Submitted:
08 June 2026
Posted:
09 June 2026
You are already at the latest version
Abstract
Background/Objectives: The homeostatic imbalance between tumor metastasis-promoting genes and -suppressing genes determines the metastatic potential of lung adenocarcinoma (LUAD) cells. However, the post-transcriptional regulation mechanism mediated by RNA-binding proteins (RBPs) in maintaining the expression balance of tumor metastasis-related genes remains unclear. Methods: The expression of MCPIP3 and IGF2BP3 in human lung cancer tissues was analyzed using bioinformatics methods and validated qRT–PCR, immunohistochemistry, and immunoblotting. In vitro cellular experiments and in vivo animal examinations were used to evaluate the effects of MCPIP3 and IGF2BP3 on metastasis. RNA-sequencing, PCRarray, RNA pull-down and mass spectrometry, RNA-EMSA, RNA immunoprecipitation (RIP), mRNA stability, luciferase assay, were performed to elucidate the mechanism. Results: MCPIP3 is expressed at low levels, while IGF2BP3 is highly expressed in LUAD tissues. Lower MCPIP3 expression and higher IGF2BP3 expression in lung cancer tissues were significantly associated with poor prognosis in LUAD patients. MCPIP3 significantly inhibited the metastasis of lung cancer cells both in vitro and in vivo, whereas IGF2BP3 promoted metastasis. Mechanistically, MCPIP3 specifically bound to the stem‒loop structure in the 3'UTR (untranslated region) of METAP2 transcript, and degraded its mRNA via its RNase domain. In contrast, by interacting with the common stem‒loop structure, IGF2BP3 could stabilize METAP2 mRNA and promote its expression. Notably, inverse correlations were observed between the expression of MCPIP3 and IGF2BP3 and the expression of METAP2 in LUAD tissues. Conclusions: MCPIP3 and IGF2BP3 antagonistically control lung cancer cell metastasis by balancing the expression of metastasis-promoting genes through mediating mRNA stability, thereby providing potential targets for lung cancer treatment.
Keywords:
MCPIP3
; IGF2BP3
; LUAD
; Metastasis
; METAP2
1. Introduction
Lung cancer is among the most common cancers worldwide. Although the 5-year survival rate for lung cancer patients has significantly improved because of surgical treatment and early diagnosis, many patients still experience cancer metastasis, which subsequently leads to organ failure and cancer-related death [1]. Therefore, further elucidation of the mechanisms underlying lung cancer cell metastasis is necessary and may provide promising therapeutic targets. The metastasis of lung cancer cells is determined by the expression of metastasis-promoting genes, such as Methionine aminopeptidase 2 (METAP2)[2], Harvey rat sarcoma viral oncogene homolog (HRAS)[3], Pinin (PNN)[4], and metastasis-inhibiting genes, such as metastasis suppressor 1 (MTSS1)[5], catenin alpha 1 (CTNNA1)[6], and cadherin 11 (CDH11)[7], in cancer cells. Lung cancer cell metastasis is usually activated either by the upregulation of metastasis-promoting genes or by the downregulation of metastasis-suppressing genes. However, the molecular mechanism controlling the expression of metastasis-related genes remains unclear, especially the mechanisms by which multiple RBPs antagonistically regulate the expression of these genes at the post-transcriptional level.
Monocyte chemotactic protein-induced protein 3 (MCPIP3), encoded by the zc3h12c gene, is a CCCH-type zinc finger RNA-binding protein belonging to the ZC3H12 family [8]. MCPIP3 has endonuclease activity and degrades mRNA through its RNase domain [9]. Extensive research has focused on the immunomodulatory function of MCPIP3, which can prevent autoimmune reactions by controlling the expression of pro-inflammatory cytokine gene [10]. MCPIP3 knockout mice develop hypertrophic lymph nodes [11]. MCPIP3 is also involved in the pathogenesis of psoriasis [12] and skin inflammation [13] through the mediation of inflammatory responses. Therefore, MCPIP3 is believed to be an important regulator of inflammation homeostasis. To date, the role of MCPIP3 in human cancer progression remains unclear. It has been reported that the expression of MCPIP3 is closely correlated with the survival of patients with lung cancer [14], colon cancer [15], and kidney cancer [16]. Moreover, MCPIP3 also is involved in regulating the metastasis of human colon cancer cells [17] and is considered as a tumor suppressor in colon cancer [18]. However, the role and mechanism of MCPIP3 in human lung cancer development, especially in metastasis progression, remain unclear.
Methionine aminopeptidase 2 (MetAP2), a metallopeptidase known to play an important role in tumor progression, is considered a key promoter of tumor metastasis [19], growth [20], and angiogenesis [21]. METAP2 may be an important functional protein for the treatment and outcome of prostate cancer patients [22]. METAP2 has been found to promote the metastasis of various human cancers, including colon cancer, breast cancer, and ovarian cancer, indicating that METAP2 is an important regulator of cancer metastasis progression [2,23,24]. METAP2 is also believed to be a promising molecular target for treating cancer metastasis [25,26]. However, the mechanism controlling METAP2 gene expression in cells, especially in lung cancer cells, remains unclear.
In this study, we discovered that MCPIP3 acts as a potent lung cancer metastasis suppressor by suppressing the expression of the tumor metastasis-promoting gene METAP2. Computational analysis revealed that MCPIP3 expression was significantly reduced among 1756 RBPs analyzed in lung cancer tissues. Low levels of MCPIP3 in lung cancer tissues were strongly correlated with poor prognosis in lung cancer patients. We found that MCPIP3 was able to inhibit the migration and invasion of lung cancer cells both in vitro and in vivo. MCPIP3 could specifically downregulate the expression of the tumor metastasis-promoting gene METAP2 by degrading its mRNA through direct binding to the stem‒loop structure in the 3’UTR. Interestingly, IGF2BP3, another typical RBP [27], was identified to bind to the stem‒loop structure of METAP2 mRNA while stabilizing its mRNA. We further found that IGF2BP3 could antagonize MCPIP3-mediated inhibition of METAP2 expression and promote lung cancer cell metastasis. Moreover, we found that the expression of MCPIP3 and IGF2BP3 was significantly negatively and positively correlated with METAP2 expression, respectively, in human lung cancer tissues. Collectively, these results revealed a post-transcriptional regulatory mechanism of metastasis-promoting genes mediated by the RBPs MCPIP3 and IGF2BP3, which antagonistically control the stability and expression of these genes in lung cancer cells, thereby providing potential molecular targets for lung cancer treatment.
2. Materials and Methods
2.1. Cell Lines and Plasmids
Human lung cancer cell lines (A549, NCI-H23) were obtained from the American Type Culture Collection (ATCC) and cultured in PRM1640 medium with 10% FBS plus 1% Peni/Stro, respectively. HEK293, and HEK293T cells were obtained from National Infrastructure of Cell Line Resource (Beijing, China). The human full-length MCPIP3 (NM_033390.2) and IGF2BP3 (NM_006547.3) coding sequence was synthesized, sequenced and inserted into pEGFP-N1 vector at NheI/SacII and NheI/SacI sites, respectively. MCPIP3 serial truncation constructs were generated by inserting the amplified fragments into pEGFP-N1 vector at NgeI and SacII sites. The luciferase reporters were constructed by inserting the full-length 3’UTRs of human MERAP2, HRAS, PNN, and CTBP1 into the pGL3 control vector between Xba I and Fse I sites, respectively. For stem–loop mutation reporters, point mutated and truncated METAP2-3’UTR (∆stem–loop) were amplified, sequenced, and inserted into pGL3 control vector using Phusion Site-Directed Mutagenesis Kit (Thermo Scientific).
2.2. Wound Healing Assay
Lung cancer cells overexpressing or knocking down MCPIP3 were seeded in 35 mm dishes and incubated at 37 ℃. After the cells grow to a monolayer, cell scratches were created using 200 μL pipette tips. The cells were continued to culture for 24-48 hours. The images of the scratch confluence were acquired using a microscope (Zeiss, Germany). The open area of the wound was measured using Zeiss ZEN software.
2.3. Transwell Invasion Assay
The transwell inserts of 24-well Transwell plates (8.0 μm pore, Corning) were precoated with 50 μL Matrigel (BD Biosciences) and incubated for 1 h at 37 ℃. Then, MCPIP3-overexpressing lung cancer cells suspended in 200 μL of serum-free medium were seeded in transwell inserts, whereas the lower wells were filled with 800 µl of conditioned medium. After 24 hours of culture, the inserts were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The remaining cells were removed by a cotton swab from the inner side of the inserts. Each inverted inserts were photographed to visualize the invading cells. The number of invaded cells were normalized to the control.
2.4. RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
Total RNA from indicated cells was extracted using TRIzol (Thermo Fisher, Waltham, USA) and reverse transcribed into cDNA for real-time qPCR using SYBR green Fast Master Mix (Roche). Relative gene expression level was calculated based on the ∆∆Ct method and normalized using GAPDH. The sequences of primers are shown in Table S1.
2.5. PCR Array Assay
The metastasis-related genes expression was quantified using the QIAGEN Human Tumor Metastasis PCR Array Kit (Cat. No. 330231, PAHS-028ZA) in MCPIP3-overexpressing A549 cells and empty vector control cells. The expression fold changes of each gene were calculated by the 2–∆∆Ct method with GAPDH as a reference. The detailed PCRarray data are shown in Table S2.
2.6. RNA Sequencing Analysis
RNA sequencing was carried out by Allwegene Technology Inc., Beijing with the PE150 sequencing strategy using an Illumina second-generation high-throughput sequencing platform. The raw reads with inferior quality or adapters were filtered. Clean reads were processed using Tophat2 and Cufflinks to complete the alignment of transcriptomes. Genes not expressed in any sample were excluded from further analysis. Differentially expressed genes and transcripts were then filtered for false discovery rate (FDR)-adjusted P values less than or equal to 0.05. The regulated genes were listed in Table S3.
2.7. mRNA Stability Assay
Actinomycin D (ActD, 5 µg/mL) and 5, 6-dichlorobenzimidazole riboside (DRB, 5 µg/mL) were added into target cells to block de novo RNA synthesis. The relative mRNA levels at different time points were measured by RT-qPCR. mRNA half-lives were determined by comparing the mRNA levels before treating with ActD and DRB.
2.8. RNA-EMSA (Electrophoretic Mobility Shift Assay)
3’-end biotin-labeled probes were incubated with total cell lysates containing MCPIP3/GFP fusion protein for 30 min at room temperature with binding buffer. RNA-protein complexes were separated by non-denaturing polyacrylamide gel (6%) electrophoresis and then transferred onto a nylon membrane and crosslinked by UV irradiation. The membrane was detected with the LightShift chemiluminescent EMSA kit (Pierce) following the manufacturer’s guide.
2.9. Western Blot Analysis
The cells or tissue samples were lysed with RIPA buffer containing PMSF and protease inhibitor cocktail (Roche), and the protein concentration was determined by BCA assay. The protein lysates were subjected to SDS-PAGE electrophoresis and transferred onto polyvinylidene membrane. The blotted membrane was then blocked with 5% fat-free milk for 1 hour and then incubated with primary antibody (dilution: 1:1000 in 5% fat-free milk) overnight at 4℃. After incubation with HRP-conjugated secondary antibodies (dilution: 1:5000 in 5% fat-free milk), the band was detected using ECL chemiluminescent detection kit (#34077, Pierce; Rockford, IL, USA).
2.10. RNA Pull-Down and Mass Spectrometry
3’-biotin-labeled RNA probes with stem‒loop or without stem‒loop were used for RNA pull-down. Whole cell lysates were obtained from A549 cells and were precleared with streptavidin-coated Dynabeads (M-280, Invitrogen), and then incubated with RNA probes and Dynabeads for 2h at 4℃. After washed six times, the beads were boiled in SDS-PAGE sample buffer. The co-precipitated proteins were separated by SDS-PAGE followed by silver staining. The unique bands were excised and in-gel digested with trypsin for mass spectrometry (LC‒MS) analysis. The mass spectrometry analysis was completed by Beijing Qinglian Biotech Co., Ltd. (Beijing, China) with RIGOL L-3000 HPLC System (RIGOL, Beijing, China). The identified proteins were listed in Table S4.
2.11. Animal Study
6-8 weeks old female BALB/c nude mice were purchased from the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences (CAMS) & Peking Union Medical College (PUMC). The mice were bred in a laminar flow hood under pathogen-free condition with a 12h light/dark cycle. All experimental procedures were approved by the Experimental Animal Care and Ethics Committee of the Institute of Microcirculation, CAMS & PUMC. A549/MCPIP3-GFP (2.0 × 106/100µL PBS), A549/GFP cells (2.0 × 106/100µL PBS); A549/shMCPIP3 (2.0 × 106/100µL PBS), A549/Scramble cells (2.0 × 106/100µL PBS), and A549/IGF2BP3-GFP (2.0 × 106/100µL PBS) were injected subcutaneously into the nude mice, respectively. Then, tumor size was measured every other day and calculated by the formula length × width × high (mm3).
2.12. Statistical Analysis
Statistical analysis between groups were analyzed by GraphPad Prism 9. Data were expressed as mean ± SD based on at least three independent repeats. The statistical analyses using Student’s t-test for comparisons between treatment and vehicle control. Spearman’s correlation analysis was used to evaluate the relationship between two different genes expression. Statistical significance was considered when P < 0.05.
3. Results
3.1. MCPIP3 Expression Is Reduced in Human Lung Cancer Tissues and Is Associated with Poor Survival
To demonstrate the expression patterns of all the RBPs in human lung cancer, we analyzed the mRNA expression of 1756 RBPs in lung cancer tissues by surveying publicly available datasets from The Cancer Genome Atlas (TCGA)[28]. MCPIP3 was found to be expressed at lower levels in lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) tissues and ranked among the lowest level RBP candidates (Figure 1A; Figure S1A). Several RBPs that have been verified to be either downregulated or upregulated in lung cancer tissues, were also identified, such as KH domain-containing, RNA-binding, signal transduction-associated protein 2 (KHDRBS2)[29], CUGBP Elav-like family member 2 (CELF2)[30], IGF2BP3[31], and tRNA (guanine-N(7)-)-methyltransferase (METTL1)[32] (Figure 1A), further confirming the reliability of our computational analysis. We further found that the expression of MCPIP3 mRNA was significantly reduced in LUAD (Figure 1B) and LUSC (Figure S1B, S1C) tissues. MCPIP3 expression was also lower in our collected LUAD tissues than in the adjacent non-cancerous tissues (Figure 1C). MCPIP3 protein expression was also reduced in LUAD tissues (Figure 1D). Western blot and immunohistochemistry (IHC) results also confirmed a low level of MCPIP3 expression in LUAD tissues (Figure 1E, F). Notably, the expression of MCPIP3 decreases with increasing lymph node metastasis in patients with LUAD (Figure 1G) and LUSC (Figure S1D). We further found that MCPIP3 expression was lower in LUAD and LUSC tissues at higher stages (Figure 1H, Figure S1E), and that the protein expression of MCPIP3 decreased with increasing cancer grade in LUAD patients (Figure 1I). These findings strongly suggest that MCPIP3 expression might be closely associated with human lung cancer metastasis. Additionally, the results of Kaplan–Meier analyses indicated that lower MCPIP3 expression in lung cancer tissues was significantly associated with poor survival in lung cancer patients (Figure 1J, K). Overall, these results suggest that MCPIP3 expression is suppressed in LUAD tissues and is strongly associated with lung cancer metastasis.
3.2. MCPIP3 Inhibits Lung Cancer Cell Metastasis by Mediating the Expression of Tumor Metastasis-Related Genes
Next, we investigated the effect of MCPIP3 on the metastasis of lung cancer cells. Exogenous MCPIP3 was expressed in A549 cells and NCI-H23 cells, respectively (Figure S2A). Ectopic expression of MCPIP3 significantly inhibited the migration (Figure 2A) and invasion (Figure 2B) of lung cancer cells in vitro. In vivo experimental results further revealed that, compared with the control treatment, the overexpression of MCPIP3 significantly suppressed the growth (Figure 2C, D) and lung metastasis of A549 tumors (Figure 2E), indicating that MCPIP3 could effectively inhibit lung cancer cell metastasis. Next, RNA sequencing (RNA-seq) was conducted to identify the downstream genes regulated by MCPIP3 in lung cancer cells. A total of 4,971 genes were commonly upregulated, while 5,290 genes were commonly downregulated by MCPIP3 (Figure S2B; Table S3). Gene Ontology (GO) analysis revealed that several terms associated with cancer cell metastasis, including cell migration, wound healing, and cell motility (Figure 2F), were significantly enriched, indicating that MCPIP3 might regulate the expression of tumor metastasis-related genes in lung cancer cells. We found that MCPIP3 could downregulate the expression of a series of metastasis-promoting genes (METAP2, HRAS, PNN, and CTBP1) and upregulate the expression of many metastasis-suppressing genes (MTSS1, CTNNA1, and CDH11) in A549 and NCI-H23 cells (Figure 2G). To further identify the genes affected by MCPIP3 in the tumor metastasis signaling pathway, a PCRarray detection was performed using the Human Tumor Metastasis RT2 Profiler PCR Array (QIAGEN). Indeed, we found that a total of 15 metastasis-promoting genes were downregulated, and 13 metastasis-suppressing genes were upregulated in A549 cells overexpressing MCPIP3 (Figure 2H). Although the upregulated and downregulated genes were not exactly the same due to differences in the sensitivities of detection methods, the trend of MCPIP3 downregulating the expression of metastasis-promoting genes and upregulating the expression of metastasis-suppressing genes was very clear. In addition, qPCR results confirmed that the mRNA expression of four metastasis-promoting genes (METAP2, HRAS, PNN, CTBP1) was downregulated, whereas the expression of four metastasis-suppressing genes (MTSS1, CTNNA1, CDH11, CD82) was upregulated in lung cancer cells overexpressing MCPIP3 (Figure 2I; Figure S2C), further validating our RNA-seq and PCRarray results. Taken together, these data demonstrate that MCPIP3 can inhibit the metastasis of human lung cancer cells by regulating the expression of tumor metastasis-related genes.
3.3. MCPIP3 Selectively Downregulated the Expression of the Metastasis-Promoting Gene METAP2 by Destabilizing Its mRNA via the RNase Domain
Next, we aimed to investigate how MCPIP3 regulates the expression of metastasis-related genes in lung cancer cells. Given that MCPIP3 is considered as an RBP, we investigated whether it directly binds to the transcripts of metastasis-related genes in lung cancer cells. RNA immunoprecipitation (RIP) was performed using specific anti-GFP antibodies and isotype IgG followed by RT‒PCR detection. We found that the metastasis-promoting genes, but not those of metastasis-suppressing genes, could be amplified in the complexes precipitated by the anti-GFP antibody (Figure 3A; Figure S3A), indicating that MCPIP3 specifically binds to metastasis-promoting genes but not metastasis-suppressing genes. Since MCPIP family proteins recognize the 3’UTRs of their target genes, we further investigated whether MCPIP3 binds to the 3’UTRs of metastasis-promoting genes. A luciferase reporter assay revealed that MCPIP3 downregulated the activity of a luciferase reporter containing the 3’UTR of metastasis-promoting genes (Figure 3B; Figure S3B), indicating that MCPIP3 binds to these mRNAs by targeting the 3’UTR. Previous studies have shown that MCPIPs can interact with the stem‒loop structure in the 3’UTR [33]. We found that similar stem‒loop structures exist in the 3’UTRs of metastasis-promoting genes by aligning the 3’UTRs sequences among different species (Figure S3C). To verify whether MCPIP3 also targets the stem‒loop structure of metastasis-promoting genes, two METAP2 3’UTR mutant reporters were constructed, in which several nucleotides were substituted; Mut1 lost the stem‒loop structure, whereas Mut2 maintained it (Figure 3C). Deletion of the stem‒loop structure prevented MCPIP3 from inhibiting luciferase activity (Figure 3D). However, restoration of the stem‒loop structure enabled MCPIP3 to suppress reporter activity, indicating that the stem‒loop conformation is crucial for MCPIP3 targeting the 3’UTRs of metastasis-promoting genes (Figure 3D). Our RNA-EMSA results showed that a unique RNA‒Protein complex formed only with stem‒loop probes but not with the mutated linearized probes (Figure 3E), indicating that a physical interaction between MCPIP3 and the RNA stem‒loop structure. RNA immunoprecipitation–chromatin immunoprecipitation (RIP–ChIP) results further confirmed that the stem‒loop sequence of METAP2 could be amplified from the complexes pulled down by the anti-GFP antibody, but not from the isotype IgG group (Figure 3F), indicating that MCPIP3 can bind to the stem‒loop sequence of METAP2 in lung cancer cells. We further found that MCPIP3 could effectively reduce the half-lives of METAP2, PNN, and CTBP1 mRNAs in lung cancer cells (Figure 3G; Figure S3D), which was consistent with previous reports [34]. Western blot analysis also confirmed that METAP2 protein expression was downregulated in a time-dependent manner in MCPIP3-overexpressing lung cancer cells (Figure 3H).
MCPIP proteins have been reported to degrade mRNA through the RNase domain. To determine whether the RNase domain is also responsible for MCPIP3-mediated mRNA degradation, two truncation mutants were generated: the amino acid 1-401 (aa 1-401) mutant, which contains the RNase domain, and the aa 401-883 mutant, which lacks the RNase domain (Figure 3I). We found that wild-type (wt) and aa 1-401 mutant MCPIP3 could effectively inhibit the expression and reduce the half-life of METAP2 mRNA, while the aa 401-883 mutant MCPIP3 completely lost this inhibitory effect (Figure 3I-K). These data suggest that the RNase domain is crucial for MCPIP3-mediated metastasis-promoting mRNA degradation. Additionally, we showed that the expression level of METAP2 was significantly negatively correlated with the survival of LUAD patients (Figure 3L). Taken together, these findings indicated that MCPIP3 can effectively reduce the stability and expression of the metastasis-promoting gene METAP2 transcript in human lung cancer cells through its RNase domain.
3.4. Knockdown of MCPIP3 Increases the Stability of METAP2 mRNA and Promotes Lung Tumor Metastasis Progression
To further confirm the inhibitory effect of MCPIP3 on lung cancer cell metastasis, MCPIP3 was knocked down in human lung cancer cells using two shRNAs. Approximately 54% and 65% of MCPIP3 protein expression was silenced by shRNA#1 and shRNA#2, respectively, in human lung cancer cells (Figure 4A). Suppression of MCPIP3 expression significantly promotes lung cancer cell migration (Figure 4B) and increased the expression of metastasis-promoting genes (Figure 4C). We further found that the half-lives of METAP2 and PNN mRNAs were increased after MCPIP3 knockdown in lung cancer cells (Figure 4D; Figure S4A). To further confirm the involvement of METAP2 in the MCPIP3-mediated inhibition of lung cancer cell metastasis, the METAP2 gene was knocked down by infecting A549 MCPIP3/shRNA#2 cells with a shRNA-expressing lentivirus (Figure 4E). Compared with MCPIP3 knockdown alone, co-knockdown of MCPIP3 and METAP2 partially restored the migration ability of A549 cells to the control group level, indicating that the metastasis-promoting gene METAP2 is indeed involved in the MCPIP3-mediated inhibition of lung cancer cell metastasis (Figure 4F; Figure S4B). In vivo experiments further demonstrated that silencing MCPIP3 promoted the growth (Figure 4G, H) and metastasis (Figure 4I) of A549 xenograft tumors. These results demonstrated that reducing MCPIP3 expression increases the expression of metastasis-promoting genes and facilitates lung cancer cell metastasis.
3.5. The RNA-Binding Protein IGF2BP3 Interacts with the Common Stem‒Loop Structure to Stabilize METAP2 mRNA and Promote Lung Cancer Metastasis
Interestingly, although several nucleotide differences exist in the stem‒loop sequences of METAP2 among human, mice, and rat, these conserved sequences can fold into a stem‒loop structure (Figure 5A), indicating that this structural stem‒loop conformation might be important for controlling METAP2 gene expression. Given that MCPIP3 can destabilize METAP2 mRNA by binding to the stem‒loop structure and METAP2 is highly expressed in human lung cancer cells, it is likely that other RBPs stabilize METAP2 mRNA and increase its expression by targeting the common stem‒loop structure. To confirm this possibility, RNA pull-down experiments combined with mass spectrometry analysis were performed using biotin-labeled stem–loop probes and linearized RNA probes (RNA was heated to 95 °C and then rapidly cooled on ice to eliminate RNA structure). Several unique protein bands were enriched by the stem–loop RNA probes compared with the linearized RNA probes group (Figure 5B). Notably, many typical RBPs, such as IGF2BP1-3, polypyrimidine tract-binding protein 1 (PTBP1), and RNA-binding motif protein 39 (RBM39), were identified based on their molecular weight and peptides number, further confirming the reliability of our RNA pull-down results (Figure 5C; Table S4). IGF2BP3 was selected to verify its interaction with METAP2 stem–loop structure. Indeed, IGF2BP3 expression was detected in proteins pulled down using stem–loop probes but not in the linearized probe group, as demonstrated by the results of RNA pull-down followed by Western blotting (Figure 5D). Moreover, RIP-CHIP results revealed that the METAP2 stem–loop sequence could be amplified from the RNAs precipitated using the anti-GFP antibody but not from the isotype IgG group (Figure 5E). These results indicate that IGF2BP3 indeed interacted with the stem–loop structure of METAP2 in human lung cancer cells. Next, we examined the genes regulated by IGF2BP3 using RNA-seq in IGF2BP3-overexpressing A549 cells. Interestingly, GO analysis revealed that the genes regulated by IGF2BP3 were significantly enriched in terms related to cell migration (Figure 5F), suggesting that IGF2BP3 may regulate the expression of metastasis-related genes in human lung cancer cells. Notably, several metastasis-promoting genes, including PNN, METAP2, and CTBP1 (Figure 5G, H; Figure S5A), were upregulated by IGF2BP3 (Table S3), whereas their expression was downregulated by MCPIP3. These findings suggested that IGF2BP3 and MCPIP3 regulate the expression of metastasis-promoting genes in human lung cancer cells in positive and negative manner, respectively. Moreover, METAP2 protein expression was also increased by IGF2BP3 in human lung cancer cells (Figure 5I). Importantly, IGF2BP3 effectively antagonized the MCPIP3-induced shortening of the half-life and reduction in the expression of METAP2 mRNA in human lung cancer cells (Figure 5J; Figure S5B). As expected, we found that IGF2BP3 overexpression significantly promoted A549 cell migration (Figure 5K). Overall, our results demonstrated that IGF2BP3 stabilizes METAP2 mRNA by targeting the stem‒loop structure and promoting lung cancer cell migration.
3.6. IGF2BP3 and MCPIP3 Potentially Antagonize Lung Cancer Cell Metastasis by Reversibly Modulating METAP2 Expression
Next, we analyzed the expression pattern of IGF2BP3 in human lung cancer tissues. Both IGF2BP3 mRNA (Figure 6A; Figure S6A) and protein (Figure 6B; Figure S6B) expression were significantly increased in LUAD and LUSC tissues. Notably, IGF2BP3 expression increased with increasing degree of lymph node metastasis in LUAD patients (Figure 6C), which was opposite to the trend observed for MCPIP3 expression (Figure 1G). Similarly, high levels of IGF2BP3 expression in cancer tissues were significantly associated with poor survival in LUAD patients (Figure 6D), whereas low levels of MCPIP3 expression were closely associated with poor survival (Figure 1J). Unsurprisingly, IGF2BP3 promoted the growth (Figure S6C) and metastasis (Figure 6E) of the A549 tumor model, which is consistent with the findings of previous reports [35]. We further discovered that METAP2 expression was significantly increased in IGF2BP3-overexpressing xenografts, but decreased in MCPIP3-overexpressing xenografts (Figure 6F). Moreover, METAP2 expression was significantly negatively correlated with MCPIP3 expression but positively correlated with IGF2BP3 expression in LUAD tissues (Figure 6G). Histological analysis also revealed that the staining intensity of METAP2 was significantly greater in MCPIP3-negative (MCPIP3-Neg) LUAD tissues than in MCPIP3-positive (MCPIP3-Pos) LUAD tissues (Figure 6H). However, the staining intensity of METAP2 protein was higher in IGF2BP3-Positive (IGF2BP3-Pos) LUAD tissues than in IGF2BP3-Negative (IGF2BP3-Neg) LUAD tissues (Figure 6H). These findings highlight that MCPIP3 and IGF2BP3 may mediate METAP2 expression in human lung cancer tissues in negative and positive manners, respectively. We also found that the expression of many metastasis-promoting genes was significantly associated with the expression of MCPIP3 and IGF2BP3 in LUAD tissues (Figure S6D). Based on these results, we proposed a model to describe the potential antagonistic effects of the RBPs MCPIP3 and IGF2BP3 in regulating lung cancer cell metastasis (Figure 6I). MCPIP3, which is expressed at a relatively low level in human lung cancer cells, destabilizes METAP2 mRNA by binding to the stem‒loop structure in the 3’UTR via its RNase domain, thereby downregulating METAP2 gene expression and inhibiting lung cancer cell metastasis. Conversely, the high expression of IGF2BP3 in lung cancer cells stabilizes METAP2 mRNA by targeting the common stem‒loop structure, and thereby promoting METAP2 gene expression as well as lung cancer metastasis (Figure 6I). In summary, these findings strongly suggest that MCPIP3 and IGF2BP3 exert distinctly opposite effects on lung cancer metastasis progression by antagonistically regulating METAP2 gene expression. In the future, effectively inducing MCPIP3 while simultaneously inhibiting IGF2BP3 expression in lung cancer cells may represent a promising strategy for controlling lung cancer metastasis.
4. Discussion
To date, the expression patterns of RBPs in human lung cancer tissues remains unclear. This study revealed for the first time the expression of all the RBPs in LUAD and LUSC tissues. We found that the expression level of RBP MCPIP3 was significantly downregulated in both LUAD and LUSC, whereas IGF2BP3 expression was significantly increased. Both RBPs were found to influence the metastasis progression of human lung cancer. In contrast, MCPIP3 significantly inhibited lung cancer cell metastasis, whereas IGF2BP3 promoted cancer cell metastasis. Both of these RBPs were able to regulate the expression of metastasis-promoting genes in human lung cancer cells. To the best of our knowledge, few studies have reported the antagonistic regulatory effects of these two RBPs on the expression of tumor metastasis-related genes, especially in human lung cancer cells. Therefore, our findings revealed a reversible regulatory mechanism mediated by RBPs in controlling the metastasis of human lung cancer cells.
METAP2 is a well-known tumor metastasis promotor involved in regulating the metastasis progression of various human cancers and is considered a promising therapeutic target for cancers [36,37]. To date, few studies have reported the molecular mechanisms regulating METAP2 gene expression, especially in human lung cancer cells. In this study, we discovered that RBP MCPIP3 downregulates METAP2 expression by degrading its mRNA through its RNase domain. Our results revealed, for the first time, the post-transcriptional regulation mechanism of METAP2 gene expression in human lung cancer cells and further elucidated the downstream targets of the MCPIP protein family. We also revealed that MCPIP3 preferentially binds to metastasis-promoting mRNAs rather than metastasis-suppressing mRNAs. This preference may result from the long-term evolution of cancer cells; however, further investigations are required to clarify its precise underlying mechanism.
The RNA secondary stem‒loop structure located in the 3’UTR is believed to be an important cis-acting element that can be recognized and bound by RBPs to control transcripts stability [38,39]. It is therefore not surprising that MCPIP3 binds to the stem‒loop structure in the 3’UTR of METAP2 mRNA, as previous studies have shown that other family members of MCPIPs also recognize stem‒loop RNA [40]. Interestingly, this study further revealed that IGF2BP3, another RBP, was also capable of binding to the same structural RNA element but instead stabilized METAP2 mRNA and promoted its expression in human lung cancer cells. It has been reported that IGF2BP3 stabilizes mRNA [41] and promotes lung cancer metastasis [35], which is consistent with our findings. Previous data from our laboratory and other groups have shown that IGF2BP1, a homolog of IGF2BP3, can bind to structural RNA [42,43], further supporting our findings. In this study, our findings revealed a novel regulatory mechanism in which IGF2BP3 and MCPIP3 antagonistically regulate the stability and expression of the metastasis-promoting gene METAP2 transcript by targeting a common stem‒loop structure in human lung cancer cells. These results provide new insights into the understanding of the regulatory mechanisms controlling METAP2 gene expression in cancer cells.
Our findings suggest that lung cancer cells are ‘smart’ cells. Specifically, we found that MCPIP3 can inhibit lung cancer cell metastasis but that its expression is suppressed. However, the expression of IGF2BP3 is increased and promotes metastasis. One consequence of this is that lung cancer cells can increase the expression of the metastasis-promoting gene METAP2, ultimately facilitating lung cancer cell metastasis. These findings lead us to speculate that RBPs in human lung cancer cells may form a ‘yin’ and ‘yang’ regulatory mechanism, which antagonistically or synergistically regulates the expression of tumor-related genes in cancer cells. Further investigation is still necessary to elucidate the molecular mechanisms through which RBPs regulate cancer progression.
5. Conclusions
In conclusion, the results of this study reveal that the RBPs MCPIP3 and IGF2BP3 antagonistically regulate the mRNA stability and expression of the metastasis-promoting gene METAP2 by binding to a common stem‒loop structure, leading to reversible regulation of human lung cancer metastasis. These results provide new insights into the regulatory mechanisms of human lung cancer metastasis and identify potential molecular targets for the future treatment of human lung cancer metastasis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: MCPIP3 expression is reduced in human lung cancer tissues; Figure S2: MCPIP3 regulates the expression of metastasis-related genes in human lung cancer cells; Figure S3: MCPIP3 selectively downregulated the expression of the metastasis-promoting gene METAP2 by destabilizing its mRNA via the RNase domain; Figure S4: MCPIP2 depletion increased proangiogenic mRNAs stability and promoted tumor angiogenesis; Figure S5: IGF2BP3 interacts with the common stem‒loop structure to stabilize METAP2 mRNA and promote lung cancer metastasis; Figure S6: IGF2BP3 and MCPIP3 potentially antagonize metastasis-promoting genes expression and lung cancer cell metastasis; Table S1: PCR Primers and RNA-EMSA Probes Sequence; Table S2: PCRarray analysis of tumor metastasis-related genes regulated by MCPIP3; Table S3: RNA-seq analysis of genes regulated by MCPIP3 or IGF2BP3 in human lung cancer cells; Table S4: Identify of proteins interacting with METAP2 RNA stem‒loop structure in A549 cells by mass spectrometry.
Author Contributions
Conceptualization, W.L.; methodology, S.S., X. L. and H. L.; validation, A.L., R.X.; formal analysis, S.S.; investigation, S.S., X.L.; resources, W.L.; data curation, B. L., W.L.; writing—original draft preparation, S.S.; writing—review and editing, W.L.; visualization, B.L.; supervision, W.L.; project administration, W.L.; funding acquisition, W.L. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by the National Natural Science Foundation of China (NSFC) (Grant Number: 82372619), grant number 82372619.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and the human tumor samples used in this study and animal study protocol were approved by the Experimental Animal Care and Ethics Committee of the Institute of Microcirculation, CAMS & PUMC (Approval No.: WXH-LZ-2023-02; approval date is 07 March 2023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data supporting the findings of this study are included in the article and/or the supplementary materials. The RNA sequencing data from this study are deposited in the Sequence Read Archive (SRA) database under projects PRJNA1119197 and PRJNA1119237. The MS data are available via ProteomeXchange with the IDs: PXD052919 and PXD052920.
Acknowledgments
We thank Qingbin Wu and Xiaochen Yuan for technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R. L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74(3), 229–263. [Google Scholar] [CrossRef]
- Weng, M.; Feng, Y.; He, Y.; Yang, W.; Li, J.; Zhu, Y.; Wang, T.; Wang, C.; Zhang, X.; Qiao, Y.; Li, Q.; Zhao, L.; Gao, S.; Zhang, L.; Wu, Y.; Zhao, R.; Wang, G.; Li, Z.; Jin, X.; Zheng, T.; Li, X. Hypoxia-Induced LIN28A mRNA Promotes the Metastasis of Colon Cancer in a Protein-Coding-Independent Manner. Front Cell Dev. Biol. 2021, 9, 642930. [Google Scholar] [CrossRef]
- Jagadeeshan, S.; Prasad, M.; Badarni, M.; Ben-Lulu, T.; Liju, V. B.; Mathukkada, S.; Saunders, C.; Shnerb, A. B.; Zorea, J.; Yegodayev, K. M.; Wainer, M.; Vtorov, L.; Allon, I.; Cohen, O.; Gausdal, G.; Friedmann-Morvinski, D.; Cheong, S. C.; Ho, A. L.; Rosenberg, A. J.; Kessler, L.; Burrows, F.; Kong, D.; Grandis, J. R.; Gutkind, J. S.; Elkabets, M. Mutated HRAS Activates YAP1-AXL Signaling to Drive Metastasis of Head and Neck Cancer. Cancer Res. 2023, 83(7), 1031–1047. [Google Scholar] [CrossRef]
- Tang, T.; Yang, L.; Cao, Y.; Wang, M.; Zhang, S.; Gong, Z.; Xiong, F.; He, Y.; Zhou, Y.; Liao, Q.; Xiang, B.; Zhou, M.; Guo, C.; Li, X.; Li, Y.; Xiong, W.; Li, G.; Zeng, Z. LncRNA AATBC regulates Pinin to promote metastasis in nasopharyngeal carcinoma. Mol. Oncol. 2020, 14(9), 2251–2270. [Google Scholar] [CrossRef]
- Chen, M.; Dong, Y.; Tian, L.; Zhou, J.; Zhu, E.; Yuan, H.; Li, X.; Wang, B. Metastasis suppressor 1 interacts with protein tyrosine phosphatase receptor-delta to regulate adipogenesis. FASEB J. 2023, 37(4), e22857. [Google Scholar] [CrossRef] [PubMed]
- Chi, Q.; Xu, H.; Song, D.; Wang, Z.; Wang, Z.; Ma, G. alpha-E-Catenin (CTNNA1) Inhibits Cell Proliferation, Invasion and EMT of Bladder Cancer. Cancer Manag Res. 2020, 12, 12747–12758. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B. T.; Li, Q. L.; Lan, T.; Xie, J.; Lu, Y. G.; Zheng, D. L.; Su, B. H. CDH11 Regulates Adhesion and Transcellular Migration of Tongue Squamous Cell Carcinoma. Onco Targets Ther. 2021, 14, 4211–4222. [Google Scholar] [CrossRef]
- Lin, R. J.; Chien, H. L.; Lin, S. Y.; Chang, B. L.; Yu, H. P.; Tang, W. C.; Lin, Y. L. MCPIP1 ribonuclease exhibits broad-spectrum antiviral effects through viral RNA binding and degradation. Nucleic Acids Res. 2013, 41(5), 3314–26. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Roske, Y.; Yamada, S.; Uehata, T.; Takeuchi, O.; Heinemann, U. PIN and CCCH Zn-finger domains coordinate RNA targeting in ZC3H12 family endoribonucleases. Nucleic Acids Res. 2021, 49(9), 5369–5381. [Google Scholar] [CrossRef]
- Guo, G.; Fu, R.; Zhang, L.; Chao, T.; Jiang, Z.; Dong, L.; Huang, R.; Lu, L.; Yang, W.; Gu, Y.; Liang, Y.; Zheng, Q. Gfi1 and Zc3h12c orchestrate a negative feedback loop that inhibits NF-kB activation during inflammation in macrophages. Mol. Immunol. 2020, 128, 219–226. [Google Scholar] [CrossRef]
- von Gamm, M.; Schaub, A.; Jones, A. N.; Wolf, C.; Behrens, G.; Lichti, J.; Essig, K.; Macht, A.; Pircher, J.; Ehrlich, A.; Davari, K.; Chauhan, D.; Busch, B.; Wurst, W.; Feederle, R.; Feuchtinger, A.; Tschop, M. H.; Friedel, C. C.; Hauck, S. M.; Sattler, M.; Geerlof, A.; Hornung, V.; Heissmeyer, V.; Schulz, C.; Heikenwalder, M.; Glasmacher, E. Immune homeostasis and regulation of the interferon pathway require myeloid-derived Regnase-3. J. Exp. Med. 2019, 216(7), 1700–1723. [Google Scholar] [CrossRef]
- Wawro, M.; Kochan, J.; Sowinska, W.; Solecka, A.; Wawro, K.; Morytko, A.; Kwiecinska, P.; Grygier, B.; Kwitniewski, M.; Fu, M.; Cichy, J.; Kasza, A. Molecular Mechanisms of ZC3H12C/Reg-3 Biological Activity and Its Involvement in Psoriasis Pathology. Int. J. Mol. Sci. 2021, 22(14). [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huang, J.; Ashraf, A.; Rahaman, O.; Lou, J.; Wang, L.; Cai, P.; Wen, J.; Anwaar, S.; Liu, X.; Ni, H.; Ganguly, D.; Zhao, J.; Yang, C. Y. The RNase MCPIP3 promotes skin inflammation by orchestrating myeloid cytokine response. Nat. Commun. 2021, 12(1), 4105. [Google Scholar] [CrossRef]
- Li, W.; Gao, L. N.; Song, P. P.; You, C. G. Development and validation of a RNA binding protein-associated prognostic model for lung adenocarcinoma. Aging 2020, 12(4), 3558–3573. [Google Scholar] [CrossRef]
- Li, T.; Hui, W.; Halike, H.; Gao, F. RNA Binding Protein-Based Model for Prognostic Prediction of Colorectal Cancer. Technol. Cancer Res. Treat. 2021, 20, 15330338211019504. [Google Scholar] [CrossRef]
- Yue, Y.; Wu, Y.; Zhao, D.; Wu, B.; Wu, X.; Liu, J.; Yang, L.; Shen, A. Pan-Cancer Analysis Predicts the Immunological and Prognostic Role of ZC3H12C in KIRC. BioMed Res. Int. 2022, 2022, 4541571. [Google Scholar]
- Du, W.; Quan, X.; Wang, C.; Song, Q.; Mou, J.; Pei, D. Regulation of tumor metastasis and CD8(+) T cells infiltration by circRNF216/miR-576-5p/ZC3H12C axis in colorectal cancer. Cell Mol. Biol. Lett. 2024, 29(1), 19. [Google Scholar] [CrossRef] [PubMed]
- Suk, F. M.; Chang, C. C.; Lin, R. J.; Lin, S. Y.; Chen, Y. T.; Liang, Y. C. MCPIP3 as a Potential Metastasis Suppressor Gene in Human Colorectal Cancer. Int. J. Mol. Sci. 2018, 19(5). [Google Scholar] [CrossRef] [PubMed]
- He, W. P.; Guo, Y. Y.; Yang, G. P.; Lai, H. L.; Sun, T. T.; Zhang, Z. W.; Ouyang, L. L.; Zheng, Y.; Tian, L. M.; Li, X. H.; You, Z. S.; Xie, D.; Yang, G. F. CHD1L promotes EOC cell invasiveness and metastasis via the regulation of METAP2. Int. J. Med. Sci. 2020, 17(15), 2387–2395. [Google Scholar] [CrossRef]
- Lin, M.; Zhang, X.; Jia, B.; Guan, S. Suppression of glioblastoma growth and angiogenesis through molecular targeting of methionine aminopeptidase-2. J. Neurooncol 2018, 136(2), 243–254. [Google Scholar] [CrossRef]
- Shimizu, S.; Kawahara, R.; Simizu, S. Methionine aminopeptidase-2 is a pivotal regulator of vasculogenic mimicry. Oncol. Rep. 2022, 47(2). [Google Scholar] [CrossRef]
- Xie, J.; Rice, M. A.; Chen, Z.; Cheng, Y.; Hsu, E. C.; Chen, M.; Song, G.; Cui, L.; Zhou, K.; Castillo, J. B.; Zhang, C. A.; Shen, B.; Chin, F. T.; Kunder, C. A.; Brooks, J. D.; Stoyanova, T.; Rao, J. In Vivo Imaging of Methionine Aminopeptidase II for Prostate Cancer Risk Stratification. Cancer Res. 2021, 81(9), 2510–2521. [Google Scholar] [CrossRef]
- Takenaga, K.; Ochiya, T.; Endo, H. Inhibition of the invasion and metastasis of mammary carcinoma cells by NBD peptide targeting S100A4 via the suppression of the Sp1/MMP-14 axis. Int. J. Oncol. 2021, 58(3), 397–408. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wu, C.; He, S.; Chen, Y.; Zhang, Y.; Xu, H.; Ling, Q.; Zhang, Z.; Sun, T.; Yao, S.; Ren, Y.; Lai, H.; Yang, G. METAP2 inhibits K48-linked ubiquitination of YTHDF2 to promote ovarian cancer progression. Oncogene 2025, 44(46), 4505–4519. [Google Scholar] [CrossRef]
- Mita, M. M.; Mita, A. C.; Carver, B. J.; Shanahan, J. M.; Mayes, B. A.; Dufour, P. J.; Browning, D.; Anderson-Villaluz, A.; Petersen, J. S.; Turnquist, D. J.; Cornelius, P. A Phase 1 Safety Study of Evexomostat (SDX-7320) in Patients with Late-Stage Cancer: An Antiangiogenic, Insulin-Sensitizing Drug Conjugate Targeting METAP2. Cancer Res. Commun. 2025, 5(6), 1008–1017. [Google Scholar] [CrossRef]
- Friese-Hamim, M.; Ortiz Ruiz, M. J.; Bogatyrova, O.; Keil, M.; Rohdich, F.; Blume, B.; Leuthner, B.; Czauderna, F.; Hahn, D.; Jabs, J.; Jaehrling, F.; Heinrich, T.; Kellner, R.; Chan, K.; Tong, A. H. Y.; Wienke, D.; Moffat, J.; Blaukat, A.; Zenke, F. T. Novel Methionine Aminopeptidase 2 Inhibitor M8891 Synergizes with VEGF Receptor Inhibitors to Inhibit Tumor Growth of Renal Cell Carcinoma Models. Mol. Cancer Ther. 2024, 23(2), 159–173. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Hu, C.; He, Y.; Guo, L.; Sun, Y.; Han, C.; Zhang, X.; Ren, J.; Han, J.; Wang, J.; Zhang, J.; Sun, Y.; Cai, S.; Jiang, D.; Yang, K.; Yang, S. IGF2BP3 as a Novel Prognostic Biomarker and Therapeutic Target in Lung Adenocarcinoma. Cells 2025, 14(15). [Google Scholar] [CrossRef]
- Sebestyen, E.; Singh, B.; Minana, B.; Pages, A.; Mateo, F.; Pujana, M. A.; Valcarcel, J.; Eyras, E. Large-scale analysis of genome and transcriptome alterations in multiple tumors unveils novel cancer-relevant splicing networks. Genome Res. 2016, 26(6), 732–44. [Google Scholar] [CrossRef] [PubMed]
- Porter, D. F.; Miao, W.; Yang, X.; Goda, G. A.; Ji, A. L.; Donohue, L. K. H.; Aleman, M. M.; Dominguez, D.; Khavari, P. A. easyCLIP analysis of RNA-protein interactions incorporating absolute quantification. Nat. Commun. 2021, 12(1), 1569. [Google Scholar] [CrossRef]
- Guo, T.; Wang, Y.; Sun, X.; Hou, S.; Lan, Y.; Yuan, S.; Yang, S.; Zhao, F.; Chu, Y.; Ma, Y.; Cheng, T.; Yu, J.; Liu, B.; Yuan, W.; Wang, X. Loss of RNA-binding protein CELF2 promotes acute leukemia development via FAT10-mTORC1. Oncogene 2024, 43(19), 1476–1487. [Google Scholar] [CrossRef]
- Wu, C.; Ma, Y.; Li, J.; Hai, B.; Zhou, Y.; Cao, F.; Kang, X.; Xu, X. The molecular mechanism of IGF2BP3 promoting the malignant progression of lung cancer. Cancer Cell Int. 2026, 26(1). [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Chen, X.; Sun, Y.; Wang, Y. METTL1 interacts with XPO5 to modulate pre-miRNA export. Nucleic Acids Res. 2026, 54(3). [Google Scholar] [CrossRef]
- Jolma, A.; Zhang, J.; Mondragon, E.; Morgunova, E.; Kivioja, T.; Laverty, K. U.; Yin, Y.; Zhu, F.; Bourenkov, G.; Morris, Q.; Hughes, T. R.; Maher, L. J., 3rd; Taipale, J. Binding specificities of human RNA-binding proteins toward structured and linear RNA sequences. Genome Res. 2020, 30(7), 962–973. [Google Scholar] [CrossRef]
- Clayer, E.; Frank, D.; Anderton, H.; Zhang, S.; Kueh, A.; Heim, V.; Nutt, S. L.; Chopin, M.; Bouillet, P. ZC3H12C expression in dendritic cells is necessary to prevent lymphadenopathy of skin-draining lymph nodes. Immunol. Cell Biol. 2022, 100(3), 160–173. [Google Scholar] [CrossRef]
- Yang, X.; Bai, Q.; Chen, W.; Liang, J.; Wang, F.; Gu, W.; Liu, L.; Li, Q.; Chen, Z.; Zhou, A.; Long, J.; Tian, H.; Wu, J.; Ding, X.; Zhou, N.; Li, M.; Yang, Y.; Cai, J. m(6) A-Dependent Modulation via IGF2BP3/MCM5/Notch Axis Promotes Partial EMT and LUAD Metastasis. Adv. Sci. (Weinh) 2023, 10(20), e2206744. [Google Scholar] [CrossRef]
- Fieni, C.; Ciummo, S. L.; Sorrentino, C.; Marchetti, S.; Vespa, S.; Lanuti, P.; Lotti, L. V.; Di Carlo, E. Prevention of prostate cancer metastasis by a CRISPR-delivering nanoplatform for interleukin-30 genome editing. Mol. Ther. 2024, 32(11), 3932–3954. [Google Scholar] [CrossRef]
- Goya Grocin, A.; Kallemeijn, W. W.; Tate, E. W. Targeting methionine aminopeptidase 2 in cancer, obesity, and autoimmunity. Trends Pharmacol. Sci. 2021, 42(10), 870–882. [Google Scholar] [CrossRef]
- Tants, J. N.; Walbrun, A.; Kollwitz, L.; Friedrich, K.; Rief, M.; Schlundt, A. Roquin exhibits opposing effects on RNA stem-loop stability through its two ROQ domain binding sites. Proc. Natl. Acad. Sci. U S A 2025, 122(15), e2424434122. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Zhang, D.; Liu, X.; Lu, W. Arid4a Suppresses Breast Tumor Metastasis by Enhancing MTSS1 Expression via mRNA Stability. Cancer Med. 2025, 14(5), e70732. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Fan, L.; Jackson, S.; Matunis, A. R.; Lou, D.; Chen, K.; Trevejo-Nunez, G. Lung Epithelial Regnase-1 Dampens Local Immune Response but Does Not Worsen Susceptibility to Klebsiella pneumoniae. Immunohorizons 2024, 8(1), 89–96. [Google Scholar] [CrossRef]
- Wang, H.; Cao, Y.; Gou, Y.; Wang, H.; Liang, Z.; Wu, Q.; Tan, J.; Liu, J.; Li, Z.; Cui, J.; Zhang, H.; Zhang, Z. IGF2BP3 promotes glutamine metabolism of endometriosis by interacting with UCA1 to enhances the mRNA stability of GLS1. Mol. Med. 2024, 30(1), 64. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Li, H.; Liu, X.; Li, A.; Xiu, R. The RNA-Binding Proteins MCPIP2 and IGF2BP1 Competitively Modulate Breast Tumor Angiogenesis by Antagonizing VEGFA mRNA Stability and Expression. FASEB J. 2025, 39(10), e70594. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Kumar, A.; Kumari, P.; Rasool, K.; Maiti, S.; Jain, N. Thermodynamics and Binding Mechanism of Insulin-like Growth Factor Binding Protein 1 (IGF2BP1) and Y3 RNA Interaction. Biochemistry 2025, 64(16), 3535–3548. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
MCPIP3 expression is reduced in human lung cancer tissues and is associated with poor survival. (A) The expression of 1,756 RNA-binding protein genes in human LUAD was profiled based on the TCGA database. (B) Expression of MCPIP3 in human LUAD and normal tissues was analyzed using UALCAN online tool (https://ualcan.path.uab.edu/analysis-prot.html). (C) MCPIP3 expression was tested by qRT-PCR in human LUAD specimens (n = 10) and their surrounding “normal” tissues (n = 10). (D) MCPIP3 protein expression in human LUAD and normal tissues was showed by the CPTAC database (http://ualcan.path.uab.edu/analysis-prot.html). (E) MCPIP3 protein expression in several LUAD tissues and their paired normal tissues was displayed by western blot. (F) Representative images of IHC staining for MCPIP3 in human normal lung tissues and LUAD tissues (left). Scale bar, 100 μm. MCPIP3 IHC staining scores of 6 pairs of matched LUAD and normal tissues (right). (G) MCPIP3 transcript expression levels were reduced with the nodal metastasis status in LUAD patients (http://ualcan.path.uab.edu/analysis-prot.html). (H, I) MCPIP3 transcript (H) and protein (I) expression was analyzed by the main pathological stages of LUAD. (J) Kaplan–Meier relapse-free survival curves of lung cancer patients grouped according to MCPIP3 expression levels (https://kmplot.com/analysis/). (K) Kaplan–Meier relapse-free survival (RFS) and overall survival (OS) curves of LUAD patients with low and high tumor MCPIP3 expression in the GSE31210 and GSE3141 datasets, respectively.
Figure 1.
MCPIP3 expression is reduced in human lung cancer tissues and is associated with poor survival. (A) The expression of 1,756 RNA-binding protein genes in human LUAD was profiled based on the TCGA database. (B) Expression of MCPIP3 in human LUAD and normal tissues was analyzed using UALCAN online tool (https://ualcan.path.uab.edu/analysis-prot.html). (C) MCPIP3 expression was tested by qRT-PCR in human LUAD specimens (n = 10) and their surrounding “normal” tissues (n = 10). (D) MCPIP3 protein expression in human LUAD and normal tissues was showed by the CPTAC database (http://ualcan.path.uab.edu/analysis-prot.html). (E) MCPIP3 protein expression in several LUAD tissues and their paired normal tissues was displayed by western blot. (F) Representative images of IHC staining for MCPIP3 in human normal lung tissues and LUAD tissues (left). Scale bar, 100 μm. MCPIP3 IHC staining scores of 6 pairs of matched LUAD and normal tissues (right). (G) MCPIP3 transcript expression levels were reduced with the nodal metastasis status in LUAD patients (http://ualcan.path.uab.edu/analysis-prot.html). (H, I) MCPIP3 transcript (H) and protein (I) expression was analyzed by the main pathological stages of LUAD. (J) Kaplan–Meier relapse-free survival curves of lung cancer patients grouped according to MCPIP3 expression levels (https://kmplot.com/analysis/). (K) Kaplan–Meier relapse-free survival (RFS) and overall survival (OS) curves of LUAD patients with low and high tumor MCPIP3 expression in the GSE31210 and GSE3141 datasets, respectively.

Figure 2.
MCPIP3 inhibits lung cancer cell metastasis by mediating the expression of tumor metastasis-related genes. (A) Representative images (up) and analysis results (down) of wound healing assays with MCPIP3-overexpressing A549 cells, MCPIP3-overexpressing NCI-H23 cells and their control groups. (B) Representative images (up) and analysis (down) of Transwell invasion assays with MCPIP3-overexpressing A549 cells, MCPIP3-overexpressing NCI-H23 cells and their control groups. (C) A549 tumor growth curves in nude mice after overexpressing MCPIP3. (D) Picture of A549 tumors expressing MCPIP3 or control vector. (E) Representative HE staining images of lung tissues from nude mice bearing A549/MCPIP3 or A549/Control vector tumors. Scale bar, 100 μm. Right panel showed the quantification of metastatic nodules. (F) GO (Gene Ontology) analysis of all regulated genes by MCPIP3 by DAVID online tool (https://davidbioinformatics.nih.gov/), and several cell motility-related terms were indicated by red arrows. (G) RNA-seq data showed that multiple metastasis-promoting genes were commonly downregulated, while the metastasis-inhibiting genes were commonly upregulated in MCPIP3-overexpressing A549 and NCI-H23 cells. (H) PCR Array data showing the metastasis promoters were downregulated, and the metastasis suppressors were upregulated in MCPIP3-overexpressing A549 cells. (I) qPCR data showing the mRNA expression levels of metastasis-promoting genes (METAP2, HRAS, PNN, CTBP1) in MCPIP3-overexpressing A549 and NCI-H23 cells.
Figure 2.
MCPIP3 inhibits lung cancer cell metastasis by mediating the expression of tumor metastasis-related genes. (A) Representative images (up) and analysis results (down) of wound healing assays with MCPIP3-overexpressing A549 cells, MCPIP3-overexpressing NCI-H23 cells and their control groups. (B) Representative images (up) and analysis (down) of Transwell invasion assays with MCPIP3-overexpressing A549 cells, MCPIP3-overexpressing NCI-H23 cells and their control groups. (C) A549 tumor growth curves in nude mice after overexpressing MCPIP3. (D) Picture of A549 tumors expressing MCPIP3 or control vector. (E) Representative HE staining images of lung tissues from nude mice bearing A549/MCPIP3 or A549/Control vector tumors. Scale bar, 100 μm. Right panel showed the quantification of metastatic nodules. (F) GO (Gene Ontology) analysis of all regulated genes by MCPIP3 by DAVID online tool (https://davidbioinformatics.nih.gov/), and several cell motility-related terms were indicated by red arrows. (G) RNA-seq data showed that multiple metastasis-promoting genes were commonly downregulated, while the metastasis-inhibiting genes were commonly upregulated in MCPIP3-overexpressing A549 and NCI-H23 cells. (H) PCR Array data showing the metastasis promoters were downregulated, and the metastasis suppressors were upregulated in MCPIP3-overexpressing A549 cells. (I) qPCR data showing the mRNA expression levels of metastasis-promoting genes (METAP2, HRAS, PNN, CTBP1) in MCPIP3-overexpressing A549 and NCI-H23 cells.

Figure 3.
MCPIP3 selectively downregulated the expression of the metastasis-promoting gene METAP2 by destabilizing its mRNA via the RNase domain. (A) RNA-IP with specific anti-GFP antibody or isotype IgG followed by RT-PCR detection showed that the metastasis-suppressing genes were precipitated by MCPIP3. (B) Luciferase reporter assay analysis showing the effects of MCPIP3 overexpression on the luciferase activities of the indicated reporters. (C) Mutation strategy of METAP2 stem–loop structure (asterisks indicate base substitution). The stem–loop structure was deleted in Mut1 (middle) and Mut2 still kept the stem–loop structure (right). (D) The relative luciferase activities of the indicated reporters were determined by reporter assay. (E) RNA-EMSA showing the unique RNA-protein complex (RPC) formed using METAP2 stem–loop probes and whole-cell lysates (WCL) extracted from A549/MCPIP3-GFP cells. (F) RNA-ChIP assay was performed with A549/MCPIP3-GFP cells. (G) Half-life of the METAP2 mRNA was reduced by MCPIP3 in A549 cells. (H) METAP2 and GAPDH protein expression were measured by western blotting in MCPIP3-overexpressing A549 and NCI-H23 cells, respectively. (I) Schematic representation of MCPIP3 truncation mutants and its domains. ZF: zinc finger. (J) The mRNA expression of four metastasis-promoting genes was measured by qPCR after overexpressing the truncations in A549 cells. (K) Half-lives of METAP2 mRNA were measured after overexpression of wild type (WT) MCPIP3 or its mutants in A549 cells. (L) Kaplan-Meier overall survival curves of LUAD patients grouped based on METAP2 mRNA expression (https://kmplot.com/analysis).
Figure 3.
MCPIP3 selectively downregulated the expression of the metastasis-promoting gene METAP2 by destabilizing its mRNA via the RNase domain. (A) RNA-IP with specific anti-GFP antibody or isotype IgG followed by RT-PCR detection showed that the metastasis-suppressing genes were precipitated by MCPIP3. (B) Luciferase reporter assay analysis showing the effects of MCPIP3 overexpression on the luciferase activities of the indicated reporters. (C) Mutation strategy of METAP2 stem–loop structure (asterisks indicate base substitution). The stem–loop structure was deleted in Mut1 (middle) and Mut2 still kept the stem–loop structure (right). (D) The relative luciferase activities of the indicated reporters were determined by reporter assay. (E) RNA-EMSA showing the unique RNA-protein complex (RPC) formed using METAP2 stem–loop probes and whole-cell lysates (WCL) extracted from A549/MCPIP3-GFP cells. (F) RNA-ChIP assay was performed with A549/MCPIP3-GFP cells. (G) Half-life of the METAP2 mRNA was reduced by MCPIP3 in A549 cells. (H) METAP2 and GAPDH protein expression were measured by western blotting in MCPIP3-overexpressing A549 and NCI-H23 cells, respectively. (I) Schematic representation of MCPIP3 truncation mutants and its domains. ZF: zinc finger. (J) The mRNA expression of four metastasis-promoting genes was measured by qPCR after overexpressing the truncations in A549 cells. (K) Half-lives of METAP2 mRNA were measured after overexpression of wild type (WT) MCPIP3 or its mutants in A549 cells. (L) Kaplan-Meier overall survival curves of LUAD patients grouped based on METAP2 mRNA expression (https://kmplot.com/analysis).

Figure 4.
Knockdown of MCPIP3 increases the stability of METAP2 mRNA and promotes lung tumor metastasis progression. (A) Knockdown of MCPIP3 protein expression was confirmed by western blotting in A549 and NCI-H23 cells, respectively. Protein expression quantification was assessed by ImageJ. (B) Representative images (up) and analysis (down) of lung cancer cell migration after MCPIP3 knockdown. (C) mRNA expression of metastasis-promoting genes was measured by qPCR in MCPIP3-silencing A549 cells. (D) Half-life of METAP2 mRNA was measured in MCPIP3-knocking down A549 cells. (E) mRNA expression of METAP2 mRNA was measured by qPCR in MCPIP3-silencing A549 cells. (F) Quantification of the cell migration abilities after co-knockdown of MCPIP3 and METAP2 in A549 cells. (G) The volumes of MCPIP3-silencing A549 xenograft tumors. H Images of MCPIP3 silenced A549 tumors are shown. (I) H&E staining of lung tissues from nude mice bearing A549/shMCPIP3 or scramble control tumors. Scale bar, 100 μm. Right: quantification of metastatic nodules was shown.
Figure 4.
Knockdown of MCPIP3 increases the stability of METAP2 mRNA and promotes lung tumor metastasis progression. (A) Knockdown of MCPIP3 protein expression was confirmed by western blotting in A549 and NCI-H23 cells, respectively. Protein expression quantification was assessed by ImageJ. (B) Representative images (up) and analysis (down) of lung cancer cell migration after MCPIP3 knockdown. (C) mRNA expression of metastasis-promoting genes was measured by qPCR in MCPIP3-silencing A549 cells. (D) Half-life of METAP2 mRNA was measured in MCPIP3-knocking down A549 cells. (E) mRNA expression of METAP2 mRNA was measured by qPCR in MCPIP3-silencing A549 cells. (F) Quantification of the cell migration abilities after co-knockdown of MCPIP3 and METAP2 in A549 cells. (G) The volumes of MCPIP3-silencing A549 xenograft tumors. H Images of MCPIP3 silenced A549 tumors are shown. (I) H&E staining of lung tissues from nude mice bearing A549/shMCPIP3 or scramble control tumors. Scale bar, 100 μm. Right: quantification of metastatic nodules was shown.

Figure 5.
The RNA-binding protein IGF2BP3 interacts with the common stem‒loop structure to stabilize METAP2 mRNA and promote lung cancer metastasis. (A) Schematic representation of METAP2 stem‒loop structure in the 3’UTRs among different species. (B) Silver staining of proteins precipitated by the human METAP2 stem–loop structure in A549 cells. (C) Top 10 proteins identified by mass spectrometry were listed. (D) RNA pull-down followed by immunoblotting analysis showing that IGF2BP3 protein was detected in the precipitants pulled down with METAP2 wild type (WT) stem–loop probe in A549 cells. (E) RIP-ChIP analysis of IGF2BP3 bound to the METAP2 stem–loop sequence in A549/IGF2BP3-GFP cells. (F) GO analysis of all regulated genes by IGF2BP3 in A549 cells. The cell motility-related terms were indicated by red arrows. (G) RNA-seq data showing the expression of the indicated metastasis-promoting genes after IGF2BP3 overexpression in A549 cells. (H) The mRNA expression of the indicated metastasis-promoting genes was measured by qPCR in IGF2BP3-overexpressing A549 cells. (I) Protein expression of METAP2 in IGF2BP3-overexpressing A549 and NCI-H23 cells as measured by western blotting. (J) Half-lives analysis of METAP2 mRNA after MCPIP3 and IGF2BP3 co-overexpression in A549 cells. (K) Representative images and analysis of cell migration after IGF2BP3 overexpression in A549 cells.
Figure 5.
The RNA-binding protein IGF2BP3 interacts with the common stem‒loop structure to stabilize METAP2 mRNA and promote lung cancer metastasis. (A) Schematic representation of METAP2 stem‒loop structure in the 3’UTRs among different species. (B) Silver staining of proteins precipitated by the human METAP2 stem–loop structure in A549 cells. (C) Top 10 proteins identified by mass spectrometry were listed. (D) RNA pull-down followed by immunoblotting analysis showing that IGF2BP3 protein was detected in the precipitants pulled down with METAP2 wild type (WT) stem–loop probe in A549 cells. (E) RIP-ChIP analysis of IGF2BP3 bound to the METAP2 stem–loop sequence in A549/IGF2BP3-GFP cells. (F) GO analysis of all regulated genes by IGF2BP3 in A549 cells. The cell motility-related terms were indicated by red arrows. (G) RNA-seq data showing the expression of the indicated metastasis-promoting genes after IGF2BP3 overexpression in A549 cells. (H) The mRNA expression of the indicated metastasis-promoting genes was measured by qPCR in IGF2BP3-overexpressing A549 cells. (I) Protein expression of METAP2 in IGF2BP3-overexpressing A549 and NCI-H23 cells as measured by western blotting. (J) Half-lives analysis of METAP2 mRNA after MCPIP3 and IGF2BP3 co-overexpression in A549 cells. (K) Representative images and analysis of cell migration after IGF2BP3 overexpression in A549 cells.

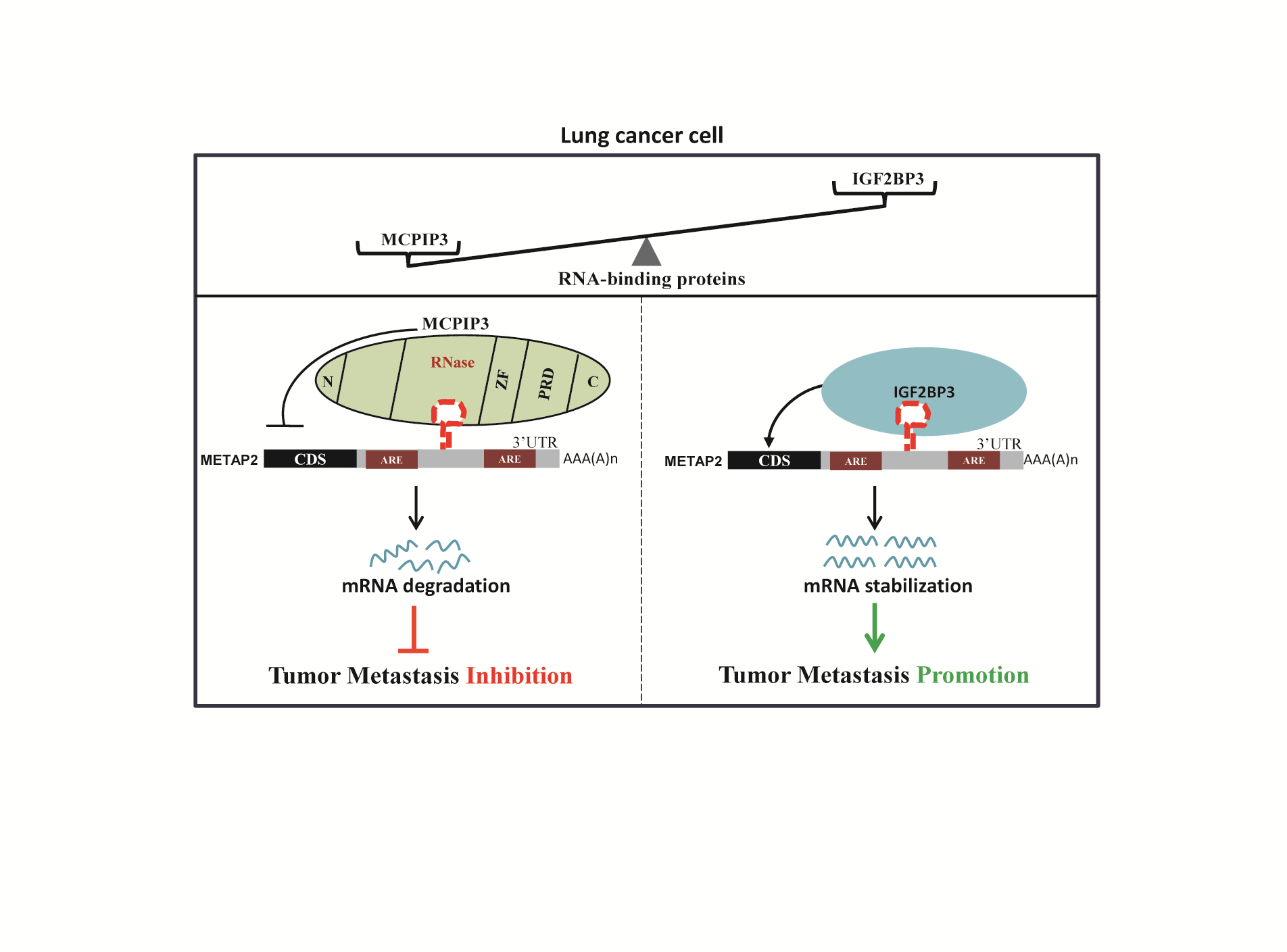
Figure 6.
IGF2BP3 and MCPIP3 potentially antagonize lung cancer cell metastasis by reversibly modulating METAP2 expression. (A) mRNA expression of IGF2BP3 in LUAD and normal lung tissues were analyzed by UALCAN online tool. (B) Protein expression of IGF2BP3 in human LUAD and normal tissues was displayed by UALCAN. (C) IGF2BP3 mRNA expression was increased with the nodal metastasis status in LUAD patients. (D) Kaplan-Meier overall survival curves of LUAD patients grouped based on IGF2BP3 mRNA expression level (https://ualcan.path.uab.edu/index.html). (E) H&E staining of lung tissues from nude mice bearing A549/IGF2BP3 tumors or A549/control vector tumors, respectively. Scale bar, 200 μm. Quantification of metastatic nodules was shown in the right panel. (F) The mRNA expression of METAP2 were measured by qRT-PCR in IGF2BP3-overexpressing and MCPIP3-overexpressing xenograft tumors and their controls, respectively. (G) Pearson’s correlation analysis between MCPIP3, IGF2BP3, and METAP2 expression in human LUAD patients (http://www.oncolnc.org/). (H) Representative images of IHC staining for MCPIP3 (left, up), IGF2BP3 (right, up), and METAP2 (lower) in human LUAD tissues. Scale bars, 50 μm. (I) A proposed model showing the underlying mechanism by which MCPIP3 and IGF2BP3 regulate the metastatic potential of lung cancer cells.
Figure 6.
IGF2BP3 and MCPIP3 potentially antagonize lung cancer cell metastasis by reversibly modulating METAP2 expression. (A) mRNA expression of IGF2BP3 in LUAD and normal lung tissues were analyzed by UALCAN online tool. (B) Protein expression of IGF2BP3 in human LUAD and normal tissues was displayed by UALCAN. (C) IGF2BP3 mRNA expression was increased with the nodal metastasis status in LUAD patients. (D) Kaplan-Meier overall survival curves of LUAD patients grouped based on IGF2BP3 mRNA expression level (https://ualcan.path.uab.edu/index.html). (E) H&E staining of lung tissues from nude mice bearing A549/IGF2BP3 tumors or A549/control vector tumors, respectively. Scale bar, 200 μm. Quantification of metastatic nodules was shown in the right panel. (F) The mRNA expression of METAP2 were measured by qRT-PCR in IGF2BP3-overexpressing and MCPIP3-overexpressing xenograft tumors and their controls, respectively. (G) Pearson’s correlation analysis between MCPIP3, IGF2BP3, and METAP2 expression in human LUAD patients (http://www.oncolnc.org/). (H) Representative images of IHC staining for MCPIP3 (left, up), IGF2BP3 (right, up), and METAP2 (lower) in human LUAD tissues. Scale bars, 50 μm. (I) A proposed model showing the underlying mechanism by which MCPIP3 and IGF2BP3 regulate the metastatic potential of lung cancer cells.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



