Submitted:
05 June 2026
Posted:
08 June 2026
You are already at the latest version
Abstract
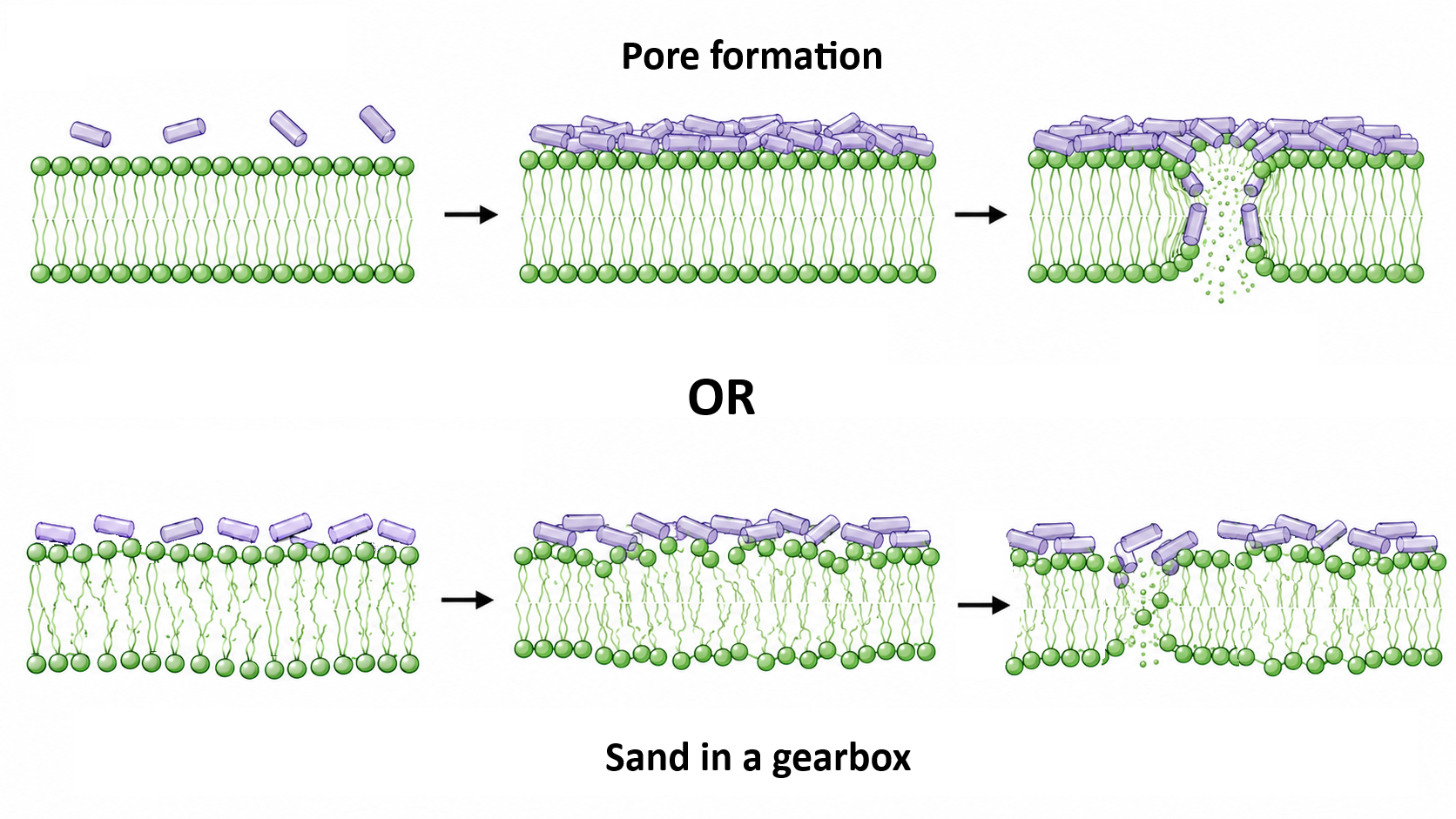
Antimicrobial peptides are promising agents for combating resistant infections. They exhibit bactericidal activity against a wide range of microbes, primarily by disrupting the permeability of the bacterial membrane and ultimately causing cell death. Effective bacterial killing requires a high number of membrane-bound peptide molecules. Therefore, it is conceivable that peptide accumulation on the membrane could also interfere with essential cellular processes by altering bilayer dynamics, a hypothesis referred to as the “sand in the gearbox” model. In this work, we systematically investigated how membrane dynamics is affected by a set of well-characterized yet highly diverse peptides: the natural AMP magainin 2, the toxin melittin, the synthetic peptides LAH4 and Killer-FLIP, and small membrane-active peptidomimetics with bactericidal activity. These effects were examined using fluorescence spectroscopy techniques, by measuring anisotropy, generalized polarization, and excimer formation of specific probes inserted at different depths within the lipid bilayer. Our results show that the activity of all compounds extends beyond membrane permeabilization, and that perturbation of membrane dynamics is a common feature among all systems analyzed. The membrane-active compounds induced a stiffening of the phospholipid bilayer by reducing lipid lateral mobility and decreasing water penetration, at least on the nanosecond timescale accessible to fluorescence measurements. Interestingly, when accounting for differences in the resulting membrane surface area coverage, the concentration range in which this behavior occurred was the same for all compounds studied. This threshold is, generally, higher than that required for membrane permeabilization and reflects near-complete coverage of the bilayer surface.
Keywords:
antimicrobial peptides
; membrane dynamics
; fluorescence spectroscopy
; peptidomimetics
1. Introduction
The emergence and global dissemination of drug-resistant pathogens have elevated antimicrobial resistance (AMR) to one of the most pressing threats to public health worldwide (https://data.who.int/dashboards/amr/overview). Antimicrobial peptides (AMPs) possess several remarkable features that make them highly promising candidates to combat the global spread of antibiotic-resistant “superbugs.” These include their rapid killing of pathogens, their generally nonspecific mechanism of action involving membrane permeabilization, which makes the development of resistance highly unlikely, and, generally, their inherent selectivity toward microbial cells [1,2].
AMPs are extremely heterogeneous in terms of secondary structure, length, and amino acid composition, and no conserved sequence motif can be identified. In the present work, we focus on linear, cationic antimicrobial peptide, which share several common physicochemical features. AMPs of this family consist of relatively short peptide sequences (typically 10–50 amino acids) enriched in basic and hydrophobic residues. Although diverse secondary structures have been reported, they are generally unstructured in aqueous solution and often fold into α-helical conformations upon interaction with phospholipid membranes [3,4,5,6], conferring an amphipathic character. The presence of basic amino acids gives AMPs an overall positive charge, that plays an essential role in driving the initial electrostatic attraction to negatively charged bacterial membranes and contributes to their selectivity toward microbial cells over zwitterionic mammalian cells [2].
It is widely accepted that the primary antimicrobial mechanism of cationic AMPs involves perturbation of the bacterial membrane, leading to dissipation of electrochemical potential, disruption of lipid asymmetry, and leakage of essential metabolites and cellular components. However, additional mechanisms may contribute to bacterial killing. Increasing evidence suggests that AMPs can also modulate the host immune response by activating leukocytes, enhancing the recruitment of immune cells (such as neutrophils, monocytes, and T lymphocytes) to the site of infection, and regulating processes including angiogenesis, apoptosis, and wound healing [7,8,9]. A hypothesis proposed by Sahl and co-workers, known as the “sand in the gearbox” model [10], suggests that rather than killing bacteria solely through pore formation, AMPs may interfere with the coordinated and highly dynamic function of membrane-bound multienzyme complexes. The centrality of the bacterial membrane to various physiological and metabolic functions of the bacterial cell, and the significantly detrimental effects that AMPs have on the structural and functional integrity of the bacterial membrane, further support the possibility that AMPs may act through such a mechanism.
Previously, we demonstrated that killing requires complete peptide coverage of the cell surface, both in Gram-negative and Gram-positive bacteria [11,12]. In parallel, using fluorescence spectroscopy, we recently showed that the activity of two natural AMPs, alamethicin and PMAP-23, is not limited to membrane permeabilization but also involves modifications in bilayer dynamics, causing a decrease of water penetration, and lipid lateral diffusion. Remarkably, these effects are not restricted to model vesicles but also occur in live bacterial cells [13].
Beyond the description of the behavior of specific AMPs, due to the intrinsic diversity of this class of molecules, it is of pivotal importance to understand whether, or to what extent, this behavior can be considered a general feature of AMPs and AMP-mimicking compounds. To this end, in this study we systematically investigated the effects of several membrane- active compounds on membrane dynamics.
First, we chose three well-characterized yet structurally diverse antimicrobial peptides: the natural AMP magainin 2, the toxin melittin, and the synthetic peptide LAH4. Magainin 2 was one of the first AMPs to be identified. It is a 23-residue cationic peptide isolated from the skin of the African frog Xenopus laevis, exhibiting strong antimicrobial and antitumor activity [14,15,16,17]. It permeabilizes negatively charged membranes by forming an amphipathic helix and creates pores via a “carpet/toroidal” mechanism.
Melittin is a 26-residue, strongly amphiphilic peptide, isolated from the European bee Apis mellifera venom, which is known for its strong hemolytic and antimicrobial activities [18,19,20]. Biophysical studies indicate that melittin exerts its antimicrobial activity via the “carpet” mechanism [21,22].
LAH4 is a synthetic peptide with both pore-forming and cell-penetrating abilities. Composed of leucines, alanines, four histidines, and terminal lysine pairs, it can have different charge and membrane orientation, based on pH changes [23,24]. Its helix lies parallel to membranes at acidic pH and becomes transmembrane at neutral or basic pH [23,25]. The peptide’s orientation is also affected by factors such as counter-ions and lipid composition [26]. Notably, a study found that in phosphatidylglycerol-bilayers, its surface-alignment is stabilized [27].
We extended this investigation to other membrane-active compounds, including the peptide Killer-FLIP [28], which exhibits remarkably potent cytotoxicity across a variety of cancer cell lines and inhibits tumor growth in vivo. The peptide was originally designed to interfere with apoptosis signaling [29], but it was demonstrated that its activity was independent of apoptosis and was accompanied by loss of membrane integrity [29].
Increasing research efforts are focused on developing synthetic molecules that emulate both the antibacterial activity and the underlying mechanisms of AMPs while circumventing some of their intrinsic limitations. Among these, we focused our attention on a class of peptidomimetics, for which it has been demonstrated that they act by disrupting the bacterial membranes, featuring positive charges and a pendant aliphatic group around a norspermidine residue (Figure 1). These compounds demonstrate strong activity against both Gram-positive and Gram-negative bacteria, including drug-resistant strains, with MIC values in the micromolar range [30]. While these molecules embody some of the physicochemical characteristics of AMPs, such as cationic charge and hydrophobicity, they differ significantly in terms of size, molecular weight and nature of chemical backbone. In this case, we aim to understand if, also under this perspective, the features of these molecules overlap those of the AMPs. The effects of all membrane-active compounds were investigated using fluorescence spectroscopy, a powerful experimental technique that allows to follow dynamic phenomena occurring on the nanosecond timescale.

Table 1.
amino-acidic sequence of the investigated peptides.
| Peptide | Sequence1 |
| Magainin 2 | GIGKFLHSAKKFGKAFVGEIMNS |
| Melittin | GIGAVLKVLTTGLPALISWIKRKRQQ-NH2 |
| LAH4 | KKALLALALHHLAHLALHLALALKKA-NH2 |
| Killer-FLIP | GRKKRRQRRRFFWSLCTA |
1 Hydrophobic, basic, His, and other residues are reported in green, blue, orange and black, respectively.
2. Results
2.1. Perturbation of Membrane Permeability
Beyond the affinity for the lipid bilayers, the activity of a membrane-permeabilizing molecules mainly depends on their abilities to insert and to form pores [31]. To determine the range of peptide concentrations that induces membrane permeability and to relate this to other possible membrane effects, we measured peptide-induced pore formation. The permeability of the bilayer induced by melittin and magainin 2 was studied by entrapping the fluorescent probe carboxyfluorescein (CF) inside lipid vesicles at a concentration high enough to induce quenching of the dye by self-association [32,33].
A fixed amount of vesicles (lipid concentration 50 µM) was titrated with increasing concentrations of peptide and the release kinetics of the probe was recorded.
We tested LAH4 activity at both pH 7.4 and 5.0 to study the effect of peptide charge on its behavior by entrapping inside the lipid vesicles calcein rather than CF, because at acidic pH the latter fluorophore is released spontaneously very quickly [34], while calcein does not suffer of this limitation [35]. We verified that peptide-induced leakage at pH 7.4 is not affected significantly by the choice of the particular dye used (Figure S1).
The membrane permeability assay provided that LAH4 exhibits the highest membrane-perturbation activity, followed by melittin and magainin 2, whose pore-forming abilities are reduced by a factor of ten relative to LAH4 (Figure 2). In addition, the membrane-perturbing behavior of LAH4 is the same at both pH’s as previously reported, as well as its affinity for negatively charged membranes except for slight variations [35]. Therefore, we can speculate that peptide activity is not influenced by its orientation on membrane, at least in POPC/POPG liposomes [27,35]. This allows to evaluate the peptide-induced effects on the bilayer without any additional complication caused by a possible variation in peptide orientation or pore forming activity.
2.2. Perturbation of Membrane Fluidity and Order: Steady-State Anisotropy
The effects of the three peptides on membrane fluidity (local lipid viscosity and packing of membrane) were assessed by steady-state fluorescence anisotropy. This spectroscopic technique allows to determine the speed and amplitude of the rotational motions of a fluorophore during its excited state lifetime. Slow (compared to the lifetime) and/or limited motions correspond to high anisotropy values (with the maximum, limiting value being 0.4), while fast and wide rotations yield anisotropy values close to zero [36].
We employed three different fluorescent probes that are located at different depths into the lipid bilayer: DPH, NBD-PE and Laurdan [37,38,39]. NBD-PE is the most superficial because the fluorophore is covalently linked to the phospholipid head group of POPE and it is located at a distance of about 19-20 Ǻ from the bilayer center [37,40,41]. Particularly, depth-depending quenching (DDQ) data indicate that the probe explores a broad distribution of positions, with a maximum at around 15 Å from the bilayer center. Molecular dynamics (MD) simulations support this observation [42], suggesting a possible downward bending of the phosphoethanolamine group, with NBD inserting slightly below the headgroups [43]. For NBD, it is well known that homo-FRET can occur when two molecules are within the Förster radius [44,45], leading to a lowering of the anisotropy; data in Figure 3 clearly show that an opposite effect is observed in our case. To further rule out this possibility, we repeated the anisotropy experiments with the membrane labeling reduced to 0.1% NBD-PE for the AMP melittin (from the 1% reported in Figure 3). The results were fully comparable, allowing us to exclude any artifacts related to homo-FRET (Figure S4). Different from NBD-PE, the hydrophobicity of DPH places the probe much deeper within the bilayer, aligned parallel to the phospholipid hydrocarbon chains. Several approaches have been employed to estimate its location: DDQ experiments reported an average depth of 7.8 Å from the center of a PC membrane [39], a result consistent with MD simulations [46,47,48], which nevertheless reveal a distribution of sites within the hydrophobic core. Neutron scattering experiments [49] further suggest that a fraction of DPH molecules may localize closer to the membrane surface, though this population was not quantified. Laurdan, in contrast, resides at an intermediate depth, just below the phospholipid headgroups. DDQ experiments place its average position at 10–11 Å from the bilayer center [50,51], in agreement with molecular dynamics (MD) simulations, which likewise reveal a distribution of positions within the bilayer [52,53].
All peptides cause an increase in the probe fluorescence anisotropy (Figure 3), slightly higher for LAH4 at neutral pH, probably for the peculiar trans-membrane configuration assumed by this peptide. Overall, these trends are indicative of a decreased mobility of the fluorophores during the excited state lifetime. The anisotropy trend is similar across the three probes, regardless of their position within the membrane, suggesting that the peptide exerted a global effect on the bilayer.
The reduction in the mobility of the probes starts in the same concentration range as the pore-forming activity of the peptide, but the main effect occurs once the AMP activity is completed.
2.3. Perturbation of Membrane Fluidity and Order: Time-Resolved Anisotropy
Increased steady-state anisotropy can be caused by a reduction in the amplitude and/or rate of the probe motions, by a shortening of the fluorescence lifetime or by both variations [54]. To discriminate among these contributions, we performed time-resolved fluorescence intensity and anisotropy experiments both with and without peptides, where an AMP concentration able to cause a leakage close to 100% was used.
Representative data for fluorescence intensity decay and time-resolved anisotropy, respectively, are reported for liposomes labelled with NBD-PE in the presence of magainin 2 (Figure 4) at the highest concentration tested in steady state anisotropy measurements; similar results were obtained for all the other peptides and dyes. Peptide addition did not cause any variation in the fluorescence intensity decays, while the anisotropy decays were significantly affected.
Time-resolved anisotropy data were satisfactorily fitted by a biexponential decay (Equation 2 in Materials and Methods), indicating that the probe motions are more complex than a simple rotation. The equation allows the calculation of the fraction residual anisotropy (f∞ in the equation), which indicates how hindered are the motions at long times. Data analysis of the biexponential fit for all the probes and peptides are reported in Supplementary Table S1. These measurements validate the results obtained with steady-state anisotropy: Although the errors are relatively high, the addition of all the tested peptides undoubtedly causes an increase in both the fraction of residual anisotropy, and the average rotational correlation time (f∞) and <ϕ> in Equation 2 and 3, respectively) (Figure 5).
This behavior indicates that the probe motions are both slower and more hindered in the presence of the peptide.
2.4. Water Penetration
Polar solutes such as water molecules can diffuse through the polar headgroup of the phospholipid membrane. The permeation of the bilayer is particularly favored at higher temperatures, when the lipid membrane undergoes the phase transition from the liquid disordered to the liquid ordered phase. Laurdan fluorescence is very sensitive to the solvent relaxation effect. When the water molecules surround the naphthalene moiety, the maximum wavelength of the fluorescence spectrum moves from about 435 nm to about 475 nm (Figure 6) [55]. These spectral variations can be quantified by calculating the generalized polarization (GP) parameter, which has positive values when the spectrum is blue shifted (apolar environment), and negative values when it is red-shifted (polar environment). GP is defined by analogy to fluorescence polarization, according to the equation 4 in Materials and Methods [56]:
Considering GP as a function of peptide concentration, both magainin 2 and melittin caused a reduction in water penetration into the lipid head group region (Figure 7). This finding is consistent with the membrane stiffening observed in the anisotropy measurements. The same behavior was observed for LAH4, at both pH values.
2.5. Lipid Lateral Mobility
Lateral diffusion is another key dynamic feature of the bilayer, as it is indirectly related to the lipid-domain formation and stability, and also affects the bidimensional diffusion of molecules in the bilayer plane. To investigate how the peptides affect this property, we exploited the photophysical properties of pyrene. This fluorophore can form excited-state dimers (or excimers) when an excited and a ground-state molecule come in contact. Excimers are fluorescent, but their emission spectrum is very different from that of the monomer and is shifted to longer wavelengths. Excimer formation depends on the local concentration of pyrene molecules and on their mobility, since it is a diffusion-limited process. Therefore, the fluorescence of lipids labelled with pyrene can give information on the lipid diffusion; in particular, we measured the ratio between the intensities of the excimer and monomer emission bands, peaked at 475 nm and 397 nm, respectively (E/M ratio). A decrease in the lateral mobility of lipids will be reflected in a lower value of the ratio between the emission bands of excimer and monomer [57,58].
Pyrene fluorescence spectra were recorded by titrating a liposome suspension (containing 3% of PC chain covalently labeled with pyrene) with increasing peptide concentrations. As an example, the relative intensity of excimer emission of pyrenyl-lipid decreased after addition of magainin 2, indicating a decrease in the lateral mobility of the phospholipids (Figure 8). E/M ratios at different peptide concentrations are reported in Figure 9 for all the peptides investigated. The data demonstrated a decrease in excimer emission, and thus a lower lipid lateral mobility after peptide addition. This effect was most evident at peptide concentrations above those required to cause leakage.
Lipid vesicles employed so far were labelled with pyrene covalently linked to the PC hydrocarbon chain. To rule out the possibility that the observed effect was due to a specific interaction of the peptide with the zwitterionic lipid, the same experiment was performed with magainin 2 and melittin by using the pyrene probe linked to the anionic lipid phosphatidylglycerol (POPG). A very similar trend was observed, indicating that the reduced mobility of the probe is not due to a direct interaction between peptide and the labelled lipid (Figure 10), but is a general AMP-induced membrane effect.
2.6. Vesicle Agglutination Experiments with Magainin 2
Peptide-induced vesicle aggregation is a common observation with cationic AMPs [57,58]. Therefore, we studied this process for one of the peptides, in order to verify if it had any role in the perturbation of membrane dynamics observed in the experiments reported in the previous subsections. Aggregation was followed by measuring the increasing of absorbance at 600 nm due to the light scattering caused by sample turbidity.
Like in the other experiments, a suspension of POPC/POPG liposomes (2:1 molar ratio, lipid concentration = 50 µM) was titrated with increasing magainin 2 concentrations (Figure 11). At approximately 5 μM of peptide, a sharp increase in apparent absorbance was observed, followed by a rapid reversal of this phenomenon. This behavior is consistent with what has been previously observed for similar systems [59,60]. When the peptide binds to the anionic membranes, its positive charges progressively neutralize the negative charges of the lipids, favoring the formation of aggregates. When more peptide molecules are added, the process is reversed, since the total net charge becomes positive, causing repulsion between vesicles, and a reversal in the aggregation.
It is thus reasonable to question if all the variations in membrane dynamics reported in the previous sections arise from the peptide induced perturbation of the bilayer, or are simply a consequence of vesicle aggregation. To clarify this point, we used steric repulsion between vesicles to reduce their aggregation, by including in the lipid mixture phospholipids derivatized at their head group with the hydrophilic polymer polyethylene glycol (PEG) (POPC, POPG, PEG-PE, DPH; 66, 31, 2, 1 mole %, respectively). This approach essentially eliminated liposome aggregation (Figure 11, left panel). By contrast, the increase in DPH anisotropy was comparable to that previously observed in POPC/POPG vesicles (Figure 11, right panel), ruling out the possibility that the peptide-induced membrane stiffening is an artefact arising from vesicle aggregation.
2.7. Bioactive Compounds: Influence on Membrane Dynamics
There is an increasing trend toward shifting from antimicrobial peptides to the design of peptidomimetics capable of perfectly mimicking the behavior of AMPs toward biological membranes. The advantage of these compounds lies in their greater stability and lower synthesis costs compared to peptides. Growing evidence indicates that these peptidomimetics behave in the same way as AMPs in terms of their ability to disrupt membranes [30]. It is worth asking whether they are also capable of employing additional or alternative mechanisms that could lead to greater efficacy in membrane permeabilization by inducing a global perturbation of the bilayer dynamics, as observed in this work for AMPs. To assess whether our findings apply more broadly to structurally different membrane-active compounds and not only to AMPs, we extended the same spectroscopic studies to other membrane-active molecular classes, including the anticancer peptide Killer-FLIP [28] and the two norspermidine-based peptidomimetics, Nor-Leu and Nor-Phe [30]. Like AMPs, they disrupt the phospholipid bilayer by creating defects, compromising its integrity and leading to membrane perturbation and ultimately cell death (see Figure S2 and S3 for the membrane-perturbing activity of these compounds).
As previously reported, we focused on steady state anisotropy (Figure 12) and generalized polarization measurements of the Laurdan probe (Figure 13). Also for these classes of molecules an increase in anisotropy is observed, indicating that the probe is surrounded by a more viscous environment after the bioactive compound addition. Of note, in these cases the effect on the membrane fluidity takes place in the same range of permeabilization, leaving open the question of whether the bactericidal properties are due to a concomitant mechanism in which both the pore formation and effect on the lipid dynamics play a role.
The generalized polarization of the probe is affected as well. The interaction of Killer-FLIP, Nor-Phe and Nor-Leu with model membranes causes a decrease in the hydration of the bilayer, as shown by the increase of GP values.
2.8. Comparison Between Membrane-Active Peptides and Peptidomimetics
Results obtained so far indicate that both membrane-active peptides and peptidomimetics induce stiffening of the entire bilayer in model membranes. However, to directly compare the effects of the two classes of compounds on membrane dynamics, further considerations are necessary. Peptides analyzed so far range from 23 residues for magainin 2, to 26 residues for melittin and LAH4, and 18 residues for killer-FLIP. Consequently, a normalization for the different surface of each molecule can help to account for effects due to mere peptide higher dimension.
From molecular dynamics simulations, it is known that the well-studied antimicrobial peptide PMAP-23, which is 23 residues long, occupies a surface area of 5.4 nm² when folded into its helical conformation [61]. The AMPs analyzed in this work are comparable in terms of length, net charge, and secondary structure; therefore, the same surface area of 5.4 nm² was assumed for the peptides investigated here, but the value was further normalized to take into account the slight difference in length between PMAP-23 and the peptides investigated in this study.
Similarly, for the nor-spermidine-based molecules, a surface area of 0.8 nm² was considered. This value was calculated for a related nor-spermidine compound containing a tryptophan residue, instead of phenylalanine or leucine [30,62].
When the concentrations of peptides and peptidomimetics were rescaled to account for the surface area of each molecule, the effects on membrane dynamics occurred within the same range for all the compounds tested, indicating a general behavior of membrane-active molecules (Figure 14), independently of possible different configurations of the analyzed peptides into the bilayer. The only compound showing a slight deviation from the others is Killer-FLIP, for which the GP parameter exhibits higher values. However, it should be noted that the membrane composition in this case differs from the others, which could account for the observed differences in terms of GP behavior, since both the Laurdan fluorescence features [63,64] and the response to the interaction with peptides [65] are strongly dependent on the membrane composition.
3. Discussion
Bacteria exploit the diversity of their lipids to modify their membranes when facing environmental stress, allowing them to maintain the membrane in an optimal state and preserve its proper function. Several theories tried to explain this peculiar behavior. The homeoviscous adaptation was introduced by Sinensky, according to which the membrane undergoes a reconstruction to keep the bilayer fluidity at a constant value despite changing conditions (mainly temperature) [66]. In the same years, McElhaney proposed that it may not be necessary to maintain a precise level of fluidity; rather, organisms might adjust the temperature range of the thermal transition so that most lipids remain in the appropriate liquid-crystalline phase [67]. Later, Silvius [68] speculated that although cells cannot grow when their membranes are in the gel state, they can tolerate a broad spectrum of fatty acid composition and fluidity within the liquid-crystalline phase. More in general, although the primary target of AMPs remains the bacterial membrane, increasing evidences indicate that they can act through multiple mechanisms to induce cell death [10,69,70].
Nowadays, it is widely accepted that membrane fluidity can change due to interactions between lipids and environmental factors like temperature, pH, pressure, ions, water, nutrients, and chemicals. Alterations in one of these factors trigger bacterial adaptive responses, making membrane adjustments a key part of the multi-factorial stress adaptation process necessary for cell survival. In the present work, we investigated the behavior of membrane-active compounds with a particular focus on this aspect.
Our previous works demonstrated that antimicrobial peptide molecules must fully cover the bacterial cell membrane in order to exert their bactericidal activity [11,12,71]. At these very high peptide concentrations, the peptides interfere with the bilayer dynamics [13]. This behavior was observed both in model vesicles and in live bacterial cells.
In the present study, we systematically extended these investigations on model membranes to membrane-active peptides and small molecules from different classes.
The peptidomimetics employed are cationic and amphiphilic. Despite their simple structures, these molecules have been shown to act similarly to antimicrobial peptides, primarily targeting the bacterial membrane. Their interaction with the bilayer is largely through binding to the membrane surface just below the head-group region, and insertion of their apolar moieties (amino acids side chain and fatty acid chain) into the bilayer core, leading to defect formation once a high level of membrane coverage is reached [72].
Anisotropy measurements conducted using three different probes, positioned at different depths and spanning the entire bilayer, indicate an overall increase in membrane viscosity.
Sporadic reports described effects on membrane dynamics similarly to those discussed here, with different antimicrobial peptides. In particular, an increase in generalized polarization is observed, which usually correlates with a reduced membrane fluidity in vesicles [73,74,75,76]. Here, we show that effects on bilayer fluidity take place at higher concentration with respect to pore formation. The evidence that the GP values, likely due to probes spread on the whole liposome, remain unchanged at the concentrations in which the pores form, strongly suggest that membrane permeabilization involves only a small fraction of the bilayer. Very likely a great part of phospholipids remains far from the pore, and this portion undergoes a reduction in fluidity, for the system here investigated only under conditions in which membrane permeabilization is already complete. However, our results extend beyond membrane hydration, providing a broader view of the overall effects on membrane dynamics. The outcomes from viscosity measurements, membrane hydration, and lipid lateral mobility are in perfect agreement, all pointing to a peptide-induced membrane stiffening, at comparable or slightly higher concentrations with respect to those needed to form pores (1–10 µM). This suggests that membrane stiffening is not caused by the formation of defects within the bilayer, but rather by a an almost complete coverage of the vesicle surface by the peptides [11,12,71]. Indeed, for several AMPs, very high peptide-to-lipid (P/L) molar ratios have been reported to induce membrane permeabilization, approaching complete membrane surface coverage in both liposomes and bacteria [77]. The effects observed here occur at even higher peptide concentrations, consistent with the “carpet” mechanism previously described for the analyzed peptides.
Nor-Leu and Nor-Phe peptidomimetics appear to affect membrane dynamics at higher concentration ranges. However, considering the difference in molecular size between these compounds and AMPs, the effective activity range becomes narrower and perfectly overlaps with that of AMPs. Thus, even for the smaller peptidomimetics, once the full coverage of the vesicle surface is most likely reached, a crucial effect is the stiffening of the entire bilayer.
Lipid domain formation is sometime recalled to explain the fluorescence signals indicating higher rigidity [74]. This alternative mechanism cannot be definitively ruled out, however, our data show that the same effect is obtained with three different probes placed at different depths into the bilayer. Excluding the unlikely possibility that all the probes are located into the more rigid domains, our evidences suggest a more general stiffening of the whole membrane.
Overall, the evidence that only minor effects on the lipid dynamics are observed at concentrations leading to pore formation, suggests that the so-called “sand in a gearbox” mechanism does not play a significant role in liposome permeabilization for the system here investigated. Of note, in bacteria changes in membrane fluidity could lead to a variations in lipid and protein packing, with possible implications on their overall physiological functioning. For this reason, the evidence here reported that pore formation and influence on the membrane fluidity are independent phenomena, taking place at different concentration ranges, strongly support the idea to perform these kind of measures in bacteria too [13].
4. Materials and Methods
4.1. Materials
POPG (1-palmitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glyc- erol)]), POPC (1-palmitoyl-2-oleoyl-sn-glycero-phosphocholine), POPS (1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-L-serine), NBD-PE (L-α-phosphatidylethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4- yl)), and PEG-PE (1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N- [methoxy(polyethylene glycol)-2000]) were purchased from Avanti Polar Lipids (Alabaster, Al); Pyr-PC (1-palmitoyl-2-(pyrene-1-yl)dec- anoyl-sn-glycero-3-phosphocholine) and Pyr-PG (1-palmitoyl-2-(pyr- ene-y-yl)decanoyl-sn-glycero-3-phosphoglycerol) were obtained from Sigma Aldrich (St. Louis, MO); DPH (diphenylhexatriene) and Laurdan (2-(dimethylamino)-6-dodecanoylnaphthalene) were purchased from Fluka (St. Louis, MO). Spectroscopic-grade chloroform and methanol (Carlo Erba, Milano, Italy) were used. Triton X-100 was purchased from Acros (Geel, Belgium).
Synthesis of Peptide and Peptidomimetics
The peptides LAH4, melittin and magainin 2 were prepared by solid-phase peptide synthesis on Millipore 9050 or ABI433 automatic peptide synthesizers using Fmoc chemistry. A fourfold excess of Fmoc-protected amino acids (Bachem, Heidelberg, Germany; Applied Biosystems, Weiterstadt, Germany) was used during chain elongation. After TFA cleavage was achieved, peptides were purified by preparative HPLC using an acetonitrile/water gradient in the presence of 0.1% TFA and a 214 nm detection wavelength. The identity and high purity of the products were confirmed by MALDI mass spectrometry and analytical HPLC. After lyophilization, the TFA counterions were exchanged in 5% acetate (v/v).
The two nor-spermidine analogues (Nor-Leu and Nor-Phe) were synthesized through two-step reactions. In the first step of the reactions, the carboxylic acid group of Boc-protected amino acids (phenylalanine or leucine) were coupled with the two primary amine groups of the lipophilic norspermidine derivatives through an amide bond using HBTU as coupling agent. In the second step of the reaction, the final compounds were achieved through deprotection of Boc groups by using trifluoroacetic acid (TFA). Finally, the compounds were characterized by 1 H NMR and HR-MS and their purity was determined by HPLC, which proved that the compounds were more than 95% pure.
KillerFLIP was purchased from Biomatik
4.2. Instrumentation
Steady-state fluorescence experiments were carried out with a Fluoromax-4 fluorimeter (Horiba, Edison, NJ). Time-resolved measurements were performed with a Lifespec-ps fluorometer (Edinburg instrument, Edinburgh, UK). Sample temperature was controlled to 25.0 ◦C (within 0.1 ◦C) with a thermostated water bath.
4.3. Liposome Preparation
Large unilamellar vesicles were prepared by dissolving in a 1:1 (vol/vol) chloroform/methanol mixture POPC/POPG lipids (2:1 molar ratio), and the appropriate fluorophore (Laurdan, NBD-PE, DPH, Pyr-PG, or Pyr- PC), or PEG-:PE (2 mol% with respect to the total lipid concentration), when needed. The fluorophore was at a ratio of 1% with respect to the total lipids, with the exception of Pyr-PG and Pyr-PC, which were 3%. Measurements with Killer-FLIP were conducted on POPS/POPC vesicles (1:1 molar ratio) to better mimic the composition of cancer cell membrane. The solvents were evaporated under reduced argon atmosphere until a thin film was formed. Complete evaporation was ensured by applying a rotary vacuum pump for at least 2 h. The lipid film was hydrated with 10 mM phosphate buffer (pH 7.4) containing 140 mM NaCl and 0.1 mM EDTA. In leakage experiments, the lipid film was hydrated with 30 mM 5,6-carboxyfluorescein (CF) or calcein solution, titrated to pH 7.4 with NaOH, and containing 10 mM phosphate buffer and 80 mM NaCl to make it isotonic to dilution buffer. The liposome suspension was vigorously stirred and 10 freeze and thaw cycles were performed. The suspension was extruded 31 times through two stacked polycarbonate membranes with 100 nm pores, to ensure a narrow size distribution. Liposomes hydrated with the CF or Calcein solutions, respectively, were separated from unencapsulated dye by gel filtration on a Sephadex G-50 medium column. The final lipid concentration was determined by the Stewart method [78].
4.4. Liposome Leakage
Perturbation of membrane permeability was determined by measuring the fractional release of the fluorophore (Calcein or CF, respectively) entrapped inside liposomes at self-quenching concentration. This quantity can be measured directly by the increase in fluorescence intensity caused by the reduction in dye self-quenching 20 minutes after peptide addition. The fractional release was calculated as follows (Equation1):
where F0 is the fluorescence of the probe before peptide addition, and F100 is the intensity after complete disruption of the vesicles caused by the addition of Triton X-100 (1 mM). The release kinetics was recorded for both probes with a FluoroMax-4 fluorimeter with excitation and emission wavelengths of 490 nm (bandwidth 1.5 nm) and 520 nm (bandwidth 0.2 nm), respectively.
4.5. Steady-State and Time-Resolved Anisotropy
Perturbation of membrane dynamics caused by peptides or peptidomimetics was analyzed by experiments of steady-state and time-resolved anisotropy of three different probes, DPH, Laurdan and NDB-PE. In all cases, the probe to phospholipids molar ratio was fixed to 1:100.
For DPH steady-state anisotropy, the experimental conditions were set as follows: excitation wavelength 372 nm, emission wavelength 450 nm, bandwidth 4 nm and 385 nm cut-off filter. For NBD-PE, the excitation wavelength was set to 460 nm, emission 530 nm, bandwidth 4 nm and 495 nm cut-off filter.
For Laurdan, the excitation wavelength was set to 372 nm, emission to 475 nm, cut-off filter 385 nm and bandwidth 3, 5 nm in excitation and emission, respectively.
Each experimental point was determined fifteen times, and the mean value was reported.
Time-resolved fluorescence measurements were performed on a Lifespec-ps setup (Edinburgh Instruments, Livingstone, UK) equipped with automatic Glan-Thompson polarizers, a cooled micro-channel plate detector and working in the time correlated single photon counting mode (TCSPC). The excitation sources were a diode laser at 440 nm (NBD-PE) and 372 nm (DPH and Laurdan), emission band pass was fixed at 8 nm, and emission wavelength 450 nm (DPH), 530 nm (NBD-PE), 475 nm (Laurdan).
Data were fitted with a biexponential decay, according to the following Equation 2:
where r0 is the limiting anisotropy, ϕ the rotational correlation times and f the fraction of residual anisotropy. The average rotational correlation time has been calculated using the following Equation 3:
4.6. Lipid Lateral Mobility
Liposomes were labelled with the fluorescent probe pyrene covalently linked to the hydrocarbon chain of phospholipids. Liposome with two different compositions were prepared: POPC/POPG/Pyr-PG (66:30:3 molar ratio) and POPC/POPG/Pyr-PC (63:33:3 molar ratio).
Pyrene fluorescence spectra (370–700 nm) were recorded at increasing peptide concentrations (excitation wavelength 328 nm, integration time 0.1 s, excitation and emission bandwidths 1 nm and 1.5 nm respectively). Excimers over monomer fluorescence intensity was measured as the ratio between fluorescence values at 475 nm and 397 nm.
4.7. Generalized Polarization
Spectra of Laurdan-labelled liposomes were recorded from 400 nm to 520 nm (excitation wavelength 372 nm, 2 nm bandwidth both in excitation and emission). The generalized polarization was calculated as follows (Equation 4):
where Iλ is the fluorescence intensity relating to a specific emission wavelength λ.
4.8. Turbidity Measurements
Peptide-induced vesicle aggregation was followed by titrating a fixed lipid concentration (50 μM) with increasing magainin 2 concentrations. The variation of the optical density at 600 nm was recorded using a Cary 100 Scan UV-visible spectrophotometer (Agilent, Santa Clara, CA). Each experimental point was determined three times, and the mean value was reported.
4.9. Software
Graphs were generated using KaleidaGraph (Synergy Software, Reading, PA, USA). The graphical abstract was prepared with the assistance of ChatGPT (OpenAI, https://chatgpt.com).
5. Conclusions
All the parameters examined in this study, membrane viscosity, hydration, and lipid lateral mobility, consistently point in the same direction, indicating that membrane-active molecules cause a global decrease in the membrane mobility on the nanosecond timescale, regardless of their composition, length, charge, or mechanism of action in vesicles.
While membrane permeabilization is achieved at relatively low peptide concentrations, alterations in membrane dynamics become evident only at higher concentrations, most likely reflecting an almost complete coverage of the bilayer surface, where alternative peptide-lipid arrangements may take place.
When normalized for molecular size and surface area, peptidomimetics show the same activity range as AMPs, confirming that full membrane coverage is required to induce perturbation and bilayer stiffening.
The workflow designed here, transferable to bacteria too [13], helps to distinguish among membrane-active compounds for which the “Sand in gearbox” mechanism could play a role.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: release of CF and calcein from liposomes after addition of LAH4; Figure S2: Membrane-perturbing activity of membrane-active compound killer FLIP and the two peptidomimetics Nor-Leu and Nor-Phe. Figure S3: Fraction of CF leakage induced by Nor Leu and Nor Phe from liposomes; S4: Comparison of Laurdan steady-state anisotropy in liposomes labeled at 1% or 0.1% with the probe. Table S1: Parameters of a biexponential fit of time-resolved anisotropy for magainin 2, melittin and LAH4 with the different probs.
Author Contributions
Conceptualization, L.S., D.R.; methodology, L.S. and D.R.; software, Z.V., A.B., C.D.S., S.B., V.S., C.A. and D.R.; validation, Z.V., A.B., C.D.S., S.B., V.S, F.C., C.A. and D.R.; formal analysis, Z.V., A.B., C.D.S., S.B., V.S, F.C., C.A., M.V. and D.R.; resources, C.H., M.M.K., Y. A., R.K.F., B.B., J.H. L.S.; data curation, Z.V., D.R.; writing—original draft preparation, D.R., G.B.; writing—review and editing, D.R., C.H., M.M.K., M.V., R.K.F., B.B., J.H. and G.B.; supervision, L.S. and G.B.; funding acquisition, L.S., G.B. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding
Acknowledgments
The authors acknowledge Paolo Calligari, Emanuela Gatto and Claudia Mazzuca for useful discussion.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMR | Antimicrobial resistance |
| AMP | Antimicrobial peptide |
| CF | carboxyfluorescein |
| DDQ | depth depending quenching |
| MD | molecular dynamics |
| GP | generalized polarization |
References
- Lazzaro, B. P.; Zasloff, M.; Rolff, J. Antimicrobial Peptides: Application Informed by Evolution. Science 2020, 368, eaau5480. [CrossRef]
- Bobone, S.; Stella, L. Selectivity of antimicrobial peptides: A complex interplay of multiple equilibria. Adv. Exp. Med. Biol. 2019, 1117, 175-214. [CrossRef]
- Zhang, L.; Rozek, A.; Hancock, R.E.W. Interaction of cationic antimicrobial peptides with lipid membranes. J. Biol. Chem. 2001, 276, 35714–35722. [CrossRef]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic pepties. Biochim. Biophys. Acta 1999, 1462, 55–70. [CrossRef]
- Reddy, K.V.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: premises and promises. Int. J. Antimic. Agents. 2004, 24, 536–547. [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed Up immunity: how antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: form follows function. Nat. Rev. Drug. Discov. 2012, 11, 37–51. [CrossRef]
- Hancock, R.E.W.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [CrossRef]
- Pag, U.; Oedenkoven, M.; Sass, V.; Shai, Y.; Shamova, O.; Antcheva, N.; Tossi, A.; Sahl, H.G. Analysis of in vitro activities and modes of action of synthetic antimicrobial peptides derived from an a-helical ‘sequence template’. J. Antimicrob. Chemother. 2008, 61, 341–352. [CrossRef]
- Roversi, D.; Luca, V.; Aureli, S.; Park, Y.; Mangoni, M.L.; Stella, L. How Many Antimicrobial Peptide Molecules Kill a Bacterium? The Case of PMAP-23. ACS Chem. Biol. 2014, 9, 2003−2007. [CrossRef]
- Savini, F.; Loffredo, M.R.; Troiano, C.; Bobone, S.; Malanovic, N.; Eichmannd, T.O.; Caprio, L.; Canale, V.C.; Park, Y.; Mangoni, M.L.; Stella, L. Binding of an antimicrobial peptide to bacterial cells: Interaction with different species, strains and cellular components. Biochim. Biophys. Acta - Biomembr. 2020, 1862, 183291. [CrossRef]
- Roversi, D.; Troiano, C.; Salnikov, E.; Giordano, L.; Riccitelli, F.; De Zotti, M.; Casciaro, B.; Loffredo, M.R.; Park, Y.; Formaggio, F.; Mangoni, M.L.; Bechinger, B.; Stella, L. Effects of antimicrobial peptides on membrane dynamics: A comparison of fluorescence and NMR experiments. Biophys. Chem. 2023, 300, 107060. [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. U.S.A. 1987, 84, 5449-5453. [CrossRef]
- Bechinger, B. (1997). Structure and functions of channel-forming peptides: magainins, cecropins, melittin and alamethicin. J. Membr. Biol. 1997, 156, 197-211. [CrossRef]
- Westerhoff, H.V.; Hendler, R.W.; Zasloff, M; Jureti, C. D. Interactions between a new class of eukaryotic antimicrobial agents and isolated rat liver mitochondria. Biochim. Biophys. Acta 1989, 975, 361-369. [CrossRef]
- Cruciani, R.A.; Barker, J.L.; Zasloff, M.; Chen, H.C.; Colamonici, O. Antibiotic magainins exert cytolytic activity against transformed cell lines through channel formation. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 3792-3796. [CrossRef]
- Habermann, E. Bee and wasp venoms. Science 1972, 177, 314–322. http://dx.doi.org/10.1126/science.177.4046.314.
- Dempsey, C.E. The actions of melittin on membranes. Biochim. Biophys. Acta 1990, 1031, 143–161. [CrossRef]
- Dempsey, C.E.; Bazzo, R.; Harvey, T.S.; Sypere, I. K.; Boheim, G.; Campbell, I.D. Contribution of proline-14 to the structure and actions of melittin. FEBS Lett. 1991, 281, 240–244. [CrossRef]
- Papo, N.; Shai, Y. New lytic peptides based on the D, L-amphipathic helix motif preferentially kill tumor cells compared to normal cells. Biochem. 2003, 42, 9346–9354. [CrossRef]
- Ladokhin, A.S.; White, S.H. “Detergent-like” permeabilization of anionic lipid vesicles by melittin. Biochim. Biophys. Acta 2001, 1514, 253–260. [CrossRef]
- Bechinger, B. Towards membrane protein design: pH-sensitive topology of histidine-containing polypeptides. J. Mol. Biol. 1996, 263, 768−775. [CrossRef]
- Georgescu, J.; Munhoz, V.H.O.; Bechinger, B. NMR Structures of the Histidine-Rich Peptide LAH4 in Micellar Environments: Membrane Insertion, pH-Dependent Mode of Antimicrobial Action, and DNA Transfection. Biophys. J. 2010, 99, 2507−2515. [CrossRef]
- Farrotti, A; Bocchinfuso, G; Palleschi, A.; Rosato, N.; Salnikov, E.; Voievoda, N.; Bechinger, B.; Stella, L. Molecular dynamics methods to predict peptide locations in membranes: LAH4 as a stringent test case. Biochim. Biophys. Acta 2015, 1848, 581-592. https://dx.doi.org/10.1016/j.bbamem.2014.11.002.
- Perrone, B.; Miles, A.J.; Salnikov, E.S.; Wallace, B.A.; Bechinger, B. Lipid interactions of LAH4, a peptide with antimicrobial and nucleic acid transfection activities. Eur. Biophys. J. 2014, 43, 499-507. http://dx.doi.org/10.1007/s00249-014-0980-y.
- Salnikov, E.; Aisenbrey, C.; Bechinger, B. Lipid saturation and head group composition have a pronounced influence on the membrane insertion equilibrium of amphipathic helical polypeptides, Biochim. Biophys. Acta-Biomembranes. 2022, 1864, 183844.
- Vaezi, Z.; Bortolotti, A.; Luca, V.; Perilli, G.; Mangoni, M. L.; Khosravi-Far, R.; Bobone, S.; Stella, L. Aggregation determines the selectivity of membrane-active anticancer and antimicrobial peptides: The case of killerFLIP. Biochim. Biophys. Acta - Biomembr. 2020, 1862, 183107. [CrossRef]
- Pennarun, B.; Gaidos, G.; Bucur, O.; Tinari, A.; Rupasinghe, C.; Jin, T.; Dewar, R.; Song, K.; Santos, M.T.; Malorni, W.; Mierke, D.; Khosravi-Far, R. killer-FLIP: a novel lytic peptide specifically inducing cancer cell death. Cell Death Dis. 2013, 4, e894. [CrossRef]
- Konai, M. M.; Adhikary, U.; Samaddar S.; Ghosh, C; Haldar J.; Structure−Activity Relationship of Amino Acid Tunable Lipidated Norspermidine Conjugates: Disrupting Biofilms with Potent Activity against Bacterial Persisters. Bioconjugate Chem., 2015, 26, 2442−2453. http://dx.doi.org/10.1021/acs.bioconjchem.5b00494.
- Carneri, F.; Troiano, C.; Giaquinto, G.; Roversi, D.; Franzyk, H.; Stella, L. Water-membrane partition and the mutant selection window of antimicrobial peptides: insights from liposome studies. Journal of Colloid and Interface Science 2025, 683, 1078-1086. [CrossRef]
- Stella, L.; Mazzuca, C.; Venanzi, M.; Palleschi, A.; Didonè, M.; Formaggio, F.; Toniolo, C.; Pispisa, B. Aggregation and water-membrane partition as major determinants of the activity of the antibiotic peptide trichogin GA IV. Biophys J. 2004, 86, 936–945. [CrossRef]
- Mazzuca, C.; Orioni, B.; Coletta, M.; Formaggio, F.; Toniolo, C.; Maulucci, G.; De Spirito, M.; Pispisa, B.; Venanzi, M.; Stella, L. Fluctuations and the rate-limiting step of peptide-induced membrane leakage. Biophys. J. 2010, 99, 1791–1800. [CrossRef]
- Allen, T.M. Theoretical and Practical Considerations in Preparing Liposomes for the Purpose of Releasing Drug in Response to Changes in Temperature and pH. In Liposome Technology 1st ed.; Gregoriadis, G.; CRC Press: Boca Raton, Florida, 1984; Vol 3, p.177-196. [CrossRef]
- Vogt, T. C.; Bechinger, B. The interactions of histidine-containing amphipathic helical peptide antibiotics with lipid bilayers. The effects of charges and pH. J. Biol. Chem. 1999, 274, 29115–29121. [CrossRef]
- Jameson, D. M. Introduction to fluorescence, 1st ed; CRC Press: Boca Raton, Florida, 2019; pp. 75-99.
- Abrams, F. S.; London, E. Extension of the parallax analysis of membrane penetration depth to the polar region of model membranes: Use of fluorescence quenching by a spin-label attached to the phospholipid polar headgroup. Biochemistry 1993, 32, 10826–10831. [CrossRef]
- De Vequi-Suplicy, C. C.; Benatti, C. R.; Lamy, M. T. Laurdan in fluid bilayers: position and structural sensitivity. J. Fluoresc. 2006, 16, 431–439. [CrossRef]
- Kaiser, R. D.; London, E. Location of diphenylhexatriene (DPH) and its derivatives within membranes: comparison of different fluorescence quenching analyses of membrane depth. Biochem. 1998, 37, 8180–8190. [CrossRef]
- Mukherjee, S.; Raghuraman, H.; Dasgupta, S.; Chattopadhyay, A. Organization and dynamics of N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-labeled lipids: a fluorescence approach. Chem. Phys. Lipids 2004, 127, 91–101. [CrossRef]
- Wolf, D. E.; Winiski, A. P.; Ting, A. E.; Bocian, K. M.; Pagano, R. E. Determination of the transbilayer distribution of fluorescent lipid analogs by nonradiative fluorescence resonance energy transfer. Biochem. 1992, 31, 2865–2873. [CrossRef]
- Kyrychenko, A.; Rodnin, M. V.; Ladokhin, A. S. Calibration of Distribution Analysis of the depth of membrane penetration using simulations and depth-dependent fluorescence quenching. J. Membr. Biol. 2015, 248, 583–594. [CrossRef]
- Filipe, H. A. L.; Santos, L. S.; Prates Ramalho, J. P.; Moreno, M. J.; Loura, L. M. S. Behaviour of NBD-head group labelled phosphatidylethanolamines in POPC bilayers: a molecular dynamics study Phys. Chem. Chem. Phys 2015, 17, 20066–20079. [CrossRef]
- Fernandes, F.; Coutinho, A.; Prieto, M.; Loura, L. M. Electrostatically driven lipid–protein interaction: Answers from FRET. Biochim. Biophys. ACTA – Biomembr. 2015, 1848, 1837-1848. [CrossRef]
- Loura, L. M. Lateral distribution of NBD-PC fluorescent lipid analogs in membranes probed by molecular dynamics-assisted analysis of Förster Resonance Energy Transfer (FRET) and fluorescence quenching. Int. J. Mol. Sci. 2012, 13, 14545-14564. [CrossRef]
- do Canto, A. M. T. M.; Robalo, J. R.; Santos, P. D.; Carvalho, A. J. P.; Ramalho, J. P. P.; Loura, L. M. S. Diphenylhexatriene membrane probes DPH and TMA-DPH: A comparative molecular dynamics simulation study. Biochim. Biophys. Acta 2016, 1858, 2647–2661. [CrossRef]
- Hurjui, I.; Neamtu, A.; Dorohoi, D. O. The interaction of fluorescent DPH probes with unsaturated phospholipid membranes: A molecular dynamics study. J. Mol. Struct. 2013, 1044, 134–139. [CrossRef]
- Paloncýová, M.; Ameloot, M.; Knippenberg, S. Orientational distribution of DPH in lipid membranes: a comparison of molecular dynamics calculations and experimental time-resolved anisotropy experiments. Phys. Chem. Chem. Phys. 2019, 21, 7594–7604. [CrossRef]
- Pebay-Peyroula, E.; Dufourc, E. J.; Szabo, A. G. Location of diphenyl-hexatriene and trimethylammonium-diphenyl-hexatriene in dipalmitoylphosphatidylcholine bilayers by neutron diffraction. Biophys. Chem. 1994, 53, 45–56. [CrossRef]
- Antollini, S. S.; Barrantes, F. J. Disclosure of discrete sites for phospholipid and sterols at the Protein−Lipid interface in native acetylcholine receptor-rich membrane. Biochemistry 1998, 37, 16653–16662. [CrossRef]
- Jurkiewicz, P.; Olzyńska, A.; Langner, M.; Hof, M. Headgroup hydration and mobility of DOTAP/DOPC bilayers: a fluorescence solvent relaxation study. Langmuir 2006, 22, 8741–8749. [CrossRef]
- Barucha-Kraszewska, J.; Kraszewski, S.; Ramseyer, C. Will C-Laurdan dethrone Laurdan in fluorescent solvent relaxation techniques for lipid membrane studies? Langmuir 2013, 29, 1174–1182. [CrossRef]
- Kraszewski, S.; De, K., Aisenbrey, C., Bechinger, B., Osella, S. Hydration- and Temperature-Dependent Fluorescence Spectra of Laurdan Conformers in a DPPC Membrane. Cell 2024, 13, 1232. [CrossRef]
- Lakowicz, J. R. Principles of fluorescence spectroscopy, 3th ed; Springer: Berlin, Germany, 2006; pp. 353-397.
- Harris, F. M.; Best, K. B.; Bell, J. D. Use of Laurdan fluorescence intensity and polarization to distinguish between changes in membrane fluidity and phospholipid order. Biochim. Biophys. Acta - Biomembr. 2002, 1565, 123–128. [CrossRef]
- Parasassi, T.; De Stasio, G.; Ravagnan, G.; Rusch, R. M.; Gratton, E. Quantitation of lipid phases in phospholipid vesicles by the generalized polarization of Laurdan fluorescence. Biophys. J. 1991, 60, 179–189. [CrossRef]
- Somerharju, P. Pyrene-labeled lipids as tools in membrane biophysics and cell biology. Chem. Phys. Lipids 2002, 116, 57–74. [CrossRef]
- Loura, L. M. S.; Fernandes, F.; Prieto, M. Membrane microheterogeneity: Förster resonance energy transfer characterization of lateral membrane domains. European Biophys. J. 2010, 39, 589–607. [CrossRef]
- Marquette, A.; Lorber, B.; Bechinger, B. Reversible liposome association induced by LAH4: a peptide with potent antimicrobial and nucleic acid transfection activities. Biophys. J. 2010, 98, 2544–2553. [CrossRef]
- Bobone, S.; Piazzon, A.; Orioni, B.; Pedersen, J. Z.; Nan, Y. H.; Hahm, K.-S.; Shin, S. Y.; Stella, L. The thin line between cell-penetrating and antimicrobial peptides: the case of Pep-1 and Pep-1-K. J. Pept. Sci. 2011, 17, 335–341. [CrossRef]
- Bocchinfuso, G.; Palleschi, A.; Orioni, B.; Grande, G.; Formaggio, F.; Toniolo, C.; Park, Y.; Hahm, K. S.; Stella, L. Different mechanism of action of antimicrobial peptides: insights from fluorescence spectoscopy experiments and molecular dynamic simulations. J. Pept. Sci. 2009, 15, 550−558. [CrossRef]
- Konai, M. M.; Ghosh, C.; Yarlagadda, V.; Samaddar, S.; Haldar, J. Membrane active phenylalanine conjugated lipophilic norspermidine derivatives with selective antibacterial activity. J. Med. Chem. 2014, 57, 9409– 9423. [CrossRef]
- Uppu, D. S. S. M., M. M. Konai, U. Baul, P. Singh, T. K. Siersma, S. Samaddar, S. Vemparala, L. W. Hamoen, C. Narayana, and J. Haldar. 2016. Isosteric Substitution in Cationic-Amphiphilic Polymers Reveals an Important Role for Hydrogen Bonding in Bacterial Membrane Interactions. Chemical Science. 2016 7, 4613-4623. https://pubs.rsc.org/en/content/articlelanding/2016/sc/c6sc00615a.
- Tortorella, A., Oliva, R., Giancola, C., Petraccone, L., Winter, R.. 2023. Bacterial Model Membranes under the Harsh Subsurface Conditions of Mars. Phys. Chem. Chem. Phys., 2024, 26, 760-769. https://pubs.rsc.org/en/content/articlehtml/2024/cp/d3cp03911k.
- Bechinger B. The SMART model: Soft Membranes Adapt and Respond, also Transiently, in the presence of antimicrobial peptides. J Pept Sci. 2015, 21, 346-55. https://onlinelibrary.wiley.com/doi/10.1002/psc.2729.
- Sinensky, M. Homeoviscous adaptation, a homeostatic process that regulates the viscosity of membrane lipids in Escherichia coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–525. [CrossRef]
- McElhaney, R.N. The effect of alterations in the physical state of the membrane lipids on the ability of Acholeplasma laidlawii B to grow at various temperatures. J. Mol. Biol. 1974, 84, 145–157. [CrossRef]
- Silvius, J.R.; Mak, N.; McElhaney, R.N. Why do prokaryotes regulate membrane lipid fluidity? In Membrane Fluidity, Biophysical Techniques and Cellular Regulation. 1st ed.; Kates, H., Kukis, A.; Humana Press: Clifton, New Jersey, 1980; pp. 213–222.
- Brogden, K. A. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238-250. [CrossRef]
- Epand R.M.; Epand R.F. Bacterial membrane lipids in the action of antimicrobial agents. J. Pept. Sci. 2011, 17, 298–305. [CrossRef]
- Loffredo, M.R.; Savini, F.; Bobone, S.; Casciaro, B.; Franzyk, H.; Mangoni, M.L.; Stella, L.. Inoculum effect of antimicrobial peptides. Proc. Natl. Acad. Sci. U.S.A. 2021 118, e2014364118. [CrossRef]
- Bortolotti, A.; Troiano, C.; Bobone, S.; Konai, M.M.; Ghosh, C.; Bocchinfuso, G.; Acharya, Y.; Santucci, V.; Bonacorsi, S.; Di Stefano, C.; Haldar, J.; Stella, L. Mechanism of lipid bilayer perturbation by bactericidal membrane-active small molecules. Biochim. Biophys. ACTA – Biomembr. 2023, 1865, 184079. [CrossRef]
- Bertrand, B.; Alejandro, C. I. R.; del Carmen Gutiérrez, M.; Rincon, G. S.; Monturiol-Gross, L.; Munoz-Garay, C. Evaluation of the antimicrobial efficiency of three novel chimeric peptides through biochemical and biophysical analyses. Arch. Biochem. Biophys. 2025, 770. 110449. [CrossRef]
- Scheinpflug, K.; Wenzel, M.; Krylova, O.; Bandow, J. E.; Dathe, M.; Strahl, H. Antimicrobial peptide cWFW kills by combining lipid phase separation with autolysis. Sci. Rep. 2017, 7, 44332. [CrossRef]
- Kikukawa, T.; Araiso, T. Changes in lipid mobility associated with alamethicin incorporation into membranes. Arch. Biochem. Biophys. 2002, 405, 214-222. [CrossRef]
- Watala, C.; Gwoździński, K. Melittin-induced alterations in dynamic properties of human red blood cell membranes. Chem.-Biol. Interact. 1992, 82, 135-149. [CrossRef]
- Melo, M.N.; Ferre, R.; Castanho, M.A.R.B. Antimicrobial peptides: linking partition, activity and high membrane bound concentrations. Nature Rev. Microbiol. 2009, 7, 245–250. http://dx.doi.org/10.1038/nrmicro2095.
- Stewart J.C.M. Colourimetric determination of phospholipids with ammonium ferrothiocyanate. Anal. Biochem. 1980, 104,10–14. [CrossRef]
Figure 1.
Chemical structure of the peptidomimetics Nor-Leu and Nor-Phe.

Figure 2.
Dye leakage from POPC/POPG (2:1 molar ratio) vesicles (lipid concentration 50 µM) induced by magainin 2, melittin and LAH4 at 298 K, in phosphate buffer 10 mM, and at pH 7.4, unless stated otherwise. The vertical dashed lines show the peptide concentration range that induced membrane leakage from >3% to >97%.
Figure 2.
Dye leakage from POPC/POPG (2:1 molar ratio) vesicles (lipid concentration 50 µM) induced by magainin 2, melittin and LAH4 at 298 K, in phosphate buffer 10 mM, and at pH 7.4, unless stated otherwise. The vertical dashed lines show the peptide concentration range that induced membrane leakage from >3% to >97%.

Figure 3.
Steady-state anisotropy of membrane probes as a function of peptide concentration for magainin 2 (upper left panel), melittin (upper right panel) and LAH4 (pH 5.0 bottom left panel; pH 7.4 bottom right panel). Lipid vesicles composition: POPC/POPG (2:1 molar ratio, 1% probe), lipid concentration 50 µM. DPH: λexc = 372 nm, λem = 450 nm; NBD: λexc = 460 nm, λem = 530 nm, Laurdan: λexc = 372 nm, λem = 475 nm. Bandwidth = 2 nm and 4 nm for excitation and emission slits, respectively. The peptide concentration range that induced membrane leakage from >3% to >97% is reported as a shaded area, for comparison.
Figure 3.
Steady-state anisotropy of membrane probes as a function of peptide concentration for magainin 2 (upper left panel), melittin (upper right panel) and LAH4 (pH 5.0 bottom left panel; pH 7.4 bottom right panel). Lipid vesicles composition: POPC/POPG (2:1 molar ratio, 1% probe), lipid concentration 50 µM. DPH: λexc = 372 nm, λem = 450 nm; NBD: λexc = 460 nm, λem = 530 nm, Laurdan: λexc = 372 nm, λem = 475 nm. Bandwidth = 2 nm and 4 nm for excitation and emission slits, respectively. The peptide concentration range that induced membrane leakage from >3% to >97% is reported as a shaded area, for comparison.

Figure 4.
Fluorescence intensity decay (left panel) and time-resolved anisotropy (right panel) of NBD-PE inserted into POPC/POPG (2:1 molar ratio) vesicles (lipid concentration = 50 μM), without peptide (black points) and with magainin 2 19 μM (blue points). Laser λexc =440 nm and λem=530 nm with filter 495 nm. Anisotropy data were fitted with biexponential decay (continuous lines).
Figure 4.
Fluorescence intensity decay (left panel) and time-resolved anisotropy (right panel) of NBD-PE inserted into POPC/POPG (2:1 molar ratio) vesicles (lipid concentration = 50 μM), without peptide (black points) and with magainin 2 19 μM (blue points). Laser λexc =440 nm and λem=530 nm with filter 495 nm. Anisotropy data were fitted with biexponential decay (continuous lines).

Figure 5.
Relative increase of fFraction of residual anisotropy (left) and rotational correlation time (right) of DPH, NBD and Laurdan after peptides addition.
Figure 5.
Relative increase of fFraction of residual anisotropy (left) and rotational correlation time (right) of DPH, NBD and Laurdan after peptides addition.

Figure 6.
Changes in the fluorescence spectra of Laurdan in POPC/POPG vesicles (lipid concentration = 50 μM, 2:1 phospholipids molar ratio, probe 1%) upon addition of peptide. λexc= 372 nm, bandwidth= 3 nm, and λem= 475 nm, bandwidth= 4 nm.
Figure 6.
Changes in the fluorescence spectra of Laurdan in POPC/POPG vesicles (lipid concentration = 50 μM, 2:1 phospholipids molar ratio, probe 1%) upon addition of peptide. λexc= 372 nm, bandwidth= 3 nm, and λem= 475 nm, bandwidth= 4 nm.

Figure 7.
Generalized polarization of Laurdan fluorescence emission resulting from vesicles titration with magainin 2 (upper left panel, lipid concentration = 50 μM, vesicle composition POPC/POPG 2:1 molar ratio, probe 1%), melittin (upper right panel) and LAH4 (bottom left panel) pH=5.0 and (bottom right panel) pH=7.4. λexc= 372 nm, bandwidth=2 nm, λem= 400-580 nm, bandwidth=1.5 nm, cut-off filter 385 nm.
Figure 7.
Generalized polarization of Laurdan fluorescence emission resulting from vesicles titration with magainin 2 (upper left panel, lipid concentration = 50 μM, vesicle composition POPC/POPG 2:1 molar ratio, probe 1%), melittin (upper right panel) and LAH4 (bottom left panel) pH=5.0 and (bottom right panel) pH=7.4. λexc= 372 nm, bandwidth=2 nm, λem= 400-580 nm, bandwidth=1.5 nm, cut-off filter 385 nm.

Figure 8.
Normalized pyrene spectra in the absence (black line) and after addition of peptide (magainin 2: 29 µM, blue line) to POPC/POPG/Pyr-PC 2:1 molar ratio vesicles (lipid concentration = 50 µM, probe 3%). Pyrene: λexc= 328 nm, bandwidth=1 nm, λem= 370-600 nm, bandwidth=1.5 nm.
Figure 8.
Normalized pyrene spectra in the absence (black line) and after addition of peptide (magainin 2: 29 µM, blue line) to POPC/POPG/Pyr-PC 2:1 molar ratio vesicles (lipid concentration = 50 µM, probe 3%). Pyrene: λexc= 328 nm, bandwidth=1 nm, λem= 370-600 nm, bandwidth=1.5 nm.

Figure 9.
Variations in the ratio of the fluorescence emission intensities of the excimer (475 nm) and monomer (397 nm) bands caused by titration of POPC/POPG/Pyr-PC 2:1 molar ratio vesicles (lipid concentration = 50 µM, probe 3%) with magainin 2 (top left panel), melittin (top right panel), and LAH4 at pH= 5.0 (bottom left panel) and pH=7.4 (bottom right panel).
Figure 9.
Variations in the ratio of the fluorescence emission intensities of the excimer (475 nm) and monomer (397 nm) bands caused by titration of POPC/POPG/Pyr-PC 2:1 molar ratio vesicles (lipid concentration = 50 µM, probe 3%) with magainin 2 (top left panel), melittin (top right panel), and LAH4 at pH= 5.0 (bottom left panel) and pH=7.4 (bottom right panel).

Figure 10.
Variations in the ratio of the fluorescence emission intensities of the excimer (475 nm) and monomer (397 nm) bands caused by titration POPC/POPG/Pyr-PG] (66:30:3) vesicles (lipid concentration = 50 μM) with magainin 2 and melittin.
Figure 10.
Variations in the ratio of the fluorescence emission intensities of the excimer (475 nm) and monomer (397 nm) bands caused by titration POPC/POPG/Pyr-PG] (66:30:3) vesicles (lipid concentration = 50 μM) with magainin 2 and melittin.

Figure 11.
Left: peptide-induced POPC/POPG (2:1 molar ratio) liposome aggregation as measured from turbidity. Optical density at 600 nm of vesicles (lipid concentration = 50 μΜ) in the presence of PEG (empty circles) and without it (full circles), titrated with increasing magainin 2 concentrations. Right: DPH anisotropy inserted into liposomes with PEG, (POPC, POPG, PEG-PE, DPH; 66, 31, 2, 1 mole %, respectively) resulting from titration with varying concentrations of magainin 2. Lines represents just a guide for the eye.
Figure 11.
Left: peptide-induced POPC/POPG (2:1 molar ratio) liposome aggregation as measured from turbidity. Optical density at 600 nm of vesicles (lipid concentration = 50 μΜ) in the presence of PEG (empty circles) and without it (full circles), titrated with increasing magainin 2 concentrations. Right: DPH anisotropy inserted into liposomes with PEG, (POPC, POPG, PEG-PE, DPH; 66, 31, 2, 1 mole %, respectively) resulting from titration with varying concentrations of magainin 2. Lines represents just a guide for the eye.

Figure 12.
Steady-state anisotropy of Laurdan resulting from vesicles titration with killer-FLIP (left panel, lipid concentration = 50 μM vesicle composition POPC/POPS 1:1 molar ratio - better mimicking the membranes in cancer cells) and peptidomimetics Nor-Leu and Nor-Phe (right panel, black: Nor-Phe, red: Nor-Leu. Vesicle composition POPC/POPG 2:1 molar ratio). Laurdan λexc= 372 nm, bandwidth= 2 nm, λem= 475 nm, bandwidth=1.5 nm, cut-off filter 385 nm.
Figure 12.
Steady-state anisotropy of Laurdan resulting from vesicles titration with killer-FLIP (left panel, lipid concentration = 50 μM vesicle composition POPC/POPS 1:1 molar ratio - better mimicking the membranes in cancer cells) and peptidomimetics Nor-Leu and Nor-Phe (right panel, black: Nor-Phe, red: Nor-Leu. Vesicle composition POPC/POPG 2:1 molar ratio). Laurdan λexc= 372 nm, bandwidth= 2 nm, λem= 475 nm, bandwidth=1.5 nm, cut-off filter 385 nm.

Figure 13.
Generalized polarization of Laurdan resulting from vesicles titration with killer-FLIP (left panel, lipid concentration = 50 μM vesicle composition POPC/POPS 1:1 molar ratio) and with peptidomimetics Nor-Leu and Nor-Phe (right panel, black: Nor-Phe, red: Nor-Leu. Vesicle composition POPC/POPG 2:1 molar ratio). Laurdan λexc= 372 nm, bandwidth= 2 nm, λem= 475 nm, bandwidth=1.5 nm, cut-off filter 385 nm.
Figure 13.
Generalized polarization of Laurdan resulting from vesicles titration with killer-FLIP (left panel, lipid concentration = 50 μM vesicle composition POPC/POPS 1:1 molar ratio) and with peptidomimetics Nor-Leu and Nor-Phe (right panel, black: Nor-Phe, red: Nor-Leu. Vesicle composition POPC/POPG 2:1 molar ratio). Laurdan λexc= 372 nm, bandwidth= 2 nm, λem= 475 nm, bandwidth=1.5 nm, cut-off filter 385 nm.

Figure 14.
Comparison of steady-state anisotropy (left panel) and GP (right panel) of Laurdan after the addition of all the AMPs and peptidomimetics used in the present work (Melittin: blue; Maginin 2: orange; LAH4 pH 5: green; LAH4 pH 7.4: light green; Killer-FLIP: light blues; Nor-leu: red; Nor-phe: black). Concentrations were normalized for the surface area occupied by AMPs or peptidomimetics. Vesicles composition: POPC/POPG at a 2:1 molar ratio for all compounds, except for Killer-FLIP, for which a POPC/POPS 1:1 molar ratio was employed.
Figure 14.
Comparison of steady-state anisotropy (left panel) and GP (right panel) of Laurdan after the addition of all the AMPs and peptidomimetics used in the present work (Melittin: blue; Maginin 2: orange; LAH4 pH 5: green; LAH4 pH 7.4: light green; Killer-FLIP: light blues; Nor-leu: red; Nor-phe: black). Concentrations were normalized for the surface area occupied by AMPs or peptidomimetics. Vesicles composition: POPC/POPG at a 2:1 molar ratio for all compounds, except for Killer-FLIP, for which a POPC/POPS 1:1 molar ratio was employed.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



