptides of mCCR6. (A) Synthesized peptides of mCCR6 (10 µg/mL respectively) were immobilized on immunoplates for 30 min at 37℃. The plates were incubated with 10 µg/mL of C6Mab-13, followed by the treatment of peroxidase-conjugated anti-rat immunoglobulins. Optical density was measured at 655 nm (OD655) using a microplate reader. (B) mCCR6 and the epitope of C6Mab-13 were schematically illustrated. The C6Mab-13 epitope of mCCR6 comprises Asp11 from ELISA experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



