Submitted:
04 June 2026
Posted:
05 June 2026
You are already at the latest version
Abstract
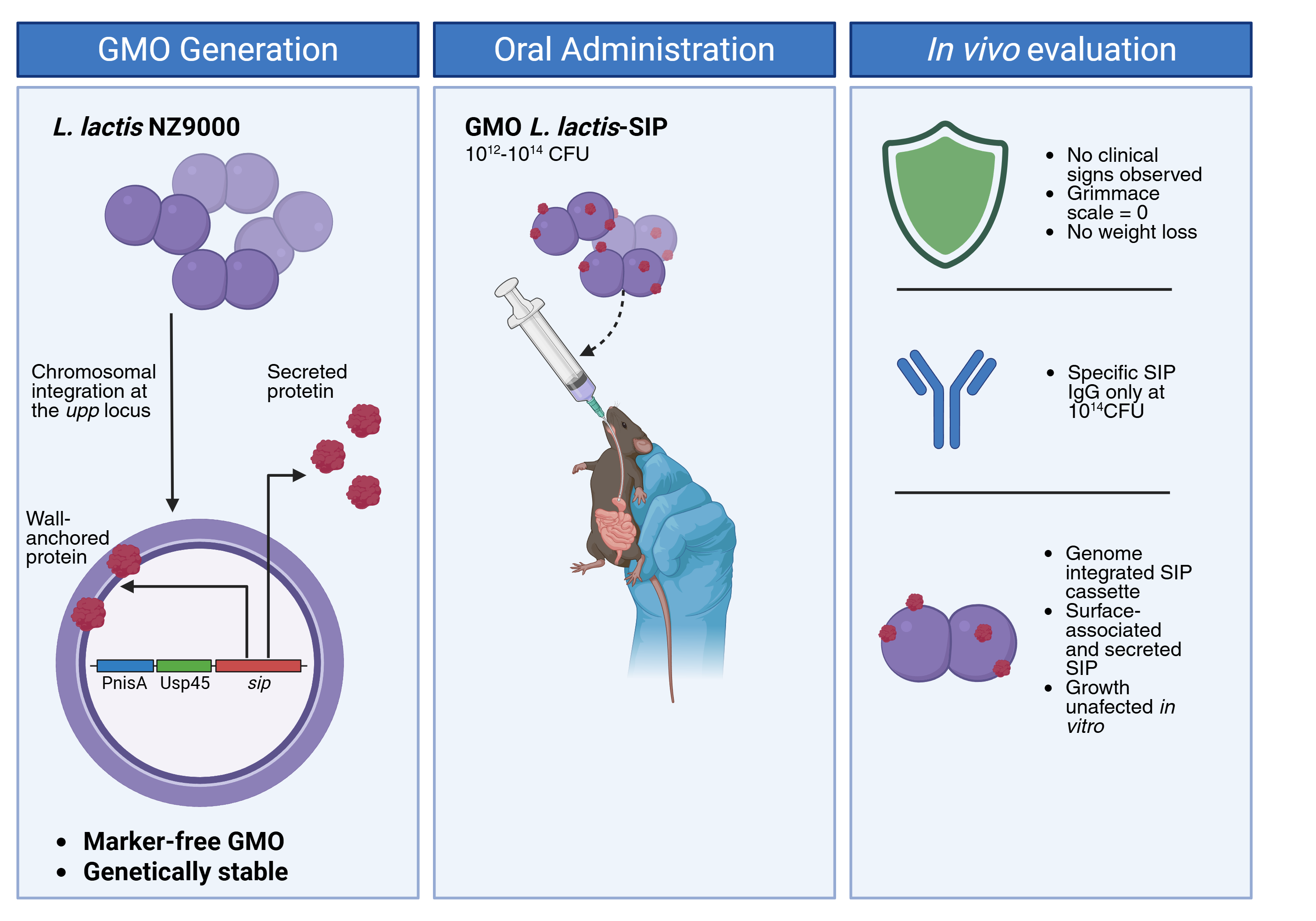
Food-grade lactic acid bacteria are increasingly recognized as promising candidates for mucosal immunization and various health applications. However, plasmid-based expression systems often face challenges related to antibiotic-resistance markers and genetic instability. To address these limitations, we developed and microbiologically characterized a marker-free strain of Lactococcus lactis subsp. cremoris NZ9000. This strain carries a chromosomally integrated expression cassette for the Surface Immunogenic Protein (SIP) of Streptococcus agalactiae, commonly known as Group B Streptococcus (GBS). Using an adapted pMBSacB sucrose-counterselection strategy, we successfully replaced the native upp locus with a nisin-inducible PnisA-Usp45-sip cassette. Whole-genome sequencing confirmed that the sip cassette was integrated at the designated locus, with no additional mutations or residual antibiotic-resistance determinants detected during our analysis. Importantly, this chromosomal insertion did not affect in vitro growth. Western blotting and confocal microscopy confirmed that the expression of rSIP is dependent on nisin and that rSIP is localized on the cell surface. In a pilot study in mice, L. lactis-SIP showed no adverse effects, including weight loss or any noticeable clinical changes. Importantly, serum analysis detected the presence of SIP-specific IgG, particularly at the highest evaluated dose of 1 × 10¹⁴ CFU. These findings support the potential of L. lactis-SIP as a genetically stable, marker-free platform for mucosal antigen delivery. This research paves the way for future evaluations of probiotic-based functional foods or live biotherapeutic formulations aimed at GBS.
Keywords:
Lactococcus lactis
; group B streptococcus
; surface immunogenic protein
; chromosomal integration
; marker-free genome editing
; pMBSacB
; mucosal antigen delivery
; live biotherapeutic
; probiotic-based platform
1. Introduction
Group B Streptococcus (GBS), scientifically known as Streptococcus agalactiae, is a Gram-positive, catalase-negative, β-hemolytic coccus that can colonize the human gastrointestinal and female genitourinary tracts [1]. GBS infection is a significant cause of neonatal morbidity and mortality, leading to conditions such as pneumonia, sepsis, meningitis, preterm delivery, and neonatal death [2]. The primary route of transmission to the fetus or newborn is through maternal rectovaginal colonization [3]. To date, ten serotypes of GBS have been identified based on their capsular polysaccharide composition (Ia, Ib, II-IX) [4]. Universal antenatal screening for maternal GBS colonization at 36-37 weeks of gestation, followed by intrapartum antibiotic prophylaxis (IAP) for colonized mothers, is the established strategy to prevent early-onset disease (EOD) [3]. Although IAP has significantly reduced the incidence of EOD, it does not prevent late-onset disease (LOD) and raises concerns regarding antimicrobial exposure. As a result, the development of an effective maternal GBS vaccine has become a global priority, with modeling studies indicating that vaccination could prevent a substantial number of annual neonatal deaths [5].
Currently, there is no licensed vaccine for GBS, although several formulations are in various stages of clinical or preclinical development [6]. Among the immunogens evaluated, the Surface Immunogenic Protein (SIP) stands out as a strong candidate due to its high conservation across all 10 GBS serotypes and demonstrated immunogenicity [7,8,9]. Recent studies suggest that SIP plays a role in GBS pathogenesis; for example, SIP suppression or loss leads to reduced adherence to human epithelial cells, decreased biofilm formation, and reduced uterine persistence in mouse models [10]. Additionally, SIP has been identified as an agonist of Toll-like receptors 2 and 4 [11], supporting its potential role in immunomodulation [12]. In silico analyses have also revealed B-cell epitopes associated with predicted immunostimulatory activity [13]. Notably, the administration of recombinant SIP combined with alum reduced vaginal GBS colonization in mice, whereas SIP alone did not [14]. These findings not only highlight SIP as a promising vaccine target but also indicate the need for efficient mucosal delivery methods and/or adjuvant strategies to enhance its effectiveness.
In response to these challenges, food-grade and probiotic-associated lactic acid bacteria have emerged as effective live delivery vehicles for antigens and bioactive molecules. This approach is particularly relevant to research in functional foods and health applications because engineered strains can potentially be formulated into fermented products, encapsulated formats, or dietary supplements and can directly interact with mucosal tissues [15,16,17,18,19]. Mucosal vaccination offers both practical and immunological benefits for pathogens that colonize mucosal surfaces, such as GBS. These advantages include the possibility of needle-free administration and the potential to stimulate local immune responses at the site of colonization [20]. However, oral antigen delivery faces significant challenges, including antigen degradation in the gastrointestinal tract, the issue of mucosal tolerance, and the necessity of effectively delivering antigens to gut-associated lymphoid tissue and professional antigen-presenting cells [21,22].
Lactococcus lactis is a non-pathogenic, Gram-positive lactic acid bacterium widely used in dairy fermentation and biotechnology. Its history as a food-grade organism, the absence of lipopolysaccharides, the availability of a fully sequenced genome, and access to various genetic tools have made it a preferred chassis for the mucosal delivery of bioactive proteins [15,23,24,25]. Within this species, the plasmid-cured MG1363 strain and its derivative, NZ9000, are commonly used for genetic manipulation and nisin-controlled expression [26,27,28,29]. Although L. lactis is often discussed in relation to probiotics, specific health benefits linked to individual strains must be demonstrated before making definite probiotic claims [30]. Therefore, in this study, NZ9000 is primarily regarded as a food-grade live-delivery platform with potential applications in probiotic-based biomedical fields.
Our research group previously evaluated an oral vaccine prototype based on Lactococcus lactis subsp. cremoris NZ9000, which expresses the SIP antigen from a plasmid (designated as L. lactis-pNZ8124-SIP). When this recombinant strain was orally administered to C57BL/6 mice, it successfully induced specific humoral and cellular immune responses against SIP. Furthermore, antibodies from vaccinated mice reduced intravaginal colonization by Group B Streptococcus (GBS) after passive transfer and demonstrated opsonophagocytic activity [31]. These findings support the use of L. lactis as a carrier for delivering oral GBS antigens; however, the current plasmid-based architecture is not optimal for further translational development.
Plasmids that carry antibiotic-resistance markers and mobile genetic elements raise regulatory and biosafety concerns, including the risk of marker spread, horizontal gene transfer, and genetic instability during bacterial replication [32,33]. To address these issues, this study aimed to construct and evaluate a marker-free L. lactis NZ9000 strain carrying a sip gene expression cassette integrated into its chromosome. This was achieved using an adapted pMBSacB sucrose-counterselection strategy.
We engineered the strain L. lactis subsp. cremoris NZ9000 to express rSIP using a nisin-inducible chromosomal cassette inserted at the upp locus. We characterized this strain using PCR, whole-genome sequencing, growth kinetics, Western blotting, confocal microscopy, and a pilot oral administration study. The study assessed clinical safety, body weight, and serum SIP-specific IgG levels. This design supports probiotic-based functional health applications by ensuring genetic stability, the absence of antibiotic-resistance markers, and the potential for mucosal antigen delivery.
2. Materials and Methods
2.1. Genome Editing Design
To integrate an rSIP expression cassette into the L. lactis NZ9000 genome, we chose the upp gene as the insertion site. The upp gene encodes uracil phosphoribosyltransferase, an enzyme involved in UMP biosynthesis and considered non-essential for L. lactis viability [34,35]. In the reference genome of L. lactis NZ9000 (GenBank accession CP002094.1), the upp gene is located on the complementary strand, spanning positions 2,145,563-2,146,198, and encodes a 636 bp open reading frame (ORF).
The editing strategy involved replacing 183 base pairs (bp) of the L. lactis NZ9000 genome. This included an 83 bp putative promoter region and the first 100 bp of the upp open reading frame (ORF). The replaced sequence was replaced with a 1,512 bp rSIP expression cassette previously used for rSIP expression in L. lactis. This cassette contained the nisin-inducible promoter PnisA (198 bp), the Usp45 secretion signal sequence (84 bp), and a fragment of the GBS sip ORF (1,230 bp; positions 76-1305 of GenBank accession KX363665.1).To facilitate allelic exchange, homology arms of 536 bp upstream and 718 bp downstream were designed based on the L. lactis NZ9000 genome. Additionally, XhoI and BamHI restriction sites were added to the 5′ and 3′ ends, respectively. The complete donor sequence, totaling 2,778 bp, was synthesized and cloned into the pUC57 vector by GenScript. Following protocol optimization for L. lactis, the pMBSacB vector [36] was subsequently employed for sucrose-counterselected allelic exchange.
2.2. Bacterial strains and growth conditions
Frozen glycerol stocks of L. lactis subsp. cremoris NZ9000 and Escherichia coli DH5α were streaked onto BHI and LB agar plates, respectively, and cultured in their corresponding broths at 30 °C for L. lactis and 37 °C for E. coli, unless indicated otherwise. Chemically competent E. coli DH5α (Thermo Scientific) was transformed following the manufacturer’s protocol. The E. coli strains carrying pMBSacB or pMBSacB-sip were incubated at 30 °C. When necessary, erythromycin was used at 1 µg/mL for L. lactis and 500 µg/mL for E. coli. BHI medium containing 0.75 M sucrose was prepared using a filter-sterilized 2 M sucrose stock solution (see Table 1).
2.3. Isolation of plasmid DNA
For plasmid extraction, E. coli DH5α::pUC57-sip was inoculated into 5 mL of LB broth supplemented with 100 µg/mL ampicillin and incubated at 37 °C for 16–18 hours with shaking at 220 rpm. E. coli DH5α::pMBSacB and E. coli DH5α::pMBSacB-sip were grown in 5 mL of LB broth supplemented with 500 µg/mL erythromycin and 3 µg/mL chloramphenicol at 30 °C for 48 hours, also with shaking at 220 rpm. Plasmid DNA was purified using the E.Z.N.A. Plasmid DNA Mini Kit I (Omega Bio-Tek) following the manufacturer’s instructions. The quality and concentration of DNA were assessed using a NanoDrop spectrophotometer (Thermo Fisher) and 1% agarose gel electrophoresis.
2.4. Subcloning and construction of the pMBSacB-sip vector
The pUC57-sip and pMBSacB vectors were digested with XhoI and BamHI (New England Biolabs) according to the manufacturer’s instructions. The desired DNA fragments were recovered from a 1% agarose gel using the PureLink Quick Gel Extraction Kit (Invitrogen). These fragments were then ligated with T4 DNA ligase (New England Biolabs) at a molar ratio of 3:1 (insert to vector) to create the pMBSacB-sip construct. The success of the ligation was confirmed by conventional PCR using SapphireAmp Fast PCR Master Mix (Takara) with T7 and M13 reverse primers at 48 °C (as indicated in Table 2). Chemically competent E. coli DH5α cells were then transformed with the ligation product. A recovered culture of 100 µL was plated on LB agar containing 500 µg/mL erythromycin and incubated at 30 °C for 24 to 48 hours.
2.5. Transformation of L. lactis with pMBSacB-sip and generation of single-crossover intermediates
Electrocompetent L. lactis NZ9000 cells were prepared using a previously described protocol with modifications [37]. For transformation and selection of single crossovers, 1 µg of pMBSacB-sip was added to 40 µL of electrocompetent L. lactis NZ9000 in a 2 mm electroporation cuvette. After incubating on ice for 15 minutes, the cells were pulsed at 2 kV for 4.5 to 5 ms, then immediately resuspended in 1 mL of BHI medium supplemented with 0.6% glycine, 20 mM MgCl₂, and 2 mM CaCl₂. This suspension was kept on ice for another 5 minutes. The cells were then incubated at 40 °C for 3 hours while shaking at 100 rpm. Following this, 100 µL of the culture was plated on BHI agar containing 1 µg/mL erythromycin and incubated at 40 °C for 8 to 24 hours. Transformant colonies were then inoculated into 3 mL of BHI broth with 5 µg/mL erythromycin and incubated at 40 °C with shaking at 100 rpm for 5 hours. Genomic DNA was purified using the GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich). Single-crossover integration was confirmed via PCR using the pMBSacB_MCS_F and KI_lactis_R primers with the SapphireAmp Fast PCR Master Mix (Takara). The amplicons of 3,993 bp or 3,017 bp indicated insertion of pMBSacB-sip through the first or second homology arm, respectively. PCR-positive cultures were promptly subjected to double-crossover selection and stored at -80 °C in broth supplemented with 20% glycerol.
2.6. Site-directed mutagenesis of the upp locus by allelic exchange and sucrose counterselection
A single-crossover strain was grown at 40 °C in brain heart infusion (BHI) broth supplemented with 5 µg/mL erythromycin and confirmed by PCR to facilitate spontaneous double-crossover resolution. One milliliter of this culture was then used to inoculate 100 mL of antibiotic-free BHI broth, followed by incubation at 30 °C for 18 to 24 hours with shaking at 100 rpm. The cultures were passaged at a 1:100 ratio in the same medium for three consecutive days. For the final transfer, the culture was diluted 1:100 into 100 mL of BHI broth supplemented with 0.75 M sucrose and incubated at 40 °C for 18 to 24 hours with shaking at 100 rpm. This step was repeated for three consecutive days. Then, serial 10-fold dilutions were plated on BHI agar containing 0.75 M sucrose and incubated at 40 °C for 18 to 24 hours. Double-crossover candidates were confirmed by endpoint PCR using the KI_lactis_F and KI_lactis_R primers. The expected amplicon sizes were 2,385 bp for L. lactis NZ9000 and 3,714 bp for L. lactis-SIP (see Table 2).
2.7. DNA sequencing
Genomic DNA was isolated from L. lactis-SIP and L. lactis NZ9000 using the GenElute Bacterial Genomic DNA Kit from Sigma-Aldrich. DNA libraries were prepared according to the manufacturer’s instructions using the Illumina DNA Prep kit. Specifically, 50 ng of genomic DNA was simultaneously fragmented and adapter-tagged using the Illumina tagmentation enzyme. The fragmented DNA was then purified with magnetic beads, followed by PCR amplification to add indices and Illumina adapters. After normalization, the libraries were pooled and sequenced on an Illumina MiSeq platform using paired-end 150 bp reads.
2.8. Bioinformatic analysis
Sequencing reads were assessed for quality using FastQC v0.12.1 [38]. Low-quality bases and adapter sequences were removed with BBDuk from the BBMap package v39.01. The filtered reads were then assembled into contigs using SPAdes v3.13.0 [39]. The qusembly w qualityas evaluated using QUAST v5.0.2 [40]. The sip gene was identified by comparing the assembled contigs with reference sequences from GenBank (accessions KX363665.1 and DQ914270.1) using BLASTn v2.14.1+ [41]. Sequencing reads were mapped to the reference sip sequence with BWA v0.7.17 [42], and the mapped reads were visualized using the Integrative Genomics Viewer (IGV) v2.17.4 [43]. Coverage analysis was conducted with BEDTools v2.31.0 [44] to confirm the presence and continuity of the integrated sip cassette.
2.9. Comparison of bacterial growth kinetics
L. lactis NZ9000 and L. lactis-SIP were cultured overnight in 2 mL of Brain Heart Infusion (BHI) broth at 30 °C with shaking at 100 rpm. The cultures were then subcultured in 200 mL of BHI broth under the same temperature and agitation conditions for 8 hours. For the culture inductions, nisin was added to a final concentration of 10 ng/mL when the cultures reached an optical density (OD600) of 0.4-0.6. Growth experiments were conducted with three independent biological replicates, and OD600 measurements were recorded every 20 minutes using a spectrophotometer.
2.10. Analysis of rSIP expression by Western blotting
Detection of rSIP expression was performed using a previously reported protocol [45] with modifications. In brief, 15 mL of L. lactis-SIP culture in BHI broth was grown to the exponential phase (OD600 between 0.4 and 0.6) at 30 °C while shaking at 100 rpm. The culture was induced with 10 ng/mL of nisin and incubated for 1 hour. The cultures were then centrifuged to separate the supernatant from the bacterial pellet. The supernatant was stored at -20 °C, while the pellet was processed for total protein extraction using B-PER reagent according to the manufacturer’s instructions. Protein concentration was measured using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific). For SDS-PAGE analysis, 50 µg of total protein was loaded per sample. The proteins were transferred to 0.45 µm nitrocellulose membranes (Amersham Protran) at 350 mA for 90 minutes. The membranes were blocked and incubated overnight at 4 °C with an in-house rabbit polyclonal anti-rSIP IgG antibody, diluted 1:5,000 in the blocking buffer. A goat anti-rabbit IgG peroxidase conjugate (Merck) was used as the secondary antibody, also diluted at 1:5,000.
2.11. Indirect immunofluorescence and confocal microscopy
Bacteria in the exponential growth phase were washed three times with phosphate-buffered saline (PBS), placed on glass slides, air-dried, and fixed with 4% paraformaldehyde in PBS for 20 minutes. To reduce autofluorescence associated with the fixative, we quenched the slides with 50 mM ammonium chloride (NH₄Cl) for 20 minutes. Following this, the slides were washed with PBS and incubated at room temperature for 3 hours with rabbit anti-SIP polyclonal serum diluted 1:100 in PBS containing 0.15% gelatin and 1% saponin. After washing, the slides were incubated for 1 hour with Alexa Fluor 488-conjugated anti-rabbit IgG (Invitrogen), diluted 1:500 in the same buffer containing 1 µM TO-PRO-3 iodide. Samples were then mounted in ProLong Gold antifade reagent and analyzed using a Leica TCS SP5 confocal laser-scanning microscope with a 100× oil-immersion objective. Confocal z-stacks were processed with Leica LAS AF and Imaris v7.4.2 [46]. The colocalization between the SIP signal and TO-PRO-3-labeled bacterial structures was evaluated using the Imaris Coloc tool, with threshold adjustments made to define the colocalized voxels.
2.12. Ethical statement
All animal experiments were conducted in accordance with international ethical standards and Chilean Animal Protection Law 20.380. The procedures involving animals were reviewed and approved by the Institutional Animal Care and Use Committee of the Universidad de Santiago de Chile (protocol no. 795/2024; approved on December 30, 2024; amended on March 12, 2025). Four-week-old non-pregnant female C57BL/6 mice were obtained from the Central Animal Facility of the Universidad de Chile and transported to the Aquaculture Biotechnology Center at the Universidad de Santiago de Chile. The animals were quarantined for 21 days prior to experimentation and were kept under controlled conditions at 24 °C with a 12-hour light/dark cycle.
2.13. Oral administration, clinical monitoring, and preliminary immunogenicity
To evaluate the safety of oral administration of L. lactis-SIP and to identify an appropriate dose for future studies, a pilot study was conducted with five groups of female C57BL/6 mice (n = 5 per group). The study compared three doses of L. lactis-SIP (1 × 10¹², 1 × 10¹³, and 1 × 10¹⁴ CFU) against two control groups: sterile M17 culture medium and L. lactis NZ9000 (1 × 10¹³ CFU). Viable counts were confirmed by serial dilution and plating on M17 agar. Formulations were administered orally in 100 µL using a Teflon gavage catheter (Craberry, Ref. I016651422) on days 1, 14, 28, and 42, following the established L. lactis-pNZ8124-SIP model [31]. Mice were randomly assigned to the experimental groups. Clinical signs were evaluated using the Mouse Grimace Scale, and body weight was recorded every three days throughout the study.
Blood samples were collected at the specified time points, and serum SIP-specific IgG levels were quantified using ELISA. On day 49, the sera were titrated thinerial dilutions (1:25, 1:50, 1:100, 1:200, and 1:400). The optimal dose was defined as the one that produced the highest IgG response without increasing clinical scores or body weight loss. Seven days after the final administration (day 49), the animals were euthanized under deep anesthesia (ketamine at 120 mg/kg and xylazine at 10 mg/kg, administered intraperitoneally), followed by cervical dislocation in accordance with the approved protocol.
2.14. ELISA
Serum samples were collected at various time points during the immunization protocol and analyzed for SIP-specific IgG antibodies by indirect ELISA. In brief, 96-well plates were coated overnight at 37 °C with recombinant SIP (rSIP) at 2 µg/mL, diluted in 100 μL of 0.1 M carbonate-bicarbonate buffer (pH 9.6). The plates were then washed twice with phosphate-buffered saline containing 0.05% Tween-20 (PBS-T) and subsequently blocked for 1 hour at room temperature using a blocking buffer consisting of 1% w/v bovine serum albumin in PBS.
Serum samples were diluted in a blocking buffer and incubated for 1 hour at 37 °C. For the initial screening, the samples were analyzed at a dilution of 1:50. Subsequently, the samples underwent endpoint titration using serial dilutions ranging from 1:25 to 1:400. After washing, the plates were incubated for 1 hour at 37 °C with a horseradish peroxidase (HRP)-conjugated rat anti-mouse IgG secondary antibody at a dilution of 1:2000. Following three washing steps, the reaction was developed with tetramethylbenzidine (TMB) substrate for 15 minutes at 37 °C and then stopped by adding 2 N sulfuric acid. Absorbance was measured at 450 nm using a microplate reader.
Samples were deemed positive when their absorbance values exceeded the mean absorbance of the negative control group plus 2 standard deviations. Endpoint antibody titers were defined as the highest serum dilution that produced an absorbance value above the established positivity threshold.
2.15. Statistical analyses
Statistical analyses were conducted using GraphPad Prism 9 (version 9.4.1). Data distribution was assessed using the Shapiro-Wilk test. Comparisons of body weight outcomes and SIP-specific IgG levels among groups were made using the Kruskal-Wallis test, followed by Dunn’s post hoc test. Data are presented as mean ± standard deviation, unless stated otherwise, and a p-value of less than 0.05 was considered statistically significant.
3. Results
3.1. Generation and Characterization of L. lactis-SIP
3.1.1. pMBSacB-sip enabled targeted knock-in of the sip expression cassette at the upp locus
We previously reported that the expression of rSIP, induced by nisin in L. lactis NZ9000, was driven by the plasmid pNZ8124, which served as a prototype for an oral GBS vaccine [31]. Building on those findings, we developed a chromosomally modified L. lactis NZ9000 strain through homologous recombination with the plasmid pMBSacB-sip (Figure 1). The rSIP expression cassette was inserted into pMBSacB using the XhoI and BamHI restriction sites. Transformation and initial single-crossover selection were conducted at 40 °C, a temperature that prevents pMBSacB replication. PCR confirmation of the integration of pMBSacB-sip into the chromosome was performed using the primers pMBSacB_MCS_F and KI_lactis_R (Figure 2A). The cultures were then passed on to encourage plasmid resolution and the second crossover. Erythromycin-sensitive colonies were screened by PCR using the KI_lactis_F and KI_lactis_R primers. The expected amplicon sizes of 2,385 bp for L. lactis NZ9000 and 3,714 bp for L. lactis-SIP confirmed the successful replacement of the target upp region with the sip expression cassette (Figure 2B).
3.1.2. The sip cassette was successfully integrated into the target locus of the L. lactis NZ9000 genome
Whole-genome sequencing confirmed the integration of the GBS sip gene into the modified L. lactis subsp. Strain. cremoris (designated L. lactis-SIP). In contrast, the unmodified L. lactis NZ9000 strain showed no evidence of this gene. The assembly quality control results indicated that there were 59 contigs for L. lactis NZ9000 and 61 contigs for L. lactis-SIP (see Table S1).
IGV visualization showed consistent read coverage across the sip cassette in L. lactis-SIP, indicating a stable presence of the inserted sequence. In contrast, the corresponding sip reference region in the unmodified strain displayed no coverage (Figure 3).
BLASTn identified the 1,512 bp sip cassette exclusively in contig #2 of L. lactis-SIP, which has a total length of 196,082 bp. This contig showed 37× coverage and 100% identity over positions 193,890-195,401. The results from PCR, IGV, and BLASTn analyses collectively confirm the successful targeted integration of the GBS sip expression cassette into the genome of L. lactis subsp. cremoris. Our analytical pipeline did not detect any residual pMBSacB-associated resistance determinants or additional insertions.
3.1.3. Insertion of the rSIP expression cassette did not alter the in vitro growth of L. lactis-SIP
Due to the potential impact of chromosomal insertion on bacterial fitness, we compared the growth kinetics of L. lactis NZ9000 and L. lactis-SIP, both with and without nisin. Three independent assays were conducted for each condition. The growth curves showed low variability, and there were no significant differences between the modified and parental strains under the tested conditions (Figure 4A). These results suggest that the insertion of the rSIP expression cassette did not significantly impede in vitro growth.
3.1.4. L. lactis-SIP expresses rSIP in a nisin-dependent manner
Western blotting was used to assess rSIP expression following nisin induction. Total protein extracts from L. lactis-SIP cultures that were induced with nisin displayed an immunoreactive band at approximately 53 kDa, which aligns with the expected molecular weight of rSIP (see Figure 4B). In contrast, protein extracts from non-induced cultures did not reveal this band. These findings support the conclusion that rSIP expression is dependent on nisin induction at 10 ng/mL.
3.1.5. rSIP is localized on the bacterial surface of L. lactis-SIP
Confocal microscopy was utilized to investigate the detection and cellular localization of rSIP in L. lactis-SIP and control bacteria (Figure 5). The strain L. lactis NZ9000 showed no significant labeling associated with SIP, while non-induced L. lactis-SIP exhibited weak or negligible signals. In contrast, induced L. lactis-SIP showed clear rSIP labeling associated with bacterial cell structures. The signal varied among individual cells, as seen in the GBS positive control (Figure S1). Three-dimensional reconstruction further confirmed the localization of surface-associated rSIP in induced L. lactis-SIP (Figure 5B). These results suggest that nisin induction enhances rSIP expression and ite surface localization, although expression is not uniform across the bacterial population.
3.1.6. Pilot safety and preliminary immunogenicity after oral administration
Clinical scores assessed using the Mouse Grimace Scale remained at zero across all groups throughout the administration schedule, with no noted changes in behavior or facial expressions. Body weight changes did not differ significantly between mice receiving L. lactis-SIP at the evaluated doses and the control groups (Figure 6). SIP-specific IgG levels were measured using ELISA. On day 28, the group that received 1 × 10¹⁴ CFU showed a significant increase in serum SIP-specific IgG compared to the M17 control group (Figure 7A), while only one mouse in each of the 1 × 10¹² and 1 × 10¹³ CFU groups exhibited detectable IgG levels. By day 42, antibody levels in the 1 × 10¹² and 1 × 10¹³ CFU groups were like those of the control group. On day 49, the 1 × 10¹⁴ CFU group again demonstrated increased antibody levels, with three out of 5 mice showing a positive response (Figure 7B). These exploratory data suggest that the highest evaluated dose was tolerated and induced a measurable, though variable, systemic humoral response; however, they should not be interpreted as evidence of protective efficacy.
4. Discussion
Engineered lactic acid bacteria are increasingly being developed as live delivery vehicles for the mucosal delivery of antigens, therapeutic proteins, and immunomodulatory molecules. In this study, we created a marker-free strain of L. lactis subsp. cremoris NZ9000, which carries a chromosomally integrated cassette that encodes the GBS Surface Immunogenic Protein. This design offers improvements over the previously reported plasmid-based model (L. lactis-pNZ8124-SIP) by eliminating the need for antibiotic resistance selection and minimizing the presence of mobile genetic elements [31]. These enhancements are directly relevant to probiotic-based health applications and enhance the safety and regulatory compliance of functional food or live biotherapeutic platforms.
The pMBSacB plasmid was initially developed for site-specific mutagenesis in GBS [36] and has also been utilized in other streptococci. In L. lactis, SacB-based counterselection systems have allowed for markerless genome editing, including the deletion of chromosomal sequences [35,47]. This work further develops this concept by integrating a functional antigen-expression cassette at the upp locus. The construct combines the nisin-responsive PnisA promoter from the NICE system [48,49] with the Usp45 secretion signal, which is widely used for extracellular delivery in L. lactis [50,51]. Targeting the upp locus was supported by previous findings that disruption of upp can be tolerated in L. lactis [35]. Therefore, this strategy provides a solid framework for generating genetically stable, marker-free L. lactis strains suitable for heterologous protein delivery.
The allelic-exchange workflow required careful optimization. Linearizing the pMBSacB-sip plasmid prior to transformation improved the recovery of PCR-positive single-crossover candidates, consistent with the principle that linear DNA can enhance recombination-mediated integration [52]. The second crossover proved more challenging, likely because cointegrate resolution can revert to either the parental or the
desired mutant genotype [53,54]. To overcome this, we conducted serial passage without antibiotics, followed by sucrose counterselection, which allowed us to recover erythromycin-sensitive candidates. Subsequent PCR and whole-genome sequencing confirmed the successful targeted replacement of the upp region with the PnisA-Usp45-sip cassette and the loss of the pMBSacB backbone. Importantly, no additional insertions or residual resistance determinants were detected using our analytical pipeline. However, performing long-read sequencing and conducting long-term passage experiments would further strengthen our conclusions regarding genome architecture and stability.
Insertion of the rSIP cassette did not significantly affect in vitro growth, indicating that the chromosomal modification and nisin-induced expression did not impose a notable growth penalty under the tested conditions. Western blot analysis confirmed a nisin-dependent band corresponding to rSIP, and confocal microscopy demonstrated that rSIP was localized on the surface. This surface localization is important because it can enhance antigen accessibility to mucosal immune cells, potentially improving host-microbe immune interactions. However, expression of rSIP was uneven across the bacterial population, and the current data does not quantify the rSIP copy number per cell or the proportion of bacteria displaying it on their surface. Future studies should aim to quantify antigen abundance, secretion into the culture supernatant, surface retention, and the stability of expression across serial passages.
The observed phenotype aligns with the expected mechanism of the NICE system: nisin activates the NisK/NisR regulatory system, which induces transcription from PnisA and facilitates the synthesis of the Usp45-rSIP fusion protein [28,29,48,49]. However, this mechanism has been inferred based on cassette design and protein detection rather than directly tested. To strengthen this mechanistic interpretation, it would be beneficial to directly measure sip transcripts, compare induced and non-induced protein levels in both pellet and supernatant fractions, and evaluate the accessibility of rSIP on intact bacteria. This distinction is crucial for translational development, as factors like antigen dose, localization, and stability are key determinants of immunogenicity in live microbial delivery systems.
The pilot study on oral administration provides an initial indication of safety. Across the various doses evaluated, clinical scores remained 0, and there were no significant differences in body weight compared with control subjects, suggesting that L. lactis-SIP was well tolerated under the experimental conditions. These findings are consistent with the established safety profile of L. lactis as a food-grade organism and align with previous delivery models based on L. lactis. However, the safety evaluation remains preliminary as it was limited to short-term clinical observations and body weight measurements. To complete the biosafety assessment, additional endpoints should be considered, including gastrointestinal signs, histopathology, bacterial shedding, persistence, dissemination, microbiota impact, and genetic stability following gastrointestinal transit.
The immunogenicity data should be interpreted with caution. A measurable serum response of SIP-specific IgG was primarily observed at a dose of 1 × 10¹⁴ CFU, with only three out of five mice showing a positive response on day 49. The need for a higher dose compared to the previous plasmid-based L. lactis-pNZ8124-SIP model [31] may be due to several factors: a lower gene copy number in the chromosomal single-copy cassette, reduced antigen production, or differences in how the antigen behaves during gastrointestinal transit. While this explanation is plausible, it remains unverified without direct quantification of rSIP expression per CFU and comparison to the plasmid-based strain under similar conditions. Thus, the current results indicate feasibility rather than optimized immunogenicity.
From a functional food and health application perspective, formulation will be crucial for future development. Strategies that use microencapsulation with alginate and chitosan have been shown to enhance the survival of lactic acid bacteria during gastrointestinal transit and to improve the delivery of probiotic or vaccine payloads to intestinal sites [55,56,57]. Applying similar strategies to L. lactis-SIP could increase the viable dose that reaches gut-associated lymphoid tissue, decrease the total CFU required for an effective response, and enhance batch-to-batch consistency. This approach aligns well with the scope of the special issue, as it connects microbial engineering with delivery systems, stability, shelf life, and formulation for practical applications.
The chromosomal integration strategy outlined here may be applicable to other heterologous proteins of similar size, provided the expression, secretion, and surface association of each antigen are empirically validated. The main advantage of this approach is not only the expression of the SIP but also the development of a marker-free, chromosomally stable L. lactis chassis. This approach eliminates the need for antibiotic resistance markers and reduces concerns about plasmid-associated horizontal gene transfer. These features are particularly important for recombinant probiotics, live biotherapeutic products, and engineered functional food platforms, where biosafety and genetic stability are critical regulatory considerations [58].
Several limitations need to be addressed before making stronger biomedical claims. The study did not assess mucosal IgA, cellular immunity, cytokine profiles, protection against GBS colonization or challenges, the duration of the immune response, or reproductive and pregnancy models relevant to maternal GBS vaccination. Additionally, the sample size was small, and the study was exploratory, leading to variability in ELISA responses. While antigen expression was demonstrated qualitatively, it was not quantified. Although these limitations do not undermine the platform’s construction, they limit the conclusions to strain generation, microbiological characterization, preliminary tolerability, and exploratory systemic IgG induction.
The work presented represents a significant advancement in probiotic-based mucosal biomedicine by substituting a plasmid-dependent expression system with a marker-free chromosomal design. The next essential experiments should compare the chromosomal and plasmid-based strains side by side. It will be important to quantify the antigen output, assess formulation strategies like microencapsulation, and evaluate protective efficacy using a GBS colonization or infection model.
5. Conclusions
We developed and characterized a marker-free genetically modified strain of L. lactis subsp. cremoris NZ9000. This strain contains a chromosomally integrated PnisA-Usp45-sip expression cassette at the upp locus. We refer to the resulting strain as L. lactis-SIP. This strain maintained its in vitro growth, successfully expressed rSIP following nisin induction, and showed surface-associated localization of rSIP.
The pilot oral administration study in mice showed that the treatment was well-tolerated. Serum-specific IgG antibodies to SIP were primarily detected at the highest dose tested. This indicates that, while the approach is feasible, there is a need to optimize both antigen output and formulation. These results position L. lactis-SIP as a stable platform for the development of probiotic-based mucosal antigen delivery systems. Future research should concentrate on several areas: quantifying the antigen, ensuring long-term genetic stability, exploring microencapsulation or functional food formulation, and conducting mucosal and cellular immune profiling. Additionally, it is crucial to assess the protective efficacy against colonization or infection by GBS.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Quality-control report of genome assemblies generated using QUAST. Figure S1: SIP surface expression.
Author Contributions
Conceptualization, CG-R, BV, MI, JAS, and AEV; methodology, JG-G, BR-R, DAS, CC, CG-R, and BV; investigation, JG-G, BR-R, DAS, CC, CG-R, and BV; formal analysis, CC, MI, JAS, and AEV; data curation, CC, MI, JAS, and AEV; writing-original draft preparation, JG-G; writing-review and editing, JG-G, BR-R, CG-R, BV, MI, JAS, and AEV; supervision, MI, JAS, and AEV; funding acquisition, MI, JAS, and AEV. All authors have read and agreed to the submitted version of the manuscript.
Funding
This research was funded by IDeA Grant ID21I10370, supported by AEV; FONDECYT 1240741; and FONDECYT 1231866, which supported MI’s and JAS’s studies, respectively. Fellowships were awarded to JG-G (ANID N°21230817).
Data Availability Statement
Data generated or analyzed during this study are included in the article and Supplementary Materials. Raw sequencing files and additional information should be made available from the corresponding author upon reasonable request or deposited in a public repository before publication.
Acknowledgments
We thank Dr. Adam Ratner from the Grossman School of Medicine, New York, United States, for providing us with the pMBsacB plasmid. Additionally, we thank the Pontificia Universidad Católica de Valparaíso’s Institute of Biology for facilitating access to the Leica SP5 confocal microscope. We would like to express our gratitude to the staff of the Research and Innovation Subdepartment within the Health Surveillance and Research Department at the Public Health Institute of Chile for their valuable technical assistance in this research.
Conflicts of Interest
CG-R and BV are employed by Liva Company, and CG-R holds stock in the company. Liva Company contributed to the study’s conceptualization and manufacture of the L. lactis-SIP strain used in this research. The remaining authors declare no competing interests.
Abbreviations
The following abbreviations are used in this manuscript:
| APC | Antigen-Presenting Cells |
| bp | base pair |
| BHI | Brain Heart Infusion |
| CFU | colony-forming units |
| EOD | Early-Onset Disease |
| FDA | Food and Drug Administration |
| GBS | Group B Streptococcus |
| GALT | Gut-Associated Lymphoid Tissue |
| GMO | Genetically Modified Organism |
| GRAS | Generally Recognized as Safe |
| IAP | Intrapartum Antibiotic Prophylaxis |
| IGV | Integrative Genomics Viewer |
| LB | Luria Bertani |
| L. lactis | Lactococcus lactis |
| LOD | Late-Onset Disease |
| OD | optical density |
| ORF | Open Reading Frame |
| PFA | paraformaldehyde |
| rSIP | recombinant Surface Immunogenic Protein |
| SIP | Surface Immunogenic Protein |
| UMP | Uridine monophosphate |
| WHO | World Health Organization |
References
- B. Armistead, E. Oler, K. Adams Waldorf, and L. Rajagopal, “The Double Life of Group B Streptococcus: Asymptomatic Colonizer and Potent Pathogen,” Jul. 26, 2019, Academic Press. [CrossRef]
- S. Shabayek and B. Spellerberg, “Group B streptococcal colonization, molecular characteristics, and epidemiology,” Front. Microbiol., vol. 9, no. MAR, pp. 1–14, 2018. [CrossRef]
- American College of Obstetricians and Gynecologists, “Committee Opinion No. 797: Prevention of Group B Streptococcal Early-Onset Disease in Newborns: Correction,” Obstetrics & Gynecology, vol. 135, no. 4, pp. 978–979, Apr. 2020. [CrossRef]
- H. C. Slotved, F. Kong, L. Lambertsen, S. Sauer, and G. L. Gilbert, “Serotype IX, a proposed new Streptococcus agalactiae serotype,” J. Clin. Microbiol., vol. 45, no. 9, pp. 2929–2936, 2007. [CrossRef]
- N. J. Russell et al., “Risk of Early-Onset Neonatal Group B Streptococcal Disease with Maternal Colonization Worldwide: Systematic Review and Meta-analyses,” Clinical Infectious Diseases, vol. 65, no. Suppl 2, pp. S152–S159, 2017. [CrossRef]
- J. M. S. Pena, P. S. Lannes-Costa, and P. E. Nagao, “Vaccines for Streptococcus agalactiae: current status and future perspectives,” Front. Immunol., vol. 15, Jun. 2024. [CrossRef]
- S. Rioux, D. Martin, H. W. Ackermann, J. Dumont, J. Hamel, and B. R. Brodeur, “Localization of surface immunogenic protein on group B streptococcus,” Infect. Immun., vol. 69, no. 8, pp. 5162–5165, 2001. [CrossRef]
- D. Martin et al., “Protection from group B streptococcal infection in neonatal mice by maternal immunization with recombinant Sip protein,” Infect. Immun., vol. 70, no. 9, pp. 4897–4901, 2002. [CrossRef]
- B. R. Brodeur et al., “Identification of group B streptococcal sip protein, which elicits cross-protective immunity,” Infect. Immun., vol. 68, no. 10, pp. 5610–5618, 2000. [CrossRef]
- K. Firestone et al., “A CRISPRi library screen in group B Streptococcus identifies surface immunogenic protein (Sip) as a mediator of multiple host interactions,” Infect. Immun., vol. 93, no. 4, Apr. 2025. [CrossRef]
- D. A. Diaz-Dinamarca et al., “Surface immunogenic protein of streptococcus group b is an agonist of toll-like receptors 2 and 4 and a potential immune adjuvant,” Vaccines (Basel)., vol. 8, no. 1, Mar. 2020. [CrossRef]
- M. M. Mosaheb, M. L. Reiser, and L. M. Wetzler, “Toll-like receptor ligand-based vaccine adjuvants require intact MyD88 signaling in antigen-presenting cells for germinal center formation and antibody production,” Front. Immunol., vol. 8, no. MAR, 2017. [CrossRef]
- V. Gent et al., “Surface protein distribution in Group B Streptococcus isolates from South Africa and identifying vaccine targets through in silico analysis,” Sci. Rep., vol. 14, no. 1, p. 22665, Sep. 2024. [CrossRef]
- D. A. Diaz-Dinamarca et al., “Oral vaccine based on a surface immunogenic protein mixed with alum promotes a decrease in Streptococcus agalactiae vaginal colonization in a mouse model,” Mol. Immunol., vol. 103, pp. 63–70, Nov. 2018. [CrossRef]
- L. G. Bermúdez-Humarán, “Lactococcus lactis as a live vector for mucosal delivery of therapeutic proteins,” Hum. Vaccin., vol. 5, no. 4, pp. 264–267, 2009. [CrossRef]
- V. Chamcha, A. Jones, B. R. Quigley, J. R. Scott, and R. R. Amara, “Oral Immunization with a Recombinant Lactococcus lactis –Expressing HIV-1 Antigen on Group A Streptococcus Pilus Induces Strong Mucosal Immunity in the Gut,” The Journal of Immunology, vol. 195, no. 10, pp. 5025–5034, Nov. 2015. [CrossRef]
- X. Peng et al., “Production and delivery of Helicobacter pylori NapA in Lactococcus lactis and its protective efficacy and immune modulatory activity,” Sci. Rep., vol. 8, no. 1, p. 6435, Apr. 2018. [CrossRef]
- S. K. Murali and T. J. Mansell, “Next generation probiotics: Engineering live biotherapeutics,” Biotechnol. Adv., vol. 72, p. 108336, May 2024. [CrossRef]
- T. Ozdemir, A. J. H. Fedorec, T. Danino, and C. P. Barnes, “Synthetic Biology and Engineered Live Biotherapeutics: Toward Increasing System Complexity,” Cell Syst., vol. 7, no. 1, pp. 5–16, Jul. 2018. [CrossRef]
- R. R. C. New, “Formulation technologies for oral vaccines,” Clin. Exp. Immunol., vol. 198, no. 2, pp. 153–169, 2019. [CrossRef]
- J. L. Owen, B. Sahay, and M. Mohamadzadeh, “New generation of oral mucosal vaccines targeting dendritic cells,” Curr. Opin. Chem. Biol., vol. 17, no. 6, pp. 918–924, 2013. [CrossRef]
- J. E. Vela Ramirez, L. A. Sharpe, and N. A. Peppas, “Current state and challenges in developing oral vaccines,” Adv. Drug Deliv. Rev., vol. 114, pp. 116–131, 2017. [CrossRef]
- U.S. Food and Drug Administration, “Early Clinical Trials with Live Biotherapeutic Products: Chemistry, Manufacturing, and Control Information. Guidance for Industry,” Silver Spring, MD, Sep. 2003. Accessed: Nov. 11, 2025. [Online]. Available: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/early-clinical-trials-live-biotherapeutic-products-chemistry-manufacturing-and-control-information.
- A. Bolotin et al., “The Complete Genome Sequence of the Lactic Acid Bacterium Lactococcus lactis ssp. lactis IL1403,” Genome Res., vol. 11, no. 5, pp. 731–753, May 2001. [CrossRef]
- D. M. Linares, J. Kok, and B. Poolman, “Genome Sequences of Lactococcus lactis MG1363 (Revised) and NZ9000 and Comparative Physiological Studies,” J. Bacteriol., vol. 192, no. 21, pp. 5806–5812, Nov. 2010. [CrossRef]
- K. H. Schleifer, J. Kraus, C. Dvorak, R. Kilpper-Bälz, M. D. Collins, and W. Fischer, “Transfer of Streptococcus lactis and Related Streptococci to the Genus Lactococcus gen. nov.,” Syst. Appl. Microbiol., vol. 6, no. 2, pp. 183–195, Sep. 1985. [CrossRef]
- M. J. Gasson, “Plasmid complements of Streptococcus lactis NCDO 712 and other lactic streptococci after protoplast-induced curing,” J. Bacteriol., vol. 154, no. 1, pp. 1–9, Apr. 1983. [CrossRef]
- O. P. Kuipers, P. G. G. A. de Ruyter, M. Kleerebezem, and W. M. de Vos, “Quorum sensing-controlled gene expression in lactic acid bacteria,” J. Biotechnol., vol. 64, no. 1, pp. 15–21, Sep. 1998. [CrossRef]
- O. P. Kuipers, M. M. Beerthuyzen, R. J. Siezen, and W. M. de Vos, “Characterization of the nisin gene cluster nisABTCIPR of Lactococcus lactis,” Eur. J. Biochem., vol. 216, no. 1, pp. 281–291, Aug. 1993. [CrossRef]
- Food and Agriculture Organization of the United Nations and World Health Organization, “Probiotics in food: Health and nutritional properties and guidelines for evaluation,” Rome, 2006. Accessed: Nov. 11, 2025. [Online]. Available: https://openknowledge.fao.org/items/1c836817-9be2-4256-83ae-0b5055039b50.
- D. A. Diaz-Dinamarca et al., “Mucosal Vaccination with Lactococcus lactis-Secreting Surface Immunological Protein Induces Humoral and Cellular Immune Protection against Group B Streptococcus in a Murine Model,” Vaccines (Basel)., vol. 8, no. 2, p. 146, Mar. 2020. [CrossRef]
- World Health Organization, “Immunization, Vaccines and Biologicals,” Group B Streptococcus (GBS). Accessed: Nov. 11, 2025. [Online]. Available: https://www.who.int/teams/immunization-vaccines-and-biologicals/diseases/group-b-streptococcus-(gbs).
- World Health Organization, “Annex 1 WHO guidelines on nonclinical,” no. 927, 2005, Accessed: Nov. 11, 2025. [Online]. Available: https://www.who.int/publications/m/item/annex1-nonclinical.p31-63.
- J. Martinussen and K. Hammer, “Cloning and characterization of upp, a gene encoding uracil phosphoribosyltransferase from Lactococcus lactis,” J. Bacteriol., vol. 176, no. 21, pp. 6457–6463, Nov. 1994. [CrossRef]
- L. Song et al., “Construction of upp deletion mutant strains of Lactobacillus casei and Lactococcus lactis based on counterselective system using temperature-sensitive plasmid,” J. Microbiol. Methods, vol. 102, pp. 37–44, Jul. 2014. [CrossRef]
- T. A. Hooven, M. Bonakdar, A. B. Chamby, and A. J. Ratner, “A counterselectable sucrose sensitivity marker permits efficient and flexible mutagenesis in streptococcus agalactiae,” Appl. Environ. Microbiol., vol. 85, no. 7, Apr. 2019. [CrossRef]
- MoBiTec GmbH, “NICE expression system: Handbook for Lactococcus lactis expression system,” Goettingen, Mar. 2015. Accessed: Nov. 11, 2025. [Online]. Available: https://www.mobitec.com/media/mobitec/old_content/NICE_Expression_System-Handbook.pdf.
- S. Andrews, “FastQC: A quality control tool for high throughput sequence data.,” Babraham Bioinformatics, Babraham Institute. Accessed: Dec. 03, 2025. [Online]. Available: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- A. Bankevich et al., “SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing,” Journal of Computational Biology, vol. 19, no. 5, pp. 455–477, May 2012. [CrossRef]
- A. Gurevich, V. Saveliev, N. Vyahhi, and G. Tesler, “QUAST: quality assessment tool for genome assemblies,” Bioinformatics, vol. 29, no. 8, pp. 1072–1075, Apr. 2013. [CrossRef]
- S. F. Altschul, W. Gish, W. Miller, E. W. Myers, and D. J. Lipman, “Basic local alignment search tool,” J. Mol. Biol., vol. 215, no. 3, pp. 403–410, Oct. 1990. [CrossRef]
- H. Li and R. Durbin, “Fast and accurate short read alignment with Burrows–Wheeler transform,” Bioinformatics, vol. 25, no. 14, pp. 1754–1760, Jul. 2009. [CrossRef]
- J. T. Robinson et al., “Integrative genomics viewer,” Nat. Biotechnol., vol. 29, no. 1, pp. 24–26, Jan. 2011. [CrossRef]
- A. R. Quinlan and I. M. Hall, “BEDTools: a flexible suite of utilities for comparing genomic features,” Bioinformatics, vol. 26, no. 6, pp. 841–842, Mar. 2010. [CrossRef]
- D. A. Díaz-Dinamarca et al., “The Optimisation of the Expression of Recombinant Surface Immunogenic Protein of Group B Streptococcus in Escherichia coli by Response Surface Methodology Improves Humoral Immunity,” Mol. Biotechnol., vol. 60, no. 3, pp. 215–225, Mar. 2018. [CrossRef]
- E. Vicencio et al., “Aggregatibacter actinomycetemcomitans Induces Autophagy in Human Junctional Epithelium Keratinocytes,” Cells, vol. 9, no. 5, p. 1221, May 2020. [CrossRef]
- Y. Xin, D. Guo, Y. Nan, P. Yang, and M. Qiao, “An efficient platform for markerless large-scale genomic deletions in Lactococcus lactis,” Food Biosci., vol. 70, p. 107057, Aug. 2025. [CrossRef]
- I. Mierau and M. Kleerebezem, “10 years of the nisin-controlled gene expression system (NICE) in Lactococcus lactis,” Appl. Microbiol. Biotechnol., vol. 68, no. 6, pp. 705–717, Oct. 2005. [CrossRef]
- I. Mierau, K. Olieman, J. Mond, and E. J. Smid, “Optimization of the Lactococcus lactis nisin-controlled gene expression system NICE for industrial applications,” Microb. Cell Fact., vol. 4, no. 1, p. 16, Dec. 2005. [CrossRef]
- Y. Le Loir, S. Nouaille, J. Commissaire, L. Brétigny, A. Gruss, and P. Langella, “Signal Peptide and Propeptide Optimization for Heterologous Protein Secretion in Lactococcus lactis,” Appl. Environ. Microbiol., vol. 67, no. 9, pp. 4119–4127, Sep. 2001. [CrossRef]
- Y. Le Loir et al., “Protein secretion in Lactococcus lactis: an efficient way to increase the overall heterologous protein production,” Microb. Cell Fact., vol. 4, no. 1, p. 2, Jan. 2005. [CrossRef]
- E. C. Conley and J. R. Saunders, “Recombination-dependent recircularization of linearized pBR322 plasmid DNA following transformation of Escherichia coli,” Mol. Gen. Genet., vol. 194, no. 1–2, pp. 211–218, Apr. 1984. [CrossRef]
- G. S. Tamura, D. S. Bratt, H. H. Yim, and A. Nittayajarn, “Use of glnQ as a Counterselectable Marker for Creation of Allelic Exchange Mutations in Group B Streptococci,” Appl. Environ. Microbiol., vol. 71, no. 1, pp. 587–590, Jan. 2005. [CrossRef]
- E. Maguin, H. Prévost, S. D. Ehrlich, and A. Gruss, “Efficient insertional mutagenesis in lactococci and other gram-positive bacteria,” J. Bacteriol., vol. 178, no. 3, pp. 931–935, Feb. 1996. [CrossRef]
- J. B. Divya and K. M. Nampoothiri, “Encapsulated Lactococcus lactis with enhanced gastrointestinal survival for the development of folate enriched functional foods,” Bioresour. Technol., vol. 188, pp. 226–230, Jul. 2015. [CrossRef]
- X. Wang et al., “Microencapsulating Alginate-Based Polymers for Probiotics Delivery Systems and Their Application,” Pharmaceuticals, vol. 15, no. 5, p. 644, May 2022. [CrossRef]
- T. Jiang et al., “Oral delivery of probiotic expressing M cell homing peptide conjugated BmpB vaccine encapsulated into alginate/chitosan/alginate microcapsules,” European Journal of Pharmaceutics and Biopharmaceutics, vol. 88, no. 3, pp. 768–777, Nov. 2014. [CrossRef]
- J. Grossolli-Galvez, M. Imarai, J. A. Soto, and A. E. Vasquez, “Lactococcus lactis as a New Strategy for Oral Vaccination: Current Insights and Future Perspectives,” Pharmaceutics, vol. 18, no. 3, p. 307, Feb. 2026. [CrossRef]
Figure 1.
Schematic representation of the allelic-exchange strategy used to generate the L. lactis-SIP strain. The pMBSacB-SIP plasmid includes the sip expression cassette (in red), homologous arms (HA) that flank the target region of the upp locus, an erythromycin resistance marker for positive selection, and SacB for sucrose counterselection. After transformation, first-crossover intermediates are selected using erythromycin at a non-permissive temperature. Plasmid resolution and the second crossover are facilitated by serial passages without antibiotics, followed by sucrose counterselection. Candidate colonies are screened for erythromycin sensitivity and confirmed through PCR and whole-genome sequencing.
Figure 1.
Schematic representation of the allelic-exchange strategy used to generate the L. lactis-SIP strain. The pMBSacB-SIP plasmid includes the sip expression cassette (in red), homologous arms (HA) that flank the target region of the upp locus, an erythromycin resistance marker for positive selection, and SacB for sucrose counterselection. After transformation, first-crossover intermediates are selected using erythromycin at a non-permissive temperature. Plasmid resolution and the second crossover are facilitated by serial passages without antibiotics, followed by sucrose counterselection. Candidate colonies are screened for erythromycin sensitivity and confirmed through PCR and whole-genome sequencing.

Figure 2.
illustrates the integration of the sip cassette into the upp locus through allelic exchange. A) PCR amplification of transformant colonies from the first step was conducted using the KI_lactis_R and pMBSacB_MCS_F primers. The molecular weight (MW) marker used was Lambda/HindIII DNA. Lanes 1-10 show transformant colonies that tested positive for the single-crossover intermediate. Lane 11 serves as a negative control, lane 12 contains pMBSacB-sip DNA, lane 13 has L. lactis NZ9000 DNA, and lane 14 is a no-template control. B) Confirmation of the double-crossover event was performed using the KI_lactis_F and KI_lactis_R primers. The MW marker for this section was a 1 kb molecular weight marker. Lane 1 is a no-template control, lane 2 contains DNA from the L. lactis-SIP mutant candidate, and lane 3 contains DNA from L. lactis NZ9000.
Figure 2.
illustrates the integration of the sip cassette into the upp locus through allelic exchange. A) PCR amplification of transformant colonies from the first step was conducted using the KI_lactis_R and pMBSacB_MCS_F primers. The molecular weight (MW) marker used was Lambda/HindIII DNA. Lanes 1-10 show transformant colonies that tested positive for the single-crossover intermediate. Lane 11 serves as a negative control, lane 12 contains pMBSacB-sip DNA, lane 13 has L. lactis NZ9000 DNA, and lane 14 is a no-template control. B) Confirmation of the double-crossover event was performed using the KI_lactis_F and KI_lactis_R primers. The MW marker for this section was a 1 kb molecular weight marker. Lane 1 is a no-template control, lane 2 contains DNA from the L. lactis-SIP mutant candidate, and lane 3 contains DNA from L. lactis NZ9000.

Figure 3.
Visualization of the sip gene from the GBS (Group B Streptococcus) in L. lactis subsp. cremoris using IGV (Integrated Genome Viewer). The top panel displays the nucleotide positions of the sip reference sequence (GenBank accession DQ914270.1). The colored panel shows whole-genome sequencing reads mapped to the sip reference sequence, colored by read strand, for both the unmodified strain (L. lactis NZ9000) and the modified strain (L. lactis-SIP). The gray panel represents a read-coverage track that indicates the depth of coverage across the region. The bottom track provides the annotation for the sip gene.
Figure 3.
Visualization of the sip gene from the GBS (Group B Streptococcus) in L. lactis subsp. cremoris using IGV (Integrated Genome Viewer). The top panel displays the nucleotide positions of the sip reference sequence (GenBank accession DQ914270.1). The colored panel shows whole-genome sequencing reads mapped to the sip reference sequence, colored by read strand, for both the unmodified strain (L. lactis NZ9000) and the modified strain (L. lactis-SIP). The gray panel represents a read-coverage track that indicates the depth of coverage across the region. The bottom track provides the annotation for the sip gene.

Figure 4.
Effect of the SIP cassette on bacterial growth and rSIP expression. A) Growth kinetics of L. lactis NZ9000 and L. lactis-SIP strains with and without nisin (10 ng/mL). B) Western blot analysis showing rSIP detection after 1 hour induction with nisin (10 ng/mL).
Figure 4.
Effect of the SIP cassette on bacterial growth and rSIP expression. A) Growth kinetics of L. lactis NZ9000 and L. lactis-SIP strains with and without nisin (10 ng/mL). B) Western blot analysis showing rSIP detection after 1 hour induction with nisin (10 ng/mL).

Figure 5.
rSIP Immunodetection by Confocal Microscopy. Bacteria in the exponential growth phase were fixed onto glass slides and labeled using a rabbit polyclonal anti-rSIP antibody (green) and TO-PRO-3 iodide to stain DNA-rich structures (red). The third column displays a digital overlay of the green and red channels, while the fourth column shows colocalized voxels in yellow. Images were captured using confocal microscopy with a 100× oil-immersion objective. GBS served as a positive control for SIP expression, and Lactis-SIP induced by GBS exhibited surface-associated rSIP labeling. The scale bar indicates 5 μm, and the blue square marks the area that is magnified in Figure 5B. Three-dimensional reconstruction was performed using Imaris software to highlight the localization of rSIP. The light blue arrowhead points to a bacterium with intense surface-associated labeling, shown with a scale bar of 2 μm.
Figure 5.
rSIP Immunodetection by Confocal Microscopy. Bacteria in the exponential growth phase were fixed onto glass slides and labeled using a rabbit polyclonal anti-rSIP antibody (green) and TO-PRO-3 iodide to stain DNA-rich structures (red). The third column displays a digital overlay of the green and red channels, while the fourth column shows colocalized voxels in yellow. Images were captured using confocal microscopy with a 100× oil-immersion objective. GBS served as a positive control for SIP expression, and Lactis-SIP induced by GBS exhibited surface-associated rSIP labeling. The scale bar indicates 5 μm, and the blue square marks the area that is magnified in Figure 5B. Three-dimensional reconstruction was performed using Imaris software to highlight the localization of rSIP. The light blue arrowhead points to a bacterium with intense surface-associated labeling, shown with a scale bar of 2 μm.

Figure 6.
Body weight monitoring following oral administration. A) Body weight measurements were taken throughout the administration period. B) The final body weight recorded on day 49 showed no significant differences between the experimental groups. Statistical analyses were performed among groups using the Kruskal-Wallis test followed by Dunn’s post hoc test. Bars represent the mean ± SEM, while dots indicate individual animals.
Figure 6.
Body weight monitoring following oral administration. A) Body weight measurements were taken throughout the administration period. B) The final body weight recorded on day 49 showed no significant differences between the experimental groups. Statistical analyses were performed among groups using the Kruskal-Wallis test followed by Dunn’s post hoc test. Bars represent the mean ± SEM, while dots indicate individual animals.

Figure 7.
Serum Levels of SIP-Specific IgG. This figure shows the serum levels of SIP-specific IgG across different experimental groups, each identified by a specific color: the M17 control group is represented in black, L. lactis NZ9000 in green, and varying concentrations of L. lactis-SIP at 1 × 10¹² CFU (red), 1 × 10¹³ CFU (blue), and 1 × 10¹⁴ CFU (orange). The red dashed line indicates the days of administration. Panel A) presents IgG levels at various time points throughout the administration schedule. Panel B) displays antibody titers measured on day 49, which is seven days after the final administration.
Figure 7.
Serum Levels of SIP-Specific IgG. This figure shows the serum levels of SIP-specific IgG across different experimental groups, each identified by a specific color: the M17 control group is represented in black, L. lactis NZ9000 in green, and varying concentrations of L. lactis-SIP at 1 × 10¹² CFU (red), 1 × 10¹³ CFU (blue), and 1 × 10¹⁴ CFU (orange). The red dashed line indicates the days of administration. Panel A) presents IgG levels at various time points throughout the administration schedule. Panel B) displays antibody titers measured on day 49, which is seven days after the final administration.


Table 1.
Bacterial strains and plasmids used in this study.
| Strain/plasmid | Characteristics | Source |
|---|---|---|
| E. coli DH5α | Chemically competent cloning host used to maintain recombinant plasmids | Lab collection |
| L. lactis NZ9000 | Derivative of MG1363; pepN::nisRK | Lab collection |
| L. lactis-SIP | Derivative of L. lactis NZ9000; Δupp::PnisA-Usp45-sip | This study |
| pUC57-sip | pUC57 plasmid carrying the synthesized upp-targeting recombination cassette | GenScript |
| pMBsacB | Temperature-sensitive pHY304-derived allelic-exchange vector carrying erythromycin resistance and sacB for sucrose counterselection | [36] |
| pMBsacB-sip | pMBSacB carrying the upp-targeting PnisA-Usp45-sip recombination cassette cloned between XhoI and BamHI sites | This study |
Table 2.
Oligonucleotides Used in This Study.
| Primer/probe | Sequence (5’- 3’) | Source |
|---|---|---|
| KI_lactis_F | ATGGAGGGTCTGGGACTCAT | This study |
| KI_lactis_R | GCGACTTTTCCTCTGTAGGAACT | This study |
| pMBsacB_MCS_F | CAATACGCAAACCGCCTCTC | [36] |
| T7 promoter | TAATACGACTCACTATAGGG | Invitrogen |
| M13 reverse | CAGGAAACAGCTATGAC | Invitrogen |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



