
Submitted:
03 March 2023
Posted:
06 March 2023
You are already at the latest version
Abstract
Sepsis is a life-threatening condition with high hospital mortality. Elevated mortality has also been observed in patients after hospital discharge due to post-sepsis syndrome (PSS). The etiology of PSS is still not entirely known, but it involves inflammation. Plasma extracellular vesicles (EVs) are recognized as a unique mechanism of intercellular communication in inflammatory processes. It has been reported that EV microRNA (miRNA) production during the acute sepsis phase may persist until after disease resolution and is associated with PSS.
We employed mass spectrometry and qPCR analysis to determine the protein and miRNA composition of plasma-derived EVs of 36 patients during sepsis-related hospitalization, immediately after ICU discharge (post-sepsis), and three, six, twelve, and up to 36 months post-sepsis.
We determined that patients’ immune system cells were the primary EV source. Fifteen differentially expressed EV miRNAs (DEmiRs) were identified in samples from septic patients compared to the control group. Predictive analyses revealed that these DEmiRs could influence inflammation by modulating pathways mediated by NF-κB, STAT3, and TLR4 signaling activation. Thirteen miRNAs (-15b-5p,-16-5p,-20a-5p,-25-3p,-27a-3p,-29a-3p,-30d-5p,-93-5p,-146a-5p,-148a-3p,-191-5p,-195-5p,-223-3p) were downregulated in the death group compared to the survivor group, making them candidate prognostic markers of ICU survival. One year after ICU discharge, the expression of miR-21-5p and miR-195-5p were decreased in the survivor group.
The miRNAs identified in the present study represent potential biomarkers for the survival prognosis of post-sepsis patients.
Keywords:
septicemia
; septic shock
; post-sepsis syndrome
; cytokines
; inflammation
1. Introduction
Sepsis is characterized by a dysregulated systemic response to microbial pathogens resulting in a disproportionate inflammatory response, multiple organ failure and/or death [1]. According to the World Health Organization [2], sepsis affects ~49 million people and accounts for 19.7% (~11 million) of annual deaths worldwide [3].
Sepsis mortality is estimated to be 25-30% [4] for patients not requiring admission to an intensive care unit (ICU) and 42% for those admitted [2]. Many septic patients die following hospital discharge due to sepsis sequelae. For example, after one year, one in three adult post-sepsis patients die [2]. After five years, sepsis patients have a mortality rate of 82–100% [5,6]. Moreover, ~50% of post-sepsis deaths are caused by recurrent infections, cardiovascular diseases [7] and post-sepsis syndrome (PSS) [2,8]. It has been reported that PSS affects one in six survivors [2,8], impairing immune, cognitive, psychiatric, cardiovascular, and renal functions and ultimately reducing the patient's quality of life. However, it remains unclear why some patients develop this condition, and others do not [6,9,10,11,12].
Sepsis stimulates immune and inflammatory responses with marked increases in plasma cytokine levels [13] that can persist at a low subclinical level even after hospital discharge [14]. Communication between immune and organ cells influences the events triggered by invasive pathogens [15]. Intercellular extracellular vesicle (EV) mediated communication exists in inflammatory and malignant diseases. These EVs are released from the surface of cells, endosomes and plasma membranes [16] and transport cargo, including [17,18] cell-originated proteins, lipids, DNA, mRNA, and microRNAs (miRNAs). Previous proteomic analyses identified cell/tissue-specific EVs [19,20]. In this sense, EV protein and miRNA content alterations have diagnostic and therapeutic potential [21,22,23,24].
EV miRNAs consist of small (20-25 nucleotides) non-coding RNA that can post-transcriptionally regulate gene expression by modulating mRNA translation and degradation [25]. Each miRNA can target hundreds of mRNAs, and multiple miRNAs can control the expression of a single mRNA [26]. The action of EV miRNAs on receptor cells varies depending on EV cell origin and gene specificity of the miRNA. Additionally, miRNA expression varies dynamically in healthy and diseased states due to their roles in intracellular processes and cell homeostasis [27].
Plasma procalcitonin (PCT) [28], C-reactive protein (CRP) [29], lactate [4,30], and interleukin-6 (IL-6) levels during the acute sepsis phase have been used as biomarkers for diagnosing and predicting septic patient outcomes [31]; however, these molecules sometimes exhibit high plasma levels in non-septic patients [32]. Interestingly, altered EV miRNA expression that persists even after sepsis resolution has been associated with increased PSS-linked patient mortality rate following hospital discharge [33]. Several studies have shown that miRNAs can regulate TLR-mediated innate immune response, NF-κB signaling, and inflammatory cytokine production [16]. Furthermore, altered miRNA expression can aggravate the disease course and enhance PSS mortality [34].
Previous investigations were performed using patient data after ICU admission and/or during hospitalization to evaluate the diagnostic and prognostic potential of EV miRNAs [15,27,33,35,36,37]. However, none of these studies followed up on the EV miRNA alterations after ICU discharge. Thus, little is known about the effect of sepsis on long-term EV miRNA expression patterns. Herein, we sought to determine EV cell origin and identify potential disease and survivor biomarkers by monitoring EV miRNA expression patterns of septic patients from disease onset up to three years following hospital discharge.
2. Results and Discussion
2.1. Clinical data and Characterization of extracellular vesicle
As shown in Table 1, no significant differences were found in the clinical characteristics (complete data in Supplementary Material Tables S2 and S3). The majority of patients were male (63.9%). Hypertension (58.3%) and diabetes (38.9%) were the most frequent comorbidity in septic patients. Pulmonary infections were the predominant cause of sepsis (33.3%), while abdominal cavity infections accounted for 25% of the cases. Of the total number of enrolled patients, 47.2% died during the study. Complementary laboratory data can be accessed in Gritte, Souza-Siqueira et al., 2022.
EVs were isolated from human plasma and analyzed using a Nanoparticle Tracking Analysis (NTA) technique, immunodetection protein CD9, CD63, HSP70, Cytochrome and Transmission Electron Microscopy (TEM) (Supplementary Material - Figure S1-S3). The EVs presented the characteristic circular shape and size range, as described by Willms, et al. [38].
We cannot exclude the possible interference of contaminants in this analysis since apolipoprotein contamination was detected in all samples analyzed. Nor can we say that all microRNAs come exclusively from EVs. Additionally, as reported by Karttunen, et al. [39], there is the possibility of plasma miRNA and lipoprotein co-precipitation.
2.2. Systemic Inflammatory mediators
Serum CRP concentrations in the survivor and death groups were significantly higher than the reference (5 mg/L) and control group values during phases A and B. The plasma CRP levels were increased in phase B of the death group compared to survivors in the same phase. In phases C-F, plasma CRP concentrations of the survivor group remained above reference values but were not significantly different from the control group (Supplementary Material Table S4).
In control groups, only IL-6 was upregulated in phases A and B of survivor patients (Figure 1D and Supplementary Material Table S5).
In over time analyses by vital status (death or survivor) of septic patients, there were differences in the plasma levels of IL-10, TNF-α, and IL-1β between the two groups. The results of these analyses are presented in Supplementary Material Table S6.
Additionally, survivor and dead septic patients were compared in phase A by vital status, and patients of the death group showed higher plasma levels of IL-6 and IL-10 (Supplementary Material Table S7).
2.3. Functional-enrichment analysis for Gene Ontology terms
Comparing the global analysis of plasma EVs by mass spectrometry and searching the Uniprot database, 125 proteins were found in the two groups (Supplementary Material Table S8). The cellular components’ analyses through PANTHER indicated that 88 (70%) of the proteins are EV marker proteins (p=8.97-57).
The biological process with the highest number of identified proteins was the response to stimuli, with 86% (n=107; p=2.02-26), followed by the regulation of biological processes, 78% (n=98; p=6.96-07). The response to stress was 71% (n=89; p=2.80-41), the immune system functioning was 61% (n=76; p=5.64-39), and the immune response was 54% (n=68; p=2.85-41).
The featured PANTHER protein class was immunoglobulin, with twenty-six representatives identified using proteomics.
For tissue specificity, the Human Protein Atlas indicated that 27% of proteins identified in proteomics are from lymphoid tissues (Supplementary Material Table S9).
Next, we sought to identify which proteins presented statistical differences among the groups. Of the 125 proteins, 48 had statistical differences in the death group compared with survivors. According to PANTHER, the biological processes with significant relevance are the response to stimulus (77%; n=37; p=1.36-07), immune system functioning (52%; n=25; p= 8.14-12) and defense response (48%; n=23; p=2.01-14).
2.4. Expression profile of DEmiRs in septic patients
Fifteen of the 84 qPCR array miRNAs were differentially expressed (DEmiRs) and downregulated in the death group compared to survivors. The mirBase IDs, p-values, and fold change of these miRNAs are summarized in Supplementary Material Table S10. The functional Enrichment (FunRich) analysis tool v3.1.4 [40] revealed that these 15 miRNAs modulate 3,219 targets and the MiRWalk 3.0. databank [41] predicted that 14 DEmiRs modulate the expression of common mRNAs.
All miRNAs were downregulated in phase B when comparing all phases of the survivor and control groups (Figure 2A-2O and Supplementary Material Table S11). However, in phase A, only miR–29a-3p, 146a-5p, and 148a-3p were not downregulated. No statistical differences were detected in Phases C and F. In Phase D, all miRNAs, except for miR-25-3p and miR-126-3p, were also downregulated. Furthermore, in Phase E, only miRNAs 21-5p and 195-5p were downregulated.
In general, miRNAs exhibited similarity in phases' expression trend (Figure 3). The expression of all these miRNAs changed over time, with differences between the death and survivor groups, except for miRNA 29a-3p and 146a-5p. Complete data can be found in Supplementary Material Table S12.
Septic patients' mortality over time followed an increase-decrease dynamic for all miRNAs expression. Although Phases A and B exhibited low expression of all miRNAs, no significant difference was detected between them. Besides miR-146a-5p, Phases C and D had increased and decreased miRNAs expression, respectively. In phase E, there was a statistically significant increase of miR-21-5p compared to the previous phase. We also observed instability in the dynamics in Phase F, in which the expression of some miRNAs was downregulated and others upregulated, but none differed between groups (Supplementary Material Table S13).
There was no difference between the survivor and the death groups for miRNAs 21-5p and 126-3p expression, and the other miRNAs were significantly lower in vital status from phase A (Supplementary Material Table S14).
Lehner, et al. [42] stated that the source of plasma EVs could promote sepsis progression or resolution [16]. Specific proteins found in EVs usually indicate their tissue or organ's origin, which can be identified by proteomic approaches [19,20]. Although protein specificity is related to its origin, the EV pool is not exclusively from one tissue only [19,20]. Herein, we identified 125 proteins in plasma EVs collected during the acute phase of sepsis in ICU-admitted patients. Of this total, 34 proteins are from lymphoid tissues, and 20 originated from B lymphocytes, strongly indicating the immune cell origin [43,44]. Many of these proteins are related to the acute inflammatory response, which significantly affects the immune response, innate immune function, and homeostasis. For example, the protein alpha-2-macroglobulin (A2MG) - detected in proteomic analysis in both groups, according to Lashin, et al. [45], is expressed in neutrophil microvesicles and has an important role as a protective factor in experimental sepsis as well as an association with high survival rates.
Intracellular LPS plays a vital role in activating the immune response in vitro and can induce increased cytotoxic activity. Upon initial events that trigger the immune response, LPS can be cell internalized by different mechanisms, such as the LPS-LBP complex. Lipopolysaccharide-Binding protein (LBP) is an acute phase protein that promotes an immune response after recognition and binding to bacterial LPS; once this complex is formed, it reaches the cytosol and activates intracellular receptors that generate cytotoxic effects. Once activated, the cell secretes IL-1α/β, IL-6, and IL-18 and undergoes pyroptosis, an inflammatory form of cell death. Macrophage pyroptosis also exacerbates the inflammation state, contributing to high mortality in sepsis [46].
We identified the presence of LBP in EVs in the survivor and death groups. During the inflammatory process, EVs have a relevant role in intercellular communication. It is plausible that EVs carry the LPS-LBP complex from one cell to another to enhance the immune response mediated by the activation of intracellular LPS signaling.
Another important mechanism in the internalization of LPS is via HMGB1, which binds to LPS and guides its internalization to macrophage lysosomes, leading to the activation of caspase-11 and pyroptosis [47]. Previously our group demonstrated upregulated gene expression of products and mediators of the non-classical (IL-1α and HMGB1 and caspase 11) and classical (IL-1β, IL-18, NLRP3, CASPASE 1) inflammation pathways in the monocytes of septic patients [48]. The increased expression of HGMB1 and LBPA proteins may be closely linked to the augmented inflammatory process and its mediators' production by activating the non-classical and classical pathways.
We reported a marked difference between the death and survivor groups regarding the septic state (Phase B) changes in the EV protein composition. It is plausible that sepsis-induced EV cargo alterations could affect cell communication and function [21,22,23].
The functional enrichment by GO indicated that biological processes with greater participation of EV proteins are related to stimulus, stress, and immune system functioning. This result may suggest that sepsis provides a notable stimulus for regulators' release through EVs, especially those from tissue and immune system cells. The expression of the 48 proteins between both groups was statistically different, and the expression of EVs proteins was continuously related to the immune system functioning, stimulus and defense response and biological process regulation.
We also monitored and compared the survivor and death groups' plasma EV miRNA expression in the ICU with a control group. Fifteen DEmiRs related to NF-κB signaling pathway regulation were identified, and all were downregulated in the death group compared to the survivors. These changes in EV miRNA expression, combined with plasma CRP and cytokine concentration variations, indicate persistent low-grade inflammation lasting up to three years post-sepsis. Concomitantly it is liked that sepsis induces epigenetic reprogramming.
Tissue-released EVs circulate in the blood, reaching distant tissues and delivering molecules that can activate or inhibit cell functions. Depending on the situation, EVs can act as a "friend" or "foe" in systemic inflammatory reactions [16]. EVs can carry pro-inflammatory miRNAs that can activate cell signaling and induce strong inflammatory processes during sepsis [43,44]. Predictive analysis of the targeted miRNA from the 15 DEmiRs described here indicated that these miRNAs are predominantly associated with inflammation, modulating the pathways mediated by NF-κB, STAT3, and TLR4 activation. Except for miR-29a-3p, the other miRNAs are negative modulators of inflammation [49].
Previous studies reported that reducing miR-15b-5p, -16-5p, -20a-5p, -146a-5p, and -223-3p expression promotes NF-κB complex migration to the nucleus and subsequent inflammatory gene expressions [33,35,49,50,51,52,53,54,55,56,57]. On the other hand, miR-29a-3p targets insulin receptor substrate-1 (IRS-1) and Grb2-associated binder-1 (Gab1) [49]. Increased Gab1 expression inhibits TNF-α-mediated NF-κB transcriptional activation and reduces pro-inflammatory cytokines expression through the NF-κB pathway. These observations could be a compensatory mechanism since only miR-29a-3p downregulation attenuates pro-inflammatory cytokines production.
The underlying mechanisms of miR-25-3p and miR-30d-5p action are still unknown. The attenuated miR-25-3p expression has been shown to increase plasma TNF-α concentrations and is linked to the degree of oxidative stress via NADPH oxidase 4 (NOX 4) [58,59]. The expression of miR-30d-5p is inversely related to plasma concentrations of CRP, IL-1β, IL-8, and IL-6. We observed reduced miRNA –25-3p and miR-30d-3p levels in the acute phase of the survivor and death groups, whereas the cytokines IL-1β, IL-8 and IL-6, and CRP levels were increased. The downregulation of miR-27a expression increases TLR4 expression, pro-inflammatory cytokine (e.g., IL-6, IL-1β, and TNF-α) concentrations, and nitric oxide production [60]. In our study, miR-27a expression was reduced during the acute phase and increased one year after ICU discharge. Previously, we showed increased gene expression of TLR4 receptors in macrophages from the same patients [48].
In the acute phase, the death group exhibited higher plasma levels of IL-6 and IL-10 and a decrease in the 13 miRNAs (except for miR126-3p and miR21-5p) (Figure 2 and Supplementary Material Table S15).
Since most deaths occur in phase A by multiple organ failure associated with higher and dysregulated inflammatory response, plasmatic cytokine concentrations were correlated to the inflammation degree, organic dysfunction gravity, and sepsis-associated mortality [31,61,62,63]. IL-12 generates isoforms (IL-12p40 and IL-12p70) in human polymorphonuclear leukocytes and monocytes/macrophages during the severe sepsis phase [64]. Overproduction of IL-12p40 leads to sepsis-related immunosuppression [64]. Even though the survivors had higher expression of the IL-12p70 isoform than the death group (Figure S4 of the Supplementary Material). This finding is associated with plasma CRP concentrations that remained elevated for up to three years, as previously reported by Iwashyna, et al. [65].
3. Experimental Section
3.1. Patient selection
Patients who developed sepsis or experienced septic shock and were admitted to the intensive care unit (ICU) of the University of Sao Paulo Hospital (HU-USP) participated in the study. Patients under 18 years of age, with HIV, chronic hepatitis, neoplasms or autoimmune or inflammatory diseases, or taking anti-allergic or anti-inflammatory drugs were excluded.
Blood samples were collected during the acute sepsis phase (by ICU day 4, phase A), immediately after ICU discharge (phase B), and three months (phase C), six months (phase D), one year (phase E), and three years (phase F) after hospital discharge. Sixteen non-septic control volunteers were paired according to age and gender with the septic patients.
Sixty-two patients were initially eligible, but nine did not meet the inclusion criteria, ten were not located after three attempts, and seven refused to participate. The remaining 36 patients were monitored and eventually divided into two outcome groups: survivor (n=19) and death (n=17). The survivor group included patients who survived up to one year after ICU discharge, and the death group included those who died in the same timeframe.
Nineteen samples from phases A–E were collected and analyzed for the survivor group. Phase F included data from eight patients because six died during phase E, three were not located, and two were readmitted to the ICU at sample collection time.
The death group consisted of 17 patients. Ten died during phase A, and seven passed away during phase B.
3.2. Blood collection and plasma separation
Blood samples (20 mL) were collected from the antecubital vein and transferred to ethylenediaminetetraacetic acid (EDTA) containing tubes. Plasma samples were obtained by centrifugation at 1,900×g for 10 minutes at 4°C and kept at -80°C until analysis.
3.3. Evaluation of inflammatory cytokines and serum C-reactive proteins levels
The plasma IL-12p70, IL-6, IL-8, IL-10, TNF-α, and IL-1β concentrations were determined using the BD CBA Human Inflammatory Cytokine kit (BD, Franklin Lakes, NJ, USA) according to the manufacturer's instructions. Data were acquired on a BD Accuri Flow Cytometer. Cytokine concentrations were calculated using the FCAP Software v3.0 (BD). The kit’s limits of detection are as follows: IL-12p70 (1.9 pg/mL), IL-6 (2.5 pg/mL), IL-8 (0.2 pg/mL), IL-10 (3.3 pg/mL), TNF-α (3.7 pg/mL), and IL-1β (7.2 pg/mL).
Serum CRP concentrations were measured using the nephelometric method at the HU-USP Clinical Analysis Laboratory.
3.4. Plasmatic extracellular vesicle isolation
Plasma EVs were isolated by ultracentrifugation (Beckman Coulter-Optima L-90K, Brea, CA, USA) using an adapted protocol from Thierry, et al. [66]. Briefly, the blood samples were defrosted, residual cells, debris, platelets, and apoptotic bodies were sedimented by centrifugation at 2,500×g for 15 minutes at 4°C [67], and the supernatant was transferred to a sterile tube. Plasma samples (2 mL) were diluted with 24 mL of Nanopure water (SW28, Beckman Coulter) and centrifuged at 100,000×g for 1 hour at 4ºC. Afterward, the supernatant was discarded, and the pellet was resuspended in 26 mL of Nanopure water and centrifuged (100,000×g, 1 hour, 4ºC). The supernatant was again discarded, and the pellet was resuspended in 1 mL of Nanopure water. Samples were stored at -80°C until the proteomics and miRNA differential expression experiments were conducted.
3.5. Proteomic analysis by mass spectrometry and data processing
Proteomic analyses were performed in random samples of six patients with sepsis (three survivors and three deaths) by the LTQ-Orbitrap Velos Electron Transfer Dissociation (ETD) Mass Spectrometer (Thermo), attached to the Easy nanoLC II (Thermo). Proteomic analyses were performed at the Facilities for Research Support Center of the Institute of Biomedical Sciences at the University of São Paulo (CEFAP-ICB-USP). Fifty µg of total EV protein was digested with trypsin, diluted in 100 µL of Nanopure water. The peptides were separated on a C18 PicoFrit column (C18 PepMap, 75 µm id × 10 cm, 3.5 µm particle size, 100 Å pore size; New Objective, Ringoes, NJ, USA) with a 115 min gradient. Trypsin-digested BSA (100 fmol) was used to verify instrumental conditions and as the standard. To identify putative proteins, we searched the Uniprot v15.0 database [69] using the term "Homo_sapiens_reviewed_uniprot" (https://www.uniprot.org). The protein content was quantified with the MaxQuant v1.6.2.3 software [70]. Results were normalized and compared using unilateral ANOVA (p≤0.05) with the Perseus v1.6.2.2 software [71]. The analysis of functional enrichment by Gene Ontology (GO) was performed using the Protein Analysis Through Evolutionary Relationships (PANTHER) v17.0 classification system [72], with the statistical overrepresentation test: Test Type = Fisher's Exact and Correction = Calculate False Discovery Rate-FDR. The proteins identified in the proteomics experiments were grouped according to cellular component, biological process, and PANTHER protein class. The Atlas of Human Protein v21.1 [73] was used for tissue and cell specificity analyses.
3.6. Plasmatic extracellular vesicles miRNA determination
EV miRNA concentrations were measured using the Human Serum & Plasma microRNA PCR array plate (MIHS-106Z, Qiagen, Hilden, Düsseldorf, Germany). The miRNA of six randomly selected phase B samples from the survivor (n=3) and death (n=3) groups. We used the miRNeasy Serum/Plasma kit (Qiagen) to purify total RNA, including miRNA. The RNA's quantity and purity were assessed spectrophotometrically at 260/280 nm using a NanoDrop 2000 spectrophotometer (Thermo). Complementary DNA (cDNA) was synthesized using the miScript II RT kit (Qiagen) with 1.5 μL of miRNA (final reaction volume=10 μL) in a LongGene MG96G thermal cycler (LongGene, Zhejiang, Hangzhou, China).
Pre-amplified miScript miRNA Quality Control PCR Array (Qiagen) plates were included to verify the quality and integrity of the samples. Pre-amplification was performed using the miScript PreAMP PCR kit (Qiagen) with 5 µL of cDNA. The pre-amplified cDNA was diluted 20-fold and stored at -80°C. For the PCR array analysis, 12.5 µL SYBR Green PCR Master Mix, 2.5 µL Universal Prime (10×), 9 µL RNase water-free and 1 µL pre-amplified cDNA (total volume=25 µL) were added to each well of the miRNA PCR Array plate. The plate was centrifuged at 1,000×g for one minute at room temperature. Data were recorded with a QuantStudio 3 device and analyzed using the QuantStudio 3 Design and Analysis Software v1.5.1 (Thermo).
The global mean normalization method was used to normalize the miRNA PCR array data [74]. Initially, data were calibrated by spiking samples with the exogenous control cel-miR-39. Then the global mean of the CT was calculated for the miRNA targets commonly expressed in all samples and presented a CT<30. The 2-∆∆CT calculation determined the relationship between the miRNA target CT and global CT [75].
The miRNAs that presented between-group expression differences in the array assay were evaluated in all phases of all samples [survivor group (n=19), phase A-E (n=19) and phase F (n=8), death group (n=17), phase A (n=10) and phase B (n=7), and control group (n=16)] using Real-Time Polymerase Chain Reaction (qPCR). In this case, the final volume of the cDNA preparation (described above) was not adjusted since pre-amplification was unnecessary. The final reaction volume (20 μL) contained 20 ng of miRNA. The cDNA was diluted in 300 µL RNase-free water for the qPCR assay.
The qPCR was performed using the miScript SYBR Green PCR kit (Qiagen) with 96-well plates. Each well contained 12.5 µL SYBR Green PCR Master Mix, 2.5 µL primer universal (10×), 2.5 µL Primer Assay (Exxtend, Paulínia, SP, Brazil), 5 µL RNase-free water and 2.5 µL cDNA. These experiments utilized the same protocol as the Array plate.
The primer sequences of the constitutive gene candidates and the algorithms of BestKeeper, GeNorm and NormFinder software were utilized to determine the comparative quantification cycle (Cq) values (Table S1).
3.7. Statistics
After analyzing data distribution (Kolmogorov-Smirnov), we applied the nonparametric Mann-Whitney test for quantitative variables of independent groups, followed by the GEE (Generalized Estimating Equations) method on data over time. In GEE's criteria, factors were considered as time (phase A-F), status vital (survivors or deaths) and interaction between both groups. The post-hoc Bonferroni Correction was performed to identify which time factor differs statistically. All data analyses were performed with the SPSS software v25 (IBN, Armonk, NY). The significance level was set at 5% (p<0.05).
4. Conclusions
Our proteomics data showed that the EVs mainly originated from immune cells. The DEmiR of EVs was downregulated in the acute phase, whereas pro-inflammatory cytokine expression was upregulated. Moreover, the expression of miRNAs was downregulated in the EVs of the death group compared to survivors, representing potential survival biomarkers. The expression of two DEmiRs remained attenuated after one year of ICU discharge. These chronic conditions may predispose patients to new infections, myocardial infarction, neoplasms, and/or organ failure and PSS, leading to death.
5. Declarations
5.1. Ethics approval and consent to participate
The study was approved by the Research Ethics Committee of the Hospital of the University of São Paulo under registration 1513/16. All patients, or their legal guardians, signed the informed consent form.
5.2. Limitations of the study
There are limitations in the study to be mentioned. The study was carried out only with patients admitted to the ICU. Knowing that approximately 55% of septic patients are seen in the medical clinic, the results may only reflect more severe states of sepsis. The patient selection was based on the hypothesis that the sickest patients have more pronounced symptoms.
The presence of lipoproteins may be indicative of contamination by lipoproteins, as well as we cannot state that miRNAs are unique to EVs. As described by Karttunen, Heiskanen, Navarro-Ferrandis, Das Gupta, Lipponen, Puhakka, Rilla, Koistinen and Pitkänen [39], there is the possibility of co-precipitation of plasmatic miRNAs.
This phenomenon may indicate that patients who die have an increased release of vesicles from the immune origin with lower content of specific miRNAs, as stated in the evaluation of the miRNAs in the acute phase. Since our proteomics data are limited to the acute sepsis phase, more studies are needed to assess whether this condition persists post-sepsis.
5.3. Data availability statements
Original datasets are available in a publicly accessible repository: The original contributions presented in the study are publicly available: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE210335. Data are available via ProteomeXchange with the identifier PXD035804.
5.4. Competing interests
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
5.5. Authors' contributions
Conceptualization, Talita Souza-Siqueira, Raquel Gritte, Laureane Masi, Fernanda Borges and Renata Gorjão; Data curation, Talita Souza-Siqueira, Raquel Gritte and Tânia Pithon-Curi; Funding acquisition, Tânia Pithon-Curi, Ricardo Azevedo and Marcel Machado; Investigation, Talita Souza-Siqueira, Raquel Gritte and Eleine Weimann; Methodology, Talita Souza-Siqueira, Raquel Gritte, Eleine Weimann, Juliana Vieira, Laureane Masi, Fernanda Borges, Gilson Murata, Laiane Oliveira, Silvana Bordin and Eliane Silva; Project administration, Talita Souza-Siqueira, Tânia Pithon-Curi, Rui Curi, Marcel Machado and Francisco Soriano; Resources, Talita Souza-Siqueira, Raquel Gritte, Rodrigo Borges, Antônio Nogueira, Rui Curi and Marcel Machado; Supervision, Laureane Masi, Fernanda Borges, Silvana Bordin, Adriana Levada-Pires, Tânia Pithon-Curi, Rui Curi, Marcel Machado and Francisco Soriano; Validation, Talita Souza-Siqueira and Raquel Gritte; Visualization, Talita Souza-Siqueira and Raquel Gritte; Writing – original draft, Talita Souza-Siqueira, Raquel Gritte, Eleine Weimann, Laureane Masi, Fernanda Borges, Renata Gorjão, Rui Curi, Marcel Machado and Francisco Soriano; Writing – review & editing, Talita Souza-Siqueira, Raquel Gritte, Eleine Weimann, Fernanda Borges, Rui Curi and Marcel Machado.
5.6. Funding
The São Paulo State Research Foundation - FAPESP (2015/04138-2, 2017/13715-9, 2018/09868-7, and 2021/08624-0), Coordination for the Improvement of Higher-Level Personnel - CAPES (88882.365194/2019-01; 88882.365195/2019-01), and the National Council for Scientific and Technological Development - CNPq (316072/2020-6) supported this study.
5.7. Acknowledgements
The authors are grateful to the nursing team and the clinical analysis laboratory, at the Hospital of University de São Paulo, for the help with patients; to Larissa Lima and Prof. Dr. Cristoforo Scavone from the Institute of Biomedical Sciences – USP, EVs centrifugation and preparation; Allan Munford and Juliana Ramos, from Qiagen, for technical-scientific advice; Thais Viggiani Santana, from CEFAP-ICB-USP, for preparing proteomics. We are also grateful to Dr. Robert Ryan for proofreading the text.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Singer, M.; Deutschman, C. S.; Seymour, C. W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G. R.; Chiche, J.-D.; Coopersmith, C. M., The third international consensus definitions for sepsis and septic shock (Sepsis-3). Jama 2016, 315, (8), 801-810.
- Organization, W. H., Global report on the epidemiology and burden of sepsis: current evidence, identifying gaps and future directions. 2020.
- Rudd, K. E.; Johnson, S. C.; Agesa, K. M.; Shackelford, K. A.; Tsoi, D.; Kievlan, D. R.; Colombara, D. V.; Ikuta, K. S.; Kissoon, N.; Finfer, S., Global, regional, and national sepsis incidence and mortality, 1990–2017: analysis for the Global Burden of Disease Study. The Lancet 2020, 395, (10219), 200-211.
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A., Sepsis and septic shock. The Lancet 2018, 392, (10141), 75-87.
- Iwashyna, T. J.; Cooke, C. R.; Wunsch, H.; Kahn, J. M., Population burden of long-term survivorship after severe sepsis in older Americans. Journal of the American Geriatrics Society 2012, 60, (6), 1070-1077.
- Quartin, A. A.; Schein, R. M.; Kett, D. H.; Peduzzi, P. N., Magnitude and duration of the effect of sepsis on survival. Jama 1997, 277, (13), 1058-1063.
- Wang, H. E.; Szychowski, J. M.; Griffin, R.; Safford, M. M.; Shapiro, N. I.; Howard, G., Long-term mortality after community-acquired sepsis: a longitudinal population-based cohort study. BMJ open 2014, 4, (1).
- Mostel, Z.; Perl, A.; Marck, M.; Mehdi, S. F.; Lowell, B.; Bathija, S.; Santosh, R.; Pavlov, V. A.; Chavan, S. S.; Roth, J., Post-sepsis syndrome–an evolving entity that afflicts survivors of sepsis. Molecular Medicine 2020, 26, (1), 1-14.
- Batista Lorigados, C.; Garcia Soriano, F.; Szabo, C., Pathomechanisms of myocardial dysfunction in sepsis. Endocrine, Metabolic & Immune Disorders-Drug Targets (Formerly Current Drug Targets-Immune, Endocrine & Metabolic Disorders) 2010, 10, (3), 274-284.
- van der Slikke, E. C.; An, A. Y.; Hancock, R. E.; Bouma, H. R., Exploring the pathophysiology of post-sepsis syndrome to identify therapeutic opportunities. EBioMedicine 2020, 61, 103044.
- Angus, D. C.; Linde-Zwirble, W. T.; Lidicker, J.; Clermont, G.; Carcillo, J.; Pinsky, M. R., Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Read Online: Critical Care Medicine| Society of Critical Care Medicine 2001, 29, (7), 1303-1310.
- Prescott, H. C.; Langa, K. M.; Liu, V.; Escobar, G. J.; Iwashyna, T. J., Increased 1-year healthcare use in survivors of severe sepsis. American journal of respiratory and critical care medicine 2014, 190, (1), 62-69.
- Delano, M. J.; Ward, P. A., The immune system's role in sepsis progression, resolution, and long-term outcome. Immunological reviews 2016, 274, (1), 330-353.
- Gritte, R. B.; Souza-Siqueira, T.; Curi, R.; Machado, M. C. C.; Soriano, F. G., Why Septic Patients Remain Sick After Hospital Discharge? Frontiers in Immunology 2021, 11, 3873.
- Reithmair, M.; Buschmann, D.; Märte, M.; Kirchner, B.; Hagl, D.; Kaufmann, I.; Pfob, M.; Chouker, A.; Steinlein, O. K.; Pfaffl, M. W., Cellular and extracellular mi RNA s are blood-compartment-specific diagnostic targets in sepsis. Journal of cellular and molecular medicine 2017, 21, (10), 2403-2411.
- Raeven, P.; Zipperle, J.; Drechsler, S., Extracellular vesicles as markers and mediators in sepsis. Theranostics 2018, 8, (12), 3348.
- Chaput, N.; Théry, C. In Exosomes: immune properties and potential clinical implementations, Seminars in immunopathology, 2011; Springer: 2011; pp 419-440.
- D’Souza-Schorey, C.; Schorey, J. S., Regulation and mechanisms of extracellular vesicle biogenesis and secretion. Essays in biochemistry 2018, 62, (2), 125-133.
- Yáñez-Mó, M.; Siljander, P. R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F. E.; Buzas, E. I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J., Biological properties of extracellular vesicles and their physiological functions. Journal of extracellular vesicles 2015, 4, (1), 27066.
- Doyle, L. M.; Wang, M. Z., Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, (7), 727.
- Escola, J.-M.; Kleijmeer, M. J.; Stoorvogel, W.; Griffith, J. M.; Yoshie, O.; Geuze, H. J., Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human B-lymphocytes. Journal of Biological Chemistry 1998, 273, (32), 20121-20127.
- de Gassart, A.; Géminard, C.; Février, B.; Raposo, G.; Vidal, M., Lipid raft-associated protein sorting in exosomes. Blood 2003, 102, (13), 4336-4344.
- Mathivanan, S.; Ji, H.; Simpson, R. J., Exosomes: extracellular organelles important in intercellular communication. Journal of proteomics 2010, 73, (10), 1907-1920.
- Heijnen, H. F.; Schiel, A. E.; Fijnheer, R.; Geuze, H. J.; Sixma, J. J., Activated Platelets Release Two Types of Membrane Vesicles: Microvesicles by Surface Shedding and Exosomes Derived From Exocytosis of Multivesicular Bodies and-Granules. Blood, The Journal of the American Society of Hematology 1999, 94, (11), 3791-3799.
- Lin, S.-L.; Miller, J. D.; Ying, S.-Y., Intronic microrna (mirna). Journal of Biomedicine and Biotechnology 2006, 2006.
- Guo, H.; Ingolia, N. T.; Weissman, J. S.; Bartel, D. P., Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, (7308), 835-840.
- Goodwin, A. J.; Guo, C.; Cook, J. A.; Wolf, B.; Halushka, P. V.; Fan, H., Plasma levels of microRNA are altered with the development of shock in human sepsis: an observational study. Critical care 2015, 19, (1), 1-10.
- Sitar, M. E.; Ipek, B. O.; Karadeniz, A., Procalcitonin in the diagnosis of sepsis and correlations with upcoming novel diagnostic markers. Int J Med Biochem 2019, 2, (3), 132-40.
- Samraj, R. S.; Zingarelli, B.; Wong, H. R., Role of biomarkers in sepsis care. Shock (Augusta, Ga.) 2013, 40, (5), 358.
- Kang, H. E.; Park, D. W., Lactate as a biomarker for sepsis prognosis? Infection & chemotherapy 2016, 48, (3), 252.
- Weidhase, L.; Wellhöfer, D.; Schulze, G.; Kaiser, T.; Drogies, T.; Wurst, U.; Petros, S., Is Interleukin-6 a better predictor of successful antibiotic therapy than procalcitonin and C-reactive protein? A single center study in critically ill adults. BMC infectious diseases 2019, 19, (1), 1-7.
- Charalampos, P.; Vincent, J.-L., Sepsis biomarkers: a review. Critical care 2010, 14, R15.
- Essandoh, K.; Fan, G.-C., Role of extracellular and intracellular microRNAs in sepsis. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2014, 1842, (11), 2155-2162.
- Szilágyi, B.; Fejes, Z.; Pócsi, M.; Kappelmayer, J.; Nagy Jr, B., Role of sepsis modulated circulating microRNAs. Ejifcc 2019, 30, (2), 128.
- Wang, J.-f.; Yu, M.-l.; Yu, G.; Bian, J.-j.; Deng, X.-m.; Wan, X.-j.; Zhu, K.-m., Serum miR-146a and miR-223 as potential new biomarkers for sepsis. Biochemical and biophysical research communications 2010, 394, (1), 184-188.
- Roderburg, C.; Luedde, M.; Vargas Cardenas, D.; Vucur, M.; Scholten, D.; Frey, N.; Koch, A.; Trautwein, C.; Tacke, F.; Luedde, T., Circulating microRNA-150 serum levels predict survival in patients with critical illness and sepsis. PloS one 2013, 8, (1), e54612.
- Wang, H.; Meng, K.; jun Chen, W.; Feng, D.; Jia, Y.; Xie, L., Serum miR-574-5p: a prognostic predictor of sepsis patients. Shock 2012, 37, (3), 263-267.
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M. J.; Vader, P., Extracellular vesicle heterogeneity: subpopulations, isolation techniques, and diverse functions in cancer progression. Frontiers in immunology 2018, 9, 738.
- Karttunen, J.; Heiskanen, M.; Navarro-Ferrandis, V.; Das Gupta, S.; Lipponen, A.; Puhakka, N.; Rilla, K.; Koistinen, A.; Pitkänen, A., Precipitation-based extracellular vesicle isolation from rat plasma co-precipitate vesicle-free microRNAs. Journal of extracellular vesicles 2019, 8, (1), 1555410.
- Fonseka, P.; Pathan, M.; Chitti, S. V.; Kang, T.; Mathivanan, S., FunRich enables enrichment analysis of OMICs datasets. Journal of Molecular Biology 2021, 433, (11), 166747.
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N., miRWalk: An online resource for prediction of microRNA binding sites. PloS one 2018, 13, (10), e0206239.
- Lehner, G.; Brandtner, A.; Joannidis, M., Microvesicles in sepsis: implications for the activated coagulation system. In Annual Update in Intensive Care and Emergency Medicine 2017, Springer: 2017; pp 29-39.
- Murao, A.; Brenner, M.; Aziz, M.; Wang, P., Exosomes in Sepsis. Frontiers in Immunology 2020, 11, 2140.
- Kalluri, R.; LeBleu, V. S., The biology, function, and biomedical applications of exosomes. Science 2020, 367, (6478).
- Lashin, H. M.; Nadkarni, S.; Oggero, S.; Jones, H. R.; Knight, J. C.; Hinds, C. J.; Perretti, M., Microvesicle subsets in sepsis due to community acquired pneumonia compared to faecal peritonitis. Shock 2018, 49, (4), 393-401.
- Gabarin, R. S.; Li, M.; Zimmel, P. A.; Marshall, J. C.; Li, Y.; Zhang, H., Intracellular and extracellular lipopolysaccharide signaling in sepsis: avenues for novel therapeutic strategies. Journal of Innate Immunity 2021, 13, (6), 321-330.
- Deng, M.; Tang, Y.; Li, W.; Wang, X.; Zhang, R.; Zhang, X.; Zhao, X.; Liu, J.; Tang, C.; Liu, Z., The endotoxin delivery protein HMGB1 mediates caspase-11-dependent lethality in sepsis. Immunity 2018, 49, (4), 740-753. e7.
- R.B. Gritte; T. Souza-Siqueira; E.B. Silva; L.C.S. Oliveira; R.C. Borges; H.H.O. Alves; L.N. Masi; G.M. Murata; R. Gorjão; A.C. Levada-Pires; A.C. Nogueira; T.C. Pithon-Curi; R. Bentes; F.G. Soriano; R. Curi; Machado, M. C. C., Evidence for Monocyte Reprogramming in a Long-Term Postsepsis Study. Critical Care Explorations 2022, 4, (8).
- Hu, Q.; Gong, W.; Gu, J.; Geng, G.; Li, T.; Tian, R.; Yang, Z.; Zhang, H.; Shao, L.; Liu, T., Plasma microRNA profiles as a potential biomarker in differentiating adult-onset Still's Disease from sepsis. Frontiers in immunology 2019, 9, 3099.
- Li, T.; Morgan, M. J.; Choksi, S.; Zhang, Y.; Kim, Y.-S.; Liu, Z.-g., MicroRNAs modulate the noncanonical transcription factor NF-κB pathway by regulating expression of the kinase IKKα during macrophage differentiation. Nature immunology 2010, 11, (9), 799-805.
- Ruan, L.; Qian, X., MiR-16-5p inhibits breast cancer by reducing AKT3 to restrain NF-κB pathway. Bioscience reports 2019, 39, (8).
- Huang, Y.; Yang, N., MicroRNA-20a-5p inhibits epithelial to mesenchymal transition and invasion of endometrial cancer cells by targeting STAT3. International journal of clinical and experimental pathology 2018, 11, (12), 5715.
- Zhao, C.; Zhao, Q.; Zhang, C.; Wang, G.; Yao, Y.; Huang, X.; Zhan, F.; Zhu, Y.; Shi, J.; Chen, J., miR-15b-5p resensitizes colon cancer cells to 5-fluorouracil by promoting apoptosis via the NF-κB/XIAP axis. Scientific reports 2017, 7, (1), 1-12.
- Gao, M.; Wang, X.; Zhang, X.; Ha, T.; Ma, H.; Liu, L.; Kalbfleisch, J. H.; Gao, X.; Kao, R. L.; Williams, D. L., Attenuation of cardiac dysfunction in polymicrobial sepsis by microRNA-146a is mediated via targeting of IRAK1 and TRAF6 expression. The Journal of Immunology 2015, 195, (2), 672-682.
- Taganov, K. D.; Boldin, M. P.; Chang, K.-J.; Baltimore, D., NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proceedings of the National Academy of Sciences 2006, 103, (33), 12481-12486.
- Zhang, Y.-G.; Song, Y.; Guo, X.-L.; Miao, R.-Y.; Fu, Y.-Q.; Miao, C.-F.; Zhang, C., Exosomes derived from oxLDL-stimulated macrophages induce neutrophil extracellular traps to drive atherosclerosis. Cell Cycle 2019, 18, (20), 2672-2682.
- Al-Banna, N.; Lehmann, C., Oxidized LDL and LOX-1 in experimental sepsis. Mediators of inflammation 2013, 2013.
- Kuhn, A. R.; Schlauch, K.; Lao, R.; Halayko, A. J.; Gerthoffer, W. T.; Singer, C. A., MicroRNA expression in human airway smooth muscle cells: role of miR-25 in regulation of airway smooth muscle phenotype. American journal of respiratory cell and molecular biology 2010, 42, (4), 506-513.
- Varga, Z. V.; Kupai, K.; Szűcs, G.; Gáspár, R.; Pálóczi, J.; Faragó, N.; Zvara, Á.; Puskás, L. G.; Rázga, Z.; Tiszlavicz, L., MicroRNA-25-dependent up-regulation of NADPH oxidase 4 (NOX4) mediates hypercholesterolemia-induced oxidative/nitrative stress and subsequent dysfunction in the heart. Journal of molecular and cellular cardiology 2013, 62, 111-121.
- Lv, Y.-n.; Ou-Yang, A.-J.; Fu, L.-s., MicroRNA-27a negatively modulates the inflammatory response in lipopolysaccharide-stimulated microglia by targeting TLR4 and IRAK4. Cellular and molecular neurobiology 2017, 37, (2), 195-210.
- Beneyto, L. A. P.; Luis, O. R.; Sánchez, C. S.; Simón, O. C.; Rentero, D. B.; Bayarri, V. M., Valor pronóstico de la interleucina 6 en la mortalidad de pacientes con sepsis. Medicina Clínica 2016, 147, (7), 281-286.
- Harbarth, S.; Holeckova, K.; Froidevaux, C.; Pittet, D.; Ricou, B.; Grau, G. E.; Vadas, L.; Pugin, J.; Network, G. S., Diagnostic value of procalcitonin, interleukin-6, and interleukin-8 in critically ill patients admitted with suspected sepsis. American journal of respiratory and critical care medicine 2001, 164, (3), 396-402.
- Song, J.; Park, D. W.; Moon, S.; Cho, H.-J.; Park, J. H.; Seok, H.; Choi, W. S., Diagnostic and prognostic value of interleukin-6, pentraxin 3, and procalcitonin levels among sepsis and septic shock patients: a prospective controlled study according to the Sepsis-3 definitions. BMC infectious diseases 2019, 19, (1), 1-11.
- Ethuin, F.; Delarche, C.; Gougerot-Pocidalo, M.-A.; Eurin, B.; Jacob, L.; Chollet-Martin, S., Regulation of interleukin 12 p40 and p70 production by blood and alveolar phagocytes during severe sepsis. Laboratory investigation 2003, 83, (9), 1353-1360.
- Iwashyna, T. J.; Ely, E. W.; Smith, D. M.; Langa, K. M., Long-term cognitive impairment and functional disability among survivors of severe sepsis. Jama 2010, 304, (16), 1787-1794.
- Thierry, C.; Amigorena, S.; Raposo, G.; Clayton, A., Isolation and characterization of exosomes from cell culture supernatants. Curr. Protoc. Cell Biol 2006, 3, 1-29.
- Mitchell, A. J.; Gray, W. D.; Hayek, S. S.; Ko, Y.-A.; Thomas, S.; Rooney, K.; Awad, M.; Roback, J. D.; Quyyumi, A.; Searles, C. D., Platelets confound the measurement of extracellular miRNA in archived plasma. Scientific reports 2016, 6, (1), 1-11.
- ThÚry, C.; Witwer, K.; Aikawa, E.; Alcaraz, M.; Anderson, J.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G., Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, (1), 1535750.
- Pundir, S.; Martin, M. J.; O'Donovan, C.; Consortium, U., UniProt tools. Current protocols in bioinformatics 2016, 53, (1), 1.29. 1-1.29. 15.
- Cox, J.; Mann, M., MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nature biotechnology 2008, 26, (12), 1367-1372.
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M. Y.; Geiger, T.; Mann, M.; Cox, J., The Perseus computational platform for comprehensive analysis of (prote) omics data. Nature methods 2016, 13, (9), 731-740.
- Thomas, P. D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L. P.; Mi, H., PANTHER: Making genome-scale phylogenetics accessible to all. Protein Science 2022, 31, (1), 8-22.
- Uhlén, M.; Fagerberg, L.; Hallström, B. M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A., Proteomics. Tissue-based map of the human proteome. Science (New York, NY) 2015, 347, (6220), 1260419-1260419.
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J., A novel and universal method for microRNA RT-qPCR data normalization. Genome biology 2009, 10, (6), 1-10.
- Livak, K. J.; Schmittgen, T. D., Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods 2001, 25, (4), 402-408.
Figure 1.
Plasma proinflammatory cytokine levels variation in the survivor and death groups. The results of phases A, B, C, D, E, and F of the survivors (blue line) and A and B of the death (red line) groups are indicated. Description of the phases: A. acute period; B. immediately after ICU discharge; C. 3 months post-ICU; D. 6 months post-ICU; E. 1-year post-ICU; and F. 3 years post-ICU. The interrupted line indicates the control data. The plasma IL-12p70 level was below the limit of detection in the death group. The row results are in Supplementary Material Tables S5 and S6.
Figure 1.
Plasma proinflammatory cytokine levels variation in the survivor and death groups. The results of phases A, B, C, D, E, and F of the survivors (blue line) and A and B of the death (red line) groups are indicated. Description of the phases: A. acute period; B. immediately after ICU discharge; C. 3 months post-ICU; D. 6 months post-ICU; E. 1-year post-ICU; and F. 3 years post-ICU. The interrupted line indicates the control data. The plasma IL-12p70 level was below the limit of detection in the death group. The row results are in Supplementary Material Tables S5 and S6.
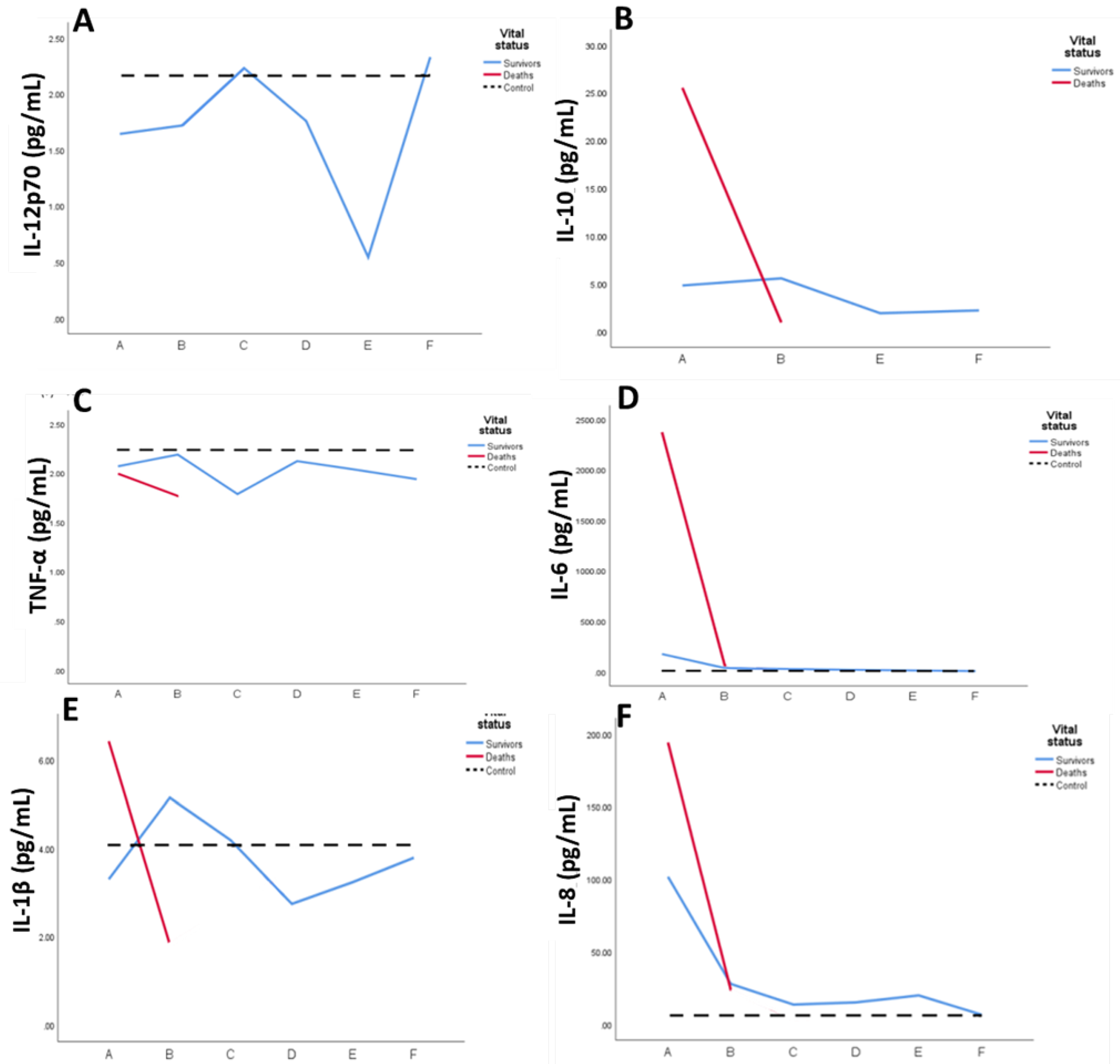
Figure 2.
Plasma EV DEmiR level variation in the death and survivor groups. The results of phases A, B, C, D, E, and F of the survivor (blue line) and A and B of the death (red line) groups are indicated. Description of the phases: A. acute period; B. immediately after ICU discharge; C. 3-month post-ICU; D. 6-month post-ICU; E. 1-year post-ICU; and F. 3-year post-ICU. The data of controls are indicated in the interrupted line. The row results are Supplementary Material Tables S11-S13.
Figure 2.
Plasma EV DEmiR level variation in the death and survivor groups. The results of phases A, B, C, D, E, and F of the survivor (blue line) and A and B of the death (red line) groups are indicated. Description of the phases: A. acute period; B. immediately after ICU discharge; C. 3-month post-ICU; D. 6-month post-ICU; E. 1-year post-ICU; and F. 3-year post-ICU. The data of controls are indicated in the interrupted line. The row results are Supplementary Material Tables S11-S13.
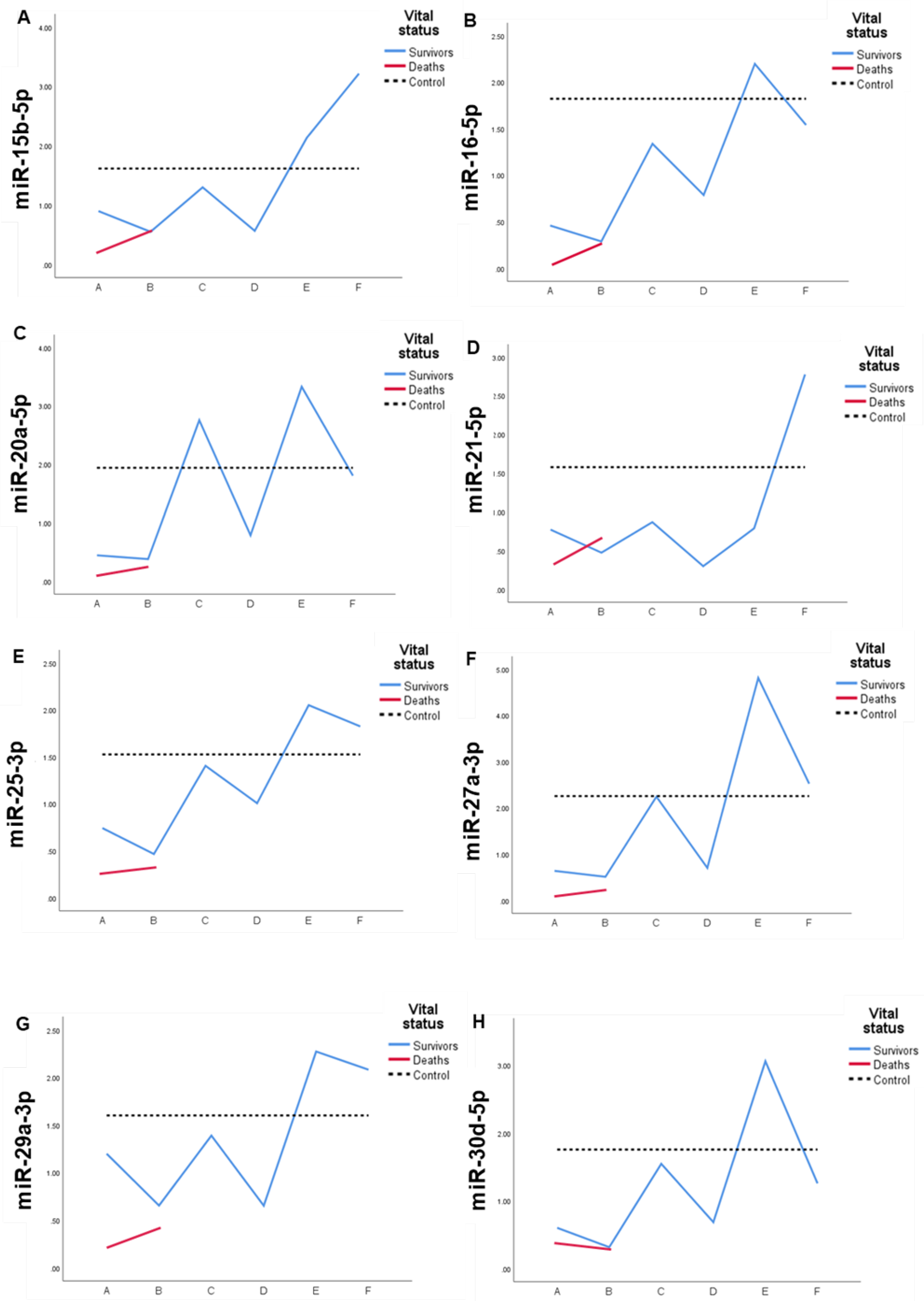
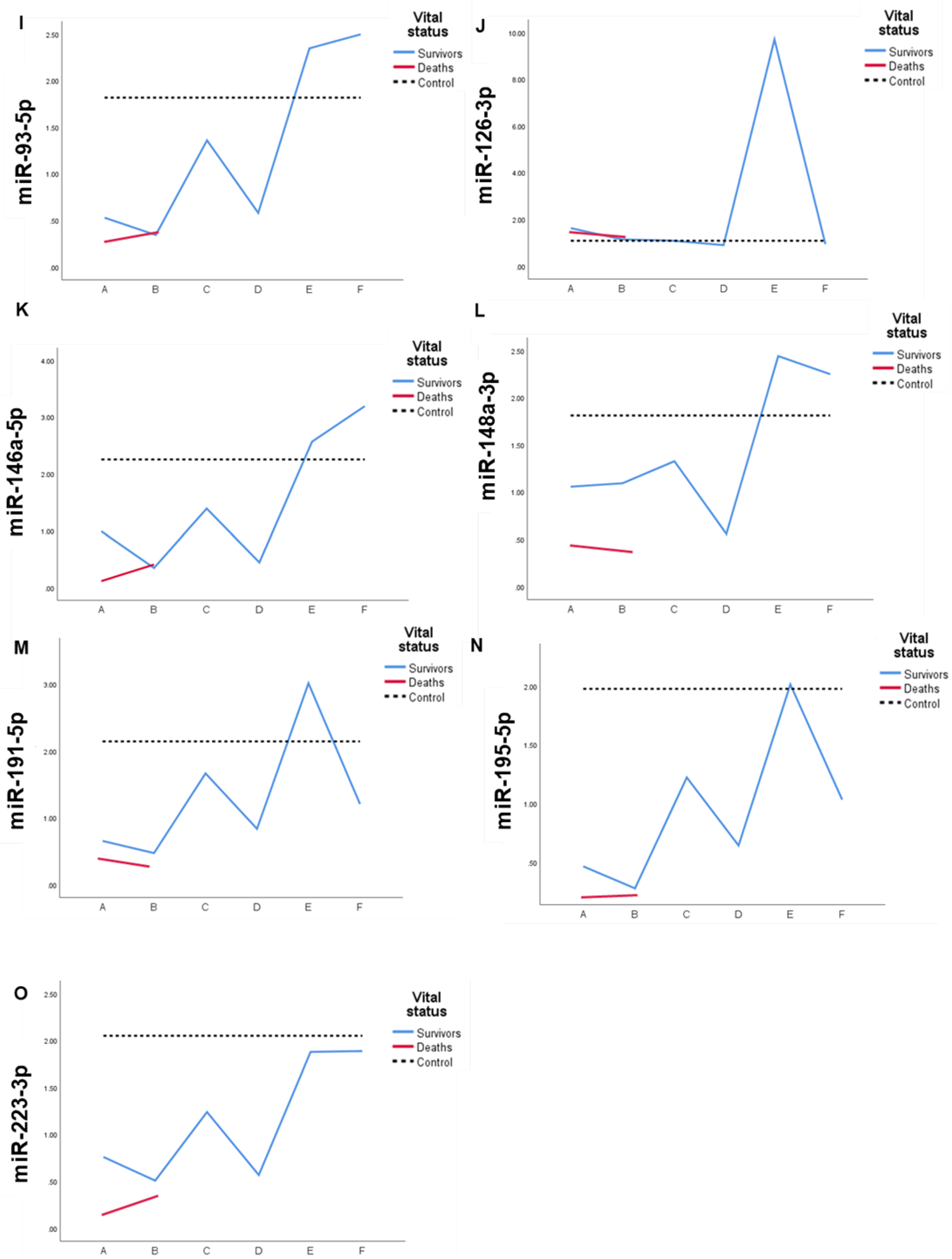
Figure 3.
Representation of EV miRNA levels variation from plasma of septic patients. The similarity in the expression profile by phase is indicated. The row results are in Supplementary Material Table S11.
Figure 3.
Representation of EV miRNA levels variation from plasma of septic patients. The similarity in the expression profile by phase is indicated. The row results are in Supplementary Material Table S11.
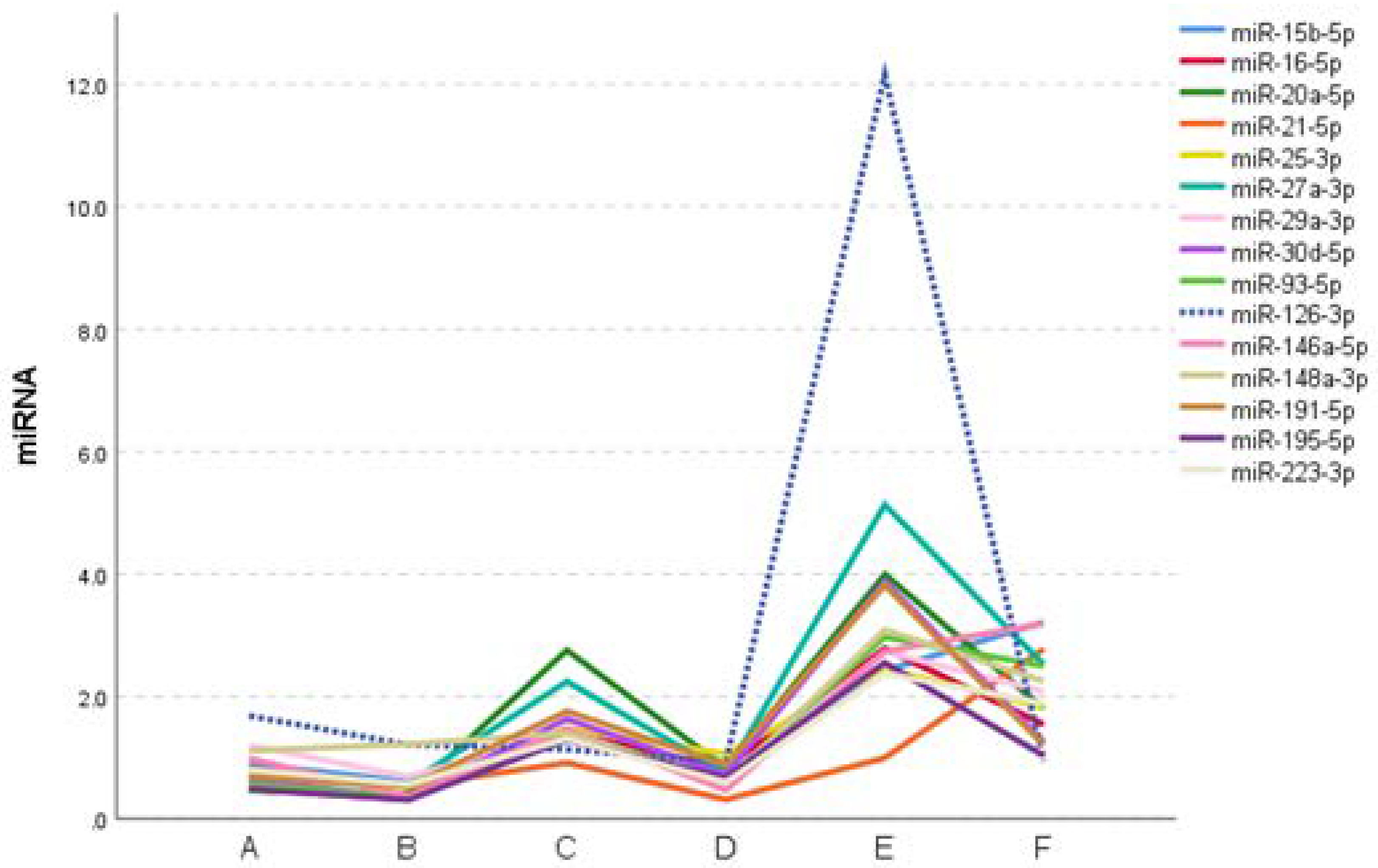

Table 1.
Clinical data of septic patients (n=36).
| number of patients (%) | |
| Gender | |
| Female | 13 (36.1) |
| Male | 23 (63.9) |
| Average (SD) | 60.2 (13.2) |
| Median (min-max) | 60.5 (34-87) |
| Comorbidities | |
| Arterial hypertension | 21 (58.3) |
| Diabetes mellitus | 14 (38.9) |
| Chronic obstructive pulmonary disease | 2 (5.6) |
| Heart disease | 9 (25.0) |
| Acute renal insufficiency | 9 (25.0) |
| Cirrhosis | 1 (2.8) |
| Congestive heart failure | 1 (2.8) |
| Chronic renal insufficiency | 2 (5.6) |
| Hyperthyroidism | 1 (2.8) |
| Dementia | 1 (2.8) |
| Chronic osteomyelitis | 1 (2.8) |
| Liver disease | 1 (2.8) |
| Sepsis focus | |
| Abdominal | 9 (25.0) |
| Leptospirosis | 1 (2.8) |
| Osteomyelitis | 1 (2.8) |
| Pancreatitis | 1 (2.8) |
| Soft parts | 1 (2.8) |
| Lungs | 12 (33.3) |
| Kidney | 6 (16.7) |
| Skin | 1 (2.8) |
| Another | 4 (11.1) |
| Vital status | |
| Survivors | 19 (52.8) |
| Deaths | 17 (47.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



