Submitted:
16 June 2026
Posted:
17 June 2026
You are already at the latest version
Abstract
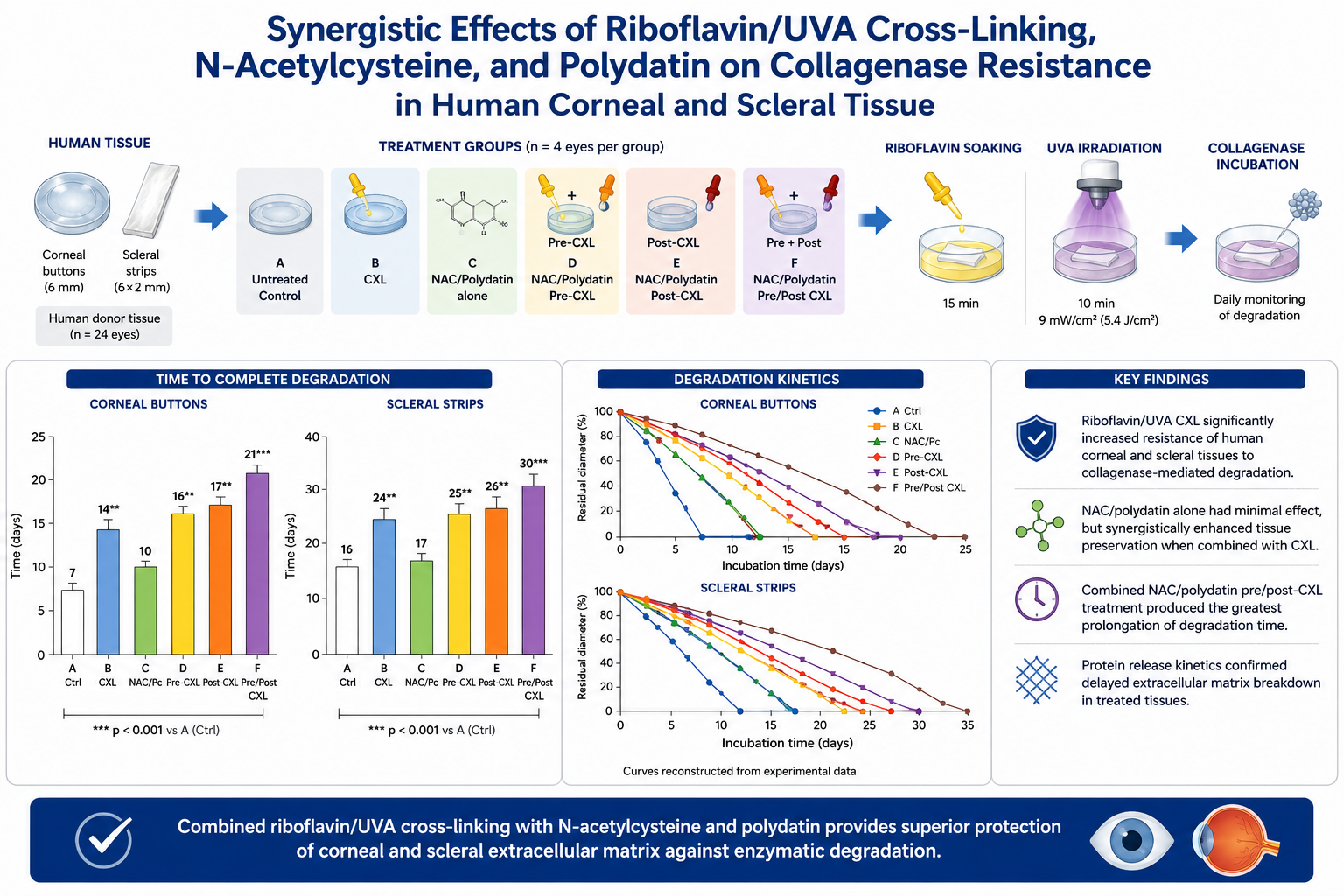
The study evaluated the resistance of human corneal and scleral tissues to collagenase-mediated degradation after riboflavin/ultraviolet-A collagen crosslinking (CXL), alone or combined with N-acetylcysteine (NAC) and Polygonum cuspidatum dry extract (Polydatin). Twenty-four human donor eyes unsuitable for transplantation provided 12 corneal buttons and 12 scleral strips, which were allocated to untreated controls, CXL alone, NAC/polydatin alone, and combined pre- and/or post-CXL treatment groups. Samples underwent accelerated CXL (9 mW/cm², 5.4 J/cm²) and were subsequently exposed to collagenase solution. Degradation time and protein release were assessed. Untreated corneal and scleral tissues were completely digested after 7 ± 2 and 16 ± 2 days, respectively. CXL significantly increased resistance to enzymatic degradation, extending digestion time to 14 ± 2 days in corneal tissue and 24 ± 2 days in scleral tissue (p < 0.01). The combination of NAC and polydatin administered before and after CXL further enhanced tissue stability, prolonging degradation time to 21 ± 1.8 days for corneas and 30 ± 2 days for sclerae (p < 0.001 vs. controls). These findings suggest that NAC and Polydatin may potentiate the extracellular matrix–stabilizing effects of CXL and warrant further investigation for corneal ectasia and progressive myopia.
Keywords:
corneal collagen crosslinking
; scleral crosslinking
; keratoconus
; progressive myopia
; collagenase
; N-acetylcysteine
; polydatin
1. Introduction
Corneal collagen crosslinking (CXL) using riboflavin and ultraviolet-A (UVA) irradiation has become an established treatment for progressive keratoconus and corneal ectatic disorders.[1] By inducing the formation of reactive oxygen species, riboflavin/UVA irradiation promotes the development of additional covalent bonds between stromal collagen fibers, resulting in increased biomechanical rigidity and stabilization of the corneal extracellular matrix. [2] Clinical studies have consistently demonstrated the efficacy of CXL in slowing or halting ectatic progression and preserving visual function in keratoconus (KC) patients.[3] Beyond its biomechanical effects, increasing evidence suggests that CXL also exerts important biochemical effects on corneal tissue. [4] Spoerl et al. demonstrated that riboflavin/UVA crosslinking significantly increases resistance of corneal collagen to pepsin, trypsin, and collagenase digestion. [5]
Increased activity of matrix metalloproteinases, collagenases and extracellular matrix degradation are known to contribute to progressive stromal weakening in keratoconus and corneal melting disorders [5]. Growing attention has recently been directed toward adjunctive biochemical strategies capable of modulating extracellular matrix degradation and oxidative stress pathways. N-acetylcysteine (NAC), a thiol-containing antioxidant and precursor of glutathione, has demonstrated anti-inflammatory and antiproteolytic properties, including modulation of matrix metalloproteinase activity. [6,7,8]
Similarly, Polydatin, a natural glucoside precursor of resveratrol extracted from Polygonum cuspidatum (Pc) , exhibits antioxidant and extracellular matrix-protective effects in several experimental models.[9] Such compounds may therefore enhance tissue resistance to enzymatic degradation and potentially synergize with the stabilizing effect induced by collagen crosslinking.[10] Although the clinical role of corneal crosslinking is now well established in KC and other corneal ectasia, analogous extracellular matrix stabilization strategies may also have potential relevance in scleral remodeling associated with progressive myopia.[11,12] Progressive scleral remodeling, collagen disorganization, scleral thinning and weakening of scleral collagen-extra-cellular- matrix (ECM) complex play a central role in myopia progression.[13,14] Progressive myopia is increasingly recognized as a disease characterized by collagen disorganization, protease activation, and biomechanical weakening of the scleral ECM, ultimately contributing to equatorial stretching, ocular expansion and axial elongation.[15] Experimental studies[16,17] have demonstrated the feasibility of scleral crosslinking and suggest that increasing scleral rigidity may potentially counteract progressive deformation of the ocular wall.[18,19,20] Human cadaver eyes scleral collagen cross-linking by ultraviolet activation of riboflavin differentially increases scleral Young Modulus (YM) more in the equatorial than posterior sclera, and most in the lateral, equatorial sclera.[21] Cross-linking might be used to arrest progressive myopia or to prevent staphyloma formation.[22]
The purpose of the present study was therefore to evaluate the resistance of human corneal buttons and scleral strips to collagenase-mediated enzymatic degradation[5] following Riboflavin/UVA CXL, either alone or combined with adjunctive NAC and Pc treatment. By investigating both corneal and scleral tissue stabilization within the same experimental model, this study aims to provide further insight into combined biomechanical and biochemical ECM stabilization strategies with potential translational implications for corneal ectatic disorders and progressive myopia.
2. Materials and Methods
2.1. Setting and Donor Tissues
This experimental laboratory study was conceived at the Siena International Crosslinking Center, Monteriggioni, Siena, Italy and conducted at the Biotecnica Associates Laboratories, Castelfidardo, Ancona, Italy, in collaboration with the Veneto Eye Bank Foundation, Venice, Italy and University of Enna Kore Ophthalmology Department. The study was authorized by the Institutional Review Board of the Siena Crosslinking Center (Sclero-Corneal Enzymatic Dygestion Study 1, Code: SCED-S1) under the tenets of Helsinky declaration and respecting good laboratory practice (GLP) rules.
Twenty-four human donor corneas unsuitable for transplantation, together with their scleral rims, were obtained from the Veneto Eye Bank Foundation, Venice, Italy, for experimental research purposes. Samples were stored at 5 ± 3°C in the storage solution (Optisol™, Bausch and Lomb Inc, Rochester, NY, USA) until the beginning of the study.
Twelve corneal buttons were punched using a 6 mm trephine to obtain homogeneous samples. Included corneal buttons measured 6 mm in diameter with an average central corneal thickness of 503 ± 27 µm. Central thickness was measured with ultrasound portable US pachymetry (Pachette 4™, DGH Technology, Inc Exton PA, USA).
Twelve scleral strips measuring 6 mm in length × 2 mm in width were dissected from the corresponding scleral rims.
2.2. Experimental Groups
Corneal buttons and scleral strips were divided into six experimental groups:
- Group A: untreated controls
- Group B: riboflavin/UVA CXL alone
- Group C: NAC/Pc incubation alone
- Group D: NAC/Pc incubation before CXL
- Group E: NAC/Pc incubation after CXL
- Group F: NAC/Pc incubation before and after CXL
Corneal buttons were divided into 6 groups (2 specimens for each group).
2.3. Crosslinking Procedure
Crosslinking (CXL) treatment consisted of 15 minutes of Riboflavin soaking followed by continuous UVA irradiation for 10 minutes at 9mW/cm², corresponding to a total Fluence of 5.4 J/cm². Isotonic dextran-free 0.1% Riboflavin solution was applied during UV-A irradiation every two minutes.[23]
NAC/Pc Solution
The adjunctive bioactive solution was prepared by dissolving a 3.7 g mixture containing NAC 400 mg and Polygonum cuspidatum (Pc), Polidatin dry extract 200 mg in 250 mL of deionized water. Samples assigned to adjunctive treatment groups were incubated in this solution 20 minutes before CXL, 20 minutes after CXL, or both pre and post CXL according to Group allocation.
2.4. Enzymatic Degradation Assay
After treatment, samples were incubated at 25 ± 2°C and 60 ± 5% relative humidity in sterile multi-well plates containing 2 mL collagenase solution prepared in phosphate-buffered saline at pH 7.5.
2.5. Statistical Analysis
Data were expressed as mean ± standard deviation and 95% confidence interval. Differences among groups were analyzed using one-way analysis of variance followed by Tukey post-hoc testing. Separate analyses were performed for corneal and scleral samples. A p value < 0.05 was considered statistically significant.
3. Results
3.1. Corneal Enzymatic Degradation
Untreated human corneal buttons underwent complete collagenase-mediated degradation after 7 ± 2 days. Riboflavin/UVA crosslinking alone significantly prolonged degradation time to approximately 14 ± 3 days, corresponding to a two-fold increase in resistance to enzymatic degradation compared with untreated controls. Incubation with NAC/Pc alone resulted in a modest prolongation of corneal preservation, increasing degradation time by approximately 3 ± 2 days compared with control corneas; however, this difference did not reach statistical significance. No differences were found in scleral strips incubation with NAC/Pc alone. Similarly, pre-incubation with NAC/Pc before CXL demonstrated only a limited not statistically significant additive effect of approximately 2 additional days compared with CXL alone. The greatest anti-degradative effect was observed in tissues receiving NAC/Pc both before and after CXL treatment, in which corneal digestion time was further prolonged by approximately 7 days compared with CXL alone, reaching approximately 21 days overall.
One-way ANOVA demonstrated significant differences among corneal experimental groups (F = 31.04, p = 3.01 × 10⁻⁸). Post-hoc analysis confirmed that the major statistically significant differences were observed between untreated controls and crosslinked groups, and between CXL alone and combined NAC/polydatin pre/post-CXL treatment. Corneal degradation kinetics are illustrated in Figure 2 and Figure 3, and displayed in Table 1.
3.2. Scleral Enzymatic Degradation
Untreated scleral strips demonstrated greater intrinsic resistance to collagenase-mediated degradation compared with corneal tissue, with complete degradation occurring after approximately 16 ± 2 days. Riboflavin/UVA crosslinking alone significantly prolonged scleral preservation to approximately 24 ± 3 days. In contrast, incubation with NAC/polydatin alone did not produce significant differences in scleral degradation time compared with untreated controls. Additional incubation with NAC/Pc before CXL demonstrated only a mild additive effect, whereas combined NAC/Pc pre- and post-CXL treatment further prolonged scleral preservation by approximately 5 days compared with CXL alone, reaching approximately 29–30 days overall.
One-way ANOVA demonstrated significant differences among scleral experimental groups (F = 29.28, p = 4.77 × 10⁻⁸). Significant post-hoc differences were primarily observed between untreated controls and crosslinked tissues and between CXL alone and combined NAC/Pc pre/post-CXL treatment. Scleral degradation kinetics are shown in Figure 4 and Figure 5 and summarized in Table 1.
3.3. Protein Release Kinetics
Protein concentration within the incubation medium progressively increased during collagenase-mediated degradation and correlated with tissue dissolution and extracellular matrix breakdown. Untreated corneal samples demonstrated earlier and greater protein release into the incubation medium, consistent with accelerated collagen degradation. Crosslinked and adjunctively treated tissues showed delayed protein release kinetics, suggesting improved preservation of stromal architecture during collagenase incubation. The greatest difference in protein release was observed between untreated controls and the NAC/polydatin pre/post-CXL group, with an 84% reduction in protein concentration at day 5 (0.63 vs 0.10 µg/mL) and a 52% reduction at day 10 (1.23 vs 0.59 µg/mL). Protein release kinetics are illustrated in Figure 6.
Riboflavin/UVA CXL markedly prolonged tissue preservation in both corneal and scleral samples. NAC/polydatin incubation alone produced only a mild, non-significant prolongation of corneal degradation time and no significant effect in scleral strips compared with untreated controls. The greatest anti-degradative effect was observed after combined NAC/polydatin incubation before and after CXL. Values are expressed as mean ± standard deviation where applicable. NS = not significant; CXL = collagen crosslinking; NAC = N-acetylcysteine; Pc = Polygonum cuspidatum dry extract.
4. Discussion
The present study demonstrated that riboflavin/UVA collagen crosslinking (CXL) significantly enhances resistance of human corneal and scleral tissues to collagenase-mediated enzymatic degradation. Furthermore, adjunctive treatment with N-acetylcysteine (NAC) and Polydatin further potentiated this anti-degradative effect when combined with CXL, whereas NAC/Pc incubation alone produced only limited or non-significant effects. These findings support the concept that photochemical collagen stabilization represents the primary mechanism of corneal and scleral tissue preservation, while adjunctive biochemical modulation may enhance extracellular matrix stabilization when associated with crosslinking treatment.
The anti-enzymatic effect observed after corneal crosslinking is consistent with the experimental findings previously reported by Spoerl et al[5] and Aldahlawi et al[25] who demonstrated increased resistance of crosslinked corneal tissue to pepsin, trypsin, and collagenase digestion.[26] The present findings further expand this anti-degradative paradigm by demonstrating similar effects in human scleral tissue and by exploring adjunctive extracellular matrix-protective strategies based on NAC and polydatin treatment.
Untreated corneal buttons underwent complete degradation after approximately 7 days, whereas crosslinked corneas remained structurally preserved for approximately 14 days, confirming a two-fold increase in resistance following riboflavin/UVA treatment. The protective effect induced by crosslinking is likely multifactorial. Riboflavin/UVA irradiation generates reactive oxygen species leading to additional intermolecular and intramolecular collagen bonds within the extracellular matrix.[27,28] These structural modifications increase tissue stiffness and reduce accessibility of collagen cleavage sites to proteolytic enzymes.[29]
A major finding of the present study was the demonstration that adjunctive incubation with NAC[6] and Polydatin[10] further enhanced resistance to enzymatic degradation in both corneal and scleral samples. A protective effect was observed in samples receiving NAC/Pc treatment both before and after crosslinking, extending corneal degradation times up to approximately 21 days and scleral degradation times up to approximately 30 days.
N-acetylcysteine[7] is a well-known antioxidant and glutathione precursor with documented anti-inflammatory and antiproteolytic properties[8]. Similarly, Polydatin has demonstrated antioxidant and extracellular matrix-protective effects in several experimental models.[9] Although the precise molecular mechanisms were not specifically investigated in the present study, the observed prolongation of tissue preservation suggests a synergistic interaction between photochemical/photodynamic collagen stabilization and biochemical modulation of extracellular matrix degradation pathways.
Although the clinical role of corneal crosslinking is now well established in Keratoconus[2,4] and iatrogenic post laser vision correction corneal ectasia[30], supplemental extracellular matrix stabilization strategies may also have potential relevance in scleral remodeling associated with progressive myopia.[11] Progressive myopia is increasingly recognized as a disease characterized by collagen disorganization, protease activation, and biomechanical weakening of the scleral extracellular matrix, ultimately contributing to eye expansion and axial elongation.[12,13]
In the present study, untreated scleral strips demonstrated greater intrinsic resistance to collagenase-mediated degradation compared with corneal tissue, likely reflecting the denser collagen architecture and lower hydration of scleral connective tissue.[14] Nevertheless, Riboflavin/UVA crosslinking significantly prolonged scleral preservation agains enzymatic degradation, while adjunctive NAC/Polydatin exposure further enhanced resistance to extracellular matrix breakdown. These findings provide preliminary experimental evidence supporting the concept that combined biomechanical and biochemical stabilization strategies may reduce scleral ECM remodeling thus enhancing its preservation.
The protein release kinetics further supported the morphologic findings. Untreated corneal samples demonstrated earlier and greater protein release into the incubation medium, whereas crosslinked and adjunctively treated tissues showed delayed protein release kinetics, suggesting slower collagen breakdown and improved preservation of stromal ultrastructure.
Several limitations of the present study should be acknowledged. First, the study was performed in vitro and therefore cannot fully reproduce the complexity of in vivo ocular remodeling, inflammatory responses, and pharmacokinetics. Second, the collagenase digestion model represents an accelerated experimental system that may not directly reflect physiologic extracellular matrix turnover. Third, the precise contribution of direct collagenase inhibition versus tissue-level collagen stabilization induced by NAC and Polydatin was not specifically investigated.
Despite these limitations, the present study provides novel evidence that combined Riboflavin/UVA crosslinking increase scleral dygestion time and adjunctive NAC/Polydatin treatment significantly enhance resistance of human corneal and scleral tissues to collagenase-mediated degradation. These findings support further translational investigation of combined extracellular matrix stabilization strategies in corneal ectatic disorders and progressive myopia.
Interestingly, NAC/polydatin incubation alone produced only limited or non-significant effects, whereas its use in association with CXL, particularly in the pre/post-CXL protocol, further prolonged tissue preservation. This pattern suggests that the bioactive compounds may not independently induce substantial collagen stabilization, but may potentiate the anti-degradative effect of photochemical crosslinking in an exposure-dependent manner. Future studies testing different concentrations and incubation times are required to confirm a true dose–response relationship.
5. Conclusions
Riboflavin/UVA collagen crosslinking significantly enhances the resistance of human corneal and scleral tissues to collagenase-mediated enzymatic degradation. Adjunctive NAC and Polydatin treatment further potentiate this anti-degradative effect, suggesting a possible synergistic strategy for extracellular matrix stabilization. These findings support future translational investigation of combined biomechanical and biochemical approaches in corneal ectasia and progressive myopia, where scleral crosslinking may potentially emerge as a clinically relevant strategy for controlling disease progression. Future studies evaluating different NAC/Polydatin concentrations and incubation times are warranted to determine whether the observed protocol-dependent potentiation reflects a true dose–response relationship.
Author Contributions
Conceptualization, C.M. and G.S.; methodology, M.C.; software, F.C.; validation, M.N., M.Z. and A.M.; formal analysis, M.C. A.A.; investigation, M.C., G.S.; data curation, M.Z.; writing—original draft preparation, C.M.; writing—review and editing, A.M.; visualization, M.Z.; supervision, M.N.; project administration, C.M.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
“The study was approved by the Institutional Review Board of the Siena Crosslinking Center under the tenets of Helsinky declaration. Name of the Study: Sclero-Corneal Enzymatic Digestion Study 1, Code: SCED-S1, 02th April 2025.” The Study was conducted in collaboration with the Veneto Eye Bank Foundation, Venice, Italy
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The corneal tissues used in the study were provided by the Fondazione Banca degli Occhi del Veneto (Venice, Italy). These tissues were deemed unsuitable for transplantation and utilized in strict compliance with the ethical and regulatory guidelines governing the use of human tissues for scientific research purposes.
Acknowledgments
During the preparation of this work, the authors used ChatGPT (OpenAI) to assist with language editing, grammar correction, and text refinement. After using this tool, the authors carefully reviewed and edited the content as needed and take full responsibility for the content of the published article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
KC: Keratoconus; UVA: Ultraviolet – A; Pc: Poligonum cuspidatum; NAC: N-acetylcysteine: CXL: Crosslinking; *NS: not statistically significant
References
- Wollensak G, Spoerl E, Seiler T. Riboflavin/ultraviolet-A–induced collagen crosslinking for the treatment of keratoconus. Am J Ophthalmol. 2003;135(5):620–627.
- Hafezi F, Kling S, Hafezi NL, Aydemir ME, Lu NJ, Hillen M, Knyazer B, Awwad S, Mazzotta C, Kollros L, Torres-Netto EA. Corneal cross-linking. Prog Retin Eye Res. 2025 Jan;104:101322. Epub 2024 Dec 15. [CrossRef] [PubMed]
- Wollensak G, Spoerl E, Seiler T. Stress-strain measurements of human and porcine corneas after riboflavin–ultraviolet-A-induced cross-linking. J Cataract Refract Surg. 2003;29(9):1780–1785.
- Raiskup F, Herber R, Lenk J, Pillunat LE, Spoerl E. Crosslinking with UV-A and riboflavin in progressive keratoconus: From laboratory to clinical practice - Developments over 25 years. Prog Retin Eye Res. 2024 Sep;102:101276. Epub 2024 Jun 1. [CrossRef] [PubMed]
- Spoerl E, Wollensak G, Seiler T. Increased resistance of crosslinked cornea against enzymatic digestion. Curr Eye Res. 2004;29(1):35–40.
- Ramaesh T, Ramaesh K, Riley SC, West JD, Dhillon B. Effects of N-acetylcysteine on matrix metalloproteinase-9 secretion and cell migration of human corneal epithelial cells. Eye (Lond). 2012 Aug;26(8):1138-44.
- Aldini G, Altomare A, Baron G, Vistoli G, Carini M, Borsani L, et al. N-Acetylcysteine as an antioxidant and disulphide breaking agent: the reasons why. Free Radic Res. 2018;52(7):751–762.
- Tenório MCDS, Graciliano NG, Moura FA, Oliveira ACM, Goulart MOF. N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants (Basel). 2021 Jun 16;10(6):967.
- Du QH, Peng C, Zhang H. Polydatin: a review of pharmacology and pharmacokinetics. Pharm Biol. 2013;51(11):1347–1354.
- Meng Q, Guo T, Li G, Sun S, He S, Cheng B, et al. Polydatin protects against oxidative stress and extracellular matrix degradation in experimental models. Oxid Med Cell Longev. 2016;2016: 1–10.
- Sun Y, Sha Y, Yang J, Fu H, Hou X, Li Z, Xie Y, Wang G. Collagen is crucial target protein for scleral remodeling and biomechanical change in myopia progression and control. Heliyon. 2024 Jul 27;10(15):e35313.
- McBrien NA, Gentle A. Role of the sclera in the development and pathological complications of myopia. Prog Retin Eye Res. 2003;22(3):307–338.
- Rada JA, Shelton S, Norton TT. The sclera and myopia. Exp Eye Res. 2006;82(2):185–200.
- Atchison DA, Jones CE, Schmid KL, Pritchard N, Pope JM, Strugnell WE, Riley RA. Eye shape in emmetropia and myopia. Invest Ophthalmol Vis Sci. 2004 Oct;45(10):3380-6.
- Holden BA, Fricke TR, Wilson DA, Jong M, Naidoo KS, Sankaridurg P, Wong TY, Naduvilath TJ, Resnikoff S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology. 2016 May;123(5):1036-42.
- Wollensak G, Spoerl E. Collagen crosslinking of human and porcine sclera. J Cataract Refract Surg. 2004 Mar;30(3):689-95.
- Wollensak G, Iomdina E. Biomechanical and histological changes after collagen crosslinking of the sclera in rabbits. Acta Ophthalmol. 2009;87(2):193–198.
- Li X, Rabinowitz YS, Rasheed K, Yang H. Long-term biomechanical effects of riboflavin/UVA-induced scleral collagen cross-linking in experimental myopia. Invest Ophthalmol Vis Sci. 2004;45(12):4341–4346.
- Zhou X, Zhang S, Zhang Q, et al. Safety and efficacy of scleral collagen crosslinking for pathological myopia. Br J Ophthalmol. 2022;106(2):245–251.
- Rong S, Wang C, Han B, Feng P, Lan W, Gao Z, Li X, Chen W. Iontophoresis-assisted accelerated riboflavin/ultraviolet A scleral cross-linking: A potential treatment for pathologic myopia. Exp Eye Res. 2017 Sep; 162:37-47.
- Gawargious BA, Le A, Lesgart M, Ugradar S, Demer JL. Differential Regional Stiffening of Sclera by Collagen Cross-linking. Curr Eye Res. 2020 Jun;45(6):718-725.
- Huang Y, Gao R, Li Z, Chen A, Jiang Q, Ding S, Chen M, Meek KM, Wang Q, Chen Z, Huang J. The effects of different riboflavin concentrations and infiltration times on rabbit scleral crosslinking. Exp Eye Res. 2025 Aug;257: 110449.
- Mazzotta C, Raiskup F, Hafezi F, Torres-Netto EA, Armia Balamoun A, Giannaccare G, Bagaglia SA. Long term results of accelerated 9 mW corneal crosslinking for early progressive keratoconus: the Siena Eye-Cross Study 2. Eye Vis (Lond). 2021 May 1;8(1):16.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976 May 7; 72:248-54.
- Aldahlawi NH, Hayes S, O'Brart DP, Meek KM. Standard versus accelerated riboflavin-ultraviolet corneal collagen crosslinking: Resistance against enzymatic digestion. J Cataract Refract Surg. 2015 Sep;41(9):1989-96.
- Aldahlawi NH, Hayes S, O'Brart DP, Meek KM. Standard versus accelerated riboflavin-ultraviolet corneal collagen crosslinking: Resistance against enzymatic digestion. J Cataract Refract Surg. 2015 Sep;41(9):1989-96.
- Weadock KS, Miller EJ, Keuffel E, Dunn MG. Effect of physical crosslinking methods on collagen-fiber durability in proteolytic solutions. J Biomed Mater Res. 1996;32(2):221–226.
- Weadock KS, Miller EJ, Bellincampi LD, Zawadsky JP, Dunn MG. Physical crosslinking of collagen fibers: comparison of ultraviolet irradiation and dehydrothermal treatment. J Biomed Mater Res. 1995;29(11):1373–1379.
- Mallya SK, Mookhtiar KA, Van Wart HE. Kinetics of hydrolysis of type I, II, and III collagens by the class I and II Clostridium histolyticum collagenases. J Protein Chem. 1992;11(1):99–107.
- Mazzotta C, Gagliano C, Borroni D, De Lossada CR, Vinciguerra R, Zagari M, Alessio G, D'Oria F. Crosslinking for post refractive surgery ectasia: application and clinical outcomes. Graefes Arch Clin Exp Ophthalmol. 2025 Dec;263(12):3461-3470.
Figure 1.
Representative experimental macroscopic appearance images showing progressive collagenase-mediated degradation of human corneal buttons (white arrows) and scleral strips (yellow arrows) in the different treatment groups following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment. Corneal buttons and scleral strips study groups incubation (a): Co: Corneal buttons; Sc: Scleral strips; C = untreated controls (cornea, withe arrow; sclera yellow arrow; XL group = only crosslinked (9mW/5.4J/cm2); IN goup = only incubated specimens with N-acetylcysteine (NAC) plus Polygonum cuspidatum (Pc) dry extract; Pre CXL = pre-CXL NAC/Pc incubated and then crosslinked samples; CXL post = crosslinked and post-CXL icubated samples; Pre CXL Post = pre-CXL Incubated, then Crosslinked and post-CXL re-incubated. (b) intermediate degradation macroscopic observation showing delayed dissolution of corneal and scleral tissues after riboflavin/UVA collagen crosslinking, particularly in samples receiving adjunctive NAC/Pc treatment; (c) late incubation phase demonstrating persistence of structurally preserved corneal and scleral samples in the NAC/Pc pre/post-crosslinking group, whereas untreated tissues had already undergone advanced extracellular matrix degradation and dissolution. The macroscopic findings are consistent with the quantitative degradation kinetics and protein release measurements reported in the study, supporting the anti-degradative effect induced by riboflavin/UVA crosslinking alone both in cornea and sclera, with further digestion time extension after pre and post crosslinking adjunctive antiprotease bioactives.
Figure 1.
Representative experimental macroscopic appearance images showing progressive collagenase-mediated degradation of human corneal buttons (white arrows) and scleral strips (yellow arrows) in the different treatment groups following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment. Corneal buttons and scleral strips study groups incubation (a): Co: Corneal buttons; Sc: Scleral strips; C = untreated controls (cornea, withe arrow; sclera yellow arrow; XL group = only crosslinked (9mW/5.4J/cm2); IN goup = only incubated specimens with N-acetylcysteine (NAC) plus Polygonum cuspidatum (Pc) dry extract; Pre CXL = pre-CXL NAC/Pc incubated and then crosslinked samples; CXL post = crosslinked and post-CXL icubated samples; Pre CXL Post = pre-CXL Incubated, then Crosslinked and post-CXL re-incubated. (b) intermediate degradation macroscopic observation showing delayed dissolution of corneal and scleral tissues after riboflavin/UVA collagen crosslinking, particularly in samples receiving adjunctive NAC/Pc treatment; (c) late incubation phase demonstrating persistence of structurally preserved corneal and scleral samples in the NAC/Pc pre/post-crosslinking group, whereas untreated tissues had already undergone advanced extracellular matrix degradation and dissolution. The macroscopic findings are consistent with the quantitative degradation kinetics and protein release measurements reported in the study, supporting the anti-degradative effect induced by riboflavin/UVA crosslinking alone both in cornea and sclera, with further digestion time extension after pre and post crosslinking adjunctive antiprotease bioactives.

Figure 2.
Resistance of human corneal buttons to collagenase-mediated degradation following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment. Mean time to complete collagenase-mediated degradation of human corneal buttons in the six experimental groups. Riboflavin/UVA collagen crosslinking significantly prolonged corneal preservation compared with untreated controls. NAC/Pc incubation alone demonstrated only a mild increase in degradation time, whereas combined NAC/Pc treatment before and after CXL produced the greatest anti-degradative effect. Individual sample values are displayed. Error bars represent standard deviation. CXL = collagen crosslinking; NAC = N-acetylcysteine; Pc = Polygonum cuspidatum (Polidatin) dry extract.
Figure 2.
Resistance of human corneal buttons to collagenase-mediated degradation following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment. Mean time to complete collagenase-mediated degradation of human corneal buttons in the six experimental groups. Riboflavin/UVA collagen crosslinking significantly prolonged corneal preservation compared with untreated controls. NAC/Pc incubation alone demonstrated only a mild increase in degradation time, whereas combined NAC/Pc treatment before and after CXL produced the greatest anti-degradative effect. Individual sample values are displayed. Error bars represent standard deviation. CXL = collagen crosslinking; NAC = N-acetylcysteine; Pc = Polygonum cuspidatum (Polidatin) dry extract.

Figure 3.
Corneal degradation kinetics during collagenase incubation. Time-course curves showing progressive reduction in residual corneal button diameter during collagenase-mediated degradation. Untreated controls demonstrated rapid extracellular matrix breakdown, whereas riboflavin/UVA crosslinked tissues exhibited delayed degradation kinetics. NAC/Pc incubation alone showed only a limited effect, while combined NAC/Pc pre/post-CXL treatment produced the greatest prolongation of corneal preservation (F = 31.04, p = 3.01 × 10⁻⁸).
Figure 3.
Corneal degradation kinetics during collagenase incubation. Time-course curves showing progressive reduction in residual corneal button diameter during collagenase-mediated degradation. Untreated controls demonstrated rapid extracellular matrix breakdown, whereas riboflavin/UVA crosslinked tissues exhibited delayed degradation kinetics. NAC/Pc incubation alone showed only a limited effect, while combined NAC/Pc pre/post-CXL treatment produced the greatest prolongation of corneal preservation (F = 31.04, p = 3.01 × 10⁻⁸).

Figure 4.
Resistance of human Scleral strips to collagenase-mediated degradation following riboflavin/UVA crosslinking and adjunctive NAC/Polydatin treatment. Mean time to complete collagenase-mediated degradation of human scleral strips in the six experimental groups. Untreated scleral tissue demonstrated greater intrinsic resistance to collagenase degradation compared with corneal tissue. Riboflavin/UVA crosslinking significantly prolonged scleral preservation, whereas NAC/Pc incubation alone did not produce substantial differences compared with untreated controls. The maximal anti-degradative effect was observed in the combined NAC/Pc (Polidatin) pre/post-CXL group. Error bars represent standard deviation.
Figure 4.
Resistance of human Scleral strips to collagenase-mediated degradation following riboflavin/UVA crosslinking and adjunctive NAC/Polydatin treatment. Mean time to complete collagenase-mediated degradation of human scleral strips in the six experimental groups. Untreated scleral tissue demonstrated greater intrinsic resistance to collagenase degradation compared with corneal tissue. Riboflavin/UVA crosslinking significantly prolonged scleral preservation, whereas NAC/Pc incubation alone did not produce substantial differences compared with untreated controls. The maximal anti-degradative effect was observed in the combined NAC/Pc (Polidatin) pre/post-CXL group. Error bars represent standard deviation.

Figure 5.
Scleral degradation kinetics during collagenase incubation. Time-course curves illustrating progressive reduction in residual scleral strip length during collagenase-mediated degradation. Scleral tissue demonstrated slower degradation kinetics compared with corneal tissue. Riboflavin/UVA crosslinking significantly prolonged tissue preservation, while adjunctive pre and post-CXL NAC/Pc (Polidatin) incubation, further enhanced resistance (F = 29.28, p = 4.77 × 10⁻⁸).
Figure 5.
Scleral degradation kinetics during collagenase incubation. Time-course curves illustrating progressive reduction in residual scleral strip length during collagenase-mediated degradation. Scleral tissue demonstrated slower degradation kinetics compared with corneal tissue. Riboflavin/UVA crosslinking significantly prolonged tissue preservation, while adjunctive pre and post-CXL NAC/Pc (Polidatin) incubation, further enhanced resistance (F = 29.28, p = 4.77 × 10⁻⁸).

Figure 6.
Protein release kinetics during collagenase-mediated degradation of human corneal tissue following riboflavin/UVA crosslinking and adjunctive NAC/polydatin treatment. Time-dependent protein concentration changes in the incubation medium measured using Bradford’s method during collagenase-mediated degradation of human corneal samples. Untreated control corneas demonstrated earlier and greater protein release, consistent with accelerated extracellular matrix degradation and collagen breakdown. Crosslinked and adjunctively treated tissues showed delayed protein release kinetics, suggesting improved preservation of stromal collagen architecture. The greatest reduction in protein release was observed in the NAC/Pc pre- and post-crosslinking group, showing an 84% lower protein concentration compared with controls at day 5 (0.10 vs 0.63 µg/mL) and a 52% lower concentration at day 10 (0.59 vs 1.23 µg/mL). Error bars represent standard deviation. CXL = collagen crosslinking; NAC = N-acetylcysteine; Pc = Polygonum cuspidatum (Polidatin) dry extract.
Figure 6.
Protein release kinetics during collagenase-mediated degradation of human corneal tissue following riboflavin/UVA crosslinking and adjunctive NAC/polydatin treatment. Time-dependent protein concentration changes in the incubation medium measured using Bradford’s method during collagenase-mediated degradation of human corneal samples. Untreated control corneas demonstrated earlier and greater protein release, consistent with accelerated extracellular matrix degradation and collagen breakdown. Crosslinked and adjunctively treated tissues showed delayed protein release kinetics, suggesting improved preservation of stromal collagen architecture. The greatest reduction in protein release was observed in the NAC/Pc pre- and post-crosslinking group, showing an 84% lower protein concentration compared with controls at day 5 (0.10 vs 0.63 µg/mL) and a 52% lower concentration at day 10 (0.59 vs 1.23 µg/mL). Error bars represent standard deviation. CXL = collagen crosslinking; NAC = N-acetylcysteine; Pc = Polygonum cuspidatum (Polidatin) dry extract.


Table 1.
Collagenase-mediated degradation times of human corneal buttons and scleral strips following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment.
Table 1.
Collagenase-mediated degradation times of human corneal buttons and scleral strips following riboflavin/UVA crosslinking and adjunctive NAC/Pc treatment.
| Group | Corneal Degradation Time | Scleral Degradation Time | |
| A Untreated controls | 7 ± 2 days | 16 ± 2 days | |
| B Riboflavin/UVA CXL alone | 14 ± 3 days | 24 ± 3 days | |
| C NAC/Pc incubation alone | 10 ± 2 days *NS | 17 ± 2 days *NS | |
| D NAC/Pc incubation pre CXL | ~16 ± 2 days | ~25 ± 2 days | |
| E NAC/Pc incubation post CXL | ~17 ± 2 days | ~26 ± 2 days | |
| F NAC/Pc incubation pre and post CXL | 21 ± 2 days | 29–30 ± 2 days |
1Pc: Poligonum cuspidatum. 2NAC: N-acetylcysteine: 3CXL: Crosslinking: *NS: not statistically significant vs control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



