Submitted:
03 June 2026
Posted:
05 June 2026
You are already at the latest version
Abstract
This comprehensive literature review delves into the multifaceted roles of RNA-binding motif protein 3 (RBM3) in cellular processes, disease pathogenesis, and therapeutic poten-tial. RBM3 has been implicated in shaping cell morphology, synaptic protection in neuro-degenerative conditions, and regulating gene expression through binding to specific RNA sequences. In cancer, RBM3 exhibits contrasting effects, influencing cell proliferation, tu-morigenic potential, and RNA splicing. Clinical studies suggest RBM3 as a predictive bi-omarker in chemotherapy response for muscle-invasive bladder cancer. Despite promising therapeutic implications in neuroprotection and cancer, challenges persist in under-standing the regulatory mechanisms and clinical behavior of RBM3. Further research is warranted to elucidate the molecular mechanisms underlying RBM3's diverse functions and its significance as a potential target for personalized medicine in cancer therapy. This review underscores the pivotal role of RBPs, particularly RBM3, in disease progression and highlights the need for continued investigation to harness their therapeutic potential effectively.
Keywords:
RNA binding motif 3 (RBM3)
; therapeutic target
; cancer biomarkers
; cell stress response
; prognostic indicators
1. Introduction
RNA-binding proteins (RBPs), as their name implies, represent a category of proteins that create complexes with various nucleic acids, including microRNAs, messenger RNAs, small inducible RNAs, small nuclear RNAs, small nucleolar RNAs, transfer RNAs, and noncoding RNAs [1]. Moreover, they may sporadically bind to single-stranded DNA [2]. It is estimated that 7.5% of the human protein-coding genome is comprised of RBPs [1]. As shown in Figure 1, these proteins are crucial in numerous cellular processes such as pre-mRNA splicing, editing, polyadenylation, localization, turnover, translation, and degradation [3]. Due to their significant roles in cellular functions, RBPs are subject to strict regulation, and any dysregulation, mutations, or mislocalization can lead to various diseases. Notably, RBPs like TDP-43, Matrin-3, and FUS are suspected to be associated with amyotrophic lateral sclerosis (ALS), a severe neurodegenerative disorder [4,5]. Additionally, RBPs have been implicated in Alzheimer’s disease, Parkinson’s disease, different types of cancer, and cardiovascular conditions [6,7,8,9]. Recently, RBPs have emerged as promising targets for therapeutic interventions, sparking renewed interest in uncovering the underlying molecular mechanisms governing their functions [10].
RNA-binding proteins engage with nucleic acids through one or more domains. Various types of domains are responsible for recognizing RNA, such as the RNA-binding domain (RBD), also referred to as the ribonucleoprotein (RNP) domain or the RNA recognition motif (RRM), the double-stranded RNA-binding domain (dsRBD), the K-homology (KH) domain (type I and type II), the zinc finger (ZnF, predominantly C-x8-X-x5-X-x3-H), the Sm domain, the DEAD/DEAH box, the cold-shock domain, the Pumilio/FBF (PUF or Pum-HD) domain, and the Piwi/Argonaute/Zwille (PAZ) domain [1]. A single domain of this kind has the capacity to bind 2–6 nucleotides. The presence of multiple copies of these domains enhances the binding of more intricate nucleotide sequences and contributes to the specificity of the sequence. The RRM domain is notably prevalent in more complex organisms [11]. Alongside these structured domains, glycine–arginine-rich (GAR/RGG/RG/GRG) motifs are frequently identified as supplementary regions in numerous human RBPs. These motifs are inherently disordered, have an affinity for RNA, and have been associated with various disorders, including neurodegenerative and neuromuscular conditions, as well as cancer [12,13,14].
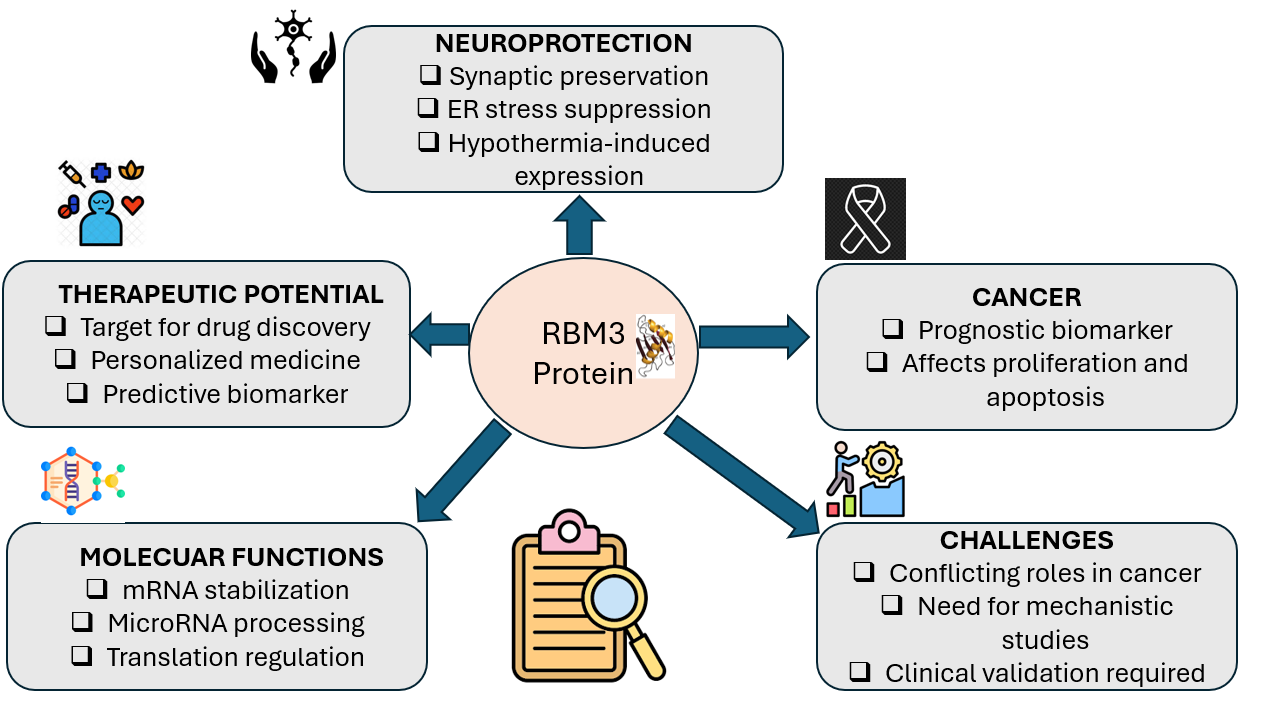
The RNA-binding motif protein 3 (RBM3) is part of the glycine–arginine (GR)-rich RBP family protein, characterized by having one N-terminal RNA-binding domain (RRM) and a highly flexible C-terminal GR-rich region [15]. Due to the presence of a single RRM domain, RBM3 falls within the class IVa-GRP subfamily, whose constituents may harbor up to three RRM domains. RBM3 gene is coded on the X chromosome [16], and its upregulation has been documented in response to cold stress conditions [17].
Utilizing cDNA selection with a YAC originating from the Xp11.2 locus, a novel gene (RBM3) has been discovered, which codes for a polypeptide exhibiting significant sequence homology to a cluster of proteins known to interact with RNA. Positioned on a YAC contig map, RBM3 is situated amidst OATL1 and GATA1/TFE3 in sub-band Xp11.23 and generates alternatively spliced transcripts across various human tissues. The primary open reading frame encompasses a 157 amino acid protein with an anticipated molecular weight of 17 kDa. The presumed RNA-binding domain of RBM3 bears strong resemblance to that of two previously identified human RNA-binding proteins - YRRM, associated with azoospermia, and hnRNP G, a glycoprotein recognized as an autoantigen. The likeness of RBM3 to an RNA binding protein from maize, AAIP, in terms of both sequence and size, indicates its potential involvement in a fundamental pathway conserved from plants to mammals [18].
Previously identified as a cold-shock protein, recent research has suggested that RBM3 also has cytoprotective functions in various stress scenarios, such as hypoxia [19] and infection-induced fever [20]. Within the realm of cellular processes, RBM3 exerts significant influence on the foundational translational apparatus, impacting overall protein synthesis, cell proliferation, and functioning as a regulator of programmed cell death. Moreover, RBM3 is associated with tumorigenesis, cancer spread, and neuroprotection [16].
Importance of RBM3 in Disease Progression
I. Molecular functions of RBM3 in Disease Pathogenesis
As stated earlier, RBM3 belongs to a small family of cold-inducible RNA-binding proteins and is involved in regulating various aspects of mRNA metabolism and possesses pleiotropic functions in cell stress, development, and oncogenesis [17,18,21]. It has been shown that at a molecular level, RBM3 facilitates global protein synthesis, enhances the stability of mRNAs containing AU-rich elements, and aids in the production of numerous microRNAs during the Dicer step [22]. These functions collectively suggest that RBM3 exerts a wide-ranging and distinct regulatory impact on the proteome. At a cellular level, initial investigations have suggested that RBM3 plays a crucial role in adaptive responses to hypothermia, potentially functioning as an mRNA chaperone that preserves translation capability until euthermic conditions are restored [23,24,25,26]. However, it has become evident that RBM3 is triggered by a diverse array of physiological stresses, including hypoxia, endoplasmic reticulum (ER) stress, excitotoxins, radiation, wasting, and certain pathological conditions [16,19,27,28]. Within these various scenarios, RBM3 exerts a potent cytoprotective effect. Within muscle cells, the induction of RBM3 opposes necrosis and apoptosis, and maintains cellular structure in the face of reactive oxygen species and conditions leading to cellular atrophy [29]. In neuronal cells, RBM3 serves as a shield against cell death brought on by endoplasmic reticulum (ER) stress, hypoxia/ischemia, as well as a range of other metabolic and disease-related stresses [16,19,30,31,32,33,34,35,36] 3. These findings suggest a broad function of RBM3 in cell protection. Supporting this idea, RBM3 is the sole transcript that is upregulated across all tissues during torpor, a condition characterized by prolonged hypothermia that spares many tissues, including muscle and brain, from the adverse effects of severe metabolic stress [37]. Apart from being influenced by physiological stressors, the expression of RBM3 displays variations in different brain regions and stages of development and has been implicated in oncogenesis as a potential proto-oncogene and a predictor of the clinical outcome in various cancers [38,39].
Multiple observations indicate that RBM3 possesses morpho-regulatory functions that are pertinent to its involvement in cell protection and significant contribution to both development and oncogenesis [40]. The protein RBM3 (P98179, RNPL) was included in the group of RNA-binding proteins that were found in the invasive pseudopodia of mesenchymal breast cancer cells. Additionally, RBM3 was present in specialized cellular regions known as spreading initiation centers (SICs) that emerge during the initial phases of cell spreading in fibroblasts and various mesenchymal cell lines [40,41,42]. Various categories of RNA-binding proteins (RNA-BPs) have been recognized within SICs and other cellular extensions, such as podosomes, pseudopodia, filopodia, growth cones, and dendritic spines [43,44,45,46,47]. The functions performed at these specific locations are diverse, encompassing the regulation of mRNA positioning and translation processes that facilitate cellular adhesion, expansion, movement, and the direction of projections, along with interactions between proteins that control the dynamic nature of the cytoskeleton. Considering this information, the presence of RBM3 within stress-induced cytoplasmic inclusions (SICs) and invading filopodia implies its involvement in shaping cell morphology and movement, potentially crucial for its stress response mechanisms and speculated role in both embryonic development and oncogenesis [47,48,49,50,51,52]. Studies have shown that the induction of RBM3 provides synaptic protection in experimental models of prion and Alzheimer’s-related neurodegeneration. Conversely, the depletion of RBM3 worsens the loss of synapses and the inadequacies in synapse regeneration in these models [53,54]. This protective effect of RBM3 on synapses, which might be linked mechanistically to the capacity for synaptic regeneration observed during torpor and cold shock, lends support to the notion that RBM3 may play a role in controlling the development and adaptability of cellular projections. Collectively, these data imply that RBM3 plays critical roles in the regulation of cellular morphology and migration, although this aspect has not yet been fully clarified [55,56].
J. Pilotte et al. provided evidence that RBM3 functions as a controller of cell spreading, polarity, and migration [57]. They also observed that RBM3 is present in various types of cell protrusions found in different cell lines and primary myoblasts. Altering RBM3 expression leads to significant modifications in cell polarity and spreading through the involvement of the RhoA-Rho associated protein kinase (ROCK) signaling pathway and the collapsin response mediator protein 2 (CRMP2). Their findings suggested that these effects may be linked to mechanisms that regulate cell motility. Also, they indicated that RBM3 facilitates directed cell migration in different scenarios by adopting a mesenchymal-like mode characterized by the extension of elongated protrusions. Conversely, reduced levels of RBM3 hinder migration and the ability of cells to elongate protrusions, prompting a shift towards a migration mode reminiscent of amoeboid motion [57].
Figure 1.
The regulation of gene expression after transcription, influenced by RNA-binding proteins. A. The RRM domain and RGG domain of RBM3 are shown. B. The posttranscriptional regulation is controlled by RNA-binding proteins. Reprinted with permission from [58].
Figure 1.
The regulation of gene expression after transcription, influenced by RNA-binding proteins. A. The RRM domain and RGG domain of RBM3 are shown. B. The posttranscriptional regulation is controlled by RNA-binding proteins. Reprinted with permission from [58].

II. Importance of RBM3 in Neuroprotection
The upregulation of Cold-inducible RNA-binding protein (CRIP) via mild hypothermia results in the inhibition of the mitochondrial apoptosis pathway. Consequently, this leads to a reduction in neuronal apoptosis, resulting in safeguarding the neurons from degeneration and subsequent apoptosis. These results validate the advantageous effects of employing mild hypothermia therapy for brain injury [59]. Recently, researchers have demonstrated the involvement of RBM3 in facilitating neuroprotection induced by hypothermia. It has been shown that a high level of RBM3 expression is associated with the development of the neurons of the murine brain. Also, mild hypothermia at 32 °C has demonstrated the capacity to significantly increase RBM3 mRNA and protein levels in both mature and young neurons. Moreover, the enhancement of the expression of RBM3 has been linked to a decrease in the degree of neuronal cell death. Furthermore, the protective impact of hypothermia has been compromised as a result of downregulation of RBM3 expression. Again, studies have demonstrated that neuronal cells acquire greater resistance to cell death by stimulating RBM3 even under normal body temperature conditions (37 °C) [60]. Peretti et al. 2015, found that upregulation of RBM3 resulted in sustained preservation of synapses during the progression of prion disease, which led to a notable extension of lifespan and mitigation of cognitive and neuronal impairments [61]. Similarly, another group reported that RBM3 provides neuroprotection by suppressing the ER stress pathway involving PERK, eIF2α, and CHOP as shown in Figure 2. In contrast, downregulation of RBM3 was linked to synaptic degeneration, negating the neuroprotective benefits of hypothermia and hastening disease progression. Overall, a potential shielding role associated with enhancing cold-stress pathways in neurodegenerative conditions was identified [62]. Furthermore, another study indicated that RBM3 modulates cell proliferation and apoptosis following spinal cord trauma [63]. The neuroprotective effect of hypothermia may involve several pathophysiological mechanisms, including an enhancement in protein misfolding and denaturation, a decrease in cell cycle progression, a suppression of translation and transcription processes, a perturbation in cellular cytoskeletal components, and changes in membrane permeability. These alterations ultimately result in elevated cytosolic levels of Na+ and H+ ions [64]. It has been shown that mild to moderate hypothermia has enhanced the expression of RBM3 [65], whereas elevated temperatures have been found to impede the expression of RBM3 [66] . A study by Chip et al. (2011) [60] has indicated that mild to moderate hypothermia at 32˚C can stimulate the induction of RBM3. The upregulation of RNA-binding motif protein 3 (RMB3) is particularly evident in specific regions such as juvenile and mature neurons under conditions of lower temperatures [60,67]. The neuroprotective efficacy of hypothermia has waned due to the suppression of RBM3 expression. However, upregulation of RBM3 has been shown to mitigate the cleavage of poly ADP ribose polymerase (PARP), prevent intranucleosomal DNA fragmentation, and inhibit the release of Lactic acid dehydrogenase (LDH) in the absence of hypothermia. These findings are consistent with a recent study indicating the crucial involvement of RBM3 in the neuroprotective mechanisms induced by hypothermia [68]. RBM3 is important in facilitating translation within neuronal cells. Moreover, RBM3 plays a crucial role in providing neuroprotection in primary neurons, PC12 cells (a type of catecholamine cell that synthesize, store and release norepinephrine and dopamine), and cortical organotypic slice cultures when exposed to hypothermic conditions [69].
III. The expression of RBM3 in cancer
RNA-binding proteins (RBPs) are responsible for governing multiple stages of protein synthesis and the regulation of post-transcriptional gene expression; therefore, they represent an important area of focus within the field of cancer research. RBPs have been demonstrated to fulfill a significant role in RNA metabolism by overseeing processes such as RNA splicing, transportation, monitoring, degradation, and translation. Disruptions in the expression of RBPs have been shown to impact the various stages of RNA metabolism, leading to altered RBP expression and malfunction in the pathogenesis of diverse diseases [70]. The development of cancer and neurodegenerative disorders can be attributed to disruptions in the process of splicing, particularly alternative splicing, which can be linked to the production of cell surface proteins. According to Kim et al. (2009), these alterations in splicing patterns may serve as potential biomarkers for both diagnosing and categorizing cancer [70]. Also, Kechavarzi and Janga (2014) performed a comparative analysis of gene expression profiles in cancerous and healthy tissues, revealing a notable upregulation of approximately 30 RNA-binding proteins (RBPs) in at least six out of the nine examined cancer cases [71]. Many studies have been conducted to examine the significance of RBM-3 in various human tissue types. RBM3 has been identified as a potential indicator for predicting treatment outcomes and prognosis in different forms of human cancer, such as prostate, breast, and ovarian cancer [72] and as shown in Figure 3. A study conducted by Jogi et al. (2009) used an antibody-based proteomics strategy to examine the levels of RBM3 protein in both tumor and normal tissue samples [73]. Based on immunohistochemistry and tissue microarrays in two distinct breast cancer patient groups, it was reported that nuclear RBM3 expression was linked to smaller, lower-grade, and ER-positive tumors. Also, in both patient cohorts, the upregulation of nuclear RBM3 protein expression was associated with improved overall survival and a lower risk of recurrence. It was also observed that elevated levels of nuclear RBM3 expression were especially prominent in hormone receptor-positive tumors, as confirmed by multivariate analysis involving tamoxifen treatment controls [74].
RBM3 has been found to serve as an independent prognostic factor in epithelial ovarian cancer patient cohorts. The expression of RBM3 has been examined in vitro using both cisplatin-sensitive and cisplatin-resistant ovarian cancer cell lines. An increased expression of RBM3 at both the protein and mRNA levels were observed in the cisplatin-sensitive cells as compared to their resistant counterparts. Silencing of RBM3 expression via small interfering RNA (siRNA) led to an enhanced cisplatin resistance, as evidenced by increased cell viability and a higher proportion of cells arrested in the G2/M-phase [74].
In addition, studies have been conducted on epithelial ovarian carcinoma to explore the molecular mechanisms underlying the prognostic significance of RBM3 in epithelial ovarian cancer (EOC). Gene analysis showed a connection between RBM3 expression and various cellular processes linked to the preservation of DNA integrity, including DNA replication, chromatin remodeling, and DNA integrity checkpoint mechanisms. Findings indicate a relationship between unfavorable prognosis in EOC patients and elevated levels of checkpoint kinase 1 and 2 homologues (Chk1 and Chk2) proteins. Thus, RBM3 may participate in cellular responses to DNA damage, potentially elucidating its cisplatin-sensitizing attributes and favorable prognostic impact in EOC. Moreover, studies have shown that Chk1 and Chk2 serve as indicators of poor prognosis and treatment response prediction in EOC [75].
In malignant melanoma, the expression of minichromosome maintenance 3 (MCM3) has been identified as an independent biomarker indicating a poor prognosis. An inverse relationship between RBM3 expression and MCM3 expression has been reported in previous studies [76,77]. These findings are similar to the conclusions drawn by Ehlén et al. (2011), who suggested that MCM3 expression is associated with unfavorable outcomes and has potential clinical use as a prognostic indicator in EOC [78]. Experimental data obtained from an in vitro study revealed a notable decrease in RBM3 gene expression as melanoma cells proliferated. Consistent with the above findings, it has been established that RBM3 expression is reduced in metastatic melanoma compared to primary tumors, where strong nuclear RBM3 expression is correlated with positive clinicopathological characteristics and extended overall survival [79].
Immunohistochemistry studies were conducted on colorectal cancer (CRC) cells to assess the prognostic significance of RBM3 in two distinct patient cohorts’ tumors, with the use of both monoclonal and polyclonal antibodies. The results showed that the nuclear expression of RBM3 was linked to enhanced overall survival in CRC patients, along with a strong correlation between the two types of antibodies [80]. In colorectal tumors, elevated levels of nuclear and cytoplasmic RBM3 expression were noted. The regulatory mechanism of RBM3 was elucidated through its binding to AU-rich sequences in the 3’UTR of cyclooxygenase-2 (COX-2). Overexpression of RBM3 induced anchorage-independent growth in non-transformed cells and increased cell proliferation, while the depletion of RBM3 resulted in decreased proliferation of colon adenocarcinoma cells [81].
Another group found that RBM3 overexpression in prostate cancer cells led to a reduction of their tumorigenic potential in vivo and their stem cell-like properties in vitro [82]. It has also been noted that the splicing of RNA in the CD44 variant v8-v10 was inhibited either by overexpressing RBM3 or by culturing cells at 32 °C, resulting in an elevation in the expression of the standard CD44 protein (CD44s) isoform. Conversely, the ratio of CD44v8-v10 to CD44s mRNA was increased by silencing RBM3 or by culturing cells in soft agar. Mechanistic analyses showed that the promotion of CD44v8-v10 hindered the expression of cyclin D1 and coincided with MMP9-mediated cleavage of CD44s. Meanwhile, the siRNA-mediated suppression of CD44v8-v10 reduced the capacity of prostate cancer cells to form colonies in soft agar. Additionally, RBM3 played a role in the stem cell-like characteristics of prostate cancer by inhibiting the splicing of CD44v8-v1096. Moreover, Jonsson et al., (2011) reported that RBM3 was significantly upregulated in various fractions and intensities in invasive carcinoma and prostatic intraepithelial neoplasia [83]. In contrast, RBM3 was seldom expressed in normal prostatic gland epithelium. Thus, the immunohistochemical evaluation of RBM3 expression may serve as an important tool for the stratification and prognostication of prostate cancer patients. Moreover, Boman et al., (2013) suggested that the assessment of RBM3 expression in samples obtained from urothelial bladder cancer could offer greater value for the early diagnosis, staging, prognosis, stratification, and monitoring of disease progression [84].
A study was conducted by Jonsson et al. to assess the clinicopathological associations and prognostic implications of RBM3 expression in tumors originating from a consecutive series of upper gastrointestinal adenocarcinomas. The findings indicated that reduced RBM3 expression is considered an independent prognostic factor for unfavorable outcomes in individuals diagnosed with adenocarcinoma affecting the upper gastrointestinal tract [85]. Also, RBM3 expression levels were found to be elevated in tumors displaying gastric intestinal metaplasia or Barrett’s esophagus as compared to those lacking precursor lesions. These findings reveal a potential role of RBM3 in diverse pathways contributing to the development of esophagogastric junction, esophageal, and gastric adenocarcinomas [85].
RBM3 plays a pivotal role in tumorigenesis across multiple cancers by modulating molecular pathways and influencing cellular processes. It stabilizes mRNA of tumorigenic factors (COX-2, IL-8, VEGF) and drives proliferation, angiogenesis, and metastasis in HCC, breast, pancreatic, and colorectal cancers. Additionally, RBM3 enhances β-catenin signaling, interacts with YAP1, and regulates apoptosis, suggesting its potential as a prognostic marker and therapeutic target [58].
Preclinical and Clinical Studies About RBM3
As discussed earlier, RBM3 has potential clinical use. A study conducted by Wahlin et al. made an argument that RBM3 may be a predictive biomarker in chemotherapy response in muscle-invasive bladder cancer. They highlighted the importance of RBM3 as a predictive biomarker and presented clinical data suggesting the importance of neoadjuvant chemotherapy in MIBC (muscle-invasive bladder cancer) patients with high nuclear RBM3 levels. Overall, high levels of RBM3 in neoplastic cells were associated with enhanced survival rates among individuals undergoing neoadjuvant chemotherapy. Also, they found out that suppression of RBM3 in malignant bladder carcinoma cells of high grade was shown to diminish their susceptibility to cisplatin and gemcitabine. Moreover, they reported that the capacity of RBM3 to serve as a prognostic indicator for the reaction to chemotherapy in muscle-invasive bladder cancer is worth considering [86]. Also, there has been an association between reduced expression of RBM3 and advanced tumor stage in non-small cell lung cancers (NSCLCs), especially in cases of lung adenocarcinomas (LUACs). The diminished presence of RBM3 is indicative of a poor prognosis in LUAC patients, correlating with a shortened overall survival [74,87]. Also, it was proposed that RBM3 may function as an independent prognostic indicator for overall survival in individuals diagnosed with primary malignant melanoma [79]. Another study also made known the significant role of nuclear RBM3 expression in breast cancer, especially in ER-positive tumors associated with low grade small tumors [88]. Similarly, it was found out that enhanced nuclear RBM3 expression correlates with enhanced overall and recurrence-free survival rates in breast cancer, and RBM3 may serve as an independent prognostic factor in breast cancer, particularly in ER-positive tumors [72]. In prostate cancer, it has been reported that enhanced expression of RBM3 in the disease may be associated with a prolonged time to disease progression; thus, RBM3 may serve as a potential biomarker for a favorable prognosis of the disease. This cohort analysis was done on 88 patients treated with radical prostatectomy for localized disease [72]. It has also been suggested that RBM3 may play a useful role as a prognostic and treatment predictive marker in EOC [74].
4. Discussion
Zhou et al. admit that in cancer, the expression level and clinical behavior of RBM3 is very conflicting; thus, further studies of the regulatory mechanisms of RBM3 in oncogenic or tumor suppression are of great need both in vitro and in vivo. As a potential biomarker, there is a need for further studies to evaluate the correlation between the expression level of RBM3 in diverse cancer stages to categorize patients for individualized prevention and treatment. In neuroprotection, though it has been shown that RBM3 is a promising therapeutic candidate for neuroprotection in prion-infected and 5XFAD models, it must be done in humans before drugs are developed to mimic the protective effects of cooling [38]. Also, there is a need for further studies to establish the importance of RBM3 as a promising target in cancer diagnosis and therapy, investigating the molecular mechanisms that promote the expression of RBM3 under adverse conditions and in cancer as well [27]. Again, there is a need for further studies for alternate explanations for the observed effects of RBM3 on tumor cells and immune cells in the tumor microenvironment. The mechanism of action of RBM3 in certain diseases, including cancer, particularly in prostate cancer, is not well-known; therefore, further investigations are needed.
Research has shown that the upregulation of RBM3 may be linked to certain diseases such as poxvirus disease [89], hepatocellular carcinoma [90], breast cancer [91], pancreatic [92] and colorectal cancer [93]. However, there is not enough evidence to prove this. Since this is a potential concern, we need more experimental and clinical data to prove or disprove this concept. This will help us to know if we can limit toxicity and achieve a good safety profile in case there are molecules that are being discovered primarily to upregulate RBM3 to treat various conditions, including cancer.
Author Contributions
Marvin A. Larbi carried out the literature review and wrote the manuscript. Dmitriy Minond and Robert Getzenberg edited the manuscript. All authors have approved the final version of the manuscript.
Funding
This research was funded by Grant 24B09 from Florida Department of Health (PI – DM).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
RBM3 RNA Binding Motif 3
ALS Amyotrophic Lateral Sclerosis
CRC Colorectal Cancer
CRIP Cold-inducible RNA-binding Protein
EOC Epithelial Ovarian Cancer
ER Endoplasmic Reticulum
PARP Poly ADP Ribose Polymerase
RBD RNA-Binding Domain
RRM RNA Recognition Motif
References
- Gerstberger, S.; Hafner, M.; Genetics, T.T.N.R.; undefined. A census of human RNA-binding proteins. nature.comS Gerstberger, M Hafner, T TuschlNature Reviews Genetics, 2014•nature.com.
- Masuzawa, T.; omega, T.O.A.C.S.; undefined. Roles of the RGG domain and RNA recognition motif of nucleolin in G-quadruplex stabilization. ACS PublicationsT Masuzawa, T OyoshiACS omega, 2020•ACS Publications 2024, 30, 2–2. [CrossRef]
- Naef, V.; De Sarlo, M.; Testa, G.; Corsinovi, D.; Azzarelli, R.; Borello, U.; Ori, M. The Stemness Gene Mex3A Is a Key Regulator of Neuroblast Proliferation During Neurogenesis. Frontiers in Cell and Developmental Biology 2020, 8. [CrossRef]
- Tyzack, G.E.; Luisier, R.; Taha, D.M.; Neeves, J.; Brain, M.M.; undefined. Widespread FUS mislocalization is a molecular hallmark of amyotrophic lateral sclerosis. academic.oup.comGE Tyzack, R Luisier, DM Taha, J Neeves, M Modic, JS Mitchell, I Meyer, L GreensmithBrain, 2019•academic.oup.com.
- Sreedharan, J.; Blair, I.P.; Tripathi, V.B.; Hu, X.; Vance, C.; Rogelj, B.; Ackerley, S.; Durnall, J.C.; Williams, K.L.; Buratti, E.; et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science 2008, 319, 1668–1672. [CrossRef]
- Qian, J.; Hassanein, M.; Hoeksema, M.D.; Harris, B.K.; Zou, Y.; Chen, H.; Lu, P.; Eisenberg, R.; Wang, J.; Espinosa, A.; et al. The RNA binding protein FXR1 is a new driver in the 3q26-29 amplicon and predicts poor prognosis in human cancers. 2015, 112, 3469–3474. [CrossRef]
- Eric Wang, A.; Lu, S.X.; Pastore, A.; Krainer, A.R.; Abdel-Wahab, O.; Wang, E.; Chen, X.; Imig, J.; Chun-Wei Lee, S.; Hockemeyer, K.; et al. Targeting an RNA-binding protein network in acute myeloid leukemia. cell.comE Wang, SX Lu, A Pastore, X Chen, J Imig, SCW Lee, K Hockemeyer, YE GhebrechristosCancer cell, 2019•cell.com 2019, 35, 369–384.e367. [CrossRef]
- Repici, M.; Hassanjani, M.; Maddison, D.C.; Garção, P.; Cimini, S.; Patel, B.; Szegö, É.M.; Straatman, K.R.; Lilley, K.S.; Borsello, T.; et al. The Parkinson’s Disease-Linked Protein DJ-1 Associates with Cytoplasmic mRNP Granules During Stress and Neurodegeneration. Molecular Neurobiology 2019, 56, 61–77. [CrossRef]
- Vanderweyde, T.; Apicco, D.J.; Youmans-Kidder, K.; Ash, P.E.A.; Cook, C.; Lummertz Da Rocha, E.; Jansen-West, K.; Frame, A.A.; Citro, A.; Leszyk, J.D.; et al. Interaction of tau with the RNA-Binding Protein TIA1 Regulates tau Pathophysiology and Toxicity. Cell Reports 2016, 15, 1455–1466. [CrossRef]
- Julio, A.R.; Backus, K.M. New approaches to target RNA binding proteins. Current Opinion in Chemical Biology 2021, 62, 13–23. [CrossRef]
- Cléry, A.; Blatter, M.; biology, F.H.T.A.C.o.i.s.; undefined. RNA recognition motifs: boring? Not quite. ElsevierA Cléry, M Blatter, FHT AllainCurrent opinion in structural biology, 2008•Elsevier.
- Côté, J.; Chemistry, S.R.J.o.B.; undefined. Tudor domains bind symmetrical dimethylated arginines. ASBMB.
- Tradewell, M.L.; Yu, Z.; …, M.T.H.m.; undefined. Arginine methylation by PRMT1 regulates nuclear-cytoplasmic localization and toxicity of FUS/TLS harbouring ALS-linked mutations. academic.oup.comML Tradewell, Z Yu, M Tibshirani, MC Boulanger, HD Durham, S RichardHuman molecular genetics, 2012•academic.oup.com.
- Thandapani, P.; O’Connor, T.R.; Bailey, T.L.; cell, S.R.M.; undefined. Defining the RGG/RG motif. cell.comP Thandapani, TR O’Connor, TL Bailey, S RichardMolecular cell, 2013•cell.com 2013. [CrossRef]
- Ciuzan, O.; Hancock, J.; Pamfil, D.; …, I.W.P.; undefined. The evolutionarily conserved multifunctional glycine-rich RNA-binding proteins play key roles in development and stress adaptation. Wiley Online LibraryO Ciuzan, J Hancock, D Pamfil, I Wilson, M LadomeryPhysiologia plantarum, 2015•Wiley Online Library 2015, 153, 1–11. [CrossRef]
- Zhu, X.; Bührer, C.; Wellmann, S. Cold-inducible proteins CIRP and RBM3, a unique couple with activities far beyond the cold. 2016, 73, 3839–3859. [CrossRef]
- Danno, S.; Nishiyama, H.; Higashitsuji, H.; …, H.Y.B.a.; undefined. Increased transcript level of RBM3, a member of the glycine-rich RNA-binding protein family, in human cells in response to cold stress. ElsevierS Danno, H Nishiyama, H Higashitsuji, H Yokoi, JH Xue, K Itoh, T Matsuda, J FujitaBiochemical and biophysical research communications, 1997•Elsevier.
- Derry, J.M.J.; Kerns, J.A.; genetics, U.F.H.m.; undefined. RBM3, a novel human gene in Xp11.23 with a putative RNA-binding domain. academic.oup.comJMJ Derry, JA Kerns, U FranckeHuman molecular genetics, 1995•academic.oup.com 1995, 4, 2307–2311.
- Wellmann, S.; Bührer, C.; …, E.M.-J.o.c.; 2004, u. Oxygen-regulated expression of the RNA-binding proteins RBM3 and CIRP by a HIF-1-independent mechanism.
- J-L Wong, J.; Au, A.Y.M.; Gao, D.; Pinello, N.; Kwok, C.-T.; Thoeng, A.; Lau, K.A.; Gordon, J.E.A.; Schmitz, U.; Feng, Y.; et al. RBM3 regulates temperature sensitive miR-142–5p and miR-143 (thermomiRs), which target immune genes and control fever. academic.oup.comJJL Wong, AYM Au, D Gao, N Pinello, CT Kwok, A Thoeng, KA Lau, JEA Gordon, U SchmitzNucleic acids research, 2016•academic.oup.com 2016, 44, 2888–2897. [CrossRef]
- Dresios, J.; Aschrafi, A.; Owens, G.C.; Vanderklish, P.W.; Edelman, G.M.; Mauro, V.P. Cold stress-induced protein Rbm3 binds 60S ribosomal subunits, alter microRNA levels, and enhances global protein synthesis. Proceedings of the National Academy of Sciences of the United States of America 2005, 102, 1865–1870. [CrossRef]
- Cok, S.J.; Acton, S.J.; Sexton, A.E.; Chemistry, A.R.M.J.o.B.; undefined. Identification of RNA-binding proteins in RAW 264.7 cells that recognize a lipopolysaccharide-responsive element in the 3-untranslated region of the murine. ASBMB.
- Nishiyama, H.; Itoh, K.; Kaneko, Y.; …, M.K.-T.J.o.c.; 1997, u. A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth.
- Nishiyama, H.; Higashitsuji, H.; Yokoi, H.; Itoh, K.; Gene, S.D.-.; 1997, u. Cloning and characterization of human CIRP (cold-inducible RNA-binding protein) cDNA and chromosomal assignment of the gene.
- Maruyama, K.; Sato, N.; research, N.O.-N.a.; 1999, u. Conservation of structure and cold-regulation of RNA-binding proteins in cyanobacteria: probable convergent evolution with eukaryotic glycine-rich RNA-binding.
- Biotechnol, J.F.-J.M.M.; 1999, u. Cold shock response in mammalian cells. 1999, 1, 243–255.
- Wellmann, S.; Truss, M.; Bruder, E.; Tornillo, L.; …, A.Z.P.; undefined. The RNA-binding protein RBM3 is required for cell proliferation and protects against serum deprivation-induced cell death. nature.comS Wellmann, M Truss, E Bruder, L Tornillo, A Zelmer, K Seeger, C BührerPediatric research, 2010•nature.com.
- Ryan, J.C.; Morey, J.S.; Ramsdell, J.S.; Neuroscience, F.M.V.D.; undefined. Acute phase gene expression in mice exposed to the marine neurotoxin domoic acid. ElsevierJC Ryan, JS Morey, JS Ramsdell, FM Van DolahNeuroscience, 2005•Elsevier.
- Dupont-Versteegden, E.E.; Nagarajan, R.; Beggs, M.L.; Bearden, E.D.; Simpson, P.M.; Peterson, C.A. Identification of cold-shock protein RBM3 as a possible regulator of skeletal muscle size through expression profiling. journals.physiology.orgEE Dupont-Versteegden, R Nagarajan, ML Beggs, ED Bearden, PM Simpson, CA PetersonAmerican Journal of Physiology-Regulatory, Integrative and, 2008•journals.physiology.org 2008, 295. [CrossRef]
- Cui, Z.; Zhang, J.; Bao, G.; Xu, G.; Sun, Y.; Wang, L.; Chen, J.; Jin, H.; Liu, J.; Yang, L.; et al. Spatiotemporal Profile and Essential Role of RBM3 Expression after Spinal Cord Injury in Adult Rats. Journal of Molecular Neuroscience 2014, 54, 252–263. [CrossRef]
- Zhao, W.; Xu, D.; Cai, G.; Zhu, X.; Qian, M.; Liu, W.; Cui, Z. Spatiotemporal pattern of RNA-binding motif protein 3 expression after spinal cord injury in rats. 2014, 34, 491–499. [CrossRef]
- Yang, H.; Ju, F.; Guo, X.; Ma, S.; Wang, L.; reports, B.C.-S.; 2017, u. RNA-binding protein RBM3 prevents NO-induced apoptosis in human neuroblastoma cells by modulating p38 signaling and miR-143.
- Chip, S.; Zelmer, A.; Ogunshola, O.; Felderhoff-Mueser, U.; Nitsch, C.; Bührer, C.; Wellmann, S. The RNA-binding protein RBM3 is involved in hypothermia induced neuroprotection. ElsevierS Chip, A Zelmer, OO Ogunshola, U Felderhoff-Mueser, C Nitsch, C Bührer, S WellmannNeurobiology of disease, 2011•Elsevier 2011, 43, 388–396. [CrossRef]
- Kita, H.; Carmichael, J.; Swartz, J.; Muro, S.; Wyttenbach, A.; Matsubara, K.; Rubinsztein, D.C.; Kato, K. Modulation of polyglutamine-induced cell death by genes identified by expression profiling. academic.oup.comH Kita, J Carmichael, J Swartz, S Muro, A Wyttenbach, K Matsubara, DC Rubinsztein, K KatoHuman molecular genetics, 2002•academic.oup.com.
- Rosenthal, L.-M.; Tong, G.; Walker, C.; Wowro, S.J.; Krech, J.; Pfitzer, C.; Justus, G.; Berger, F.; Rose, K.; Schmitt, L. Neuroprotection via RNA-binding protein RBM3 expression is regulated by hypothermia but not by hypoxia in human SK-N-SH neurons. Taylor & FrancisLM Rosenthal, G Tong, C Walker, SJ Wowro, J Krech, C Pfitzer, G Justus, F BergerHypoxia, 2017•Taylor & Francis 2017, Volume 5, 33–43. [CrossRef]
- Smart, F.; Aschrafi, A.; Atkins, A.; Owens, G.C.; Pilotte, J.; Cunningham, B.A.; Vanderklish, P.W. Two isoforms of the cold-inducible mRNA-binding protein RBM3 localize to dendrites and promote translation. 2007, 101, 1367–1379. [CrossRef]
- Tong, G.; Endersfelder, S.; Rosenthal, L.; research, S.W.-B.; 2013, u. Effects of moderate and deep hypothermia on RNA-binding proteins RBM3 and CIRP expressions in murine hippocampal brain slices.
- Zhou, R.; Lu, X.; Zhang, C.; Oncotarget, D.Y.-.; 2017, u. RNA binding motif protein 3: a potential biomarker in cancer and therapeutic target in neuroprotection.
- Pilotte, J.; Cunningham, B.; research, G.E.-B.; 2009, u. Developmentally regulated expression of the cold-inducible RNA-binding motif protein 3 in euthermic rat brain.
- Mardakheh, F.K.; Paul, A.; Kü, S.; McCarthy, A.; Yuan, Y.; Correspondence, C.J.M. Global analysis of mRNA, translation, and protein localization: local translation is a key regulator of cell protrusions. cell.comFK Mardakheh, A Paul, S Kümper, A Sadok, H Paterson, A Mccarthy, Y Yuan, CJ MarshallDevelopmental cell, 2015•cell.com 2015, 35, 344–357. [CrossRef]
- Caillier, A.; Huot, M.-E.; Bergeman, J.; Houle, F.; Gagné, L.M.G.; Huot, M.-É. Localized translation regulates cell adhesion and transendothelial migration. journals.biologists.comJ Bergeman, A Caillier, F Houle, LM Gagné, MÉ HuotJournal of Cell Science, 2016•journals.biologists.com 2016. [CrossRef]
- Hoog, C.L.d.; Foster, L.J.; Cell, M.M.; undefined. RNA and RNA binding proteins participate in early stages of cell spreading through spreading initiation centers. cell.comCL de Hoog, LJ Foster, M MannCell, 2004•cell.com.
- Vanderklish, P.; Genes, G.E.-.; Behavior, B.a.; 2005, u. Differential translation and fragile X syndrome. 2005, 4, 360–384. [CrossRef]
- Antar, L.N.; Afroz, R.; Dictenberg, J.B.; Carroll, R.C.; Bassell, G.J. Metabotropic glutamate receptor activation regulates fragile x mental retardation protein and FMR1 mRNA localization differentially in dendrites and at synapses. Soc NeuroscienceLN Antar, R Afroz, JB Dictenberg, RC Carroll, GJ BassellJournal of Neuroscience, 2004•Soc Neuroscience 2004. [CrossRef]
- Leung, K.; Horck, F.v.; Lin, A.; …, R.A.-N.; 2006, u. Asymmetrical β-actin mRNA translation in growth cones mediates attractive turning to netrin-1.
- Yasuda, K.; Zhang, H.; Loiselle, D.; …, T.H.-J.o.C.; 2013, u. The RNA-binding protein Fus directs translation of localized mRNAs in APC-RNP granules.
- Mili, S.; Moissoglu, K.; Nature, I.M.-.; 2008, u. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions.
- Hörnberg, H.; Holt, C. RNA-binding proteins and translational regulation in axons and growth cones. Frontiers in Neuroscience 2013. [CrossRef]
- Cervero, P.; Himmel, M.; Krüger, M.; biology, S.L.E.j.o.c.; undefined. Proteomic analysis of podosome fractions from macrophages reveals similarities to spreading initiation centres. ElsevierP Cervero, M Himmel, M Krüger, S LinderEuropean journal of cell biology, 2012•Elsevier.
- Andersson, M.K.; Ståhlberg, A.; Arvidsson, Y.; Olofsson, A.; Semb, H.; Stenman, G.; Nilsson, O.; Åman, P. The multifunctional FUS, EWS and TAF15 proto-oncoproteins show cell type-specific expression patterns and involvement in cell spreading and stress response. 2008, 9. [CrossRef]
- Caillier, A.; Huot, M.-E.; Bergeman, J.; Houle, F.; Gagné1, L.M.G.; Huot, M.-É. Localized translation regulates cell adhesion and transendothelial migration. 2016. [CrossRef]
- Hoog, C.d.; Foster, L.; Cell, M.M.-.; 2004, u. RNA and RNA binding proteins participate in early stages of cell spreading through spreading initiation centers.
- Abstract, G.; Bastide, A.; Peretti, D.; Knight, J.R.P.; Smales, C.M.; Mallucci, G.R.;Willis, A.E. RTN3 is a novel cold-induced protein and mediates neuroprotective effects of RBM3. 2017, 27, 638–650. [CrossRef]
- Peretti, D.; Bastide, A.; Radford, H.; Verity, N.; Molloy, C.; Martin, M.G.; Moreno, J.A.; Steinert, J.R.; Smith, T.; Dinsdale, D.; et al. RBM3 mediates structural plasticity and protective effects of cooling in neurodegeneration. 2015, 518, 236–239. [CrossRef]
- Von Der Ohe, C.G.; Darian-Smith, C.; Garner, C.C.; Craig Heller, H. Ubiquitous and temperature-dependent neural plasticity in hibernators. Soc NeuroscienceCG von der Ohe, C Darian-Smith, CC Garner, HC HellerJournal of Neuroscience, 2006•Soc Neuroscience 2006. [CrossRef]
- Popov, V.; Bocharova, L.; Neuroscience, A.B.-.; 1992, u. Repeated changes of dendritic morphology in the hippocampus of ground squirrels in the course of hibernation.
- Pilotte, J.; Kiosses, W.; Chan, S.W.; Makarenkova, H.P.; Dupont-Versteegden, E.; Vanderklish, P.W. Morphoregulatory functions of the RNA-binding motif protein 3 in cell spreading, polarity and migration. Scientific Reports 2018 8:1 2018, 8, 1–19. [CrossRef]
- Zhou, R.B.; Lu, X.L.; Zhang, C.Y.; Yin, D.C. RNA binding motif protein 3: A potential biomarker in cancer and therapeutic target in neuroprotection. 2017, 8, 22235–22250. [CrossRef]
- Polderman, K.H. Keeping a cool head: How to induce and maintain hypothermia. Critical care medicine 2004, 32, 2558–2560. [CrossRef]
- Chip, S.; Zelmer, A.; Ogunshola, O.O.; Felderhoff-Mueser, U.; Nitsch, C.; Bührer, C.; Wellmann, S. The RNA-binding protein RBM3 is involved in hypothermia induced neuroprotection. 2011, 43, 388–396. [CrossRef]
- Peretti, D.; Bastide, A.; Radford, H.; Verity, N.; Nature, C.M.-.; 2015, u. RBM3 mediates structural plasticity and protective effects of cooling in neurodegeneration.
- SiesjÖ, B.K.; Bengtsson, F.; Grampp, W.; Theander, S. Calcium, excitotoxins, and neuronal death in the brain. Annals of the New York Academy of Sciences 1989, 568, 234–251. [CrossRef]
- Balogh, G.T.; Vukics, K.; Könczöl, Á.; Kis-Varga, Á.; Gere, A.; Fischer, J. Nitrone derivatives of trolox as neuroprotective agents. Bioorganic and Medicinal Chemistry Letters 2005, 15, 3012–3015. [CrossRef]
- Brown, R.S. Zinc finger proteins: getting a grip on RNA. Current opinion in structural biology 2005, 15, 94–98. [CrossRef]
- Danno, S.; Nishiyama, H.; Higashitsuji, H.; …, H.Y.-B.a.; 1997, u. Increased transcript level of RBM3, a member of the glycine-rich RNA-binding protein family, in human cells in response to cold stress.
- Raff, M.C.; Barres, B.A.; Burne, J.F.; Coles, H.S.; Ishizaki, Y.; Jacobson, M.D. Programmed cell death and the control of cell survival: lessons from the nervous system. 1993, 262, 695–700. [CrossRef]
- Shintani, Y.; Terao, Y. 4 Molecular Mechanisms Underlying the Neuroprotective Effect of Hypothermia in Cerebral Ischemia.
- Chip, S.; Zelmer, A.; Ogunshola, O.; O; Felderhoff-Mueser, U.; Nitsch, C.; Bührer, C.; ; Wellmann, S. The RNA-binding protein RBM3 is involved in hypothermia induced neuroprotection. 2011, 43, 388–396. [CrossRef]
- Shintani, Y.; Terao, Y. 4 Molecular Mechanisms Underlying the Neuroprotective Effect of Hypothermia in Cerebral Ischemia.
- Kim, M.Y.; Hur, J.; reports, S.J.J.B.M.B.; undefined. Emerging roles of RNA and RNA-binding protein network in cancer cells. koreascience.krMY Kim, J Hur, SJ JeongBMB reports, 2009•koreascience.kr.
- Kechavarzi, B.; Janga, S.C. Dissecting the expression landscape of RNA-binding proteins in human cancers. Genome Biology 2014, 15. [CrossRef]
- Jögi, A.; Brennan, D.; Rydén, L.; Magnusson, K.; Pathology, M.F.-M.; 2009, u. Nuclear expression of the RNA-binding protein RBM3 is associated with an improved clinical outcome in breast cancer.
- Salomonsson, A.; Micke, P.; Mattsson, J.S.M.; La Fleur, L.; Isaksson, J.; Jönsson, M.; Nodin, B.; Botling, J.; Uhlén, M.; Jirström, K.; et al. Comprehensive analysis of RNA binding motif protein 3 (RBM3) in non-small cell lung cancer. Cancer Medicine 2020, 9, 5609–5609. [CrossRef]
- Ehlén, Å.; Brennan, D.J.; Nodin, B.; O’Connor, D.P.; Eberhard, J.; Alvarado-Kristensson, M.; Jeffrey, I.B.; Manjer, J.; Brändstedt, J.; Uhlén, M.; et al. Expression of the RNA-binding protein RBM3 is associated with a favourable prognosis and cisplatin sensitivity in epithelial ovarian cancer. 2010, 8. [CrossRef]
- Ehlén, Õ.; Nodin, B.; Rexhepaj, E.; …, J.B.T.; undefined. RBM3-regulated genes promote DNA integrity and affect clinical outcome in epithelial ovarian cancer. ElsevierÕ Ehlén, B Nodin, E Rexhepaj, J Brändstedt, M Uhlén, M Alvarado-Kristensson, F PonténTranslational oncology, 2011•Elsevier.
- Baldi, A.; Battista, T.; De Luca, A.; Santini, D.; Rossiello, L.; Baldi, F.; Giorgio Natali, P.; Lombardi, D.; Picardo, M.; Felsani, A.; et al. Identification of genes down-regulated during melanoma progression: a cDNA array study. 2003, 12, 213–218. [CrossRef]
- Nodin, B.; Fridberg, M.; Jonsson, L.; Bergman, J.; Uhlén, M.; Jirström, K. High MCM3 expression is an independent biomarker of poor prognosis and correlates with reduced RBM3 expression in a prospective cohort of malignant melanoma. Diagnostic Pathology 2012, 7. [CrossRef]
- Ehlén, Õ.; Nodin, B.; Rexhepaj, E.; …, J.B.-T.; 2011, u. RBM3-regulated genes promote DNA integrity and affect clinical outcome in epithelial ovarian cancer.
- Jonsson, L.; Bergman, J.; Nodin, B.; Manjer, J.; Pontén, F.; Uhlén, M.; Jirström, K. Low RBM3 protein expression correlates with tumour progression and poor prognosis in malignant melanoma: An analysis of 215 cases from the Malmö Diet and Cancer Study. 2011, 9. [CrossRef]
- Hjelm, B.; Brennan, D.J.; Zendehrokh, N.; Eberhard, J.; Nodin, B.; Gaber, A.; Pontén, F.; Johannesson, H.; Smaragdi, K.; Frantz, C.; et al. High nuclear RBM3 expression is associated with an improved prognosis in colorectal cancer. 2011, 5, 624–635. [CrossRef]
- Sureban, S.M.; Ramalingam, S.; Natarajan, G.; May, R.; Subramaniam, D.; Bishnupuri, K.S.; Morrison, A.R.; Dieckgraefe, B.K.; Brackett, D.J.; Postier, R.G.; et al. Translation regulatory factor RBM3 is a proto-oncogene that prevents mitotic catastrophe. 2008, 27, 4544–4556. [CrossRef]
- Zeng, Y.; Wodzenski, D.; Gao, D.; Shiraishi, T.; research, N.T.-C.; 2013, u. Stress-response protein RBM3 attenuates the stem-like properties of prostate cancer cells by interfering with CD44 variant splicing.
- Jonsson, L.; Gaber, A.; Ulmert, D.; Uhlén, M.; Bjartell, A.; Jirström, K. High RBM3 expression in prostate cancer independently predicts a reduced risk of biochemical recurrence and disease progression. 2011, 6. [CrossRef]
- Boman, K.; Segersten, U.; Ahlgren, G.; Eberhard, J.; Uhlén, M.; Jirström, K.; Malmström, P.U. Decreased expression of RNA-binding motif protein 3 correlates with tumour progression and poor prognosis in urothelial bladder cancer. BMC Urology 2013, 13. [CrossRef]
- Jonsson, L.; Hedner, C.; Gaber, A.; Korkocic, D.; Nodin, B.; Uhlén, M.; Eberhard, J.; Jirström, K. High expression of RNA-binding motif protein 3 in esophageal and gastric adenocarcinoma correlates with intestinal metaplasia-associated tumours and independently predicts a reduced risk of recurrence and death. Biomarker Research 2014, 2, 11–11. [CrossRef]
- Wahlin, S.; Boman, K.; Moran, B.; Nodin, B.; Gallagher, W.M.; Karnevi, E.; Jirström, K. Pre-clinical and clinical studies on the role of RBM3 in muscle-invasive bladder cancer: longitudinal expression, transcriptome-level effects and modulation of chemosensitivity. BMC Cancer 2022, 22, 1–15. [CrossRef]
- Melling, N.; Bachmann, K.; Hofmann, B.; El Gammal, A.T.; Reeh, M.; Mann, O.; Moebius, C.; Blessmann, M.; Izbicki, J.R.; Grupp, K. Prevalence and clinical significance of RBM3 immunostaining in non-small cell lung cancers. Journal of Cancer Research and Clinical Oncology 2019, 145, 873–879. [CrossRef]
- Li, Z.; Guo, Q.; Zhang, J.; Fu, Z.; Wang, Y.; Wang, T.; Tang, J. The RNA-Binding Motif Protein Family in Cancer: Friend or Foe? Frontiers in Oncology 2021, 11, 757135–757135. [CrossRef]
- Dellis, S.; Strickland, K.C.; McCrary, W.J.; Patel, A.; Stocum, E.; Wright, C.F. Protein interactions among the vaccinia virus late transcription factors. Virology 2004, 329, 328–336. [CrossRef]
- Miao, X.; Zhang, N. Role of RBM3 in the regulation of cell proliferation in hepatocellular carcinoma. Experimental and Molecular Pathology 2020, 117, 104546–104546. [CrossRef]
- Chen, P.; Yue, X.; Xiong, H.; Lu, X.; Ji, Z. RBM3 upregulates ARPC2 by binding the 3’UTR and contributes to breast cancer progression. 2019, 54, 1387–1397. [CrossRef]
- Karnevi, E.; Dror, L.B.; Mardinoglu, A.; Elebro, J.; Heby, M.; Olofsson, S.-E.; Nodin, B.; Eberhard, J.; Gallagher, W.; Uhlén, M.; et al. Translational study reveals a two-faced role of RBM3 in pancreatic cancer and suggests its potential value as a biomarker for improved patient stratification. Oncotarget 2018, 9, 6188–6200. [CrossRef]
- Venugopal, A.; Subramaniam, D.; Balmaceda, J.; Roy, B.; Dixon, D.A.; Umar, S.; Weir, S.J.; Anant, S. RNA binding protein RBM3 increases β-catenin signaling to increase stem cell characteristics in colorectal cancer cells. Molecular Carcinogenesis 2016, 55, 1503–1516. [CrossRef]
- Zhou, R.B.; Lu, X.L.; Zhang, C.Y.; Yin, D.C. RNA binding motif protein 3: a potential biomarker in cancer and therapeutic target in neuroprotection. 2017, 8, 22235. [CrossRef]
- Van Pelt, D.W.; Hettinger, Z.R.; Dupont-Versteegden, E.E. Cold shock RNA-binding protein RBM3 as a potential therapeutic target to prevent skeletal muscle atrophy. Journal of Orthopedics and Orthopedic Surgery 2020, 1.
- Ávila-Gómez, P.; Vieites-Prado, A.; Correa-Paz, C.; Del Pozo-Filíu, L.; Palomar-Alonso, N.; Campos, F.; López-Arias, E. Therapeutic modulation of protein RBM3 for ischemic stroke treatment. Frontiers in Pharmacology 2025, 16. [CrossRef]
- Wahlin, S.; Boman, K.; Moran, B.; Nodin, B.; Gallagher, W.M.; Karnevi, E.; Jirström, K. Pre-clinical and clinical studies on the role of RBM3 in muscle-invasive bladder cancer: longitudinal expression, transcriptome-level effects and modulation of chemosensitivity. BMC Cancer 2022, 22, 131–131. [CrossRef]
- Zhang, S.; Lv, C. Niu, Y.; Li, C.; Li, X.; Shang, Y.; Zhang, Y.; Zhang, Y.; Zhang, Y.; Zeng, Y. RBM3 suppresses stemness remodeling of prostate cancer in bone microenvironment by modulating N6-methyladenosine on CTNNB1 mRNA. 2023, 14, 1–14. [CrossRef]
- Zeng, Y.; Wodzenski, D.; Gao, D.; Shiraishi, T.; Terada, N.; Li, Y.; Griend, D.J.V.; Luo, J.; Kong, C.; Getzenberg, R.H.; et al. Stress-response protein RBM3 attenuates the stem-like properties of prostate cancer cells by interfering with CD44 variant splicing. 2013, 73, 4123–4133. [CrossRef]
Figure 2.
A model illustrating how RBM3 provides neuroprotection by suppressing the ER stress pathway involving PERK, eIF2α, and CHOP. RBM3 functions as an inhibitor of PERK through a mechanism that depends on NF90. By blocking the PERK-eIF2α-CHOP pathway involved in ER stress, RBM3 helps prevent cell apoptosis. This figure is being printed with permission from [38].
Figure 2.
A model illustrating how RBM3 provides neuroprotection by suppressing the ER stress pathway involving PERK, eIF2α, and CHOP. RBM3 functions as an inhibitor of PERK through a mechanism that depends on NF90. By blocking the PERK-eIF2α-CHOP pathway involved in ER stress, RBM3 helps prevent cell apoptosis. This figure is being printed with permission from [38].

Figure 3.
Role of RBM3 in tumorigenesis.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



