Submitted:
09 June 2026
Posted:
09 June 2026
You are already at the latest version
Abstract
Immune checkpoint inhibitors (ICIs) are now being introduced into perioperative treatment for several solid tumors. This strategy is usually explained by tumor reduction before surgery or by the elimination of minimal residual disease (MRD) after surgery. However, these explanations may not be sufficient to understand why the timing of ICI treatment, especially before lymph node (LN) removal, is important. In this review, we discuss tumor-draining lymph nodes (tdLNs) from two different aspects. tdLNs are anatomical routes for regional and distant metastasis, but they are also sites where tumor antigens are presented and tumor-specific T cell responses are generated. In particular, tdLNs may maintain stem-like or progenitor-exhausted CD8⁺ T cells, which can respond to PD-1 blockade and give rise to more differentiated exhausted T cells in the tumor. From this viewpoint, neoadjuvant ICI may be effective because the primary tumor, antigen flow, dendritic cell migration, and tdLN-based T cell priming are preserved. We also discuss the possible role of adjuvant ICI in controlling MRD, micrometastases, and metastatic-site draining LNs, and consider future implications for LN surgery and postoperative immune surveillance.
Keywords:
tumor-draining lymph node
; perioperative immunotherapy
; immune checkpoint inhibitor
; neoadjuvant immunotherapy
; adjuvant immunotherapy
; T cell exhaustion
; progenitor-exhausted T cells
; TPEX/TEX
; lymph node surgery
; immune surveillance
1. Introduction: Perioperative Immunotherapy Across Solid Tumors
Immune checkpoint inhibitors (ICIs) have become an important component of cancer treatment, initially in advanced or metastatic disease and more recently in earlier-stage, potentially curable tumors. This shift has led to growing interest in neoadjuvant and perioperative immunotherapy across several solid tumors. Conventional neoadjuvant chemotherapy or radiotherapy is generally used to reduce tumor burden and facilitate definitive local treatment. In contrast, neoadjuvant ICI may have an additional immunological role because it is administered while the primary tumor, tumor antigens, lymphatic drainage, and regional immune structures are still present [1,2,3]. This treatment window also allows early pathological response assessment and analysis of tumor tissues during therapy, providing information on mechanisms of response and resistance [4].
Clinical evidence for perioperative ICI has increased in multiple tumor types. In melanoma, the SWOG S1801 trial showed that pembrolizumab given before and after surgery improved event-free survival compared with the same drug given only in the adjuvant setting, indicating that treatment timing itself can affect clinical outcome [5]. The NADINA trial further supported the value of a neoadjuvant approach by showing favorable outcomes with neoadjuvant ipilimumab plus nivolumab followed by surgery and response-driven adjuvant therapy in resectable stage III melanoma [6]. In non-small cell lung cancer (NSCLC), KEYNOTE-671 demonstrated clinical benefit from perioperative pembrolizumab combined with chemotherapy in resectable disease [7]. Together, these findings suggest that perioperative ICI is not a disease-specific strategy, but an emerging therapeutic framework for solid tumors [4].
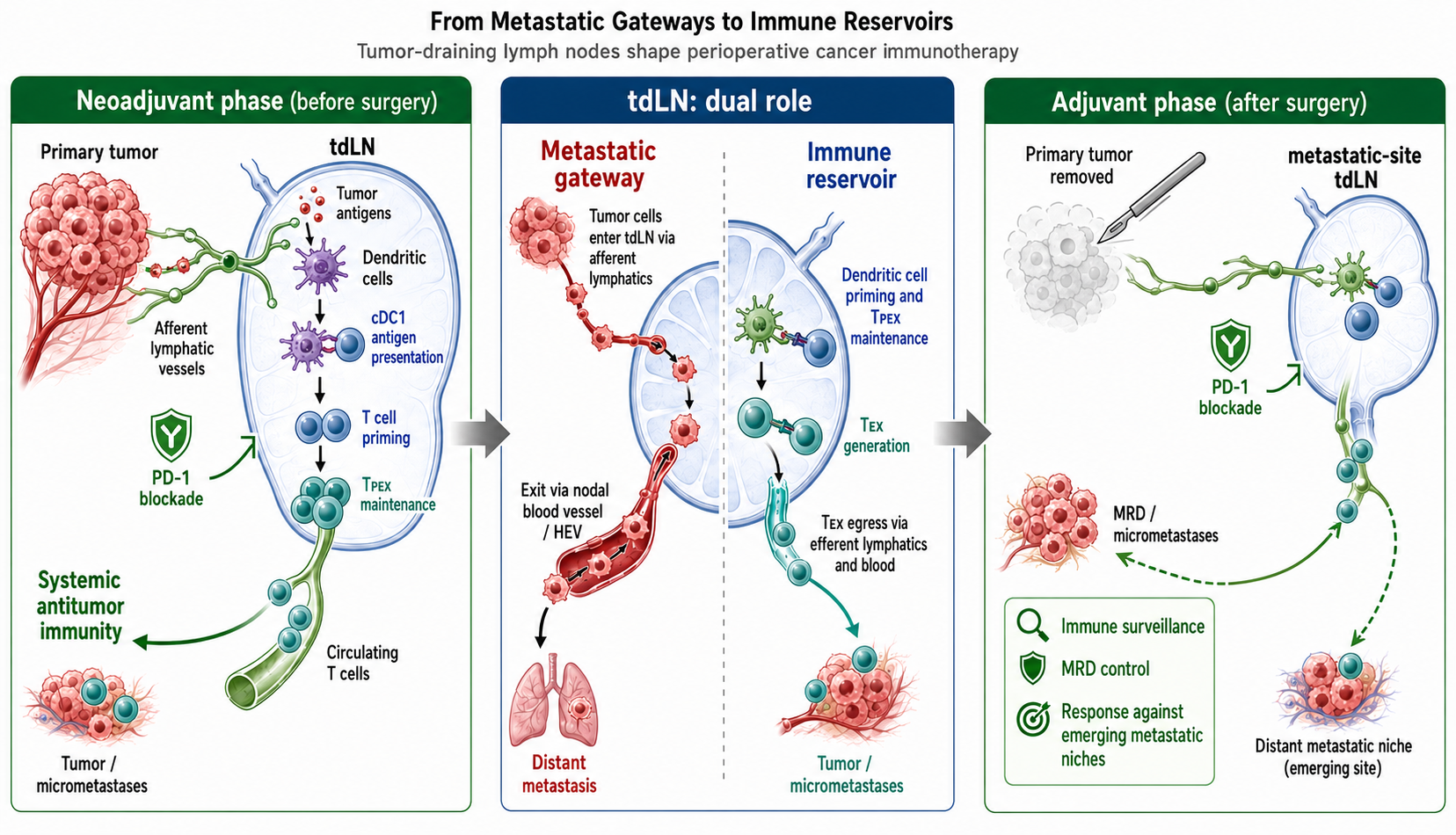
These clinical results raise an important mechanistic question: why should the administration of ICIs before the removal of the primary tumor and regional lymph nodes (LNs) be advantageous? One possible explanation is that the primary tumor and tumor-draining lymph nodes (tdLNs) are not merely surgical targets, but also active sites involved in the generation and regulation of antitumor immunity [8,9,10]. tdLNs are traditionally regarded as sites of lymphatic metastasis, and experimental studies have shown that tumor cells within LNs can enter nodal blood vessels and disseminate to distant organs [11,12]. At the same time, tdLNs are sites where tumor antigens are presented and tumor-specific T cell responses are initiated or maintained. This dual role of tdLNs as metastatic gateways and immune reservoirs is illustrated in Figure 1. In particular, studies of PD-1/PD-L1 blockade have shown that tdLNs can be critical for therapeutic efficacy and that checkpoint interactions within tdLNs regulate antitumor T cell immunity [8,9]. Therefore, perioperative immunotherapy should be considered not only as systemic drug treatment given around surgery, but also as a timing-dependent immune intervention that is influenced by whether the primary tumor and tdLN network remain intact.
2. tdLNs as Metastatic Gateways
Lymphatic spread is a common route of dissemination in many solid tumors. Tumor cells can access lymphatic vessels in the primary tumor or peritumoral tissue and are transported by lymphatic flow to regional draining LNs. This process is not only a passive movement of cancer cells. Tumor-derived signals, including lymphangiogenic factors, can remodel lymphatic vessels and alter the premetastatic environment of draining LNs. As a result, tdLNs may provide a specialized niche in which disseminated tumor cells (DTCs) are retained, adapt to local stromal and immune conditions, and develop into nodal metastases [13,14].
In clinical oncology, LN metastasis has long been used as an indicator of disease progression, prognosis, and treatment selection. Regional LNs are therefore important targets for surgical dissection and radiotherapy. However, this conventional view does not fully capture the biological role of nodal metastases. Recent experimental studies suggest that tumor cells growing in LNs may acquire access to the nodal vasculature and subsequently contribute to distant organ seeding [11]. In a related study, LN blood vessels, including high endothelial venules (HEVs), were shown to serve as a vascular route by which tumor cells can leave the LN and enter the systemic circulation [12]. These findings indicate that nodal metastases may function not only as regional disease deposits, but also as intermediates between lymphatic and hematogenous dissemination. Achen and Stacker integrated these observations into a model in which tumor cells enter LNs through afferent lymphatics, move through the nodal tissue, approach blood vessels within the node, and then disseminate systemically [15]. This model is useful for understanding tdLNs as anatomical structures that connect local tumor drainage, regional metastasis, and distant spread. Although the relative contribution of LN-derived dissemination may vary among tumor types and clinical settings, the concept broadens the role of tdLNs beyond that of passive filters or staging markers.
This metastatic function of tdLNs has direct relevance to perioperative immunotherapy. The same lymphatic route that allows tumor cells to reach regional LNs also carries tumor antigens and antigen-presenting cells to immune priming sites. Thus, tdLNs have a dual role. They can support tumor progression as metastatic intermediates, but they can also participate in the induction and maintenance of antitumor immunity. This dual nature creates an important therapeutic dilemma in the ICI era. Removal of metastatic LNs remains necessary for oncologic control, but the timing and extent of LN removal may influence the immune responses generated during neoadjuvant treatment. Understanding tdLNs as metastatic gateways is therefore essential before considering their role as immune reservoirs.
3. tdLNs as Immune Reservoirs
The role of tdLNs is not limited to metastatic spread. The lymphatic route that permits tumor cells to reach regional LNs also delivers soluble tumor antigens, cell debris, and antigen-bearing dendritic cells (DCs) from the tumor to the draining LN. In this anatomical setting, tdLNs provide a structured environment in which antigen-presenting cells interact with T cells and initiate tumor-specific immune responses. Therefore, tdLNs should be considered not only as possible sites of tumor cell deposition, but also as important sites for the generation and maintenance of systemic antitumor immunity [10,16].
DCs are central to this process. Among them, conventional type 1 DCs (cDC1s) are particularly important for CD8⁺ T cell immunity because they efficiently present tumor-derived antigens through the MHC class I pathway. Tumor antigens may be acquired in the tumor microenvironment and then carried to tdLNs by migratory DCs, or they may be transferred to LN-resident antigen-presenting cells. Within tdLNs, cDC1-dependent antigen presentation supports the activation and expansion of tumor-reactive CD8⁺ T cells. In addition to their role in initial priming, cDC1s also help preserve a population of proliferative, tumor antigen-specific TCF1+ CD8⁺ T cells in tdLNs, suggesting that DC–T cell interactions in LNs contribute to the durability of antitumor immunity [17,18].
Recent studies have shown that tdLNs contain CD8⁺ T cell populations exhibiting stem-like features, known as progenitor-exhausted T cells (TPEX). These cells retain proliferative potential and differ from more differentiated exhausted T cells (TEX) that accumulate within tumors. They are important because they can serve as a source of newly generated effector-like progeny during an ongoing antitumor response. Thus, tdLNs are not simply places where T cell priming begins. They also provide a niche in which less differentiated tumor-specific T cells can be maintained before giving rise to more differentiated T cells that migrate to tumor sites [19].
This function is closely linked to the activity of PD-1/PD-L1 blockade. Studies in experimental tumor models have shown that intact tdLNs are required for optimal therapeutic effects of PD-1/PD-L1 blockade [8]. Other work has demonstrated that checkpoint interactions within tdLNs can suppress tumor-specific T cell responses, indicating that the draining LN itself is an important site of action for immune checkpoint therapy [9]. In addition, tumor-specific memory-like CD8⁺ T cells in draining LNs, now often interpreted as TPEX, have been shown to respond to PD-1/PD-L1 blockade and contribute to antitumor immunity [20].
Taken together, these findings support the view that tdLNs act as immune reservoirs. They receive tumor antigens and DCs, support cDC1-dependent CD8⁺ T cell priming, maintain less differentiated TCF1+ tumor-specific T cells, and provide a cellular source that can be mobilized by PD-1/PD-L1 blockade. This concept is important for perioperative immunotherapy because neoadjuvant ICI is administered while the primary tumor, antigen flow, lymphatic drainage, and tdLN-based immune reactions are still preserved.
4. The TPEX/TEX Axis as the Cellular Basis of PD-1 Blockade
T cell exhaustion was originally described as a state induced by persistent antigen stimulation, particularly in chronic viral infection and cancer. It is characterized by sustained expression of inhibitory receptors, altered transcriptional regulation, reduced cytokine production, impaired proliferative capacity, and metabolic adaptation. However, exhaustion should not be understood simply as irreversible T cell failure. Rather, it represents a heterogeneous differentiation program that allows antigen-experienced T cells to persist under conditions of chronic stimulation, although their function and developmental potential change over time [21,22,23].
A key advance in this field has been the recognition that exhausted CD8⁺ T cells are not a uniform population. Less differentiated TPEX cells retain self-renewal capacity and can generate more differentiated progeny. These cells commonly express transcriptional regulators such as TCF1 and show greater proliferative potential than terminally TEX cells. In contrast, more differentiated TEX cells are enriched within the tumor microenvironment, display stronger cytotoxic or effector-associated features, and are positioned closer to direct tumor cell killing. However, they have limited ability to expand and are more prone to terminal dysfunction under persistent antigen exposure [23,24,25,26]. This differentiation hierarchy and its relationship to PD-1 blockade are summarized in Figure 2.
This developmental organization is important for understanding the mechanism of PD-1 blockade. Earlier interpretations often emphasized the functional recovery of TEX cells already present in tumors. This view is partly correct, but it is incomplete. Studies in chronic antigen models and cancer models have shown that the cells most capable of expanding after PD-1 pathway inhibition are not necessarily the most terminally differentiated TEX cells. Instead, the response is largely supported by TPEX cells or less differentiated exhausted T cells with preserved proliferative potential [24,25,27]. These cells can expand after checkpoint blockade and supply more differentiated effector-like progeny that participate in tumor control.
At the same time, PD-1 blockade does not fully reset terminally TEX cells to a naïve or memory-like state. TEX cells acquire transcriptional and epigenetic features that are only partially reversible, and this limits the durability of functional restoration after PD-1 inhibition [21,28]. Therefore, the therapeutic effect of PD-1 blockade should be viewed less as a complete rejuvenation of terminal TEX cells and more as a shift in the balance of the TEX compartment. By releasing inhibitory signaling, PD-1 blockade can increase the contribution of TPEX cells, promote their expansion, and enhance their differentiation into effector-like TEX cells.
This interpretation has direct relevance to perioperative immunotherapy. If the antitumor response depends on the continued presence of a TPEX pool, then the anatomical sites that maintain these cells become important. tdLNs, tertiary lymphoid structures (TLS), and selected intratumoral niches may all contribute to the preservation and differentiation of less exhausted tumor-specific CD8⁺ T cells. In this model, the efficacy of PD-1 blockade depends not only on the number of TEX cells within the tumor, but also on whether a renewable source of tumor-reactive T cells is available. The TPEX/TEX axis therefore provides a cellular framework for understanding why immune activation before removal of the primary tumor and tdLN network may be clinically important.
5. Why Neoadjuvant ICI Works: Activating Immunity Before Tumor and tdLN Removal
The advantage of neoadjuvant immune checkpoint inhibition cannot be explained only by reduction of tumor volume before surgery. A more important feature may be the timing of immune activation. Before resection, the primary tumor remains as a source of tumor antigens, lymphatic drainage from the tumor bed is still preserved, DCs can traffic to tdLNs, and regional LNs can still support tumor-specific T cell priming. Therefore, neoadjuvant ICI is administered at a stage when the primary tumor, draining LNs, and lymphatic connections that support systemic antitumor immunity remain anatomically and immunologically intact [1,3].
This timing may explain why neoadjuvant ICI can generate broader immune effects than treatment given only after surgery. In preclinical models, immunotherapy before tumor removal showed superior control of distant disease compared with the same treatment given after surgery, suggesting that the presence of the tumor during treatment can be immunologically useful rather than merely detrimental [29]. Clinical and translational studies also support this concept. In melanoma, neoadjuvant checkpoint blockade induced stronger tumor-infiltrating T cell responses and greater TCR clonal expansion than adjuvant treatment alone [30]. In resectable NSCLC, neoadjuvant PD-1 blockade was associated with early expansion of tumor mutation-associated T cell clones in peripheral blood, indicating that local treatment of the tumor-bearing host can produce systemic immune changes [31].
The role of tdLNs is especially important in this context. If PD-1/PD-L1 blockade acts, at least in part, by enhancing immune reactions within tdLNs, then surgical removal of these LNs before immune activation may reduce the opportunity to generate or amplify tumor-specific T cell responses. Experimental studies have shown that tdLNs are required for optimal responses to PD-1/PD-L1 pathway inhibition [8]. In addition, PD-1/PD-L1 interactions within tdLNs can restrain antitumor T cell immunity, supporting the idea that the draining LN is not simply a metastatic site but also a site where checkpoint blockade can act [9].
From this perspective, neoadjuvant ICI should be viewed as an intervention that uses the intact tumor–lymphatic–tdLN axis to initiate or amplify systemic antitumor immunity before surgery. This does not mean that all regional LNs should be preserved when they contain clinically significant metastases. Rather, it suggests that the timing of ICI relative to tumor resection and LN dissection may influence the quality and magnitude of the immune response. The main value of neoadjuvant ICI may therefore lie not only in preoperative tumor shrinkage, but also in the generation of systemic immune pressure against residual tumor cells, micrometastases, and early metastatic niches.
The immunological advantage of neoadjuvant ICI is therefore closely related to the preservation of the primary tumor, lymphatic drainage, DC migration, and tdLN-based T cell priming before surgery. In contrast, the role of adjuvant ICI must be understood in a different postoperative context, in which macroscopic disease has been removed but MRD, dormant tumor cells, and micrometastatic lesions may persist. These distinct roles of neoadjuvant and adjuvant ICI are summarized in Figure 3.
6. Why Adjuvant ICI Matters: MRD, Micrometastases, and Metastatic-Site tdLNs
As summarized in Figure 3, adjuvant immune checkpoint inhibition acts in a different anatomical and immunological context from neoadjuvant treatment. After curative-intent surgery, visible tumor lesions and regional tdLNs may have been removed. However, surgical clearance of macroscopic disease does not necessarily mean biological eradication of cancer. Minimal residual disease (MRD) may persist as circulating tumor cells (CTCs), DTCs, or small micrometastatic deposits that remain below the limit of clinical detection. These residual cancer cells provide the substrate for later relapse and distant metastasis, but their interaction with the immune system is likely to be variable and highly dependent on anatomical site, antigen availability, and local inflammatory context [32].
A very small number of transformed tumor cells or DTCs may not immediately induce a productive adaptive immune response. For T cell immunity to be initiated, tumor antigens must be released, captured by antigen-presenting cells, transported to an appropriate lymphoid site, and presented in a context that supports T cell priming rather than tolerance. Experimental studies have shown that insufficient antigen availability can result in immunological ignorance, whereas increased antigen expression can overcome this barrier [33]. In addition, sporadic tumors may induce tolerance even when they express recognizable antigens [34]. These observations are consistent with the cancer-immunity cycle and cancer immunoediting models, in which antigen release, LN priming, effector T cell trafficking, immune equilibrium, and immune escape occur as dynamic and sequential processes rather than as a single event [35,36].
Tumor dormancy further complicates the timing of adjuvant immunotherapy. DTCs can remain clinically silent for long periods, either as non-proliferating cells, as small lesions restricted by the microenvironment, or as tumor cell populations held in partial check by immune pressure [37,38]. Recent work has also suggested that dormant DTCs may avoid immune attack in part because they are rare and therefore infrequently encountered by effector immune cells [39]. From this perspective, adjuvant ICI may not act at one defined moment after surgery. Instead, it may provide sustained immune pressure during a period in which dormant or slowly expanding tumor cells intermittently become immunologically visible.
This concept is also relevant to metastatic-site draining LNs. If micrometastatic lesions grow in distant organs, they may establish new routes of antigen drainage to LNs located outside the original surgical field. These LNs could then become sites where tumor antigens are presented and new or recalled T cell responses are generated. Because T cell state and function are strongly influenced by anatomical location and tissue context, the immune response to postoperative micrometastases may depend on where and when these metastatic-site immune niches develop [40]. Thus, adjuvant ICI can be viewed as a strategy to maintain or redirect antitumor immunity against evolving metastatic foci, rather than only as a drug given to eliminate residual cells immediately after surgery.
Clinical trial designs also reflect this uncertainty in timing, although they do not directly define the optimal biological duration of treatment. In resected melanoma, adjuvant pembrolizumab in KEYNOTE-054 was administered for a fixed period and improved recurrence-free survival [41]. In renal cell carcinoma, KEYNOTE-564 showed that adjuvant pembrolizumab after nephrectomy improved survival outcomes in high-risk patients [42]. Perioperative trials such as KEYNOTE-671 in NSCLC and KEYNOTE-689 in head and neck squamous cell carcinoma (HNSCC) also include postoperative ICI phases after surgery [7,43]. These regimens suggest that continued checkpoint blockade after surgery can be clinically beneficial, but the duration of treatment has largely been determined by trial design rather than by direct measurement of MRD activity, metastatic-site tdLN priming, or T cell reactivation.
Therefore, the rationale for adjuvant ICI should be framed more broadly than immediate killing of residual tumor cells. It may function as an immune surveillance strategy during a period when MRD, dormant disseminated cells, and early micrometastases are biologically unstable and not continuously visible to the immune system. The key unresolved question is when and where adjuvant ICI exerts its most important effect: on residual cells at the surgical site, on occult micrometastases, or within newly established draining LNs at distant anatomical sites. Defining biomarkers that capture MRD, tumor dormancy, antigen release, and metastatic-site immune priming will be necessary to determine which patients need prolonged adjuvant treatment and when treatment can be safely discontinued.
7. Cancer-Type-Specific Clinical Evidence
The clinical development of perioperative immune checkpoint inhibition has progressed across several solid tumors, but the biological meaning of this strategy is not identical among cancer types. Differences in tumor antigenicity, lymphatic anatomy, timing of surgery, use of chemotherapy, pattern of relapse, and dependence on regional LN management may influence how the tdLN–TPEX/TEX axis contributes to therapeutic efficacy. Representative clinical examples are summarized in Table 1.
In melanoma, neoadjuvant ICI provides one of the clearest examples of how pathological response can guide postoperative treatment. The NADINA trial evaluated neoadjuvant ipilimumab plus nivolumab followed by surgery and response-driven adjuvant therapy in resectable stage III melanoma [6]. This approach is particularly informative because melanoma is generally highly immunogenic and frequently contains pre-existing tumor-reactive T cells. In this setting, preserved tdLNs may support rapid immune activation before surgery, and pathological response can be used as an early indicator of whether systemic antitumor immunity has been effectively induced.
In NSCLC, perioperative chemoimmunotherapy has been evaluated in trials such as KEYNOTE-671 and CheckMate 77T [7,44]. These studies combine neoadjuvant chemotherapy with PD-1 blockade, followed by surgery and postoperative ICI. The immunological interpretation may differ from melanoma because chemotherapy can contribute to tumor cell death, antigen release, and remodeling of the inflammatory environment. In addition, mediastinal LNs may provide an anatomical site for antigen presentation and T cell priming before resection. Thus, perioperative ICI in NSCLC may act through both tumor debulking and systemic immune induction before surgery, followed by continued immune pressure against MRD after surgery.
In HNSCC, including oral squamous cell carcinoma (OSCC), the role of regional LNs is particularly important. KEYNOTE-689 showed the clinical benefit of adding neoadjuvant and adjuvant pembrolizumab to surgery and standard postoperative therapy in resectable locally advanced disease [43]. From an immunological viewpoint, cervical tdLNs may be especially relevant because they are directly connected to the primary tumor through lymphatic drainage and are frequently included in surgical management. These nodes may function not only as metastatic sites but also as immune reservoirs in which TPEX cells are maintained and mobilized by PD-1 blockade. Therefore, HNSCC provides a clinically important model for considering how neck dissection, nodal re
Triple-negative breast cancer (TNBC) represents another pattern of perioperative immunotherapy. In KEYNOTE-522, pembrolizumab was combined with neoadjuvant chemotherapy and continued after surgery [45]. In this setting, chemotherapy is not only cytotoxic but may also enhance antitumor immunity by increasing antigen release, inducing inflammatory signals, and altering the tumor microenvironment. PD-1 blockade may then amplify T cell responses generated during this period. The tdLN–TPEX/TEX axis in TNBC may therefore operate in close association with chemotherapy-induced immune remodeling rather than through checkpoint blockade alone.
In muscle-invasive bladder cancer, the NIAGARA trial evaluated durvalumab with neoadjuvant gemcitabine/cisplatin followed by radical cystectomy and adjuvant durvalumab [46]. This strategy illustrates how perioperative PD-L1 blockade can be combined with standard cisplatin-based chemotherapy. Before cystectomy, the primary tumor and pelvic lymphatic drainage remain present, allowing the possibility of treatment-induced antigen release and immune activation. After surgery, continued durvalumab may help maintain immune surveillance against residual disease. Thus, bladder cancer provides an example in which neoadjuvant chemoimmunotherapy and adjuvant immune maintenance are combined within one perioperative strategy.
Mismatch repair-deficient (dMMR) colon cancer provides a distinct model. In NICHE-2, short-course neoadjuvant nivolumab plus ipilimumab induced marked pathological responses in locally advanced dMMR colon cancer [47]. The strong response in this setting is likely related to high neoantigen burden and pre-existing immune recognition. Because dMMR tumors may already contain active tumor-reactive immune responses, a short period of checkpoint blockade before surgery may be sufficient to rapidly amplify antitumor immunity. This tumor type therefore highlights how intrinsic tumor antigenicity can strongly influence the magnitude and speed of response to neoadjuvant ICI.
Renal cell carcinoma differs from these examples. KEYNOTE-564 supports the clinical value of adjuvant pembrolizumab after nephrectomy in high-risk disease, whereas PROSPER RCC did not show a clear benefit for perioperative nivolumab [42,48]. These results suggest that the optimal timing of ICI may vary according to tumor biology and clinical context. In renal cell carcinoma, current evidence more strongly supports postoperative immune surveillance against MRD than neoadjuvant immune priming before nephrectomy. This contrast is important because it shows that the perioperative ICI paradigm should not be applied uniformly across all tumors.
Taken together, these clinical examples suggest that the tdLN–TPEX/TEX axis may operate differently depending on tumor type. In highly immunogenic tumors such as melanoma and dMMR colon cancer, neoadjuvant ICI may rapidly amplify pre-existing tumor-reactive immunity. In tumors treated with chemoimmunotherapy, such as NSCLC, TNBC, and bladder cancer, chemotherapy-induced antigen release and inflammatory remodeling may cooperate with checkpoint blockade. In HNSCC, the close relationship between the primary tumor and cervical tdLNs makes LN management a central issue. In renal cell carcinoma, the current evidence emphasizes postoperative MRD control rather than a clear neoadjuvant advantage. These differences support the need for cancer-type-specific perioperative strategies rather than a single model of ICI timing for all solid tumors.
8. Reconsidering LN Surgery in the ICI Era
LN surgery has traditionally had three major purposes in solid tumors: pathological staging, regional disease control, and removal of clinically or microscopically involved LNs. This principle remains essential in the era of immunotherapy. Clinically metastatic LNs should not be left untreated simply because tdLNs may have immunological functions. Rather, the emerging question is more specific: when ICIs are given before surgery, could the timing and extent of LN removal influence the antitumor immune response induced by treatment?
This question arises because tdLNs are not passive anatomical structures. Experimental studies have shown that tdLNs can be required for optimal responses to PD-1/PD-L1 blockade, and that checkpoint interactions within tdLNs can regulate tumor-specific T cell immunity [8,9]. Recent reviews have therefore emphasized the clinical dilemma that tdLNs can act both as metastatic sites and as sites where antitumor immunity is initiated or maintained [49,50]. From this viewpoint, removal or irradiation of uninvolved or minimally involved lymphatic basins before immune activation could theoretically reduce the lymphoid sites available for antigen presentation, T cell priming, and maintenance of less differentiated tumor-reactive T cells. However, this idea remains a biological hypothesis and must be balanced against established oncologic indications for nodal treatment.
Melanoma provides the clearest clinical example of how LN surgery has already been de-escalated in selected settings. In MSLT-II, immediate completion LN dissection after a positive sentinel LN improved regional control and provided additional staging information, but did not improve melanoma-specific survival compared with nodal observation [51]. This result supported a more selective approach to completion LN dissection in patients with limited sentinel node involvement. In the immunotherapy era, melanoma has moved further toward response-adapted surgery. In the PRADO study, pathological response after neoadjuvant ipilimumab plus nivolumab was used to guide subsequent surgery and adjuvant therapy, allowing treatment de-escalation in major responders and escalation in poor responders [52]. The phase 3 NADINA trial further supported neoadjuvant ipilimumab plus nivolumab followed by response-driven adjuvant therapy in resectable stage III melanoma [6]. These studies suggest that pathological response after neoadjuvant ICI can become a decision-making tool for tailoring the extent of subsequent treatment. However, this approach is best established in melanoma and should not be directly extrapolated to other tumor types without disease-specific evidence.
OSCC and other head and neck cancers require a more cautious interpretation. In early node-negative oral cancer, elective neck dissection has been shown to improve overall and disease-free survival compared with therapeutic neck dissection performed after nodal relapse [53]. This finding is important because it shows that occult nodal disease can be clinically meaningful and that elective nodal treatment should not be abandoned without strong evidence. At the same time, sentinel node biopsy has been evaluated as a less invasive staging strategy in selected patients with early oral cancer. The SENT trial showed that sentinel node biopsy can provide reliable staging of the clinically N0 neck in T1–T2 oral cancer, with high sentinel node identification and negative predictive value [54]. These data indicate that LN management in oral cancer can be individualized in some settings, but always within a framework that preserves oncologic safety.
The introduction of perioperative ICI in HNSCC adds another layer to this discussion. KEYNOTE-689 showed that neoadjuvant and adjuvant pembrolizumab added to surgery and standard postoperative therapy improved event-free survival in resectable locally advanced disease [43]. This trial does not by itself define how neck dissection should be modified after neoadjuvant ICI. However, it creates a new clinical context in which cervical LNs are exposed to checkpoint blockade before surgical removal. In this setting, the neck specimen may provide information not only about residual metastatic disease, but also about immune activation, pathological regression, T cell states, DC function, and the presence or loss of tumor-reactive T cell reservoirs. Thus, neck dissection after neoadjuvant ICI may become both a therapeutic procedure and a translational window for understanding nodal immune responses.
A practical framework is therefore needed. First, clinically involved nodes should remain targets for adequate surgical or radiation-based control. Second, elective treatment of clinically uninvolved nodal basins should be guided by tumor type, stage, anatomical risk, and available evidence, rather than relying solely on immunological speculation. Third, when neoadjuvant ICI is used, pathological response in both the primary tumor and LNs should be evaluated systematically. Fourth, immune monitoring of resected LNs may help distinguish nodes that were mainly metastatic deposits from nodes that retained active immune function. Such monitoring could include spatial analysis of DCs, TPEX-like CD8⁺ T cells, TCR clonality, TLS-like structure, and checkpoint ligand expression.
In summary, the ICI era does not eliminate the need for LN surgery. Instead, it changes the questions that LN surgery must answer. The goal is no longer only to determine whether nodal metastasis is present or absent, but also to understand how nodal disease, nodal immune activity, and treatment-induced pathological response should guide subsequent therapy. Future strategies may integrate sentinel node approaches, response-adapted surgery, pathological regression scoring, and immune profiling. The central challenge is to preserve oncologic safety while avoiding unnecessary disruption of lymphoid structures that may contribute to systemic antitumor immunity.
9. Postoperative Lymphatic Dysfunction, Lymphedema, and Immune Surveillance
Postoperative lymphatic dysfunction is usually discussed as a complication that affects quality of life, physical function, and long-term survivorship. In head and neck cancer, lymphedema can involve both external soft tissues and internal structures, and may contribute to discomfort, dysphagia, speech problems, cosmetic changes, and functional impairment [55,56]. LN dissection and radiotherapy are major contributors to this condition. Radiotherapy can damage lymphatic vessels, promote tissue fibrosis, and impair lymphatic drainage, whereas surgery can physically interrupt lymphatic channels and remove regional LNs [57]. These clinical consequences are well recognized, but their possible immunological implications in the era of immune checkpoint inhibition have been less extensively discussed.
The lymphatic system is not only a drainage pathway for interstitial fluid. It also provides an anatomical route for antigen transport, DC migration, and immune cell trafficking. Afferent lymphatic vessels carry tissue-derived antigens and antigen-bearing DCs to draining LNs, where adaptive immune responses can be initiated or regulated [58]. In cancer, lymphatic vessels can therefore influence both metastasis and antitumor immunity. Recent reviews have emphasized that lymphatic transport shapes the delivery of tumor antigens, the movement of immune cells, and the local conditions under which immunotherapy acts [59,60]. From this perspective, postoperative disruption of lymphatic flow may alter not only fluid balance, but also the immune communication between peripheral tissues, residual tumor sites, and regional lymphoid structures. This proposed cascade, linking local cancer therapy to lymphatic disruption, tissue remodeling, and altered immune surveillance, is summarized in Figure 4.
Lymphedema is also associated with chronic inflammatory and immune alterations. Lymphatic dysfunction can change the local immune environment by affecting leukocyte trafficking, antigen clearance, inflammatory cell accumulation, and immune regulation [61,62]. Experimental models have shown that CD4+ T cells are activated after lymphatic injury and can contribute to the initiation of lymphedema [63]. Regulatory T cells (Tregs) may also accumulate in lymphedematous tissues and participate in local immune suppression [64]. These observations suggest that lymphatic failure is not a purely mechanical disorder, but a condition in which persistent immune activation and local immune regulation coexist.
Fibrosis is another important component of postoperative lymphatic dysfunction. In lymphedema, chronic tissue swelling is frequently accompanied by extracellular matrix remodeling, soft tissue thickening, and fibrotic change. Th2-skewed immune responses have been implicated in the development of tissue fibrosis and impaired lymphatic function after lymphatic injury [65]. In addition, lymph leakage or impaired lymphatic integrity may influence macrophage differentiation and favor anti-inflammatory or tissue-remodeling phenotypes [66]. These changes could theoretically affect local immune surveillance by modifying antigen availability, stromal architecture, immune cell access, and the balance between inflammatory and suppressive immune pathways.
The relevance of these processes to cancer recurrence remains uncertain. At present, there is limited direct clinical evidence showing that postoperative lymphedema itself increases the risk of local recurrence or distant metastasis. Therefore, postoperative lymphatic dysfunction should not be presented as an established cause of cancer relapse. A more appropriate interpretation is that lymphatic injury, chronic inflammation, fibrosis, altered macrophage polarization, and impaired immune cell trafficking may create a postoperative tissue environment that could influence immune surveillance. This possibility is particularly relevant in patients treated with perioperative ICI, because the efficacy of immunotherapy depends on antigen transport, DC function, T cell priming, and T cell access to sites of residual disease.
Thus, postoperative lymphatic dysfunction should be considered as a component of postoperative immune ecology. After surgery and radiotherapy, the local tissue environment is no longer the same as the preoperative tumor-draining system. Lymphatic drainage routes may be interrupted, LNs may be removed, fibrotic barriers may develop, and immune cell migration may be altered. These changes could affect how residual tumor antigens are transported, how DCs reach lymphoid tissues, and how tumor-reactive T cells enter or survey postoperative tissues. In the ICI era, lymphedema and fibrosis should therefore be viewed not only as survivorship problems, but also as potential modifiers of local and regional immune surveillance.
Future studies should examine postoperative lymphatic dysfunction using immunological endpoints. These may include spatial analysis of lymphatic vessels, DCs, macrophage subsets, T cell infiltration, Treg accumulation, fibrosis-associated stromal changes, and checkpoint ligand expression in postoperative tissues. Clinical studies should also evaluate whether the severity of lymphedema or fibrosis correlates with immune-related biomarkers, response to adjuvant ICI, or patterns of recurrence. Such studies would help determine whether postoperative lymphatic dysfunction is only a consequence of local therapy or whether it also modifies the immune environment in which perioperative immunotherapy must operate.
10. Future Directions: Immune-Guided Surgery and Response-Adapted Perioperative Therapy
The next step in perioperative immunotherapy is to move from uniform treatment schedules toward strategies guided by immune status, pathological response, and residual disease risk. Neoadjuvant ICI provides a unique opportunity because tumor tissues, draining LNs, and blood samples can be analyzed before treatment, during treatment, and at surgery. These paired samples may help identify where immune activation occurs, which T cell populations expand, and whether the response is maintained after tumor removal [3].
First, immune monitoring of tdLNs should be developed as a routine research component of neoadjuvant ICI trials. Current assessment of LNs is mainly focused on the presence or absence of metastatic tumor cells. In the ICI era, this may be insufficient. tdLNs should also be evaluated for immune activity, including TCF1+ or TPEX-like CD8⁺ T cells, DC subsets, TCR clonality, B cell-rich lymphoid aggregates, and spatial organization of immune cells. Studies showing stem-like CD8⁺ T cell populations and cDC1-dependent maintenance of tumor-reactive T cells in tdLNs provide a rationale for this approach [17,19]. TCR sequencing may also help determine whether tumor-reactive clones are expanded locally, recruited from outside the tumor, or replaced after PD-1 blockade [67]. In addition, TLS and B cell-rich immune niches should be incorporated into spatial immune profiling because they have been associated with immunotherapy response in several cancer settings [68,69].
Second, pathological response should be integrated with immune biomarkers to guide postoperative treatment. In melanoma, pathological response after neoadjuvant therapy is strongly associated with clinical outcome and has already been used to adapt subsequent surgery and adjuvant therapy [52,70]. The NADINA trial further supports the clinical relevance of a neoadjuvant approach followed by response-driven postoperative management [6]. These studies suggest that major responders may not require the same intensity or duration of adjuvant therapy as poor responders. However, pathological regression alone may not fully reflect the quality of systemic immunity. Future response-adapted strategies should therefore combine pathological response with immune features such as tdLN T cell states, TCR clonal expansion, DC activation, circulating immune markers, and MRD status.
Third, LN surgery should be reconsidered within an immune-guided framework. This does not mean that metastatic LNs should be preserved when oncologic control requires their removal. Rather, it means that the timing, extent, and purpose of LN surgery should be discussed together with the immunological role of tdLNs. Recent discussions have emphasized that tdLNs can be both sites of tumor spread and sites of antitumor immune activation [49,50]. Therefore, future trials should ask whether LN dissection before, during, or after ICI affects immune priming, T cell expansion, pathological response, and recurrence patterns. In selected settings, sentinel node strategies, response-based nodal surgery, or limited nodal dissection may be evaluated, but only when oncologic safety is maintained.
Fourth, the duration of adjuvant ICI should be refined using biomarkers rather than fixed schedules alone. At present, many adjuvant or perioperative ICI regimens use predefined treatment periods, often based on clinical trial design. However, the biological window during which residual tumor cells become immunologically visible may differ among patients. MRD detected by circulating tumor DNA may provide one approach to identifying patients with persistent microscopic disease or early molecular relapse [71]. In the future, MRD-guided strategies could help determine whether adjuvant ICI should be continued, stopped, intensified, or combined with other therapies. Such decisions may be particularly important for patients with dormant micrometastases or evolving metastatic-site draining LNs, in whom immune activation may occur only after a variable period of tumor growth or antigen release.
Finally, perioperative immunotherapy should be studied as an integrated anatomical and immunological process. The primary tumor, tdLNs, metastatic sites, blood, bone marrow, and postoperative lymphatic environment should not be analyzed separately. Instead, they should be considered as connected compartments that shape antigen flow, DC migration, T cell priming, T cell exhaustion, and immune surveillance. Future trials should therefore include coordinated sampling of tumors, LNs, blood, MRD markers, and postoperative tissues. Key immune monitoring targets for each anatomical compartment are summarized in Table 2. Such studies may allow perioperative ICI to evolve from a fixed drug schedule into a personalized strategy that combines immune monitoring, response-adapted therapy, MRD-guided treatment duration, and immune-guided surgery.
11. Conclusions
Perioperative immunotherapy is changing the way solid tumors are treated, but its full significance cannot be understood only by focusing on tumor shrinkage or postoperative eradication of residual disease. The timing of immune checkpoint inhibition relative to surgery is immunologically important because the primary tumor, lymphatic drainage, DC migration, and tdLNs remain connected before resection. In this setting, tdLNs are not only anatomical sites of metastatic spread, but also immune organs that support antigen presentation, T cell priming, and the maintenance of less differentiated tumor-reactive CD8⁺ T cells.
This dual role creates both an opportunity and a dilemma. On one hand, tdLNs may act as metastatic gateways and must be managed according to established oncologic principles. On the other hand, they may serve as immune reservoirs that contribute to the response to PD-1/PD-L1 blockade. Therefore, perioperative immunotherapy should be considered as a strategy that acts through the entire tumor–lymphatic–tdLN axis, rather than only within the tumor microenvironment.
Future treatment strategies should integrate pathological response, immune monitoring of tdLNs, TCR clonality, DC function, MRD assessment, and postoperative lymphatic status. Such an approach may allow clinicians to select patients who require intensified adjuvant therapy, identify those in whom treatment can be safely reduced, and design LN surgery that preserves oncologic safety while taking immune function into account. Reframing tdLNs as both metastatic gateways and immune reservoirs may therefore provide a useful framework for optimizing perioperative immunotherapy across solid tumors.
Author Contributions
Conceptualization, K.K.; literature review, K.K., Y.K. and K.N.; writing—original draft preparation, K.K.; writing—review and editing, K.K., Y.K. and K.N.; visualization, K.K.; supervision, K.K.; project administration, K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors thank colleagues and collaborators for helpful discussions on tumor immunology and perioperative cancer immunotherapy. During the preparation of this manuscript, the authors used ChatGPT (OpenAI, GPT-5.5) for assistance with language editing, manuscript structuring, and preparation of draft conceptual figure illustrations. The authors reviewed, edited, and approved all generated content and take full responsibility for the final content of the manuscript. The authors acknowledge the Immuno Editorial Office for considering an article processing charge waiver for this invited review.
Conflicts of Interest
The authors declares no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ICI | immune checkpoint inhibitor |
| MRD | minimal residual disease |
| LN | lymph node |
| tdLN | tumor-draining lymph node |
| CD8 | cluster of differentiation 8 |
| PD-1 | programmed cell death protein 1 |
| TPEX | progenitor-exhausted T cell |
| TEX | exhausted T cell |
| NSCLC | non-small cell lung cancer |
| PD-L1 | programmed death-ligand 1 |
| HEV | high endothelial venule |
| DC | dendritic cell |
| cDC1 | conventional type 1 dendritic cell |
| DTC | disseminated tumor cell |
| MHC | major histocompatibility complex |
| TCF1 | T cell factor 1 |
| TLS | tertiary lymphoid structure |
| TCR | T cell receptor |
| CTC | circulating tumor cell |
| HNSCC | head and neck squamous cell carcinoma |
| OSCC | oral squamous cell carcinoma |
| TNBC | triple-negative breast cancer |
| dMMR | mismatch repair-deficient |
| RCC | renal cell carcinoma |
| Treg | regulatory T cell |
| CD4 | cluster of differentiation 4 |
| Th2 | T helper 2 |
References
- Topalian, S.L.; Taube, J.M.; Pardoll, D.M. Neoadjuvant checkpoint blockade for cancer immunotherapy. Science 2020, 367, eaax0182. [Google Scholar] [CrossRef]
- Versluis, J.M.; Long, G.V.; Blank, C.U. Learning from clinical trials of neoadjuvant checkpoint blockade. Nat. Med. 2020, 26, 475–484. [Google Scholar] [CrossRef]
- Topalian, S.L.; Forde, P.M.; Emens, L.A.; Yarchoan, M.; Smith, K.N.; Pardoll, D.M. Neoadjuvant immune checkpoint blockade: A window of opportunity to advance cancer immunotherapy. Cancer Cell 2023, 41, 1551–1566. [Google Scholar] [CrossRef]
- Awada, G.; Cascone, T.; van der Heijden, M.S.; Blank, C.U.; Kok, M.; Chalabi, M. The rapidly evolving paradigm of neoadjuvant immunotherapy across cancer types. Nat. Cancer 2025, 6, 967–987. [Google Scholar] [CrossRef]
- Patel, S.P.; Othus, M.; Chen, Y.; Wright, G.P., Jr.; Yost, K.J.; Hyngstrom, J.R.; Hu-Lieskovan, S.; Lao, C.D.; Fecher, L.A.; Truong, T.G.; et al. Neoadjuvant-Adjuvant or Adjuvant-Only Pembrolizumab in Advanced Melanoma. N Engl. J. Med. 2023, 388, 813–823. [Google Scholar] [CrossRef]
- Blank, C.U.; Lucas, M.W.; Scolyer, R.A.; van de Wiel, B.A.; Menzies, A.M.; Lopez-Yurda, M.; Hoeijmakers, L.L.; Saw, R.P.M.; Lijnsvelt, J.M.; Maher, N.G.; et al. Neoadjuvant Nivolumab and Ipilimumab in Resectable Stage III Melanoma. N Engl. J. Med. 2024, 391, 1696–1708. [Google Scholar] [CrossRef]
- Wakelee, H.; Liberman, M.; Kato, T.; Tsuboi, M.; Lee, S.H.; Gao, S.; Chen, K.N.; Dooms, C.; Majem, M.; Eigendorff, E.; et al. Perioperative Pembrolizumab for Early-Stage Non-Small-Cell Lung Cancer. N Engl. J. Med. 2023, 389, 491–503. [Google Scholar] [CrossRef]
- Fransen, M.F.; Schoonderwoerd, M.; Knopf, P.; Camps, M.G.; Hawinkels, L.J.; Kneilling, M.; van Hall, T.; Ossendorp, F. Tumor-draining lymph nodes are pivotal in PD-1/PD-L1 checkpoint therapy. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Dammeijer, F.; van Gulijk, M.; Mulder, E.E.; Lukkes, M.; Klaase, L.; van den Bosch, T.; van Nimwegen, M.; Lau, S.P.; Latupeirissa, K.; Schetters, S.; et al. The PD-1/PD-L1-Checkpoint Restrains T cell Immunity in Tumor-Draining Lymph Nodes. Cancer Cell 2020, 38, 685–700.e688. [Google Scholar] [CrossRef]
- Delclaux, I.; Ventre, K.S.; Jones, D.; Lund, A.W. The tumor-draining lymph node as a reservoir for systemic immune surveillance. Trends Cancer 2024, 10, 28–37. [Google Scholar] [CrossRef]
- Pereira, E.R.; Kedrin, D.; Seano, G.; Gautier, O.; Meijer, E.F.J.; Jones, D.; Chin, S.M.; Kitahara, S.; Bouta, E.M.; Chang, J.; et al. Lymph node metastases can invade local blood vessels, exit the node, and colonize distant organs in mice. Science 2018, 359, 1403–1407. [Google Scholar] [CrossRef]
- Brown, M.; Assen, F.P.; Leithner, A.; Abe, J.; Schachner, H.; Asfour, G.; Bago-Horvath, Z.; Stein, J.V.; Uhrin, P.; Sixt, M.; et al. Lymph node blood vessels provide exit routes for metastatic tumor cell dissemination in mice. Science 2018, 359, 1408–1411. [Google Scholar] [CrossRef]
- Karaman, S.; Detmar, M. Mechanisms of lymphatic metastasis. J. Clin. Invest 2014, 124, 922–928. [Google Scholar] [CrossRef]
- Ji, H.; Hu, C.; Yang, X.; Liu, Y.; Ji, G.; Ge, S.; Wang, X.; Wang, M. Lymph node metastasis in cancer progression: molecular mechanisms, clinical significance and therapeutic interventions. Signal Transduct. Target Ther. 2023, 8, 367. [Google Scholar] [CrossRef]
- Achen, M.G.; Stacker, S.A. Exit Stage Left: A Tumor Cell’s Journey from Lymph Node to Beyond. Trends Cancer 2018, 4, 519–522. [Google Scholar] [CrossRef]
- du Bois, H.; Heim, T.A.; Lund, A.W. Tumor-draining lymph nodes: At the crossroads of metastasis and immunity. Sci. Immunol. 2021, 6, eabg3551. [Google Scholar] [CrossRef]
- Schenkel, J.M.; Herbst, R.H.; Canner, D.; Li, A.; Hillman, M.; Shanahan, S.L.; Gibbons, G.; Smith, O.C.; Kim, J.Y.; Westcott, P.; et al. Conventional type I dendritic cells maintain a reservoir of proliferative tumor-antigen specific TCF-1(+) CD8(+) T cells in tumor-draining lymph nodes. Immunity 2021, 54, 2338–2353.e2336. [Google Scholar] [CrossRef]
- Pittet, M.J.; Di Pilato, M.; Garris, C.; Mempel, T.R. Dendritic cells as shepherds of T cell immunity in cancer. Immunity 2023, 56, 2218–2230. [Google Scholar] [CrossRef]
- Connolly, K.A.; Kuchroo, M.; Venkat, A.; Khatun, A.; Wang, J.; William, I.; Hornick, N.I.; Fitzgerald, B.L.; Damo, M.; Kasmani, M.Y.; et al. A reservoir of stem-like CD8(+) T cells in the tumor-draining lymph node preserves the ongoing antitumor immune response. Sci. Immunol. 2021, 6, eabg7836. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, X.; Wang, Z.; Chen, X.; Wang, L.; Lu, Y.; Xiong, D.; Liu, Q.; Tian, Y.; Lin, H.; et al. The primordial differentiation of tumor-specific memory CD8(+) T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell 2022, 185, 4049–4066.e4025. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- van der Leun, A.M.; Thommen, D.S.; Schumacher, T.N. CD8(+) T cell states in human cancer: insights from single-cell analysis. Nat. Rev. Cancer 2020, 20, 218–232. [Google Scholar] [CrossRef]
- Chow, A.; Perica, K.; Klebanoff, C.A.; Wolchok, J.D. Clinical implications of T cell exhaustion for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2022, 19, 775–790. [Google Scholar] [CrossRef]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; et al. Subsets of exhausted CD8(+) T cells differentially mediate tumor control and respond to checkpoint blockade. Nat. Immunol. 2019, 20, 326–336. [Google Scholar] [CrossRef]
- Siddiqui, I.; Schaeuble, K.; Chennupati, V.; Fuertes Marraco, S.A.; Calderon-Copete, S.; Pais Ferreira, D.; Carmona, S.J.; Scarpellino, L.; Gfeller, D.; Pradervand, S.; et al. Intratumoral Tcf1(+)PD-1(+)CD8(+) T Cells with Stem-like Properties Promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity 2019, 50, 195–211.e110. [Google Scholar] [CrossRef]
- Beltra, J.C.; Manne, S.; Abdel-Hakeem, M.S.; Kurachi, M.; Giles, J.R.; Chen, Z.; Casella, V.; Ngiow, S.F.; Khan, O.; Huang, Y.J.; et al. Developmental Relationships of Four Exhausted CD8(+) T Cell Subsets Reveals Underlying Transcriptional and Epigenetic Landscape Control Mechanisms. Immunity 2020, 52, 825–841.e828. [Google Scholar] [CrossRef]
- Im, S.J.; Hashimoto, M.; Gerner, M.Y.; Lee, J.; Kissick, H.T.; Burger, M.C.; Shan, Q.; Hale, J.S.; Lee, J.; Nasti, T.H.; et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature 2016, 537, 417–421. [Google Scholar] [CrossRef]
- Pauken, K.E.; Sammons, M.A.; Odorizzi, P.M.; Manne, S.; Godec, J.; Khan, O.; Drake, A.M.; Chen, Z.; Sen, D.R.; Kurachi, M.; et al. Epigenetic stability of exhausted T cells limits durability of reinvigoration by PD-1 blockade. Science 2016, 354, 1160–1165. [Google Scholar] [CrossRef]
- Liu, J.; Blake, S.J.; Yong, M.C.; Harjunpää, H.; Ngiow, S.F.; Takeda, K.; Young, A.; O’Donnell, J.S.; Allen, S.; Smyth, M.J.; et al. Improved Efficacy of Neoadjuvant Compared to Adjuvant Immunotherapy to Eradicate Metastatic Disease. Cancer Discov. 2016, 6, 1382–1399. [Google Scholar] [CrossRef]
- Blank, C.U.; Rozeman, E.A.; Fanchi, L.F.; Sikorska, K.; van de Wiel, B.; Kvistborg, P.; Krijgsman, O.; van den Braber, M.; Philips, D.; Broeks, A.; et al. Neoadjuvant versus adjuvant ipilimumab plus nivolumab in macroscopic stage III melanoma. Nat. Med. 2018, 24, 1655–1661. [Google Scholar] [CrossRef]
- Forde, P.M.; Chaft, J.E.; Smith, K.N.; Anagnostou, V.; Cottrell, T.R.; Hellmann, M.D.; Zahurak, M.; Yang, S.C.; Jones, D.R.; Broderick, S.; et al. Neoadjuvant PD-1 Blockade in Resectable Lung Cancer. N Engl. J. Med. 2018, 378, 1976–1986. [Google Scholar] [CrossRef]
- Badia-Ramentol, J.; Linares, J.; Gómez-Llonin, A.; Calon, A. Minimal Residual Disease, Metastasis and Immunity. Biomolecules 2021, 11, 130. [Google Scholar] [CrossRef]
- Spiotto, M.T.; Yu, P.; Rowley, D.A.; Nishimura, M.I.; Meredith, S.C.; Gajewski, T.F.; Fu, Y.-X.; Schreiber, H. Increasing Tumor Antigen Expression Overcomes “Ignorance” to Solid Tumors via Crosspresentation by Bone Marrow-Derived Stromal Cells. Immunity 2002, 17, 737–747. [Google Scholar] [CrossRef]
- Willimsky, G.; Blankenstein, T. Sporadic immunogenic tumours avoid destruction by inducing T-cell tolerance. Nature 2005, 437, 141–146. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology meets immunology: the cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef]
- Aguirre-Ghiso, J.A. Models, mechanisms and clinical evidence for cancer dormancy. Nat. Rev. Cancer 2007, 7, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.S.; Bragado, P.; Aguirre-Ghiso, J.A. Mechanisms of disseminated cancer cell dormancy: an awakening field. Nat. Rev. Cancer 2014, 14, 611–622. [Google Scholar] [CrossRef]
- Goddard, E.T.; Linde, M.H.; Srivastava, S.; Klug, G.; Shabaneh, T.B.; Iannone, S.; Grzelak, C.A.; Marsh, S.; Riggio, A.I.; Shor, R.E.; et al. Immune evasion of dormant disseminated tumor cells is due to their scarcity and can be overcome by T cell immunotherapies. Cancer Cell 2024, 42, 119–134.e112. [Google Scholar] [CrossRef]
- Schenkel, J.M.; Pauken, K.E. Localization, tissue biology and T cell state - implications for cancer immunotherapy. Nat. Rev. Immunol. 2023, 23, 807–823. [Google Scholar] [CrossRef]
- Eggermont, A.M.M.; Blank, C.U.; Mandala, M.; Long, G.V.; Atkinson, V.; Dalle, S.; Haydon, A.; Lichinitser, M.; Khattak, A.; Carlino, M.S.; et al. Adjuvant Pembrolizumab versus Placebo in Resected Stage III Melanoma. N Engl. J. Med. 2018, 378, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Tomczak, P.; Park, S.H.; Venugopal, B.; Ferguson, T.; Symeonides, S.N.; Hajek, J.; Chang, Y.H.; Lee, J.L.; Sarwar, N.; et al. Overall Survival with Adjuvant Pembrolizumab in Renal-Cell Carcinoma. N Engl. J. Med. 2024, 390, 1359–1371. [Google Scholar] [CrossRef]
- Uppaluri, R.; Haddad, R.I.; Tao, Y.; Le Tourneau, C.; Lee, N.Y.; Westra, W.; Chernock, R.; Tahara, M.; Harrington, K.J.; Klochikhin, A.L.; et al. Neoadjuvant and Adjuvant Pembrolizumab in Locally Advanced Head and Neck Cancer. N Engl. J. Med. 2025, 393, 37–50. [Google Scholar] [CrossRef]
- Cascone, T.; Awad, M.M.; Spicer, J.D.; He, J.; Lu, S.; Sepesi, B.; Tanaka, F.; Taube, J.M.; Cornelissen, R.; Havel, L.; et al. Perioperative Nivolumab in Resectable Lung Cancer. N Engl. J. Med. 2024, 390, 1756–1769. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Cortes, J.; Dent, R.; McArthur, H.; Pusztai, L.; Kummel, S.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N.; et al. Overall Survival with Pembrolizumab in Early-Stage Triple-Negative Breast Cancer. N Engl. J. Med. 2024, 391, 1981–1991. [Google Scholar] [CrossRef]
- Powles, T.; Catto, J.W.F.; Galsky, M.D.; Al-Ahmadie, H.; Meeks, J.J.; Nishiyama, H.; Vu, T.Q.; Antonuzzo, L.; Wiechno, P.; Atduev, V.; et al. Perioperative Durvalumab with Neoadjuvant Chemotherapy in Operable Bladder Cancer. N Engl. J. Med. 2024, 391, 1773–1786. [Google Scholar] [CrossRef]
- Chalabi, M.; Verschoor, Y.L.; Tan, P.B.; Balduzzi, S.; Van Lent, A.U.; Grootscholten, C.; Dokter, S.; Buller, N.V.; Grotenhuis, B.A.; Kuhlmann, K.; et al. Neoadjuvant Immunotherapy in Locally Advanced Mismatch Repair-Deficient Colon Cancer. N Engl. J. Med. 2024, 390, 1949–1958. [Google Scholar] [CrossRef]
- Allaf, M.E.; Kim, S.E.; Master, V.; McDermott, D.F.; Harshman, L.C.; Cole, S.M.; Drake, C.G.; Signoretti, S.; Akgul, M.; Baniak, N.; et al. Perioperative nivolumab versus observation in patients with renal cell carcinoma undergoing nephrectomy (PROSPER ECOG-ACRIN EA8143): an open-label, randomised, phase 3 study. Lancet Oncol. 2024, 25, 1038–1052. [Google Scholar] [CrossRef] [PubMed]
- Saddawi-Konefka, R.; Schokrpur, S.; Gutkind, J.S. Let it be: Preserving tumor-draining lymph nodes in the era of immuno-oncology. Cancer Cell 2024, 42, 930–933. [Google Scholar] [CrossRef]
- von Renesse, J.; Lin, M.-C.; Ho, P.-C. Tumor-draining lymph nodes – friend or foe during immune checkpoint therapy? Trends Cancer 2025, 11, 676–690. [Google Scholar] [CrossRef]
- Faries, M.B.; Thompson, J.F.; Cochran, A.J.; Andtbacka, R.H.; Mozzillo, N.; Zager, J.S.; Jahkola, T.; Bowles, T.L.; Testori, A.; Beitsch, P.D.; et al. Completion Dissection or Observation for Sentinel-Node Metastasis in Melanoma. N Engl. J. Med. 2017, 376, 2211–2222. [Google Scholar] [CrossRef]
- Reijers, I.L.M.; Menzies, A.M.; van Akkooi, A.C.J.; Versluis, J.M.; van den Heuvel, N.M.J.; Saw, R.P.M.; Pennington, T.E.; Kapiteijn, E.; van der Veldt, A.A.M.; Suijkerbuijk, K.P.M.; et al. Personalized response-directed surgery and adjuvant therapy after neoadjuvant ipilimumab and nivolumab in high-risk stage III melanoma: the PRADO trial. Nat. Med. 2022, 28, 1178–1188. [Google Scholar] [CrossRef]
- D’Cruz, A.K.; Vaish, R.; Kapre, N.; Dandekar, M.; Gupta, S.; Hawaldar, R.; Agarwal, J.P.; Pantvaidya, G.; Chaukar, D.; Deshmukh, A.; et al. Elective versus Therapeutic Neck Dissection in Node-Negative Oral Cancer. N Engl. J. Med. 2015, 373, 521–529. [Google Scholar] [CrossRef]
- Schilling, C.; Stoeckli, S.J.; Haerle, S.K.; Broglie, M.A.; Huber, G.F.; Sorensen, J.A.; Bakholdt, V.; Krogdahl, A.; von Buchwald, C.; Bilde, A.; et al. Sentinel European Node Trial (SENT): 3-year results of sentinel node biopsy in oral cancer. Eur. J. Cancer 2015, 51, 2777–2784. [Google Scholar] [CrossRef]
- Deng, J.; Ridner, S.H.; Dietrich, M.S.; Wells, N.; Wallston, K.A.; Sinard, R.J.; Cmelak, A.J.; Murphy, B.A. Prevalence of secondary lymphedema in patients with head and neck cancer. J. Pain Symptom Manag. 2012, 43, 244–252. [Google Scholar] [CrossRef]
- Deng, J.; Wulff-Burchfield, E.M.; Murphy, B.A. Late Soft Tissue Complications of Head and Neck Cancer Therapy: Lymphedema and Fibrosis. J. Natl. Cancer Inst. Monogr. 2019. [Google Scholar] [CrossRef]
- Allam, O.; Park, K.E.; Chandler, L.; Mozaffari, M.A.; Ahmad, M.; Lu, X.; Alperovich, M. The impact of radiation on lymphedema: a review of the literature. Gland Surg. 2020, 9, 596–602. [Google Scholar] [CrossRef]
- Arasa, J.; Collado-Diaz, V.; Halin, C. Structure and Immune Function of Afferent Lymphatics and Their Mechanistic Contribution to Dendritic Cell and T Cell Trafficking. Cells 2021, 10, 1269. [Google Scholar] [CrossRef]
- Karakousi, T.; Mudianto, T.; Lund, A.W. Lymphatic vessels in the age of cancer immunotherapy. Nat. Rev. Cancer 2024, 24, 363–381. [Google Scholar] [CrossRef]
- Sun, M.; Angelillo, J.; Hugues, S. Lymphatic transport in anti-tumor immunity and metastasis. J. Exp. Med. 2025, 222. [Google Scholar] [CrossRef]
- Yuan, Y.; Arcucci, V.; Levy, S.M.; Achen, M.G. Modulation of Immunity by Lymphatic Dysfunction in Lymphedema. Front. Immunol. 2019, 10–2019. [Google Scholar] [CrossRef]
- Kataru, R.P.; Baik, J.E.; Park, H.J.; Wiser, I.; Rehal, S.; Shin, J.Y.; Mehrara, B.J. Regulation of Immune Function by the Lymphatic System in Lymphedema. Front Immunol. 2019, 10, 470. [Google Scholar] [CrossRef]
- García Nores, G.D.; Ly, C.L.; Cuzzone, D.A.; Kataru, R.P.; Hespe, G.E.; Torrisi, J.S.; Huang, J.J.; Gardenier, J.C.; Savetsky, I.L.; Nitti, M.D.; et al. CD4+ T cells are activated in regional lymph nodes and migrate to skin to initiate lymphedema. Nat. Commun. 2018, 9, 1970. [Google Scholar] [CrossRef]
- García Nores, G.D.; Ly, C.L.; Savetsky, I.L.; Kataru, R.P.; Ghanta, S.; Hespe, G.E.; Rockson, S.G.; Mehrara, B.J. Regulatory T Cells Mediate Local Immunosuppression in Lymphedema. J. Invest Dermatol. 2018, 138, 325–335. [Google Scholar] [CrossRef]
- Avraham, T.; Zampell, J.C.; Yan, A.; Elhadad, S.; Weitman, E.S.; Rockson, S.G.; Bromberg, J.; Mehrara, B.J. Th2 differentiation is necessary for soft tissue fibrosis and lymphatic dysfunction resulting from lymphedema. Faseb J. 2013, 27, 1114–1126. [Google Scholar] [CrossRef]
- Herrada, A.A.; Olate-Briones, A.; Lazo-Amador, R.; Liu, C.; Hernández-Rojas, B.; Riadi, G.; Escobedo, N. Lymph Leakage Promotes Immunosuppression by Enhancing Anti-Inflammatory Macrophage Polarization. Front. Immunol. 2022, 13–2022. [Google Scholar] [CrossRef]
- Yost, K.E.; Satpathy, A.T.; Wells, D.K.; Qi, Y.; Wang, C.; Kageyama, R.; McNamara, K.L.; Granja, J.M.; Sarin, K.Y.; Brown, R.A.; et al. Clonal replacement of tumor-specific T cells following PD-1 blockade. Nat. Med. 2019, 25, 1251–1259. [Google Scholar] [CrossRef]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Skaarup Larsen, M.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J.; et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef]
- Menzies, A.M.; Amaria, R.N.; Rozeman, E.A.; Huang, A.C.; Tetzlaff, M.T.; van de Wiel, B.A.; Lo, S.; Tarhini, A.A.; Burton, E.M.; Pennington, T.E.; et al. Pathological response and survival with neoadjuvant therapy in melanoma: a pooled analysis from the International Neoadjuvant Melanoma Consortium (INMC). Nat. Med. 2021, 27, 301–309. [Google Scholar] [CrossRef]
- Zheng, J.; Qin, C.; Wang, Q.; Tian, D.; Chen, Z. Circulating tumour DNA-Based molecular residual disease detection in resectable cancers: a systematic review and meta-analysis. EBioMedicine 2024, 103, 105109. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Dual roles of tumor-draining lymph nodes as metastatic gateways and immune reservoirs. Tumor-draining lymph nodes (tdLNs) have two opposing but closely connected biological roles in cancer. (A) As metastatic gateways, tumor cells from the primary lesion can enter afferent lymphatic vessels and migrate to regional tdLNs, where they may survive and form nodal metastases. Tumor cells within metastatic LNs can further gain access to nodal blood vessels, including high endothelial venules (HEVs), and disseminate to distant organs. Thus, tdLNs may serve not only as regional metastatic sites but also as anatomical intermediates linking lymphatic spread to systemic dissemination. (B) As immune reservoirs, the same lymphatic route transports tumor antigens and antigen-bearing dendritic cells (DCs) from the tumor bed to tdLNs. Within tdLNs, conventional type 1 DC (cDC1)-mediated antigen presentation promotes tumor-specific T cell priming and supports the maintenance of progenitor-exhausted T cells (TPEX). These cells can exit tdLNs and give rise to more differentiated exhausted T cells (TEX) that traffic to primary tumors and micrometastatic lesions. This dual role provides a conceptual basis for understanding why the timing of immune checkpoint inhibition relative to tumor and LN removal may influence antitumor immunity.
Figure 1.
Dual roles of tumor-draining lymph nodes as metastatic gateways and immune reservoirs. Tumor-draining lymph nodes (tdLNs) have two opposing but closely connected biological roles in cancer. (A) As metastatic gateways, tumor cells from the primary lesion can enter afferent lymphatic vessels and migrate to regional tdLNs, where they may survive and form nodal metastases. Tumor cells within metastatic LNs can further gain access to nodal blood vessels, including high endothelial venules (HEVs), and disseminate to distant organs. Thus, tdLNs may serve not only as regional metastatic sites but also as anatomical intermediates linking lymphatic spread to systemic dissemination. (B) As immune reservoirs, the same lymphatic route transports tumor antigens and antigen-bearing dendritic cells (DCs) from the tumor bed to tdLNs. Within tdLNs, conventional type 1 DC (cDC1)-mediated antigen presentation promotes tumor-specific T cell priming and supports the maintenance of progenitor-exhausted T cells (TPEX). These cells can exit tdLNs and give rise to more differentiated exhausted T cells (TEX) that traffic to primary tumors and micrometastatic lesions. This dual role provides a conceptual basis for understanding why the timing of immune checkpoint inhibition relative to tumor and LN removal may influence antitumor immunity.

Figure 2.
The tdLN–TPEX/TEX axis as the cellular basis of PD-1 blockade. This schematic illustrates a proposed cellular model in which PD-1 blockade acts primarily by mobilizing a renewable supply of tumor-reactive T cells from tumor-draining lymph nodes (tdLNs), rather than by simply restoring terminally exhausted T cells (TEX) within the tumor. In tdLNs, cDC1-mediated presentation of tumor antigens supports the generation and maintenance of TCF1+ progenitor-exhausted T cells (TPEX), a less differentiated CD8⁺ T cell population that expresses PD-1 but retains proliferative capacity. PD-1 blockade can promote expansion of this TPEX pool and facilitate the continuous supply of activated tumor-specific T cells through blood and lymphatic circulation. After entering the tumor microenvironment, these cells can differentiate along an exhaustion continuum, including intermediate TEX cells, effector-like TEX cells, and terminal TEX cells. Effector-like TEX cells may contribute directly to tumor control, whereas terminal TEX cells are more differentiated, have limited proliferative potential, and are less likely to be fully reverted by PD-1 blockade. This model emphasizes the importance of maintaining tdLN-based TPEX reservoirs for sustained antitumor immunity during immune checkpoint therapy.
Figure 2.
The tdLN–TPEX/TEX axis as the cellular basis of PD-1 blockade. This schematic illustrates a proposed cellular model in which PD-1 blockade acts primarily by mobilizing a renewable supply of tumor-reactive T cells from tumor-draining lymph nodes (tdLNs), rather than by simply restoring terminally exhausted T cells (TEX) within the tumor. In tdLNs, cDC1-mediated presentation of tumor antigens supports the generation and maintenance of TCF1+ progenitor-exhausted T cells (TPEX), a less differentiated CD8⁺ T cell population that expresses PD-1 but retains proliferative capacity. PD-1 blockade can promote expansion of this TPEX pool and facilitate the continuous supply of activated tumor-specific T cells through blood and lymphatic circulation. After entering the tumor microenvironment, these cells can differentiate along an exhaustion continuum, including intermediate TEX cells, effector-like TEX cells, and terminal TEX cells. Effector-like TEX cells may contribute directly to tumor control, whereas terminal TEX cells are more differentiated, have limited proliferative potential, and are less likely to be fully reverted by PD-1 blockade. This model emphasizes the importance of maintaining tdLN-based TPEX reservoirs for sustained antitumor immunity during immune checkpoint therapy.

Figure 3.
Distinct immunological roles of neoadjuvant and adjuvant immune checkpoint inhibition. Neoadjuvant and adjuvant immune checkpoint inhibition (ICI) have different immunological roles during perioperative treatment. Before surgery, the primary tumor, tumor antigens, lymphatic drainage, dendritic cell migration, and tumor-draining lymph node (tdLN)-based T cell priming remain intact. Neoadjuvant ICI can therefore activate systemic antitumor immunity through the preserved tumor–lymphatic–tdLN axis. Surgery removes the primary tumor and may partially or extensively remove regional tdLNs, while also altering lymphatic architecture and allowing pathological response assessment. After surgery, minimal residual disease (MRD), circulating tumor cells (CTCs), disseminated tumor cells (DTCs), and micrometastases may remain. Some dormant or residual tumor cells may become immunologically visible only later, and emerging metastatic lesions may establish new drainage to metastatic-site lymph nodes. Adjuvant ICI may therefore maintain immune surveillance and immune pressure against MRD and developing metastatic niches during this uncertain postoperative period.
Figure 3.
Distinct immunological roles of neoadjuvant and adjuvant immune checkpoint inhibition. Neoadjuvant and adjuvant immune checkpoint inhibition (ICI) have different immunological roles during perioperative treatment. Before surgery, the primary tumor, tumor antigens, lymphatic drainage, dendritic cell migration, and tumor-draining lymph node (tdLN)-based T cell priming remain intact. Neoadjuvant ICI can therefore activate systemic antitumor immunity through the preserved tumor–lymphatic–tdLN axis. Surgery removes the primary tumor and may partially or extensively remove regional tdLNs, while also altering lymphatic architecture and allowing pathological response assessment. After surgery, minimal residual disease (MRD), circulating tumor cells (CTCs), disseminated tumor cells (DTCs), and micrometastases may remain. Some dormant or residual tumor cells may become immunologically visible only later, and emerging metastatic lesions may establish new drainage to metastatic-site lymph nodes. Adjuvant ICI may therefore maintain immune surveillance and immune pressure against MRD and developing metastatic niches during this uncertain postoperative period.

Figure 4.
Postoperative lymphatic dysfunction as a component of postoperative immune ecology. Lymph node dissection and radiotherapy can disrupt regional lymphatic networks after cancer treatment. This disruption may impair lymph flow, residual antigen transport, dendritic cell (DC) migration, and T cell trafficking. Chronic lymphatic dysfunction can subsequently promote tissue remodeling, including lymphedema, fibrosis, persistent inflammation, macrophage polarization, and regulatory T cell (Treg) accumulation. These changes may alter the local immune environment by affecting immune surveillance, residual antigen drainage, and the response to adjuvant immune checkpoint inhibition (ICI). However, direct clinical evidence linking postoperative lymphedema or fibrosis to increased cancer recurrence remains limited. Therefore, postoperative lymphatic dysfunction should be regarded as a potential modifier of local immune surveillance and postoperative immune ecology, rather than as an established cause of recurrence.
Figure 4.
Postoperative lymphatic dysfunction as a component of postoperative immune ecology. Lymph node dissection and radiotherapy can disrupt regional lymphatic networks after cancer treatment. This disruption may impair lymph flow, residual antigen transport, dendritic cell (DC) migration, and T cell trafficking. Chronic lymphatic dysfunction can subsequently promote tissue remodeling, including lymphedema, fibrosis, persistent inflammation, macrophage polarization, and regulatory T cell (Treg) accumulation. These changes may alter the local immune environment by affecting immune surveillance, residual antigen drainage, and the response to adjuvant immune checkpoint inhibition (ICI). However, direct clinical evidence linking postoperative lymphedema or fibrosis to increased cancer recurrence remains limited. Therefore, postoperative lymphatic dysfunction should be regarded as a potential modifier of local immune surveillance and postoperative immune ecology, rather than as an established cause of recurrence.


Table 1.
Cancer-type-specific patterns of perioperative or adjuvant immune checkpoint inhibitor strategies viewed through the tdLN–TPEX/TEX axis.
Table 1.
Cancer-type-specific patterns of perioperative or adjuvant immune checkpoint inhibitor strategies viewed through the tdLN–TPEX/TEX axis.
| Cancer type | Representative strategy | Main clinical implication | Immunological interpretation |
|---|---|---|---|
| Melanoma | NADINA: neoadjuvant ipilimumab plus nivolumab followed by surgery and response-driven adjuvant therapy [6] | Neoadjuvant ICI enables pathological response-guided reduction or escalation of adjuvant therapy | High tumor antigenicity and preserved tdLNs may allow strong systemic T cell priming before surgery. Pathological response can serve as an early readout of effective T cell activation. |
| NSCLC | KEYNOTE-671: perioperative pembrolizumab plus chemotherapy [7]; CheckMate 77T: perioperative nivolumab plus chemotherapy [44] | Perioperative chemoimmunotherapy improves event-free survival and may reduce systemic relapse | The primary tumor and mediastinal tdLNs may support systemic antitumor T cell induction before resection, while postoperative ICI may maintain immune pressure against MRD. |
| HNSCC | KEYNOTE-689: neoadjuvant and adjuvant pembrolizumab added to surgery and standard adjuvant therapy [43] | Perioperative pembrolizumab improves event-free survival in resectable locally advanced HNSCC | Cervical tdLNs may function as immune reservoirs where TPEX cells are maintained and mobilized by PD-1 blockade, supporting continuous TEX supply to tumor and micrometastatic sites. |
| TNBC | KEYNOTE-522: neoadjuvant pembrolizumab plus chemotherapy followed by adjuvant pembrolizumab [45] | Perioperative pembrolizumab improves pathological complete response, event-free survival, and overall survival in high-risk early-stage TNBC | Chemotherapy-induced tumor cell death, antigen release, and inflammatory remodeling may cooperate with PD-1 blockade to enhance T cell priming and systemic antitumor immunity. |
| Muscle-invasive bladder cancer | NIAGARA: neoadjuvant durvalumab plus gemcitabine/cisplatin followed by radical cystectomy and adjuvant durvalumab [46] | Perioperative PD-L1 blockade improves event-free survival and overall survival in cisplatin-eligible muscle-invasive bladder cancer | Neoadjuvant chemoimmunotherapy may promote antigen release and immune activation before cystectomy, whereas adjuvant durvalumab may sustain immune surveillance against MRD. |
| dMMR colon cancer | NICHE-2: short-course neoadjuvant nivolumab plus ipilimumab before surgery [47] | Short-course neoadjuvant dual checkpoint blockade induces marked pathological responses in locally advanced dMMR colon cancer | High neoantigen burden and pre-existing immune recognition in dMMR tumors may permit rapid expansion of tumor-specific T cells when checkpoint inhibition is delivered before tumor removal. |
| Renal cell carcinoma | KEYNOTE-564: adjuvant pembrolizumab after nephrectomy [42]; PROSPER RCC: perioperative nivolumab before and after nephrectomy [48] | Adjuvant pembrolizumab improves disease-free and overall survival, whereas perioperative nivolumab has not shown recurrence-free survival benefit | In RCC, the current evidence supports postoperative MRD control more strongly than neoadjuvant immune priming. The negative perioperative nivolumab result suggests that optimal timing, regimen, and immune context may differ from other solid tumors. |
Table 2.
Key immune monitoring targets for perioperative immunotherapy.
| Compartment | Suggested analyses | Biological meaning |
|---|---|---|
| Primary tumor | CD8 T cells, TEX, TLS, PD-L1, antigen presentation machinery | Local immune activation and tumor immune contexture |
| tdLN | TPEX / TCF1+ CD8 T cells, cDC1, TCR clonality, PD-1/PD-L1 axis | Immune reservoir and priming capacity |
| Blood | TCR expansion, Ki-67+ CD8 T cells, ctDNA, cytokines | Systemic immune activation and MRD |
| Metastatic site | Tumor antigen expression, local draining LN response, T cell infiltration | Metastatic-site immune reactivation |
| Postoperative tissue | lymphatic vessels, fibrosis, macrophages, Tregs, T cell trafficking | Postoperative immune ecology |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



