Submitted:
12 June 2026
Posted:
15 June 2026
You are already at the latest version
Abstract
Mucosal surfaces constitute the principal immunological interface between mammals and their environments, continuously integrating dietary antigens, the commensal microbiota, and exposure to pathogens. These barrier sites are organized into compartmentalized immune networks that coordinate epithelial surveillance, antigen sampling, immunoglobulin A (IgA) diversification, and tissue-resident immune responses. However, prevailing paradigms in mucosal immunology are derived predominantly from murine and human systems and often assume broad conservation of mucosal immune organization across mammals. Emerging comparative evidence challenges this assumption as ruminants, monogastrics, carnivores, and reservoir wildlife species exhibit substantial divergence in mucosal architecture, Peyer’s patch ontogeny, epithelial sensing pathways, and immune–microbiota interactions, reflecting adaptation to distinct ecological and dietary pressures
We propose that mucosal immunity serves as an ecological filter shaping microbial persistence, pathogen shedding, and zoonotic transmission at barrier sites where pathogen emergence initiates. Failure to account for species-specific mucosal immune organization may limit cross-species extrapolation of vaccine responses, microbiome-targeted therapeutics, and zoonotic risk prediction. We further argue that variation in mucosal compartmentalization contributes directly to reservoir competence by influencing immune tolerance, microbial filtering, IgA repertoire diversification, and pathogen persistence within host populations. Finally, we identify major unresolved questions and highlight the need for comparative spatial immunology and integrated mucosal multi-omics to develop predictive models of host–pathogen interaction within a One Health framework.
Keywords:
mucosal immunology
; barrier immunity
; zoonotic transmission
; host-microbiota interactions
; One Health
; reservoir competence
1. Introduction
Mucosal surfaces of the gastrointestinal, respiratory, and urogenital tracts represent the principal immunological interface between mammals and their environments. Hence, they function continuously in integrating signals from commensal microbiota, dietary antigens, and invading pathogens across highly specialized barrier tissues (Peterson & Artis, 2014; Zheng et al., 2020). However, rather than functioning singly as passive physical barriers, these surfaces serve as dynamic immunological ecosystems in which epithelial cells, stromal populations, resident leukocytes, and organized lymphoid tissues collectively maintain tissue homeostasis and coordinate organismal protective immune responses (Belkaid & Hand, 2014; Krausgruber et al., 2020).
Central to this organization is the concept of mucosal immune compartmentalization, whereby anatomically distinct mucosal sites develop specialized inductive, and effector immune programs shaped by local antigenic exposure, microbial ecology, and tissue-specific regulatory networks. Within the intestine, in some animals, Peyer’s patches function as major inductive sites that coordinate antigen sampling through specialized microfold (M) cells and initiate mucosal B-cell responses that culminate in secretory immunoglobulin A (SIgA) production (Reboldi & Cyster, 2016; Mabbott et al., 2013; Mabbott & Hase, 2024). The produced SIgA, a dominant antibody isotype at mucosal surfaces, plays a central role in microbial immune exclusion, bacterial coating, and maintenance of host–microbiota homeostasis (Brandtzaeg, 2013; Pietrzak et al., 2020). Additional evidence demonstrates concordance among microbiota-derived signals regulating immune cell trafficking, lymphoid organization, and tissue-resident immune specialization within mucosal compartments, emphasizing the reciprocal relationship between mucosal immunity and microbial ecology (Zheng et al., 2020; Kabbert et al., 2025).
Importantly, studies have shown that compartmentalized immune programs are not uniformly conserved across mammalian species but are shaped by evolutionary history, ecological niche, diet, microbial exposure, and host physiology (Youngblut et al., 2019; Kuthyar & Reese, 2021). Current paradigms in mucosal immunology are largely based on murine and human models; however, accumulating comparative evidence indicates that these models inadequately capture the diversity of mucosal immune organization across mammalian lineages (Beura et al., 2016; Gray & Farber, 2022). Ruminants, for example, possess an ileal Peyer’s patch that functions as a primary lymphoid organ, supporting antigen-independent B-cell repertoire diversification during fetal development (Yasuda et al., 2006). This feature is absent in conventional murine systems and has major implications for pre-immune mucosal immunity (Farukawa et al., 2021). Similarly, laboratory mice maintained under specific pathogen-free conditions have altered microbial exposure histories and immune developmental trajectories compared with wild mammals, limiting the ecological and translational relevance of many experimentally derived mucosal immune models (Abolins et al., 2017; Rosshart et al., 2017; Yeung et al., 2020). These limitations, therefore, represent not only technical constraints, but also broader conceptual gaps in understanding how mammalian mucosal immune systems have adapted to distinct ecological pressures and host–microbiota relationships across species.
Within a One Health framework, most zoonotic emergence occurs at the intersection of animal, human, and environmental health, making this gap significant. About 60% of emerging infectious diseases originate in animal reservoirs, with mucosal surfaces as primary sites for pathogen acquisition, replication, and transmission (Jones et al., 2008; Rahman et al., 2020). Consequently, species-specific differences in barrier immunity, tissue-resident immune organization, microbial ecology, and mucosal tolerance mechanisms are likely to influence reservoir competence, pathogen persistence, and shedding dynamics (Zhang et al., 2025). Bats, for instance, serve as reservoir hosts for numerous pandemic-capable RNA viruses, including coronaviruses and filoviruses, yet the organization of their mucosal immune systems remains poorly characterized despite increasing evidence for unique innate immune regulatory adaptations (Pavlovich et al., 2018; Zhang et al., 2013). Similarly, in livestock species, gastrointestinal mucosal surfaces are a major reservoir for enteric zoonotic pathogens, including Salmonella enterica, Campylobacter jejuni, and enterohemorrhagic Escherichia coli, yet the comparative mucosal determinants governing colonization and pathogen shedding remain incompletely understood (Scallan et al., 2011; Kempf et al., 2022).
This review consolidates comparative evidence across mammalian systems to argue that mucosal immune compartmentalization is an evolutionarily and ecologically calibrated feature of host defense (Zhou et al., 2025). We examine the structural and functional diversity of mucosal immune organization across mammalian lineages, the role of host–microbiota co-evolution in calibrating tissue-specific immune states, and the implications of these differences for understanding pathogen emergence and cross-species transmission. Finally, we highlight major knowledge gaps and discuss how comparative spatial immunology, single-cell mucosal profiling, and integrated multi-omics approaches may help establish predictive frameworks for host-pathogen interactions within a One Health context.
2. Structural and Functional Organization of Mammalian Mucosal Immunity
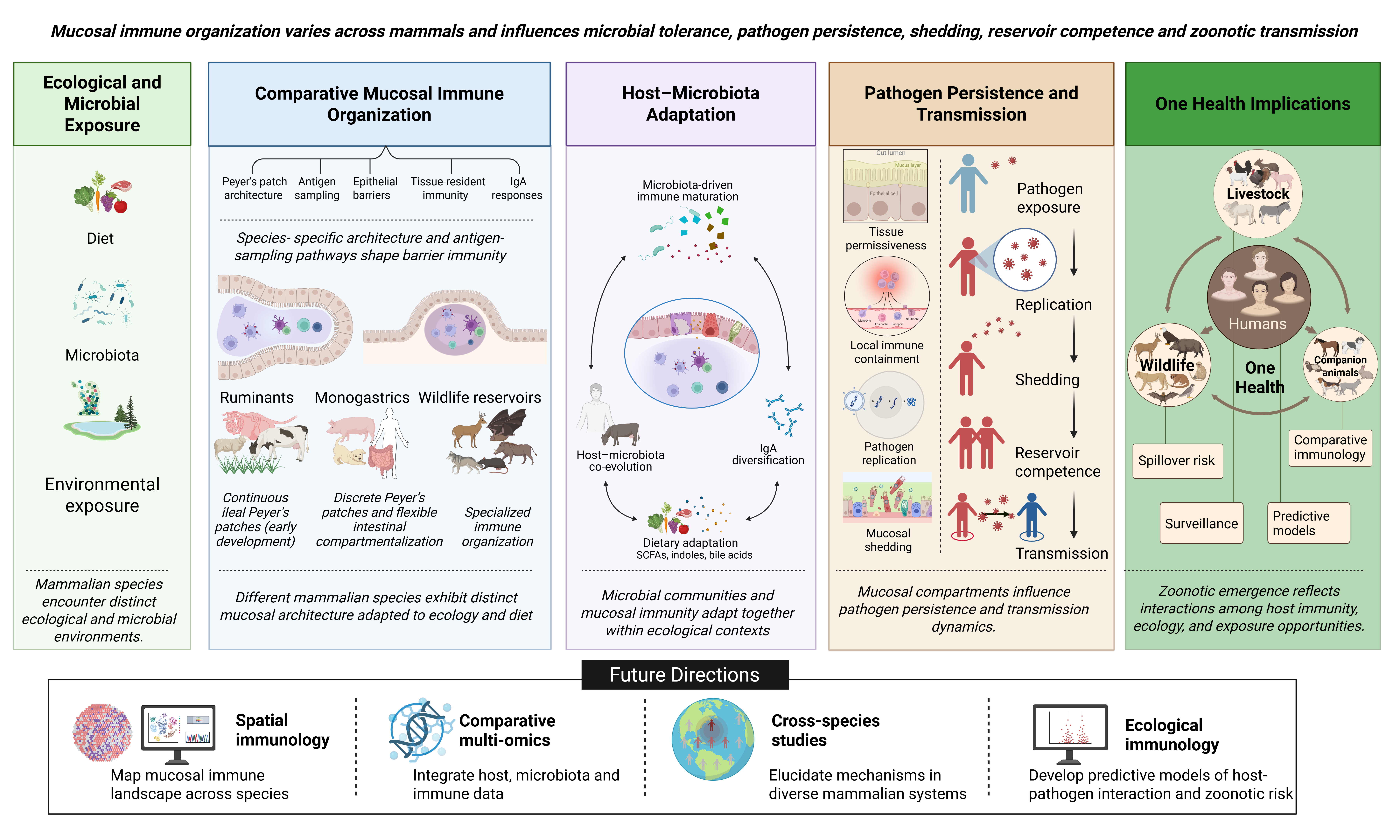
Mucosal immune systems are compartmentalized across barrier tissues, enabling localized adaptation to distinct microbial, dietary, and environmental pressures encountered at gastrointestinal, respiratory, and other mucosal surfaces (Peterson & Artis, 2014; Zheng et al., 2020; Figure 1). Collectively, these tissues form the mucosa-associated lymphoid tissue (MALT) system, including gut-associated lymphoid tissue (GALT), nasal-associated lymphoid tissue (NALT), and bronchus-associated lymphoid tissue (BALT), which function as specialized sites for antigen surveillance, lymphocyte activation, and regulation of barrier homeostasis (Mabbott et al., 2013; Reboldi & Cyster, 2016; Silva-Sanchez & Randall, 2020). Although these compartments share conserved immunological functions, their structural organization, immune cell composition, and developmental regulation differ substantially across anatomical sites and mammalian species, reflecting adaptation to distinct ecological niches and patterns of host–microbe interaction (Arroyo Portilla et al., 2021; Gray & Farber, 2022). Increasing evidence further demonstrates active regulation of compartment-specific immune responses through localized signaling networks that shape tissue-resident immunity, inflammatory responsiveness, and microbial tolerance at barrier surfaces (Krausgruber et al., 2020; Zheng et al., 2020). Consequently, understanding the comparative organization of mucosal immune tissues is essential for interpreting species-specific immune function and for identifying how barrier immune architecture may influence pathogen susceptibility, microbial colonization, and zoonotic transmission dynamics across mammals.
2.1. Comparative Organization of Mucosal Immune Tissues
Mucosal lymphoid tissues exhibit substantial anatomical and functional heterogeneity across barrier sites and mammalian species, reflecting adaptation to distinct physiological demands, microbial landscapes, and antigenic pressures encountered at mucosal surfaces (Zheng et al., 2020; Zhou et al., 2025). Within the gastrointestinal tract, GALT constitutes the most extensively developed mucosal immune compartment and includes Peyer’s patches, isolated lymphoid follicles, mesenteric lymph nodes, and dispersed immune populations throughout the lamina propria and epithelial layers (Mabbott et al., 2013; Reboldi & Cyster, 2016). Among these structures, Peyer’s patches serve as major inductive sites for antigen sampling, germinal center formation, IgA class-switch recombination, and the generation of tissue-adapted lymphocyte populations that maintain intestinal immune homeostasis (Reboldi & Cyster, 2016; Ullrich et al., 2021; Kabbert et al., 2025). The intestinal mucosa further contains specialized macrophages, dendritic cells, innate lymphoid cells, and intraepithelial lymphocytes that collectively regulate epithelial integrity, microbial tolerance, and inflammatory responsiveness under conditions of continuous antigen exposure (Zheng et al., 2020; Arroyo Portilla et al., 2021). In parallel, epithelial and stromal cell networks actively shape compartment-specific immunity through localized cytokine production, metabolic regulation, and microenvironmental signaling pathways (Peterson & Artis, 2014; Krausgruber et al., 2020).
In contrast to the densely colonized intestinal environment, respiratory mucosal tissues are adapted to continual exposure to inhaled particulates and airborne pathogens while maintaining the integrity of highly delicate gas-exchange surfaces. NALT functions as a major inductive site for upper airway immune surveillance, whereas BALT may occur constitutively or emerge as inducible lymphoid aggregates following infection or chronic inflammatory stimulation (Allie & Randall, 2017; Seefeld et al., 2024). Compared with GALT, respiratory mucosal compartments generally harbor lower microbial densities and rely on rapid antiviral responses, coupled with tightly regulated inflammatory mechanisms, to limit tissue damage and preserve pulmonary function (Allie & Randall, 2017; Gray & Farber, 2022).
Comparative analyses demonstrate that the organization and functional specialization of mucosal immune tissues vary considerably across mammalian lineages (Table 1). Ruminants possess extensive ileal Peyer’s patches that function as primary lymphoid organs during early development and support antigen-independent B-cell diversification, whereas rodents rely predominantly on bone marrow-mediated B-cell maturation pathways (Yasuda et al., 2006). Similarly, constitutive BALT is prominent in several livestock and wildlife species but remains limited in healthy adult humans and laboratory mice, where inducible forms are more commonly associated with pulmonary inflammation (Allie & Randall, 2017). Additional interspecies variation in epithelial organization, microbiota composition, lymphoid tissue distribution, and tissue-resident immune populations further reflects adaptation to species-specific diets, ecological niches, and host–microbe interactions (Arroyo Portilla et al., 2021; Zheng et al., 2020). Collectively, these observations demonstrate that mucosal immune organization is evolutionarily shaped rather than universally conserved across mammals, underscoring the importance of comparative mucosal immunology for understanding species-specific immune function, pathogen susceptibility, and barrier-associated determinants of zoonotic transmission within a One Health framework (Table 1).
2.2. Peyer’s Patches and Antigen Sampling Mechanisms
Peyer’s patches (PPs) are highly organized lymphoid structures that function as major inductive sites for intestinal immunity, coordinating adaptive immune responses against commensal microbiota, dietary antigens, and invading pathogens. As secondary lymphoid organs, Peyer’s patches develop during gestation through tightly regulated lymphoid organogenesis and serve as sites where naïve lymphocytes encounter cognate antigens within the intestinal mucosa (Reboldi & Cyster, 2016; Kabbert et al., 2025). Owing to continuous exposure to luminal microorganisms and environmental antigens, Peyer’s patches maintain persistent germinal center activity that supports B-cell proliferation, affinity maturation, and clonal selection within structurally organized dark and light zones (Ullrich et al., 2021). This highly coordinated organization enables the intestinal immune system to balance tolerance toward harmless luminal antigens while simultaneously generating protective responses against enteric pathogens, thereby preserving epithelial and immunological homeostasis under conditions of constant antigenic stimulation (Reboldi & Cyster, 2016; Zheng et al., 2020).
Structurally, Peyer’s patches consist of B-cell follicles, germinal centers, T-cell zones, and specialized follicle-associated epithelium (FAE) that overlies the lymphoid tissue. The FAE contains microfold (M) cells, specialized epithelial cells that mediate transcytosis of luminal antigens and microorganisms from the intestinal lumen to underlying antigen-presenting cells, including dendritic cells and macrophages (Mabbott et al., 2013). In contrast to conventional enterocytes, M cells possess reduced microvilli, a thinner glycocalyx, and diminished mucus coverage, which are structural adaptations that facilitate efficient antigen uptake and transport across the epithelial barrier (Mabbott et al., 2013; Rios et al., 2016). Following antigen capture, dendritic cells process and present antigens to naïve T-lymphocytes, thereby promoting B-cell activation, class-switch recombination, and differentiation into IgA-secreting plasma cells within the germinal centers of gut-associated lymphoid tissue (Brandtzaeg, 2013; Pietrzak et al., 2020). Secretory IgA subsequently contributes to immune exclusion by limiting microbial adherence, neutralizing pathogens, and restricting excessive inflammatory responses while maintaining tolerance toward commensal microbiota and dietary antigens (Brandtzaeg, 2013; Pietrzak et al., 2020).
Antigen sampling represents a central mechanism through which intestinal immune tissues continuously monitor luminal contents while preserving barrier integrity. In addition to M-cell-mediated transcytosis, dendritic cells and macrophages may extend transepithelial dendrites between epithelial cells to directly capture luminal antigens (Rios et al., 2016). These coordinated antigen-sampling pathways facilitate rapid immune recognition and communication between epithelial barriers and underlying immune compartments, supporting both protective immunity and mucosal tolerance (Mabbott & Hase, 2024). Recent evidence further demonstrates that microbiota-derived signals actively regulate lymphocyte trafficking, germinal center dynamics, and immune organization within Peyer’s patches, highlighting the reciprocal relationship between intestinal microbial ecology and mucosal immune architecture (Kabbert et al., 2025; Zheng et al., 2020). Collectively, Peyer’s patches function as highly specialized immunological interfaces that integrate antigen surveillance, adaptive immune induction, and microbiota-responsive regulation to maintain intestinal homeostasis at the host–microbe interface.
2.3. Tissue-Resident and Compartment-Specific Immunity
Mucosal immune compartments exhibit substantial functional specialization across anatomical sites, enabling localized adaptation to the distinct microbial, dietary, and environmental pressures encountered at barrier surfaces (Peterson & Artis, 2014; Zheng et al., 2020). Resident immune populations are shaped by local epithelial, stromal, and microbial microenvironments and acquire tissue-specific functional programs that coordinate immune surveillance, pathogen clearance, tissue repair, and preservation of barrier integrity (Krausgruber et al., 2020; Arroyo Portilla et al., 2021). Such regional specialization is particularly important at mucosal interfaces, where continuous exposure to commensal microorganisms, dietary antigens, and environmental particulates requires tight regulation of inflammatory responses to maintain tissue homeostasis while preserving effective antimicrobial defense (Peterson & Artis, 2014; Zheng et al., 2020).
Tissue-resident memory T (TRM) cells represent a specialized lineage of noncirculating memory T-lymphocytes that establish long-term residence within epithelial and subepithelial compartments of mucosal tissues (Gray & Farber, 2022). Unlike circulating memory T-cells, TRM populations remain strategically positioned at barrier surfaces, where they provide rapid localized protection following recurrent microbial exposure and contribute to epithelial immune surveillance and barrier maintenance (Marchesini et al., 2024; Xie et al., 2025). Within intestinal, respiratory, and oral mucosal tissues, TRM cells interact closely with epithelial cells, dendritic cells, macrophages, and stromal populations to coordinate localized immune defense programs and rapid antimicrobial responses (Krausgruber et al., 2020; Gray & Farber, 2022). Intraepithelial lymphocytes (IELs) further support the elimination of infected or damaged epithelial cells while contributing to tissue repair and immune regulation. Although these resident immune populations are essential for frontline defense, persistent activation of localized immune programs may also contribute to chronic inflammatory pathology under dysregulated conditions (Tian et al., 2025).
Innate lymphoid cells (ILCs) similarly represent key components of tissue-resident immunity and contribute to epithelial maintenance, cytokine-mediated regulation, and early antimicrobial defense at mucosal barriers (Peterson & Artis, 2014). ILC populations exhibit pronounced tissue-specific specialization and respond rapidly to local cytokine and microbial signals, thereby functioning as central regulators of regional immune homeostasis (Zheng et al., 2020). In the intestinal mucosa, ILCs support epithelial regeneration, microbial tolerance, and maintenance of barrier integrity during continuous antigen exposure, whereas respiratory ILC populations participate more prominently in rapid inflammatory and antiviral responses following inhalation of pathogens and environmental particulates (Allie & Randall, 2017; Zheng et al., 2020). Increasing evidence further demonstrates that coordinated interactions among tissue-resident lymphocytes, epithelial cells, stromal populations, and local microbiota collectively shape distinct immunological landscapes across mucosal compartments (Krausgruber et al., 2020; Zheng et al., 2020).
Functional specialization is particularly evident across intestinal, respiratory, and oral mucosal tissues. The intestinal mucosa is primarily adapted to maintain tolerance toward dense commensal microbial communities and dietary antigens through coordinated activity of epithelial barriers, secretory IgA, antimicrobial peptides, and regulatory immune populations (Brandtzaeg, 2013; Peterson & Artis, 2014; Pietrzak et al., 2020). In contrast, respiratory mucosal tissues are optimized for rapid pathogen detection and tightly regulated inflammatory responsiveness because of continual exposure to airborne microorganisms and environmental particulates (Allie & Randall, 2017). Oral mucosal tissues similarly possess specialized immune programs adapted to constant polymicrobial exposure and mechanical stress within highly colonized epithelial environments. Collectively, these compartmentalized immune adaptations underscore the remarkable plasticity of mucosal immunity across mammals and highlight how tissue-specific immune organization influences microbial colonization, pathogen persistence, and susceptibility to zoonotic transmission (Krausgruber et al., 2020; Zheng et al., 2020; Arroyo Portilla et al., 2021; Table 1).
3. Ecological and Microbial Shaping of Mucosal Immunity
The mucosal surface hosts dense, metabolically active microbial communities essential for health, supplying nutrients, processing complex dietary polysaccharides, and supporting robust immune function and neurodevelopment (Beurel, 2024; Luo et al., 2024). In mammalian species, microbiota are not merely passive entities; they actively instruct both innate and adaptive immunity through metabolites (such as SCFAs and indole-3-lactic acid) and signaling, facilitating immune cell differentiation, regulatory T-cell induction, and IgA synthesis (Wang et al., 2025; Groen et al., 2026; Figure 1). The reciprocal relationship is especially evident in the gastrointestinal tract, where microbial abundance, nutritional complexity, and environmental exposure exert species-specific selective pressures on mucosal immune structure (Beurel, 2024; Wang et al., 2025). Comparative evidence increasingly suggests that microbiota-immune interactions are not dictated by a universal mammalian model but rather demonstrate ecological adaptation to host nutrition, digestive physiology, and environmental context (Ullah et al., 2024; Wang et al., 2025; Zhao et al., 2026). Recognizing this diversity is critical for understanding interspecies variation in pathogen susceptibility, microbial persistence, and zoonotic transmission (Zhao et al., 2026).
3.1. Microbiota-Driven Immune Maturation
Early-life microbial colonization plays a fundamental role in the establishment and maturation of mucosal immunity. Although the mammalian intrauterine environment was traditionally considered sterile, recent studies detecting microbial DNA within the placenta, amniotic fluid, umbilical cord, and meconium have challenged this long-standing paradigm and suggest that host–microbiota interactions may begin before birth (Groen et al., 2026). Following birth, progressive microbial colonization provides essential developmental signals that regulate epithelial differentiation, lymphoid tissue organization, and immune education at mucosal surfaces (Pirker & Vogl, 2024; Groen et al., 2026). Germ-free mammalian models consistently exhibit profound deficiencies in Peyer’s patch maturation, isolated lymphoid follicle development, secretory IgA production, and T-cell compartmentalization, underscoring the indispensable role of commensal microorganisms in shaping barrier immunity (Iliev et al., 2025). Microbial-associated molecular patterns (MAMPs) engage epithelial and immune pattern-recognition receptors, including Toll-like receptors (TLRs) and NOD-like receptors, to stimulate epithelial turnover, antimicrobial peptide secretion, and development of organized mucosal lymphoid structures (Groen et al., 2026; Iliev et al., 2025).
Comparative evidence further demonstrates that mammalian species differ substantially in how microbiota direct immune maturation and compartmentalization (Iliev et al., 2025). In monogastric species such as rodents and humans, early-life microbial exposure strongly influences immune programming in the distal small intestine, where segmented filamentous bacteria and Clostridial taxa promote the differentiation of T helper 17 (Th17) cells and regulatory T-cell populations (Groen et al., 2026). In contrast, microbial colonization in ruminants is closely associated with foregut fermentation systems, in which the rumen microbiome exerts major immunological and metabolic influences prior to the stabilization of distal intestinal microbial communities (Li et al., 2023; Luo et al., 2024). These observations suggest that immune education in ruminants may occur across anatomically distinct mucosal compartments, including the rumen epithelium, rather than being predominantly concentrated in distal intestinal tissues, as is commonly described in murine models (Li et al., 2023; Tardiolo et al., 2025).
3.2. IgA Diversification and Microbial Selection
Secretory immunoglobulin A (SIgA) represents one of the principal mechanisms through which mucosal immune systems regulate microbial ecology at barrier surfaces. Produced predominantly by plasma cells within mucosal lymphoid tissues and transported across epithelial barriers via the polymeric immunoglobulin receptor (pIgR), SIgA functions as a central mediator of immune exclusion while simultaneously preserving symbiotic host–microbiota relationships (León & Francino, 2022; Siniscalco et al., 2024). Through coating luminal microorganisms, SIgA spatially restricts epithelial adherence, interferes with virulence-associated mechanisms, dampens excessive inflammatory stimulation, and facilitates clearance of potentially invasive organisms from the gastrointestinal tract (León & Francino, 2022; Gupta et al., 2023). Consequently, SIgA functions not only as an immune effector molecule but also as an ecological selector that shapes microbial niche occupancy, regulates community composition, and contributes to long-term maintenance of mucosal homeostasis (Pinheiro et al., 2023; Zhang et al., 2023; Siniscalco et al., 2024). Comparative evidence further demonstrates substantial species-level variation in IgA induction pathways, repertoire diversification, and functional deployment across mammals (Pinheiro et al., 2023; Zhang et al., 2023; Siniscalco et al., 2024; Figure 1). In many monogastric species, IgA diversification depends largely on organized germinal center activity within Peyer’s patches and mesenteric lymphoid tissues, where both T-cell-dependent and T-cell-independent pathways generate antigen-adapted repertoires directed against commensal and pathogenic microorganisms (Carreto-Binaghi et al., 2024). In contrast, ruminants possess continuous ileal Peyer’s patches that function as primary lymphoid organs during early development and generate extensive B-cell diversity through mechanisms that may favor broad repertoire formation over highly antigen-specific selection (Sarma et al., 2023). These developmental differences shape mucosal microbial landscapes across mammals, affecting filter efficiency, tolerance, pathogen control, and the risk of dysbiosis. These findings emphasize SIgA as an immunological and ecological regulator linked to species-specific microbes and host pressures (Sarma et al., 2023; Carreto-Binaghi et al., 2024; Siniscalco et al., 2024).
3.3. Dietary Ecology and Environmental Antigen Exposure
Dietary ecology and environmental antigen exposure exert major influences on microbiota composition and mucosal immune specialization across mammalian species (Tardiolo et al., 2025). Because ruminants and monogastric mammals possess fundamentally distinct digestive physiologies and microbial ecosystems, their mucosal immune systems encounter markedly different metabolic and antigenic environments (Colella et al., 2023; Zulfiqar et al., 2025). In ruminants, microbial fermentation of dietary fiber generates large quantities of short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate, which function not only as major metabolic substrates but also as immunoregulatory signaling molecules that influence epithelial integrity, inflammatory tone, and barrier homeostasis (Zulfiqar et al., 2025). Continuous exposure to dense microbial fermentation products within foregut compartments is therefore associated with tolerogenic immune programming that limits excessive inflammation within highly colonized microbial environments (Almansour et al., 2025; Kim et al., 2025).
In contrast, monogastric omnivores such as humans and pigs experience greater dietary heterogeneity and more variable antigen exposure, necessitating increased compartmental flexibility to discriminate among harmless dietary antigens, commensal microorganisms, and invading pathogens (Colella et al., 2023; Reichel et al., 2026). These ecological differences directly influence the balance between inflammatory responsiveness and immune tolerance at barrier surfaces, thereby contributing to species-specific patterns of microbial colonization, immune compartmentalization, and susceptibility to dysbiosis (Colella et al., 2023; Zhao et al., 2026). Environmental conditions further amplify these interspecies differences. Domestication, intensive farming practices, urbanization, and altered environmental microbial diversity can substantially reshape the antigenic landscapes encountered at mucosal surfaces (Kauer et al., 2025). Grazing mammals, for example, experience continual exposure to soil-associated microorganisms, helminths, and environmental fungi at scales distinct from those encountered by indoor, captive, or predatory species (Kauer et al., 2025). Such ecological variation influences microbial diversity, epithelial barrier function, inflammatory thresholds, and pathogen susceptibility across mammalian hosts (Kauer et al., 2025; Zhao et al., 2026).
3.4. Host-Microbiota Co-Evolution Across Species
Mammals and their microbiota have evolved through long-standing reciprocal interactions that extend beyond passive microbial colonization (Groen et al., 2026). Host genomes encode structural, metabolic, and immunological traits that shape microbial niche selection, whereas resident microbial communities exert selective pressures on epithelial barrier function and immune regulation. Over evolutionary time, these interactions have contributed to species-specific patterns of mucosal compartmentalization, including variation in lymphoid tissue distribution, epithelial specialization, and antimicrobial immune programs across mammals (Beurel, 2024; Wang et al., 2025). Comparative evidence suggests that these adaptations are closely linked to ecological lifestyle and microbial exposure. In ruminants, specialized mucosal immune structures may have evolved to maintain tolerance toward dense fermentative microbial communities while preserving surveillance against translocating pathogens (Tardiolo et al., 2025; Souza et al., 2026). Likewise, bats exhibit distinct microbiota-associated immune regulatory features that may contribute to viral tolerance and persistent asymptomatic carriage of zoonotic viruses (Crowley et al., 2024; Roffler et al., 2024). These findings suggest that co-evolved host–microbiota systems influence microbial exclusion, pathogen accommodation, and reservoir competence across species. However, these ecological equilibria may be disrupted by anthropogenic pressures, including antibiotic exposure, habitat fragmentation, urbanization, and intensive livestock farming (White et al., 2023; Kauer et al., 2025). Such disturbances can alter mucosal barrier integrity, inflammatory tone, and pathogen susceptibility, potentially favoring pathogen amplification and zoonotic spillover (Kauer et al., 2025). Collectively, these observations highlight the importance of host–microbiota co-evolution in shaping mucosal immune organization and the dynamics of emerging infectious diseases within a One Health framework.
4. Mucosal Compartmentalization in Pathogen Susceptibility and Zoonotic Risk
Mucosal surfaces are key biological decision points in zoonotic infection because they determine whether pathogen exposure is blocked, converted into productive replication, tolerated as persistent carriage, or amplified through shedding. This section argues that zoonotic susceptibility is not only species-specific but also tissue-specific: a host may resist infection at one mucosal site while permitting replication or transmission from another. Using respiratory, gastrointestinal, and secretory epithelial sites as examples, the section explains how epithelial barriers, interferon responses, local antibodies, and tissue-resident immune cells shape pathogen entry and containment in the airway (Mitsi et al., 2023; Noh & Rha, 2024), how microbiota and nutrient ecology influence intestinal persistence and shedding (Ruddle et al., 2023), how immune tolerance can sustain asymptomatic carriage (Di Luccia et al., 2025), and how mammary epithelial tropism can create an unexpected shedding route during zoonotic infection (Caserta et al., 2024).
4.1 Barrier Immunity and Pathogen Entry
Barrier immunity functions as the first selective filter in the mucosal compatibility cascade. Pathogen exposure does not inevitably result in infection; successful entry depends on whether the exposed tissue provides accessible epithelial surfaces, compatible receptors, activating proteases, and a local immune environment permissive for early replication (Abdelwhab & Mettenleiter, 2023; Lean et al., 2023; Noh & Rha, 2024; Shi et al., 2024). Within the respiratory tract, epithelial junctions, mucociliary clearance, mucins, antimicrobial peptides, commensal microorganisms, immunoglobulins, dendritic cells, and type I/type III interferon responses all form a layered defense network that restricts pathogen attachment, invasion, and early replication (Noh & Rha, 2024). These defenses are both structural and immunological: interferons induce antiviral programs through interferon-stimulated genes, whereas airway antibodies and tissue-resident lymphocytes provide localized immune memory that may not be reflected in peripheral circulation (Mitsi et al., 2023; Noh & Rha, 2024). Consequently, the site of exposure strongly influences infection outcomes because the nasal epithelium, tracheobronchial mucosa, and lower airways differ in receptor landscapes, protease availability, and local immune pressures (Noh & Rha, 2024).
Influenza A viruses illustrate how receptor compatibility shapes mucosal entry. Human-adapted strains preferentially bind α2,6-linked sialic acids enriched within the mammalian respiratory tract, whereas avian-adapted strains preferentially bind α2,3-linked sialic acids classically associated with the avian intestinal tract (Abdelwhab & Mettenleiter, 2023; Petro-Turnquist et al., 2024). Swine are therefore important in influenza ecology because their respiratory tract may contain both receptor types, permitting infection by avian, swine, and human influenza viruses and creating opportunities for reassortment (Petro-Turnquist et al., 2024; Madapong et al., 2025). However, receptor distribution alone does not determine mixing-vessel potential, which additionally depends on host contact patterns, viral diversity, co-infection opportunities, and tissue permissiveness (Abdelwhab & Mettenleiter, 2023; Madapong et al., 2025).
Coronaviruses further demonstrate that receptor presence alone is insufficient for productive infection. ACE2 expression varies substantially across tissues and host species, meaning that susceptibility depends partly on the anatomical localization of receptors rather than simply their presence (Lean et al., 2023). SARS-CoV-2 additionally requires spike activation by host proteases, and Omicron variants exhibit enhanced infection of nasal epithelial cells through metalloproteinase-dependent pathways that favor upper-airway replication while partially evading local antiviral restriction (Shi et al., 2024). Thus, productive mucosal entry depends on coordinated compatibility among receptor access, protease activation, epithelial permissiveness, and immune evasion mechanisms.
Coronaviruses reinforce the same argument: receptor presence alone is not enough. ACE2 mapping shows that receptor expression varies across species and tissues, meaning that coronavirus susceptibility depends partly on where receptors are expressed, not simply whether a species carries them (Lean et al., 2023). SARS-CoV-2 also requires spike activation by host proteases, and Omicron’s enhanced infection of nasal epithelial cells through a metalloproteinase-dependent route shows how variant-specific adaptation can increase upper-airway infectivity while evading local antiviral restriction (Shi et al., 2024). Thus, productive mucosal entry requires the convergence of receptor access, protease availability, epithelial permissiveness, and immune evasion. The route of exposure further determines which mucosal barriers pathogens must overcome. Nipah virus may enter via respiratory or gastrointestinal routes, while infected Pteropus bats shed virus in saliva, urine, and feces, creating multiple opportunities for mucosal exposure (Scotto et al., 2026). In contrast, Salmonella primarily follows an intestinal route involving epithelial adhesion, invasion of epithelial or phagocytic cells, and interaction with host pattern-recognition receptors (Lamichhane et al., 2024).
- Mucosal Determinants of Pathogen Replication and Shedding
Following entry, zoonotic risk depends on whether a mucosal compartment supports replication to levels sufficient for shedding and onward transmission. Productive infection is shaped by epithelial permissiveness, pathogen immune evasion, interferon timing, local antibody activity, microbiota-mediated resistance, and tissue-resident immune memory (Noh & Rha, 2024; Mitsi et al., 2023; Ruddle et al., 2023; Di Luccia et al., 2025). Importantly, infection, disease, and transmission are not synonymous outcomes, as pathogens may bind receptors or enter cells yet still fail to replicate efficiently within the tissue compartments required for sustained shedding.
Respiratory virus studies illustrate this principle. Recent clade 2.3.4.4b H5N1 viruses attach to and replicate within human nasal and tracheobronchial epithelial cultures more efficiently than earlier H5N1 strains, suggesting increased mammalian adaptation potential (Bauer et al., 2026). However, bovine H5N1 transmitted inefficiently between experimentally exposed ferrets despite features associated with mammalian infection, demonstrating that receptor binding and tissue tropism alone do not guarantee efficient host-to-host transmission (Eisfeld et al., 2024). Similarly, BANAL-related bat coronaviruses may exhibit substantial genomic similarity to high-risk coronaviruses yet replicate poorly in human nasal epithelial cells and show reduced transmission in animal models, emphasizing that spillover risk requires functional evidence of replication and shedding rather than genomic similarity alone (Peña-Hernández et al., 2024).
The H5N1 outbreak in dairy cattle further demonstrates how transmission biology may shift when pathogens exploit unexpected shedding compartments. In infected cattle, viral RNA and infectious virus were consistently detected in milk, with marked tropism for milk-secreting mammary epithelial cells and higher viral detection in milk than in nasal secretions, urine, or feces (Caserta et al., 2024). Experimental infection studies in lactating goats similarly demonstrated H5N1-associated mastitis, high viral loads in milk, transmission to suckling offspring, and replication within both respiratory and mammary epithelial tissues (Alkie et al., 2025). These findings expand influenza ecology beyond the classical respiratory–intestinal framework and highlight how secretory epithelial tissues may become epidemiologically important when they generate high-volume infectious output (Caserta et al., 2024; Alkie et al., 2025).
Within the gastrointestinal tract, shedding dynamics reflect the balance between pathogen invasion, microbiota-mediated colonization resistance, nutrient availability, and local immune regulation. Salmonella must overcome colonization resistance imposed by commensal microorganisms, which restrict expansion through nutrient competition, short-chain fatty acid production, antimicrobial peptides, and immune stimulation (Ruddle et al., 2023; Lamichhane et al., 2024). The pathogen may bypass these restrictions through epithelial invasion, intracellular survival, virulence-associated secretion systems, and exploitation of metabolic niches such as diet-derived L-arabinose (Ruddle et al., 2023). In typhoid models, early Peyer’s patch invasion and hypoxia-associated immune signatures further link local mucosal invasion to progression toward systemic disease (Bossel Ben-Moshe et al., 2025).
Ultimately, local mucosal immunity determines whether pathogen replication is contained or progresses to sustained shedding. In SARS-CoV-2 infection, combined infection and vaccination induce stronger airway mucosal antibodies and memory B-cell responses than vaccination alone, whereas parenteral vaccination may not consistently establish tissue-resident spike-specific T cells within the respiratory mucosa (Mitsi et al., 2023). Respiratory containment also depends on the coordinated activity of IgA, tissue-resident memory T-cells, mucosal memory B cells, MAIT cells, interferons, mucins, and antimicrobial peptides (Noh & Rha, 2024). Collectively, these examples demonstrate that mucosal shedding reflects a dynamic contest between pathogen replication capacity and the speed, localization, and effectiveness of compartment-specific immune containment mechanisms (Table 2).
4.2. Reservoir Competence and Transmission Dynamics
Reservoir competence emerges when a host maintains a pathogen without developing disease severe enough to interrupt persistence or transmission. This state reflects the balance between resistance mechanisms that reduce pathogen burden and tolerance mechanisms that limit tissue damage despite continued infection (Irving et al., 2021; Di Luccia et al., 2025). At mucosal surfaces, tolerance becomes epidemiologically important when clinically silent hosts continue disseminating pathogens through secretions, excretions, contaminated food sources, or close-contact interfaces (Di Luccia et al., 2025; Scotto et al., 2026).
Salmonella superspreaders illustrate how mucosal tolerance can simultaneously facilitate persistence and transmission. During chronic enteric infection, regulatory T-cells preserve intestinal barrier integrity and limit immunopathology without fully eliminating pathogen burden, thereby allowing asymptomatic hosts to continue shedding Salmonella (Di Luccia et al., 2025). Treg-mediated control, including suppression of cytotoxic CD4+ T-cell expansion by T-bet-expressing colonic regulatory T-cells, may therefore explain how a relatively small subset of high-burden carriers contributes disproportionately to disease spread despite limited clinical signs (Di Luccia et al., 2025).
Bats extend this principle at an evolutionary scale. Their reservoir capacity appears linked to an immune equilibrium in which enhanced antiviral defenses coexist with restrained inflammatory responses, including constitutive interferon signaling alongside dampened STING, NLRP3 inflammasome, PYHIN, and IL-1β pathways (Irving et al., 2021). This balance may permit prolonged persistence of viruses such as Hendra, Nipah, Marburg, Ebola, SARS-related coronaviruses, and MERS-related coronaviruses with limited disease expression, thereby supporting long-term reservoir status across bat populations (Irving et al., 2021).
However, reservoir competence translates into spillover risk only when pathogen shedding intersects with susceptible host interfaces. Nipah virus illustrates this transition: Pteropus bats may harbor infection asymptomatically while shedding virus through saliva, urine, and feces, and human outbreaks in Bangladesh and India have been linked to consumption of raw date palm sap or fruit contaminated with bat excreta (Scotto et al., 2026). Similarly, leptospirosis transmission depends not only on infected rodent reservoirs but also on environmental persistence, shaped by rainfall, sanitation, exposure to contaminated water, and host contact patterns (Soni et al., 2024). These examples demonstrate that transmission dynamics are governed not simply by reservoir infection, but by the convergence of pathogen maintenance, shedding intensity, environmental persistence, and ecological exposure opportunities.
4.3. Comparative Implications for Zoonotic Spillover
Comparative evidence across livestock, wildlife, companion animals, and humans demonstrates that zoonotic spillover is not determined solely by exposure (Abdelwhab & Mettenleiter, 2023). A pathogen becomes a spillover threat only when host ecology aligns with tissue permissiveness, immune evasion or tolerance, efficient shedding, and sustained opportunities for contact (Abdelwhab & Mettenleiter, 2023; Caserta et al., 2024). Consequently, host species should not be classified simply as “susceptible” or “resistant,” but rather according to the mucosal compartments that permit replication, the routes through which pathogens are shed, and the ecological interfaces that facilitate transmission (Abdelwhab & Mettenleiter, 2023; Caserta et al., 2024; Soni et al., 2024; Di Luccia et al., 2025).
In livestock systems, agricultural species may function as amplification hosts when mucosal or secretory tissues support high-level pathogen replication and shedding (Caserta et al., 2024). Swine illustrate the importance of receptor compatibility and reassortment potential, whereas H5N1 infection in dairy cattle demonstrates how nonclassical secretory tissues can generate unexpected transmission routes through milk-associated shedding (Caserta et al., 2024; Petro-Turnquist et al., 2024; Alkie et al., 2025; Madapong et al., 2025). These observations suggest that livestock surveillance should extend beyond infection detection to include tissue tropism, routes of shedding, farm management practices, and the likelihood of sustained animal-to-animal or animal-to-human exposure. Similarly, wildlife reservoirs reflect the convergence of tissue permissiveness, immune tolerance, shedding dynamics, population ecology, and environmental mobility. Bats, rodents, and wild birds each represent distinct reservoir strategies in which viral persistence, excretory shedding, migratory behavior, or synanthropic ecology facilitate pathogen maintenance and transmission across species interfaces (Irving et al., 2021; Scotto et al., 2026; Soni et al., 2024; Abdelwhab & Mettenleiter, 2023). Consequently, this highlights that wildlife surveillance should prioritize identifying which hosts support persistence, which tissues support shedding, and the ecological interactions that permit multi-species or unispecific spillover
Companion animals, however, occupy a distinct intermediary position because they share close human contact while also interacting with household, farm-associated, food-associated, or wildlife-linked environments. Hence, their epidemiological importance lies less in the primary reservoir’s functions but more in revealing high-risk interfaces where pathogen adaptation, environmental contamination, feeding practices, and close human exposure intersect (Lamichhane et al., 2024; Caserta et al., 2024; Hwang et al., 2025). As such, companion animals may serve as important sentinels of spillover risk at the boundary between domestic, agricultural, and wildlife ecosystems. In humans, mucosal immunity determines whether zoonotic exposure is eliminated locally or progresses to infection, disease, and onward transmission. Systemic immune measurements may not accurately reflect mucosal protection, hence, spillover preparedness should incorporate mucosal correlates of protection, including local antibodies, tissue-resident memory cells, interferon responsiveness, and durability of vaccine-induced mucosal immunity (Mitsi et al., 2023; Noh & Rha, 2024). These observations show that zoonotic risk can be reframed as a tissue-level process with population-level consequences. Future spillover prediction should therefore integrate receptor mapping, primary epithelial infection models, mucosal immune profiling, microbiome analysis, shedding quantification, and ecological exposure data to distinguish hosts that merely encounter pathogens from those capable of maintaining, amplifying, and transmitting them across species boundaries (Table 2).
5. Emerging Technologies and Knowledge Gaps in Comparative Mucosal Immunology
Despite major advances in mucosal immunology, the field remains constrained by a persistent disconnect between mechanistic resolution and ecological relevance. Much of the current understanding of barrier immunity derives from reductionist experimental systems dominated by laboratory rodents maintained under controlled environmental conditions, whereas zoonotic emergence and host–microbiota interactions occur within ecologically complex and evolutionarily diverse mammalian systems (Beura et al., 2016; Abolins et al., 2017). Consequently, fundamental aspects of comparative mucosal immune organization, tissue specialization, and host–microbiota adaptation across wildlife, livestock, companion animals, and humans remain incompletely understood. This limitation has become increasingly important within a One Health framework because spillover risk, reservoir competence, and pathogen persistence are now recognized as tissue-level processes shaped by highly specialized mucosal interfaces rather than by pathogen exposure alone. Emerging spatial, single-cell, and systems-level technologies therefore provide an opportunity to move comparative mucosal immunology beyond descriptive anatomy toward a more predictive form of ecological immunology capable of resolving how barrier tissues regulate host–pathogen interactions across species.
5.1. Single-Cell and Spatial Mucosal Immunology
Recent advances in single-cell transcriptomics, spatial transcriptomics, high-dimensional imaging, and multiplexed tissue analysis are transforming the study of mucosal immunity by enabling simultaneous characterization of epithelial, stromal, microbial, and immune-cell interactions within intact tissue microenvironments (Liu et al., 2021; FitzPatrick et al., 2025; Kang et al., 2025) These approaches have demonstrated that barrier tissues contain highly specialized cellular niches whose organization and function cannot be fully resolved through bulk immunological analyses alone (Krausgruber et al., 2020). Structural cell populations, including epithelial and stromal cells, are now recognized as active regulators of localized immune responses through cytokine production, chemokine gradients, metabolic signaling, and control of immune-cell residency (Krausgruber et al., 2020; Gray & Farber, 2022).
Within mucosal systems, spatial organization is particularly important because immune function depends not only on cellular composition but also on anatomical positioning relative to epithelial barriers, microbiota, vascular networks, and lymphoid structures (Arroyo Portilla et al., 2021; Gray & Farber, 2022). Tissue-resident memory T cells (TRM), innate lymphoid cells (ILCs), macrophages, dendritic cells, and epithelial subsets exhibit compartment-specific transcriptional programs that vary substantially across intestinal, respiratory, oral, and secretory tissues (Arroyo Portilla et al., 2021; Gray & Farber, 2022). Spatially resolved technologies, therefore, provide an opportunity to define comparative “mucosal immune landscapes” across species and ecological settings rather than relying on simplified binary models of immune activation or tolerance.
Importantly, these technologies provide an opportunity to resolve mucosal immunity at the level of tissue microenvironments rather than bulk cellular averages. By integrating spatial organization with transcriptional and functional profiling, single-cell and spatial approaches can reveal how epithelial niches, resident immune populations, stromal networks, and local microbial communities collectively shape compartment-specific immune responses across species. Such analyses may be particularly valuable for understanding why certain tissues support pathogen persistence, asymptomatic carriage, or high-efficiency shedding, thereby linking mucosal immune architecture to zoonotic transmission dynamics.
5.2. Comparative Omics and Systems Immunology
Comparative omics approaches increasingly demonstrate that mucosal immunity functions as an integrated ecological network involving host genetics, microbiota composition, epithelial metabolism, immune signaling, and environmental exposures (Fu et al., 2023; Zhang et al., 2024). Metagenomics, metatranscriptomics, metabolomics, and single-cell immune profiling now enable simultaneous interrogation of microbial communities and host immune states across barrier tissues, providing systems-level insight into host–microbiota co-adaptation (Fu et al., 2023; Bishop et al., 2022). These technologies are particularly valuable in comparative studies because they allow immune function to be interpreted within a physiological and ecological context rather than as isolated molecular pathways.
Emerging evidence further suggests that species-specific immune phenotypes are strongly shaped by differences in microbiota composition, dietary ecology, fermentation biology, and environmental microbial exposure (Tardiolo et al., 2025; Zhao et al., 2026). In ruminants, mucosal immunity is closely linked to foregut fermentation systems and microbial metabolite production, whereas monogastric species exhibit distinct patterns of microbial organization and immune compartmentalization (Li et al., 2023; Zulfiqar et al., 2025). Comparative systems immunology therefore provides an opportunity to distinguish conserved from lineage-specific immune programs across mammals and may help explain why some species function as tolerant reservoirs while others develop severe inflammatory pathology following infection. Integrated multi-omics frameworks may further improve understanding of zoonotic susceptibility by combining epithelial receptor distribution, microbiome composition, tissue-specific immune signatures, metabolite networks, and pathogen replication phenotypes within unified analytical models (Fu et al., 2023; Zhang et al., 2024). Such approaches move comparative mucosal immunology beyond descriptive characterization toward a more mechanistic understanding of pathogen emergence, host adaptation, and transmission potential across ecological systems.
5.3. Lack of Cross-Species Mechanistic Studies
Despite growing recognition of comparative diversity in mucosal immunity, mechanistic studies remain heavily concentrated in laboratory mice and selected human cohorts. Although murine models have provided foundational insights into lymphoid organization, epithelial biology, and immune regulation, laboratory mice maintained under specific pathogen-free conditions differ substantially from wild mammals in microbial exposure history, immune maturation, tissue-resident immune composition, and inflammatory responsiveness (Abolins et al., 2017; Beura et al., 2016; Rosshart et al., 2017). As a result, many paradigms considered canonical in mucosal immunology may partially reflect laboratory adaptation rather than broadly conserved mammalian biology.
This limitation is particularly important in a One Health context because many reservoir hosts and agricultural species possess mucosal architectures, microbial ecologies, and immunological pressures that are absent from murine systems. Ruminants exhibit specialized ileal Peyer’s patches that function as primary lymphoid organs during development, bats display distinctive antiviral and inflammatory regulatory programs associated with viral tolerance, and wildlife species encounter environmental and microbial exposures fundamentally different from those of laboratory animals (Yasuda et al., 2006; Irving et al., 2021; Pavlovich et al., 2018). Yet direct comparative studies examining how tissue architecture, microbiota composition, and immune specialization interact across mammalian species remain limited. Consequently, the field faces a major translational bottleneck since ecological observations continue to accumulate more rapidly than mechanistic understanding. Addressing this gap will require comparative tissue atlases, standardized mucosal sampling strategies, wildlife-compatible immunological tools, and analytical frameworks that integrate ecological, microbiological, and molecular data across species. Such efforts will be essential for determining which aspects of mucosal immunity are evolutionarily conserved and which represent lineage-specific adaptations shaped by ecology, microbial exposure, and host–pathogen interactions.
5.4. Challenges in Translating Findings Across Mammals
A major challenge in comparative mucosal immunology is that immune mechanisms identified in one species may not translate directly across mammals because mucosal tissues are shaped by evolutionary history, ecological niche, microbial exposure, and physiological specialization. Even broadly conserved pathways, including interferon signaling, IgA-mediated immunity, epithelial sensing, and tissue-resident immune maintenance, may function differently depending on tissue architecture, microbial ecology, and metabolic environment (Peterson & Artis, 2014; Gray & Farber, 2022; Wang et al., 2025). Consequently, extrapolation from experimentally tractable species to livestock, wildlife reservoirs, or humans often oversimplifies the ecological and immunological complexity underlying barrier immunity (Abolins et al., 2017; Rosshart et al., 2017).
Microbiome variability further complicates translation across species. Host-associated microbial communities differ substantially across diet, geography, domestication, captivity, and antibiotic exposure, meaning that immune phenotypes observed under controlled laboratory conditions may not accurately reflect those observed in natural ecosystems (White et al., 2023; Kauer et al., 2025). Similarly, ruminants, monogastric mammals, bats, and wildlife reservoirs exhibit distinct microbial ecologies and mucosal immune adaptations associated with differences in fermentation biology, environmental exposure, and pathogen tolerance (Irving et al., 2021; Tardiolo et al., 2025; Zhao et al., 2026). Anthropogenic environmental change may further reshape mucosal inflammatory thresholds and pathogen susceptibility, thereby influencing pathogen persistence and spillover dynamics across species (White et al., 2023; Kauer et al., 2025). These limitations suggest that future progress in One Health immunology will depend on moving beyond species-isolated experimental systems, and toward ecologically integrated comparative frameworks (Figure 2). Predictive mucosal immunology will likely require integrating spatial tissue biology, comparative multi-omics, microbiome ecology, epithelial infection models, and longitudinal environmental surveillance to identify the tissue-level determinants of pathogen emergence across interconnected human, animal, and environmental systems (Table 1 and Table 2). Ultimately, the next generation of comparative mucosal immunology must move beyond descriptive characterization of barrier tissues toward mechanistic ecological models that explain how host-microbiota-pathogen interactions shape zoonotic risk across mammals.
6. Conclusions and Future Perspectives
Comparative evidence across mammalian systems increasingly demonstrates that mucosal immunity is not a universally conserved barrier defense program but an evolutionarily and ecologically specialized system, shaped by host anatomy, microbial exposure, diet, and environmental pressure (Brown & Esterházy, 2021; Wang et al., 2025; Zhou et al., 2025). Across gastrointestinal, respiratory, and urogenital tissues, mucosal compartments function as critical interfaces where microbial tolerance, pathogen restriction, immune homeostasis, and zoonotic transmission are simultaneously regulated. From a One Health perspective, these interfaces are particularly important because controlling infection within livestock and wildlife reservoirs not only protects animal health, but also reduces opportunities for pathogen amplification and spillover into human populations (World Health Organization, 2022; Marzouk & Alajaji, 2025). Consequently, spillover risk cannot be understood through pathogen exposure alone, but must instead be interpreted through tissue permissiveness, immune regulation, shedding biology, and ecological contact networks operating at mucosal surfaces (Murphy et al., 2022; Gleeson & Monteiro, 2024; Table 2).
The findings synthesized throughout this review further challenge long-standing reliance on generalized murine or human-centric models of barrier immunity. Although conserved mechanisms of epithelial defense, IgA-mediated immunity, and tissue-resident immune surveillance exist across mammals, comparative studies increasingly demonstrate that mucosal immune organization is strongly shaped by microbial ecology, environmental exposure, and host-specific physiological adaptation (Abolins et al., 2017; Rosshart et al., 2017; Brown & Esterházy, 2021; Gray & Farber, 2022). Structural variation in lymphoid architecture, epithelial specialization, microbiota composition, and immune tolerance programs suggests that barrier immunity emerges from dynamic host–microbe–environment interactions rather than from a fixed universal blueprint (Table 1). Understanding why some hosts function as asymptomatic reservoirs, whereas others develop severe inflammatory disease, will therefore require comparative and spatial approaches capable of resolving tissue-level immune organization across species.
Advances in spatial immunology, comparative multi-omics, and epithelial infection models are now beginning to resolve mucosal immunity at the level of tissue microenvironments and ecological systems (Krausgruber et al., 2020; Fu et al., 2023). These developments carry important implications for vaccine design, surveillance, and zoonotic disease control because immune protection is often most effective when localized at pathogen entry sites. Consequently, mucosal vaccination remains a major translational objective in both human and veterinary medicine, although progress in mammalian systems remains constrained by the intrinsic tolerogenic nature of mucosal tissues (Nochi et al., 2018; Niewiesk, 2014; Hui et al., 2026). Emerging delivery systems, including nanotechnology-based carriers, cationic liposomes, and environmentally responsive mucosal adjuvants, may improve antigen retention, epithelial uptake, and localized immune activation at barrier surfaces (Bernasconi et al., 2016; Singh et al., 2025). However, major gaps remain in understanding how developmental exposure, microbial ecology, environmental disruption, and species-specific immune specialization collectively shape mucosal responsiveness, pathogen persistence, and transmission across mammals. Addressing these challenges will require integrated comparative frameworks that combine ecological biology, microbiome science, systems immunology, and longitudinal surveillance to improve prediction, prevention, and control of emerging infectious diseases at the animal–human–environment interface.
CRediT authorship contribution statement: Babatunde Ibrahim Olowu: Conceptualization, Visualization, Writing – original draft, Writing – review and editing; Olalekan Toheeb Okewale: Writing – original draft, Writing – review and editing; Abdulhakeem Opeyemi Azeez: Writing – original draft, Writing – review and editing; Adeyemi Shammah Oluwaseyi: Visualization; Kehinde Samuel Adebayo: Writing – original draft; Nahimah Opeyemi Idris: Writing – original draft; Oyeyebi Dolapo Qudus: Writing – original draft; Al-Amin Adebare Olojede: Writing – original draft; Henrietta Elizabeth Nduka: Writing – review and editing; Zaynab Adeshewa Omishakin: Writing – original draft.
Funding statement: This study has received no external funding.
Declaration of competing interest: The authors declare that they have no known competing financial interests or personal relationships that could have influenced the work reported in this paper.
Data availability: The datasets supporting this review are included within the manuscript; original sources are cited in the text and can be consulted for further details.
Ethics declaration: Not applicable
References
- Abdelwhab, E. M.; Mettenleiter, T. C. Zoonotic animal influenza virus and potential mixing vessel hosts. Viruses 2023, 15(4), 980. [Google Scholar] [CrossRef] [PubMed]
- Abolins, S.; King, E. C.; Lazarou, L.; Weldon, L.; Hughes, L.; Drescher, P.; Raynes, J. G.; Hafalla, J. C. R.; Viney, M. E.; Riley, E. M. The comparative immunology of wild and laboratory mice, Mus musculus domesticus. Nat. Commun. 2017, 8, 14811. [Google Scholar] [CrossRef] [PubMed]
- Al Nabhani, Z.; Dulauroy, S.; Marques, R.; Cousu, C.; Al Bounny, S.; Déjardin, F.; Sparwasser, T.; Bérard, M.; Cerf-Bensussan, N.; Eberl, G. A Weaning Reaction to Microbiota Is Required for Resistance to Immunopathologies in the Adult. Immunity 2019, 50(5), 1276–1288.e5. [Google Scholar] [CrossRef] [PubMed]
- Alkie, T. N.; Embury-Hyatt, C.; Signore, A. V.; Ramos, D.; Moffat, E.; Raj, S.; Berhane, Y. Dairy cow-and avian-origin clade 2.3. 4.4 b H5N1 induce severe mastitis in lactating goats and transmission to suckling goats. Cell Rep. 2025, 44(10). [Google Scholar] [CrossRef] [PubMed]
- Allie, S. R.; Randall, T. D. Pulmonary immunity to viruses. Clin. Sci. 2017, 131(14), 1737–1762. [Google Scholar] [CrossRef] [PubMed]
- Almansour, N.; Al-Rashed, F.; Choudhry, K.; Alqaderi, H.; Sindhu, S.; Al-Mulla, F.; Ahmad, R. Gut microbiota: a promising new target in immune tolerance. Front. Immunol. 2025, 16, 1607388. [Google Scholar] [CrossRef] [PubMed]
- Arroyo Portilla, C.; Tomas, J.; Gorvel, J. P.; Lelouard, H. From Species to Regional and Local Specialization of Intestinal Macrophages. Front. Cell Dev. Biol. 2021, 8, 624213. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.; Leijten, L.; Iervolino, M.; Chopra, V.; van Dijk, L.; Power, M.; van Riel, D. Attachment and replication of clade 2.3. 4.4 b influenza A (H5N1) viruses in human respiratory epithelium: an in-vitro study. Lancet Microbe 2026, 7(1). [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T. W. Role of the microbiota in immunity and inflammation. Cell 2014, 157(1), 121–141. [Google Scholar] [CrossRef] [PubMed]
- Bello, M. B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B. P. H.; Omar, A. R. Diagnostic and Vaccination Approaches for Newcastle Disease Virus in Poultry: The Current and Emerging Perspectives. In BioMed Research International; 2018; pp. 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, V.; Norling, K.; Bally, M.; Höök, F.; Lycke, N. Y. Mucosal Vaccine Development Based on Liposome Technology. J. Immunol. Res. 2016, 2016, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Beura, L. K.; Hamilton, S. E.; Bi, K.; Schenkel, J. M.; Odumade, O. A.; Casey, K. A.; Thompson, E. A.; Fraser, K. A.; Rosato, P. C.; Filali-Mouhim, A.; Sekaly, R. P.; Jenkins, M. K.; Vezys, V.; Haining, W. N.; Jameson, S. C.; Masopust, D. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532(7600), 512–516. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E. Stress in the microbiome-immune crosstalk. Gut Microbes 2024, 16(1). [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.L.; Drikic, M.; Wacker, S.; et al. Moving beyond descriptive studies: harnessing metabolomics to elucidate the molecular mechanisms underpinning host-microbiome phenotypes. Mucosal Immunol. 2022, 15, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Bossel Ben-Moshe, N.; Hen-Avivi, S.; Levy Efrati, L.; Veinman, L.; Hill, J.; O’Connor, D.; Avraham, R. Salmonella Typhi gut invasion drives hypoxic immune subsets associated with disease outcomes. Nat. Commun. 2025, 16(1), 6755. [Google Scholar] [CrossRef] [PubMed]
- Boyaka, P. N.; Fujihashi, K. Host defenses at mucosal surfaces. In Clinical immunology; Elsevier, 2019; pp. 285–298. [Google Scholar]
- Brandtzaeg, P. Secretory IgA: Designed for Anti-Microbial Defense. Front. Immunol. 2013, 4, 222. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P.; Johansen, F. E. Mucosal B cells: phenotypic characteristics, transcriptional regulation, and homing properties. Immunol. Rev. 2005, 206, 32–63. [Google Scholar] [CrossRef] [PubMed]
- Brook, C. E.; Dobson, A. P. Bats as 'special' reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23(3), 172–180. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.; Esterházy, D. Intestinal immune compartmentalization: Implications of tissue specific determinants in health and disease. Mucosal Immunol. 2021, 14(6), 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Bunker, J. J.; Flynn, T. M.; Koval, J. C.; Shaw, D. G.; Meisel, M.; McDonald, B. D.; Ishizuka, I. E.; Dent, A. L.; Wilson, P. C.; Jabri, B.; Antonopoulos, D. A.; Bendelac, A. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43(3), 541–553. [Google Scholar] [CrossRef] [PubMed]
- Canesso, M. C. C.; Moreira, T. G.; Faria, A. M. C. Compartmentalization of gut immune responses: Mucosal niches and lymph node peculiarities. Immunol. Lett. 2022, 251–252, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Carreto-Binaghi, L. E.; Sztein, M. B.; Booth, J. S. Role of cellular effectors in the induction and maintenance of IgA responses leading to protective immunity against enteric bacterial pathogens. Front. Immunol. 2024, 15, 1446072. [Google Scholar] [CrossRef] [PubMed]
- Caserta, L. C.; Frye, E. A.; Butt, S. L.; Laverack, M.; Nooruzzaman, M.; Covaleda, L. M.; Diel, D. G. Spillover of highly pathogenic avian influenza H5N1 virus to dairy cattle. Nature 2024, 634(8034), 669–676. [Google Scholar] [CrossRef] [PubMed]
- Colella, M.; Charitos, I. A.; Ballini, A.; Cafiero, C.; Topi, S.; Palmirotta, R.; Santacroce, L. Microbiota revolution: How gut microbes regulate our lives. World J. Gastroenterol. 2023, 29(28), 4368. [Google Scholar] [CrossRef] [PubMed]
- Crowley, D. E.; Falvo, C. A.; Benson, E.; Hedges, J.; Jutila, M.; Ezzatpour, S.; Aguilar, H. C.; Ruiz-Aravena, M.; Ma, W.; Schountz, T.; Rynda-Apple, A.; Plowright, R. K. Bats generate lower affinity but higher diversity antibody responses than those of mice, but pathogen-binding capacity increases if protein is restricted in their diet. PLoS Biol. 2024, 22(9), e3002800. [Google Scholar] [CrossRef] [PubMed]
- Di Luccia, B.; Massis, L. M.; Butler, D. S.; Narasimhan, R.; Ruddle, S. J.; Pham, T. H.; Monack, D. M. Salmonella-superspreader hosts require gut regulatory T cells to maintain a disease-tolerant state. J. Exp. Med. 2025, 222(11), e20242431. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A. J.; Biswas, A.; Guan, L.; Gu, C.; Maemura, T.; Trifkovic, S.; Kawaoka, Y. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature 2024, 633(8029), 426–432. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.A. Immunology: Comparative Immunology of Mammals; eLS, Ed.; 2012. [Google Scholar] [CrossRef]
- FitzPatrick, M.E.B.; Antanaviciute, A.; Dunstan, M.; et al. Immune–epithelial–stromal networks define the cellular ecosystem of the small intestine in celiac disease. Nat. Immunol. 2025, 26, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhu, F.; Xu, C. J.; Li, Y. Metabolomics meets systems immunology. EMBO Rep. 2023, 24(4), e55747. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Ito, S.; Suzuki, S.; Fuchimoto, D.; Onishi, A.; Niimi, K.; Usami, K.; Wu, G.; Bazer, F. W.; Ogasawara, K.; Watanabe, K.; Aso, H.; Nochi, T. Organogenesis of Ileal Peyer's Patches Is Initiated Prenatally and Accelerated Postnatally With Comprehensive Proliferation of B Cells in Pigs. Front. Immunol. 2020, 11, 604674. [Google Scholar] [CrossRef] [PubMed]
- Ghaffar, K. A.; Marasini, N.; Giddam, A. K.; Batzloff, M. R.; Good, M. F.; Skwarczynski, M.; Toth, I. Liposome-based intranasal delivery of lipopeptide vaccine candidates against group A streptococcus. Acta Biomater. 2016, 41, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, P. J.; Monteiro, R. C. The Role of Mucosal Immunity: What Can We Learn From Animal and Human Studies? Semin. Nephrol. 2024, 44(5), 151566. [Google Scholar] [CrossRef] [PubMed]
- Gray, J. I.; Farber, D. L. Tissue-Resident Immune Cells in Humans. Annu. Rev. Immunol. 2022, 40, 195–220. [Google Scholar] [CrossRef] [PubMed]
- de Groen, P.; Gouw, S. C.; Hanssen, N. M. J.; Nieuwdorp, M.; Rampanelli, E. Early-Life Gut Microbiota: Education of the Immune System and Links to Autoimmune Diseases. Microorganisms 2026, Vol. 14(14(1)), 210. [Google Scholar] [CrossRef]
- Gupta, S.; Gupta, S. L.; Singh, A.; Oswal, N.; Bal, V.; Rath, S.; George, A.; Basu, S. IgA Determines Bacterial Composition in the Gut. Crohn’s Colitis 2023, 360(5(3)). [Google Scholar] [CrossRef]
- Hoces, D.; Arnoldini, M.; Diard, M.; Loverdo, C.; Slack, E. Growing, evolving and sticking in a flowing environment: Understanding IgA interactions with bacteria in the gut. Immunology 2020, 159(1), 52–62. [Google Scholar] [CrossRef] [PubMed]
- Hodgins, D. C.; Chattha, K.; Vlasova, A.; Parreño, V.; Corbeil, L. B.; Renukaradhya, G. J.; Saif, L. J. Mucosal Veterinary Vaccines. In Mucosal Immunology; Elsevier, 2015; pp. 1337–1361. [Google Scholar] [CrossRef]
- HogenEsch, H.; O’Hagan, D. T.; Fox, C. B. Optimizing the utilization of aluminum adjuvants in vaccines: You might just get what you want. npj Vaccines 2018, 3(1), 51. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Tian, X.; Ding, S.; Gao, G.; Gao, S.; Sun, A.; Zhao, T.; Wang, H. Advances and prospects of mucosal vaccination in the prevention and control of avian influenza. Front. Immunol. 2026, 17, 1766957. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Yoon, S. W.; Ga, E.; Choi, J.; Moon, S.; Bae, E.; Na, W. Intranasal NS1-truncated live attenuated canine influenza vaccine confers superior protection compared to inactivated vaccine in beagles. Vet. Res. 2025, 56(1), 178. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I. D.; Blander, J. M.; Collins, N.; Guo, C. J.; Longman, R. S.; Sonnenberg, G. F.; Zeng, M. Y.; Artis, D. Microbiota-mediated mechanisms of mucosal immunity across the lifespan. Nat. Immunol. 2025, 26(10), 1645. [Google Scholar] [CrossRef] [PubMed]
- Irving, A. T.; Ahn, M.; Goh, G.; Anderson, D. E.; Wang, L. F. Lessons from the host defences of bats, a unique viral reservoir. Nature 2021, 589(7842), 363–370. [Google Scholar] [CrossRef] [PubMed]
- Jones, K. E.; Patel, N. G.; Levy, M. A.; Storeygard, A.; Balk, D.; Gittleman, J. L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451(7181), 990–993. [Google Scholar] [CrossRef] [PubMed]
- Kabbert, J.; Kaminski, A.; Pabst, O.; Ugur, M. Intestinal microbiota regulates naïve lymphocyte migration in Peyer's patches. Front. Immunol. 2025, 16, 1717788. [Google Scholar] [CrossRef] [PubMed]
- Kang, D. H.; Kim, Y.; Lee, J. H.; Kang, H. S.; Chung, C. Spatial Transcriptomics in Lung Cancer and Pulmonary Diseases: A Comprehensive Review. Cancers 2025, 17(12), 1912. [Google Scholar] [CrossRef] [PubMed]
- Kauer, L.; Sapountzis, P.; Imholt, C.; Berens, C.; Kuehn, R. Microbial exchange at the wildlife-livestock interface: insights into microbial composition, antimicrobial resistance and virulence factor gene dynamics in grassland ecosystems. Anim. Microbiome 2025, 7(1), 84. [Google Scholar] [CrossRef] [PubMed]
- Kempf, F.; La Ragione, R.; Chirullo, B.; Schouler, C.; Velge, P. Super Shedding in Enteric Pathogens: A Review. Microorganisms 2022, 10(11), 2101. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ndwandwe, C.; Devotta, H.; Kareem, L.; Yao, L.; O’Mahony, L. Role of the microbiome in regulation of the immune system. Allergol. Int. 2025, 74(2), 187–196. [Google Scholar] [CrossRef] [PubMed]
- Krausgruber, T.; Fortelny, N.; Fife-Gernedl, V.; Senekowitsch, M.; Schuster, L. C.; Lercher, A.; Nemc, A.; Schmidl, C.; Rendeiro, A. F.; Bergthaler, A.; Bock, C. Structural cells are key regulators of organ-specific immune responses. Nature 2020, 583(7815), 296–302. [Google Scholar] [CrossRef] [PubMed]
- Kuthyar, S.; Reese, A. T. Variation in Microbial Exposure at the Human-Animal Interface and the Implications for Microbiome-Mediated Health Outcome. mSystems 2021, 6(4), e0056721. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, B.; Mawad, A. M.; Saleh, M.; Kelley, W. G.; Harrington, P. J.; Lovestad, C. W.; Helmy, Y. A. Salmonellosis: an overview of epidemiology, pathogenesis, and innovative approaches to mitigate the antimicrobial-resistant infections. Antibiotics 2024, 13(1), 76. [Google Scholar] [CrossRef] [PubMed]
- Lean, F. Z.; Cox, R.; Madslien, K.; Spiro, S.; Nymo, I. H.; Bröjer, C.; Núñez, A. Tissue distribution of angiotensin-converting enzyme 2 (ACE2) receptor in wild animals with a focus on artiodactyls, mustelids and phocids. One Health 2023, 16, 100492. [Google Scholar] [CrossRef] [PubMed]
- León, E. D.; Francino, M. P. Roles of Secretory Immunoglobulin A in Host-Microbiota Interactions in the Gut Ecosystem. Front. Microbiol. 2022, 13, 880484. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Shi, B.; Na, R. The Colonization of Rumen Microbiota and Intervention in Pre-Weaned Ruminants. Anim. An. Open Access J. From MDPI 2023, 13(6), 994. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.; Sun, Y.; Cui, H.; Zhu, S. J.; Qiu, H.-J. Mucosal vaccines: Strategies and challenges. Immunol. Lett. 2020, 217, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, M.; Deng, Y.; Su, G.; Enninful, A.; Guo, C. C.; Tebaldi, T.; Zhang, D.; Kim, D.; Bai, Z.; Norris, E.; Pan, A.; Li, J.; Xiao, Y.; Halene, S.; Fan, R. High-Spatial-Resolution Multi-Omics Sequencing via Deterministic Barcoding in Tissue. Cell 2020, 183(6), 1665–1681.e18. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Zhu, J.; Li, K.; Li, Y.; Li, J.; Chen, Y.; Shi, H. Crosstalk between innate immunity and rumen-fecal microbiota under the cold stress in goats. Front. Immunol. 2024, 15, 1363664. [Google Scholar] [CrossRef] [PubMed]
- Mabbott, N. A.; Hase, K. Editorial: Immunological consequences of antigen sampling at mucosal surfaces, volume II. Front. Immunol. 2024, 15, 1499174. [Google Scholar] [CrossRef] [PubMed]
- Mabbott, N. A.; Donaldson, D. S.; Ohno, H.; Williams, I. R.; Mahajan, A. Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013, 6(4), 666–677. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A. J.; Yilmaz, B.; Limenitakis, J. P.; Ganal-Vonarburg, S. C. IgA Function in Relation to the Intestinal Microbiota. Annu. Rev. Immunol. 2018, 36, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Madapong, A.; Wiggins, J.; DeBeauchamp, J.; Webby, R. J.; Weaver, E. A. Systemic and Mucosal Immune Responses Induced by Adenoviral-Vectored Consensus H5 Influenza A Vaccines in Mice and Swine. Vaccines 2025, 13(9), 928. [Google Scholar] [CrossRef] [PubMed]
- Marchesini Tovar, G.; Gallen, C.; Bergsbaken, T. CD8+ Tissue-Resident Memory T Cells: Versatile Guardians of the Tissue. J. Immunol. 2024, 212(3), 361–368. [Google Scholar] [CrossRef] [PubMed]
- Marzouk, E.; Alajaji, A. I. Preventive Immunology for Livestock and Zoonotic Infectious Diseases in the One Health Era: From Mechanistic Insights to Innovative Interventions. Vet. Sci. 2025, 12(10), 1014. [Google Scholar] [CrossRef] [PubMed]
- Mettelman, R. C.; Allen, E. K.; Thomas, P. G. Mucosal immune responses to infection and vaccination in the respiratory tract. Immunity 2022, 55(5), 749–780. [Google Scholar] [CrossRef] [PubMed]
- Mitsi, E.; Diniz, M. O.; Reiné, J.; Collins, A. M.; Robinson, R. E.; Hyder-Wright, A.; Ferreira, D. M. Respiratory mucosal immune memory to SARS-CoV-2 after infection and vaccination. Nat. Commun. 2023, 14(1), 6815. [Google Scholar] [CrossRef] [PubMed]
- Mowat, A. M.; Agace, W. W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14(10), 667–685. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C. C.; Rawls, J. F. Commensal Microbiota Regulate Vertebrate Innate Immunity-Insights From the Zebrafish. Front. Immunol. 2019, 10, 2100. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K. M.; Weaver, C.; Berg, L. J.; Janeway, C. A.; Barton, G. Janeway’s immunobiology, 10th edition, international student edition; W.W. Norton & Company, 2022. [Google Scholar]
- Neish, A. S. Mucosal Immunity and the Microbiome. Ann. Am. Thorac. Soc. 2014, 11 (Suppl 1), S28. [Google Scholar] [CrossRef] [PubMed]
- Niewiesk, S. Maternal Antibodies: Clinical Significance, Mechanism of Interference with Immune Responses, and Possible Vaccination Strategies. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Nochi, T.; Jansen, C. A.; Toyomizu, M.; Eden, W. V. The Well-Developed Mucosal Immune Systems of Birds and Mammals Allow for Similar Approaches of Mucosal Vaccination in Both Types of Animals. Front. Nutr. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Noh, H. E.; Rha, M. S. Mucosal immunity against SARS-CoV-2 in the respiratory tract. Pathogens 2024, 13(2), 113. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C. R.; Holmes, E. C.; Morens, D. M.; Park, E. C.; Burke, D. S.; Calisher, C. H.; Laughlin, C. A.; Saif, L. J.; Daszak, P. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol. Mol. Biol. Rev. MMBR 2008, 72(3), 457–470. [Google Scholar] [CrossRef] [PubMed]
- Pavlovich, S. S.; Lovett, S. P.; Koroleva, G.; Guito, J. C.; Arnold, C. E.; Nagle, E. R.; Kulcsar, K.; Lee, A.; Thibaud-Nissen, F.; Hume, A. J.; Mühlberger, E.; Uebelhoer, L. S.; Towner, J. S.; Rabadan, R.; Sanchez-Lockhart, M.; Kepler, T. B.; Palacios, G. The Egyptian Rousette Genome Reveals Unexpected Features of Bat Antiviral Immunity. Cell 2018, 173(5), 1098–1110.e18. [Google Scholar] [CrossRef] [PubMed]
- Peña-Hernández, M. A.; Alfajaro, M. M.; Filler, R. B.; Moriyama, M.; Keeler, E. L.; Ranglin, Z. E.; Wilen, C. B. SARS-CoV-2-related bat viruses evade human intrinsic immunity but lack efficient transmission capacity. Nat. Microbiol. 2024, 9(8), 2038–2050. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L. W.; Artis, D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14(3), 141–153. [Google Scholar] [CrossRef] [PubMed]
- Petro-Turnquist, E.; Pekarek, M. J.; Weaver, E. A. Swine influenza A virus: challenges and novel vaccine strategies. Front. Cell. Infect. Microbiol. 2024, 14, 1336013. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int. J. Mol. Sci. 2020, 21(23), 9254. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.; de Sousa-Pereira, P.; Esteves, P. J. The IgA of hares (Lepus sp.) and rabbit confirms that the leporids IgA explosion is old and reveals a new case of trans-species polymorphism. Front. Immunol. 2023, 14, 1192460. [Google Scholar] [CrossRef] [PubMed]
- Pirker, A. L.; Vogl, T. Development of systemic and mucosal immune responses against gut microbiota in early life and implications for the onset of allergies. Front. Allergy 2024, 5, 1439303. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M. T.; Sobur, M. A.; Islam, M. S.; Ievy, S.; Hossain, M. J.; El Zowalaty, M. E.; Rahman, A. T.; Ashour, H. M. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms 2020, 8(9), 1405. [Google Scholar] [CrossRef] [PubMed]
- Randeni, N.; Bordiga, M.; Xu, B. A Comprehensive Review of the Triangular Relationship among Diet–Gut Microbiota–Inflammation. Int. J. Mol. Sci. 2024, 25(17), 9366. [Google Scholar] [CrossRef] [PubMed]
- Reboldi, A.; Cyster, J. G. Peyer's patches: organizing B-cell responses at the intestinal frontier. Immunol. Rev. 2016, 271(1), 230–245. [Google Scholar] [CrossRef] [PubMed]
- Reichel, P. E.; Chakraborty, S.; Tay, H. L.; Hogan, S. P. Dietary Antigen Interaction with Intestinal Epithelial Cells. Curr. Allergy Asthma Rep. 2026, 26(1), 13. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M. The intestinal epithelial barrier in the control of homeostasis and immunity. Trends Immunol. 2011, 32(6), 256–264. [Google Scholar] [CrossRef] [PubMed]
- Rios, D.; Wood, M. B.; Li, J.; Chassaing, B.; Gewirtz, A. T.; Williams, I. R. Antigen sampling by intestinal M cells is the principal pathway initiating mucosal IgA production to commensal enteric bacteria. Mucosal Immunol. 2016, 9(4), 907–916. [Google Scholar] [CrossRef] [PubMed]
- Roffler, A. A.; Maurer, D. P.; Lunn, T. J.; Sironen, T.; Forbes, K. M.; Schmidt, A. G. Bat humoral immunity and its role in viral pathogenesis, transmission, and zoonosis. Front. Immunol. 2024, 15, 1269760. [Google Scholar] [CrossRef] [PubMed]
- Rosshart, S. P.; Vassallo, B. G.; Angeletti, D.; Hutchinson, D. S.; Morgan, A. P.; Takeda, K.; Hickman, H. D.; McCulloch, J. A.; Badger, J. H.; Ajami, N. J.; Trinchieri, G.; Pardo-Manuel de Villena, F.; Yewdell, J. W.; Rehermann, B. Wild Mouse Gut Microbiota Promotes Host Fitness and Improves Disease Resistance. Cell 2017, 171(5), 1015–1028.e13. [Google Scholar] [CrossRef] [PubMed]
- Ruddle, S. J.; Massis, L. M.; Cutter, A. C.; Monack, D. M. Salmonella-liberated dietary L-arabinose promotes expansion in superspreaders. Cell Host Microbe 2023, 31(3), 405–417. [Google Scholar] [CrossRef] [PubMed]
- Sarma, K.; Sasan, J. S.; Suri, S.; Devi, J.; Pura, R. S. Gut Associated Lymphoid Tissue (Peyer’s patches) in the intestine of the adult Bakerwali goat and non-descript goats of Jammu region. J. Livest. Sci. 2023, 14, 304–307. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R. M.; Angulo, F. J.; Tauxe, R. V.; Widdowson, M. A.; Roy, S. L.; Jones, J. L.; Griffin, P. M. Foodborne illness acquired in the United States--major pathogens. Emerg. Infect. Dis. 2011, 17(1), 7–15. [Google Scholar] [CrossRef] [PubMed]
- Schoultz, I.; Claesson, M. J.; Dominguez-Bello, M. G.; Fåk Hållenius, F.; Konturek, P.; Korpela, K.; Laursen, M. F.; Penders, J.; Roager, H.; Vatanen, T.; Öhman, L.; Jenmalm, M. C. Gut microbiota development across the lifespan: Disease links and health-promoting interventions. J. Intern. Med. 2025, 297(6), 560–583. [Google Scholar] [CrossRef] [PubMed]
- Scotto, G.; Fazio, V.; Moula Ali, A. M.; Bavisetty, S. C. B.; Franza, A.; Massa, S. Nipah Virus Encephalitis: Pathogenetic Aspects and Current Therapeutic Strategies. Pathogens 2026, 15(4), 443. [Google Scholar] [CrossRef] [PubMed]
- Seefeld, M. L.; Templeton, E. L.; Lehtinen, J. M.; Sinclair, N.; Yadav, D.; Hartwell, B. L. Harnessing the potential of the NALT and BALT as targets for immunomodulation using engineering strategies to enhance mucosal uptake. Front. Immunol. 2024, 15, 1419527. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Li, T.; Lai, K. K.; Johnson, R. F.; Yewdell, J. W.; Compton, A. A. Omicron Spike confers enhanced infectivity and interferon resistance to SARS-CoV-2 in human nasal tissue. Nat. Commun. 2024, 15(1), 889. [Google Scholar] [CrossRef] [PubMed]
- Shrwani, K. J.; Mahallawi, W. H.; Mohana, A. I.; Algaissi, A.; Dhayhi, N.; Sharwani, N. J.; Gadour, E.; Aldossari, S. M.; Asiri, H.; Kameli, N.; Asiri, A. Y.; Asiri, A. M.; Sherwani, A. J.; Cunliffe, N.; Zhang, Q. Mucosal immunity in upper and lower respiratory tract to MERS-CoV. Front. Immunol. 2024, 15, 1358885. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sanchez, A.; Randall, T. D. Anatomical Uniqueness of the Mucosal Immune System (GALT, NALT, iBALT) for the Induction and Regulation of Mucosal Immunity and Tolerance. In Mucosal Vaccines; 2020; pp. 21–54. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, A.; Sharma, S.; Sharma, A. Mucosal Vaccination in Veterinary Medicine. In Mucosal Vaccine Delivery Systems: The Future of Immunization (Part 2); Sharma, S., Alam, A., Sharma, A., Eds.; BENTHAM SCIENCE PUBLISHERS, 2025; pp. 107–140. [Google Scholar] [CrossRef]
- Siniscalco, E. R.; Williams, A.; Eisenbarth, S. C. All roads lead to IgA: Mapping the many pathways of IgA induction in the gut. In Immunological Reviews; WEBSITE:WEBSITE:PERICLES; STRING:PUBLICATION: WGROUP, 2024; Volume 326, 1, pp. 66–82. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Mehl, F.; Zeichner, S. L. Vaccine Strategies to Elicit Mucosal Immunity. Vaccines 2024, 12(2), 191. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.; Eyre, M. T.; Souza, F. N.; Diggle, P. J.; Ko, A. I.; Begon, M.; Costa, F. Disentangling the influence of reservoir abundance and pathogen shedding on zoonotic spillover of the Leptospira agent in urban informal settlements. Front. Public Health 2024, 12, 1447592. [Google Scholar] [CrossRef] [PubMed]
- Souza, J. M.; Ribeiro, P. H. C.; Millen, D. D. Review: Shifts of rumen microbiota by feeding non-fibrous carbohydrates to improve cattle performance. Front. Microbiol. 2026, 17, 1735296. [Google Scholar] [CrossRef] [PubMed]
- Suk, J. S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L. M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef] [PubMed]
- Tada, R.; Muto, S.; Iwata, T.; Hidaka, A.; Kiyono, H.; Kunisawa, J.; Aramaki, Y. Attachment of class B CpG ODN onto DOTAP/DC-chol liposome in nasal vaccine formulations augments antigen-specific immune responses in mice. BMC Res. Notes 2017, 10(1), 68. [Google Scholar] [CrossRef] [PubMed]
- Tardiolo, G.; La Fauci, D.; Riggio, V.; Daghio, M.; Di Salvo, E.; Zumbo, A.; Sutera, A. M. Gut Microbiota of Ruminants and Monogastric Livestock: An Overview. Animals 2025, Vol. 15(15(5)). [Google Scholar] [CrossRef]
- Taylor, L. H.; Latham, S. M.; Woolhouse, M. E. Risk factors for human disease emergence. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2001, 356(1411), 983–989. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, J.; Wu, L.; Zhang, C.; Zheng, F.; Yang, Y.; Lu, G.; Xie, D. Tissue-Resident Memory T Cells in Rheumatoid Immune Diseases: Pathogenic Mechanisms and Therapeutic Strategies. Biomedicines 2025, 13(12), 2945. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Arbab, S.; Tian, Y.; Chen, Y.; Liu, C. Q.; Li, Q.; Li, K. Crosstalk between gut microbiota and host immune system and its response to traumatic injury. Front. Immunol. 2024, 15, 1413485. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, L.; Lueder, Y.; Juergens, A. L.; Wilharm, A.; Barros-Martins, J.; Bubke, A.; Demera, A.; Ikuta, K.; Patzer, G. E.; Janssen, A.; Sandrock, I.; Prinz, I.; Rampoldi, F. IL-4-Producing Vγ1+/Vδ6+ γδ T Cells Sustain Germinal Center Reactions in Peyer's Patches of Mice. Front. Immunol. 2021, 12, 729607. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Meng, Q.; Chen, Y.; Liu, Y.; Li, X.; Zhou, J.; Ma, Y.; Yu, Z.; Chen, X. Interaction between gut microbiota and immunity in health and intestinal disease. Front. Immunol. 2025, 16, 1673852. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Amato, K. R.; Decaestecker, E.; McKenzie, V. J. Editorial: Impact of anthropogenic environmental changes on animal microbiomes. Front. Ecol. Evol. 2023, 11. [Google Scholar] [CrossRef]
- World Health Organization. Global antimicrobial resistance and use surveillance system (GLASS) report 2022. 2022. Available online: https://www.who.int/publications/i/item/9789240062702.
- Xie, D.; Lu, G.; Mai, G.; Guo, Q.; Xu, G. Tissue-resident memory T cells in diseases and therapeutic strategies. MedComm 2025, 6(1), e70053. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Jenne, C. N.; Kennedy, L. J.; Reynolds, J. D. The sheep and cattle Peyer's patch as a site of B-cell development. Vet. Res. 2006, 37(3), 401–415. [Google Scholar] [CrossRef] [PubMed]
- Yeung, F.; Chen, Y. H.; Lin, J. D.; Leung, J. M.; McCauley, C.; Devlin, J. C.; Hansen, C.; Cronkite, A.; Stephens, Z.; Drake-Dunn, C.; Fulmer, Y.; Shopsin, B.; Ruggles, K. V.; Round, J. L.; Loke, P.; Graham, A. L.; Cadwell, K. Altered Immunity of Laboratory Mice in the Natural Environment Is Associated with Fungal Colonization. Cell Host Microbe 2020, 27(5), 809–822.e6. [Google Scholar] [CrossRef] [PubMed]
- Youngblut, N.D.; Reischer, G.H.; Walters, W.; et al. Host diet and evolutionary history explain different aspects of gut microbiome diversity among vertebrate clades. Nat. Commun. 2019, 10, 2200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K. A.; Fang, X.; Wynne, J. W.; Xiong, Z.; Baker, M. L.; Zhao, W.; Tachedjian, M.; Zhu, Y.; Zhou, P.; Jiang, X.; Ng, J.; Yang, L.; Wu, L.; Xiao, J.; Feng, Y.; Chen, Y.; Wang, J. Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 2013, 339(6118), 456–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jiang, A.; Yu, H.; Dong, B. Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian. Int. J. Mol. Sci. 2023, 24(7), 6270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, Z.; Hong, W.; He, X.; Wei, X. Respiratory mucosal immunity: Biological functions, diseases, prevention and therapy. Genes Dis. 2026, 102200. [Google Scholar] [CrossRef]
- Zhang, Y.; Thomas, J. P.; Korcsmaros, T.; Gul, L. Integrating multi-omics to unravel host-microbiome interactions in inflammatory bowel disease. Cell Reports. Med. 2024, 5(9), 101738. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hong, W.; Zhang, Y.; Li, X.; Que, H.; Wei, X. Mucosal immunity and vaccination strategies: Current insights and future perspectives. Mol. Biomed. 2025, 6(1), 57. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ren, Z.; Xu, Q.; Zhu, T.; Hu, H.; Fu, Y.; Jiang, J.; Zhai, Q. Factors influencing gut microbial colonization: A host-microbe-environment interaction perspective. Curr. Res. Food Sci. 2026, 12, 101361. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30(6), 492–506. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, Y.; Zhu, Z.; Lu, C.; Zhang, C.; Zeng, L.; Xie, F.; Zhang, L.; Zhou, F. Mucosal immune response in biology, disease prevention and treatment. Signal Transduct. Target. Ther. 2025, 10(1), 7. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, Z.; Huang, S.; Shi, Y. Dietary fiber derived short-chain fatty acids as a critical driver of the gut–bone axis in animal bone health: A review. Anim. Nutr. 2025, 22, 242–258. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Ecological and microbial exposures shape mucosal immune adaptation through coordinated microbiota–immune interactions, barrier regulation, antigen sensing, and IgA-mediated microbial selection. Comparative differences in mucosal organization across mammalian species influence barrier homeostasis, microbial tolerance, pathogen persistence, and transmission dynamics.
Figure 1.
Ecological and microbial exposures shape mucosal immune adaptation through coordinated microbiota–immune interactions, barrier regulation, antigen sensing, and IgA-mediated microbial selection. Comparative differences in mucosal organization across mammalian species influence barrier homeostasis, microbial tolerance, pathogen persistence, and transmission dynamics.

Figure 2.
Emerging technologies in comparative mucosal immunology. Spatial transcriptomics, single-cell profiling, and integrative multi-omics approaches enable high-resolution analysis of tissue-specific microbiota–immune interactions, epithelial specialization, and compartmentalized immune organization across mammalian mucosal systems.
Figure 2.
Emerging technologies in comparative mucosal immunology. Spatial transcriptomics, single-cell profiling, and integrative multi-omics approaches enable high-resolution analysis of tissue-specific microbiota–immune interactions, epithelial specialization, and compartmentalized immune organization across mammalian mucosal systems.


Table 1.
Comparative organization and functional specialization of mucosal immune tissues across mammalian species.
Table 1.
Comparative organization and functional specialization of mucosal immune tissues across mammalian species.
| Mammalian Group | Representative Species | Mucosal Lymphoid Features | Lymphoid Organization | BALT/NALT Characteristics | Mucosal Immune Adaptations | Ecological and Microbial Influences | Translational Relevance | References |
| Ruminants | Cattle, sheep, goats | Extensive and highly developed GALT associated with continuous microbial and dietary antigen exposure | Continuous ileal Peyer’s patches function as primary lymphoid organs supporting antigen-independent B-cell diversification during early development | Constitutive BALT commonly observed | Strong mucosal IgA responses, extensive intraepithelial lymphocyte populations, tolerance-oriented intestinal immunity | Fermentation-associated microbial ecosystems shape immune maturation and epithelial homeostasis | Major reservoirs for enteric zoonotic pathogens including Salmonella enterica, enterohemorrhagic Escherichia coli, and Campylobacter jejuni | Brandtzaeg, 2013; Zheng et al., 2020 |
| Carnivores | Dogs, cats, wild canids and felids | Moderately developed GALT adapted to protein-rich diets and comparatively lower intestinal fermentation | Discrete Peyer’s patches and organized lymphoid follicles support localized antigen surveillance and IgA induction | Respiratory lymphoid tissues adapted for rapid inflammatory responses | Enhanced epithelial antimicrobial defense and tissue-resident immune responsiveness | Environmental exposure and dietary specialization shape intestinal microbial diversity | Comparative models for respiratory and gastrointestinal mucosal disease; potential zoonotic transmission interfaces | Zheng et al., 2020; Gray & Farber, 2022 |
| Rodents | Mice, rats | Highly characterized experimental mucosal immune systems containing organized GALT and diffuse leukocyte populations | Multiple Peyer’s patches support antigen sampling, germinal center formation, and mucosal B-cell activation | Constitutive BALT limited in healthy laboratory mice | Well-characterized tissue-resident lymphocyte populations and inducible mucosal immune responses | Specific pathogen-free housing alters microbial exposure and immune maturation relative to wild rodents | Widely used translational models, although ecological and immunological limitations remain | Reboldi & Cyster, 2016; Abolins et al., 2017; Rosshart et al., 2017 |
| Primates | Humans, non-human primates | Complex compartmentalized intestinal and respiratory mucosal immune networks | Peyer’s patches function as adaptive immune inductive sites supporting mucosal IgA production | NALT and inducible BALT contribute to respiratory immune surveillance | Highly specialized tissue-resident memory T-cell populations and compartment-specific immune regulation | Diet, environmental exposure, and microbiota diversity strongly influence mucosal immune homeostasis | Central to translational immunology, vaccine development, and emerging infectious disease susceptibility | Zheng et al., 2020; Gray & Farber, 2022 |
| Herbivorous Hindgut Fermenters | Horses, rabbits | Enlarged intestinal immune surveillance systems associated with hindgut fermentation | Peyer’s patches contribute to microbial monitoring and mucosal immune induction | Respiratory mucosal tissues adapted to environmental particulate exposure | Immune tolerance mechanisms support coexistence with dense fermentative microbiota | Fermentation-associated microbial ecosystems influence epithelial and immune regulation | Relevant for comparative gastrointestinal immunology and environmental pathogen exposure | Zheng et al., 2020; Arroyo Portilla et al., 2021 |
| Chiropterans (Bats) | Fruit bats, insectivorous bats | Comparative mucosal immune architecture remains poorly characterized despite major zoonotic relevance | Limited characterization of intestinal lymphoid organization and Peyer’s patch structure | Respiratory mucosal organization remains incompletely defined | Unique antiviral tolerance-associated innate immune regulation | High colony density, viral exposure, and metabolic adaptation likely shape mucosal immune specialization | Reservoir hosts for coronaviruses, filoviruses, and paramyxoviruses | Zhang et al., 2013; Pavlovich et al., 2018 |
| Wildlife Reservoir Mammals | Wild rodents, ungulates, mesocarnivores | Highly variable mucosal immune organization shaped by ecological exposure and chronic pathogen pressure | Heterogeneous lymphoid organization across species | Inducible mucosal lymphoid tissues frequently associated with environmental exposure | Balance between pathogen tolerance and inflammatory control | Natural microbial exposure generates immunologically experienced mucosal systems | Important reservoirs for vector-borne, enteric, and respiratory zoonotic pathogens | Krausgruber et al., 2020; Zheng et al., 2020 |
Table 2.
Mucosal determinants of pathogen persistence, shedding, and zoonotic transmission across host systems.
Table 2.
Mucosal determinants of pathogen persistence, shedding, and zoonotic transmission across host systems.
| Pathogen / pathogen group | Host system | Dominant mucosal / epithelial compartment | Key tropism determinants | Local mucosal immune determinants | Shedding route | Zoonotic / One Health implication | References |
| Influenza A viruses | Wild birds, swine, poultry, cattle, humans | Respiratory tract; avian intestinal tract; mammary epithelium in dairy cattle | α2,3- and α2,6-linked sialic acid receptor distribution; host proteases; tissue permissiveness | Airway interferons, mucosal antibodies, tissue-resident lymphocytes, epithelial restriction factors | Respiratory secretions, feces in birds, milk in infected dairy ruminants | Receptor compatibility and tissue tropism shape cross-species infection; swine may support reassortment, while dairy cattle H5N1 shows that secretory tissues can create unexpected transmission routes | Abdelwhab & Mettenleiter, 2023; Caserta et al., 2024; Eisfeld et al., 2024; Alkie et al., 2025; Bauer et al., 2026 |
| SARS-CoV-2 and related coronaviruses | Humans, bats, susceptible wildlife and domestic species | Nasal, airway, and respiratory epithelium | ACE2 distribution, spike activation, TMPRSS2 or alternative protease use, variant-specific epithelial adaptation | Type I/III interferons, airway IgA, mucosal memory B cells, TRM cells, mucins, antimicrobial peptides | Respiratory droplets and aerosols | Receptor presence alone is insufficient; productive spillover depends on tissue receptor expression, protease availability, immune evasion, and efficient replication in transmission-relevant mucosa | Lean et al., 2023; Shi et al., 2024; Noh & Rha, 2024; Peña-Hernández et al., 2024 |
| Nipah virus | Pteropusbats; spillover hosts including pigs and humans | Respiratory and gastrointestinal mucosa; secretory and excretory interfaces | Exposure route, epithelial access, viral entry compatibility, tissue permissiveness | Reservoir tolerance, innate antiviral regulation, local barrier immunity | Saliva, urine, feces; contaminated food or environmental interfaces | Spillover occurs when reservoir shedding intersects with human or livestock exposure. | Scotto et al., 2026 |
| Salmonella enterica | Livestock, poultry, humans, chronic carriers | Intestinal epithelium, Peyer’s patches, phagocytic cells | Adhesion, epithelial invasion, virulence systems, nutrient niche exploitation | Colonization resistance, microbiota-derived metabolites, antimicrobial peptides, Tregs, mucosal IgA | Fecal shedding and foodborne contamination | Disease and transmission diverge; tolerant superspreaders may shed high pathogen burdens with limited clinical signs | Lamichhane et al., 2024; Di Luccia et al., 2025. |
| Leptospira spp. | Rodents and other mammals | Renal tubules with environmental mucosal exposure through skin or mucosa | Renal colonization, environmental persistence, water-mediated exposure | Host tolerance, local inflammatory control, barrier breach during exposure | Urine and contaminated water or soil | Spillover reflects reservoir abundance, shedding intensity, rainfall, sanitation, and human environmental exposure | Soni et al., 2024 |
| Bat-borne RNA viruses | Bats | Respiratory, oral, gastrointestinal, and excretory mucosal interfaces | Viral compatibility with epithelial tissues; reservoir-specific immune regulation | Constitutive interferon activity, restrained inflammatory pathways, altered inflammasome/STING/PYHIN signaling | Saliva, urine, feces, respiratory secretions depending on virus | Reservoir competence depends on immune tolerance plus shedding at ecological interfaces linking bats to humans, livestock, or food systems | Irving et al., 2021; Roffler et al., 2024 |
| Companion animal respiratory viruses | Dogs and other companion animals | Upper and lower respiratory mucosa | Respiratory epithelial tropism and local replication competence | Mucosal antibodies, local cellular immunity, vaccine-induced respiratory protection | Respiratory secretions | Companion animals may act as bridge hosts or sentinels at household–farm–wildlife interfaces | Hwang et al., 2025; |
| Foodborne enteric pathogens broadly | Livestock, poultry, wildlife, humans | Gastrointestinal mucosa | Epithelial adherence, invasion, microbiota disruption, nutrient availability | Colonization resistance, IgA, antimicrobial peptides, epithelial barrier function, inflammatory tone | Fecal shedding, contaminated food, water, or farm environments | Transmission risk depends on intestinal persistence, shedding load, food-chain contamination, and host-management practices | Ruddle et al., 2023; Lamichhane et al., 2024; Kauer et al., 2025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



