Submitted:
28 May 2026
Posted:
01 June 2026
You are already at the latest version
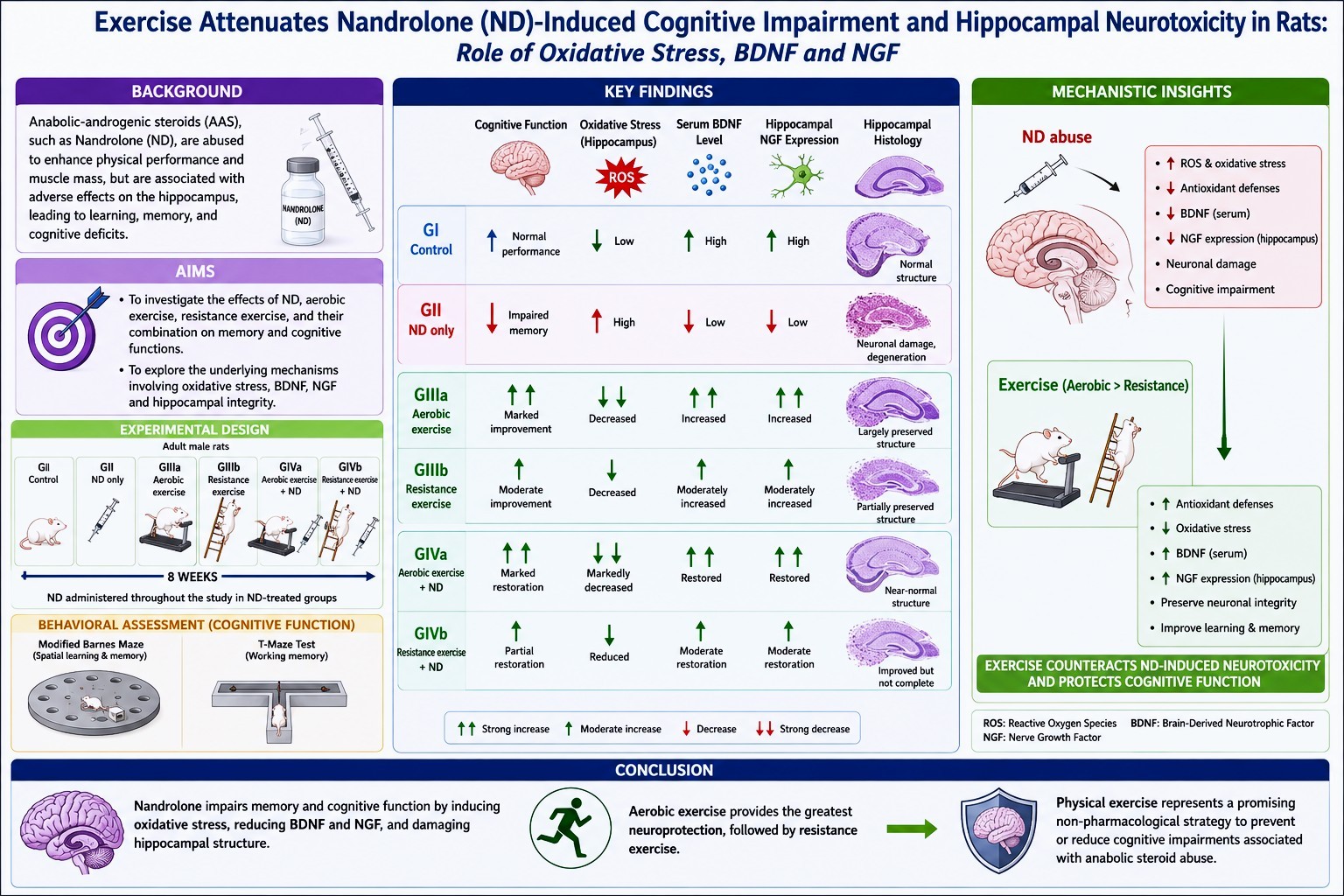
Abstract
Anabolic-androgenic steroids (AAS), such as ND, are commonly used to enhance physical performance and muscle mass. However, its abuse has been associated with numerous adverse health effects, particularly on the hippocampus, affecting learning, working memory, and cognitive functions. The current study aims to investigate the potential effects of ND, aerobic exercise, resistance exercise, and the combination of ND with aerobic or resistance exercise on memory and cognitive functions. Moreover, the study aims to explore the possible underlying mechanisms of changes in memory and cognitive function. Adult male rats were divided into four groups: control (GI), only ND-treated (GII), only exercise (GIIIa & GIIIb), and combined exercise + ND-treated (GIVa & GIVb). Exercise included aerobic treadmill training and ladder-climbing modalities for 8 weeks, either alone or with ND. Cognitive function was evaluated using the Modified Barnes Maze and T-maze tests. Hippocampal integrity was assessed through oxidative stress markers, serum brain-derived neurotrophic factor (BDNF) levels, hippocampal nerve growth factor (NGF) expression, and histological examination. ND significantly impaired spatial learning and memory, increased oxidative stress, lowered serum BDNF levels, and reduced NGF expression in the hippocampus. Exercise markedly attenuated these harmful effects. Aerobic exercise gave the most pronounced improvements. It restored cognitive function, enhanced antioxidant defenses, increased serum BDNF levels, increased hippocampal NGF expression, and preserved hippocampal structure. Resistance training also had neuroprotective effects, but to a lesser extent than aerobic exercise. In brief, both aerobic and anaerobic exercise have been shown to exert protective effects against ND-induced neurotoxicity by enhancing antioxidant defenses and promoting neuronal integrity. The monitored neuroprotective benefits suggest that physical exercise could serve as a possible non-pharmacological approach to prevent or decrease cognitive impairments associated with anabolic steroid abuse.
Keywords:
hippocampus
; anabolic steroids
; oxidative stress
; exercise
; cognition
1. Introduction
The hippocampus is a brain region primarily responsible for memory building. In rats, it consists of several subregions, including the dentate gyrus and the CA1, CA2, CA3, and CA4 subfields. The dentate gyrus contains a layer of densely packed granule cells, an adjacent subgranular zone, and an additional molecular layer. Granule cells play a vital role in the formation of new memories and are characterized by their distinct morphology and prominent granules [1].
Anabolic-androgenic steroids (AAS) abuse is a public health problem for both athletes and non-athletes, including adolescents and young adults [2,3]. Globally, prevalence rates are notable among 20% of US athletes [4], 22% of UAE gym users [5], 13% of Iranian youth bodybuilders [6], and 28% of Riyadh gym users [7]. In Egypt, AAS use was reported among 32% of supplement users and 57% of athletes [8]. AAS are synthetic derivatives of testosterone [9]. Nandrolone decanoate (ND) is frequently abused for its strong anabolic effects [10]. Clinically, AAS are used to treat osteoporosis and muscle-wasting diseases [11]. However, at high doses, AAS can alter central nervous system structure and function [12], particularly affecting the hippocampus, a key region for cognition and behavior [13].
Experimental studies indicate that administration of AAS can induce anxiety, depression, memory deficits, and structural changes in critical brain regions, including the hippocampus, amygdala, and prefrontal cortex [14]. In humans, prolonged use of these substances is associated with increased amygdala volume, cortical thinning, and reduced grey matter [14,15]. ND administration can also increase the levels of pro-inflammatory cytokines, such as TNF-α and IL-1β [16].
Physical exercise confers significant cognitive and mental health benefits. It reduces the risk of depression and improves the overall brain health [17]. Exercise promotes neuroplasticity by increasing brain-derived neurotrophic factor (BDNF), improving memory, and stimulating neurogenesis [18]. Aerobic training has been associated with increased hippocampal volume [19]. Appropriately timed exercise may also cause protection against inflammation associated with certain diseases [20]. The effects of these benefits depend on the type, intensity, and duration of exercise [21]. According to World Health Organization guidelines, any level of physical activity is preferable to inactivity [22]. Therefore, the current study aimed to investigate the effects of different exercise modalities (moderate aerobic and resistance exercise training) on cognitive functions, oxidative stress markers, BDNF levels and expression, and hippocampal morphological changes in rats, and to clarify the underlying mechanisms with and without chronic supra-physiological ND administration.
2. Materials and Methods
2.1. Drugs and Chemicals
ND (Deca-Durabolin 200 mg/mL in arachis oil), Alpha MD, Lynnwood, USA. SOD (CUSABIO, CSB-E08555r), MDA (MYBIOSOURCE, MBS268427), and BDNF ELISA (CUSABIO, CSB-E04504r) kits were used. Routine histology reagents (10% formalin and H&E stains) were used. The recombinant nerve growth factor (NGF) protein was obtained from Abcam (UK; ab269181).
2.2. Study Design
Thirty-six adult male albino rats (200-250 g, 9 weeks old) were obtained from the Laboratory Animal House, Faculty of Veterinary Medicine, Zagazig University. Animals were housed in standard steel cages under monitored temperature (24 ± 2 °C) and a natural light/dark cycle. Rats were acclimatized for 2 weeks before starting the experiment [23]. All procedures followed the Guidelines for the Care and Use of Laboratory Animals (1996) and were approved by the Institutional Animal Care and Use Committee of Zagazig University, ZU-IACUC (No. ZU-IACUC/3/F/281/2026). Rats were randomly assigned to four experimental groups using a computer-generated randomization method to minimize sampling bias. The sample size was chosen based on previous comparable experimental studies and in accordance with ethical principles to minimize animal use. The control group (GI, n=6) received 0.15 mL of a vehicle (Arachis oil, IM) 5 times/week for 8 weeks. The ND group (GII, n=6) received 15 mg of ND /kg/day of ND IM, 5 times/week for 8 weeks [24]. The exercise group (GIII, n=12) was subdivided into two equal subgroups: GIIIa (aerobic treadmill training) and GIIIb (anaerobic ladder-climbing training). The exercise + ND group (GIV, n=12) was subdivided into two equal subgroups: GIVa (aerobic exercise + ND) and GIVb (anaerobic exercise + ND). Behavioral assessments were conducted by investigators blinded to group allocation whenever feasible. Biochemical, histopathological, immuno-histochemical evaluation, and morphometric measurements were performed by independent investigators blinded to treatment conditions. The exclusion criteria consisted of rats weighing less than 200g, severe illness, accidental injury, or inability to complete the assigned exercise protocol.
2.3. Exercise Training Protocols
2.3.1. Aerobic Training Rats
Subgroups GIIIa and GIVa underwent a one-week habituation period to become familiar with the treadmill environment. During this period, they were gently trained to run at a low speed (5m/min for 10min), using minimal tactile stimulation (a gentle push with a small stick) as needed to encourage forward movement. Then rats ran on a motorized treadmill 5 times/week for 8 weeks at progressively increasing intensity (10–20 m/min; 25–30 min/session) without electrical shock [25,26].
2.3.2. Resistance (Anaerobic) Training Rats
Subgroups GIIIb and GIVb were first habituated to the resistance-training apparatus for 1 week, which consisted of a 1-meter ladder inclined at 80°. A dark, enclosed box was placed at the top to encourage climbing and to provide a safe resting area between trials [26]. After habituation, they completed 8 climbs/session, 5 times/week for 8 weeks, with progressively increasing tail-attached loads [24,26].
2.4. Behavioral Assessment:
2.4.1. Barnes Maze Test
Spatial learning and memory were evaluated following standard habituation (Day 0), acquisition (Days 1–3), and probe trials (Days 4 and 9; PT1 and PT2, respectively). Each rat underwent two sessions. There was a 4-day rest (days 5, 6, 7, and 8). Escape latency and error frequency were recorded [27].
2.4.2. T-Maze Spontaneous Alternation Test
Working memory was evaluated over two consecutive days following established spontaneous alternation methods [28]. A percentage of correct choice (alternation) per animal was calculated as follows: Number of correct choices (Alternations) / Total possible alternations × 100
2.4.3. Rotarod Test
Motor coordination and endurance were assessed using an accelerating rotarod (5–40 rpm). Latency to fall was recorded [29].
2.5. Biochemical Assays
Twenty-four hours after behavioral testing, overnight-fasted rats were anesthetized, and blood was collected from the orbital sinus. Serum was separated (3000 rpm, 15 min) and stored at −20 °C [30]. Serum superoxide dismutase (SOD) activity was measured using a sandwich ELISA [31]. Serum Malondialdehyde (MDA) was determined by competitive ELISA [32]. Brain BDNF was quantified in brain homogenates using a quantitative sandwich ELISA [33].
2.6. Histopathology and Immunohistochemistry Study
Each brain was bisected into two hemispheres; one hemisphere was fixed in 10% formalin, paraffin-embedded, sectioned (4 µm), and stained with H&E [34]. Immunohistochemistry for NGF was performed using antigen retrieval, monoclonal primary antibody incubation, and the DAKO EnVision+ system was used as a detection system, and Diaminobenzidine (DAB) was applied as a chromogen to visualize the immunoreactivity, followed by counterstaining with Mayer’s hematoxylin. [35].
2.7. Statistical Analysis:
Researchers of the current study collected, tabulated, and statistically analyzed all data using IBM SPSS Statistics for Windows, Version 23.0 (Armonk, NY: IBM Corp, 2015). They used the Shapiro-Wilk test to assess the normality of quantitative variables and reported descriptive statistics as mean ± standard deviation and ranges. To compare more than two normally distributed groups, they used ANOVA. If the F-test was significant, they used the LSD test for each pair to identify significant groups. All tests considered a two-sided p-value < 0.05 statistically significant, and a p-value ≥ 0.05 as statistically non-significant. Effect sizes were calculated using eta-squared (η²).
3. Results
3.1. Behavioral Assessment:
3.1.1. Modified Barnes Maze Test:
Table 1 shows the Escape Latency Across Acquisition Trials (AT1–AT6). Across all acquisition trials, chronic ND administration (GII) induced a severe and persistent impairment in spatial learning, as evidenced by a significant prolongation in escape latency compared to GI (p<0.001 for all trials). This impairment was strong and did not attenuate with repeated training, indicating that ND induced a marked impairment in the hippocampal-dependent learning. Contrarily, both aerobic (GIIIa) and anaerobic (GIIIb) exercise markedly improved learning performance. Exercise significantly shortened the escape latencies compared with ND-treated rats across all trials (p<0.001). In the early learning phase (AT1–AT2), exercised rats performed better than controls (p<0.05), suggesting that exercise facilitated acquisition. No significant difference was detected between the effects of aerobic and anaerobic exercises (p > 0.05). In the combined treatment groups (GIVa & GIVb), aerobic and anaerobic exercise partially improve ND-induced impairment. Rats receiving ND combined with exercise (GIVa and GIVb) showed a significant decrease in escape latency compared with ND alone (p<0.001). However, their performance stayed less than that of the exercise-only groups (GIIIa & GIIIb) (p<0.001 in most trials). Aerobic training (GIVa) generally exerted a more protective effect than anaerobic training (GIVb), as evidenced by longer latencies across several trials. In all Barnes maze escape latency trials (AT1–AT6), ND treatment and exercise modality consistently yielded large effect sizes (η² = 0.972–0.982), indicating potent, biologically significant, and consistent effects on spatial learning acquisition and cognitive performance.
Table 2 shows the Number of Errors Across Acquisition Trials (AT1–AT6). The error analysis results aligned with the latency results. GII showed the highest error counts across all trials, far outmatching those of controls (p<0.001), reflecting an impaired spatial search strategy and reduced learning aptitude. Exercise only groups (GIIIa & GIIIb) showed a markedly reduced number of errors relative to both GI and GII across nearly all trials (p<0.05 vs GI; p<0.001 vs GII). Aerobic and anaerobic training did not differ significantly (p>0.05). In the combined groups, both GIVa and GIVb demonstrated intermediate performance. While errors were significantly fewer than in GII rats (p<0.001), they remained elevated compared with exercise-only subgroups (GIIIa & GIIIb) (p<0.001). GIVb made more errors than GIVa, indicating higher neuroprotection for aerobic training when combined with ND than anaerobic training. In all Barnes maze tests, the number of errors across trials (AT1–AT6), ND treatment and exercise modality showed a consistently large effect size (η² = 0.900–0.987), indicating robust, biologically meaningful, and consistent effects on spatial learning acquisition and cognitive performance.
Table 3 shows the Probe trials (PT1 and PT2), which measure spatial reference memory and retention of the learned escape location, demonstrated a compatible pattern across all experimental groups. ND administration produced a considerable impairment in memory performance, as evidenced by the markedly prolonged escape latencies in both PT1 and PT2 compared with controls (p<0.001). This shows a significant disruption of hippocampal-dependent spatial memory recollection following chronic ND exposure. In contrast, both aerobic and anaerobic exercise produced strong memory-enhancing effects. Rats in subgroups GIIIa and GIIIb showed the shortest latencies in both probe trials, compared to those in GI and GII (p<0.001 for all). These data suggest that regular physical exercise raises the quality of memory and recovery capabilities, with significant efficacy for both aerobic and anaerobic training. When exercise was combined with ND in GIVa and GIVb, escape latencies improved compared with ND alone, indicating partial protection against ND-induced cognitive impairment. However, these ND and exercise-combined subgroups (GIVa & GIVb) showed longer latencies than the exercise-only subgroups (GIIIa and GIIIb) (P>0.001 for both). Aerobic training (GIVa) significantly exceeded anaerobic training (GIVb), indicating a greater neuroprotective effect of aerobic exercise against ND-induced neurotoxicity. In both probe trials (PT1 and PT2), ND treatment and exercise modality showed large effect sizes (η² = 0.988–0.991), indicating a potent and significant effect on spatial memory retrieval and retention.
Table 4 shows the Number of Errors in Probe Trial (PT1 & PT2). Performance of rats during PT1 and PT2 was fully consistent with the latency outcomes, further confirming the effects of ND and exercise on spatial memory accuracy. Rats administered ND alone (GII) exhibited the highest number of errors in both probe trials, exceeding those of all other groups (p<0.001 for all). This elevated error rate indicated a significant impairment in spatial memory recall, as reflected in a decrease in the rats’ ability to correctly identify the target hole, consistent with ND-induced hippocampal dysfunction. Conversely, both aerobic and anaerobic exercise subgroups (GIIIa and GIIIb) showed the most accurate performance, achieving the fewest errors across both Probe trials (p<0.001 vs GI and GII). This improvement indicates the strong memory-enhancing effect of routine physical training and its role in improving spatial search strategies. In contrast, the combination of exercises with ND treatment in both subgroups GIVa and GIVb showed significantly fewer errors than GII, indicating partial neuroprotection conferred by exercises against ND-induced impairment. While error rates remained higher than those observed in GIIIa and GIIIb. Moreover, GIVa continually exceeded GIVb, suggesting that aerobic training provides superior protection of navigational accuracy during AAS exposure. The number of errors across both probe trials (PT1 and PT2), ND exposure, and exercise modality showed large effect sizes (η² = 0.982–0.989), indicating highly significant effects on spatial memory retrieval and retention.
3.1.2. The Modified T-Maze Test:
Table 5 shows the results of the Modified T-Maze Test, indicating significant differences in working memory performance among groups. In GII, rats showed significantly lower correct-choice percentages than in GI (p<0.001). Moreover, both exercise modalities (GIIIa and GIIIb) showed significant improvements in memory accuracy (p<0.001 & p<0.05, respectively), with no significant difference between the two modalities (P>0.05). In contrast, the combination of exercises with ND (GIVa and GIVb) partially improved performance (P<0.001); however, performance remained below the levels observed in GIIIa and GIIIb (P<0.001 vs both GIIIa and GIIIb). Aerobic exercise in GIVa showed better outcomes than anaerobic exercise in GIVb (P<0.05).
3.1.3. Rotarod Test Latency:
Table 5 shows that rotarod performance indicated significant differences in motor coordination and balance among groups. ND exposure significantly lowered latency to fall in GII compared with GI (P<0.001). In contrast, GIIIa and GIIIb showed the longest latencies, indicating superior motor performance (p<0.001 vs all groups). Combined training and ND (GIVa & GIVb) enhanced performance relative to ND alone (GII) (P<0.001 for both), but did not reach the level of exercise-only groups and remained less than GI. Also, there were insignificant differences in performance between GIVa and GIVb. ND exposure and exercise modality resulted in large effect sizes for both Modified T-Maze performance (η² = 0.884) and Rotarod Latency (η² = 0.968). These findings demonstrated significant impacts on memory, cognitive flexibility, motor coordination, and neuromuscular performance.
3.2. Biochemical Assays:
Table 6 shows the results of the biochemical assays: GII showed a significant reduction in Neuroplasticity Marker levels, BDNF compared with GI (P<0.05), indicating impaired neuroplasticity. Conversely, both GIIIa and GIIIb showed higher BDNF levels exceeding those of GI and GII (p<0.001 for all). In contrast, GIVa and GIVb partially restored BDNF but remained significantly lower than exercise alone (P<0.001 for all), demonstrating that training reduces but does not fully reverse the neurotoxic effects of ND. Oxidative stress marker, serum MDA levels showed their highest levels in GII (p<0.001 vs all groups), indicating severe oxidative stress, while GIIIa and GIIIb showed markedly reduced MDA levels (P<0.0001 vs all groups), demonstrating the powerful antioxidant effects of both exercise modalities. The combination of exercise and ND in subgroups GIVa & GIVb showed a significant reduction in oxidative stress compared with GII; however, they still exhibited elevated oxidative stress compared with GIIIa and GIIIb (P<0.001), indicating partial protection. Moreover, the ND significantly suppressed serum SOD activity in GII (P<0.001), whereas exercise modalities in both GIIIa and GIIIb dramatically increased SOD values compared with all other groups, with aerobic training showing a superior antioxidant effect compared with anaerobic training (P<0.001). In contrast, combined aerobic and anaerobic training with ND in GIVa and GIVb improved SOD levels relative to ND alone in GII (P<0.001 for all), but remained significantly less than both GIIIa and GIIIb (P<0.001). ND exposure and exercise modality showed large effect sizes across all biochemical parameters, including serum BDNF levels, MDA, and SOD (η² = 0.997, 0.992, and 0.998, respectively). These findings indicate significant effects on neuroplasticity, oxidative stress, lipid peroxidation, and antioxidant defense.
3.3. Histopathological Study:
Figure 1 and Figure 2 show the histopathological examination of the dentate gyrus in all experimental groups at low and high magnifications.
Figure 3 exhibited immunohistochemical examination of the DG in all experimental groups.
Table 7 shows the results of measuring hippocampal GCL thickness, which was markedly reduced in the GII compared with all other groups (p<0.05). Aerobic exercise alone (GIIIa) produced the highest mean thickness and did not differ significantly from GI, indicating preservation of hippocampal architecture. Similarly, resistance training (GIIIb) maintained thickness at levels comparable to those of GI. In contrast, combined treatment subgroups (GIVa and GIVb) exhibited partial structural recovery. GIVa and GIVb showed significantly greater thickness than GII (p<0.05) but remained significantly reduced compared with exercise-only groups, with GIVb displaying the most considerable reduction among the combined groups. Moreover, GII showed a profound reduction in the number of NGF-immunostained cells compared with all other groups (p<0.05). Aerobic exercise in GIIIa markedly increased neuronal count, which was significantly higher than GI, GII, and the combined treatment subgroups (GIVa & GIVb) (p<0.05). Also, resistance exercise (GIIIb) significantly increased NGF expression compared with GII, to a lesser extent than aerobic training (GIVa). The combined exercise with ND (GIVa and GIVb) improved NGF expression compared with GII (p<0.05); however, both subgroups remained significantly lower than GIIIa and GIIIb, indicating partial neurotrophic protection conferred by exercise against chronic ND treatment.
4. Discussion
Several limitations should be regarded when analyzing the findings of the present study. First, the inclusion of only adult male rats limits the generalizability of the results to females, particularly given potential sex-related differences in the hormonal regulation of oxidative stress responses and hippocampal neuroplasticity. The second is that the relatively small sample size may reduce statistical power to detect relations between ND exposure and exercise modality. The third is that although the study integrated behavioral, biochemical, histological, and immunohistochemical outcomes, additional molecular analyses are needed to clarify the underlying mechanisms. Finally, the findings should be interpreted with caution on the human relevance, as experimental ND dosing paradigms may not fully reflect the patterns of AAS abuse in clinical or athletic settings. The present findings show that prolonged ND exposure induces pronounced cognitive dysfunction accompanied by redox imbalance and structural alterations within the hippocampus. ND-treated animals exhibited impaired spatial acquisition, reduced working memory, and reduced cognitive performance across behavioral paradigms, suggesting widespread disruption of hippocampal-dependent processing. Similar patterns of morphological and cognitive impairment have been described in humans exposed to supra-physiological chronic AAS abuse [36,37]. In contrast, animals that participated in regular physical activity showed a potent protective effect. Both aerobic and resistance exercises enhanced memory and cognitive performance, with aerobic exercise showing the highly detectable benefits. These results are consistent with extensive evidence indicating that regular physical training enhances hippocampal plasticity and brain perfusion, and promotes adult neurogenesis, particularly learning and memory consolidation [38,39].
A comparable study has further suggested that endurance-based exercise may result in a greater cognitive improvement than anaerobic modality, especially in domains such as spatial navigation and executive performance [40]. The greatest outcomes observed in the aerobic group, therefore, appear biologically significant and compatible with the known sensitivity of hippocampal circuits to maintained aerobic stimulation [41]. The cognitive impairment associated with ND administration may be attributed to steroid-induced alterations in hippocampal circuitry. Previous studies have shown that supra-physiological doses of AAS affect dopaminergic signaling and modify dendritic spine density within hippocampal subfields, possibly compromising the neural substrates underlying spatial and short-term memory [42]. Biochemical analyses further support ND-induced neurotoxicity, characterized by elevated MDA levels and reduced SOD activity, demonstrating a shift toward a peroxidative state. In contrast, both exercise modalities reduced lipid peroxidation and enhanced endogenous antioxidant activity. This antioxidant support is well documented in the literature, which defines regular exercise as a potent modulator of redox homeostasis [43,44]. The lowered oxidative burden in exercised groups provides a reasonable explanation for the enhanced behavioral and structural outcomes. Furthermore, a decline in hippocampal BDNF was observed in GII. This reduction in BDNF may reflect a stress-like response induced by AAS exposure, which has been linked to increased endogenous corticosterone. Elevated corticosterone is known to suppress hippocampal BDNF expression, as demonstrated under immobilization stress conditions [45]. Conversely, the exercise groups showed elevated BDNF levels, a finding supported by accumulating evidence indicating a beneficial role of physical activity in enhancing brain functional plasticity [46]. Exercise also increases levels of growth factors beyond BDNF, including insulin-like growth factor-1 (IGF-1) [47]. Nay et al. [48] demonstrated that four weeks of treadmill training increased hepatic IGF-1 mRNA expression in rats. Moreover, resistance exercise has been associated with higher circulating IGF-1 levels, which can cross the blood–cerebrospinal fluid barrier and promote neurogenesis, particularly in the hippocampus [49]. In the present study, histopathological investigations corroborated the proposed mechanisms and revealed reduced hippocampal thickness, neuronal loss, and degenerative cytological features in GII. Exercises, particularly aerobic training, preserved tissue morphology and reduced the degenerative changes. It was consistent with prior findings that physical activity ameliorates structural deterioration and supports hippocampal integrity [24,50].
Moreover, the immunohistochemical investigation of NGF expression suggested an additional mechanism underlying the neurobiological alterations underlying these behavioral changes. ND markedly suppressed NGF immunoreactivity, consistent with diminished neurotrophic signaling, whereas both exercise paradigms restored NGF expression, with aerobic exercise producing the greatest effect. These observations are consistent with previous reports demonstrating that exercise promotes neurotrophin expression and maintains neuronal integrity, whereas AAS exposure suppresses neurotrophic pathways [51,52]. Concurrently, these findings indicate that ND abuse exerts its cognitive and structural toxicity through varied mechanisms, including oxidative stress, neurotrophic suppression, and hippocampal degeneration. Contrarily, regular physical exercise, particularly aerobic training, counteracts these harmful effects by enhancing antioxidant defenses, preserving neuronal integrity, and upregulating BDNF.
5. Conclusions
Collectively, our study demonstrated that the convergence of behavioral, biochemical, histological, and immunohistochemical findings supports the potential of exercise as an accessible, non-pharmacological approach to reduce the neurological consequences of AAS misuse. Further studies are needed to examine the effects of varying doses and durations of AAS exposure to better understand the dose–response relationship in neurocognitive and structural brain alterations. Additionally, future research on female rats is recommended to assess sex-related differences in responses to AAS and exercise on brain function and to investigate the potential synergistic benefits of combining exercise with other neuroprotective agents to antagonize AAS-induced neurotoxicity.
Author Contributions
Conceptualization, Wesam M.; methodology, Wesam M., Abeer K., Lamiaa A., Enas M., Rasha E., and Ayman M.; software, Saleh A.; validation, Hadeel E., Lamiaa A., and Abeer K.; formal analysis, Lamiaa A. and Abeer K.; investigation, Enas M., Saleh A.; resources, Wesam M., Enas M., and Hadeel E.; data curation, Lamiaa A.; writing—original draft preparation, Wesam M., Enas M.; writing—review and editing, Enas M., Rasha E., and Ayman M.; visualization, Wesam M., Enas M., and Hadeel E.; supervision, Saleh A., Wesam M., Enas M., and Hadeel E.; project administration, Wesam M., Enas M., and Hadeel E.; funding acquisition, no external funding. All authors have read and agreed to the published version of the manuscript.
Funding
“This research received no external funding.”.
Institutional Review Board Statement
All experimental procedures involving animals in the current study were approved and conducted in accordance with the ARRIVE Essential 10 and the institutional guidelines for the Care and Use of Laboratory Animals by the Institutional Animal Care and Use Committee, Zagazig University (ZU-IACUC) (Approval No. ZU-IACUC/3/F/281/2026).
Informed Consent Statement
“Not applicable.”.
Data Availability Statement
Data will be available from the corresponding author upon reasonable request.
Acknowledgments
The researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University (www.qu.edu.sa) for financial support (QU-APC-2026).
Conflicts of Interest
The authors declare no conflicts of interest.
Artificial Intelligence Disclosure Statement
The authors used ChatGPT (OpenAI, USA; version 5.5, 2025) as an AI-assisted writing tool to improve the manuscript’s English, grammar, and clarity. The authors take full responsibility for the content of the manuscript, including the accuracy of the data, interpretations, and conclusions.
References
- Saab, B.J.; Georgiou, J.; Nath, A.; Lee, F.J.; Wang, M.; Michalon, A.; et al. NCS-1 in the dentate gyrus promotes exploration, synaptic plasticity, and rapid acquisition of spatial memory. Neuron 2009, 63(5), 643–656. [Google Scholar] [CrossRef]
- Khalili, S.; Khoshravesh, S.; Barati, M.; Hazavehei, S.M.M.; Rezapour, R. Androgenic-anabolic steroids use among bodybuilders in western Iran: Application of ridge logistic regression model. BMC Sports Sci. Med. Rehabil. 2023, 15(1), 7. [Google Scholar] [CrossRef]
- Perry, J.C.; Schuetz, T.M.; Memon, M.D.; Faiz, S.; Cancarevic, I. Anabolic steroids and cardiovascular outcomes: The controversy. Cureus 2020, 12(7), e9333. [Google Scholar] [CrossRef] [PubMed]
- Alsaeed, I.; Alabkal, J.R. Usage and perceptions of anabolic-androgenic steroids among male fitness centre attendees in Kuwait: A cross-sectional study. Subst. Abus. Treat. Prev. Policy 2015, 10(1), 33. [Google Scholar] [CrossRef] [PubMed]
- Al-Falasi, O.; Al-Dahmani, K.; Al-Eisaei, K.; Al-Ameri, S.; AlMaskari, F.; Nagelkerke, N.; et al. Knowledge, attitude and practice of anabolic steroids use among gym users in Al-Ain district, United Arab Emirates. Open Sports Med. J. 2008, 2, 75–81. [Google Scholar] [CrossRef]
- Bahrke, M.S.; Yesalis, C.E.; Brower, K.J. Anabolic-androgenic steroid abuse and performance-enhancing drugs among adolescents. Child. Adolesc. Psychiatr. Clin. N Am. 1998, 7(4), 821–838. [Google Scholar] [CrossRef]
- Alhekail, O.; Almeshari, A.; Alabdualkarim, B.; Alkhalifa, M.; Almarek, N.; Alzuman, O.; et al. Prevalence and patterns of the use of protein supplements among gym users in Riyadh, Saudi Arabia. Int. J. Pharm. Res. Allied Sci. 2018, 7(1), 80–86. [Google Scholar]
- Abd El-Raouf, M.; Araby, E. Anabolic steroids use and perceived side effects among gymnasium attendees in Benha and Shebin Al-Kom cities. Egypt J. Community Med. 2022, 40(2), 114–120. [Google Scholar] [CrossRef]
- Saddick, S.Y. Effect of nandrolone decanoate induced-oxidative stress on rat testes, prostate, and seminal vesicle: Biochemical, morphometric and histopathological studies. Saudi J. Biol. Sci. 2021, 28(1), 196–203. [Google Scholar] [CrossRef]
- Patanè, F.G.; Liberto, A.; Maria Maglitto, A.N.; Malandrino, P.; Esposito, M.; Amico, F.; et al. Nandrolone decanoate: Use, abuse and side effects. Medicina 2020, 56(11), 606. [Google Scholar] [CrossRef]
- Wenbo, Z.; Yan, Z. The uses of anabolic androgenic steroids among athletes: Its positive and negative aspects—a literature review. J. Multidiscip. Healthc. 2023, 16, 4293–4305. [Google Scholar] [CrossRef]
- Windfeld-Mathiasen, J.; Christoffersen, T.; Strand, N.A.W.; Dalhoff, K.; Andersen, J.T.; Horwitz, H. Psychiatric morbidity among men using anabolic steroids. Depress Anxiety 2022, 39(12), 805–812. [Google Scholar] [CrossRef] [PubMed]
- Joksimovic, J.; Selakovic, D.; Matovic, M.; Zaletel, I.; Puskas, N.; Rosic, G. The role of neuropeptide-Y in nandrolone decanoate-induced attenuation of antidepressant effect of exercise. PLoS ONE 2017, 12(6), e0178922. [Google Scholar] [CrossRef]
- Scarth, J.P.; Bjørnebekk, A. Androgen abuse and the brain. Curr. Opin. Endocrinol. Diabetes Obes. 2021, 28(6), 604–614. [Google Scholar] [CrossRef]
- Kanayama, G.; Kaufman, M.J.; Pope, H.G., Jr. Public health impact of androgens. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25(3), 218–223. [Google Scholar] [CrossRef]
- El-Shamarka, M.E.S.; Sayed, R.H.; Assaf, N.; Zeidan, H.M.; Hashish, A.F. Combined neurotoxic effects of cannabis and nandrolone decanoate in adolescent male rats. Neurotoxicology 2020, 76, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Zahid, M.F.; Zain, S.; Kabir, A.; Hassan, S.U. The neuroprotective effects of exercise on cognitive decline: A preventive approach to Alzheimer disease. Cureus 2020, 12(2), e6958. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, L.; Polverino, A.; Montuori, S.; Foti, F.; Ferraioli, G.; Sorrentino, P.; et al. Effects of physical exercise on cognitive functioning and wellbeing: Biological and psychological benefits. Front Psychol. 2018, 9, 509. [Google Scholar] [CrossRef]
- Bredin, S.S.D.; Warburton, D.E.R.; Lang, D.J. The health benefits and challenges of exercise training in persons living with schizophrenia: A pilot study. Brain Sci. 2013, 3(2), 821–848. [Google Scholar] [CrossRef]
- Tari, A.R.; Walker, T.L.; Huuha, A.M.; Sando, S.B.; Wisloff, U. Neuroprotective mechanisms of exercise and the importance of fitness for healthy brain ageing. Lancet 2025, 405(10484), 1093–1118. [Google Scholar] [CrossRef]
- Vanhees, L.; Geladas, N.; Hansen, D.; Kouidi, E.; Niebauer, J.; Reiner, Ž.; et al. Importance of characteristics and modalities of physical activity and exercise in the management of cardiovascular health in individuals with cardiovascular risk factors: Recommendations from the EACPR (Part II). Eur. J. Prev. Cardiol. 2012, 19(5), 1005–1033. [Google Scholar] [CrossRef]
- Gallardo-Gómez, D.; del Pozo-Cruz, J.; Noetel, M.; Álvarez-Barbosa, F.; Alfonso-Rosa, R.M.; del Pozo Cruz, B. Optimal dose and type of exercise to improve cognitive function in older adults: A systematic review and Bayesian model-based network meta-analysis of RCTs. Ageing Res. Rev. 2022, 76, 101591. [Google Scholar] [CrossRef]
- Buchheister, S.; Bleich, A. Health monitoring of laboratory rodent colonies: Talking about (R)evolution. Animals 2021, 11(5), 1410. [Google Scholar] [CrossRef] [PubMed]
- Diniz e Magalhães, C.O.; Oliveira, L.R.S.; Santos, S.T.; Mendes, B.F.; Dias, I.R.; Cruz, P.P.; et al. Nandrolone decanoate reduces the positive effects of resistance training on cognition, anxious behavior, and hippocampal morphology in rats. Res. Soc. Dev. 2022, 11(8), e10511830600. [Google Scholar] [CrossRef]
- De Assis, G.G.; de Souza, E.O.N.; de Almeida-Neto, P.F.; Ceylan, H.I.; Bragazzi, N.L. A proposal for a noxious stimuli-free, moderate-intensity treadmill running protocol to improve aerobic performance in experimental research on rats. Metabolites 2024, 14(10), 534. [Google Scholar] [CrossRef] [PubMed]
- Segabinazi, E.; Gasperini, N.F.; Faustino, A.M.; Centeno, R.; Santos, A.S.D.; Almeida, W.; et al. Comparative overview of the effects of aerobic and resistance exercise on anxiety-like behavior, cognitive flexibility, and hippocampal synaptic plasticity parameters in healthy rats. Braz. J. Med. Biol. Res. 2020, 53(11), e9816. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents: Methodological consideration. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392(1), 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.C.; Lerner, F.M.; Couto E Silva, A.; Possoit, H.E.; Hsieh, T.H.; Neumann, J.T.; et al. Utilizing the modified T-maze to assess functional memory outcomes after cardiac arrest. J. Vis. Exp. 2018, (131), 56694. [Google Scholar] [CrossRef]
- Ataie, A.; Sabetkasaei, M.; Haghparast, A.; Hajizadeh Moghaddam, A.; Ataee, R.; Nasiraei Moghaddam, S. Curcumin exerts neuroprotective effects against homocysteine intracerebroventricular injection-induced cognitive impairment and oxidative stress in rat brain. J. Med. Food 2010, 13(4), 821–826. [Google Scholar] [CrossRef]
- Kale, V.P.; Patel, S.G.; Gunjal, P.S.; Wakchaure, S.U.; Sundar, R.S.; Ranvir, R.K.; et al. Effect of repeated freezing and thawing on 18 clinical chemistry analytes in rat serum. J. Am. Assoc. Lab Anim. Sci. 2012, 51(4), 475–478. [Google Scholar]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem Biophys. 1984, 21(2), 130–132. [Google Scholar]
- Nielsen, F.; Mikkelsen, B.B.; Nielsen, J.B.; Andersen, H.R.; Grandjean, P. Plasma malondialdehyde as biomarker for oxidative stress: Reference interval and effects of lifestyle factors. Clin. Chem. 1997, 43(7), 1209–1214. [Google Scholar] [CrossRef]
- Lim, Y.; Zhong, J.H.; Zhou, X.F. Development of mature BDNF-specific sandwich ELISA. J. Neurochem. 2015, 134(1), 75–85. [Google Scholar] [CrossRef]
- Soliman, M.; Kefafy, M.; Mansour, M.; Ali, A.; Esa, W.I. Histological study on the possible protective effect of pentoxifylline on pancreatic acini of L-arginine-induced acute pancreatitis in adult male albino rats. Menoufia Med. J. 2014, 27(4), 801. [Google Scholar]
- Cattoretti, G.; Pileri, S.; Parravicini, C.; Becker, M.H.G.; Poggi, S.; Bifulco, C.; et al. Antigen unmasking on formalin-fixed, paraffin-embedded tissue sections. J. Pathol. 1993, 171(2), 83–98. [Google Scholar] [CrossRef]
- Bertozzi, G.; Salerno, M.; Pomara, C.; Sessa, F. Neuropsychiatric and behavioral involvement in AAS abusers: A literature review. Medicina 2019, 55(7), 396. [Google Scholar] [CrossRef]
- Kaufman, M.J.; Janes, A.C.; Hudson, J.I.; Brennan, B.P.; Kanayama, G.; Kerrigan, A.R.; et al. Brain and cognition abnormalities in long-term anabolic-androgenic steroid users. Drug Alcohol Depend. 2015, 152, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Koo, J.H.; Cho, J.Y.; Kang, E.B. Neuroprotective effect of treadmill exercise against blunted brain insulin signaling, NADPH oxidase, and tau hyperphosphorylation in rats fed a high-fat diet. Brain Res. Bull. 2018, 142, 374–383. [Google Scholar] [CrossRef]
- Sanaeifar, F.; Pourranjbar, S.; Pourranjbar, M.; Ramezani, S.; Mehr, S.R.; Wadan, A.H.S.; et al. Beneficial effects of physical exercise on cognitive-behavioral impairments and brain-derived neurotrophic factor alteration in the limbic system induced by neurodegeneration. Exp. Gerontol. 2024, 195, 112539. [Google Scholar] [CrossRef]
- Zhang, M.; Jia, J.; Yang, Y.; Zhang, L.; Wang, X. Effects of exercise interventions on cognitive functions in healthy populations: A systematic review and meta-analysis. Ageing Res. Rev. 2023, 92, 102116. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. U S A 2011, 108(7), 3017–3022. [Google Scholar] [CrossRef]
- Wood, R.I.; Serpa, R.O. Anabolic-androgenic steroid abuse and cognitive impairment: Testosterone impairs biconditional task performance in male rats. Behav. Brain Res. 2020, 379, 112339. [Google Scholar] [CrossRef]
- Hyatt, H.W.; Smuder, A.J.; Sollanek, K.J.; Morton, A.B.; Roberts, M.D.; Kavazis, A.N. Comparative changes in antioxidant enzymes and oxidative stress in cardiac, fast twitch and slow twitch skeletal muscles following endurance exercise training. Int. J. Physiol. Pathophysiol. Pharmacol. 2016, 8(4), 160–168. [Google Scholar]
- Powers, S.K.; Goldstein, E.; Schrager, M.; Ji, L.L. Exercise training and skeletal muscle antioxidant enzymes: An update. Antioxidants 2023, 12(1), 39. [Google Scholar] [CrossRef] [PubMed]
- Costine, B.A.; Oberlander, J.G.; Davis, M.C.; Penatti, C.A.; Porter, D.M.; Leaton, R.N.; et al. Chronic anabolic androgenic steroid exposure alters corticotropin releasing factor expression and anxiety-like behaviors in the female mouse. Psychoneuroendocrinology 2010, 35(10), 1473–1485. [Google Scholar] [CrossRef]
- Gajewski, P.D.; Falkenstein, M. Physical activity and neurocognitive functioning in aging: A condensed updated review. Eur. Rev. Aging Phys. Act. 2016, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhao, X.; Li, B.; Cai, Y.; Zhang, S.; Wan, Q.; et al. Comparative efficacy of various exercise interventions on cognitive function in patients with mild cognitive impairment or dementia: A systematic review and network meta-analysis. J. Sport Health Sci. 2022, 11(2), 212–223. [Google Scholar] [CrossRef]
- Nay, K.; Smiles, W.J.; Kaiser, J.; McAloon, L.M.; Loh, K.; Galic, S.; et al. Molecular mechanisms underlying the beneficial effects of exercise on brain function and neurological disorders. Int. J. Mol. Sci. 2021, 22(8), 4052. [Google Scholar] [CrossRef]
- Trejo, J.L.; Llorens-Martín, M.V.; Torres-Alemán, I. The effects of exercise on spatial learning and anxiety-like behavior are mediated by an IGF-I-dependent mechanism related to hippocampal neurogenesis. Mol. Cell Neurosci. 2008, 37(2), 402–411. [Google Scholar] [CrossRef] [PubMed]
- Novaes Gomes, F.G.; Fernandes, J.; Vannucci Campos, D.; Cassilhas, R.C.; Viana, G.M.; D’Almeida, V.; et al. The beneficial effects of strength exercise on hippocampal cell proliferation and apoptotic signaling is impaired by anabolic androgenic steroids. Psychoneuroendocrinology 2014, 50, 106–117. [Google Scholar] [CrossRef]
- Hong, Y.P.; Lee, H.C.; Kim, H.T. Treadmill exercise after social isolation increases the levels of NGF, BDNF, and synapsin I to induce survival of neurons in the hippocampus, and improves depression-like behavior. J. Exerc Nutr. Biochem. 2015, 19(1), 11–18. [Google Scholar] [CrossRef]
- Selakovic, D.; Joksimovic, J.; Zaletel, I.; Puskas, N.; Matovic, M.; Rosic, G. The opposite effects of nandrolone decanoate and exercise on anxiety levels in rats may involve alterations in hippocampal parvalbumin-positive interneurons. PLoS ONE 2017, 12(12), e0189595. [Google Scholar] [CrossRef]
Figure 1.
Shows representative photomicrographs from sections of the hippocampus in all groups of rats stained with H&E. The dentate gyrus (DG) below the hippocampal sulcus (HS) shows three layers [granule cell layer (GCL), subgranular zone, and molecular cell layer (MCL)]. The GCL has closely arranged neurons in the control group (GI), whereas the GCL shows a marked reduction in thickness, accompanied by numerous dilated blood capillaries (DBC), indicating neurodegenerative changes in the ND-treated group (GII). In contrast, the DG shows a marked increase in GCL thickness in the aerobic subgroup (GIIIa), with a dense neuronal arrangement, and a normal thickness in the anaerobic subgroup (GIIIb), with closely packed neurons. Also, the DG shows improvement in the aerobic+ND subgroup (GIVa), evidenced by a considerable increase in GCL thickness, and a mild improvement in the anaerobic+ND subgroup (GIVb), as indicated by increased GCL thickness relative to GII. (H&E, X200) .
Figure 1.
Shows representative photomicrographs from sections of the hippocampus in all groups of rats stained with H&E. The dentate gyrus (DG) below the hippocampal sulcus (HS) shows three layers [granule cell layer (GCL), subgranular zone, and molecular cell layer (MCL)]. The GCL has closely arranged neurons in the control group (GI), whereas the GCL shows a marked reduction in thickness, accompanied by numerous dilated blood capillaries (DBC), indicating neurodegenerative changes in the ND-treated group (GII). In contrast, the DG shows a marked increase in GCL thickness in the aerobic subgroup (GIIIa), with a dense neuronal arrangement, and a normal thickness in the anaerobic subgroup (GIIIb), with closely packed neurons. Also, the DG shows improvement in the aerobic+ND subgroup (GIVa), evidenced by a considerable increase in GCL thickness, and a mild improvement in the anaerobic+ND subgroup (GIVb), as indicated by increased GCL thickness relative to GII. (H&E, X200) .

Figure 2.
Shows representative photomicrographs from higher magnifications of the hippocampal sections in all groups of rats stained with H&E. The DG shows normal thickness of the GCL, with intact closely arranged granular neurons that exhibit normal nuclear morphology in GI, while shows a marked reduction in the thickness of the GCL, accompanied by an increased number of degenerated neurons (black arrows) at the MCL, indicating neurodegenerative changes in GII. In contrast, the DG shows a marked increase in GCL thickness, with dense, intact granular neurons in GIIIa, and a normal GCL thickness, with closely packed, intact neurons in GIIIb. Also, the DG shows a moderate improvement in GIVa, evidenced by increased GCL thickness and fewer degenerated neurons (black arrows), whereas it shows a mild improvement in GIVb, demonstrated by increased GCL thickness and fewer degenerated neurons (black arrows) compared to GII. (H&E, X400).
Figure 2.
Shows representative photomicrographs from higher magnifications of the hippocampal sections in all groups of rats stained with H&E. The DG shows normal thickness of the GCL, with intact closely arranged granular neurons that exhibit normal nuclear morphology in GI, while shows a marked reduction in the thickness of the GCL, accompanied by an increased number of degenerated neurons (black arrows) at the MCL, indicating neurodegenerative changes in GII. In contrast, the DG shows a marked increase in GCL thickness, with dense, intact granular neurons in GIIIa, and a normal GCL thickness, with closely packed, intact neurons in GIIIb. Also, the DG shows a moderate improvement in GIVa, evidenced by increased GCL thickness and fewer degenerated neurons (black arrows), whereas it shows a mild improvement in GIVb, demonstrated by increased GCL thickness and fewer degenerated neurons (black arrows) compared to GII. (H&E, X400).

Figure 3.
Shows representative high-magnification photomicrographs of hippocampal sections from all rat groups, immunostained for nerve growth factor (NGF). The DG shows preserved standard architecture in GI, with normal thickness of the GCL that exhibit mild cytoplasmic immunoexpression (CIE) of NGF in few scattered granule cells (red stars) and the neuropil (green stars) and negative CIE of NGF in most of the granule cells (yellow stars), whereas it shows a marked reduction in the thickness of the GCL and their neuronal density in GII, accompanied with a weak CIE of NGF in few granule cells (red stars) and negative in most of the granule cells (yellow stars), indicating neurodegenerative changes in GII. At the same time, the DG shows preserved architecture, with increased cellularity in the dentate granular layer and moderate CIE of NGF in several neurons (red stars) and in the neuropil (green stars), whereas other neuronal elements appear negative (yellow stars) in GIIIa. Also, GIIIb shows a mild focal increase in GCL cellularity, with mild positive CIE of NGF in scattered neurons (red stars) and neuropil (green stars), while most neuronal elements show negative CIE of NGF (yellow stars). In contrast, the DG shows a moderate increase in GCL cellularity, with weak CIE of NGF in a few neurons (red stars) and the neuropil (green stars), while most neuronal elements remain negative (yellow stars) in GIVa. Also, the DG shows a mild focal increase in the cellularity of the dentate GCL with weak CIE of NGF in a few neurons (red stars) and the neuropil (green stars), while most of the neuronal elements remain negative (yellow stars), showing a mild improvement in GIVb compared to GII. (NGF, X400).
Figure 3.
Shows representative high-magnification photomicrographs of hippocampal sections from all rat groups, immunostained for nerve growth factor (NGF). The DG shows preserved standard architecture in GI, with normal thickness of the GCL that exhibit mild cytoplasmic immunoexpression (CIE) of NGF in few scattered granule cells (red stars) and the neuropil (green stars) and negative CIE of NGF in most of the granule cells (yellow stars), whereas it shows a marked reduction in the thickness of the GCL and their neuronal density in GII, accompanied with a weak CIE of NGF in few granule cells (red stars) and negative in most of the granule cells (yellow stars), indicating neurodegenerative changes in GII. At the same time, the DG shows preserved architecture, with increased cellularity in the dentate granular layer and moderate CIE of NGF in several neurons (red stars) and in the neuropil (green stars), whereas other neuronal elements appear negative (yellow stars) in GIIIa. Also, GIIIb shows a mild focal increase in GCL cellularity, with mild positive CIE of NGF in scattered neurons (red stars) and neuropil (green stars), while most neuronal elements show negative CIE of NGF (yellow stars). In contrast, the DG shows a moderate increase in GCL cellularity, with weak CIE of NGF in a few neurons (red stars) and the neuropil (green stars), while most neuronal elements remain negative (yellow stars) in GIVa. Also, the DG shows a mild focal increase in the cellularity of the dentate GCL with weak CIE of NGF in a few neurons (red stars) and the neuropil (green stars), while most of the neuronal elements remain negative (yellow stars), showing a mild improvement in GIVb compared to GII. (NGF, X400).


Table 1.
Escape latency (Sec) across acquisition trials (AT1–AT6).
| Group | AT1 | AT2 | AT3 | AT4 | AT5 | AT6 |
|---|---|---|---|---|---|---|
| GI | 52.67±3.26 | 35.33±5.06 | 41.17±2.64 | 32.67±2.58 | 37.33±3.20 | 33.33±1.97 |
| GII | 117.83±2.14* | 111.0±7.23* | 113.67±3.61* | 115.0±4.29* | 115.0±4.29* | 99.83±7.98* |
| GIIIa | 43.17±1.17*,$ | 24.0±1.79*,$ | 38.33±2.87$ | 28.17±2.32$ | 28.0±2.83*, $ | 19.83±2.32*, $ |
| GIIIb | 44.33±1.63*, $ | 25.83±2.32*, $ | 38.50±2.66$ | 30.83±3.31$ | 30.83±3.31*, $ | 22.33±1.51*, $ |
| GIVa | 69.83±3.43*, $, #, ^ | 49.0±3.89*, $, #,^ | 56.67±4.55*,$,#,^ | 45.33±3.78*,$,#, ^ | 44.5±3.39*, $, #, ^ | 42.67±4.59*, $, #, ^ |
| GIVb | 86.50±7.56*, $, #, ^, † | 56.33±4.18*, $, #,^, † | 65.0±7.32*,$,#,^, † | 51.67±7.55*,$, #, ^ | 59.3±7.86*, $, #, ^, † | 49.5±4.14*, $, #, ^, † |
| η² (Eta-squared) | 0.982 | 0.951 | 0.930 | 0.980 | 0.975 | 0.972 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 2.
Number of errors across acquisition trials (AT1–AT6).
| Group | AT1 | AT2 | AT3 | AT4 | AT5 | AT6 |
|---|---|---|---|---|---|---|
| GI | 7.17±0.76 | 5.5±1.05 | 4.83±0.75 | 3.67±0.82 | 4.0±0.89 | 3.5±0.55 |
| GII | 12.83±0.98* | 11.33±0.82* | 11.67±1.03* | 13.0±0.89* | 11.0±0.89* | 11.18±0.98* |
| GIIIa | 4.33±0.52*,$ | 2.67±0.82*,$ | 2.17±0.41*, $ | 1.33±0.52*, $ | 2.17±0.41*, $ | 1.17±0.41*, $ |
| GIIIb | 4.83±0.98*,$ | 2.83±0.98*,$ | 2.33±0.52*, $ | 1.50±0.55*, $ | 2.50±0.55*, $ | 1.50±0.55*, $ |
| GIVa | 7.83±0.98$,#,^ | 7.50±1.05*,$,#,^ | 6.17±1.17*, $, #, ^ | 5.0±0.89*, $, #, ^ | 5.5±0.55*, $, #, ^ | 4.33±0.52*, $, #, ^ |
| GIVb | 8.50±1.05*,$,#,^ | 8.0±1.26*,$,#,^ | 7.5±1.05*, $, #, ^,† | 6.0±0.89*, $, #, ^, † | 6.5±1.04*, $, #, ^, † | 6.0±0.89*, $, #, ^, † |
| η² (Eta-squared) | 0.925 | 0.900 | 0.954 | 0.981 | 0.964 | 0.987 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 3.
Shows the Escape Latency Across Probe Trial (PT1 & PT2).
| Group | PT1 Latency | PT2 Latency |
|---|---|---|
| GI | 24.17 ± 2.04 | 22.83 ± 3.25 |
| GII | 116.17 ± 4.22* | 117.17 ± 3.43* |
| GIIIa | 17.33 ± 2.06*, $ | 14.67 ± 2.58*, $ |
| GIIIb | 17.83 ± 1.47*, $ | 16.33 ± 3.14 *, $ |
| GIVa | 32.50 ± 2.59*, $, #, ^ | 29.50 ± 1.05*, $, #, ^ |
| GIVb | 41.83 ± 5.42*, $, #, ^, † | 35.17 ± 5.74*, $, #, ^, † |
| η² (Eta-squared) | 0.988 | 0.991 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 4.
Shows the Number of Errors Across Probe Trial (PT1 & PT2).
| Group | PT1 Errors | PT2 Errors |
|---|---|---|
| GI | 3.17 ± 1.17 | 2.17 ± 0.75 |
| GII | 12.33 ± 0.52* | 11.17 ± 0.75* |
| GIIIa | 1.17 ± 0.41*, $ | 0.33 ± 0.52*, $ |
| GIIIb | 1.17 ± 0.41*, $ | 0.50 ± 0.55*, $ |
| GIVa | 4.50 ± 0.84*, $, #, ^ | 3.33 ± 0.52*, $, #, ^ |
| GIVb | 4.50 ± 0.83*, $, #, ^, † | 4.17 ± 1.17*, $, #, ^, † |
| η² (Eta-squared) | 0.982 | 0.989 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 5.
Percentage of Correct Choice in T-maze and Latency to Fall in Rotarod Results.
| Group | T-maze Correct Choices (%) | Rotarod Latency (sec) |
|---|---|---|
| GI | 65.0 ± 5.48 | 46.5 ± 3.78 |
| GII | 23.33 ± 5.16* | 35.0 ± 4.05* |
| GIIIa | 81.67 ± 7.53* $ | 59.33 ± 0.82* $ |
| GIIIb | 78.33 ± 4.08* $ | 58.67 ± 1.97* $ |
| GIVa | 48.33 ± 7.53* $ # ^ | 48.17 ± 3.89$ # ^ |
| GIVb | 40.0 ± 8.94* $ # ^ † | 45.83 ± 3.66$ # ^ † |
| η² (Eta-squared) |
0.968 | 0.884 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 6.
Shows the serum levels of BDNF, MDA, and SOD.
| Group | BDNF (ng/ml) | MDA (nmol/ml) | SOD (U/ml) |
|---|---|---|---|
| GI | 1.53 ± 0.38 | 1.96 ± 0.31 | 42.47 ± 4.04 |
| GII | 0.69 ± 0.22* | 7.09 ± 0.36* | 28.89 ± 6.73* |
| GIIIa | 6.50 ± 0.21*$ | 2.98 ± 0.34*$ | 159.53 ± 6.26*$ |
| GIIIb | 6.41 ± 0.13*$# | 2.96 ± 0.46*$# | 138.89 ± 6.54*$# |
| GIVa | 4.27 ± 0.18*$#^ | 4.26 ± 0.25*$#^ | 108.70 ± 5.41*$#^ |
| GIVb | 4.16 ± 0.12*$#^† | 4.88 ± 0.34*$#^† | 93.11 ± 6.26*$#^† |
| η² (Eta-squared) | 0.997 | 0.992 | 0.998 |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Table 7.
The granule cell layer (GCL) thickness and number of immunostained neurons/ 10 HPF.
| Group | GCL Thickness (Mean ± SD) |
Immunostained Neurons /10 HPF (Mean ± SD) |
|---|---|---|
| GI | 69.1 ± 22.6 | 25.4 ± 3.5 |
| GII | 26.6 ± 6.7 * | 7.8 ± 1.6 * |
| GIIIa | 74.1 ± 14.4 $ | 48.2 ± 7.3 *$ |
| GIIIb | 69.9 ± 14.6 $ | 31.1 ± 6.4 $# |
| GIVa | 54.0 ± 15.1 *$#^ | 25.7 ± 6.7 $ |
| GIVb | 48.4 ± 10.4 *$#^† | 15.8 ± 5.7 *$#^† |
Data are expressed as mean ± SD (n = 6). Significance = p<0.05; * Significant vs GI; $ Significant vs GII; # Significant vs GIIIa; ^ Significant vs GIIIb; † Significant vs GIVa.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



