Submitted:
15 June 2026
Posted:
01 July 2026
You are already at the latest version
Abstract
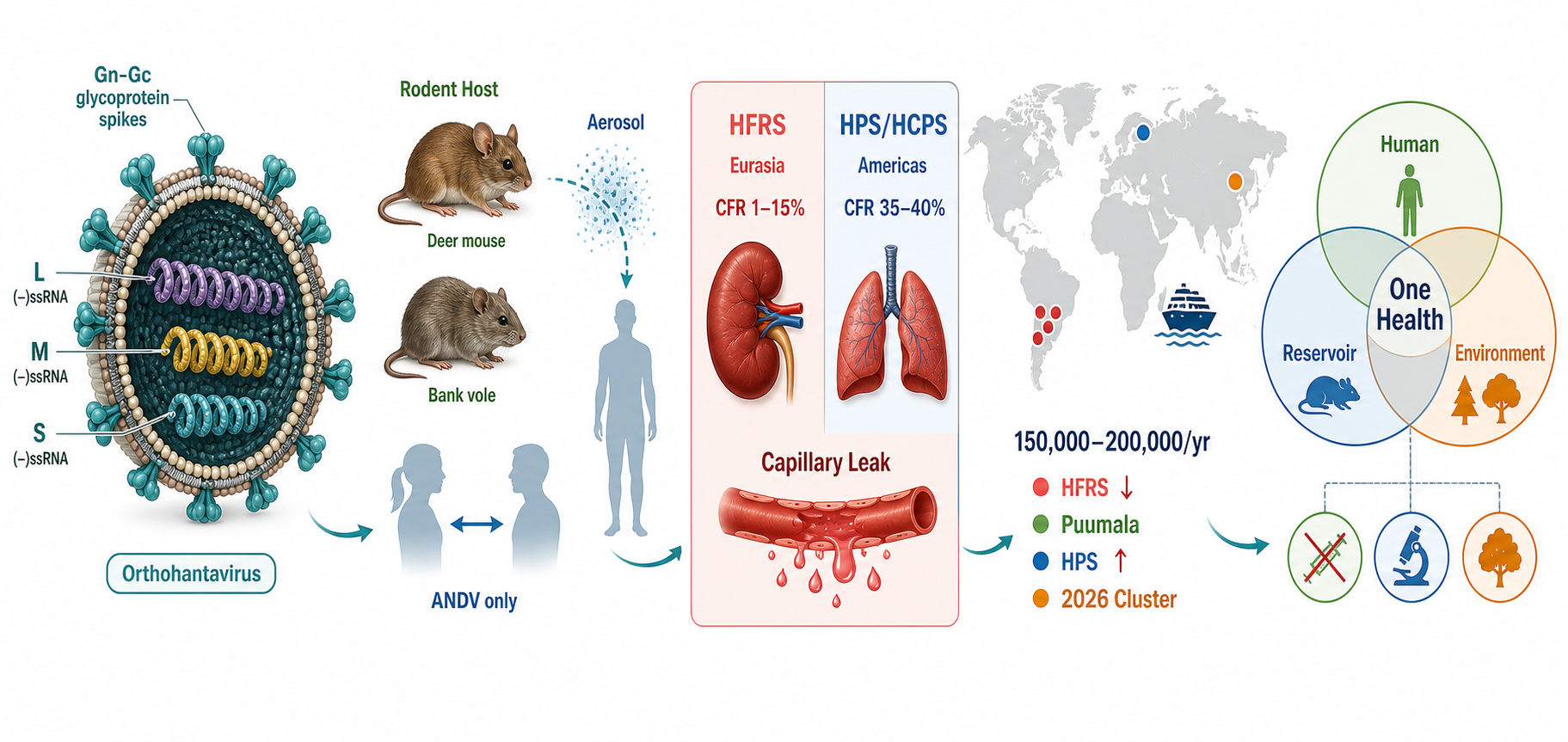
Hantaviruses (family Hantaviridae, genus Orthohantavirus) are globally distributed zoonotic RNA viruses maintained in rodent, insectivore, and chiropteran reservoirs, with humans infected mainly through inhalation of aerosolized excreta. They cause two major syndromes: hemorrhagic fever with renal syndrome (HFRS), endemic in Europe and Asia, and hantavirus cardiopulmonary syndrome (HCPS), reported principally in the Americas and associated with higher case fatality. Surveillance and seroprevalence data indicate that the true burden is underestimated because subclinical infection and clinical overlap with dengue, leptospirosis, scrub typhus, and other acute febrile illnesses limit recognition. Recent trends include climate-associated shifts in rodent reservoir dynamics, land-use-driven expansion of human–rodent interfaces, and renewed concern regarding Andes virus, the only orthohantavirus with documented person-to-person transmission. No broadly licensed antiviral therapy is available, and vaccine access remains geographically limited. This review synthesizes current evidence on hantavirus taxonomy, epidemiology, transmission, pathogenesis, diagnosis, clinical management, vaccines, and prevention, while framing control within a One Health approach that integrates human, animal, and environmental surveillance.
Keywords:
hantavirus
; orthohantavirus
; hemorrhagic fever with renal syndrome
; hantavirus cardiopulmonary syndrome
; One Health
; emerging infectious disease
; rodent reservoir
; Andes virus
; epidemiology
; zoonosis
1. Introduction
Zoonotic infectious diseases account for approximately 60% of known human infectious diseases and approximately 75% of recently emerging infectious diseases, with rodent-associated pathogens constituting a disproportionately large segment of this burden [1]. Hantaviruses represent a paradigmatic example of a rodent-borne viral zoonosis: globally distributed, capable of causing severe and frequently fatal disease, and intimately linked to the ecology of their reservoir hosts [2]. First scientifically described in the context of Korean Hemorrhagic Fever during the Korean War era (with the causative agent—Hantaan virus—isolated in 1978 by Ho-Wang Lee and colleagues), hantaviruses have since been recognized as a family of negative-sense, tripartite RNA viruses now classified within the order Bunyavirales, family Hantaviridae [3].
The revised International Committee on Taxonomy of Viruses (ICTV) 2024 taxonomy recognizes Hantaviridae as a family of negative-sense RNA viruses with genomes of approximately 10.5–14.6 kb, maintained in and transmitted by fish, reptiles, and mammals, with multiple orthohantavirus species capable of causing human disease [4]. The genomic architecture consists of three negative-sense single-stranded RNA segments: the small (S) segment encoding the nucleoprotein (N), the medium (M) segment encoding the glycoprotein precursor (GPC) that is cleaved into Gn and Gc surface glycoproteins, and the large (L) segment encoding the RNA-directed RNA polymerase [4,5].
Global estimates of annual hantavirus incidence range from 10,000 to over 100,000 cases, with the highest burden borne by East and Southeast Asia, Eastern Europe, and the Andean regions of South America [6]. Yet these figures almost certainly underestimate the true burden: serological data from a landmark 2024 meta-analysis derived a pooled global seroprevalence of 2.93%, implying that millions of prior infections—including subclinical ones—have gone undiagnosed and unreported [7]. Furthermore, evolving climatic and ecological conditions are reshaping the distribution and abundance of reservoir rodent populations, creating conditions conducive to novel and recurrent outbreaks [8].
The One Health framework—which integrates human, animal, and environmental health provides the most appropriate conceptual and operational structure for addressing hantavirus risk. As hantavirus transmission is fundamentally driven by rodent ecology, land-use change, agricultural practices, and climate variability, a silo-based, human-health-only response is structurally inadequate [9]. This review aims to synthesize the current state of hantavirus epidemiology, virology, pathogenesis, clinical management, and prevention, while explicitly embedding findings within a One Health perspective.
2. Virology and Taxonomy
2.1. Taxonomic Classification
According to the 2024 ICTV profile of Hantaviridae, the family belongs to the order Bunyavirales and contains two subfamilies: Mammantavirinae (containing the genus Orthohantavirus) and Repantavirinae (containing genera associated with reptiles and fish) [4]. The genus Orthohantavirus contains all species of public health relevance. Current taxonomy recognizes over 40 distinct orthohantavirus species, with new species continuously being described through wildlife surveillance programs [4]. The species demarcation criterion is a pairwise evolutionary distance (PED) exceeding 0.1 in the nucleoprotein coding region.
Pathogenic species of primary concern include: Hantaan orthohantavirus (HTNV), Puumala orthohantavirus (PUUV), Dobrava-Belgrade orthohantavirus (DOBV), Seoul orthohantavirus (SEOV), and Saaremaa orthohantavirus (SAAV) causing HFRS in Eurasia; and Sin Nombre orthohantavirus (SNV), Andes orthohantavirus (ANDV), and Laguna Negra orthohantavirus (LANV) causing HCPS in the Americas [5,10].
2.2. Genome and Replication
Orthohantavirus virions are pleomorphic, enveloped particles of 80–160 nm diameter [4]. Replication occurs in the cytoplasm following receptor-mediated endocytosis, with CD55 (decay-accelerating factor), β3 integrins, and protocadherin-1 (PCDH1) identified as key entry receptors depending on the virus species and cell type [5]. The RNA-dependent RNA polymerase initiates replication through cap-snatching, a mechanism in which the viral polymerase cleaves 5′-capped host mRNA fragments to prime viral RNA synthesis, and viral assembly occurs at the Golgi apparatus before exocytosis. The endothelium is the primary target of pathogenic orthohantaviruses, and endothelial infection leads to increased vascular permeability without direct cytolytic destruction—a central feature distinguishing hantavirus pathogenesis from many other hemorrhagic fever viruses [10].
2.3. Host–Virus Co-Evolution
A defining characteristic of orthohantaviruses is their long-term co-evolutionary relationship with specific reservoir rodent or insectivore species, which maintain persistent, generally asymptomatic infection [2]. Each orthohantavirus is typically associated with a single primary reservoir: HTNV with Apodemus agrarius (striped field mouse), PUUV with Myodes glareolus (bank vole), SEOV with Rattus norvegicus (brown rat), ANDV with Oligoryzomys longicaudatus (long-tailed pygmy rice rat), and SNV with Peromyscus maniculatus (North American deer mouse) [2,10]. Intraspecific horizontal transmission among reservoir rodents occurs through biting and is influenced by rodent population density, seasonality, and mast events [11].
3. Global Epidemiology
3.1. Burden of Disease
Hantavirus infection imposes a substantial yet incompletely characterized global health burden. The World Health Organization estimates between 10,000 and over 100,000 infections annually [6]. A 2024 systematic review and meta-analysis by Tortosa et al., encompassing 110 studies with 81,815 participants, estimated the global pooled seroprevalence at 2.93% (95% CI: 2.34–3.67%), representing the most comprehensive assessment of hantavirus burden to date [7]. Regional seroprevalence varied: the Americas, 2.43% (95% CI: 1.71–3.46%); Europe, 2.98% (95% CI: 2.19–4.06%); with China representing the dominant Asian burden [7].
China reports the highest absolute HFRS burden globally. Data from 31 provincial-level administrative divisions for 2014–2023 documented 91,388 HFRS cases, with an average annual incidence of 0.65 per 100,000 population [12]. The incidence fluctuated between 0.37 and 0.86 per 100,000, peaking in 2018, with the disease concentrated in male agricultural workers aged 20–60 years. The causative agents are principally HTNV in rural forested zones and SEOV in peri-urban settings [12].
3.2. European Epidemiology
In Europe, HFRS is predominantly caused by PUUV and DOBV. PUUV, maintained by the bank vole Myodes glareolus, causes nephropathia epidemica (NE)—a generally milder form of HFRS with a CFR below 0.3%—concentrated in Scandinavia, Germany, the Baltic states, and Russia. In Finland, incidence rates can surge to over 25 per 100,000 population during peak vole years [8]. A 2026 pan-European analysis identified climate variability, bank vole population dynamics, and socioeconomic factors as the key determinants of spatiotemporal hantavirus risk across the continent [8]. Mast events (periods of heavy beech and oak seed production) drive bank vole population explosions, which in turn precipitate PUUV transmission peaks, with a characteristic 12–18 month lag between mast event and human cases [11].
3.3. Americas Epidemiology
In the Americas, HCPS presents as an acute respiratory distress syndrome with the highest CFR of any hantavirus disease (20–50%). Sin Nombre virus (SNV), the principal agent in North America, causes disease following inhalation of deer mouse excreta; although case numbers remain relatively limited compared with Old World HFRS, individual outbreaks are associated with high clinical severity [10]. ANDV is responsible for the majority of HCPS cases in Argentina, Chile, and Uruguay, and represents a unique epidemiological and public health concern because of its documented capacity for person-to-person transmission [14,15]. A 2019 cluster in Patagonia, Argentina, involving 29 confirmed HCPS cases and 11 deaths illustrated the outbreak potential of ANDV under conditions of extended close contact [15].
Recent multi-country events further demonstrate the international public health relevance of ANDV. In 2026, WHO reported a hantavirus outbreak linked to MV Hondius cruise ship travel, with more than 600 contacts identified across 32 countries, territories, and areas by 22 May 2026 [13]. CDC communications confirmed Andes virus as the causative agent and documented cases and deaths among passengers and crew [17]. These events underline how travel, close-contact settings, and delayed recognition can internationalize a historically regional zoonotic risk.
3.4. Seroprevalence in Emerging Regions
A 2025 cross-sectional seroprevalence study in West Kazakhstan documented the burden in a previously understudied region [16]. In Central Asia more broadly, Seoul virus—with its globally distributed reservoir Rattus norvegicus—represents an underappreciated source of human HFRS, theoretically present wherever the brown rat is found, encompassing all inhabited continents [10].

Table 1.
Major orthohantaviruses of public health importance: reservoir hosts, geographic distribution, and associated clinical syndromes.
Table 1.
Major orthohantaviruses of public health importance: reservoir hosts, geographic distribution, and associated clinical syndromes.
| Virus Species | Primary Reservoir | Geographic Distribution | Clinical Syndrome | CFR (%) |
| Hantaan orthohantavirus (HTNV) | Apodemus agrarius | China, Republic of Korea, Russia | HFRS (severe) | 5–15 |
| Puumala orthohantavirus (PUUV) | Myodes glareolus | Scandinavia, Central/North Europe | HFRS/NE (mild–moderate) | <0.3 |
| Seoul orthohantavirus (SEOV) | Rattus norvegicus | Global (cosmopolitan) | HFRS (mild–moderate) | 1–5 |
| Dobrava-Belgrade orthohantavirus (DOBV) | Apodemus flavicollis | Balkans, Central Europe | HFRS (severe) | 3–12 |
| Sin Nombre orthohantavirus (SNV) | Peromyscus maniculatus | North America | HCPS | 20–40 |
| Andes orthohantavirus (ANDV) | Oligoryzomys longicaudatus | Argentina, Chile, Uruguay | HCPS; P2P ᵃ | 30–50 |
| Laguna Negra orthohantavirus (LANV) | Calomys laucha | Paraguay, Bolivia, Argentina | HCPS | 15–25 |
CFR = case fatality rate; HFRS = hemorrhagic fever with renal syndrome; HCPS = hantavirus cardiopulmonary syndrome; NE = nephropathia epidemica; P2P = person-to-person. ᵃ Person-to-person transmission is documented for ANDV only and requires sustained close contact [14,15,17]. Data from [4,6,10,12].
4. Transmission Dynamics
4.1. Rodent-to-Human Transmission
The primary route of human infection is inhalation of virus-laden aerosols generated from dried rodent urine, feces, or saliva, making enclosed, poorly ventilated spaces associated with rodent infestations (agricultural storage facilities, rural cabins, construction sites) the highest-risk settings [6]. Direct contact with rodents or their excreta, contamination of mucosal membranes, and, rarely, rodent bites also contribute to transmission [6]. No arthropod vector has been identified; hantaviruses are therefore distinct from arboviruses, and insect repellents do not provide specific protection against hantavirus transmission.
The duration and efficiency of viral shedding in reservoir rodents varies by species and virus. In Myodes glareolus infected with PUUV, peak viral RNA levels are detectable in excreta for 11–28 days following infection, though RNA can persist in saliva and feces for up to 84 and 44 days, respectively [2]. Significant gender dimorphism in seroprevalence—consistently higher in males—reflects differential occupational and behavioural exposure rather than intrinsic biological susceptibility [7].
4.2. Person-to-Person Transmission: The Andes Virus Exception
Among the 40+ recognized orthohantavirus species, ANDV stands unique in its demonstrated capacity for person-to-person transmission [14,15]. This mode of transmission has been confirmed through epidemiological clustering analyses, genomic sequencing demonstrating phylogenetic linkage between human cases, and immunohistochemical evidence for salivary and respiratory routes of spread [15]. A 2020 study by Alonso et al. confirmed person-to-person ANDV transmission in a 2014 Argentine cluster using full-length genome sequencing—providing the most definitive molecular evidence to date [14]. Person-to-person transmission appears to occur primarily during the prodromal phase of illness, through prolonged close contact (particularly household and sexual partners), rather than via casual or community contact [14,15]. No documented presymptomatic person-to-person transmission has been confirmed as of 2026 [17].
The current understanding, as reflected in May 2026 CDC emergency guidance issued in the context of an ANDV-associated cluster on a cruise ship, is that person-to-person ANDV transmission, while real, remains relatively rare and requires sustained close contact [17]. This epidemiological profile distinguishes ANDV from SARS-CoV-2 and other easily transmitted respiratory pathogens, though it demands heightened infection control precautions in clinical and household settings when ANDV is suspected.
5. Pathogenesis and Immunology
Orthohantaviruses are non-cytolytic in their reservoir hosts but cause severe immunopathology in incidental human hosts. The primary target cell is the microvascular endothelium; hantaviruses infect endothelial cells via integrin and PCDH1-mediated entry without lysing them, instead dysregulating tight junction integrity and inducing vascular hyperpermeability [5,10]. The resultant pathophysiological consequence is plasma leakage: into the retroperitoneal space and renal interstitium in HFRS, or into the pulmonary vasculature in HCPS.
Immunopathological mechanisms involve vigorous CD8+ T-cell responses, dysregulated pro-inflammatory cytokine production (notably TNF-α, IFN-γ, and IL-6), and impaired regulatory T-cell activity [10]. Thrombocytopenia—resulting from both platelet consumption and megakaryocyte dysfunction—is a near-universal early feature and a critical diagnostic marker [10]. In severe HCPS, a cardiogenic-depressant component involving reduced cardiac output, elevated pulmonary capillary wedge pressure, and systemic vasoconstriction compounds the non-cardiogenic capillary leak, resulting in a hemodynamic profile distinct from classic septic shock [10].
The incubation period ranges from one to six weeks (typically two to four weeks), during which the virus disseminates silently via mononuclear cells acting as Trojan horses to the target endothelium [6,10]. Clinical presentation follows a distinct phasic pattern for HFRS (febrile, hypotensive, oliguric, diuretic, and convalescent phases) and a biphasic pattern for HCPS (febrile prodrome followed by abrupt cardiopulmonary decompensation) [10].
6. Clinical Features and Diagnosis
6.1. Hemorrhagic Fever with Renal Syndrome
HFRS presents as an acute febrile illness characterized by the sequential development of renal impairment, hemorrhagic manifestations, and electrolyte disturbances [10]. Initial symptoms are non-specific: high fever, myalgia, headache, and retroorbital pain. Petechiae, conjunctival hemorrhage, and thrombocytopenia typically emerge within the first week, followed by acute kidney injury (AKI) manifesting as oliguria, proteinuria, and elevated creatinine. Severe HFRS (principally caused by HTNV and DOBV) may produce hemorrhagic shock, hemorrhage of visceral organs, and multi-organ failure [10]. The milder NE form caused by PUUV is characterized predominantly by self-limiting AKI with rare hemorrhage and a CFR below 0.3% [8].
6.2. Hantavirus Cardiopulmonary Syndrome
HCPS has a dramatic clinical course: a 3–7-day febrile prodrome of myalgia, headache, and gastrointestinal symptoms is followed by abrupt onset of non-cardiogenic pulmonary edema, tachycardia, and hemodynamic collapse [10]. The hallmark radiographic finding is bilateral interstitial infiltrates or pulmonary edema without cardiomegaly. Thrombocytopenia, neutrophilia with left shift, hemoconcentration, and elevated lactate are characteristic laboratory findings. ECMO (extracorporeal membrane oxygenation) has been used as a bridge to hemodynamic recovery in refractory HCPS, and early transfer to ECMO-capable centers is now recommended in severe cases [10].
6.3. Diagnosis
Diagnosis is primarily serological: detection of IgM and IgG antibodies against hantavirus nucleoprotein by enzyme-linked immunosorbent assay (ELISA) or immunofluorescence assay (IFA) [10]. IgM antibodies are usually detectable at or shortly before symptom onset, enabling acute-phase diagnosis. Reverse-transcription polymerase chain reaction (RT-PCR) on blood, plasma, or tissue permits species-level identification during the viremic phase (typically the first 4–7 days of illness) and is valuable for outbreak investigation and contact tracing [10,12]. Cross-reactive antibody responses across orthohantavirus species can complicate serological species attribution in regions of co-circulation, necessitating molecular confirmation. Rapid point-of-care diagnostics and multiplex acute-febrile-illness panels remain insufficiently deployed in many endemic and emerging settings, contributing to underascertainment.
Table 2.
Comparative clinical and diagnostic features of hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (HCPS).
Table 2.
Comparative clinical and diagnostic features of hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (HCPS).
| Feature | HFRS | HCPS |
| Causative viruses | HTNV, SEOV, PUUV, DOBV, SAAV | SNV, ANDV, LANV |
| Endemic region | Europe, Asia | Americas |
| Target organ(s) | Kidney, vascular endothelium | Lung, cardiovascular system |
| CFR range | 1–15% (HTNV/DOBV); <0.3% (PUUV) | 20–50% (SNV, ANDV) |
| Hallmark lab finding | Thrombocytopenia + AKI | Thrombocytopenia + hypoxia |
| P2P transmission | Not documented | ANDV only |
| Key diagnostic test | IgM ELISA + RT-PCR | IgM ELISA + RT-PCR |
| Antiviral treatment | IV ribavirin (early HFRS; HTNV) | No approved antiviral |
| Critical supportive care | Fluid balance; renal replacement therapy | Lung-protective ventilation; ECMO |
7. Treatment, Vaccines, and Prevention
7.1. Antiviral Therapy
No broadly licensed, specifically approved antiviral therapy exists for hantavirus disease as of 2026 [10,18]. Intravenous ribavirin has demonstrated efficacy in early HFRS—particularly in reducing CFR in HTNV-associated disease when administered within the first four days of fever—and is used in several Asian and European countries, though it has not demonstrated convincing benefit in HCPS [10]. The absence of an approved antiviral for HCPS means that management remains supportive: careful hemodynamic monitoring, lung-protective ventilation, vasopressors for shock, and ECMO for refractory cases [10,19].
Recent immunotherapeutic approaches have explored monoclonal antibodies, convalescent plasma, and computationally derived inhibitor molecules [18,19]. Passive immunization with high-titer convalescent plasma from ANDV survivors has shown some signal in observational data from Chile and Argentina, though phase III trial data are lacking. The 2024 Frontiers editorial by Ul Qamar et al. highlighted advances in RNA-based therapeutics and immunoinformatics-guided drug discovery for hantaviruses, though none have reached clinical translation [19].
7.2. Vaccines
Inactivated bivalent vaccines (incorporating HTNV and SEOV antigens) are licensed and used in China and the Republic of Korea, where they have contributed to reductions in HFRS incidence among vaccinated populations [20]. However, limitations include variable protection across strains, requirement for three-dose primary series with boosters, and limited immunogenicity data in elderly populations [20]. A 2025 study by Aram et al. in the MDPI journal Vaccines investigated a modified vaccinia Ankara (MVA)-vectored vaccine incorporating a chimeric SEOV-HTNV nucleoprotein antigen, demonstrating induction of both humoral and cellular immunity with reduction in viral load in a type-I interferon receptor-deficient mouse model (A129) [21].
A 2025 comprehensive review in Vaccines by Chai et al. outlined the trajectory of orthohantavirus vaccine development, from first-generation inactivated platforms to emerging DNA, mRNA, virus-like particle (VLP), and recombinant subunit candidates targeting Gn/Gc glycoproteins [20]. The lack of universally accepted immunological correlates of protection and the absence of a standardized animal challenge model that recapitulates human HCPS have historically impeded clinical development [20].
7.3. Prevention and Rodent Control
In the absence of a universally available vaccine or antiviral, primary prevention relies on interrupting human-rodent contact. Evidence-based measures include: rodent-proofing of dwellings and food storage; avoidance of sweeping or vacuuming activities in rodent-infested environments (which aerosolize infectious particles); use of N95 respirators and gloves when cleaning potentially contaminated spaces; and rodenticide-based population management in high-transmission settings [6,17]. For healthcare workers caring for suspected ANDV patients, enhanced contact and droplet precautions—including N95 respirators, gowns, gloves, and eye protection—are recommended during close-contact procedures [17].
8. One Health Perspectives
8.1. The One Health Framework Applied to Hantavirus
The One Health approach, formally defined as an integrated, unifying framework that aims to sustainably balance and optimize the health of people, animals, and ecosystems, is particularly well-suited to the hantavirus system [9]. Hantavirus transmission is fundamentally driven by ecological determinants—rodent population abundance, species composition, habitat type, and climate—that cannot be addressed by human health systems acting in isolation [9,11]. Effective hantavirus risk reduction requires simultaneous and coordinated action across veterinary, environmental, and public health disciplines.
8.2. Climate Change and Shifting Rodent Ecology
Climate change is reshaping the epidemiology of hantavirus through multiple interconnected pathways [8,11]. Warmer temperatures and altered precipitation patterns affect the phenology and abundance of plant food sources for rodent reservoir populations: mast years driven by elevated beech and oak seed production are associated with population explosions of Myodes glareolus in Europe, directly preceding PUUV outbreak peaks [8,11]. In the Americas, drought-rainfall cycles can alter vegetation productivity and rodent breeding success, creating periodic increases in human-rodent contact [11].
Milder winters in Europe are contributing to higher orthohantavirus prevalence and an expanded geographic risk range by reducing winter die-off of bank vole populations and extending the transmission season [8]. Modeling analyses suggest that projected climate trajectories under shared socioeconomic pathway (SSP) scenarios will further expand the European risk zone for PUUV infections northward and into higher altitudes through 2100 [8].
8.3. Land-Use Change and Urbanization
Deforestation, agricultural expansion, and urbanization alter the composition and distribution of small mammal communities, often favoring the commensal and opportunistic rodent species that serve as primary hantavirus reservoirs [9]. Poorly managed periurban settlements with inadequate waste disposal and sanitation infrastructure create ecological conditions conducive to Rattus norvegicus and Rattus rattus proliferation, expanding the geographic footprint of SEOV [9]. Conversely, forest fragmentation in South America places agricultural communities at the forest-edge interface, increasing exposure to ANDV-infected pygmy rice rats [2].
8.4. Integrated Surveillance
A One Health surveillance architecture for hantavirus requires parallel monitoring of (i) rodent population abundance and seroprevalence in wildlife reservoir communities; (ii) genomic surveillance of circulating hantavirus strains for evolutionary change and geographic spread; and (iii) human epidemiological surveillance with case confirmation by molecular and serological methods [9]. Few countries currently implement all three components in a coordinated fashion. WHO and FAO have called for expansion of integrated One Health surveillance platforms, particularly in endemic countries with limited laboratory capacity [6]. The use of environmental RNA detection from water sources and dusty environments as a sentinel surveillance tool for rodent-borne pathogens represents a nascent but promising complementary approach.
8.5. Global Connectivity and Emerging Risk
The May 2026 outbreak of Andes virus-associated HCPS cases linked to MV Hondius cruise ship travel, with subsequent contact tracing across multiple countries, illustrates how global travel and connectivity can amplify the international public health implications of historically geographically restricted zoonoses [13,17]. Imported hantavirus cases present diagnostic challenges in non-endemic settings and require heightened clinician awareness. As international wildlife tourism, adventure travel, and global cargo movement expand rodent range and human-rodent contact opportunities, the zoonotic hantavirus risk profile becomes genuinely global rather than regionally constrained.
9. Research Gaps and Future Directions
Despite substantial research progress, critical gaps persist that limit hantavirus prevention and control:
First, the correlates of protective immunity against hantavirus—both humoral (neutralizing antibody titers against Gn/Gc) and cellular (CD8+ T-cell responses)—remain incompletely characterized. This gap impedes rational vaccine development and endpoint selection for clinical trials [20].
Second, the mechanisms and risk factors governing the unique person-to-person transmissibility of ANDV—and the absence of this property in other orthohantaviruses—remain poorly understood at the molecular level [14,15]. Identification of specific viral or host determinants could inform antiviral targeting and infection control strategies.
Third, the global burden of hantavirus is systematically underestimated due to diagnostic underascertainment, particularly in settings where hantavirus co-circulates with other febrile illnesses (dengue, leptospirosis, typhoid) generating clinical diagnostic confusion. Expanded point-of-care diagnostic capacity and multiplex syndromic testing platforms are research and implementation priorities [7].
Fourth, although climate-driven projections of increased risk are increasingly supported by observational data, operational ecological early warning systems that translate mast event data, rodent population monitoring, and climate forecasts into actionable public health risk alerts have not been implemented at scale. Investment in transdisciplinary modeling and decision-support tools is warranted [8,11].
Fifth, next-generation vaccine platforms—including mRNA vaccines, recombinant VLPs, and MVA-vectored constructs—offer accelerated development timelines and improved cross-reactive immunogenicity compared to first-generation inactivated vaccines, but require phase II/III efficacy data to support regulatory approval [20,21].
Sixth, the burden of hantavirus infection in South and Southeast Asia, including the Indian subcontinent, remains poorly characterized because of limited surveillance infrastructure and clinical overlap with dengue, leptospirosis, scrub typhus, and other acute febrile illnesses. Region-specific seroepidemiological studies and inclusion of hantavirus in multiplex acute-febrile-illness algorithms are warranted [7,10].
10. Conclusions
Hantavirus infection remains a globally distributed, clinically severe, and incompletely controlled zoonotic threat. With no broadly approved antiviral and limited vaccine access, prevention through a One Health lens—integrating ecological monitoring, rodent control, climate-informed risk assessment, and coordinated human–animal–environmental surveillance—represents the most viable pathway to reducing the burden of hantavirus disease. The unique person-to-person transmission of Andes virus and the climate-mediated expansion of reservoir rodent distributions underscore the dynamic and evolving nature of this threat. Sustained investment in surveillance infrastructure, diagnostic capacity, and translational research for antivirals and next-generation vaccines is urgently warranted to anticipate and contain this neglected yet recurrently emerging pathogen.
Author Contributions
Conceptualization, S.P. and K.G.S.; methodology, S.P. and K.G.S.; literature search and investigation, S.K., H.J., P., S.D.P., V.P., S.L.R., and R.M.; writing—original draft preparation, S.P. and K.G.S.; writing—review and editing, S.K., H.J., K.G.S., P., S.D.P., V.P., S.L.R., and R.M.; supervision, S.P. and K.G.S.; project administration, S.P. and K.G.S. All authors have read and agreed to the submitted version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable. This review article did not involve human participants, animals, or primary data collection.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable. No new data were created or analyzed in this review.
Acknowledgments
The authors acknowledge K.G. Sruthi, PhD Scholar, Department of Clinical Microbiology, School of Allied Health Sciences, Malla Reddy University, Hyderabad, Telangana, India, for helping refine the manuscript content and improve the clarity and scientific presentation of the review.
Conflicts of Interest
The authors declare no conflicts of interest.:.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Full Term |
| AKI | Acute kidney injury |
| ANDV | Andes orthohantavirus |
| CFR | Case fatality rate |
| DOBV | Dobrava-Belgrade orthohantavirus |
| ECMO | Extracorporeal membrane oxygenation |
| ELISA | Enzyme-linked immunosorbent assay |
| GPC | Glycoprotein precursor |
| HCPS | Hantavirus cardiopulmonary syndrome |
| HFRS | Hemorrhagic fever with renal syndrome |
| HTNV | Hantaan orthohantavirus |
| ICTV | International Committee on Taxonomy of Viruses |
| IFA | Immunofluorescence assay |
| LANV | Laguna Negra orthohantavirus |
| MVA | Modified vaccinia Ankara |
| NE | Nephropathia epidemica |
| P2P | Person-to-person |
| PCDH1 | Protocadherin-1 |
| PUUV | Puumala orthohantavirus |
| RT-PCR | Reverse-transcription polymerase chain reaction |
| SAAV | Saaremaa orthohantavirus |
| SEOV | Seoul orthohantavirus |
| SNV | Sin Nombre orthohantavirus |
| VLP | Virus-like particle |
| WHO | World Health Organization |
References
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Schmaljohn, C.S. A brief history of bunyaviral family Hantaviridae. Diseases 2023, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Bradfute, S.B.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Laenen, L.; Tischler, N.D.; Maes, P. ICTV Virus Taxonomy Profile: Hantaviridae 2024. J. Gen. Virol. 2024, 105, 001975. [Google Scholar] [CrossRef] [PubMed]
- Vial, P.A.; Ferrés, M.; Vial, C.; Klingström, J.; Ahlm, C.; López, R.; Le Corre, N.; Mertz, G.J. Hantavirus in humans: A review of clinical aspects and management. Lancet Infect. Dis. 2023, 23, e371–e382. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Hantavirus Fact Sheet. WHO; Geneva, 2026. Available online: https://www.who.int/news-room/fact-sheets/detail/hantavirus (accessed on 8 June 2026).
- Tortosa, F.; Perre, F.; Tognetti, C.; Lossetti, L.; Carrasco, G.; Guaresti, G.; Iglesias, A.; Espasandin, Y.; Izcovich, A. Seroprevalence of hantavirus infection in non-epidemic settings over four decades: A systematic review and meta-analysis. BMC Public Health 2024, 24, 2553. [Google Scholar] [CrossRef] [PubMed]
- Tersago, K.; Verhagen, R. A pan-European assessment of multi-sector drivers of human hantavirus risk: Climate, biodiversity, and socio-economic factors as key determinants. Environ. Res. 2026. [Google Scholar] [CrossRef]
- Duong, V.; Morand, S.; Buchy, P. Editorial: Rodent-borne diseases: ‘One Health’ perspectives. Front. Vet. Sci. 2025, 12, 1602402. [Google Scholar] [CrossRef] [PubMed]
- Noack, D.; Goeijenbier, M.; Reusken, C.; Koopmans, M.P.G.; Rockx, B.H.G. Orthohantavirus pathogenesis and cell tropism. Front. Cell. Infect. Microbiol. 2020, 10, 399. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Stenseth, N.C. The ecological dynamics of hantavirus diseases: From environmental variability to disease prevention largely based on data from China. PLoS Negl. Trop. Dis. 2019, 13, e0006901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Y.; Zhao, J.; China CDC HFRS Surveillance Team. Hemorrhagic fever with renal syndrome and diversity and distribution of hantaviruses—China, 2014–2023; China CDC Wkly, 2025; p. PMC12228097. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Hantavirus outbreak linked to cruise ship travel, Multi-locations. Disease Outbreak News. 28 May 2026. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2026-DON604 (accessed on 8 June 2026).
- Alonso, D.O.; Pérez-Sautu, U.; Bellomo, C.M.; Prieto, K.; Iglesias, A.; Coelho, R.; Periolo, N.; Domenech, I.; Talmon, G.; Hansen, R.; et al. Person-to-person transmission of Andes virus in hantavirus pulmonary syndrome, Argentina, 2014. Emerg. Infect. Dis. 2020, 26, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, E.; Navarrete, M.; Mendez, C.; Zaror, L.; Mansilla, C.; Tapia, M.; Carrasco, C.; Salazar, P.; Murua, R.; Padula, P.; et al. Immunocytochemical and ultrastructural evidence supporting that Andes hantavirus is transmitted person-to-person through the respiratory and/or salivary pathways. Front. Microbiol. 2020, 10, 2992. [Google Scholar] [CrossRef] [PubMed]
- Gubareva, U.; Horth, R.; Nabirova, D.; Tukhanova, N.; Utegenova, E.; Shapiyeva, Z.; Turliyev, Z.; Tleumbetova, N.; Maykanov, N.; Smagul, M.; et al. Hantavirus antibody seroprevalence and risk factors among adults in West Kazakhstan, 2023. Front. Public Health 2025, 12, 1519117. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. 2026 Multi-country Hantavirus Cluster Linked to Cruise Ship. Health Alert Network (HAN) Health Advisory. 8 May 2026. Available online: https://www.cdc.gov/han/php/notices/han00528.html (accessed on 8 June 2026).
- Ul Qamar, M.T.; Ahmad, S.; Khan, A.; Wei, D. Editorial: Immunotherapeutics development against hantaviruses. Front. Immunol. 2024, 15, 1377137. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and therapeutics against hantaviruses. Front. Microbiol. 2020, 10, 2989. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Wang, L.; Du, H.; Jiang, H. Achievement and challenges in orthohantavirus vaccines. Vaccines 2025, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- Aram, M.; Graham, V.; Kennedy, E.; Rayner, E.; Hewson, R.; Dowall, S. A multi-valent hantavirus vaccine based on recombinant modified vaccinia Ankara reduces viral load in a mouse infection model. Vaccines 2025, 13, 270. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



