Submitted:
11 June 2026
Posted:
12 June 2026
You are already at the latest version
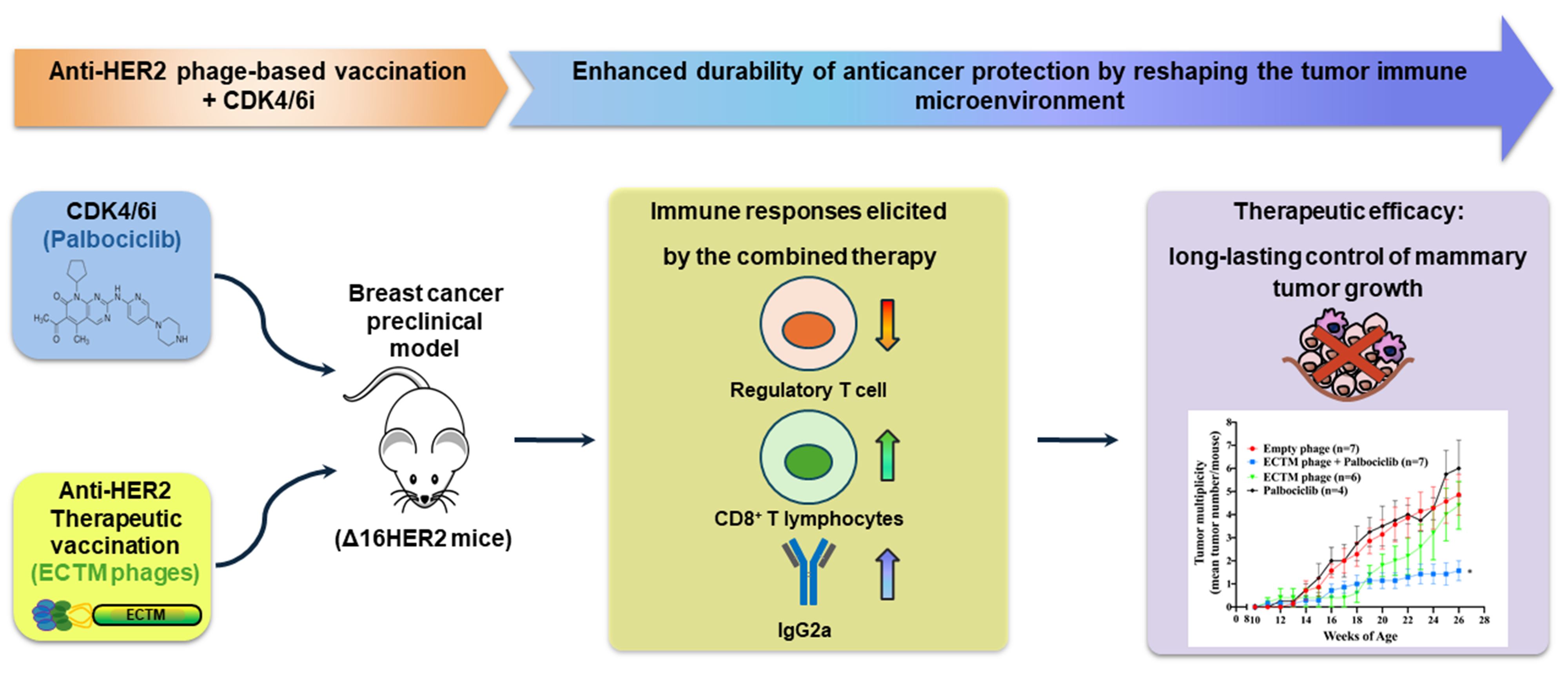
Abstract
Background/Objectives: Cancer vaccines represent the next frontier in immunotherapy, aiming to elicit long-lasting protective anti-tumor protective immune responses. Human epidermal growth factor receptor 2 (HER2) is a well-established therapeutic target in breast cancer. Active immunization with HER2-displaying M13 bacteriophages can induce a therapeutic immune response against HER2-positive breast cancer, offering a promising alternative to trastuzumab. However, the duration of anticancer immune protection triggered by anti-HER2 phage-based vaccines is limited by tumor-immune suppressive mechanisms. Methods: In this study, two vaccination cycles with ECTM phages displaying the extracellular (EC) and transmembrane (TM) domains of human HER2 were combined with palbociclib, a CDK4/6 inhibitor, to enhance antitumor im-munity in the clinically relevant Δ16HER2 transgenic preclinical model of breast cancer. Results: The proposed combination treatment resulted in a better and long-lasting control of tumor growth rate and multiplicity than either palbociclib or phage vaccination alone, correlating with a significantly stronger anti-HER2 humoral response (IgG2a isotype). Analysis of the tumor immune infiltrate revealed an increased presence of CD8⁺ T cells concomitant with a reduction in FoxP3⁺ regulatory T cells (Tregs) in tumors explanted from mice receiving the combination therapy. Conclusions: These preclinical results provide a rationale for the clinical translation of CDK4/6 inhibitors combined with anti-HER2 active immunotherapies in breast cancer, as they may yield sustained antitumor responses by reverting the immunosuppressive tumor environment.
Keywords:
bacteriophages
; breast cancer
; cancer vaccines
; HER2
; combination therapy
; CDK4/6 inhibitors
; palbociclib
1. Introduction
Global breast cancer incidence and associated mortality is projected to rise by 2050, underscoring the urgent need for novel and more effective therapeutic strategies [1]. The human epidermal growth factor receptor 2 (HER2) is overexpressed in 20% of breast cancers and it is associated with poor prognosis, although HER2-targeted therapies dramatically improved survival outcomes [2]. Indeed, the effectiveness of anti-HER2 targeted drugs, including the monoclonal antibody trastuzumab, is often compromised by drug resistance mechanisms [3]. Moreover, trastuzumab therapy is associated with cardiotoxicity [4], which increases significantly with age [5]. Anti-HER2 vaccination represents a promising alternative to therapeutic monoclonal antibodies, being potentially able to induce durable immunologic and clinical responses. However, cancer vaccine efficacy is limited by tumor immune suppressive mechanisms and currently no anti-HER2 cancer vaccine has been approved yet for human use [6]. Among cancer vaccine platforms, phage-displayed vaccines hold considerable potential, combining the intrinsic immunogenicity of viral particles with the high specificity of phage-displayed antigens [7,8,9]. In particular, M13 filamentous bacteriophages represent reliable immunogen carriers, as they are nonpathogenic and nonlytic viruses that infect and replicate exclusively in Escherichia coli cells harboring an F′ episome. Phages are internalized and processed by antigen-presenting cells, thereby inducing both B cell- and T cell-mediated immune responses, and they are immunogenic even in the absence of adjuvants [10]. Previous reports on anti-MAGE (Melanoma Antigen Gene) vaccination indicate that engineered filamentous bacteriophage virions can increase the immunogenicity of delivered tumor-associated antigens [11]. Phage vaccination was also shown to be safe and effective in evoking tumor-specific immune responses in malignant melanoma animal models [12,13] and multiple myeloma patients [14]. Consistently, we reported that vaccines based on filamentous bacteriophage M13 virions, engineered to display on their surface antigenic portions of HER2, can overcome tolerance against HER2 self-antigen and elicit a protective immune response. Indeed, they were able to prevent breast cancer development in Δ16HER2 mice, that spontaneously develop HER2-positive and estrogen receptor-positive mammary carcinomas [15,16] and exhibit immunological tolerance to human HER2 antigen [17]. However, when tested in therapeutic settings, the duration of anticancer immune protection triggered by these phage-based vaccines was limited by tumor immune evasion mechanisms [18]. Alterations of the cell cycle are present in virtually all human cancers. Cyclin E-cyclin-dependent kinase 2 (CDK2) and cyclin D-CDK4/6 complexes are crucial drivers of the cell cycle, as they promote the G1-to-S transition through phosphorylation of the retinoblastoma protein (pRb). Pharmacologic inhibitors of CDK4/6, by restoring the cell-cycle control, have shown significant activity against several tumors [19]. Recently, some CDK4/6 inhibitors, including palbociclib and abemaciclib, have been approved by the Food and Drug Administration (FDA) and the European Medicines Agency (EMA) for the treatment of hormone receptor (HR)-positive, HER2-negative breast cancer in combination with endocrine therapy, and are under clinical evaluation in other breast cancer subtypes [20]. In HER2-positive breast cancer, a combination of CDK4/6 inhibitors with HER2-targeted therapies showed promise in preclinical studies, and it is currently being tested in multiple clinical trials [21,22]. Of note, beyond inducing tumor cell cycle arrest, CDK4/6 inhibitors also promote antitumor immunity by increasing the functional capacity of tumor cells to present antigen and reducing the immunosuppressive regulatory T cells (Treg) population through the selective suppression of their proliferation [23]. Treg cells are a CD4+ T-cell subset essential for maintaining immune tolerance. In the tumor microenvironment, Treg cells contribute to tumor immune evasion and their downregulation by CDK4/6 inhibitors was demonstrated in both preclinical models and breast cancer patients [24,25]. Moreover, CDK4/6 inhibition skews newly activated CD8+T cells toward a memory phenotype as reported by Heckler et al. [25]. Thus, treatment with CDK4/6 inhibitors can re-educate the anti-cancer immune response in breast cancer patients by downregulating immunosuppressive Treg cells and activating effector T cells [24]. In this study, we demonstrate that combining a phage-based anti-HER2 vaccine with the CDK4/6 inhibitor palbociclib significantly improves the therapeutic control of HR⁺/HER2⁺ breast cancer in Δ16HER2 transgenic mice compared with either monotherapy. The combination treatment resulted in more durable tumor growth suppression and reduced tumor multiplicity, which were associated with enhanced anti-HER2 humoral immunity, decreased HER2 activation in mammary tumors, and a profound reshaping of the tumor immune microenvironment. These findings support the concept that CDK4/6 inhibition can enhance the efficacy of active anti-HER2 immunization by simultaneously overcoming immune tolerance to HER2 and mitigating tumor-driven immunosuppression. Another aspect that deserves to be clarified is the impact of phage-based vaccines on gut microbioma. Although it has been recently reported that a phage-based vaccine against SARS-CoV-2 did not affect the gut microbiota [26], phages can potentially manipulate it. This is a double-edged sword, because intestinal microbiota can influence vaccine responses [27] and enhance patient responses to immunotherapy [28]. In this study, ECTM-phage vaccine did not affect overall the relative abundance of most bacterial families. However, a selective increase in Lachnospiracea, positively correlated with a stronger therapeutic response to trastuzumab [29], was observed in vaccinated mice.
2. Materials and Methods
2.1. Phage Production and Purification
Engineered M13 bacteriophages, displaying on their surface the EC and TM domains of human HER2 (ECTM phages) in fusion with the coat protein pIII, were produced as previously described [17,18,30]. ECTM phages were released by TG1 bacterial cells, after transformation with recombinant phagemid vector pIF6 followed by isopropyl β-d-1-thiogalactopyranoside (IPTG) induction and super-infection with M13K07 helper phages. In brief, Escherichia coli TG1 cells were transformed with recombinant pIF6-phagemid, grown overnight in 2x TY medium with 100 µg/mL ampicillin and 1 % glucose at 37 °C, and then, when they reached the log phase of growth (optical density (O.D.) corresponding to 0.4 – 0.5 at 600 nm), they were superinfected with the helper phage M13K07 at a multiplicity of infection (M.O.I.) of 20:1, incubated for 30 minutes at 37 °C without shaking, followed by 30 minutes at 37 °C with shaking. TG1 cells were then centrifuged for 10 minutes at 2465 x g, and the cell pellet was resuspended in 2x TY medium supplemented with 100 µg/mL ampicillin, 70 µg/mL kanamycin and 200 µmol/L of IPTG and grown overnight at 30 °C. Polyethylene glycol (PEG)-based precipitation was used to purify M13 recombinant phages, which were harvested from the supernatant after centrifugation of the infected culture for 30 minutes at 352 x g. In details, phages were precipitated with 3:10 v/v ratio of PEG- NaCl (20 % PEG, 2.5 M NaCl) through incubation for 2 hours in ice, pelleted by centrifugation at 4 °C for 2 hours at 3220 x g and suspended in 2 mL of PBS. They were subsequently filtered through a 0.22 µm Millipore filter and then quantified by colony forming assay before being used as anti-HER2 vaccines in ∆16HER2 transgenic mice. The concentration of virion particles was verified by UV absorption spectrometry using the formula: virions/mL = [(A269 – A320) x 16 x 1016] / (number of bases/virion).
2.2. Mice Immunization and Palbociclib Treatment
Δ16HER2 transgenic mice (FVB/NHsd-Tg(D16HER2-LUC)6157Acam) were treated in accordance with the U.K. Animals (Scientific Procedures) Act, 1986 and associated guidelines, EU Directive 2010/63/EU for animal experiments and with the 3Rs principles. All animal experiments were authorized by the Italian Ministry of Health (#773/2023-PR) and by the Animal Research Committee of the University of Camerino (OPBA). Δ16HER2 female mice were vaccinated by intraperitoneal (i.p.) injection of ECTM-phages (1x1011 CFU/mouse in 0.1 mL PBS) at 12, 14, 16 weeks of age (first vaccination cycle), and at 20, 22, and 24 weeks of age (second vaccination cycle). Empty phages were used as control. Palbociclib (100 mg/Kg) was administered to Δ16HER2 mice by oral gavage with plastic feeding tubes for 5 days a week, from 15 to 24 weeks of age, alone or in combination with anti-HER2 phage vaccination. Blood was collected from the retro-orbital plexus under anesthesia, before and 2 weeks after the 3rd and the 6th booster doses, and left to clot at room temperature for 20 minutes. Serum was separated from blood by subsequent centrifugation at 3000 rpm at 4°C for 10 minutes and analyzed by FACS. Animals were weekly monitored for tumor onset, growth, and multiplicity by palpation until 26 weeks of age. Progressively growing masses 1mm mean diameters were regarded as tumors.
Body weight and food consumption did not significantly differ between treated and control mice (data not shown), suggesting the absence of a significant systemic toxicity of the administered therapies.
2.3. Cell Lines
SK-BR-3 were cultured in Dulbecco’s Modified Essential Medium (DMEM, CORNING, Mediatech, New York, NY, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, Life Technologies, Carlsbad, CA, USA) and 1% penicillin–streptomycin (P/S) (Gibco, Life Technologies). SK-BR-3 cells were kindly provided by the laboratory of Dr. B. Belletti (Division of Molecular Oncology, CRO of Aviano, IRCCS, National Cancer Institute, Aviano, Italy) and tested for mycoplasma contamination with negative results. CAM3 cells, Δ16HER2-expressing epithelial tumor cell lines established from a mammary carcinoma spontaneously arisen in a Δ16HER2 female [31], were cultured in DMEM supplemented with 20% FBS and 1% P/S. Cells were cultured at 37 °C under humidified atmosphere with 5% CO2.
2.4. Analysis of Antibody Response
Sera collected from immunized mice were analyzed by flow cytometry (BD FACSCalibur), using SK-BR-3 cells, to verify the presence of HER2-specific antibodies. Briefly, SK-BR-3 cells were detached and dispensed at a density of 0.5 x 106 cells per tube. After a 5-minute centrifugation at 800 rpm at 4°C, the obtained cell pellet was resuspended and washed twice in staining buffer (2% FBS-containing 1× PBS, pH 7.4, and 0.1% NaN3). Cells were incubated with sera of vaccinated mice (1:40 dilution in staining buffer) for 1 hour at 4°C. After incubation, cells were washed three times and incubated on ice in dark with goat anti-mouse IgG (H+L) secondary antibody-FITC (Thermo Fisher Scientific, 1:200 dilution in staining buffer). Samples were washed twice, resuspended in 600 μL of staining buffer and analyzed using BD FACSCalibur. Cell Quest Pro (version 6.0.2) and FlowJo version 8.7 were used as acquisition and analysis software, respectively. Analysis of anti-HER2 antibody titer and identification of different IgG isotypes (IgA, IgM, IgG1, IgG2a, IgG2b, and IgG3) in immune sera (diluted 1:200) was also performed by ELISA. Briefly, sera were incubated on plates coated with human HER2 (Sino Biological, 100 ng/well) protein, and binding was detected with HRP-conjugated rat anti-mouse-IgG (Merck) or with biotinylated rat anti-mouse IgG1, IgG2a, IgG2b or IgG3 (BD Biosciences), followed by HRP-conjugated streptavidin. Signals were developed with TMB (Tebu Bio) and analyzed using a 680XR microplate reader (BioRad). For antibody-dependent cellular cytotoxicity (ADCC), 1 x 104 CAM3 target cells stained with 2 µM CFSE (Molecular Probes) were cultured overnight with splenocytes from untreated wild-type FVB female mice, used as effector cells, at different effector:target (E:T) ratios (200:1, 100:1, and 50:1 E:T ratio) in the presence of a 1:50 dilution of sera from vaccinated mice. Cells were then collected and stained with 1 μg/mL 7-Amino-ActinomycinD (7-AAD, BD Biosciences), acquired by FACS on a BD FACSVerse and analyzed using Flowjo version 10. ADCC was calculated as described in [32].
2.5. Measurements of Cytokines in Sera
The pro- and anti-inflammatory cytokines IL-1α, IL-1β, IL-6, IL-10, IFNγ, and TNF-α were estimated in mouse sera by using multiplex immunoassay (Q-Plex Mouse Cytokine Panel (6-Plex), Quansys Biosciences, Technogenetics Srl., Milan, Italy), Q-View Imager LS, Q-View software Version 3.11, and following the manufacturer’s instructions.
2.6. Western Blot Analysis
Tumors were homogenized in RIPA buffer (0.1 % SDS, 1% NP40, 0.5 % CHAPS) supplemented with protease inhibitors (Sigma-Aldrich, St. Louis, MO). For western blot analysis, an equal amount of protein lysates was separated onto Criterion™ TGX™ precast gels (4-20 % gradient precast SDS-PAGE; Bio-Rad, Hercules, CA, USA) and transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore, Burlington, MA, USA) using Criterion™ Blotter (Bio-Rad). Membranes were blocked with EveryBlot Blocking Buffer (Bio-Rad, Hercules, CA, USA) and then incubated overnight with primary antibodies at 4 °C. Primary antibodies to vinculin (sc-73614), HER2 (sc-284, lot #I0507), and p-HER2 (sc-12352-R, lot #D2512) were from Santa Cruz Biotechnology. Secondary antibodies conjugated with peroxidase were from Sigma-Aldrich (Sigma-Aldrich/Merck, Darmstadt, Germany). Secondary antibody binding was performed at room temperature for 1 hour. After TBS-T washing, membranes were incubated with PierceTM ECL Western Blotting Substrate (Thermo Scientific, Boston, MA, USA), and the immunoreactive proteins were detected with ChemiDoc™ XRS-System (Bio-Rad, Hercules, CA, USA). Densitometry analysis was performed through ImageJ software (Version: 2.1.0/1.53C).
2.7. Histological Analysis
Heart, kidney and liver samples were fixed in 4% neutral buffered paraformaldehyde solution and embedded into paraffin. Slides were cut and stained with Hematoxylin (Bio-Optica) and Eosin (Bio-Optica) for histological examination. Section of heart were processed for Picro-Sirius Red (Direct Red 80, Sigma Aldrich) used for histological visualization of collagen I and III fibers to evaluate fibrotic process as previously described [33]. The sections were observed, and images were captured with the microscope by DS-Ri2 NIKON camera and area of fibrosis around cardiomyocyte was measured using a NIS Elements Nikon image analyzer software.
2.8. Immunohistochemical and Immunofluorescence Analyses
Tumor samples were fixed in 10% neutral buffered formalin and embedded into paraffin; slides were cut and stained with Hematoxylin (Bio-Optica) and Eosin (Bio-Optica) for histological examination. For immunohistochemistry or immunofluorescence, slides were deparaffinized, serially rehydrated and, after the appropriate antigen retrieval procedure, stained with the following primary antibodies: rabbit anti-mouse CD3 antibody (ab16669, Abcam), rabbit anti-mouse CD8 antibody (98941, Cell signaling) and rat anti-mouse FOXP3 antibody (14-5773-82, e-Bioscience). Immunostainings were developed with streptavidin peroxidase methods and the DAB Chromogen system (Dako). After chromogen incubation, slides were counterstained in Hematoxylin (Bio-Optica). For the evaluation of CD8 positive cells, whole-tumor sections were scanned using Nanozoomer scanner (Hamamatsu Photonics) and analysed using Qu-Path software (v0.3.2.). For immunofluorescence, immunostainings were developed using anti-rabbit Alexa Fluor 488 (A11008, ThermoFisher), anti-rat Alexa Fluor 546 (A11081, ThermoFisher) as secondary antibodies. Nuclei were stained with Dapi (Sigma). Images were acquired with a Zeiss LSM800 confocal microscope.
2.9. Microbial Community Analysis of the Mouse Gut Microbiome
Mouse feces (250-300 mg per each mouse) were dissolved in 0.5 mL of the SLX buffer supplied by the E.Z.N.A. Kit (Omega Bio-tek Inc., Norcross, GA, USA). After the addition of glass beads, samples were homogenized by vortexing for 10 minutes. Chromosomal DNA extraction was then completed according to the instructions given by the kit manufacturer. DNA concentration was estimated by NanoDrop (ThermoFisher Scientific, Waltham, MA, USA). The V3–V4 hypervariable regions of 16S rDNA were amplified by PCR using universal primers 341F 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG-3′ and 805R 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC-3′ and amplicon libraries were sequenced on an Illumina MiSeq platform (Illumina Inc., San Diego, CA, USA) using a 2 × 250 bp paired-end sequencing and following the manufacturer's standard protocols [34]. Briefly, raw sequencing reads were subjected to quality control to remove low-quality bases, adapter contamination, and short reads using Trimmomatic. High-quality paired-end reads were subsequently merged into full-length amplicon sequences using FLASH, based on the overlap between forward and reverse reads. The resulting merged sequences were further processed with VSEARCH to identify and remove potential chimeric sequences. After chimera filtering, sequences were clustered into Operational Taxonomic Units (OTUs) at a 97% sequence similarity threshold [35]. The NCBI 16S RefSeq [36] database was employed for taxonomic classification.
2.10. Statistical Analyses
Statistical analysis was carried out with GraphPad Prism 10 Software (San Diego, CA, USA), using the most appropriate test, as specified in each figure. p < 0.05 was used as the critical level of significance.
3. Results
3.1. Therapeutic Anti-HER2 Phage-Based Vaccination Delays Tumor Onset but Does Not Maintain Long-Term Control
In Δ16HER2 transgenic female mice, which spontaneously develop multiple HR⁺/HER2⁺ mammary adenocarcinomas starting at 12 weeks of age with a 100% penetrance [15,37], we first evaluated the therapeutic efficacy of an optimized anti-HER2 phage-based vaccination schedule. ECTM-phages displaying the EC and TM domains of human HER2 on M13 virions were administered intraperitoneally according to a 3×2 protocol, consisting of three doses at 12, 14 and 16 weeks of age followed by three additional doses at 20, 22 and 24 weeks (1×10¹¹ CFU/mouse per injection), while empty phages were used as control (Figure 1A). This protocol significantly delayed tumor onset and reduced tumor growth rate compared with control mice. At 16 weeks of age, 80% of vaccinated mice were tumor-free, whereas all control mice bore at least one palpable tumor (Figure 1B). Tumor multiplicity and mean tumor diameter were also lower in ECTM-vaccinated animals at 16–18 weeks, indicating a marked therapeutic effect (Figure 1C–D). However, this protection was not durable: by 20 weeks of age only 20% of vaccinated mice remained tumor-free, and by 25 weeks all immunized animals had developed multiple tumors, with tumor multiplicity no longer efficiently controlled (Figure 1B–C). Analysis of the humoral response showed that the first vaccination cycle induced a robust increase in anti-HER2 antibodies, as measured by flow cytometry on SK-BR-3 cells, with titers peaking two weeks after the third dose and declining thereafter (Figure 1E). The second vaccination cycle boosted anti-HER2 antibody levels again and the sera from vaccinated mice were able to mediate ADCC against Δ16HER2-derived CAM3 cells in vitro (Figure 1F). Multiplex cytokine analysis of sera collected two weeks after the second vaccination cycle revealed increased levels of IFN-γ and TNF-α, together with elevated IL-10 and IL-6, in ECTM-vaccinated mice compared with controls (Figure 1G).
3.2. Combination of ECTM-Phage Vaccination with Palbociclib Enhances Tumor Control and Modulates HER2 Signaling
We next investigated whether combining ECTM-phage vaccination with the CDK4/6 inhibitor palbociclib could improve therapeutic efficacy in Δ16HER2 mice. Palbociclib was administered by oral gavage at 100 mg/kg/day, 5 days per week, from 15 to 24 weeks of age, either alone or in combination with the ECTM vaccination following the 3×2 schedule; empty phages were used as control (Figure 2A). The start of palbociclib treatment was delayed relative to the first vaccine dose to avoid interference with T-cell expansion. Treatment with palbociclib alone did not significantly reduce tumor growth rate or tumor multiplicity compared with control mice (Figure 2B–C). In contrast, the combination of palbociclib with ECTM-phage vaccination resulted in a significantly better and more durable control of both tumor growth and multiplicity than either monotherapy. Tumor diameters in the combination group remained significantly lower than those in control and palbociclib-treated mice throughout the observation period, and tumor multiplicity was significantly reduced in the combination group compared with both control and palbociclib alone (Figure 2B–C). Measurement of anti-HER2 antibodies by ELISA in sera collected at 26 weeks of age showed that total anti-HER2 IgG titers were comparably increased in mice vaccinated with ECTM-phages alone and in those receiving the combination therapy, relative to empty-phage controls (Figure 2D). By contrast, analysis of IgG subclasses revealed that the combination regimen induced a higher proportion of IgG2a isotype antibodies compared with vaccination alone, whereas other isotypes were not markedly altered (Figure 2E). Western blot analysis showed that total HER2 levels were reduced in tumors explanted from mice receiving ECTM-phage vaccine alone or in combination with palbociclib, suggesting that anti-HER2 antibodies induced by vaccination could promote HER2 internalization and degradation. HER2 downregulation was also associated with a decreased HER2 activation, as indicated by the ratio of phosphorylated HER2 to total HER2 (Figure 2F–G).
To evaluate the toxicity of palbociclib and ECTM-phage vaccination, administered either as monotherapy or in combination, we carried out histopathological analyses of explanted kidneys, liver, and heart, common target organs of anticancer drugs. Histopathological analysis of liver (Figure S1A) and kidney (Figure S1B) tissues did not reveal any parenchymal alterations across the different experimental groups. Analysis of heart sections from ECTM-phage vaccinated mice did not show any signs of significant cardiotoxicity. However, heart sections from Δ16HER2 mice treated palbociclib, either alone or in combination with ECTM-phage vaccine, exhibited an increased fibrotic area characterized by collagen fiber deposition compared with control or ECTM-phage vaccinated mice (Figure S2A–B).
3.3. Combination Therapy Reshapes Intratumoral CD8⁺ T-Cell and Treg Compartments
To characterize the immune correlates of treatment efficacy, we analyzed tumor-infiltrating lymphocytes by immunohistochemistry and immunofluorescence. Immunostaining for CD8+ cells showed that tumors from mice treated with ECTM-phages or palbociclib alone displayed a modest increase in CD8⁺ T-cell infiltration compared with controls, whereas tumors from mice receiving the combination therapy exhibited a marked enrichment of CD8⁺ T cells within the tumor microenvironment (Figure 3A-D). Quantitative analysis confirmed that CD8⁺ T-cell density was significantly higher in the combination group than in control and palbociclib-treated mice (Figure 3E).
Treg cells were evaluated by confocal microscopy as CD3⁺FoxP3⁺ cells in peritumoral and intratumoral areas. In control tumors, Treg cells were mainly localized in the stromal area at the tumor border, with some infiltration into the tumor core (Figure 4A). ECTM-phage vaccination did not substantially reduce Treg abundance at the tumor margins but decreased the number of Treg cells within the tumor parenchyma (Figure 4B). Palbociclib treatment slightly reduced Treg cells at the tumor border and more markedly within the tumor core (Figure 4C). Notably, the combination of ECTM-phages and palbociclib led to a strong depletion of Treg cells, with only a few CD3⁺FoxP3⁺ cells detectable at the tumor border and virtually none within the tumor core (Figure 4D).
3.4. Anti-HER2 ECTM-Phage Vaccination Selectively Modulates the Gut Microbiome
Finally, we assessed the impact of ECTM-phage vaccination on the gut microbiome. 16S rRNA gene next-generation sequencing of fecal samples from control and vaccinated mice (n = 4 per group) showed that ECTM-phage treatment did not markedly alter overall bacterial family richness or the relative abundance of most families (Figure 5). However, vaccinated mice displayed a significant increase in the relative abundance of Lachnospiraceae and a concomitant decrease in Muribaculaceae compared with controls (Figure 5).
4. Discussion
Here, we show that CDK4/6 inhibition can enhance the therapeutic durability of anti-HER2 vaccination by reshaping the tumor immune microenvironment and by overcoming the tumor-induced immunosuppression. Indeed, the combination of a phage-based anti-HER2 vaccine with the CDK4/6 inhibitor palbociclib resulted in a long-lasting control of mammary tumor growth in Δ16HER2 transgenic mice, associated with enhanced intratumoral CD8⁺ T-cell infiltration and a profound depletion of regulatory T cells (Treg), while a robust anti-HER2 humoral responses was preserved.
4.1. Phage-Based Anti-HER2 Vaccination as a Pharmaceutically Relevant Platform
From a pharmaceutics perspective, filamentous M13 bacteriophages represent a versatile vaccine platform that combines high antigen density, intrinsic immunogenicity, and relative ease of large-scale production using Escherichia coli hosts. In our model, ECTM-phages displaying the extracellular and transmembrane domains of human HER2 [17,18] elicited robust HER2-specific antibody responses and significantly delayed tumor onset in Δ16HER2 mice, which are immunotolerant to human HER2 and spontaneously develop HR⁺/HER2⁺ mammary carcinomas [15,16,17,37]. These findings confirm and extend previous evidence that phage-based vaccines can break tolerance against self-tumor antigens and provide therapeutic benefit in established disease. However, despite optimization of the vaccination schedule and dose (3×2 protocol with increased phage load), ECTM-phage vaccination alone was ultimately unable to maintain long-term tumor control, as all immunized mice eventually developed multiple tumors. The kinetic analysis of the humoral response showed that anti-HER2 antibody titers peaked shortly after the first vaccination cycle and then declined, with a partial boost following the second cycle, paralleled by changes in pro- and anti-inflammatory cytokines. This transient pattern suggests that, in the context of an advanced, immunosuppressive tumor microenvironment, active vaccination alone is insufficient to sustain durable immune pressure on tumor cells. These observations underscore both the potential and the limitations of phage-based cancer vaccines as pharmaceutically relevant immunotherapeutics: while they offer an attractive, modular platform for antigen delivery, their full therapeutic impact in established tumors is likely to require rational combination with agents that recondition the host immune milieu.
4.2. CDK4/6 Inhibition Cooperates with Vaccination to Reprogram the Tumor Immune Microenvironment
CDK4/6 inhibitors were initially developed as cell cycle-targeted agents, but accumulating evidence indicates that they exert broad immunomodulatory effects, including enhancement of antigen presentation, promotion of memory-like CD8⁺ T cells, and selective inhibition of Treg cell proliferation [23,24,25]. In our study, palbociclib administered as monotherapy at a clinically relevant schedule did not significantly reduce tumor growth or multiplicity compared with controls in Δ16HER2 mice, representing an aggressive breast cancer preclinical model. Nonetheless, palbociclib alone modestly increased intratumoral CD8⁺ T-cell infiltration and reduced Treg abundance within the tumor core, in line with its reported capacity to modulate T-cell subsets [23]. When palbociclib was combined with ECTM-phage vaccination, these immune effects were markedly amplified. Tumors from mice receiving the combination therapy displayed an enrichment of CD8⁺ T cells and a depletion of CD3⁺FoxP3⁺ Treg cells at both the tumor border and within the tumor parenchyma. This shift in the effector/Treg balance was accompanied by a qualitative change in the humoral response, with a higher proportion of anti-HER2 IgG2a antibodies in the combination group compared with vaccination alone, an isotype associated with enhanced Fc-mediated effector functions in therapeutic settings [38]. Overall, these data support a model in which palbociclib creates a more permissive immune landscape by limiting Treg-mediated suppression and promoting CD8⁺ T-cell infiltration in which phage-induced anti-HER2 effector responses can be sustained and efficiently translated into superior tumor control. This cooperative interaction between cell cycle inhibition and active vaccination highlights the potential of CDK4/6 inhibitors as immunomodulatory partners for cancer vaccines, beyond their direct antiproliferative effects.
Analysis of organs toxicity revealed that both the CDK4/6 inhibitor palbociclib and ECTM phage vaccination were well tolerated, with no evidence of liver or kidney damage, supporting the acceptable safety profile of both therapeutic approaches. Of note, phage-based anti-HER2 vaccination was not associated with cardiotoxicity. In contrast, treatment with palbociclib, either alone or in combination with ECTM phage vaccination, was associated with increased cardiomyopathy characterized by collagen fiber deposition between cardiomyocytes. This finding may be related to the cardiovascular toxicity of CDK4/6 inhibitors previously reported in women with breast cancer undergoing pharmacological treatment [39].
4.3. Exploratory Evidence for Microbiome Modulation by Phage-Based Vaccination
An additional, exploratory aspect of our work concerns the impact of anti-HER2 phage vaccination on the gut microbiome. This part of the study did not include palbociclib as the main objective was to exclude harms to microbiome caused by the phage-based vaccine. ECTM-phage immunization did not markedly alter overall microbiome diversity or the relative abundance of most bacterial families, indicating that systemic administration of bacteriophages at the tested dose and schedule does not grossly disrupt gut microbial communities. However, we observed a selective increase in the relative abundance of Lachnospiracea and a decrease in Muribaculaceae in vaccinated mice compared with controls. Notably, Lachnospiraceae have been reported to be enriched in responders to Trastuzumab in HER2-positive breast and gastrointestinal cancers, suggesting a potential link between this family and improved outcome of HER2-targeted therapies [28,29]. Conversely, Muribaculaceae have been implicated in pro-inflammatory states [40], although their role in tumor immunity remains incompletely understood. Given the small sample size and the observational nature of the analysis, these findings should be interpreted as hypothesis-generating and do not allow causal inferences about the contribution of specific taxa to the efficacy of vaccination. Nevertheless, they raise the intriguing possibility that phage-based vaccines could exert dual immunomodulatory actions by directly priming anti-HER2 immune responses and indirectly shaping the gut microbiota in a manner that may favor antitumor immunity. Future studies integrating metagenomic and metabolomic profiling with longitudinal immune monitoring will be essential to clarify the interplay between bacteriophage-based vaccination, alone or in combination with CDK4/6 inhibitors, and the gut microbiome.
4.4. Limitations of the Study
This study has several limitations that should be acknowledged. First, all experiments were conducted in a single transgenic mouse model of HR⁺/HER2⁺ breast cancer, which, although highly relevant and well characterized, cannot fully recapitulate the molecular and immunological heterogeneity of human HER2-positive tumors. Second, we focused on a single CDK4/6 inhibitor (palbociclib) and one dosing schedule; whether other CDK4/6 inhibitors with distinct pharmacokinetic and pharmacodynamic profiles, or alternative sequencing regimens, might further optimize the balance between tumor cell-cycle arrest and T-cell expansion remains to be determined. Finally, the microbiome analysis was exploratory and limited to 16S rRNA gene sequencing at the family level in a small number of animals, precluding definitive conclusions about the biological significance of the observed compositional changes. Larger cohorts, higher-resolution microbiome profiling, and interventional approaches (e.g., fecal microbiota transfer or antibiotic modulation) will be required to validate the impact of phage-based vaccination on gut microbial communities and their relevance for antitumor immunity.
4.5. Translational Implications and Pharmaceutics Perspective
Despite these limitations, our findings have important translational implications. Palbociclib is already approved for HR⁺/HER2⁻ breast cancer and is under clinical evaluation in combination with HER2-targeted therapies in HER2-positive disease, providing a clinically tractable backbone for combinatorial strategies [41]. Our data suggest that integrating a phage-based anti-HER2 vaccine into CDK4/6 inhibitor–containing regimens may offer a means to induce durable, antigen-specific immune memory while simultaneously alleviating tumor-driven immunosuppression. This is a crucial point, considering that up to now clinical trials evaluating breast cancer vaccines have provided limited evidence of clinical benefits mainly due to the immune-suppressive microenvironment [42]. From a pharmaceutics standpoint, the use of filamentous phages as modular, genetically programmable vaccine carriers opens opportunities for scalable manufacturing, rapid antigen redesign, and flexible combination with existing targeted agents. Collectively, these results provide a preclinical rationale for the clinical translation of CDK4/6 inhibitors in combination with phage-based anti-HER2 active immunotherapies and motivate further investigation into the role of the gut microbiome as a potential modulator of treatment outcome.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Morphology of peripheral organs; Figure S2: Evaluation of cardiac fibrosis.
Author Contributions
Conceptualization, J.W., A.A., M.I., and C.M.; methodology, J.W., M.G.; formal analysis, J.W., A.L., M.I., L.C., D.T. and C.M.; investigation, J.W., A.L., D.T., L.C., C.C., A.D.L., M.G., D.L., F.O., D.B., VV..; resources, B.B., F.Olivieri, S.M.; writing—original draft preparation, J.W., M.F., M.I., C.M.; writing—review and editing, J.W., A.L., M.F., M.I., C.M., F.C., L.C., B.B.; supervision, C.M., M.I., F.C.; project administration, C.M..; funding acquisition, C.M., M.F. M.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by Italian Ministry of Health (#773/2023-PR) and by the Animal Research Committee of the University of Camerino (OPBA).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
J. Wang was supported by Fondazione Umberto Veronesi. The authors thank Andrea Amoroso (Charles River) at the Experimental Animal Models for Aging Unit, Scientific Technological Area, IRCCS INRCA, for his expert technical assistance in this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Freihat, O.; Sipos, D.; Kovacs, A. Global burden and projections of breast cancer incidence and mortality to 2050: a comprehensive analysis of GLOBOCAN data. Front Public Health 2025, 13, 1622954–1622966. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.M.; Shastry, M.; Hamilton, E. Targeting HER2-positive breast cancer: advances and future directions. Nat. Rev. Drug Discov. 2023, 22(2), 101–126. [Google Scholar] [CrossRef] [PubMed]
- Tabish, J.; Nusrat, J.; Shazia, S.; Gowhar, M.; Ayesha, J.; Showkat, A. A.; Manzoor, A. M. HER2-targeted therapy resistance in breast cancer: Molecular mechanisms, therapeutic evolution, and precision oncology approaches. Adv. Cancer Biol. – Metastasis 2026, 17, 100183. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, J.; Xie, Y.; Zhou, D.; Guo, M.; Qin, Y.; Yi, K.; Tian, J.; You, T. Interventions for prevention and treatment of trastuzumab-induced cardiotoxicity: an umbrella review of systematic reviews and meta-analyses. Front Pharmacol. 2024, 15, 1479983. [Google Scholar] [CrossRef] [PubMed]
- Dall, P.; Lenzen, G.; Göhler, T.; Lerchenmüller, C.; Feisel-Schwickardi, G.; Koch, T.; Eggert, J.; Heilmann, V.; Schindler, C.; Wilke, J.; et al. Trastuzumab in the treatment of elderly patients with early breast cancer: Results from an observational study in Germany. J. Geriatr. Oncol. 2015, 6, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Solinas, C.; Aiello, M.; Migliori, E.; Willard-Gallo, K.; Emens, L.A. Breast cancer vaccines: Heeding the lessons of the past to guide a path forward. Cancer Treat. Rev. 2020, 84, 101947–101965. [Google Scholar] [CrossRef] [PubMed]
- Ragothaman, M.; Yoo, S. Y. Engineered Phage-Based Cancer vaccines: current advances and future directions. Vaccines 2023, 11, 919–947. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, R.; D’Apice, L.; Prisco, A.; De Berardinis, P. Arming Filamentous Bacteriophage, a Nature-Made Nanoparticle, for New Vaccine and Immunotherapeutic Strategies. Pharmaceutics 2019, 11(9), 437–459. [Google Scholar] [CrossRef] [PubMed]
- Goracci, M.; Pignochino, Y.; Marchiò, S. Phage Display-Based nanotechnology applications in Cancer immunotherapy. Molecules 2020, 25(4), 843–871. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Shahgolzari, M.; Yavari, A.; Hefferon, K. Bacteriophages as Targeted Therapeutic Vehicles: Challenges and Opportunities. Bioengineering 2025, 12(5), 469–517. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, R.; Pisu, P.; D'Apice, L.; Pizzella, L.; Romano, C.; Cortese, G.; Giorgini, A.; Santoni, A.; Velotti, F.; De Berardinis, P. The use of filamentous bacteriophage fd to deliver MAGE-A10 or MAGE-A3 HLA-A2-restricted peptides and to induce strong antitumor CTL responses. J. Immunol. 2008, 180(6), 3719–3728. [Google Scholar] [CrossRef] [PubMed]
- Brišar, N.; Šuster, K.; Brezar, S.K.; Vidmar, R.; Fonović, M.; Cör, A. An Engineered M13 Filamentous Nanoparticle as an Antigen Carrier for a Malignant Melanoma Immunotherapeutic Strategy. Viruses 2024, 16(2), 232–248. [Google Scholar] [CrossRef] [PubMed]
- Brišar, N.; Šuster, K.; Brezar, S.K.; Cör, A. A virus based vaccine combined with IL12 gene therapy eradicates aggressive melanoma. Sci. Rep. 2025, 15(1), 18786–18798. [Google Scholar] [CrossRef] [PubMed]
- Roehnisch, T.; Then, C.; Nagel, W.; Blumenthal, C.; Braciak, T.; Donzeau, M.; Böhm, T.; Flaig, M.; Bourquin, C.; Oduncu, F.S. Phage idiotype vaccination: first phase I/II clinical trial in patients with multiple myeloma. J. Transl. Med. 2014, 12, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Marchini, C.; Gabrielli, F.; Iezzi, M.; Zenobi, S.; Montani, M.; Pietrella, L.; Kalogris, C.; Rossini, A.; Ciravolo, V.; Castagnoli, L.; et al. The human splice variant Δ16HER2 induces rapid tumor onset in a reporter transgenic mouse. PLoS One. 2011, 6(4), e18727. [Google Scholar] [CrossRef] [PubMed]
- Andreani, C.; Bartolacci, C.; Wijnant, K.; Crinelli, R.; Bianchi, M.; Magnani, M.; Hysi, A.; Iezzi, M.; Amici, A.; Marchini, C. Resveratrol fuels HER2 and ERα-positive breast cancer behaving as proteasome inhibitor. Aging 2017, 9(2), 508–523. [Google Scholar] [CrossRef] [PubMed]
- Bartolacci, C.; Andreani, C.; Curcio, C.; Occhipinti, S.; Massaccesi, L.; Giovarelli, M.; Galeazzi, R.; Iezzi, M.; Tilio, M.; Gambini, V.; et al. Phage-Based Anti-HER2 Vaccination Can Circumvent Immune Tolerance against Breast Cancer. Cancer Immunol. Res. 2018, 6(12), 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lamolinara, A.; Conti, L.; Giangrossi, M.; Cui, L.; Morelli, M.B.; Amantini, C.; Falconi, M.; Bartolacci, C.; Andreani, C.; et al. HER2-Displaying M13 Bacteriophages induce Therapeutic Immunity against Breast Cancer. Cancers 2022, 14, 4054–4071. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, A.; Rosen, L.S.; Tolaney, S.M.; Tolcher, A.W.; Goldman, J.W.; Gandhi; Papadopoulos, L.; Beeram, K.P.; Rasco, M.; Hilton, D.W.J.F.; et al. Efficacy and Safety of Abemaciclib, an Inhibitor of CDK4 and CDK6, for Patients with Breast Cancer, Non-Small Cell Lung Cancer, and Other Solid Tumors. Cancer Discov. 2016, 6(7), 740–753. [Google Scholar] [CrossRef] [PubMed]
- Shanabag, A.; Armand, J.; Son, E.; Yang, H.W. Targeting CDK4/6 in breast cancer. Exp. Mol. Med. 2025, 57(2), 312–322. [Google Scholar] [CrossRef] [PubMed]
- Gianni, L.; Bisagni, G.; Colleoni, M.; Del Mastro, L.; Zamagni, C.; Mansutti, M.; Zambetti, M.; Frassoldati, A.; De Fato, R.; Valagussa, P.; Viale, G. Neoadjuvant treatment with trastuzumab and pertuzumab plus palbociclib and fulvestrant in HER2-positive, ER-positive breast cancer (NA-PHER2): an exploratory, open-label, phase 2 study. Lancet Oncol. 2018, 19(2), 249–256. [Google Scholar] [CrossRef] [PubMed]
- Ciruelos, E.; Villagrasa, P.; Pascual, T.; Oliveira, M.; Pernas, S.; Paré, L.; Escrivá-de-Romaní, S.; Manso, L.; Adamo, B.; Martínez, E.; et al. Palbociclib and Trastuzumab in HER2-Positive Advanced Breast Cancer: Results from the Phase II SOLTI-1303 PATRICIA Trial. Clin. Cancer Res. 2020, 26(22), 5820–5829. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; DeCristo, M.J.; Watt, A.C.; BrinJones, H.; Sceneay, J.; Li, B.; Khan, N.; Ubellacker, J.M.; Xie, S.; Metzger-Filho, O.; et al. CDK4/6 inhibition triggers anti-tumour immunity. Nature 2017, 548(7668), 471–475. [Google Scholar] [CrossRef] [PubMed]
- Scirocchi, F.; Scagnoli, S.; Botticelli, A.; Di Filippo, A.; Napoletano, C.; Zizzari, I.G.; Strigari, L.; Tomao, S.; Cortesi, E.; Rughetti, A.; et al. Immune effects of CDK4/6 inhibitors in patients with HR+/HER2-metastatic breast cancer: Relief from immunosuppression is associated with clinical response. EBioMedicine 2022, 79, 104010. [Google Scholar] [CrossRef] [PubMed]
- Heckler, M.; Ali, L.R.; Clancy-Thompson, E.; Qiang, L.; Ventre, K.S.; Lenehan, P.; Roehle, K.; Luoma, A.; Boelaars, K.; Peters, V.; et al. Inhibition of CDK4/6 Promotes CD8 T-cell Memory Formation. Cancer Discov. 2021, 11(10), 2564–2581. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Jain, S.; Sha, J.; Batra, H.; Ananthaswamy, N.; Kilgore, P.B.; Hendrix, E.K.; Hosakote, Y.M.; Wu, X.; Olano, J.P.; et al. A Bacteriophage-Based, Highly Efficacious, Needle- and Adjuvant-Free, Mucosal COVID-19 Vaccine. mBio 2022, 13(4), e0182222. [Google Scholar] [CrossRef] [PubMed]
- Ardura-Garcia, C.; Curtis, N.; Zimmermann, P. Systematic review of the impact of intestinal microbiota on vaccine responses. npj Vaccines 2024, 9(1), 254. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Lau, H.C.; Yu, J. Modulating gut microbiome in cancer immunotherapy: Harnessing microbes to enhance treatment efficacy. Cell Rep. Med. 2024, 5(4), 101478. [Google Scholar] [CrossRef] [PubMed]
- Di Modica, M.; Arlotta, V.; Sfondrini, L.; Tagliabue, E.; Triulzi, T. The Link Between the Microbiota and HER2+ Breast Cancer: The New Challenge of Precision Medicine. Front Oncol. 2022, 12, 947188. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, F.; Salvi, R.; Garulli, C.; Kalogris, C.; Arima, S.; Tardella, L.; Monaci, P.; Pupa, S.M.; Tagliabue, E.; Montani, M.; et al. Identification of relevant conformational epitopes on the HER2 oncoprotein by using Large Fragment Phage Display (LFPD). PLoS ONE 2013, 8(3), e58358. [Google Scholar] [CrossRef] [PubMed]
- Tilio, M.; Gambini, V.; Wang, J.; Garulli, C.; Kalogris, C.; Andreani, C.; Bartolacci, C.; Elexpuru Zabaleta, M.; Pietrella, L.; Hysi, A.; et al. Irreversible inhibition of Δ16HER2 is necessary to suppress Δ16HER2-positive breast carcinomas resistant to Lapatinib. Cancer Lett. 2016, 381(1), 76–84. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Bolli, E.; Di Lorenzo, A.; Franceschi, V.; Macchi, F.; Riccardo, F.; Ruiu, R.; Russo, L.; Quaglino, E.; Donofrio, G.; Cavallo, F. Immunotargeting of the xCT Cystine/Glutamate Antiporter Potentiates the Efficacy of HER2-Targeted Immunotherapies in Breast Cancer. Cancer Immunol. Res. 2020, 8(8), 1039–1053. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Tomassoni, D.; Martinelli, I.; Bellitto, V.; Nittari, G.; Amenta, F.; Tayebati, S.K. Protective effects of the R-(+)-thioctic acid treatment: possible anti-inflammatory activity on heart of hypertensive rats. BMC Complement Med. Ther. 2024, 24(1), 281. [Google Scholar] [CrossRef] [PubMed]
- 16s Metagenomic Library Prep Guide. Part # 15044223 B. Available online: https://support.illumina.com/documents/documentation/ (accessed on 2 August 2024 and 18 November 2024).
- Petrilli, R.; Fabbretti, A.; Cerretani, A.; Pucci, K.; Pagliaretta, G.; Picciolini, M.; Napolioni, V.; Falconi, M. Selection, Identification and Functional Performance of Ammonia-Degrading Microbial Communities from an Activated Sludge for Landfill Leachate Treatment. Microorganisms 2023, 11(2), 311–326. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [PubMed]
- Segatto, I.; Zompit, M.M.; Citron, F.; D’Andrea, S.; Vinciguerra, G.L.R.; Perin, T.; Berton, S.; Mungo, G.; Schiappacassi, M.; Marchini, C.; et al. Stathmin Is Required for Normal Mouse Mammary Gland Development and Δ16HER2-Driven Tumorigenesis. Cancer Res. 2019, 79, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, N.; Segués, A.; Huang, S.; Waterfall, M.; Sijts, A.J.A.M.; Zaiss, D.M. Mouse IgG2a Isotype Therapeutic Antibodies Elicit Superior Tumor Growth Control Compared with mIgG1 or mIgE. Cancer Res. Commun. 2023, 3(1), 109–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, Z.; Chen, P.; Tian, L.; Fang, F.; Li, J. Cardiovascular toxicity of cyclin-dependent kinase 4 and 6 inhibitors. J. Cardio-Oncol. 1(1), 20–27. [CrossRef]
- Jang, S.; Kim, Y.J.; Park, J.; Kim, D.; Kim, T.H.; Lee, S.; Kim, D.J.; Ryu, C.M.; Seo, H.W. A Muribaculaceae-enriched microbiota exacerbates TLR4-dependent Acinetobacter baumannii-induced hyperinflammatory sepsis. Nat. Commun. 2026. [Google Scholar] [CrossRef] [PubMed]
- Metzger, O.; Mandrekar, S.; Goel, S.; Gligorov, J.; Lim, E.; Ciruelos, E.; Loibl, S.; Dockter, T.; Gonzàlez Farré, X.; et al. Palbociclib for Hormone-Receptor-Positive, HER2-Positive Advanced Breast Cancer. N Engl. J. Med. 2026, 394(5), 451–462. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Y.; Yu, K.D. Breast Cancer Vaccines: Disappointing or Promising? Front Immunol. 2022, 13, 28386. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Therapeutic phage-based vaccination (protocol 3x2) against HER2+/HR+ breast cancer in Δ16HER2 mice. A. Experimental workflow: ECTM-phages (1x1011 CFU/mouse) were injected intraperitoneally (i.p.) at 12, 14, 16 weeks of age (first vaccination cycle), and at 20, 22, and 24 weeks of age (second vaccination cycle). Empty phages were used as control. Bleeding occurred at the indicated times for antibody analysis. B. Kaplan–Meier curves; Log Rank test (* p< 0.05 ECTM versus (vs) empty phages (control)). C. Tumor multiplicity curves. D. Tumor growth curves; **** p < 0.0001 control vs. ECTM phages. E. Antibody detection. Sera of mice were collected at the indicated times and analyzed by FACS using SK-BR-3 cells; data represent Mean fluorescence intensity (MFI) ± SE; One-way ANOVA with Dunnett’s multiple comparison test (* p < 0.05 control vs ECTM phages (two weeks after the first vaccination cycle); * p < 0.05 control vs ECTM phages (two weeks after the second vaccination cycle)). F. ADCC assay was performed using CAM3 target cells incubated with a 1:50 dilution of sera from vaccinated mice (harvested at 26 weeks of age; n = 3) and splenocytes as effector cells at different effector/target cell ratios (200:1, 100:1 and 50:1). Results are shown as the mean ± SEM of the percentage of ADCC induced by different sera; Student’s t-test (* p < 0.05; ** p < 0.01). G. Quantification of cytokines in mouse sera by using a multiplex immunoassay. Sera were harvested from control and vaccinated mice, before immunization and two weeks after the second vaccination cycle. Data are shown as mean ± SEM. Unpaired t-test, control vs ECTM phage post-Vax (IL-6: p = 0.000069; IL-10 p = 0.000091; TNF-α p = 0.000095; IFN-γ p = 0.003474).
Figure 1.
Therapeutic phage-based vaccination (protocol 3x2) against HER2+/HR+ breast cancer in Δ16HER2 mice. A. Experimental workflow: ECTM-phages (1x1011 CFU/mouse) were injected intraperitoneally (i.p.) at 12, 14, 16 weeks of age (first vaccination cycle), and at 20, 22, and 24 weeks of age (second vaccination cycle). Empty phages were used as control. Bleeding occurred at the indicated times for antibody analysis. B. Kaplan–Meier curves; Log Rank test (* p< 0.05 ECTM versus (vs) empty phages (control)). C. Tumor multiplicity curves. D. Tumor growth curves; **** p < 0.0001 control vs. ECTM phages. E. Antibody detection. Sera of mice were collected at the indicated times and analyzed by FACS using SK-BR-3 cells; data represent Mean fluorescence intensity (MFI) ± SE; One-way ANOVA with Dunnett’s multiple comparison test (* p < 0.05 control vs ECTM phages (two weeks after the first vaccination cycle); * p < 0.05 control vs ECTM phages (two weeks after the second vaccination cycle)). F. ADCC assay was performed using CAM3 target cells incubated with a 1:50 dilution of sera from vaccinated mice (harvested at 26 weeks of age; n = 3) and splenocytes as effector cells at different effector/target cell ratios (200:1, 100:1 and 50:1). Results are shown as the mean ± SEM of the percentage of ADCC induced by different sera; Student’s t-test (* p < 0.05; ** p < 0.01). G. Quantification of cytokines in mouse sera by using a multiplex immunoassay. Sera were harvested from control and vaccinated mice, before immunization and two weeks after the second vaccination cycle. Data are shown as mean ± SEM. Unpaired t-test, control vs ECTM phage post-Vax (IL-6: p = 0.000069; IL-10 p = 0.000091; TNF-α p = 0.000095; IFN-γ p = 0.003474).

Figure 2.
Therapeutic phage-based vaccination in combination with palbociclib against HER2+/HR+ breast cancer in Δ16HER2 mice. A. Experimental workflow: palbociclib (100 mg/Kg) was administered daily by oral gavage from 15 to 24 weeks of age; ECTM-phages (1x1011 CFU/mouse) were injected intraperitoneally (i.p.) at 12, 14, 16 weeks of age (first vaccination cycle), and at 20, 22, and 24 weeks of age (second vaccination cycle), alone or in combination with palbociclib (combo); empty phages were used as control. Bleeding occurred at the indicated times for antibody analysis. B. Tumor growth curves; One-way ANOVA with Tukey’s multiple comparison test (** p < 0.01 control vs ECTM phages; ** p < 0.01 control vs combo). C. Tumor multiplicity curves; One-way ANOVA with Tukey’s multiple comparison test (* p < 0.05 combo vs control; ** p < 0.01 combo vs palbociclib). D. Anti-HER2 antibody detection. Sera of mice collected at 26 weeks of age were diluted 1:200 and analyzed by ELISA; data are expressed as mean ± SEM; Student’s t-test (** p< 0.01 empty phage (control) vs ECTM phages; ** p < 0.01 empty phage (control) vs combo). E. Analysis of antibody isotypes induced by vaccination with ECTM-phages alone or in combination with palbociclib (combo) (n = 3 mice per group) by ELISA. Sera were collected from mice after the second vaccination cycle (at 26 weeks of age) and diluted 1:200. Data are expressed as mean ± SEM. One-way ANOVA test followed by Tukey’s multiple comparison test (* p< 0.05 empty phage (control) vs combo). F. Representative western blot analysis of HER2, phospho (p)-HER2, and vinculin (loading control) in tumors explanted from 30-week-old Δ16HER2 mice receiving the indicated treatments (n = 3 mice/group). Equal amounts of protein (20 μg) were loaded. G. Densitometric quantification of HER2 expression normalized with vinculin (upper panel) and of pHER2/HER2 from two independent experiments were shown. Data are expressed as mean ± SEM. One-way ANOVA test followed by Tukey’s multiple comparison test (* p≤ 0.05; ** p ≤ 0.01).
Figure 2.
Therapeutic phage-based vaccination in combination with palbociclib against HER2+/HR+ breast cancer in Δ16HER2 mice. A. Experimental workflow: palbociclib (100 mg/Kg) was administered daily by oral gavage from 15 to 24 weeks of age; ECTM-phages (1x1011 CFU/mouse) were injected intraperitoneally (i.p.) at 12, 14, 16 weeks of age (first vaccination cycle), and at 20, 22, and 24 weeks of age (second vaccination cycle), alone or in combination with palbociclib (combo); empty phages were used as control. Bleeding occurred at the indicated times for antibody analysis. B. Tumor growth curves; One-way ANOVA with Tukey’s multiple comparison test (** p < 0.01 control vs ECTM phages; ** p < 0.01 control vs combo). C. Tumor multiplicity curves; One-way ANOVA with Tukey’s multiple comparison test (* p < 0.05 combo vs control; ** p < 0.01 combo vs palbociclib). D. Anti-HER2 antibody detection. Sera of mice collected at 26 weeks of age were diluted 1:200 and analyzed by ELISA; data are expressed as mean ± SEM; Student’s t-test (** p< 0.01 empty phage (control) vs ECTM phages; ** p < 0.01 empty phage (control) vs combo). E. Analysis of antibody isotypes induced by vaccination with ECTM-phages alone or in combination with palbociclib (combo) (n = 3 mice per group) by ELISA. Sera were collected from mice after the second vaccination cycle (at 26 weeks of age) and diluted 1:200. Data are expressed as mean ± SEM. One-way ANOVA test followed by Tukey’s multiple comparison test (* p< 0.05 empty phage (control) vs combo). F. Representative western blot analysis of HER2, phospho (p)-HER2, and vinculin (loading control) in tumors explanted from 30-week-old Δ16HER2 mice receiving the indicated treatments (n = 3 mice/group). Equal amounts of protein (20 μg) were loaded. G. Densitometric quantification of HER2 expression normalized with vinculin (upper panel) and of pHER2/HER2 from two independent experiments were shown. Data are expressed as mean ± SEM. One-way ANOVA test followed by Tukey’s multiple comparison test (* p≤ 0.05; ** p ≤ 0.01).

Figure 3.
CD8+ T cells evaluation on tumor sections by immunohistochemistry. Representative images of tumor sections: CD8+ T lymphocytes were stained in tumors explanted from Δ16HER2 mice receiving empty phages (control) (A), palbociclib (B) or ECTM-phages alone (C) or ECTM-phages in combination with palbociclib (combo) (D). Scale bar, 100 μm; original magnification, ×40. E. Quantification of CD8+ T cells in sections of tumors explanted from Δ16HER2 mice receiving the indicated treatments. ** p < 0.01 combo vs empty phage (control); * p < 0.05 combo vs palbociclib). Data are expressed as mean ± SEM; n = 4-6. .
Figure 3.
CD8+ T cells evaluation on tumor sections by immunohistochemistry. Representative images of tumor sections: CD8+ T lymphocytes were stained in tumors explanted from Δ16HER2 mice receiving empty phages (control) (A), palbociclib (B) or ECTM-phages alone (C) or ECTM-phages in combination with palbociclib (combo) (D). Scale bar, 100 μm; original magnification, ×40. E. Quantification of CD8+ T cells in sections of tumors explanted from Δ16HER2 mice receiving the indicated treatments. ** p < 0.01 combo vs empty phage (control); * p < 0.05 combo vs palbociclib). Data are expressed as mean ± SEM; n = 4-6. .

Figure 4.
Evaluation of Treg abundance in tumors by immunofluorescence. Representative images of immunofluorescence analysis of T lymphocytes and Tregs, stained for membranous CD3 (green) and nuclear Foxp3 (red), in tumors explanted from Δ16HER2 mice receiving empty phage (control) (A), ECTM-phage vaccine (B), palbociclib (C), or the combination therapy (combo) (D) (n = 4-6 tumors/group). Both peritumoral (upper panels) and intratumoral (lower panels) areas were analyzed and quantified (E,F). In the A-D left panels, the tumor border is indicated by the white line; the tumor core is indicated by the white asterisk, and Treg cells are circled; nuclei are stained with DAPI (blu). Scale bar 50 μm.
Figure 4.
Evaluation of Treg abundance in tumors by immunofluorescence. Representative images of immunofluorescence analysis of T lymphocytes and Tregs, stained for membranous CD3 (green) and nuclear Foxp3 (red), in tumors explanted from Δ16HER2 mice receiving empty phage (control) (A), ECTM-phage vaccine (B), palbociclib (C), or the combination therapy (combo) (D) (n = 4-6 tumors/group). Both peritumoral (upper panels) and intratumoral (lower panels) areas were analyzed and quantified (E,F). In the A-D left panels, the tumor border is indicated by the white line; the tumor core is indicated by the white asterisk, and Treg cells are circled; nuclei are stained with DAPI (blu). Scale bar 50 μm.

Figure 5.
Impact of ECTM-phage vaccination on gut microbiome. The abundance of bacterial families in the gut microbiome of control or ECTM-phage-vaccinated mice is shown. DNA was extracted from the fecal matter of individual mice (n = 4/group) left untreated (control) or immunized with ECTM-phage, and species identification was carried out by 16S rRNA gene next-generation sequencing (NGS). Data are expressed as mean ± SEM. Two-way ANOVA followed by Šidák’s multiple comparison test (*** p < 0.001; **** p < 0.0001 control vs ECTM-phage immunized group).
Figure 5.
Impact of ECTM-phage vaccination on gut microbiome. The abundance of bacterial families in the gut microbiome of control or ECTM-phage-vaccinated mice is shown. DNA was extracted from the fecal matter of individual mice (n = 4/group) left untreated (control) or immunized with ECTM-phage, and species identification was carried out by 16S rRNA gene next-generation sequencing (NGS). Data are expressed as mean ± SEM. Two-way ANOVA followed by Šidák’s multiple comparison test (*** p < 0.001; **** p < 0.0001 control vs ECTM-phage immunized group).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



