
Submitted:
01 January 2023
Posted:
06 January 2023
You are already at the latest version
Abstract
During Sep. 2011, forty-four Leymus chinensis culms with stroma development were collected from Inner Mongolia. Stromata surrounded the flag leaf sheathes, white, at the final stage of development. Perithecium but not asci were observed under microscope. Nine epichloid endophyte strains were obtained from Tongliao subdivision, and sixteen from Horqin. Morphological and phylogenetic properties of these strains were investigated. Morphological properties of these strains were similar among these 27 strains. Colonies could grow to 28.5~41.1 mm/21d at 25℃; phialides 24.1~26.4 μm long, 2.1~2.5 μm at base, tapering to less than 1.0 μm at tip; conidia reniform to elliptical, 4.7~5.5×2.8~3.5 μm, indicating typical properties of Epichloë endophytes. Based on the sequences of tefA and tubB gene fragments, strains isolated from Inner Mongolia formed a distinct subclade in E. bromolica/ E. yangzii clade, with a bootstrap value of 66% and 64% respectively. Considering morphological and phylogenetic properties, we tend to determine strains isolated from Leymus chinensis as a new taxon. The detailed taxonomic position of this taxon needs further investigations on hybridization or genomic sequence.
Keywords:
Leymus chinensis
; fungal endophyte
; stromata
; morphological properties
; phylogenetic properties
1. Introduction
Endophytic fungi spend part or the whole life cycle in plants, and form systemic and asymptomatic associations throughout the aerial parts of the host plant [1]. Epichloid grass endophytes (Ascomycota, Hypocreales, Clavicipitaceae) are novel microbial resources characterized by strict host specificity, and until now, were recorded on at least 9tribes of cold-season grasses, including some economic grasses distributed all over the world [2,3]. Previously, cold-season grass endophytic fungi were divided into sexual and asexual grass endophytes. The former belongs entirely to the genus Epichloë, while the latter imperfect fungi belong to the very heterogeneous Acremonium [4]. In 1996, grass endophytic fungi in the genus Acremonium were transferred to the newly established genus Neotyphodium [5], and subsequently the genus Neotyphodium was merged into the genus Epichloë in 2014, which is also endophytic fungus of the cold-season grass [6]. Nowadays, cold-season grasses endophytic fungi are all in the genus Epichloë, so they are also called “Epichloid endophytes”.
As a symbiosis with cold-season grasses, Epichloid endophytes improve the resistance of host plants to biotic and abiotic stresses [1,7]. They are usually found only in plants, and most can be dispersed vertically through seeds. A small number of members can also form stromata under the flag leaves during the plant’s flower stage, allowing natural hybridization and horizontal transmission [6]. Most of the original Neotyphodium members such as E. coenophialum, E. festucae var. lolii, E. aoteroae cannot form stromata on the plant surface and can only be vertically dispersed through seeds [1]; E. poae do not form ascomycetes and ascospores and can only be transmitted horizontally by conidia [3], while E. stromatolonga can form stromata without identifying sexual stage, and the horizontal transmissibility of conidia forming on the surface of stromata has not been confirmed [8]. On the other hand, the largely heterogeneous species E. typhina on orchard grass, timothy and some other plants has not been observed or isolated from seeds of their host plants [6].
Researches of endophytic fungi in China mainly focuses on cold-season grasses, especially Achnatherum spp., Leymus spp., and Elymus spp., which have attracted much attention in the northern grassland region [9,10,11]. In 2006, we reported the endophytic fungus Epichloë yangzii forming stromata on Roegneria spp. plants in China [12]. Due to its characteristics in phylogenetic analysis and its ability to show weak hybridization ability in artificial hybridization experiments with E. brimicola from several European Bromus spp. [13], Leuchtmann et al. suggested to combining the two groups with the name Epichloë brimicola [6]. But the taxonomy of this fungal endophyte still remains a controversy.
Leymus chinensis is tolerant against drought, cold, alkali and trample on by livestock, as well as has a high grass yield, good palatability, rich protein content, and wide adaptability. Thus, it’s not only an important grass forage in China, but also a widely used substance in artificial grassland construction and ecological construction [14,15]. The endophyte in L. chinensis was firstly detected by Nan Zhibiao from the seeds in Qinghai in 1996 [16]. In 2013, the plant ecology research team of Nankai University reported the asymptomatic endophytic fungi obtained from L. chinensis grown in eastern Inner Mongolia. They concluded that those fungal isolates were close to the E. yangzii we found from Roegneria spp. through mycological and molecular systematic properties. They identified these strains as Choke disease endophyte Epichloë bromicola, according to Leuchtmann et al. [11]. However, they did not find or make any symptom on host plants neither in natural grassland, nor under artificially controlled conditions [6]. Therefore, we believe that the identification of the symptomatic endophytic fungus obtained from Leymus chinensis in eastern Mongolia is still a new task.
During field investigations in Liaoning, Hebei, Inner Mongolia and Shanxi from 2010 to 2011, we collected stromata on Leymus plants in Inner Mongolia. Fungal strains were isolated. Their morphological and phylogenetic properties were analyzed, their taxonomic position was discussed and revealed different insights from previous study.
2. Materials and Methods
2.1. Collection and storage of plant samples
On early September 2011, endophytic fungal resources were investigated in southeast Inner Mongolia. In order to minimize damage to vegetation, only one sample was collected among a 4 m2 area, and only one tiller from a plant if not necessary; when the plants were rare, only one plant was collected within about 30~50 m2, and the others were only recorded and not collected. Generally, 5~15 cm culm with inflorescence was cut into an envelope; When the plant does not form a spike, only 2 leaf parts were selected; When it was absolutely necessary, aerial parts but not root of tiny number of plants were collected.
Plants with stromata were collected from Tongliao district and Horqin Shadi. The aerial part of the plant was taken and brought back. Species of the grass with stromata was identified as L. chinensis by morphological properties, microscopic characteristics of leaf epidermis as described by Zhang et al. elsewhere [17]. Samples were brought back to the lab for immediate processing or preserved at 4°C for further use.
2.2. Observation of and isolation of stromata
Fresh stromata were sliced off, crushed in water. Microtome sections were cut by hand to observe the morphological characteristics of stromata.
The culms bearing stroma were cut into 5~10 mm long segments and disinfected by double treatments of 75% alcohol and NaOCl (1% available Cl). After rinsing in sterile water the samples were placed on potato-dextrose agar (PDA) and incubated at ca. 25 C in dark [18].。
2.3. Morphological characteristics of the endophytic fungus
Growth rate, shape and colour of colonies on the same plates were determined on PDA plates after 3 wk incubation. Mycological properties of hyphae, conidia and phialides were observed and recorded respectively under the light microscope with 20 repeats.
2.4. Molecular phylogenetic analysis
Basing on their morphology, representative strains of Ltla101, Ltla103, Ltla104 (Tongliao district) and Ltla201, Ltla204, Ltla205 (Horqin Shadi) were selected to extracted genomic DNA from fresh mycelium by IQS method [18].
Selective primers were designed by different base pairs of tubB and tefA (Table 1). Primers were synthesized by Shanghai Generay Biotech Co. Ltd. The PCR products were sequenced at Shanghai Majorbio Bio-Pharm Technology Co., Ltd.
The resulting sequence was blasted in NCBI and confirmed to be correct. The corresponding sequences of representative Epichloë species were selected, download and organized with TBtools [19]. Alignments, model testing and then Maximum Likelihood (ML) analyses were performed by MEGA X [20].

Table 1.
PCR primers used for amplifying of tefA and tubB fragments.
| Gene | Primer | Sequence of Primer(5’- 3’ ) | Reference |
| tefA | tef1-exon1d | GGG TAA GGA CGA AAA GAC TCA | [21] |
| tef1-exon6u | CGG CAG CGA TAA TCA GGA TAG | [21] | |
| KF-3D | TCG AGA AGG TAA GAC ATT CTT C | [22] | |
| KF-6D | GTT CGA GAA GGT GAG ATA TAT C | [23] | |
| tubB | tub2-exon1d | GAG AAA ATG CGT GAG ATT GT | [21] |
| tub2-exon4u | TGG TCA ACC AGC TCA GCA CC | [21] | |
| ZB-1D | GTT TCG TCC GAG TTC TCG ACA | This study | |
| ZB-1U | GAG AAA ATG CGT GAG ATT GTA AGT T | This study |
3. Results
3.1. Sampling and morphological characteristics of the stromata
On September 2011, we collected a total of 2252 grasses from Inner Mongolia. Most species were few (<10%) while 99% of Achnatherum sibiricum were infected by endophytes. Forty -four samples of Tongliao district and Horqin Shadi were collected. This study mainly focused on these samples.
Stromata surrounded the flag leaf sheathes, white, at the final stage of development, suppressed the development of host inflorescence (Figure 1A). They even collapsed from the sheath. Stromata usually developed on the leaf sheath by encompassing a part of it (bottom parts of leaf blade were involved) thus inhibiting plants heading. The asymptomatic L. chinensis plants nearby had their flowers fall down, no fresh inflorescence was observed.
The mycelia layer of stromata was thick, loose and homogenous under microscope (Figure 1B). In microtome sections, perithecia buried deep inside stromata without aperture. No ascus was observed within perithecium.
3.2. Isolation, cultivation and morphological characteristics of the endophytic fungus.
Typical, white and minute colonies were obtained from PDA plates after 1 wk at 25 °C from culms. After three repeats of single conidium isolation we obtained 9 fungal isolates from Tongliao district and 16 from Horqin Shadi. Purified strains were inoculated in PDA medium, grown for 21 d, and stored at 4 °C.
A: Aging stromata on Leymus chinensis; B: Cross section of the stromata; C, D: Colony properties (PDA, 21d, 25C), E: conidia and phialides
The morphological characteristics of the isolated strains from Tongliao district and Horqin Shadi were similar on PDA medium. Colonies were white, cottony, centrally raised, circular with a regular margin (Figure 1C). Colony reverses pale brown and lighter toward the margins (Figure 1D).
Colony diam 28.5~41.1 mm after 21 d at 25 ℃. aerial mycelium smooth, septated and ca. 2 μm wide. Sporulation in culture abundant. Phialides discrete, arising solitary from the aerial mycelium, septated at the base, hyaline, smooth, 24.1~26.4 μm long, 2.1~2.5 μm at base, tapering to less than 1.0 μm at tip. Conidia hyaline, lunate to reniform, 4.7~5.5×2.8~3.5 μm (Figure 1E). Characteristics in cultivation and morphology of these strains were identical to those of epichloë endophytes (Table 2).
Morphology of strains in this study were conducted for 25 repeats; Information and data on other strains were modified from Kang et al. 2011 [22].
3.3. Phylogenetic analyses of fungal isolates
DNA fragments of tefA, tubB genes from 6 strains obtained from L. chinensis were amplified by primers (Table 1) and sequenced. Single copies of 790 bp and 870 bp sequences with each of 3 base differences were obtained respectively. DNA sequences of tefA or tubB fragments were 99.83% and 99.85% identical.
The resulting sequences were blasted in NCBI to confirm the identify of epichloid endophytic fungi. According to the blast results, sequences of representative species were selected to construct a phylogenetic tree. Based on the tefA and tubB fragments, the Kimura 2-parameter model was used to construct ML (maximum likelihood) tree. ML trees based on tefA and tubB gene sequences were similar: most members of a particular Epichloë species tended to cluster together with a high bootstrap value, in accordance with previous reports. These strains tended to separate into Epichloë typhina complex (ETC) and E. bromicola/E. yangzii (EBY) clades. The strains of L. chinensis in Inner Mongolia formed independent cluster in the EBY clade with the bootstrap values of 99% and 97%, respectively. These suggest that these strains form a separate taxonomic unit among epichloid endophytes (Figure 2).
3.4. Taxonomy and origin of isolated strains
Basing on morphological and phylogenetic analysis, all the strains obtained from L. chinensis should be an independent taxon (Table 2, Figure 2).
This taxon has a similar genetic origin with E. sinifestucae from Festuca parvigluma(syn. N. sinofestucae), E. bromicola(syn. E. yangzii) from R. kamoji and E. sinica(syn. N. sinicum)in our country (Figure 2). Similarly, E. bromicola from Bromus erectus in European, E. siegellii(syn. N. siegellii) from meadow fescue (Festuca pratense) in European., and E. chisosa(syn. N. chisocum)from Stipa eminence in the North American, are also associated in its origin (Figure 2).
4. Discussion
Leymus spp. are important forage grasses in northern China, and also widely distributed in northeast, northwest and north China as ground-cover plants. Particularly, they are also important plants compatible with wheat (Triticum aestivum L.) in artificial hybridization tests. Therefore, we focused on the survey of seed transmitted fungal endophyte within these plants. Stomata bearing plants of Tongliao district and Horqin Shadi were collected and isolated 9 and 16 strains, respectively. These strains had similar morphological and phylogenetic characteristics, with slightly faster growth rates than common members of Epichloë spp.. The characteristics of conidia and phialides were typical of epichloid endophytes (Figure 1, Table 2). Combined with phylogenetic analysis, these isolated strains are typical epichloid endophytic fungi. This is the first report that typical symptomatic epichloid endophytes have been isolated from wild L. chinensis (Figure 1A) grown in natural grasslands and sandy lands.
L. chinensis (Triticeae) is rarely recorded to be infected by endophytes [16,24]. In 1996, Nan et al. firstly found 12% L. chinensis seeds from Qinhai were infected, confirming the distribution of endophytic fungi in grass L. chinensis grown in Northwest China[16]. In 2006, Gao et al. investigated the endophyte resources of grasses in Inner Mongolia, and also found endophyte in L. chinensis [24]. However, mycological properties were not tested in the early stage. In 2013, Gao et al. carefully investigated L. chinensis in a large area from central to northeastern Inner Mongolia, and reported three morphotypes of endophytes from asymptomatic L. chinensis plants. But none of them was reported to be a new taxon [11]. On September 2011, we surveyed endophyte resource in Inner Mongolia, and collected a total of 2252 grasses. Most species were lower than 10% in infection level while 99% of A. sibiricum were infected, which was coincidence with previous study [16,24].
Tongliao district and Horqin Shadi are located between 43° and 44° N in southeastern Inner Mongolia, and belong to the arid and semi-arid region of the middle temperate zone. L. chinensis usually begin flower and fruit on June, the followers wither away on August, in Tongliao, southeast area of Inner Mongolia. On the sampling date of September, the leaf sheath only surrounded with white and collapsed stromata, the stromata were at the final stage (Figure 1A). Observing the cross section of the stromata, perithecia with no aperture was observed under microscope (Figure 1B), and no ascus was observed. Therefore, we speculate that it is possible that the ascus has been ejected from the perithecium and the perithecium collapses, or that the ascus is not developed due to unfertile stromata. We obtained endophytic fungi E. stromatolonga Ji et al. that can form stromata but no mature stromata from Calamagrostis epigeios grown from Nanjing and Huangshan in eastern China [8], and whether this is the another case or not, this stromata needs to be further investigated.
In this study, 25 endophytic fungi strains were isolated from Tongliao district and Horqin Shadi in Inner Mongolia. Colony morphology, growth rate, size and shape of phialides and conidia, were typical of epicholid endophytes, but these strains exhibited difference from previous reported species (Table 2) [2]. The strains from Tongliao district and Horqin Shadi shared nearly similar identity in tefA and tubB fragment. Their sequences diversity between strains was far less than that of their previous relatives [11,16,24]. The two sampling sites of this study are actually distributed in the Horqin region. Nankai research group surveyed a wide range of sampling sites, which can better reflect the fact that L. chinensis and its endophytic fungi in the central and eastern regions of Inner Mongolia. The endophytic fungal strains of L. chinensis exhibit richer genetic diversity, together with the long-term distribution and long-term evolution of L. chinensis in eastern Inner Mongolia. [16,24]。
In the phylogenetic analysis based on tefA and tubB fragments, each species formed small independent branches with high bootstrap values, consistent with previous reports [11]. All the strains of Inner Mongolia L. chinensis formed an independent branche on the EBY clade, with bootstrap values of 99% and 97% for tefA and tubB respectively, showing significant differences from other strains (Figure 2). Due to the absence of stromata morphology and live plants, we could not conduct artificial hybridization experiments to determine the strains of L. chinensis. The relationships between these strains and strains of Roegneria spp. and F. parvigluma which were most closely related in systematics were not settled down. Therefore, the taxonomic status of the L. chinensis strain was not conclusively determined in this study. Strains of E. sinica (syn. N. sinicum), E. sinofestucae (syn. N. sinofestucae) species are interspecific hybrids [16], but not for L. chinensis strains. Combing morphological and phylogenetical properties, L. chinensis strains could be a new taxon apart from E. bromicola (syn. E. yangzii) and E. sinica (syn. N. sinicum).
Nankai group investigated fugal diversity in middle and north Inner Mongolia steppe. In 2013, 96 fungal isolates of 3 morphotype (MT) were obtained from of L. chinensis. They believed that MT1 was similar to our E. yangzii strains obtained from Roegneria spp., and proposed them as Epichloë bromicola [11]. This is based on the academic view of Leuchtmann’s team in Europe, which is different from ours.
At the 9th (2010) and 10th (2012) the International Symposium on Fungal Endophytes of Grasses, Leuchtmann and we discussed the taxonomic relationship between E. bromicola and E. yangzii, but failed to gain a consistent insight. Leuchtmann et al. adhered to the most basic European tradition of “being able to hybridize is the same species, and vice versa”, and used this as the only criteria to classify fungi [13]. We believe that reproductive isolation is indeed important and agree that the individuals with reproductive isolation are different species, but we believe that the individuals that can be artificially crossed are not necessarily the same species. Allopatric isolation is another “natural isolation” in addition to reproductive isolation, which can cause the separated population to form their own evolutionary process. Moreover, artificial hybridization is not natural hybridization, and strains that can be artificially hybridized in the laboratory may not meet in nature. Especially for plant endophytes, variations in host species and stromata formation times can also constitute allopatric isolation. It is conceivable that successful hybridization can be mated when mating with artificial cultures, but in nature they cannot be crossed due to the time and place of stromata formation. In other words, two strains with a certain hybridization compatibility may not have the opportunity to hybridize under natural conditions. In fact, the affinity between E. bromicola isolated from Bromus erectus in Europe, and E. yangzii isolated from Roegneria spp. in the Far East of Asia, is not strong. Hybridization between the two strains, even under artificial conditions, was not frequent, and only a small number of perithecia, even unfertilized were formed in our repeated inoculating experiments (Wang Zhiwei and Li Wei, 2005, unpublished data). There are 77 species of Bromus plants in China, but distribution of Bromus erectus is rare [25]. Also, Bromus spp. (tribe Bromeae) have a slightly distant plant taxonomic relationship with Roegneria spp. (tribe Triticeae) in the family Poaceae. Although Roegneria spp. are widely distributed in humid areas, the flowering stage varies with region, and it may overlap with Bromus erectus in time. However, the Far East and Europe are separated by thousands of mountains and rivers, and the two places are separated by plateaus, Gobi, especially arid areas with altitudes of more than 4,000 m such as the Himalayas, Kunlun Mountains and Tianshan Mountains. It is really hard that Bromus erectus and Roegneria spp. endophytic fungi isolated from the Far East of Asia are able to communicate. Roegneria is a large genus with a lot of members distributed in the eastern end of Eurasia (north of South Ridge in China, the Korean Peninsula, the Japanese Islands, etc.), while there are few at the western end of Eurasia. The European taxonomic system merges the Roegneria genus into the genus Agropyron. This physical barrier also has a certain impact on the distribution of Roegneria spp. Thus, geographical barriers also play an important role in the distribution and formation of species. Therefore, in terms of fungal and plant classification, the ability to hybridize between strains and plants is not the only criterion for species judgment. Other isolations, such as temporal isolation and geographical isolation, are also the driving forces that in fact promote the evolution of species.
On the contrary, with the continuous discovery of various strains and classification, there are more and more taxonomic groups, and hybridization between strains is becoming more and more unrealistic. This will undoubtedly hinder the development of fungal endophyte research. Conversely, if strains from various plants are grouped together as a result of artificial hybridization, as advocated by Prof. Leuchtmann, the generalized E. bromicola [6] species will contain more and more members, and its heterogeneity will increase, which will eventually lag the discovery on this species. The classification of organisms is originally artificial classification, which is relatively based on subjective factors, so there is more factors for discussion. The reason why people want to talk about the classification of various organisms is to promote the better development of research in this field. One species with high heterogeneity among the endophytic is Epichloë typhina (syn. N. typhinum). This is a relatively classic species that was recorded and studied earlier, but compared with other endophytic fungi, the biological characteristics of the strains within this species are very different, and the molecular systematics characteristics are also relatively complex. E. typhina, which is known as ETC (Epichloë typhina-complex, or Epichloë typhina-clade) in phylogenetic analysis, is lagged behind in research. In fact, there are very few researchers concerning about E. typhina. Whether this is related to the excessive heterogeneity of E. typhina is worth speculating. Therefore, in order to promote the research of grass endophytes, we still advocate that factors such as spatiotemporal isolation and host isolation should also be included in the classification. Moreover, in addition to spatiotemporal isolation and host species, there are many differences between E. bromicola, E. yangzii and E. sinica (unpublished data). Therefore, we tend to not classify these L. chinensis strains as E. bromicola.
Epichloë spp. have a smaller distribution than the previous asexual Neotyphodium spp. [2]. In addition to the genotype of fungi, whether or not to form stromata on host plants is also affected by environmental conditions [1]. Therefore, the formation of stromata of Epichloë on host plants may not occur every year. Most grasses endophytic fungi are host-specific, and one plant corresponds to one endophytic fungus [2]. Endophytic fungi can form stromata on L. chinensis, indicating that in the long-term evolutionary process, L. chinensis and endophytic fungi formed new associations.
Typical Epichloë species could develop stromata around plant sheath/culm and is considered to be the sexual stage of the pre-Neotyphodium spp.. Until now, five stromata bearing species have been reported in China, E. yangzii in Roegneria spp.[12], E. stromatolonga in Calamagrostis epigeios[8], E. sibirica in Achnatherum sibiricum [26], E. sylvatica in Brachypodium sylvaticum [27], E. liyangensis in Poa pratensis ssp. pratensis [22]. E. yangzii was considered to be hybrid ancentor of E. sincia [23]. In addition to the middle and lower basin of the Yangtze River, our research group also collected Roegneria spp. plants from Shandong, Liaoning and other regions. However, the distribution range of Calamagrostis epigeios and Poa pratensis ssp. pratensis that can form stromata is relatively limit [8,22]. In our 20 years’ investigation of grasses endophytes resources of in more than 10 provinces, municipalities and autonomous regions in East China and North China, only a small number of stromata bearing L. chinensis in Tongliao District and Horqin Shadi were collected. L. chinensis is widely distributed in northeast, northwest and north of China, and the investigation of endophytes requires further research. In this study, 44 stromata bearing L. chinensis samples of Tongliao District and Horqin Shadi in Inner Mongolia were collected. It was the first report of stromata on wild L. chinensis plants, and it was also the sixth grass with stromata reported in China.
Until now, no records about toxic symptoms caused by L. chinensis had been documented, indicating neither ergot nor indole-diterpenoids alkaloids were estimated within this association. L. chinensis has the advantages of drought tolerance, cold resistance, alkali resistance, and resistance to trampling by cattle and horses [14,15], and the plants that bearing stromata tend to have strong vegetative growth, stems, leaves and root systems. For a long time, L. chinensis has been a dominant grass species in northern China, and it plays a pivotal role in the vegetation in arid and semi-arid areas. It is worth considering whether the discovery of L. chinensis with stromata in Tongliao district and the Horqin Shadi is related to the windy and dry environment and its surroundings. Therefore, whether the endophytic fungus of L. chinensis is a new type of microbial resource is expected to be further studied.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Sampling, Yong Wang and Tao Lu; methodology, Baohu Liu; software, Yanling Ji; data curation, X.X.; writing—original draft preparation, Hongxia Zhang; writing—Yanling Ji, funding acquisition, Zhiwei Wang. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by National Science Foundation of China, grant number 30800156, 30970081, 31372365” and “The APC was funded by National Science Foundation of China”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annual Review of Plant Biology 2004, 55, 315-340. [CrossRef]
- Wang Z; Ji Y; Chen Y; Kang Y. Endophytic fungi resources and species diversity in grass family. Acta Ecologia Sinica 2010, 30, 4771-4781.
- Tadych, M.; Bergen, M.S.; White, J.F., Jr. Epichloë spp. associated with grasses: new insights on life cycles, dissemination and evolution. Mycologia 2014, 106, 181-201. [CrossRef]
- Morgan-Jones, G.; Gams, W. Notes on hyphpmycetes. XLI. An endophyte of Festuca arundinacea and the anamorph of Epichloë typhina, new taxa in one of the two new sections of Acremonium. Mycotaxon 1982, 15, 311-318.
- Glenn, A.E.; Bacon, C.W.; Price, R.; Hanlin, R.T. Molecular phylogeny of Acremonium and its taxonomic implications. Mycologia 1996, 88, 369-383. [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F.; Tadych, M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202-215. [CrossRef]
- Wang Z; Ji Y; Chen Y. Grass endophytes and their potential applications in agriculture. Journal of Nanjing Agricultural University 2011, 34, 144-154.
- Ji, Y.; Zhan, L.; Kang, Y.; Sun, X.; Yu, H.; Wang, Z. A new stromata-producing Neotyphodium species symbiotic with clonal grass Calamagrostis epigeios (L.) Roth. grown in China Mycologia 2009, 101, 200-205. [CrossRef]
- Li, B.; Zheng, X.; Shahe, D.; Sun, S. An investigation of endophyte-grasses in north-west of China. Grassland of China 1996, 2, 29-32.
- Li, C.; Nan, Z.; Paul, V.H.; Dapprich, P.D.; Liu, Y. A new Neotyphodium species symbiotic with drunken horse grass (Achnatherum inebrians) in China. Mycotaxon 2004, 90, 141-147.
- Zhu, M.; Ren, A.; Wen, W.; Gao, Y. Diversity and taxonomy of endophytes from Leymus chinensis in the Inner Mongolia steppe of China. FEMS Microbiol Lett 2013, 340, 135-145. [CrossRef]
- Li, W.; Ji, Y.; Yu, H.; Wang, Z. A new species of Epichloë symbiotic with Chinese grasses. Mycologia 2006, 98, 560-570. [CrossRef]
- Lembicz, M.; Górzyńska, K.; Leuchtmann, A. Choke disease caused by Epichloë bromicola in the grass Agropyron repens in Poland. Plant Disease 2010, 94, 1372. [CrossRef]
- Gao, Y.; Wang, D.; Ba, L.; Bai, Y.; Liu, B. Interactions between herbivory and resource availability on grazing tolerance of Leymus chinensis. Environmental and Experimental Botany 2008, 63, 113-122. [CrossRef]
- Wang Y; He X; Zhou G. Study on the responses of Leymus chinensis steppe to grazing in Songnen Plain. Acta Agrestia Sinica 2002, 45-49.
- Nan Z. lncidence and distribution of endophytic fungi in seeds of some native and introduced grasses in China. Acta Prataculturae Sinica 1996, 1-8.
- Zhang H; Ji Y; Lu T; Wang Y; Liu B; Wang Z. Identification of grass without inflorescence collected from Korqin, Inner Mongolia. Journal of Nanjing Agricultural University 2014, 37, 60-67.
- Wang Y; Ji Y; Wang H; Chen Y; Wang Z. Grass endophytic fungi researches 13:Endophytic fungi identification and rapid extraction of genomic DNA. Journal of Nanjing Agricultural University 2012, 35, 52-56.
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Molecular Plant 2020, 13, 1194-1202. [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547-1549. [CrossRef]
- Gentile, A.; Rossi, M.S.; Cabral, D.; Craven, K.D.; Schardl, C.L. Origin, divergence, and phylogeny of epichloe endophytes of native Argentine grasses. Molecular phylogenetics and evolution 2005, 35, 196-208. [CrossRef]
- Kang, Y.; Ji, Y.L.; Zhu, K.R.; Wang, H.; Miao, H.M.; Wang, Z.W. A new Epichloë species with interspecific hybrid origins from Poa pratensis ssp. pratensis in Liyang, China Mycologia 2011, 103, 1341-1350. [CrossRef]
- Kang, Y.; Ji, Y.L.; Sun, X.H.; Zhan, L.H.; Li, W.; Yu, H.S.; Wang, Z.W. Taxonomy of Neotyphodium endophytes of Chinese native Roegneria plants. Mycologia 2009, 101, 211-219. [CrossRef]
- Wei, Y.K.; Gao, Y.B.; Xu, H.; Su, D.; Zhang, X.; Wang, Y.H.; Lin, F.; Chen, L.; Nie, L.Y.; Ren, A.Z. Occurrence of endophytes in grasses native to northern China. Grass and Forage Science 2006, 61, 422-429. [CrossRef]
- Liu, l. Flora reipublicae popularis sinicae; Science Press: Beijing, 2002; Volume 9.
- Wei, Y.; Gao, Y.; Zhang, X.; Su, D.; Wang, Y.; Xu, H.; Lin, F.; Ren, A.; Chen, L.; Nie, L. Distribution and diversity of Epichloë/Neotyphodium fungal endophytes from different populations of Achnatherum sibiricum (Poaceae) in the Inner Mongolia Steppe, China. Fungal Diversity 2007, 24, 329-345.
- Ji Y; Sun X; Wang Z. A survey of the gramineous plant endophytes in Huangshan Geopark. Journal of Nanjing Agricultural University 2011, 34, 147-150.
Figure 1.
Morphological properties of host and fungal endophyte in Leymus chinensis.
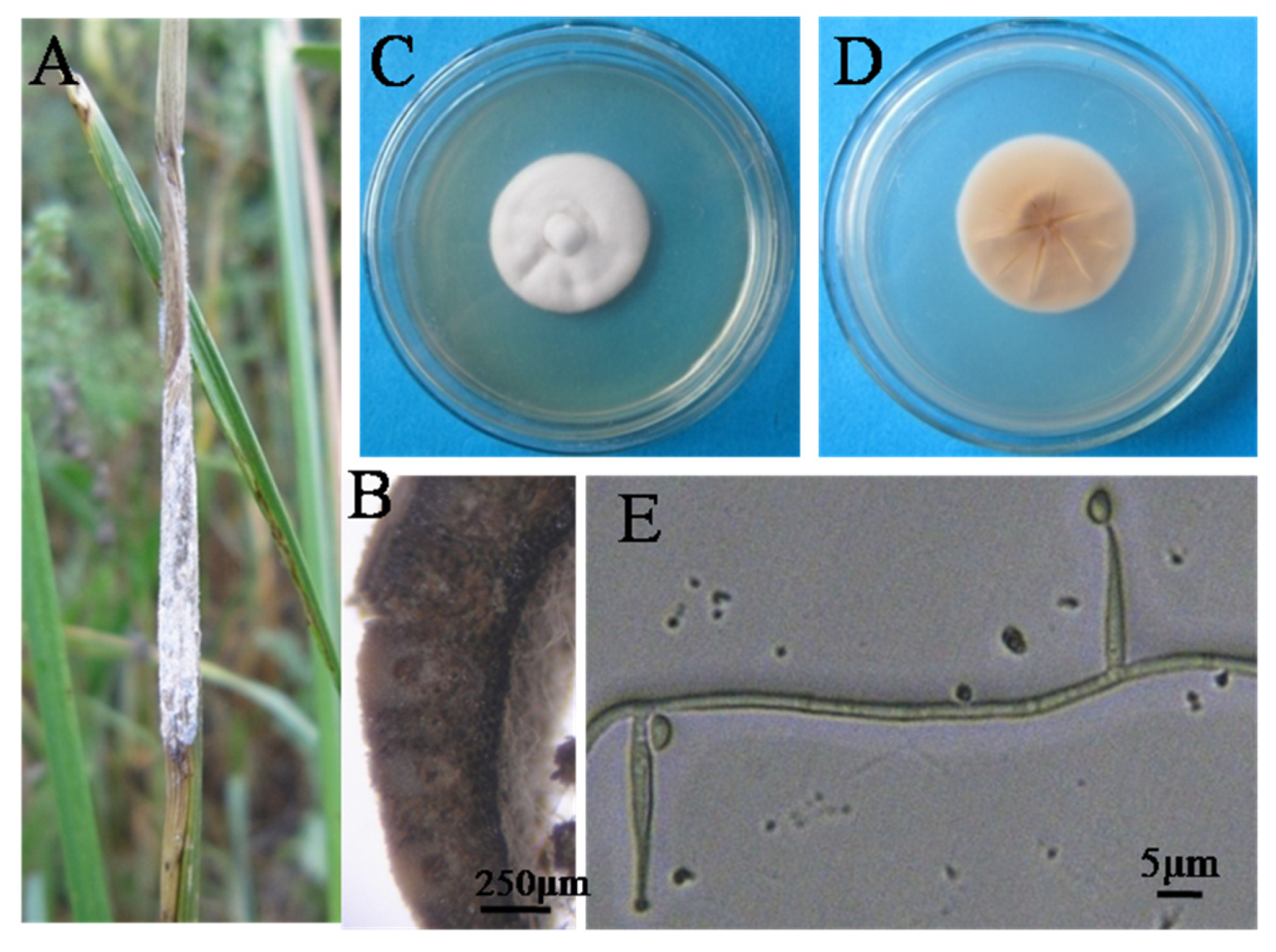
Figure 2.
Epichloë spp. tefA and tubB phylograms based on Maximum Likelihood (ML). Numbers (>60%) at branches are the percentage of trees containing the corresponding clade based on 1000 bootstrap replications. ▲: isolates in Nankai University; Circles represent isolates in this study.
Figure 2.
Epichloë spp. tefA and tubB phylograms based on Maximum Likelihood (ML). Numbers (>60%) at branches are the percentage of trees containing the corresponding clade based on 1000 bootstrap replications. ▲: isolates in Nankai University; Circles represent isolates in this study.
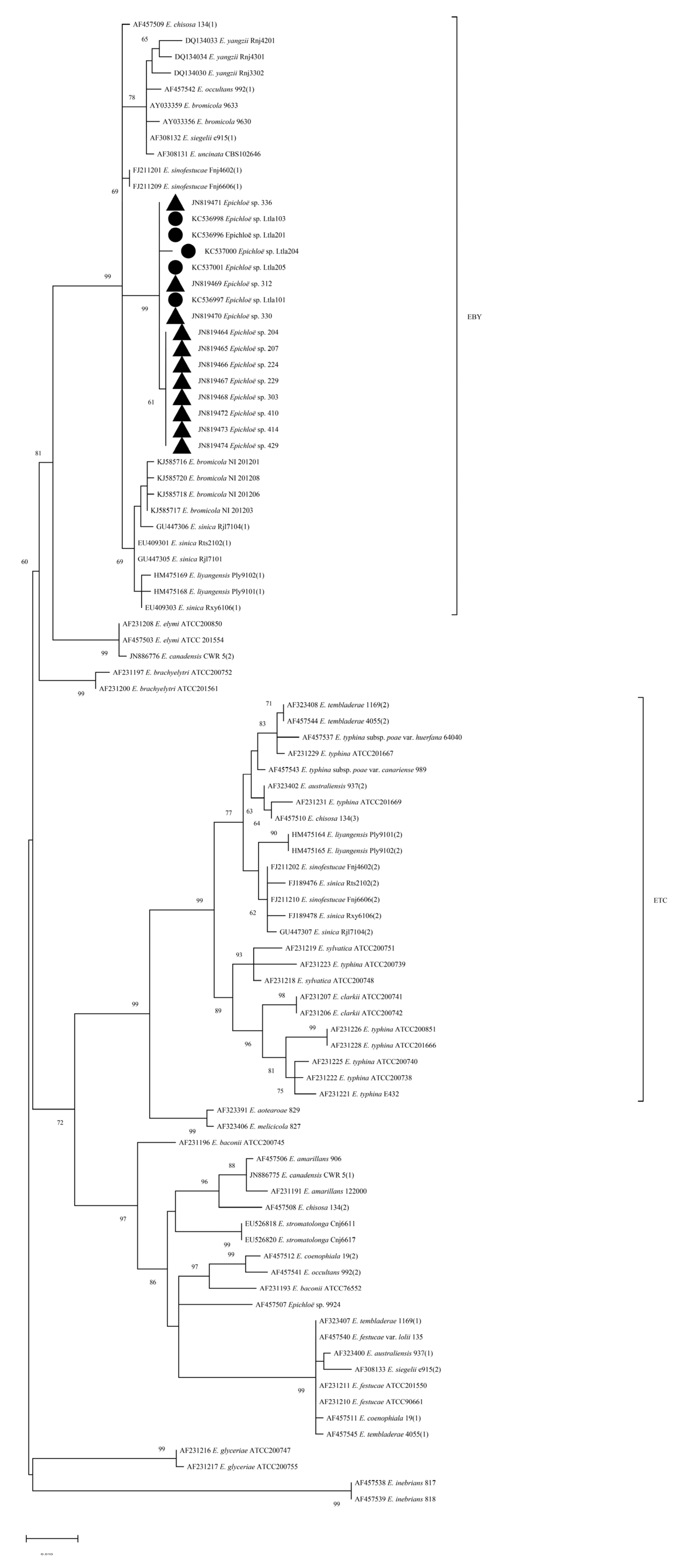
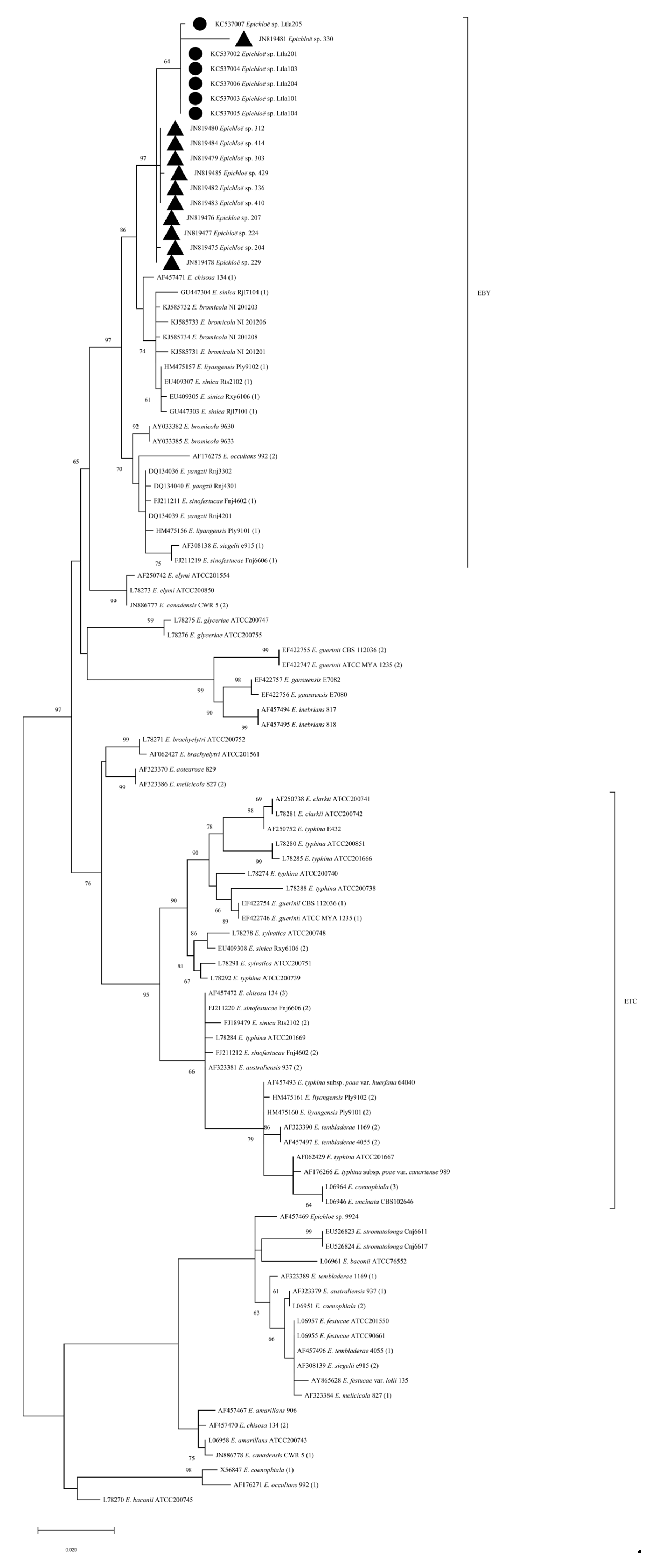
Table 2.
Table 2. Morphological comparison of Leymus chinensis isolates and some epichloid endophytes.
Table 2.
Table 2. Morphological comparison of Leymus chinensis isolates and some epichloid endophytes.
| Fungal Isolate | Host | Geographic origin | Conidium | Phialide | Growth rate(mm/week) | |||
|---|---|---|---|---|---|---|---|---|
| Length(μm) | Width(μm) | Length(μm) | Base(μm) | Tip(μm) | ||||
| E. sibirica | Achnatherum sibiricum | China | 3.0-4.6 | 1.7-2.2 | 24.1-40.5 | 1.6-2.2 | 0.8-1.0 | 2.3-3.0 |
| E. stromatolonga | Calamagrostis epigeios | China | 4.4-5.3 | 2.8-3.8 | 16.5-25.8 | 2.2-2.9 | <1.0 | 10.4-13.4 |
| E. typhina | Dactylis glomerata | Switzerland | - | - | - | - | - | - |
| E. sylvatica | Brachypodium sylvaticum | Switzerland | 5.0±1.1 | 2.0±0.5 | 15.0-33.0 | 1.0-1.7 | 0.5 | 8.3-15.0 |
| E. baconii | Agrostis capillaris | England | - | - | - | - | - | - |
| E. clarkii | Holus lanatus | England | - | - | - | - | - | - |
| E. amarillans | Agrostis hiemalis | N. America | - | - | - | - | - | - |
| E. festucae | Festuca spp. | Switzerland | 4.7±0.6 | 2.2±0.3 | 12.0-25.0 | 1.5-2 | <1 | 7.0-18.7 |
| E. glyceriae | Glyceria striata | N. America | 3.8-6.2 | 2.2-2.8 | 31.0±5.0 | 2.5 | 1 | 17.3-21.0 |
| E. brachyelytri | Brachyelytrum erectum | N. America | 4.1±0.6 | 2.8±0.3 | 16.0±3.0 | 2.0-2.8 | 0.5 | 5.3-17.0 |
| E. elymi | Elymus virginicus | N. America | 4.0±0.4 | 2.2±0.2 | 17.0±3.0 | 2 | 0.5 | 13.7-20.0 |
| E. yangzii | Roegneia kamoji | China | 4.7-5.2 | 2.0-2.9 | 16.5-25.8 | 2.2-2.9 | <1.0 | 5.8-18.1 |
| E. bromicola | Bromus ramosus | Switzerland | 3.8±0.4 | 2.0±0.3 | 8.0-23.0 | 1.2-2.9 | 0.5 | 16.0-17.3 |
| E. poae | Poa secunda subsp. juncifolia | S. America | 3.0–4.4 | 1.7–3.2 | 9.9–24.0 | 1.2–2.0 | 0.4–0.8 | 11.0-12.3 |
| E. canadensis | Elymus canadensis | N. America | 5.75–8 | 2.5–4 | 12.5–41.5 | 1.5–3.0 | 0.5 | 6.7-12.0 |
| E. liyangensis | Poa pratensis ssp. pratensis | China | 5.9–6.4 | 3.0–3.2 | 22.1–24.5 | 2.3–2.5 | 1 | 12.1-16.6 |
| Epichloë sp. (MTI) | Leymus chinensis | China | 5.3±0.1× | 3.5±0.1 | 29.0–31.0 | 1.4–3.0 | 1.7±0.07 | |
| Strains in this study | Leymus chinensis | China | 4.7~5.5 | 2.8~3.5 | 24.1~26.4 | 2.1~2.5 | <1.0 | 9.5~13.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



